Embed Size (px)
Citation preview

EFFECTIVE CONCENTRATION AND DETECTION OF HUMAN ENTERIC
VIRUSES IN HAWAIIAN ENVIRONMENTAL WATERS
A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAIʻI AT MĀNOA IN PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
MICROBIOLOGY
MAY 2012
By
Christina Connell
Thesis Committee:
Yuanan Lu, Chairperson Roger Fujioka Hongwei Li
08 Fall

i
ACKNOWLEDGEMENT
The completion of this thesis would not have been
possible without the support of several people, to whom the
author is immeasurably indebted.
First, the author would like to thank her superb
advisor, Dr. Yuanan Lu, for his genuine guidance, support,
encouragement, and understanding throughout the duration of
this study. His invaluable attention and direction, as well
as his immense knowledge, are extremely admirable and will
forever be appreciated.
The author also wishes to thank her dedicated lab
members for their gracious assistance and camaraderie. Dr.
Roger Fujioka and Dr. Hongwei Li are gratefully
acknowledged for provision of their time and support
through serving as thesis committee members. And finally,
profound gratitude is expressed toward the author’s
incredibly inspiring family and friends.

ii
ABSTRACT
Health risks associated with sewage-contaminated
recreational waters are of important public health concern.
Reliable water monitoring systems are therefore crucial.
Current recreational water quality criteria rely
predominantly on the enumeration of bacterial indicators,
while potentially dangerous viral pathogens often remain
undetected. Human enteric viruses have been proposed as
alternative indicators; however, their detection is often
hindered by low viral concentrations present in the aquatic
environment.
Reported here are novel and effective laboratory
protocols for enhanced enteric virus detection in Hawaiian
environmental waters. First, a fine-tuned, highly optimized
assay for the detection of enterovirus, an important
enteric virus subset, was developed by comparatively
evaluating eighteen published enterovirus primer pairs for
detection sensitivity. The primer set exhibiting the lowest
detection limit under optimized conditions, EQ-1/EQ-2, was
validated through testing urban wastewater, and then
utilized in a field survey of 22 recreational bodies of
water located around the island of Oahu, Hawaii. Eleven
sites tested positive for enterovirus, indicating fecal
contamination in a significant portion of Hawaiian waters.

iii
Additionally, the filter-feeding phenomenon of
indigenous bivalve mollusks was explored as a natural
bioconcentration technique to infer microbial quality of
the surrounding waters. Shellfish were collected from 12
coastal locations and dissected for subsequent nucleic acid
extraction from internal tissues. Optimized RT-PCR/PCR
protocols were then applied to test for the presence of
various enteric viruses, including enterovirus, adenovirus,
norovirus genogroups I and II, and F-specific RNA
coliphage. Shellfish collected from around the island
tested positive for several enteric virus types, indicating
that these animals are indeed natural and competent
bioindicators of water quality. The extremely sensitive and
innovative techniques implemented here are valuable
resources to aid accurate reflection of microbial
contamination in Hawaii’s environmental waters.

iv
TABLE OF CONTENTS
ACKNOWLEDGEMENT ........................................... i ABSTRACT ................................................. ii LIST OF TABLES ............................................ v LIST OF FIGURES .......................................... vi CHAPTER 1: INTRODUCTION AND OVERALL PROPOSED STUDY ........ 1 BACKGROUND INFORMATION ................................... 1 STUDY DESIGN ............................................ 17 SPECIFIC AIMS ........................................... 19 SIGNIFICANCE ............................................ 20 REFERENCES .............................................. 21
CHAPTER 2: ENTEROVIRUS METHOD OPTIMIZATION AND SEWAGE .... 27 VALIDATION INTRODUCTION ............................................ 27 MATERIALS AND METHODS ................................... 29 RESULTS ................................................. 37 DISCUSSION .............................................. 42 REFERENCES .............................................. 44
CHAPTER 3: HUMAN ENTEROVIRUS OCCURRENCE IN HAWAIIAN ...... 47 ENVIRONMENTAL WATERS INTRODUCTION ............................................ 47 MATERIALS AND METHODS ................................... 48 RESULTS ................................................. 54 DISCUSSION .............................................. 58 REFERENCES .............................................. 61
CHAPTER 4: SHELLFISH-MEDIATED BIOACCUMULATION OF HUMAN ... 63 ENTERIC VIRUSES IN OAHU’S MARINE ENVIRONMENT INTRODUCTION ............................................ 63 MATERIALS AND METHODS ................................... 68 RESULTS ................................................. 77 DISCUSSION .............................................. 86 REFERENCES ............................................. 100
CHAPTER 5: CONCLUSIONS .................................. 105

v
LIST OF TABLES
1. Enterovirus primer sets employed in comparative ....... 32 analysis
2. PCR condition brackets included in optimization ....... 34 assay 3. Optimized amplification conditions and detection ...... 39 limits of EnV primer sets employed in comparative
analysis 4. Enterovirus detection in Hawaiian environmental ....... 55 waters 5. Optimized PCR reaction components for enteric virus ... 72
detection 6. Optimized PCR cycle conditions for enteric virus ...... 73
detection 7. Primer sets employed in bioaccumulation study ......... 74 8. Enteric virus detection in shellfish .................. 79 9. Enteric virus detection in corresponding water ........ 80 samples 10. Enterovirus detection correlation between ............ 81 shellfish and water samples 11. Enteric virus strains detected in Hawaiian ........... 84 marine environment

vi
LIST OF FIGURES
1. Agarose gel depicting enterovirus detection from ...... 41 urban sewage 2. Geographic locations of environmental water ........... 49 sampling sites on Oahu, HI 3. Nucleotide sequence analysis of EnV isolated from ..... 57
wastewater (multiple clones) and environmental water samples
4. Geographic locations of twelve sample collection ...... 69 sites included in shellfish-mediated bioaccumulation
study 5. Agarose gel depicting adenovirus detection from ....... 82
shellfish collected at Kualoa Park 6. Agarose gel depicting enterovirus detection from ...... 82 samples collected at Ko Olina Lagoon 3 7. Agarose gel depicting norovirus genogroup I ........... 83 detection from samples collected at Ko Olina Lagoon 3 8. Agarose gel depicting enterovirus detection from ...... 83 samples collected at Kahala Beach

1
CHAPTER 1
INTRODUCTION AND OVERALL PROPOSED STUDY
BACKGROUND INFORMATION
Sewage-associated recreational waterborne disease
Worldwide, various recreational activities involving
water exposure are quite popular. However, the usage of
recreational waters places the public at risk of
contracting various illnesses associated with waterborne
pathogens. Results of several epidemiological studies have
shown significantly increased illness incidents, including
gastrointestinal, respiratory, ear, ocular, and skin
infections, among those involved in water-based
recreational activities (Sinclair 2009). Since reporting
began in 1978, levels of recreational waterborne disease
are at their highest (Reynolds 2009).
Of paramount public health concern are waters
contaminated by human sewage (Bonkosky 2009). The world’s
oceans receive billions of liters of virus-laden treated
and untreated wastewater on any given day (Rao 1986), and
the negative impacts of fecal-oral waterborne disease
affect more people across the globe than other types of
infectious waterborne disease (Jofre and Blanch 2010). As
demonstrated in 2010 at Rancho Santa Margarita, California,

