Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE MONTES CLAROS
PROGRAMA DE PÓS GRADUAÇÃO STRICTO SENSU
EM CIÊNCIAS BIOLÓGICAS
MOSCAS ECTOPARASITAS DE MORCEGOS EM FLORESTAS TROPICAIS
SECAS BRASILEIRAS
PEDRO FONSECA DE VASCONCELOS
Montes Claros. Minas Gerais
2014

PEDRO FONSECA DE VASCONCELOS
MOSCAS ECTOPARASITAS DE MORCEGOS EM FLORESTAS TROPICAIS
SECAS BRASILEIRAS
Dissertação apresentada ao programa de
Pós-Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros, como requisito necessário
para a conclusão do curso de Mestrado em
Ciências Biológicas.
Orientador:
Prof. Dr. Magno Augusto Zazá Borges
Co-orientador:
Prof. Dr. Gustavo Graciolli
Montes Claros, Minas Gerais
2014

PEDRO FONSECA DE VASCONCELOS
MOSCAS ECTOPARASITAS DE MORCEGOS EM FLORESTAS TROPICAIS
SECAS BRASILEIRAS
Dissertação apresentada ao programa de
Pós-Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros, como requisito necessário
para a conclusão do curso de Mestrado em
Ciências Biológicas.
Data da aprovação: ___ de ____________ de 2014
Orientador:___________________________________________
Prof. Dr. Magno Augusto Zazá Borges (UNIMONTES)
Examinadores:________________________________________
Prof. Dr. Lemuel Olívio Leite (UNIMONTES)
________________________________________
Prof. Dr. Daniel Sobreira Rodrigues (EPAMIG)
Montes Claros. Minas Gerais
2014

À minha mãe, meu pai, Kit, João e Carol pelo incentivo diário...
A todos que me ajudaram;
DEDICO

AGRADECIMENTOS
Agradeço a todos aqueles que de alguma maneira, direta ou indiretamente,
contribuíram para a realização deste trabalho.
Ao meu orientador, o Prof. Dr. Magno Augusto Zazá Borges ou Magoo, por me
proporcionar realizar uma parte do projeto que tracei para minha vida. Não posso deixar
de agradecer pela paciência, orientação e por acreditar no meu trabalho. E ao meu co-
orientador, o Prof. Dr. Gustavo Graciolli, por me ajudar e ensinar a identificar os
estreblídeos e nicteribiideos. E principalmente, por ser solícito sempre que precisei,
tirando todas as minhas dúvidas e me acolhendo em Campo Grande – MS.
Agradeço à Luiz Falcão por me proporcionar a oportunidade de enveredar pelos
rumos da pesquisa, me apresentar ao fantástico mundo dos morcegos e possibilitar que
eu realizasse meu projeto. Além disso, pelos conselhos, orientações e pela amizade
durante esses anos de trabalho.
Às pessoas que ajudaram nas coletas de dados (Carine, Gustavo, Hugo e
Paulim), em especial Maria Fernanda (Odetinha) que foi companheira sempre. A
Raphael Neiva por ceder os dados coletados no Parque Estadual da Mata Seca. A Éder
Barbier pelos artigos enviados e ajuda nas análises parasitológicas. E a Karol pela ajuda.
Agradeço ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq - 563304/2010-3 e 562955/2010-0), Fundação de Amparo à Pesquisa de Minas
Gerais (FAPEMIG CRA - APQ-00001-11) e o Inter-American Institute for Global
Change Research (IAI-CRN II-021) pelo financiamento do projeto. E ao Instituto
Estadual de Floresta e ICMBio, por possibilitar a realização da pesquisa de campo.
Agradeço à CAPES pela bolsa de mestrado e ao Programa de Pós-Graduação em
Ciências Biológicas da Universidade Estadual de Montes Claros, pela estrutura

oferecida.
Aos professores de mestrado, especialmente Marcílio, Ronaldo, Mário e Og
pelos ensinamentos.
Agradeço a galera do LEBC pela companhia durante todos esses anos e pelos
momentos de risada. Em especial a Créu, Jamilli, Jú, Mano Brown, Michelle, Aniele e
Quedinha. E aos amigos da biologia Bárbara, Lucas (Lôra), Raissinha, Sarinha e Sara.
Aos colegas de mestrado, em especial a André e Lukete, que se tornaram meus
amigos, e com eles pude compartilhar as angústias de um pós-graduando e me divertir
nos momentos de descontração.
Aos membros da banca examinadora pelas correções e sugestões. Especialmente
ao Prof. Lemuel, por acompanhar este projeto e contribuir para enriquecê-lo.
Não posso deixar de agradecer os meus amigos de longa data, que fora do
mestrado sempre me apoiaram e incentivaram: Aninha, Brunim, Fernando, Gui,
Mateusão, Rennan e Victor. Também agradeço aos familiares que me entenderam nos
momentos de ausência nas festas, em especial minha avó Zelita e minhas tias.
Por fim, e não menos importante, agradeço aos meus pais (Rozânia e Heron) por
todos os dias me incentivarem a correr atrás do meu sonho, por serem meu exemplo de
dedicação e me manterem forte durante toda minha caminhada. Aos meus irmãos, Kit e
João, por me fazerem rir nos dias mais tristes, me aconselharem sempre que preciso e
serem as pessoas com quem posso contar para tudo. À minha namorada Carol, que
entendeu minha ausência todos os dias que precisei ir ao campo ou que estava atarefado,
pelas conversas, pelo companheirismo em todos os momentos, e por ser minha alegria.
Obrigado a todos e peço desculpas aqueles que não me lembrei!
OBRIGADO!

LISTA DE FIGURAS
Figura 1: Mapa com os pontos de coletas das Unidades de Conservação do Norte de
Minas Gerais com Florestas Tropicais Secas, na área II ............................................... 20
Figura 2: Mapa dos pontos de coletas na Serra do Cipó, município de Santana do
Riacho, localizado na região central do estado de Minas Gerais ................................ 21

LISTA DE TABELA
Tabela 1. Associações morcegos-moscas ectoparasitas em três diferentes Florestas
Tropicais Secas Brasileiras .................................................................................... 23

SUMÁRIO
RESUMO ....................................................................................................................... 10
ABSTRACT ................................................................................................................... 11
INTRODUÇÃO .............................................................................................................. 12
MÉTODO ....................................................................................................................... 15
Área de estudo ............................................................................................................ 15
Desenho amostral ........................................................................................................ 16
Coleta de dados ........................................................................................................... 17
Análise dos dados ....................................................................................................... 18
Sazonalidade e sucessão ecológica ............................................................................. 19
RESULTADOS .............................................................................................................. 20
DISCUSSÃO .................................................................................................................. 27
Componentes da comunidade e índices parasitológicos ............................................. 27
Especificidade ............................................................................................................. 33
Sazonalidade e sucessão ecológica ............................................................................. 35
CONCLUSÃO ................................................................................................................ 37
REFERÊNCIAS ............................................................................................................. 38

RESUMO
Moscas da família Streblidae e Nycteribiidae, são ectoparasitas especializados
no hábito parasitário de morcegos, que podem ser influenciados por fatores ligados ao
ambiente de seus hospedeiros. Em Florestas Tropicais Secas (FTS) trabalhos sobre estas
moscas ainda não foram realizados. As FTS são ambientes altamente sazonais e
encontram-se distribuídas em forma de mosaicos por todo Brasil. Devido ao uso
intensivo da terra por atividades antrópicas, estes ambientes encontram-se em diferentes
estágios de sucessão ecológica. Neste trabalho foram estudados aspectos parasitológicos
(composição da comunidade, prevalência, intensidade média de infestação e
especificidade) na interação moscas-morcegos em três áreas de FTS brasileiras.
Ademais, testamos a hipótese de que a sazonalidade e sucessão ecológica influenciam
no parasitismo de Artibeus planirostris, espécie mais abundante no estudo, nestes
ambientes. As associações aqui encontradas são comuns para o Brasil, entretanto alguns
são novos registros para a região. A maioria das moscas foram consideradas altamente
específicas, reforçando que tais ectoparasitas possuem baixa capacidade de sobreviver
em hospedeiros que não são habituais. Portanto, as barreiras de dispersão exercem forte
pressão nesta interação hospedeiro-parasita. Quanto a sazonalidade e sucessão
ecológica, não encontramos resultado significativo no parasitismo de A. planirostris.
Sendo que, aspectos como o abrigo e plasticidade alimentar do hospedeiro, e
especificidade do parasita, podem ser responsáveis por tal resultado.
Palavras-chave: Streblidae, Nycteribiidae, interação hospedeiro-parasita, Artibeus
planirostris.

ABSTRACT
Flies from the family Streblidae and Nycteribiidae are specialized ectoparasites in the
parasitic habit of bats, which can be influenced by factors related to their host
environment. In Tropical Dry Forest (TDF) studies of these flies have not been realized.
The TDF are highly seasonal environments and are distributed in the form of mosaic
across Brazil. Due to the intensive use of the land by human activities, these
environments are at different stages of ecological succession. In this work
parasitological aspects (community composition, prevalence, mean intensity of
infestation and specificity) in the interaction bat flies-bats in three areas of Brazilian
TDF were studied. Furthermore, we tested the hypothesis that seasonality and
ecological succession influence the parasitism of Artibeus planirostris, most abundant
species in the study, in these environments. The associations found here are common to
Brazil, though some are new records for the region. Most flies were considered highly
specific, reinforcing that these ectoparasites have low ability to survive on hosts that are
not usual. Therefore, dispersal barriers have a strong pressure in this host-parasite
interaction. As for seasonality and ecological succession, we found no significant effect
on parasitism of A. planirostris. Being that, aspects like shelter and feeding plasticity of
host, and specificity parasite, may be responsible for this result.
Keywords: Streblidae, Nycteribiidae, host-parasite interaction, Artibeus planirostris.

INTRODUÇÃO
Moscas de morcego (Diptera: Hippoboscoidea) são ectoparasitos especializados
em morcegos (Mammalia: Chiroptera), apresentando hábito hematofágico (sugando
sangue na pele ou membranas das asas do hospedeiro) (Dick & Patterson 2006). Essas
moscas são divididas em duas famílias, Streblidae e Nycteribiidae, distribuídas
mundialmente (Dittmar et al. 2006). O tipo de reprodução desses parasitos é por
viviparidade adenotrófica. Os ovos ficam retidos na mãe e ao eclodirem as larvas se
alimentam de secreções de glândulas hipertrofiadas, sendo depositadas no abrigo, como
larvas de terceiro instar prestes a empupar. Desta maneira, a chance de sobrevivência da
pupa é maior, visto que ela fica protegida do hospedeiro (Kim 1985, Dick & Patterson
2006). A mosca adulta eclode do pupário de três a quatro semanas, após esse período o
adulto emerge e coloniza o hospedeiro imediatamente para sua alimentação (Ching &
Marshall 1968). As famílias Streblidae e Nycteribiidae são altamente especializadas no
hábito de vida parasitário (Allen 1967, Dick & Patterson 2007). Por isso, a morfologia
de ambas está intimamente relacionada aos seus hospedeiros. Deste modo, vários
fatores podem influenciar na abundância e distribuição dos ectoparasitos de morcegos,
tais como sexo e reprodução do hospedeiro (Marshall 1982).
Esses dípteros possuem distribuição biogeográfica ampla, em especial nas
regiões tropicais, porém as espécies diferem entre os hemisférios leste e oeste (Dittmar
et al. 2006). Os fatores que influenciam o aumento ou diminuição da diversidade e,
dispersão desses ectoparasitas estão relacionados ao fato de seus hospedeiros
(morcegos) serem altamente diversos e com ampla distribuição geográfica (Wenzel
1976, Rui & Graciolli 2005). Além disso, a variação geográfica das espécies de
morcegos é um dos fatores determinantes da especificidade neste sistema hospedeiro-
parasito (Dick 2007).

Os nicteribiídeos (Nycteribiidae) compreendem 274 espécies de 11 gêneros,
divididos em três subfamílias (Nycteribiinae, Cyclopodiinae e Archinycteribiinae)
(Guimarães & D’Andretta 1956, Graciolli & Dick 2006). Apenas dois gêneros da
subfamília Nycteribiinae foram encontrados para as Américas, Basilia Miranda–
Ribeiro, 1903 e Hershkovitizia Guimarães & D’Andretta, 1956. No Brasil, o gênero
mais abundante é Basilia com 24 espécies registradas, e Hershkovitizia possui quatro
espécies descritas no país (Graciolli et al. 2007).
Já os estreblídeos (Streblidae) estão representados por 237 espécies, distribuídas
em 33 gêneros e cinco subfamílias (Brachytarsininae, Ascodipterinae, Nycterophiliinae,
Trichobiinae e Streblinae). Somente as subfamílias Nycterophiliinae, Trichobiinae e
Streblinae estão presentes nas Américas, as demais são restritas ao Velho Mundo
(Guerrero 1997, Dick & Graciolli 2006). E no Brasil, a família Streblidae é representada
por 70 espécies (Prevedello et al. 2005, Dias et al. 2009, Soares et al. 2013).
Estudos sobre Streblidae e Nicteribiidae têm aumentado no Brasil nos últimos
anos, mas ainda estão restritos a regiões de Cerrado e Mata Atlântica. Esses, por sua
vez, raramente lançam mão de ferramentas ecológicas (índices de prevalência e
intensidade média de infestação) e evolutivas (especificidade) para discutir a relação
morcego-mosca. Portanto, expandir estudos desta natureza para outras regiões e tipos de
formação florestal pode ajudar a entender se existem padrões quanto ao parasitismo de
Streblidae e Nycteribiidae em morcegos. Bem como, permite avaliar o quanto o
ambiente em que estão inseridos influenciam neste sistema de trabalho (hospedeiro-
parasita).
Em Florestas Tropicais Secas (FTS) esta relação morcego-mosca ainda não foi
estudada. Estas florestas são caracterizadas pela sazonalidade bem marcada, onde
durante a estação seca (de 3 a 5 meses) há alta deciduidade foliar e baixas temperaturas,

exercendo grande influencia sobre a produção vegetal (Sánchez-Azofeifa et al. 2005,
Pezzini, 2008). As FTS brasileiras encontram-se fragmentadas em todo o país, com
distribuição nos biomas da Caatinga e Cerrado (Espírito-Santo et al. 2009). Mesmo
sendo consideradas de grande importância para a conservação, tais áreas sofreram com
uso da terra por atividades agrárias e atualmente encontram-se em diferentes estágios de
sucessão (Quesada et al. 2009).
Com base no exposto, este trabalho objetiva caracterizar as associações
morcego-mosca em FTS, discutindo os componentes da comunidade destas moscas.
Para isto, avaliamos os índices de prevalência e de intensidade média de infestação bem
como a especificidade do sistema (hospedeiro-parasita). Em adição, este trabalho possui
como objetivo responder as seguintes perguntas: A sazonalidade e a sucessão ecológica
influenciam no parasitismo de morcegos por moscas ectoparasitas em FTS? Para
responder as perguntas propostas predizemos que: (i) haverá aumento no parasitismo na
estação seca, pois neste período a escassez de alimentos para os morcegos acarretará em
comprometimento na condição corporal do morcego favorecendo o parasitismo; (ii) o
estágio tardio de regeneração da área terá menor parasitismo que os demais, devido a
estas áreas serem mais preservadas haverá menor comprometimento da condição
corporal do hospedeiro.

MÉTODO
Área de estudo
O estudo foi conduzido em três ares de Florestas Tropicais Secas (FTS) no
estado de Minas Gerais (MG). As amostragens foram realizadas nas regiões norte e
central de MG entre os anos de 2008 e 2013.
Área I- Parque Estadual da Mata Seca (PEMS): é uma unidade de proteção
integral, sob responsabilidade do Instituto Estadual de Florestas (IEF). Possui área de
15.281 ha e está localizado no vale médio do rio São Francisco, norte do estado de
Minas Gerais, entre as coordenadas 14°48’36” – 14°56’59” S e 137 43°55’12” –
44°04’12” W. A vegetação original do parque é a FTS, associada a solos planos e
nutritivos (IEF 2000). O clima da região é Aw (Köppen) caracterizado por uma severa
estação seca durante o inverno. A temperatura e a precipitação média anual são de 24ºC
(Antunes 1994) e 871 mm, respectivamente (Madeira et al. 2009). A principal atividade
econômica na área antes da implementação do parque foi a pecuária. Aproximadamente
1.525 ha do parque são de pastagens em diferentes estágios de regeneração natural e
áreas remanescentes de floresta primária e secundária (Madeira et al. 2009).
Área II: realizamos as coletas em três unidades de conservação do norte de MG:
Parque Estadual Lagoa do Cajueiro (20.500 ha), nas Reservas Biológicas do Jaíba
(6.358 ha) e Serra Azul (7.200 ha). Estas áreas localizadas entre os municípios de
Matias Cardoso e Jaíba, foram criadas como medidas compensatórias à expansão do
Projeto Jaíba. A vegetação é caracterizada por FTS, encontrando-se na faixa de transição
do Cerrado, Caatinga e Mata Atlântica (IEF 2000). O clima que predomina nesta região
é o Aw (segundo classificação de Köppen) em que no inverno há uma estação seca bem
definida. Em nível anual o índice pluviométrico é de 871 mm e a temperatura média é

de 24ºC (Antunes 1994). Antes da criação destas unidades de conservação, estas áreas
eram destinadas principalmente à criação de gados, de maneira que, atualmente, é
composta por mosaicos florestais oriundos de diversos estágios florestais de
regeneração.
Área III- Serra do Cipó: localizada no município de Santana do Riacho, na
região central do estado de MG. A Serra do Cipó está inserida na porção sul da Cadeia
do Espinhaço, sendo que as áreas de FTS encontram-se associadas a afloramentos
calcários. A vegetação predominante na área é o Cerrado e Campo Rupestre. O clima da
região é mesotérmico (Cwb na classificação de Köppen), onde os invernos são secos e
os verões chuvosos. A precipitação média anual é de 1.500 mm e temperatura anual
média de 17,4 a 19,8ºC (Giulietti et al. 1997).
Desenho amostral
Para a diferenciação entre os estágios determinamos, previamente, a estrutura
vertical (número de estratos) e horizontal (densidade) das árvores nas áreas amostradas,
bem como o uso histórico da terra (idade de diferentes fragmentos) (veja Arroyo-Mora
et al. 2005, Madeira et al. 2010). Segue a definição de cada estágio de regeneração
(inicial, intermediário e tardio), com base na estratificação vertical. O inicial é
caracterizado por estrato herbáceo-arbustivo, com manchas esparsas de vegetação
lenhosa de porte mais alto formando um dossel descontínuo. O estágio intermediário
possui dois estratos; um composto de árvores decíduas de crescimento rápido que
formam um dossel fechado e, outro composto por lianas, árvores adultas pertencentes a
espécies tolerantes à sombra e árvores juvenis. O tardio caracteriza-se pela presença de
três estratos verticais; o primeiro constituído por árvores de grande porte que formam

um dossel bastante fechado, o segundo é formado por árvores juvenis de diferentes
idades e tamanhos, e o terceiro é composto por espécies herbáceas e arbustivas típicas
de sub-bosque.
Área I: para esta área, utilizamos os dados previamente coletados (2008) em um
estudo realizado pelo Laboratório de Ecologia e Controle Biológico (LECB) da
Universidade Estadual de Montes Claros, onde estão depositados os espécimes-
testemunho.
Para captura de morcegos, nas demais áreas, utilizamos 10 redes de neblina por
noite, dispostas ao nível do solo. Distribuímos as redes distribuídas arbitrariamente
dentro da mata, em possíveis rotas de voos, abrindo-as logo no período crepuscular.
Estas permaneceram assim dispostas durante cinco horas, sendo visitadas a cada 30
minutos.
Área II: Escolhemos três pontos de coleta em cada UC, totalizando nove pontos
de amostragem (três em cada estágio sucessional) (Figura 1). Realizamos seis coletas
neste local, no período de outubro de 2012 a agosto de 2013, totalizando 54 noites de
coleta. O esforço amostral total foi de 24.300m2.h (Straube & Bianconi 2002).
Área III: De maneira semelhante à Área II, escolhemos um total de nove pontos
amostrados (três em cada estágio sucessional) (Figura 2). O período de coleta foi entre
janeiro de 2012 e setembro de 2013, totalizando quatro campanhas. O esforço amostral
total foi de 8.910 m2.h (Straube & Bianconi 2002).
Coleta de dados
Os morcegos capturados foram acondicionados em sacos de algodão e
identificamos ainda no campo utilizando chaves taxonômicas (Vizotto & Taddei 1973,
Medellín et al. 1997, Gregorin & Taddei 2002, Reis et al. 2007). Utilizamos anilhas

metálicas para marcação dos animais e depois os indivíduos foram soltos. O uso de
anilha é importante para monitorar os morcegos.
Os ectoparasistos (Streblidae e Nicteribiidae) foram coletados por catação
manual com auxílio de uma pinça, durante inspeção do hospedeiro. Posteriormente,
acondicionamos em tubos eppendorf contendo álcool 70%. E levamos para
identificação no LECB, onde estão depositados os espécimes-testemunho. Utilizamos a
chave de Guerrero (1994a, 1994b, 1995, 1996 e 1997) para identificar os estreblídeos e,
Guimarães & D’Andretta e Guimarães (1966, 1977) para os nicteribiídeos.
Análise dos dados
Para cada associação hospedeiro-parasito calculamos a prevalência e a
intensidade média de infestação e, analisamos os componentes da comunidade (cada
espécie de parasito encontrada em uma única espécie de hospedeiro), seguindo Bush et.
al (1997). Prevalência (de hospedeiros infestados / número de hospedeiros examinados
x 100) e intensidade média de infestação (número de parasitos / número de hospedeiros
infestados) permitem inferir como uma determinada espécie de morcego está parasitada
por seus ectoparasitos. Com auxílio do software Quantitative Parasitology 3.0 (Rózsa et
al. 2000) obtivemos os intervalos de confiança para estes índices. Consideramos
associação como primária, quando a prevalência foi ≥ 5%. As demais associações foram
consideradas não-primárias, provavelmente por contaminação da coleta ou distúrbios de
transferência (Dick 2007).
Além disso, utilizamos um índice de especificidade (STD*) proposto por Poulin
& Moullit (2005), que engloba informações filogenéticas e ecológicas (prevalência),
calculando um único valor para cada espécie de parasito em uma assembleia de