2
a single ruptured sewer line can rapidly release hundreds
of thousands of gallons of untreated sewage into coastal
waters, resulting in fecal pollution along miles of beaches
(Devine 2011).
Current recreational water quality monitoring standards
In order to secure public protection from diseases
associated with fecally-contaminated recreational waters,
effective systems for monitoring water quality are of
crucial importance (U.S. EPA 2011). Because it is not
practically feasible to screen for every possible pathogen
of concern, indicators of human fecal pollution are often
used to assess microbial water quality (Jofre and Blanch
2010). The criteria for an ideal indicator organism are as
follows (Bitton 2005):

3
1. The indicator should be part of the human intestinal
microflora.
2. Its presence should correlate with the presence of
fecally-borne pathogens.
3. It must be present in greater numbers than pathogens
of concern.
4. It should be at least equally resistant as pathogens
of concern to environmental factors and wastewater
treatment plant disinfection practices.
5. It should not multiply in the environment.
6. It should be easily, rapidly, and inexpensively
detectable.
7. It should be non-pathogenic.

4
Currently, widely accepted microorganisms for water
quality assessment are bacterial indicators, including E.
coli and enterococci (Turbow 2003, Choi 2005).
Epidemiological data has linked increased risk of
contracting gastrointestinal illness with exposure to
waters with elevated concentrations of these fecal
indicator bacteria, and established detection assays are
inexpensive, standardized, and widely available (Lees 2000,
Boehm 2003). Bacterial indicators have been used to infer
microbial water quality for decades (Noble 2001); however,
several drawbacks are associated with this indicator
system, and many bacterial indicators fail to fit all of
the criteria listed above for ideal indicator organisms.
For instance, the presence of fecal indicator bacteria is
not limited to human feces; they may be present in the
excrement of a wide variety of other warm-blooded animals
such sea gulls and pigeons, both of which congregate near
shorelines (Boehm 2003). Also, these indicator bacteria may
multiply in the environment after excretion from their
host, yielding inaccurate estimations of true fecal
pollution levels (Fong and Lipp 2005). Epidemiological
studies yield inconsistent results as to which indicators
are most correlated with illness incidents (Snyder 2009),

5
and waterborne outbreaks have occurred in waters free of
standard indicator organisms (Papapetropoulou 1998).
Tropical beaches in the United States receive more
visitors than all temperate beaches combined (Leeworthy
2001). The state of Hawaii has over 400 public beaches,
stretching along roughly 300 miles of Pacific Ocean
Coastline (Dorfman 2011). Therefore, in tropical regions
such as Hawaii, Guam, Puerto Rico, and south Florida,
reliable water quality monitoring methods are of utmost
importance. However, current monitoring techniques
utilizing bacterial indicators are especially questionable
in such regions. Recreational water quality standards
utilizing bacterial indicators have been developed using
data from temperate regions and are then applied worldwide.
However, these standards are not applicable in the tropics,
as bacterial indicators are known to proliferate naturally
in these environments, especially in soil (Fujioka,
University of Hawaii). As soil is frequently introduced
into environmental waters, these organisms then become
members of the normal aquatic microbial community, which in
turn may lead to inaccurate assessments of water pollution
levels (Bernhard 2000, Byamukama 2005, Boehm 2003). In
addition to soil, enterococcus is reported to reproduce in
biofilms found in drainage pipes, concrete channels, river

6
rocks, and beach sand; enterococci levels are also often
increased during rainstorms or high surf events
(nonindicative of fecal contamination). For these reasons,
Hawaii utilizes a secondary bacterial indicator,
Clostridium perfringens, to help trace human fecal
pollution. However, a robust, consistent monitoring method
has not yet been developed (HI DOH 2008).
Another major inadequacy of current water quality
management procedures is that bacterial indicators fail to
reliably reflect the presence of pathogenic viruses
(Gabrieli 2007, Sinclair 2009, Terio 2010). Over 100 types
of viral pathogens may be present in sewage-impacted water
(Lipp 2001). Viruses survive longer than indicator bacteria
in both fresh and marine water environments and are
generally more resistant than bacterial indicators during
conventional wastewater treatment processes (Fong and Lipp
2005). This is of significant concern, as viral outbreaks
have been linked to waters that meet state or local
bacterial water quality criteria (Sinclair 2009). Of all
gastrointestinal illnesses contracted in sewage-impacted
recreational waters, viruses are thought to be the primary
etiologic agents (US EPA 2011). Several viruses have been
associated with such outbreaks, including coxsackieviruses,
adenoviruses, echoviruses, hepatitis A virus, astroviruses,

7
and noroviruses (Sinclair 2009). Although most viral
illnesses contracted are mild, serious disease can occur,
including aseptic meningitis, encephalitis, poliomyelitis,
hepatitis, myocarditis and diabetes (Pond 2005). In 1995,
an adenovirus outbreak was associated with a contaminated
swimming pool in Greece that tested negative for three
different bacterial indicators – total coliform, fecal
coliform, and fecal streptococci (Papapetropoulou 1998).
Similarly, in 2001, a community-wide enterovirus outbreak
in Germany was tied to bathing in a particular pond that,
despite weekly testing, never contained higher-than-normal
levels of total coliforms, fecal coliforms, enterococci, or
Staphylococcus aureus (Hauri 2005). In 1987, 18 of 41 water
samples collected from Oak Creek, Arizona tested positive
for enterovirus or rotavirus, while the level of fecal
indicators remained within the regulatory range (Rose
1987). A study of the coastal waters in Santa Monica Bay,
California, found no significant relationship between the
presence of enteroviruses and any single routinely
monitored bacterial indicator (Noble 2001). Clearly,
bacterial indicators cannot be solely relied upon to
accurately assess microbial water quality. Alternative
monitoring systems are crucial in order to improve the
surveillance of recreational waters and secure protection

8
to the public from waterborne disease (Lees 2000, Gersberg
2006).
Human enteric viruses as alternative indicators
Human enteric viruses, represented by the
astroviruses, rotaviruses, noroviruses, adenoviruses, and
picornaviruses, are commonly harbored in environmental
waters and are suggested as possible alternative indicators
of microbial water quality (US EPA 2011, Lipp 2001, Griffin
2003). According to the US EPA, human enteric viruses cause
a large proportion of illnesses from human-impacted
recreational waters. The presence of human enteric viruses
in recreational waters has been linked with increased
disease risk around the world, including studies in
Australia, United Kingdom, Ireland, Canada, Israel, New
Orleans, and Hong Kong (Sinclair 2009, Terio 2010). Enteric
virus-associated waterborne outbreaks have been associated
with contaminated swimming pools, lakes, ponds, rivers,
reservoirs, oceans, fountains, hot springs, and drinking
water (Fong 2005). Enteric viruses are primarily
transmitted via the fecal-oral route, and viral particles
are shed in extremely high numbers from infected
individuals, typically between 105 and 1011 virions per gram
of stool (Fong 2005). Although most enteric virus