hospedeiros. STD* mede as distâncias taxonômicas (ou seja, gênero, família, ordem,
classe e filo) de todas as espécies de hospedeiros utilizados por um dado parasito, dando
peso para cada nível taxonômico. Os valores dos níveis taxonômicos variam de um 1 a
5. Dessa forma, quanto maior o valor desta distância, maior o valor deste índice, assim,
STD* é inversamente proporcional à especificidade. Portanto, quando este índice possui
valor igual a 0, o parasito é considerado monoxênico, ou seja, utiliza uma única espécie
de hospedeiro. Se o valor for 1, o parasito é oligoxênico, parasitando indivíduos
congêneres. E consideramos o parasito como não-específico, quando o valor for maior
que 1,podendo chegar até 5 quando as espécies hospedeiras são de classes diferentes.
Utilizamos a classificação proposta por Reis et al.(2007) para inferirmos as distâncias
taxonômicas dos morcegos.
Sazonalidade e sucessão ecológica
Para testar as hipóteses propostas neste estudo (sazonalidade e sucessão
ecológica), construímos Modelos Lineares Generalizados (GLM’s), com distribuição do
tipo binomial negativa. Onde as variáveis explicativas foram estação (seca e chuvosa) e
estágio sucessional (inicial, intermediário e tardio), e a variável resposta abundância de
parasitos por indivíduo. Nesta análise, utilizamos apenas Artibeus planirostris, por ser a
espécie mais abundante. Além disso, as análises foram realizadas apenas para
indivíduos das áreas II e III, por possuírem o desenho amostral adequado para testar tais
hipóteses. Todas as análises foram realizadas no software R.
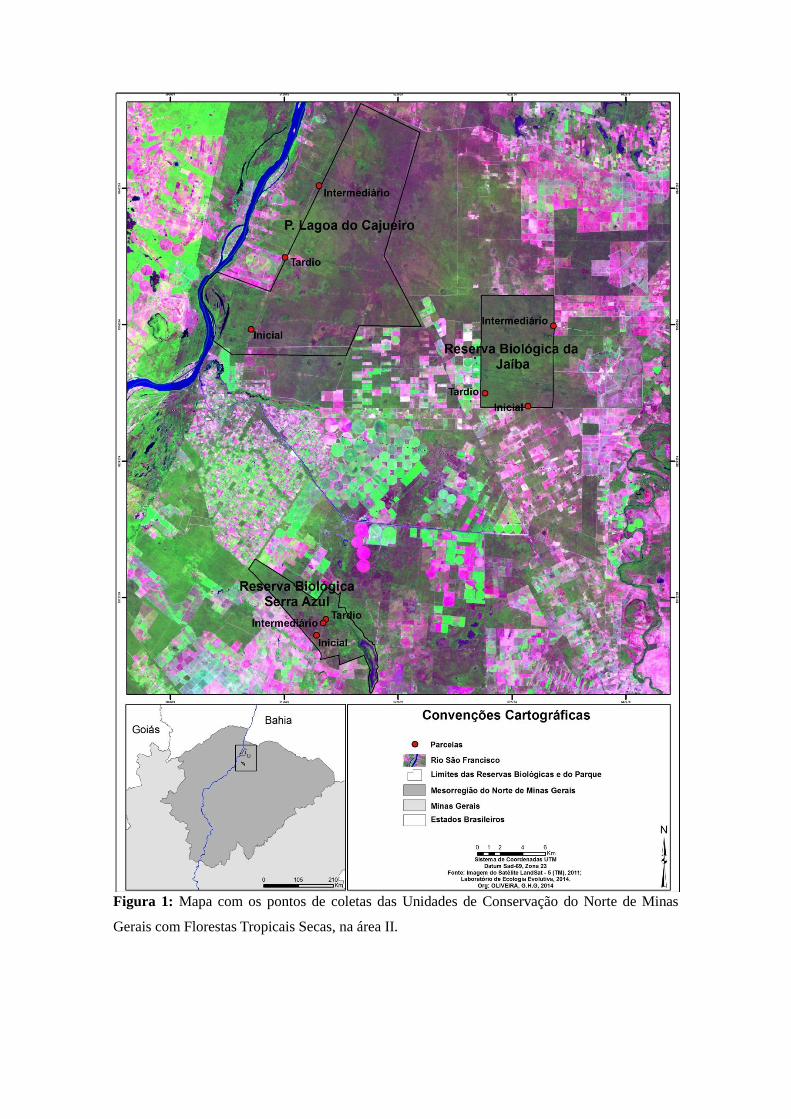
Figura 1: Mapa com os pontos de coletas das Unidades de Conservação do Norte de Minas
Gerais com Florestas Tropicais Secas, na área II.
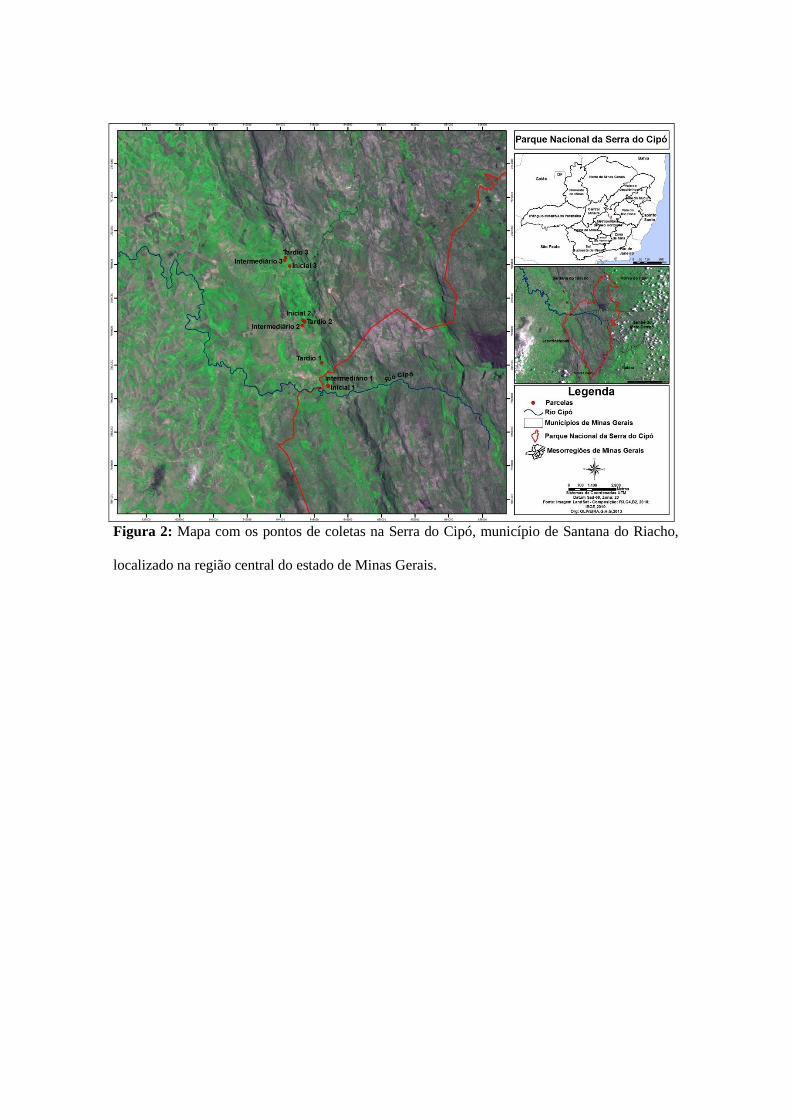
Figura 2: Mapa com os pontos de coletas na Serra do Cipó, município de Santana do Riacho,
localizado na região central do estado de Minas Gerais.