9
infections are primarily associated with diarrhea and self-
limiting gastroenteritis, they may also cause hepatitis,
conjunctivitis, and respiratory infections. Additionally,
enteric virus infections may cause aseptic meningitis,
encephalitis, and paralysis in immunocompromised
individuals, resulting in high mortality rates (Fong 2005).
Another concerning characteristic of human enteric viruses
is their low infectious dose; for instance, exposure to a
single rotavirus places an individual at a 31% risk of
infection, and 1 viral plaque forming unit (PFU) is enough
to establish infection in 1% of healthy adults (Fong and
Lipp 2005).
Common wastewater treatment processes fail to
completely inactivate enteric viruses (Fujioka 1980),
rendering recreational waters in areas such as Hawaii,
where primary-treated sewage is discharged into the sea on
a normal basis, vulnerable to viral contamination. Because
of their small size, enteric viruses are easily transported
from original sources of pollution via tidal currents,
water circulation, and prevailing winds (Rao 1986).
Additionally, their resistant protein coat enables
prolonged persistence in the environment under a wide range
of water temperatures, salinities, and pH values (Lipp
2001, Rajtar 2008). The tendency of many viruses to

10
associate with various solids in the aquatic environment,
such as sand, clays (montmorillonite, kaolinite, bentonite,
illite), live organisms (algae, bacteria), silts,
sediments, and other protective particles is another
survival-prolonging factor (Lipp 2001, Rao 1986). These
suspended solids and sediments offer safe havens from
enzymes, UV irradiation, and other degrading factors (Fong
and Lipp 2005). Following release into the environment,
enteric viruses can survive within the water column or
associated with particulate matter for weeks to months
(Lees 2000); some viruses have even been reported to
survive and remain infectious for up to 130 days in
seawater, up to 120 days in freshwater and sewage, and up
to 100 days in soil at 20 to 30°C (Fong and Lipp 2005). In
similar environments, fecal indicator bacteria do not
survive nearly as long. Additionally, unlike bacterial
indicators, enteric viruses are obligate intracellular
parasites and are therefore unable to replicate in the
aquatic environment outside of their respective human hosts
(Lees 2000). Due to large viral loads released into sewage-
impacted waters, increased environmental persistence
compared to indicator bacteria, and the significant role
viruses play in waterborne disease, enteric viruses show
promising potential to be used as alternative indicators

11
for a more accurate depiction of recreational water quality
(Fong and Lipp 2005).
Enteric viruses included in study
Adenovirus
Adenoviruses (AdV) belong to the Adenoviridae family,
genus Mastadenovirus, containing all human serotypes.
Adenoviral particles are icosahedrons, 80-90 nm in size,
and contain double-stranded DNA. Adenovirus is responsible
for causing a wide range of human illnesses, including
respiratory infections, ocular infections, enteric
infections, encephalitis, pneumonia, and genitourinary
infections. Infections are generally mild; however, a
number of fatal cases have been reported (Pond 2005).
Specific gastroenteritis-causing adenoviruses transmitted
via the fecal-oral route include adenoviruses 40 and 41
(Sinclair 2009); these two serotypes are estimated to cause
5-20% of acute gastroenteritis cases among infants and
young children (Kokkinos 2011).
Highly resistant to physical/chemical agents and
adverse pH conditions, AdV can persist in the aquatic
environment for prolonged periods. AdV are reported to be
60 times more resistant than RNA viruses to UV irradiation

12
(Thurston-Enriquez 2003, Fong and Lipp 2005), which is a
commonly used disinfection procedure at sewage treatment
plants, including the Sand Island Wastewater Treatment
Plant here in Hawaii. In raw sewage from around the world,
AdV are consistently more prevalent than enteroviruses
(Jiang 2001). Adenoviruses are frequently present in
polluted environmental waters and are easily detectable by
PCR; they have therefore been suggested as alternative
indicators of human fecal pollution (Lees 2000, Choi 2005).
Enterovirus
Enteroviruses (EnV) are single-stranded, positive-
sense RNA viruses belonging to the Picornaviridae family.
Enteroviral particles are icosahedral, nonenveloped, and
~30 nm in diameter. EnV, including coxsackievirus,
poliovirus, echovirus, and the numbered enteroviruses, are
the most commonly detected enteric viruses in polluted
waters and are estimated to cause 30 - 50 million
infections in the US annually (Donaldson 2002, Gregory
2006). Enteroviral epidemics are predominantly waterborne
(Rajtar 2008). The EnV disease spectrum is wide, including
gastroenteritis, respiratory infection, diabetes, heart
disease, bronchiolitis, conjunctivitis, meningitis,
paralysis, and the common cold (Fong 2005). EnV can remain

13
viable at extremely low temperatures (between minus 20°C and
70°C) for years, and for weeks at 4°C (Rajtar 2008). Because
these viruses are common, fecally shed in extremely high
numbers from infected individuals, highly tolerant to
salinity and temperature fluctuations, and stable in the
environment for extended time periods, they have been
suggested as a parameter for evaluating viral pollution of
environmental waters (Hot 2003, Gregory 2006). The
availability of permissive cell lines for determining EnV
infectivity greatly enhances the attractiveness of using
this important enteric virus subset as an alternative
indicator of water quality (Fong 2005).
Norovirus
Noroviruses (NoV), members of the Caliciviridae family
and Norovirus genus, are 27-32 nm in diameter, single-
stranded, non-enveloped RNA viruses. They are the most
common cause of acute, nonbacterial gastroenteritis,
causing ~23 million cases of acute onset vomiting and
diarrhea in the United States per year (Jothikumar 2005,
Mead 1999). Infections also sometimes include fever,
headache, and malaise. Two human genogroups are of interest
here, norovirus genogroups I (NoVI) and II (NoVII), based
on genetic divergence in the polymerase and capsid regions

14
(Boxman 2006). NoVII is primarily transmitted via person-
to-person (Siebenga 2007), while NoVI is more often
transmitted via food or environmental contamination
(Maalouf 2010). Waterborne outbreaks of norovirus-
associated gastroenteritis are well documented (Kokkinos
2011). Norovirus is the specific enteric virus suggested by
the EPA to be used as an alternative water quality
indicator (US EPA 2011). An efficient cell culture system
for isolating and propagating norovirus has not been
established (Sinclair 2009).
F-specific RNA coliphage
Although not classified as human enteric viruses,
male-specific FRNA coliphages have been considered suitable
indicators of fecal pollution in the environment (Umesha
2008). These viruses are quite prevalent in wastewater and
exhibit relatively high resistance to disinfection
procedures (Bitton 2005). Using only bacteria as hosts for
replication, these FRNA bacteriophages infect E. coli
carrying the F plasmid, which codes for the F or sex pilus,
to which the phages attach (Bitton 2005, Griffin 2003).
This group of viruses resembles human enteric viruses in
size, shape, and general composition (Sobsey 2006). F-
specific RNA coliphages are easily enumerated and have

15
shown slow elimination kinetics, also representative of
human enteric viruses (Hernroth 2002). FRNA coliphages may
be useful for fecal source tracking, as they distinguish
human fecal waste from non-human fecal waste (Sobsey 2006).
1.5 Virological analysis of environmental waters
The basic steps of virological analysis of
environmental waters include sampling, virus concentration,
and detection (either through cell culture or molecular
biology assays) (Fong and Lipp 2005). Although human
enteric viruses are suggested as alternative indicators of
microbial water quality, the low concentration of these
viruses present in environmental waters presents a major
problem (Fout 2003, Griffin 2003, Fong 2005). Therefore,
extremely sensitive detection methods are absolutely
essential if enteric viruses are to be effectively utilized
as alternative indicators of fecal contamination (Ijzerman
1997). Early studies utilized cell culture techniques for
the detection and isolation of enteric viruses; however,
these assays are laborious, expensive, and time-consuming.
Additionally, traditional cell lines are not established
for all enteric virus subsets of interest (Fong and Lipp
2005). RT-PCR-based assays, due to their rapidity,
sensitivity, specificity, and relative ease-of-use, are

16
often favored by environmental virologists (Schwab 1996).
These molecular assays have greatly enhanced environmental
enteric virus detection, especially for those viruses for
which no cell culture system is available (Rodriguez 2009).
However, the presence of inhibitory compounds, which can
lead to false-negative viral detection, presents an
additional detection barrier (Fout 2003). Detection
challenges may be overcome by improved methods for viral
concentration, efficient inhibitor removal during nucleic
acid extraction, and extremely sensitive molecular
detection assays (Karamoko 2005).

17
STUDY DESIGN
The goal of this study is the development and
exploration of novel, highly sensitive methods for
effective enteric virus detection in the Hawaiian aquatic
environment. These approaches are geared toward an ultimate
purpose, involving the utilization of human enteric viruses
as alternative microbial water quality indicators in order
to protect the public from waterborne disease.
First, a side-by-side comparison and optimization of
previously published EnV detection protocols was conducted,
yielding a fine-tuned, highly sensitive and optimized RT-
PCR procedure for the effective detection of EnV from
environmental water samples. As confirmation of the newly
developed protocol, it was applied to urban wastewater
samples at various treatment stages for method validation.
Next, in order to utilize the assay in a practical manner,
an EnV surveillance study of recreational waters around the
island of Oahu was conducted.
Finally, as a technique to enhance viral concentration
from coastal water samples, the bioaccumulation phenomenon
of bivalve shellfish was explored. Indigenous mollusks were
collected from multiple sites around the island of Oahu and
screened for the presence of various enteric viruses, using
individually optimized PCR protocols. Direct water samples

18
were simultaneously collected and processed in order to
compare enteric viral detection efficiency. Positive
enteric virus detection from sewage, environmental water,
and shellfish samples was confirmed through DNA sequencing.

19
SPECIFIC AIMS
Specific aims of this study are as follows:
1. Establishment of a highly sensitive RT-PCR protocol for
EnV detection, accomplished by screening and optimizing
conditions for previously published EnV primer sets;
2. Validation of the established protocol through testing
urban wastewater collected at various treatment stages;
3. Utilization of optimized protocol through environmental
surveillance study: screen 22 local recreational bodies
of water for the presence of EnV;
4. Shellfish-mediated bioaccumulation study to explore a
natural means of enhanced viral concentration. Apply
optimized detection protocols for EnV and other enteric
viruses to nucleic acids extracted from shellfish
collected at 12 Oahu beaches.

20
SIGNIFICANCE
The innovative viral concentration and detection
techniques described here may enrich alternative water
quality monitoring methods not only in the state of Hawaii,
but worldwide. These findings contribute to a better
understanding of the prevalence of fecal contamination in
Hawaiian environmental waters. The development and
utilization of novel enteric virus detection assays may be
of great interest to those interested in recreational water
quality, including environmental virologists, public health
officials, researchers, regulators, and even the general
public.

21
REFERENCES
Bernhard AE, Field KG (2000) Identification of nonpoint sources of fecal pollution in coastal waters by using host-specific 16S ribosomal DNA genetic markers from fecal anaerobes. Appl Environ Microbiol 66:1587-1594 Bitton G (2005) Microbial indicators of fecal contamination: application to microbial source tracking. Report submitted from Department of Environmental Engineering Sciences, University of Florida to Florida Stormwater Association Boehm AB, Fuhrman JA, Mrse RD, Grant SB (2003) Tiered approach for identification of a human fecal pollution source at a recreational beach: Case study at Avalon Bay, Catalina Island, California. Environ Sci Technol 37: 673-680 Bonkosky M, Hernandez-Delgado EA, Sandoz B, Robledo IE, Norat-Ramirez J, Mattei H (2009) Detection of spatial fluctuations of non-point source fecal pollution in coral reef surrounding waters in southwestern Puerto Rico using PCR-based assays Boxman ILA, Tilburg JJHC, te Loeke NAJM, Vennema H, Jonker K, de Boer E, Koopmans M (2006) Detection of noroviruses in shellfish in the Netherlands. International J Food Microbiol 108: 391-396 Byamukama D, Kansiime F, Mach RL, Farnleitner AH (2000) Determination of Escherichia coli contamination with chromocult coliform agar showed a high level of discrimination efficiency for differing fecal pollution levels in tropical waters of Kampala, Uganda Choi S, Jiang SC (2005) Real-time PCR quantification of human adenoviruses in urban rivers indicates genome prevalence but low infectivity. Appl Environ Microbiol 71: 7426-7433 Devine J (2011) Sources of beachwater pollution. Natural Resources Defense Council Donaldson KA, Griffin DW, Paul JH (2002) Detection, quantitation, and identification of enteroviruses from surface waters and sponge tissue from the Florida Keys using real-time RT-PCR. Water Res 36: 2505-2514

22
Dorfman M and Rosselot KS (2011) Testing the waters: A guide to water quality at vacation beaches. Twenty-first annual report. Natural Resources Defense Council Fong T, Lipp EK (2005) Enteric viruses of humans and animals in aquatic environments: health risks, detection, and potential water quality assessment tools. Microbiol Mol Bio Rev 69: 357-371 Fout GS, Martinson BC, Moyer MWN, Dahling DR (2003) A multiplex reverse transcription-PCR method for detection of human enteric viruses in groundwater. Appl Environ Microbi 69: 3158-3164 Fujioka RS, Loh PC, Lau LS (1980) Survival of human enteroviruses in the Hawaiian ocean environment: Evidence for virus-inactivating microorganisms. Appl Environ Microbiol 39: 1105-1110 Fujioka R, Loh P, Katz A, Seifried S, Steward G, Tice A, Wilcox B. Microbial pathogens in tropical coastal waters: an ecosystem approach to determine risk and prevent waterborne diseases. Pacific Research Center for Marine Biomedicine, University of Hawaii Gabrieli R, Macaluso A, Lanni L, Saccares S, Di Giamberardino F, Cencioni B, Petrinca AR, Divizia M (2007) Enteric viruses in molluscan shellfish. New Microbiologica 30: 471-475 Gersberg RM, Rose MA, Robles-Sikisaka R, Dhar AK (2006) Quantitative detection of hepatitis A virus and enteroviruses near the United States-Mexico border and correlation with levels of fecal indicator bacteria. Appl Environ Microbiol 72: 7438-7444 Gregory JB, Litaker RW, Noble RT (2006) Rapid one-step quantitative reverse transcriptase PCR assay with competitive internal positive control for detection of enteroviruses in environmental samples. Appl Environ Microbiol 72: 3960-3967 Griffin DW, Donaldson KA, Paul JH, Rose JB (2003) Pathogenic human viruses in coastal waters. Clin Microbiol Rev 16: 129-143 Hauri AM, Schimmelpfennig M, Walter-Domes M, Letz A, Diedrich S, Lopez-Pila J, Schreier E (2005) An outbreak of