RESULTADOS
Amostramos 33 espécies de morcegos (n=882), pertencentes a cinco famílias
(Tabela). A família mais abundante foi Phyllostomidae (n=853), seguida por Molossidae
(n=14), Vespertilionidae (n=8), Emballuronidae (n=3) e Noctilionidae (n=1). Apenas as
espécies de duas destas famílias (Phyllostomidae e Vespertiolionidae) estavam
parasitadas por 38 espécies de dípteros das famílias Streblidae e Nycteribiidae (Tabela).
A maioria dos dípteros ectoparasitos encontrados foi estreblídeos (n=1.096),
compreendendo 36 espécies. Destes, Trichobius joblingi, Trichobius costalimai e
Megistopoda aranea, foram as mais abundantes com 298, 200 e 107 indivíduos
coletados, respectivamente. Somente duas espécies representaram a família
Nycteribiidae (n=2), Basilia juquiensis e Basilia mimoni.
A intensidade média de infestação variou de 1,0 a 10,7. Os morcegos
Phyloderma stenops (9,00) e Phyllostomus hastatus (7,41) foram os que apresentaram
maiores valores, quando parasitados por Trichobius longipes e Trichobius costalimai,
respectivamente. Para a prevalência a variação foi de 0 à 100%,com 74% de associações
primárias.
Para o cálculo de especificidade excluímos as associações não primárias. Dos
parasitos com associações primárias, cerca de73% obtiveram valor zero para o STD* e
foram considerados monoxênicos. Ou seja, estão associadas a um único hospedeiro.
Trichobius cf. handleyi foi a única espécie oligoxênica deste estudo, apresentando valor
um (01) para este índice. Esta, parasitou duas espécies congêneres, Micronycteris sp. e
Micronycteris minuta. Aproximadamente 24% tiveram valor superior a 1 para STD*,
indicando que estas espécies utilizam mais de uma espécie de morcegos como
hospedeiro.
As hipóteses propostas neste trabalho não foram corroboradas (p>0,05), em
nenhuma das áreas. Portanto, a estação e o estágio sucessional não são fatores
determinantes do parasitismo em Artibeus planirostris na FTS.
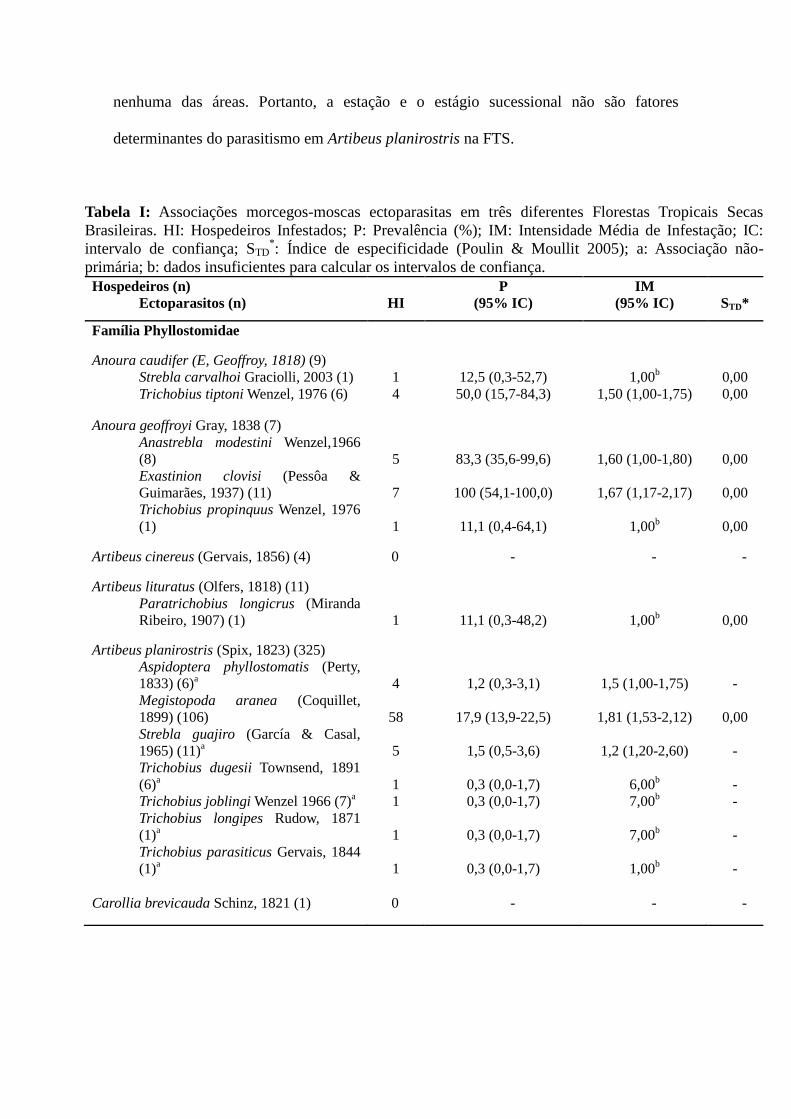
Tabela I: Associações morcegos-moscas ectoparasitas em três diferentes Florestas Tropicais Secas
Brasileiras. HI: Hospedeiros Infestados; P: Prevalência (%); IM: Intensidade Média de Infestação; IC:
intervalo de confiança; STD*: Índice de especificidade (Poulin & Moullit 2005); a: Associação não-
primária; b: dados insuficientes para calcular os intervalos de confiança.
Hospedeiros (n)
Ectoparasitos (n)
HI
P
(95% IC)
IM
(95% IC)
STD*
Família Phyllostomidae
Anoura caudifer (E, Geoffroy, 1818) (9)
Strebla carvalhoi Graciolli, 2003 (1)
Trichobius tiptoni Wenzel, 1976 (6)
1
4
12,5 (0,3-52,7)
50,0 (15,7-84,3)
1,00b
1,50 (1,00-1,75)
0,00
0,00
Anoura geoffroyi Gray, 1838 (7)
Anastrebla modestini Wenzel,1966
(8)
Exastinion clovisi (Pessôa &
Guimarães, 1937) (11)
Trichobius propinquus Wenzel, 1976
(1)
5
7
1
83,3 (35,6-99,6)
100 (54,1-100,0)
11,1 (0,4-64,1)
1,60 (1,00-1,80)
1,67 (1,17-2,17)
1,00b
0,00
0,00
0,00
Artibeus cinereus (Gervais, 1856) (4) 0 - - -
Artibeus lituratus (Olfers, 1818) (11)
Paratrichobius longicrus (Miranda
Ribeiro, 1907) (1)
1
11,1 (0,3-48,2)
1,00b
0,00
Artibeus planirostris (Spix, 1823) (325)
Aspidoptera phyllostomatis (Perty,
1833) (6)a
Megistopoda aranea (Coquillet,
1899) (106)
Strebla guajiro (García & Casal,
1965) (11)a
Trichobius dugesii Townsend, 1891
(6)a
Trichobius joblingi Wenzel 1966 (7)a
Trichobius longipes Rudow, 1871
(1)a
Trichobius parasiticus Gervais, 1844
(1)a
4
58
5
1
1
1
1
1,2 (0,3-3,1)
17,9 (13,9-22,5)
1,5 (0,5-3,6)
0,3 (0,0-1,7)
0,3 (0,0-1,7)
0,3 (0,0-1,7)
0,3 (0,0-1,7)
1,5 (1,00-1,75)
1,81 (1,53-2,12)
1,2 (1,20-2,60)
6,00b
7,00b
7,00b
1,00b
-
0,00
-
-
-
-
-
Carollia brevicauda Schinz, 1821 (1) 0 - - -
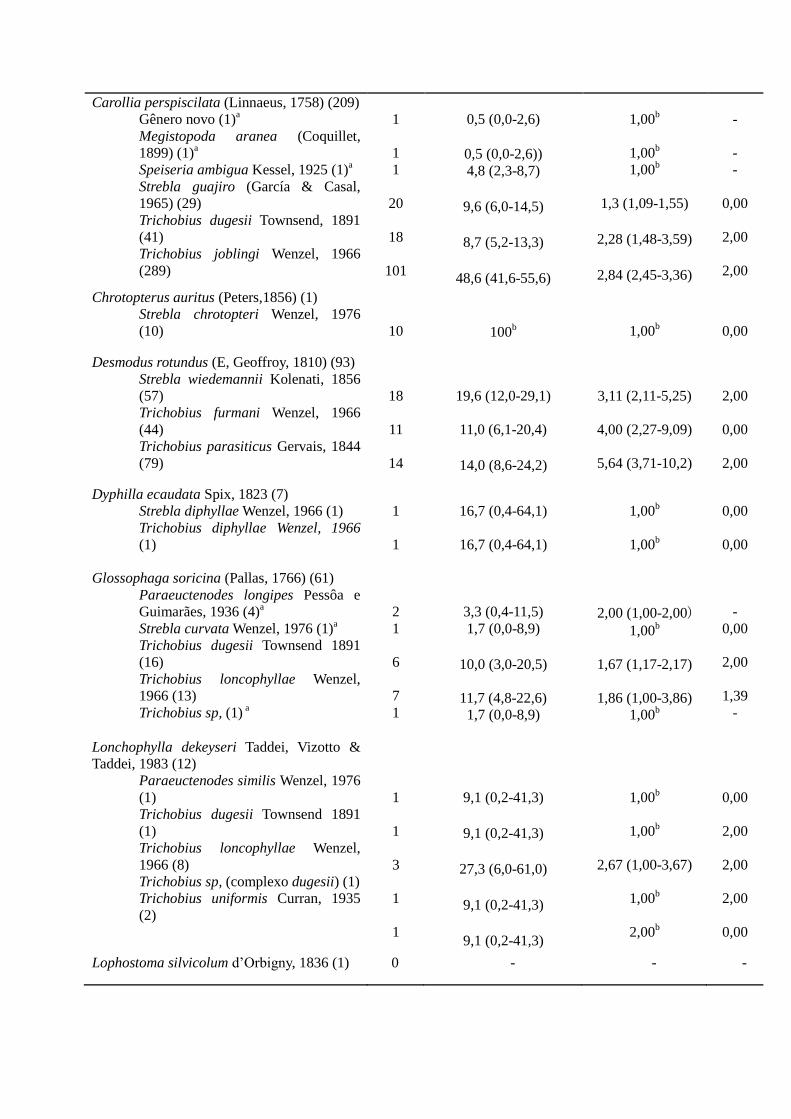
Carollia perspiscilata (Linnaeus, 1758) (209)
Gênero novo (1)a
Megistopoda aranea (Coquillet,
1899) (1)a
Speiseria ambigua Kessel, 1925 (1)a
Strebla guajiro (García & Casal,
1965) (29)
Trichobius dugesii Townsend, 1891
(41)
Trichobius joblingi Wenzel, 1966
(289)
1
1
1
20
18
101
0,5 (0,0-2,6)
0,5 (0,0-2,6)) 4,8 (2,3-8,7)
9,6 (6,0-14,5)
8,7 (5,2-13,3)
48,6 (41,6-55,6)
1,00b
1,00b
1,00b
1,3 (1,09-1,55)
2,28 (1,48-3,59)
2,84 (2,45-3,36)
-
-
-
0,00
2,00
2,00
Chrotopterus auritus (Peters,1856) (1)
Strebla chrotopteri Wenzel, 1976
(10)
10
100b
1,00b
0,00
Desmodus rotundus (E, Geoffroy, 1810) (93)
Strebla wiedemannii Kolenati, 1856
(57)
Trichobius furmani Wenzel, 1966
(44)
Trichobius parasiticus Gervais, 1844
(79)
18
11
14
19,6 (12,0-29,1)
11,0 (6,1-20,4)
14,0 (8,6-24,2)
3,11 (2,11-5,25)
4,00 (2,27-9,09)
5,64 (3,71-10,2)
2,00
0,00
2,00
Dyphilla ecaudata Spix, 1823 (7)
Strebla diphyllae Wenzel, 1966 (1)
Trichobius diphyllae Wenzel, 1966
(1)
1
1
16,7 (0,4-64,1)
16,7 (0,4-64,1)
1,00b
1,00b
0,00
0,00
Glossophaga soricina (Pallas, 1766) (61)
Paraeuctenodes longipes Pessôa e
Guimarães, 1936 (4)a
Strebla curvata Wenzel, 1976 (1)a
Trichobius dugesii Townsend 1891
(16)
Trichobius loncophyllae Wenzel,
1966 (13)
Trichobius sp, (1) a
2
1
6
7
1
3,3 (0,4-11,5)
1,7 (0,0-8,9)
10,0 (3,0-20,5)
11,7 (4,8-22,6) 1,7 (0,0-8,9)
2,00 (1,00-2,00) 1,00
b
1,67 (1,17-2,17)
1,86 (1,00-3,86) 1,00
b
-
0,00
2,00
1,39
-
Lonchophylla dekeyseri Taddei, Vizotto &
Taddei, 1983 (12)
Paraeuctenodes similis Wenzel, 1976
(1)
Trichobius dugesii Townsend 1891
(1)
Trichobius loncophyllae Wenzel,
1966 (8)
Trichobius sp, (complexo dugesii) (1)
Trichobius uniformis Curran, 1935
(2)
1
1
3
1
1
9,1 (0,2-41,3)
9,1 (0,2-41,3)
27,3 (6,0-61,0)
9,1 (0,2-41,3)
9,1 (0,2-41,3)
1,00b
1,00b
2,67 (1,00-3,67)
1,00b
2,00b
0,00
2,00
2,00
2,00
0,00
Lophostoma silvicolum d’Orbigny, 1836 (1) 0 - - -
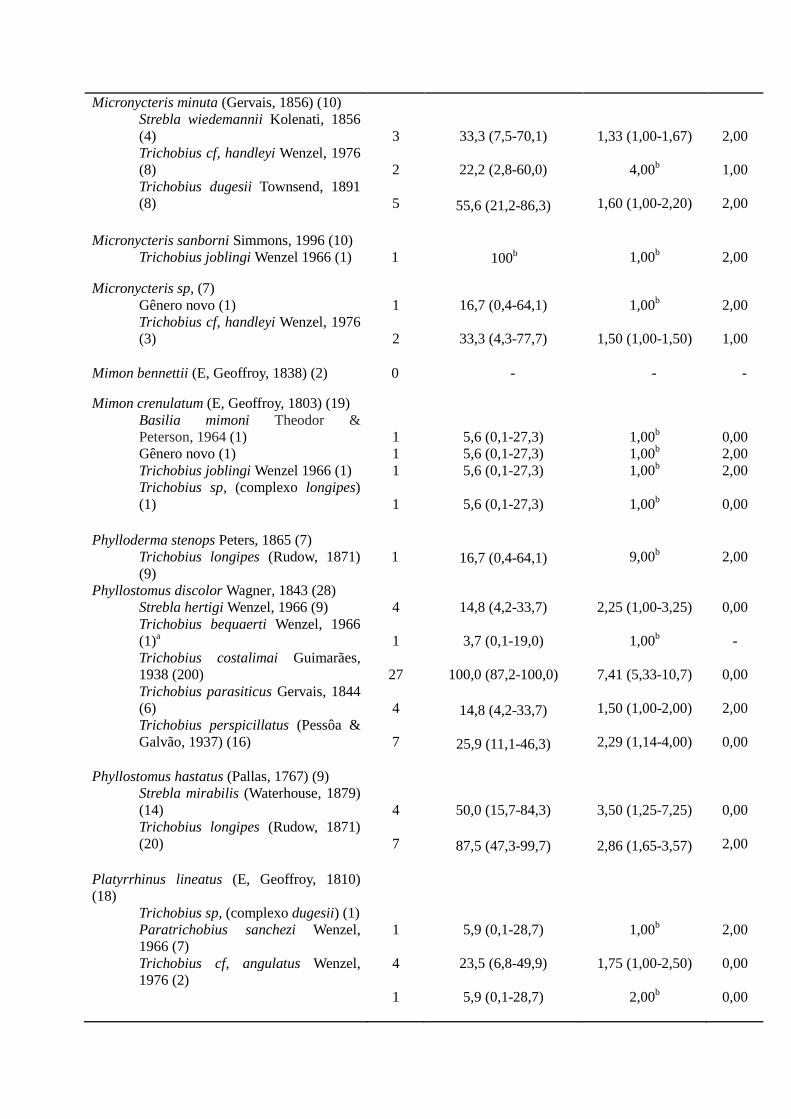
Micronycteris minuta (Gervais, 1856) (10)
Strebla wiedemannii Kolenati, 1856
(4)
Trichobius cf, handleyi Wenzel, 1976
(8)
Trichobius dugesii Townsend, 1891
(8)
3
2
5
33,3 (7,5-70,1)
22,2 (2,8-60,0)
55,6 (21,2-86,3)
1,33 (1,00-1,67)
4,00b
1,60 (1,00-2,20)
2,00
1,00
2,00
Micronycteris sanborni Simmons, 1996 (10)
Trichobius joblingi Wenzel 1966 (1)
1
100b
1,00b
2,00
Micronycteris sp, (7)
Gênero novo (1)
Trichobius cf, handleyi Wenzel, 1976
(3)
1
2
16,7 (0,4-64,1)
33,3 (4,3-77,7)
1,00b
1,50 (1,00-1,50)
2,00
1,00
Mimon bennettii (E, Geoffroy, 1838) (2) 0 - - -
Mimon crenulatum (E, Geoffroy, 1803) (19)
Basilia mimoni Theodor &
Peterson, 1964 (1)
Gênero novo (1)
Trichobius joblingi Wenzel 1966 (1)
Trichobius sp, (complexo longipes)
(1)
1
1
1
1
5,6 (0,1-27,3) 5,6 (0,1-27,3) 5,6 (0,1-27,3)
5,6 (0,1-27,3)
1,00b
1,00b
1,00b
1,00b
0,00
2,00
2,00
0,00
Phylloderma stenops Peters, 1865 (7)
Trichobius longipes (Rudow, 1871)
(9)
1
16,7 (0,4-64,1)
9,00b
2,00
Phyllostomus discolor Wagner, 1843 (28)
Strebla hertigi Wenzel, 1966 (9)
Trichobius bequaerti Wenzel, 1966
(1)a
Trichobius costalimai Guimarães,
1938 (200)
Trichobius parasiticus Gervais, 1844
(6)
Trichobius perspicillatus (Pessôa &
Galvão, 1937) (16)
4
1
27
4
7
14,8 (4,2-33,7)
3,7 (0,1-19,0)
100,0 (87,2-100,0)
14,8 (4,2-33,7)
25,9 (11,1-46,3)
2,25 (1,00-3,25)
1,00b
7,41 (5,33-10,7)
1,50 (1,00-2,00)
2,29 (1,14-4,00)
0,00
-
0,00
2,00
0,00
Phyllostomus hastatus (Pallas, 1767) (9)
Strebla mirabilis (Waterhouse, 1879)
(14)
Trichobius longipes (Rudow, 1871)
(20)
4
7
50,0 (15,7-84,3)
87,5 (47,3-99,7)
3,50 (1,25-7,25)
2,86 (1,65-3,57)
0,00
2,00
Platyrrhinus lineatus (E, Geoffroy, 1810)
(18)
Trichobius sp, (complexo dugesii) (1)
Paratrichobius sanchezi Wenzel,
1966 (7)
Trichobius cf, angulatus Wenzel,
1976 (2)
1
4
1
5,9 (0,1-28,7)
23,5 (6,8-49,9)
5,9 (0,1-28,7)
1,00b
1,75 (1,00-2,50)
2,00b
2,00
0,00
0,00