23
viral meningitis associated with a public swimming pond. Epidemiol Infect 133: 291–298
Hawaii State Department of Health (2008) 2006 State of Hawaii water quality monitoring and assessment report: integrated report to the U.S. Environmental Protection Agency and the U.S. Congress pursuant to sections 303(D) and 305(B) Clean Water Act (P.L. 97-117). Honolulu, HI.
Hernroth BE, Conden-Hansson ACC, Rehnstam-Holm AS, Girones R, Allard AK (2002) Environmental factors influencing human viral pathogens and their potential indicator organisms in the blue mussel, Mytilus edulis: the first Scandinavian report. Appl Environ Microbiol 68: 4523-4533 Hot D, Legeay O, Jacques J, Gantzer C, Caudrelier, Y et al (2003) Detection of somatic phages, infectious enteroviruses and enterovirus genomes as indicators of human enteric viral pollution in surface water. Water Res 37: 4703-710 Ijzerman MM, Dahling DR, Fout GS (1997) A method to remove environmental inhibitors prior to the detection of waterborne enteric viruses by reverse transcription-polymerase chain reaction. J Virol Methods 63: 145-153 Jiang S, Noble R, Chu Weiping (2001) Human adenoviruses and coliphages in urban runoff-impacted coastal waters of Southern California. Appl Environ Microbiol 67: 179-184 Jofre J and Blanch AR (2010) Feasibility of methods based on nucleic acid amplification techniques to fulfill the requirements for microbiological analysis of water quality. J of Appl Microbiol 109: 1853-1867 Jothikumar N, Lowther JA, Henshilwood K, Lees DN, Hill VR, Vinje J (2005) Rapid and sensitive detection of noroviruses by using TaqMan-based one-step reverse transcription-PCR assays and application to naturally contaminated shellfish samples. Appl Environ Microbiol 71: 1870-1875 Karamoko Y, Ibenyassine K, Aitmhand R, Idaomar M, Ennaji MM (2005) Adenovirus detection in shellfish and urban sewage in Morocco (Casablanca region) by the polymerase chain reaction. J Virol Methods 126: 135-137 Kokkinos P, Ziros P, Meri D, Filippidou S, Kolla S, Galanis A, Vantarakis A (2011) Environmental surveillance. An additional/alternative approach for virological

24
surveillance in Greece? Int J Environ Rese Public Health 8: 1914-1922 Lees D (2000) Viruses and bivalve shellfish. International J Food Microbiol 59: 81-116 Leeworthy VR and Wiley PC (2001) Marine recreation participation and use, national survey on recreation and the environment. National Ocean Service, National Ocean and Atmospheric Administration: Silver Spring, MD Lipp EK, Lukasik J, Rose JB (2001) Human enteric viruses and parasites in the marine environment. Methods in Microbiology 30: 559-588 Maalouf H, Zakhour M, Le Pendu J, Le Saux JC, Atmar RL, Le Guyader FS (2010) Distribution in tissue and seasonal variation of norovirus genogroup I and II ligans in oysters. Appl Environ Microbiol 76:5621-5630 Mead PS, Slutsker L, Dietz V, McCaig LF, Bresee JS, Shapiro C, Griffin PM, Tauxe RV (1999) Food- related illness and death in the United States. Emerg Infect Dis 5: 607–625 Noble RT, Fuhrman JA (2001) Enteroviruses detected by reverse transcriptase polymerase chain reaction from the coastal waters of Santa Monica Bay, California: low correlation of bacterial indicator levels. Hydrobiologica 460: 175-184 Papapetropoulou M and Vantarakis AC (1998) Detection of adenovirus outbreak at a municipal swimming pool by nested PCR amplification. J Infect 36: 101–103 Pond, Kathy (2005) Water recreation and disease - Plausibility of associated infections: acute effects, sequelae and mortality. World Health Organization Rajtar B, Majek M, Polanski L, Polz-Dacewicz M (2008) Enteroviruses in water environment – a potential threat to public health. Ann Agric Environ Med 15: 199-203 Rao VC, Metcalf TG, Melnick JL (1986) Human viruses in sediments, sludges, and soils. Bulletin of the World Health Organization 64: 1-14 Reynolds KA (2009) Recreational waterborne disease at an all-time high. Water Conditioning and Purification April 2009: 56-58

25
Rodriguez RA, Pepper IL, Gerba CP (2009) Applicaton of PCR-based methods to assess the infectivity of enteric viruses in environmental samples. Appl Environ Microbiol 75: 297-307 Rose, J.B., Mullinax, R.L., Singh, S.N., Yates, M.V. and Gerba, C.P. (1987) Occurrence of rotaviruses and enteroviruses in recreational waters of Oak Creek, Arizona. Water Res 11, 1375–1381
Schwab KJ, Leon RD, Sobsey MD (1996) Immunoaffinity concentration and purification of waterborne enteric viruses for detection by reverse transcriptase PCR. Appl Environ Microbiol 62: 2086-2094 Siebenga JH, Vennema DP, Zheng J, Vinje, BE, Lee XL et al (2009) Norovirus illness is a global problem: emergence and spread of norovirus GII.4 variants, 2001-2007. J Infect Dis 200:802-812 Sinclair RG, Jones EL, Gerba CP (2009) Viruses in recreational water-borne disease outbreaks: a review. J of Appl Microbiol 107: 1769-1780 Sobsey MD, Love DC, Lovelace GL (2006) F+ RNA coliphages as source tracking viral indicators of fecal contamination. NOAA/UNH Cooperative Institute for Coastal and Estuarine Environmental Technology (CICEET) Terio V, Di Pinto A, Di Pinto P, Martella V, Tantillo G (2010) RNA extraction method for the PCR detection of hepatitis A virus in shellfish. International Journal of Food Microbiology 142: 198-201 Thurston-Enriquez JA, Haas CN, Jacangelo J, Riley K, Gerba CP (2003) Inactivation of feline calicivirus and adenovirus type 40 by UV radiation. Appl Environ Microbiol 69: 577–582 Turbow DJ, Osgood ND, Jiang SC (2003) Evaluation of recreational health risk in coastal waters based on enterococcus densities and bathing patterns. Environmental Health Perspectives 11: 598-603 Umesha KR, Bhavani NC, Venugopal MN, Karunasagar I, Krohne G, Karunasagar I (2008) Prevalence of human pathogenic enteric viruses in bivalve molluscan shellfish and cultured shrimp in south west coast of India. International J Food Microbiology 122: 279-286

26
U.S. Environmental Protection Agency (EPA) (2011) Recreational Water Quality Criteria. Office of Water. 66 p Viau EJ, Lee D, Boehm AB (2011) Swimmer risk of gastrointestinal illness from exposure to tropical coastal waters impacted by terrestrial dry-weather runoff. Environmental Sci Technol 45: 7158-7165