Sturnira lilium (E, Geoffroy, 1810) (1) 0 - - -
Tonatia saurophila Koopman & Wiliiams,
1951 (2)
0 - - -
Uroderma magnirostrum Davis, 1968 (1) 0 - - -
Família Vespertilionidae
Lasiurus blossevillii (Lesson & Garnot,
1826) (1)
0 - - -
Histiotus velatus (I, Geoffroy, 1824) (1) 0 - - -
Myotis sp, (3) 0 - - -
Myotis nigricans (Schinz, 1821) (2)
Basilia juquiensis Guimarães,
1943 (1)
1
100,0 (2,5-100,0)
1,00b
0,00
Família Molossidae
Molossops teminckii (Burmeister, 1854) (13) 0 - - -
Molossus molossus (Pallas, 1766) (1) 0 - - -
Família Emballuronidae
Peropteryx macrotis (Wagner, 1843) (3) 0 - - -
Família Noctiolinidae
Noctilio albiventris Desmarest, 1818 (1) 0 - - -
Total (882)

DISCUSSÃO
Componentes da comunidade e índices parasitológicos
A espécie Artibeus planirostris estava parasitada por Megistopoda aranea
(n=106). Esta espécie normalmente parasita morcegos do gênero Artibeus (Guerrero
1994). A associação entre M. aranea e A. planirostris já foi registrada para o estado de
Minas Gerais (MG), em área de Cerrado (Komeno & Linhares 1999). Bem como, para
regiões de Cerrado em Goiás (GO), no Mato Grosso do Sul (MS) (Graciolli et al. 2010,
Erickson et al. 2011) e Maranhão (MA) (Santos et al. 2013). A prevalência de M.
aranea (17,9%) foi menor quando comparada a trabalhos desenvolvidos no Cerrado
(Erickson et al. 2011, Santos et al. 2013), já a intensidade de infestação (1,81) foi
semelhante.
Carollia perspiscilata encontrou-se parasitada por Trichobius joblingi (n=289),
Trichobius dugesii (n=41) e Strebla guajiro (n=29). As espécies T. joblingi e S. guajiro,
são associações comuns em outros estudos no Cerrado em MG (Komeno & Linhares
1999, Azevedo & Linardi 2002, Teixeira & Ferreira 2010), MS (Erickson et al. 2011) e
MA (Santos et al. 2013). E na Mata Atlântica em São Paulo (SP) (Graciolli et al.
2006b), Paraná (PR) (Prevedello et al. 2005) e MG (Moras et al. 2013). E T. joblingi
também foi registrada no MS em uma área de transição Cerrado-Floresta Estacional
(Graciolli et al. 2006a). A prevalência e a intensidade de infestação para T. joblingi e S.
guajiro são semelhantes aos estudos realizados no Cerrado por Erickson et al. (2011) e
Santos et al. (2013). Porém, quando comparadas ao estudo de Teixeira & Ferreira
(2010), são diferentes. Para T. joblingi ambos os índices parasitológicos foram maiores.
Em S. guajiro a prevalência foi menor e a intensidade média de infestação maior. T.
dugesii foi encontrada pela primeira vez em C. perspiscilata no Brasil, possuindo

registros para outras regiões da América do Sul (Guerrero 1995). Speiseria ambigua é
comumente encontrado neste hospedeiro (Dias et al. 2009, Teixeira & Ferreira 2010,
Erickson et al. 2011, Soares et al. 2013, Santos et al. 2013), porém devido a baixa
amostragem desta espécie não foi considerada uma associação primária.
Trichobius parasiticus (n=79), Strebla wiedemannii (n=57) e Trichobius furmani
(n=44) estavam presentes em Desmodus rotundus. S. wiedemannii, diferente das outras
espécies de moscas, possui registro para MG (Komeno& Linhares 1999, Azevedo &
Linardi 2002, Teixeira & Ferreira 2010), e também é comumente encontrado sobre D.
rotundus em outras regiões do país (Graciolli & Coelho 2001, Graciolli & Linardi 2002,
Prevedello et al. 2005, Rios et al. 2008, Graciolli et al. 2010, Aguiar &Antonini 2011,
Erickson et al. 2011). Este ectoparasito possui taxa de prevalência (19,6%) menor em
FTS comparado a outros estudos (Rios et al. 2008, Aguiar & Antonini 2011, Erickson et
al. 2011). Sendo semelhante apenas ao estudo realizado por Teixeira & Ferreira (2010).
A intensidade média de infestação (3,11) foi semelhante a estes trabalhos, exceto para
região de Caatinga na Bahia (BA) (Rios et al. 2008) que foi maior. T. parasiticus
também foi amplamente amostrado sobre este hospedeiro em outros estudos (Graciolli
& Linardi 2002, Rios et al. 2008, Dias et al. 2009, Graciolli et al. 2010. Aguiar &
Antonini 2011, Soares et al. 2013, Santos et al. 2013). Entretanto, sua prevalência
(14%) foi menor e intensidade de infestação (5,64) maior, quando comparadas a estudos
na BA (Rios et al. 2008) e DF (Aguiar & Antonini 2011). Ao compararmos com o
trabalho feito no MA (Santos et al. 2013), a prevalência e intensidade de infestação
foram menores. T. furmani substitui T. parasiticusem D. rotundus na região sudeste do
Brasil (Graciolli & Linardi 2002, Bertola et al. 2005). Entretanto, como mostramos
aqui, em FTS de MG ambas as espécies ocorrem. Também possuem registros de T.
furmani para o DF (Aguiar & Antonini 2001, Graciolli & Coelho 2001). A prevalência