27
CHAPTER 2
ENTEROVIRUS METHOD OPTIMIZATION AND SEWAGE VALIDATION INTRODUCTION
The fecally-derived enteric viruses have been proposed
as alternative water quality monitoring indicators.
However, their effective detection is hindered by extremely
low concentrations in diluted recreational waters.
Therefore, a pressing need exists for the development of
sensitive and efficient viral monitoring assays. Optimized
protocols for the detection of several enteric viruses,
including AdV, NoVI, NoVII, and F+, have already been
established in this laboratory (Tong & Lu 2011, Tong et al.
2011). Discussed here is the development of a highly
sensitive protocol for the detection of another important
enteric virus subset, EnV.
Several molecular assays exist for the environmental
detection of EnV, but little is known about their
detection limits and sensitivities. Therefore, eighteen
published EnV primer sets have been analyzed here in a
side-by-side comparative study, and PCR conditions have
been individually optimized for peak sensitivity. The
primer set exhibiting the lowest detection limit under
optimized conditions was then validated through testing
urban wastewater.

28
Of various polluted waters, sewage contains the
highest concentrations of microbial pathogens and fecal
indicators (Jofre and Blanch 2010). The microbial content
of raw or inadequately treated sewage reflects the health
status of the population from where it is derived (Pond
2005). Because multiple enteroviral strains are fecally
shed in high loads from infected individuals (Hot 2003),
urban wastewater was used as the nucleic acid source for
optimization of EnV molecular amplification. Wastewater was
obtained from Sand Island Wastewater Treatment Plant, which
receives sewage and other wastewater from businesses and
residences of the City of Honolulu, including Waikiki. This
treatment facility is the largest treatment facility on the
island of Oahu, responsible for processing approximately
85% of the island’s wastewater. As of 2007, approximately
412,000 people were served by the Sand Island plant,
equating to an estimate of 66 million gallons of raw
influent per day. This facility utilizes an advanced
primary treatment, disinfecting sewage via ultraviolet (UV)
radiation before releasing it 1.7 miles offshore into
Mamala Bay via the Sand Island outfall. This outfall rests
approximately 230 feet below the surface of the ocean (US
EPA 2007, Tong 2011).

29
MATERIALS AND METHODS
Wastewater sample collection
Wastewater samples were collected in 2-L sterile,
polypropylene containers from the Sand Island Wastewater
Treatment Plant at the following three treatment stages:
raw influent, post-primary clarification/pre-UV
disinfection, and post-UV disinfection/effluent. Samples
were transported on ice to a BSL-2 laboratory located at
the University of Hawaii at Manoa and processed
immediately.
Sample concentration and nucleic acid extraction
Sewage samples were processed using a filtration-based
method adapted from Tong and Lu (2011). In order to enhance
viral absorption to membrane filters, MgCl2 solution was
mixed into samples prior to filtration at a final
concentration of 25 mM. 100 mL of sewage from each of the
three stages were filtered through 0.45-µM pore size, type
HA membranes (Millipore Corporation, MA) on a filtration
manifold under vacuum. (It should be noted that this was
the maximum passable volume before filtration ceased due to
membrane clogging.) RNA was extracted from the recovered
membranes using the PowerWater RNA Isolation Kit, supplied

30
by MoBio Laboratories, CA. RNA was stored at -80°C until RT-
PCR was to be performed.
cDNA preparation via RT-PCR
Because enteroviruses are RNA viruses, the extracted
RNA must first be converted to cDNA via RT-PCR before PCR
is performed. Seven microliters of RNA extracted from each
of the three sewage stages were used as RT-PCR templates,
performed with the DyNAmo cDNA synthesis kit (New England
Biolabs, NEB, MA) according to the manufacturer’s
instructions. Random hexamers were used as primers. cDNA
was stored at -20°C until PCR performance.
Comparative analysis of published enterovirus primer sets
While several RT-PCR protocols have already been
established for the detection of EnV, little is known about
their comparative detection sensitivities, which is of
utmost importance when assessing microbial water quality.
Therefore, eighteen published primer sets, specific for
amplifying various regions of the EnV genome, were selected
in this study in a comparative evaluation of detection
sensitivity (Table 1). The primer sets chosen are specific
for all pathogenic but highly diverse human enteroviruses,
with the exception of EvVP1F/EvVP1R, which specifically

31
selects for EV71, causative agent of hand, foot, and mouth
disease in children (Tan 2006). All primers were
synthesized by Integrated DNA Technologies (IDT, IA).
Initial PCR test
All primer sets were initially tested under standard
PCR conditions using single-source cDNA from wastewater
influent as the nucleic acid template. Five microliters of
cDNA was added to 20 µL PCR mix containing 1X Taq reaction
buffer (NEB, MA), 2.0 mM MgCl2 solution (NEB, MA), 200 nM of
each dNTP (Sigma-Adrich, MO), 400 nM of forward and reverse
primers (Integrated DNA Technologies, IA), and 2 units of
Taq DNA polymerase (provided by Dr. Tung Hoang, University
of Hawaii at Manoa). Reaction tubes were placed in a
Mastercycler® Gradient (Eppendorf, Germany) for an initial
denaturation at 94°C for 5 min., followed by 40 cycles of
denaturation at 94°C for 30 sec., annealing at 56°C for 20
sec., and extension at 72°C for 30 sec., completed by a
final extension at 72°C for 5 min.

32
Table 1. Enterovirus primer sets employed in comparative analysis
Primer Sequence (5' - 3') Amplicon size (bp) Ref.
EV1/EV2 CGGCCCCTGAATGCGGC / CACCGGATGGCCAATCCA 196
Gomara et. al 2006 EntAF/R TNCARGCWGCNGARACNGG / ANGGRTTNGTNGMWGTYTGCCA 414
EntBF/R GCNGYNGARACNGGNCACAC / CTNGGRTTNGTNGANGWYTGCC 397
EntCF/R TNACNGCNGTNGANACHGG / TGCCANGTRTANTCRTCCC 395
EQ-1/EQ-2 ACATGGTGTGAAGAGTCTATTGAGCT / CCAAAGTAGTCGGTTCCGC 142 Dierssen et. al 1998
EvVP1F/R GAGAGTTCTATAGGGGACAGT / AGCTGTGCTATGTGAATTAGGAA 204 Tan et. al 2006
2AB/2C3A GAIGYIATGGARCARGG / GGICCYTGRAAIARIGCYTC 1200 Bessaud et. al 2008
EV1/EV2 GGCCCCTGAATGCGGCTAAT / CAATTGTCACCATAAGCAGCCA 54 Hymas et. al 2008
Lees3/4 CATTCAGGGGCCGGAGGA / AAGCACTTCTGTTTCC 256 Lees et. al 1994
P1/P3 CAAGCACTTCTGTTTCCCCGG / ATTGTCACCATAAGCAGCCA 440 Zoll et. al 1991
P2/P3 TCCTCCGGCCCCTGAATGCG / ATTGTCACCATAAGCAGCCA 155
EV-L/-R CCTCCGGCCCCTGAATG / ACCGCGATGGCCAATCCAA 197 Chung et. al 1996
Abba1/2 TGTCACCATAAGCAGCC / TCCGGCCCCTGAATGCGGCT 149 Abbaszadegan et. al 1993
EVZ1/Z2 CAAGCACTTCTGTTTCCCCGG / ACCCATAGTAGTCGGTTCCGC 388 Zhang et. al 2010
EVF/EVR CCTGAATGCGGCTAATCC / ATTGTCACCATAAGCAGCCA 144 Jothikumar et. al 2010
ev1q/ev2q GATTGTCACCATAAGCAGC / CCCCTGAATGCGGCTAATC 146 Fuhrman et. al 2005
Ent1/Ent2 CGGGTACCTTTGTACGCCTGT / ATTGTCACCATAAGCAGCCA 534 Puig et. al 1994
EvUp/Dwn TGTCACCATAAGCAGCC / TCCGGCCCCTGAATGCGGCT 149 Reynolds et. al 1998