(11%) foi semelhante à encontrada por Aguiar & Antonini (2011), mas a intensidade
média de infestação (4,00) foi maior.
Trichobius dugesii (n=16) e Trichobius loncophyllae (n=13) parasitaram
Glossophaga soricina. A mosca T. loncophyllae já foi registrada nesta associação para
uma região de Cerrado em MG (Azevedo & Linardi 2002). T. dugesii não foi
encontrada no estudo anteriormente citado, sendo, portanto, este o seu primeiro registro
para o estado. Porém, esta mosca é encontrada em várias regiões de Cerrado no DF, GO,
MS e MA (Graciolli & Coelho 2001, Graciolli et al. 2010, Erickson et al. 2011,Santos
et al. 2013, respectivamente). Além disso, no Rio Grande do Sul (RS) (Graciolli & Rui
2001) em áreas de fragmentos de Mata Atlântica e no MA (Dias et al. 2009) em
fragmentos de floresta amazônica. Quanto às taxas de prevalência e intensidade de
infestação de T. dugesii sobre G. soricina, estas se mostraram semelhantes quando
comparadas a outros trabalhos (Erickson et al. 2011, Santos et al. 2013).
Phyllostomus discolor estava parasitado por Trichobius costalimai (n=200),
Trichobius perspiscillatus (n=16), Strebla hertigi (n=9) e Trichobius parasiticus (n=6).
Dentre estas associações, T. parasiticus ainda não foi descrita, sendo que este é um
parasito de Phyllostomus hastatus (Guerrero 1995). As demais foram encontradas em
área de Mata Atlântica no MA (Dias et al. 2009) e, apenas T. costalimai e T.
perspiscillatus em Pernambuco (PE) (Soares et al. 2013). Ademais, T. costalimai possui
registro para o GO em área de Cerrado (Graciolli et al. 2010). S. hertigi e T. costalimai
foram descritas para o Cerrado no MA (Santos et al. 2013). Os índices parasitológicos
de T. costalimai quando comparados a este estudo do MA, são maiores em FTS (100% e
7.41). Para S. hertigi a prevalência (14.8%) é semelhante e a intensidade média (2.25)
maior. Todas estas espécies são novos registros para o sudeste do Brasil.
Platyrrhinus lineatus foi parasitado por Paratrichobius sanchezi (n=7),

Trichobius cf. angulatus (n=2) e Trichobius sp. (complexo dugesii) (n=1). A presença
destas moscas em P. lineatus é um novo registro para o sudeste brasileiro. Além disso, P.
sanchezi é uma nova associação com P. lineatus, parasitando geralmente indivíduos do
gênero Artibeus (Guerrero 1994). Trichobius angulatus já foi registrada por Erickson et
al. (2011), em uma área de Cerrado no MS. A prevalência deste parasito (5,9%) foi
menor quando comparada ao estudo anteriormente citado. Já a intensidade média de
infestação (2,00) foi semelhante, mesmo não sendo possível calcular o intervalo de
confiança. A identificação de Trichobius sp. (complexo dugesii) foi difícil devido ao
material coletado estar danificado e a semelhança das espécies presentes neste
complexo.
Basilia mimoni (n=1), T. joblingi (n=1) e Trichobius sp. (complexo longipes)
(n=1) parasitaram Mimon crenulatum. A associação entre o nicteribiídeo B. mimoni e M.
crenulatum foi encontrada no MA em uma região de Mata Atlântica (Dias et al. 2013) e
uma de Cerrado (Santos et al. 2013), e é a única comum.Este é o primeiro registro
destas espécies no sudeste do país. Os índices parasitológicos são menores do que os
apresentados por Santos et al. (2013).
Artibeus lituratus foi parasitada apenas por Paratrichobius longicrus (n=1).
Normalmente, este parasito costuma se associar a morcegos do gênero Artibeus e
Plathyrrhinus (Wenzel et al. 1966). Esta associação entre P. longicrus e A. lituratus é
comum, com registros para várias regiões do Brasil. No Cerrado foi encontrada em MG
(Azevedo & Linardi 2002) e no DF (Graciolli & Aguiar 2002). Na Mata Atlântica
possui registros para o RS (Graciolli & Rui 2001, Rui & Graciolli 2005, Camilotti et al.
2010), SP (Bertola et al. 2005, Graciolli et al. 2006b), PR (Prevedello et al. 2005,
Anderson & Ortêncio-Filho 2006), PE (Soares et al. 2013) e em MG (Moras et al.
2013). E em áreas de Florestas com Araucárias no PR (Graciolli & Bianconi 2007). A

prevalência de P. longicrus (11,1%) foi menor que outros estudos (Graciolli & Rui
2001, Rui & Graciolli 2005, Anderson & Ortêncio-Filho 2006, Graciolli & Bianconi
2007), porém maior que encontrado por Bertola et al. (2005) (1,5%) e Camilotti et al.
2006 (2,6%), em áreas de Mata Atlântica. Entretanto a intensidade média de infestação
foi semelhante quando comparados a outros em Mata Atlântica (Bertola et al. 2005, Rui
& Graciolli 2005, Camilotti et al. 2006).
T. loncophyllae (n=8), Trichobius uniformis (n=2), T. dugesii (n=1),
Paraeuctenodes similis (n=1) e Trichobius sp. (complexo dugesii) (n=1) estavam
associados à Loncophylla dekeyseri. Sendo que, todos estes registros são novos para a
região sudeste do Brasil. T. loncophyllae sobre L. dekeyseri já foi registrada no DF em
uma região de Cerrado (Graciolli & Coelho 2001). As demais associações não são
comuns a L. dekeyseri. T. uniformis foi encontrada em Loncophylla robusta, mas é
comum em G. soricina, bem como T. dugesii (Guerrero 1994, 1995). Quanto a espécie
P. similis, normalmente esta encontra-se associada à C. perspiscillata, possuindo
registros para o Brasil (Guerrero 1996). Indivíduos dos gêneros Glossophaga e
Loncophyllae pertencem a mesma grupo funcional (nectarívoro) e, portanto, possuem
características em comum, o que pode determinar seu parasitismo por estas espécies de
ectoparasitos.
A espécie Micronycteris minuta estava parasitada por T. dugesii (n=8),
Trichobius cf. handleyi (n=8) e S. wiedemannii (n=4). E Micronycteris sp. estava
associado à Trichobius cf. handleyi (n=3). Segundo Wenzel (1976), este díptero está
relacionado á morcegos do gênero Micronycteris. Além disso, T. handleyii já foi descrita
em M. minuta (Guerrero 1995) para o Novo Mundo, mas pela primeira vez no Brasil.
As outras associações não são comuns. Porém, T. dugesii é comum em indivíduos do
gênero Micronycteris.

T. joblingi (n=1) foi a única espécie de mosca presente em Micronycteris
sanborni. Esta associação é comum em outras espécies do gênero Micronycteris
(Guerrero 1995). Entretanto, este é o primeiro registro de um estreblídeo em M.
sanborni.
O morcego Anoura caudifer estava parasitado por Trichobius tiptoni (n=6) e
Strebla carvalhoi (n=1). A mosca T. tiptoni já foi encontrada em diversas localidades do
Brasil. Na Mata Atlântica no RS (Rui & Graciolli 2001, Graciolli & Rui 2005), SP
(Bertola et al. 2005) e MG (Teixeira & Ferreira 2010). E no Cerrado em MG (Komeno
& Linhares 1999, Moras et al. 2013), GO (Graciolli et al. 2010) e no MS (Erickson et
al. 2011). Em outros estudos (Teixeira & Ferreira 2010, Erickson et al. 2011), a
prevalência para T. tiptoni foi menor do que a encontrada aqui (50%). Entretanto, a
intensidade média de infestação (1,5) foi marginalmente semelhante, oscilando dentro
do intervalo de confiança. Strebla carvalhoi foi descrita por Graciolli (2003) no PR, em
associação a A. caudifer. E, este é o primeiro registro desta associação para o sudeste do
país.
P. hastatus foi parasitada por Trichobius longipes (n=20) e Strebla mirabilis
(n=14). Trichobius longipes já foi encontrada sobre P. hastatus em outro estudo no
estado de MG (Azevedo & Linardi 2002), bem como no estado do MA (Dias et al.
2013, Santos et al.2013). A prevalência (87,5%) foi maior do que a encontrada por
Santos et al. (2013) no MA. Mas, a intensidade de infestação (2,86) foi parecida. S.
mirabilis não foi encontrada em associação com P. hastatus no sudeste do país, apenas
no DF (Graciolli & Coelho 2001).
As moscas Exastinion clovisi (n=11), Anastrebla modestini (n=8) e Trichobius
propinquus (n=1) estavam parasitando Anoura geoffroyi. As espécies E. clovisi e A.
modestini foram registradas em regiões de Cerrado (Graciolli & Coelho 2001, Graciolli

et al. 2010) e Mata Atlântica (Graciolli & Rui 2001, Bertola et al. 2005). Ambas
espécies possuem registros para MG em Mata Atlântica (Moras et al. 2013), sendo que
E. clovisi, foi registrada também em estudo no Cerrado (Komeno & Linhares 1999). Já
T. propinquus foi encontrada apenas em uma área de Cerrado do DF (Graciolli &
Coelho 2001). Esta mosca é um novo registro para a região sudeste do Brasil.
O hospedeiro Dyphilla ecaudata estava parasitado por Strebla diphyllae (n=1) e
Trichobius diphyllae (n=1). Tipicamente, estas moscas são espécies-tipo deste morcego,
(Guerrero 1995, 1996). Entretanto, são consideradas raras, pela baixa amostragem.
T. longipes (n=9) foi a única mosca presente em Phylloderma stenops neste
estudo. Esta mosca geralmente é parasita de P. hastatus (Graciolli & Carvalho 2001).
Este é o primeiro registro desta associação. Porém, estreblídeos do gênero Trichobius
pertencentes ao complexo longipes, são comuns para grandes Phyllostominae (Guerrero
1995), o que pode explicar esta associação.
Myotis nigricans foi parasitada apenas pelo nicteribiídeo Basilia juquiensis
(n=2). Esta é uma associação comum, possuindo registro para região sul do Brasil
(Graciolli 2004). Além disso, recentemente foi encontrada no estado de MG em uma
área de Mata Atlântica (Moras et al. 2013).
A mosca Strebla chrotopteri (n=10) estava associada a Chrotopterus auritus.
Esta associação é comum, sendo registrada em área de Mata Atlântica em SP (Graciolli
et al. 2006b) e de Cerrado no MS (Erickson et al. 2011). A prevalência (100%) e
intensidade de média de infestação foram superiores as dos estudos citados. Porém,
apenas um indivíduo hospedeiro foi capturado e para comparação deveríamos conter
mais exemplares.
Especificidade

As moscas ectoparasitos de morcego, em sua grande maioria, são consideradas
monoxênicas, com especificidade variando entre 71 a 88% em alguns estudos (Wenzel
et al. 1966, Marshall 1981, 1982, Dick & Gettinger 2005, Dick 2007, Dick & Patterson
2007, Santos et al. 2013). Este padrão se repete neste estudo em FTS (73%). Deste
modo, estes dados indicam que neste tipo de ambiente, estes ectoparasitos, são
altamente específicos. De fato, este é um resultado esperado, devido a baixa capacidade
de uma mosca sobreviver em um morcego que não é o seu hospedeiro habitual (Fritz
1983).
Entretanto, o principal fator ligado ao elevado grau de especificidade é a
imunocompatibilidade entre parasitos e morcegos (Dick & Patterson 2007). Estas
moscas podem ser capazes de mimetizar sinais moleculares do seu hospedeiro, evitando
ação do sistema imunológico do morcego. Isto pode ter levado a eventos de co-
especiação neste sistema de estudo (mosca-morcego), devido a mudanças nas bases
filogenéticas e ecológicas imediatas (Dick & Patterson 2007).
Além disso, existem barreiras de dispersão, que impedem que um parasito
colonize um novo hospedeiro. Tais barreiras podem ser classificadas como filtros de
encontro, compatibilidade e reprodutivo, que atuam em conjunto impedindo a utilização
de hospedeiros não-primários (Dick & Patterson 2007). O filtro de encontro ocorre
quando um parasito não consegue encontrar e colonizar um hospedeiro por razões
comportamentais ou ecológicas (Combes 1991 apud Dick & Patterson 2007). Já o filtro
de compatibilidade acontece quando um parasito não consegue sobreviver em um novo
hospedeiro por razões morfológicas, fisiológicas ou imunológicas (Combes 1991 apud
Dick & Patterson 2007). E o filtro reprodutivo atua quando há isolamento de um
parasito, por não encontrar algum indivíduo do sexo oposto de sua espécie, impedindo

uma chance de reprodução (Dick & Patterson 2007).
Mesmo com este padrão de especificidade dominante 27% das moscas
ectoparasitas de morcego utilizaram mais de uma espécie de hospedeiro, sejam elas
parasitos oligoxênicos ou polixênicos. Esta baixa especificidade ocorre devido a quebra
das barreiras de dispersão citadas anteriormente. Além disso, a mobilidade destas
moscas pode ser um fator determinante neste processo (Marshall 1981). Os
ectoparasitos menos específicos deste estudo, pertencem a família Streblidae (gêneros
Trichobius e Strebla) e possuem asas desenvolvidas e funcionais, facilitando na
dispersão para outros hospedeiros.
Os fatores ligados à história evolutiva neste sistema mosca-morcego precisam
ser elucidados para entender os mecanismos que levam a especificidade em parasitos
obrigatórios, em FTS. Fatores estes que podem estar ligados ao hospedeiro (tipo de
abrigo e compartilhamento de espécies no mesmo, disponibilidade de recurso, tamanho
corporal) e ao parasito (mobilidade, imunocompatibilidade com o hospedeiro).
Sazonalidade e sucessão ecológica
O parasitismo por moscas em A. planirostris, espécie mais abundante neste
estudo, não aumentou na estação seca, como era esperado, ou seja, a sazonalidade não
influencia tais hospedeiros em FTS. Este resultado pode ser explicado pela amplitude
alimentar utilizada por essa espécie de morcego, mesmo em períodos de escassez, já que
morcegos frugívoros podem se tornar generalistas nestas épocas (Flemming 1988).
Portanto, em períodos secos esses hospedeiros não ficam debilitados na falta de seu
recurso alimentar principal, não havendo aumento nas cargas parasitárias. É importante
estender esta análise para indivíduos de diferentes guildas alimentares, para verificar se
a sazonalidade pode influenciar outros grupos de hospedeiros.

O mesmo resultado, não significativo, foi encontrado para o parasitismo em A.
planirostris no estágio tardio, não havendo influencia da sucessão ecológica na
associação entre mosca e morcego. Esperávamos que com uma maior complexidade
ambiental, os hospedeiros teriam mais acesso ao seu alimento e estariam mais aptos a
escaparem do parasitismo, seja por mecanismos comportamentais (“grooming” e
mudança de abrigo) ou fisiológicos (sistema imunológico). Entretanto, o alto grau de
especialização permite que parasitos vivam em hospedeiros que estejam sob condições
de estresse, devido a evolução de mecanismos para co-existência (Combes 2001).
Assim, moscas ectoparasitos de morcego conseguem se alimentar de um hospedeiro
mesmo que este esteja sob condições de estresse alimentar. Ademais, morcegos
possuem grande capacidade de mobilidade e a distância dos fragmentos pode não ser
suficiente para amostrar indivíduos de diferentes populações. Talvez por isso, não foi
possível verificar algum padrão de parasitismo em função do estágio sucessional.
Além disso, em ambos os casos (sazonalidade e sucessão ecológica) devemos
levar em conta o fato de que os indivíduos de A. planirostris podem pertencer as
mesmas populações e possuir fidelidade ao abrigo. Como o ciclo de vida das moscas
estão diretamente ligados ao local de abrigo dos hospedeiros (Patterson et al. 2007), a
fidelidade a estes locais favorece a infestação por estes ectoparasitos (Bordes et al.
2008). Sendo assim, os morcegos amostrados manteriam os mesmos níveis de
parasitismo. Entretanto, esta é uma hipótese a ser tratada em estudos posteriores.

CONCLUSÃO
Os resultados apresentados aqui são importantes para iniciar estudos sobre o
parasitismo de morcegos por moscas das famílias Streblidae e Nycteribiidae em FTS.
Entender os fatores que moldam esta relação são importantes e estão sendo explorados
em vários trabalhos. Entretanto, alguns aspectos ainda permanecem obscuros. Fatores
como, a imunocompatibilidade entre parasito e hospedeiro, precisam ser elucidados para
esclarecer se de fato houve co-especiação a tais níveis neste sistema. Estudos em longo
prazo também auxiliarão a esclarecer se aspectos temporais e espaciais, como os
testados aqui são relevantes na relação mosca ectoparasito e morcego. Além disso, é
necessário abranger estudos para diversas regiões e formações florestais, para comparar
e entender se o parasitismo é influenciado por características locais ou regionais.

REFERÊNCIAS
Allen G.M. 1967. The parasites of bats. Cambridge: Museum of Comparative Zoology,
p.368.
Aguiar L.M.S. & Antonini Y. 2011. Descriptive ecology of bat flies (Diptera:
Hippoboscoidea) associated with vampire bats (Chiroptera: Phyllostomidae) in the
cerrado of Central Brazil. Memórias do Instituto Oswaldo Cruz, 106: 170-176.
Anderson. R. & Ortêncio-Filho H. 2006. Dípteros ectoparasitas (Diptera, Streblidae) de
filostomídeos (Chiroptera, Mammalia) do Parque Municipal no Cinturão Verde de
Cianorte, Paraná, Brasil e sua incidência ao longo das estações do ano. Chiroptera
Neotropical, 12: 238-243.
Antunes F. Z.1994. Caracterização climática. Informe Agropecuário, 17: 15-19.
Azevedo A.A. & Linardi P.M. 2002. Streblidae (Diptera) of phyllostomid bats from
Minas Gerais, Brazil. Memórias do Instituto Oswaldo Cruz, 97: 421-422.
Bertola P.B., Aires C.C., Favorito S.E., Graciolli G., Amaku M. & Pinto-da-Rocha R.
2005. Bat flies (Diptera: Streblidae, Nycteribiidae) parasitic on bats (Mammalia:
Chiroptera) at Parque Estadual da Cantareira, São Paulo, Brazil: parasitism rates and
host-parasite associations. Memórias do Instituto Oswaldo Cruz, 100: 25-32.
Bordes F., Morand S. & Guerrero R. 2008. Bat fly species richness in Neotropical bats:
correlation with host ecology and host brain. Oecologia, 158: 109-116.

Bush A.O., Lafferty K.D., Lotz J.M., Shostak A.W. 1997. Parasitology meets ecology
on its own terms: Margolis et al. revisited. Journal of Parasitology, 83: 575-583.
Ching L.M. & Marshall, A.G. 1968. The breeding biology of the bat-fly Eucampispoda
sundaicum Theodor, 1955 (Diptera: Nycteribiidae). Malayan Nature Journal, 21: 171–
180.
Camilotti V.L., Graciolli G., Weber M.M., Arruda J.L.S. & Cáceres N.C. 2010. Bat flies
from the deciduous Atlantic Forest in southern Brazil: Host-parasite relationships and
parasitism rates. Acta Parasitologica, 55 (2): 194–200.
Combes C. 2001. Parasitism: the ecology and evolution of intimate interactions.
University of Chicago Press, Chicago.
Dias, P.A., Santos, C.L.C., Rodrigues, F.S., Rosa, L.C., Lobato, K.S. & Rebêlo, J.M.M.
2009. Espécies de moscas ectoparasitas (Diptera, Hippoboscoidea) de morcegos
(Mammalia, Chiroptera) no estado do Maranhão. Revista Brasileira de Entomologia, 53:
128-133.
Dick, C.W. & Gettinger, D. 2005. A faunal survey of streblid bat flies (Diptera:
Streblidae) associated with bats in Paraguay. Journal of Parasitology, 91: 1015-1024.
Dick C.W. & Patterson B.D. 2006. Bat flies: obligate ectoparasites of bats. Em S.
Morand, B.R. Krasnov, R. Poulin, Micromammals and macroparasites: from

evolutionary ecology to management, Springer-Verlag, p. 179-194.
Dick C.W. 2007. High host specificity of obligate ectoparasites. Ecological Entomology,
32: 446 – 450
Dick C.W. & Patterson B.D. 2007. Against all odds: Explaining high host specificity in
dispersal-prone parasites. International Journal for Parasitology, 37: 871-876.
Dittmar K., Porter M.L., Murray S., Whiting M.F. 2006. Molecular phylogenetic
analysis of nycteribiid and streblid bat flies (Diptera: Brachycera, Calyptratae):
implications for host associations and phylogeographic origins. Molecular
Phylogenetics and Evolution, 38: 155 170.
Espírito-Santo M.M., Sevilha A.C., Anaya F.C., Barbosa R., Fernandes G.W., Sanchez-
Azofeifa G.A., Scariot A., Noronha S.E.D. & Sampaio C.A. 2009. Sustainability of
tropical dry forests: Two case studies in southeastern and central Brazil. Forest Ecology
and Management, 258: 922-930.
Fleming T.H. 1988. The short-tailed fruit bat. A study in plant-animal interactions.
University of Chicago Press, Chicago, 365 pp.
Fritz, G.N. 1983. Biology and ecology of bat flies (Diptera: Streblidae) on bats of the
genus Carollia. Journal of Medical Entomology, 20: 1–10.
Guimarães L.R. & D'Andretta M.A.V. 1956. Sinopse dos Nycteribiidae (Diptera) do