33
EnV detection was analyzed by gel electrophoresis. 10
µL PCR product + 2 µL 6x loading dye was loaded into the
wells of an ethidium-bromide stained 2% agarose gel in 0.5x
TBE buffer, to which 120V was applied until sufficient
fragment migration had occurred. A 50-bp DNA ladder (NEB,
MA) was used for indication of PCR product fragment size.
The Molecular Imager Gel Doc XR+ system (BioRad
Laboratories, Inc., CA) was used to visualize results under
UV light.
PCR condition optimization
PCR conditions for all primer sets that successfully
detected EnV from untreated wastewater were then adjusted
for optimal sensitivity. Optimization brackets included
annealing temperature, MgCl2 concentration, primer
concentration, and the presence or absence of 0.1 µg/µL
molecular biology grade, protease/nuclease-free, fraction V
BSA (NEB, MA) (Table 2).

34
Table 2. PCR condition brackets included in optimization assay
Condition Test Range
Tanneal 50 - 60°Ca, 2° increments
[MgCl2] 1.5, 2.0, 3.0, 4.0 mM
[Primer] 200, 400, 600, 800, 1000 nM
BSA Presence/Absence (0.1 µg/µL)
a 40-50°C was included if reported Tanneal in literature was <50°C

35
Detection limit comparison using optimized PCR conditions
Using the final optimized conditions, primer set
detection limits were determined by PCR using 10-fold
serial dilutions of influent sewage cDNA template.
Detection limits were denoted by the highest dilution
yielding a clear, positive detection signal, visualized by
performing gel electrophoresis after PCR amplification.
Sewage validation
Once PCR conditions had been individually optimized
for all successful primer sets and detection limits had
been determined, the primer set exhibiting the highest
sensitivity, EQ-1/EQ-2, was confirmed using cDNA obtained
from the three sewage stages described earlier (raw
influent, post-clarification/pre-UV disinfection, and post-
UV disinfection/effluent). PCR and subsequent gel
electrophoresis were performed under optimized conditions
as described above.
PCR product sequencing and analysis
In order to confirm true EnV detection and identify
enteroviral strains present in Honolulu wastewater,
selected positive DNA fragments amplified by primer set EQ-
1/EQ-2 were subjected to DNA sequencing. DNA bands were

36
excised from the 2% agarose gel and recovered using the
QIAquick Gel Extraction kit (Qiagen, CA), according to the
manufacturer’s instructions. Recovered DNA samples from
sewage were eluted using 30 μL EB buffer and cloned into
pCR®2.1-TOPO® vectors using the TOPO TA Cloning® kit
(Invitrogen, CA) according to the manufacturer’s
instructions. 8 positive clones from a single influent
sewage sample were submitted with the M13 forward primer,
provided by the commercial kit, to the College of Natural
Sciences Advanced Studies of Genomics, Proteomics and
Bioinformatics (ASGPB, University of Hawaii at Manoa) for
DNA sequencing. Resulting genomic sequences were aligned
and compared with all available EnV sequences listed in the
National Center for Biotechnology Information (NCBI)
databank using the Basic Local Alignment Search Tool
(BLAST).

37
RESULTS
RT-PCR condition optimization
Of the initial 18 primer sets tested, only 7 generated
PCR products of the expected size from untreated sewage,
indicating positive EnV detection (EQ-1/EQ-2, Primer
1/Primer 3, Primer 2/Primer 3, EV-L/EV-R, EVZ1/EVZ2,
EVF/EVR, ev1qia/ev2qia). Conditions for these 7 pairs were
then optimized for their use in conventional PCR. Optimal
annealing temperatures, salt concentrations, primer
concentrations, and BSA presence/absence for these 7 primer
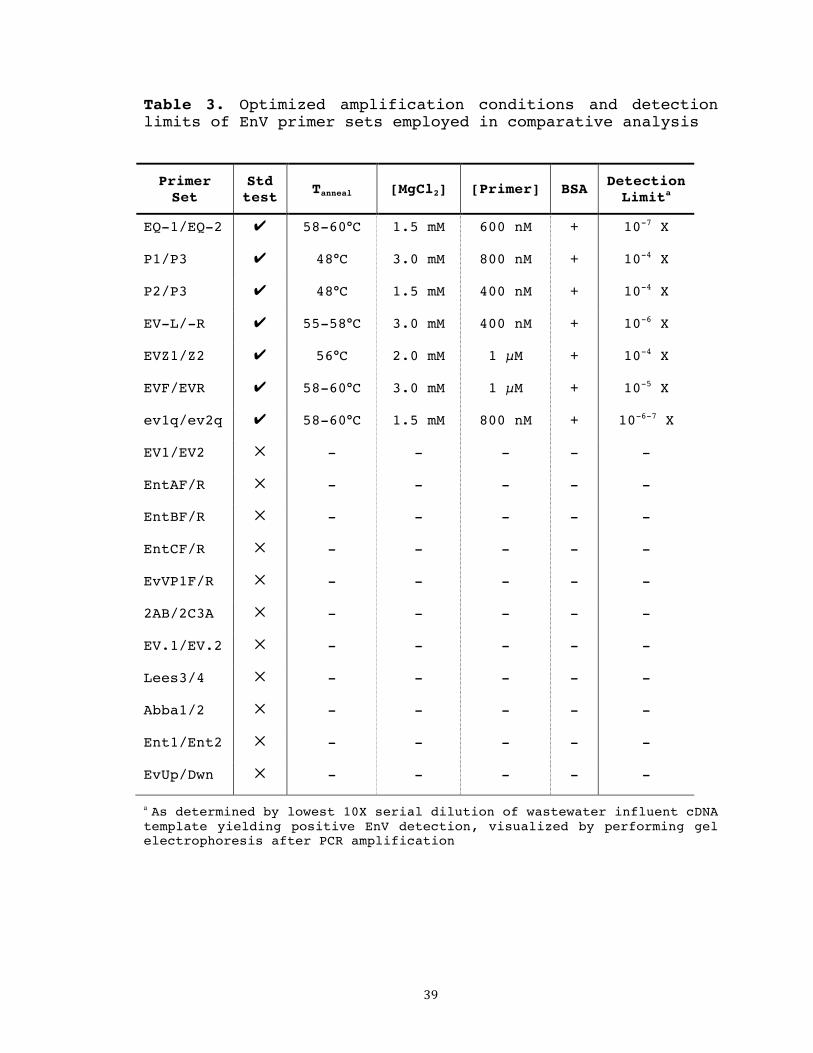
sets are summarized in Table 3. It was found that the
addition of BSA increased detection strength of all 7 sets
of primer pairs. Although the exact mechanism by which BSA
enhances PCR reactions is unknown, it is thought to relieve
any lingering inhibitory effects (Kreader 1996).
Determination of detection sensitivities
Detection limits significantly varied among these
seven primer sets, differing by as much as 1000-fold (Table
3). The primer set exhibiting the highest sensitivity, EQ-
1/EQ-2, with a detection limit of 10-7 X, was selected for
further experimentation. This primer set generates a 142
base pair amplicon within the highly conserved 5’ UTR