Novo Mundo. Arquivos de Zoologia. São Paulo, 10: 1-184.
Guerrero R. 1994a. Catalogo de los Streblidae (Diptera: Pupipara) parasitos de
murciélagos (Mammalia: Chiroptera) del Nuevo Mundo. II. Los grupos: pallidus,
caecus, major, uniformis, y longipes del genero Trichobius Gervais, 1844. Acta
Biologica Venezuelana, 15: 1–18.
- 1994b. Catalogo de los Streblidae (Diptera: Pupipara) parasitos de murciélagos
(Mammalia: Chiroptera) del Nuevo Mundo. IV. Trichobiinae com alas desarrolladas.
Boletin Entomologia Venezuelana, 9: 161–192.
- 1995. Catalogo de los Streblidae (Diptera: Pupipara) parasitos de murciélagos
(Mammalia: Chiroptera) del Nuevo Mundo. V. Trichobiinae con alas reducidas o
ausentes y miscelaneos. Boletin Entomologia Venezuelana, 10: 135–160.
- 1996. Catalogo de los Streblidae (Diptera: Pupipara) parasitos de murciélagos
(Mammalia: Chiroptera) del Nuevo Mundo. VI. Streblinae. Acta Biologica
Venezuelana, 16: 1–26.
- 1997. Catalogo de los Streblidae (Diptera: Pupipara) parasitos de murciélagos
(Mammalia: Chiroptera) del Nuevo Mundo. VII. Lista de espécies, hospedadores y
paises. Acta Biológica Venezuelana, 17: 9–24.
Graciolli G. & Carvalho C.J.B. 2001. Moscas ectoparasitas (Diptera, Hippoboscoidea)
de morcegos (Mammalia, Chiroptera) do Estado do Paraná, Brasil. II. Streblidae. Chave

pictórica para as espécies. Revista Brasileira de Zoologia, 18: 907-960.
Graciolli G. & Coelho D.C. 2001. Streblidae (Diptera, Hippoboscoidea) sobre morcegos
filostomídeos (Chiroptera, Phyllostomidae) em cavernas do Distrito Federal. Revista
Brasileira de Zoologia, 18: 965-970.
Graciolli G. & Rui A.M. 2001. Streblidae (Diptera, Hippoboscoidea) em morcegos
filostomídeos (Chiroptera, Phyllostomidae) no noroeste do Rio Grande do Sul, Brasil.
Iheringia, Série Zoologia, 90: 85-92.
Graciolli G. & Linardi P. 2002. Some Streblidae and Nycteribiidae (Diptera:
Hippoboscoidea) from Maracá Island, Roraima, Brazil. Memórias do Instituto Oswaldo
Cruz, 97: 139-141.
Graciolli G. & Aguiar L.S. 2002. Ocorrência de moscas ectoparasitas (Diptera,
Streblidae e Nycteribiidae) de morcegos (Mammalia, Chiroptera) no Cerrado de
Brasília, Distrito Federal, Brasil. Revista Brasileira de Zoologia, 19: 177–181.
Graciolli G. 2003. Uma nova espécie de Strebla Wiedemann, 1824 (Diptera, Streblidae,
Streblinae) sobre Anoura caudifer (E. Geoffroy, 1818) (Chiroptera, Phyllostomidae,
Glossophaginae). Revista Brasileira de Entomologia, 47: 435-436.
Graciolli G. & Dick C.W. 2006. Checklist of world Nycteribiidae (Diptera:
Hippoboscoidea). National Science Foundation, 9p. Disponível em
http://fm1.fieldmuseum.org/aa/Files/cdick/ Nycteribiidae_Checklist_2oct06.pdf.

Graciolli G., Cáceres N.C. & Bornschein M.R. 2006a. Novos registros de moscas
ectoparasitas (Diptera, Streblidae e Nycteribiidae) de morcegos (Mammalia, Chiroptera)
em áreas de transição Cerrado-floresta estacional no Mato Grosso do Sul, Brasil.
Disponível em: http://www.biotaneotropica.org.br/v6n2/pt/abstract?short-
communication+bn03206022006.
Graciolli G., Passos F.C., Pedro W.A. & Lim B.K. 2006b. Moscas ectoparasitas
(Diptera: Streblidae) de morcegos filostomídeos (Mammalia, Chiroptera) na Estação
Ecológica dos Caetetus, São Paulo, Brasil. Revista Brasileira de Zoologia, 23: 298-299.
Graciolli G., Zortea M. & Carvalho L.F.A.C. 2010. Bat flies (Diptera, Streblidae and
Nycteribiidae) in a Cerrado area of state of Goiás, Brazil. Revista Brasileira de
Entomologia, 54: 511-514.
Gregorin R. & Taddei V.A. 2002. Chave Artificial para a Identificação de Molossídeos
Brasileiros (Mammalia, Chiroptera). Mastozoología Neotropical 9: 13-32.
IEF - Instituto Estadual de Florestas. 2000. Parecer técnico para a criação do Parque
Estadual da Mata Seca. Instituto Estadual de Florestas, Belo Horizonte.
Kim K.C. 1985. Coevolution of parasitic arthropods and mammals. John Wiley and
Sons, New York, p.800.
Komeno C.A. & Linhares A.X. 1999. Batflies parasitic on some phyllostomid bats in

Southeastern Brazil: parasitism rates and host-parasite relationships. Memórias do
Instituto Oswaldo Cruz, 94: 151–156.
Marshall, A.G. 1981. The Ecology of Ectoparasitic Insects. Academic Press, London,
459 pp.
Marshall A.G. 1982. Ecology of insects ectoparasites on bats, pp. 369-400. In: T.H.
Kunz (ed.). Ecology of bats. Plenum press, New York, USA, 69–401.
Medellín R.A., Arita H.T., Sánchez H. 1997. Identificación de los Murciélagos de
México, clave de campo. Asociación Mexicana de Mastozoología, México, 83 pp.
Moras L.M., Bernardi L.F.O., Graciolli G. & Gregorin R. 2013. Bat flies (Diptera:
Streblidae, Nycteribiidae) and mites (Acari) associated with bats (Mammalia:
Chiroptera) in a high-altitude region in southern Minas Gerais, Brazil. Acta
Parasitologica, 58 (4), 556–563.
Patterson B.D., Dick C.W. & Dittmar K. 2007. Roosting habits of bats affect their
parasitism by bat flies (Diptera: Streblidae). Journal of Tropical Ecological, 23: 177–
189.
Pezzini, F.F. 2008. Fenologia e características reprodutivas em comunidades arbóreas
em três estágios sucessionais em Floresta Estacional Decidual do norte de Minas Gerais.
Dissertação. Universidade Federal de Minas Gerais.

Prevedello J.A., Graciolli G. & Carvalho C.J.B. 2005. A Fauna de dípteros (Streblidae e
Nycteribiidae) ectoparasitos de morcegos (Chiroptera) do Estado do Paraná, Brasil:
composição, distribuição e áreas prioritárias para novos estudos. Biociências, 13
(2):193-209.
Poulin, R. & Mouillot, D. 2005. Combining phylogenetic and ecological information
into a new index of host specificity. Journal of Parasitology, 91: 511 – 514.
Quesada M., Sanchez-Azofeifa G.A., Alvarez-Añorve M., Stoner K.E., Avila-Cabadilla
L., Calvo-Alvarado J., Castillo A., Espírito-Santo M.M., Fagundes M. & Fernandes
G.W. 2009. Succession and management of tropical dry forests in the Americas: Review
and new perspectives. Forest Ecology and Management, 258: 1014-1024.
R Development Core Team (2010) R: a language and environment for statistical
computing. Vienna: R Foundation for Statistical Computing. ISBN 3–900051–07–0.
Available: http://www.R-project.org.
Reis NR, Peracchi AL, Pedro WA, de Lima IP (2007) Morcegos do brasil. Londrina:
Universidade Estadual de Londrina. 253 p.
Rios G.F.P., Sá-Neto R.J. & Graciolli G. 2008. Fauna de dípteros parasitas de morcego
em uma área de Caatinga no nordeste do Brasil. Chiroptera Neotropical, 14: 339-345.
Rózsa L., Reiczigel J. & Majoros G. 2000. Quantifying parasites in samples of hosts.
Journal of Parasitology, 86: 228-232.

Rui A.M. & Graciolli G. 2005. Moscas ectoparasitas (Diptera: Streblidae) de morcegos
(Chiroptera, Phyllostomidae) no sul do Brasil: associações hospedeiro-parasitos e taxas
de infestação. Revista Brasileira de Zoologia, 22: 438-445.
Sánchez-Azofeifa G.A., Quesada M., Rodriguez J.P., Nassar J.M., Stoner K.E., Castillo,
A., Garvin T., Zent E.L., Calvo-Alvarado J.C., Kalacska M.E.R., Fajardo L., Gamon
J.A. & Cuevas-Reyes P. 2005. Research priorities for neotropical dry forests.
Biotropica, 37: 477–485.
Santos C.L.C., Pereira A.C.N., Bastos V.J.C., Graciolli G. & Rebêlo J.M.M. 2013.
Parasitism of ectoparasitic flies on bats in the northern Brazilian cerrado. Acta
Parasitologica, 58 (2), 207–214.
Soares F.A.M., Graciolli G., Alcântra D.M.C., Pinto-Ribeiro C.E.B., Valença G.C. &
Ferrari, S.F. 2013. Bat flies (Diptera: Streblidae) ectoparasites of bats at
an Atlantic Rainforest site in northeastern Brazil. Biota Neotropical, 13: 242-246.
Speakman J.R. & Racey P.A. 1986. The influence of body condition sexual
development of male brown long-eared bats (Plecotus auritus) in the wild. Journal of
Zoology, 210: 515-525.
Straube F.C. & Bianconi G.V. 2002. Sobre a grandeza e a unidade utilizada para estimar
esforço de captura com utilização de redes-de-neblina. Chiroptera Neotropical, 8: 150-
152.

Teixeira A.L.M. & Ferreira R.L. 2010. Fauna de dipteros parasitas (Diptera: Streblidae)
e taxas de infestação em morcegos presentes em cavidades artificiais em Minas Gerais.
Chiroptera Neotropical, 16: 748-754.
Wenzel R.L., Tipton V.J. & Kiewlicz, A. 1966. The streblid batflies of Panama (Diptera:
Calyptera: Streblidae). Em: (Eds. R.L. Wenzel, V.J. Tipton) Ectoparasites of Panama.
Field Museum of Natural History, Chicago, 677–723.
Wenzel R.L. 1976. The streblidae batflies of Venezuela (Diptera: Streblidae). Em: (Eds.
R.L. Wenzel, V.J. Tipton) Ectoparasites of Panama. Field Museum of Natural History,
Chicago, 405–675.