38
region of the EnV genome, including parts of domains IV and
V of the internal ribosomal entry site (Dierssen 1998).
39
Table 3. Optimized amplification conditions and detection limits of EnV primer sets employed in comparative analysis
Primer Set
Std test Tanneal [MgCl2] [Primer] BSA Detection
Limita
EQ-1/EQ-2 ✔ 58-60°C 1.5 mM 600 nM + 10-7 X
P1/P3 ✔ 48°C 3.0 mM 800 nM + 10-4 X
P2/P3 ✔ 48°C 1.5 mM 400 nM + 10-4 X
EV-L/-R ✔ 55-58°C 3.0 mM 400 nM + 10-6 X
EVZ1/Z2 ✔ 56°C 2.0 mM 1 µM + 10-4 X
EVF/EVR ✔ 58-60°C 3.0 mM 1 µM + 10-5 X
ev1q/ev2q ✔ 58-60°C 1.5 mM 800 nM + 10-6-7 X
EV1/EV2 ✕ - - - - -
EntAF/R ✕ - - - - -
EntBF/R ✕ - - - - -
EntCF/R ✕ - - - - -
EvVP1F/R ✕ - - - - -
2AB/2C3A ✕ - - - - -
EV.1/EV.2 ✕ - - - - -
Lees3/4 ✕ - - - - -
Abba1/2 ✕ - - - - -
Ent1/Ent2 ✕ - - - - -
EvUp/Dwn ✕ - - - - -
a As determined by lowest 10X serial dilution of wastewater influent cDNA template yielding positive EnV detection, visualized by performing gel electrophoresis after PCR amplification

40
Sewage validation
Primer set EQ-1/EQ-2’s optimized PCR conditions were
confirmed using urban wastewater retrieved from the Sand
Island Wastewater Treatment Plant, resulting in positive
EnV detection bands of the expected size (142 bp) at all
three treatment stages tested (Figure 1).
PCR product sequencing and analysis
Sequencing and BLAST analysis from selected EnV-
positive sewage samples revealed high sequence homology
with a variety of EnV strains listed in the NCBI database,
as is to be expected when using a primer set broadly
reactive for all enterovirus types. PCR sequencing results
will be examined in greater detail in the following
chapters.

41
Figure 1. Agarose gel depicting enterovirus detection from urban sewage. Amplified with primer set EQ-1/EQ-2. Detection from 100 mL of raw influent, post-primary clarification/pre-UV disinfection, and post-disinfection/effluent treatment stages. M = 50bp DNA ladder. (-) = no template control.

42
DISCUSSION
Reported here is the development of a rapid, user-
friendly method for the effective concentration and
detection of enteroviruses from Hawaiian environmental
waters. As previously discussed, because reliance on
bacterial indicators fails to reflect the presence of
potentially problematic viral pathogens, a need for
alternative monitoring parameters exists. EnV has already
been discussed as a potential alternative indicator of
microbial water quality; the highly optimized detection
protocol established and validated here is a substantial
step toward making this goal a reality.
By using urban wastewater as our nucleic acid source
for protocol optimization, as opposed to a single clinical
sample, primer efficacy was optimized for a broad genotypic
range of all human enteroviruses. As enteric viruses are
known to exhibit resistance to common wastewater treatment
practices, it was no surprise that EnV was detected at all
three treatment stages tested, including the effluent that
is discharged into the open ocean.
By comparing detection efficiencies of presently
available primer sets in a side-by-side manner, we were
able to establish EQ-1/EQ-2 to be a fine-tuned, and highly
sensitive protocol for the effective detection of human

43
enteroviruses. Under the described conditions, this
optimized protocol is 103- to 107-fold more sensitive than
all other protocols tested, suggesting its suitability to
detect viral pathogens present in water environments at low
concentrations.
The establishment of a highly sensitive EnV detection
reported here represents an extremely useful tool for
environmental virologists and is an important stepping-
stone leading toward the concrete establishment of
alternative model systems for water quality monitoring.

44
REFERENCES
Abbaszadegan M, Huber MS, Gerba CP, Pepper IL (1993) Detection of enteroviruses in groundwater with the polymerase chain reaction. Appl Environ Microbiol 59: 1318-1324 Bessaud M, Jegouic S, Joffret ML, Barge C, Balanant J et al (2008) Characterization of the genome of human enteroviruses: Design of generic primers for amplification and sequencing of different regions of the viral genome. J Virol Methods 149: 277-284 Chung H, Jaykus L, Sobsey MD (1996) Detection of human enteric viruses in oysters by in vivo and in vitro amplification of nucleic acids. Appl Environ Microbiol 62: 3772-3778 Dierssen U, Rehren F, Henke-Gendo C, Harste G, Heim A (1998) Rapid routine detection of enterovirus RNA in cerebrospinal fluid by a one-step real-time RT-PCR assay. J Clin Virol 42: 58-64 Fuhrman JA, Liang X, Noble RT (2005) Rapid detection of enteroviruses in small volumes of natural waters by real-time quantitative reverse transcriptase PCR. Appl Environ Microbiol 71: 4523-4530 Gomara MI, Megson B, Gray J (2006) Molecular detection and characterization of human enteroviruses directly from clinical samples using RT-PCR and DNA sequencing. J Med Virol 78: 243-253 Hot D, Legeay O, Jacques J, Gantzer C, Caudrelier, Y et al (2003) Detection of somatic phages, infectious enteroviruses and enterovirus genomes as indicators of human enteric viral pollution in surface water. Water Res 37: 4703-710 Hymas WC, Aldous WK, Taggart EW, Stevenson JB, Hillyard DR (2008) Description and validation of a novel real-time RT-PCR enterovirus assay. Clin Chem 54: 406-413 Jofre J and Blanch AR (2010) Feasibility of methods based on nucleic acid amplification techniques to fulfill the requirements for microbiological analysis of water quality. J of Appl Microbiol 109: 1853-1867

45
Jothikumar N, Sobsey MD, Cromeans TL (2010) Development of an RNA extraction protocol for detection of waterborne viruses by reverse transcriptase quantitative PCR (RT-qPCR). J Virol Methods 169: 8-12 Kreader CA (1996) Relief of amplification inhibition in PCR with bovine serum albumin or T4 gene 32 protein. Appl Environ Micriobiol 62:1102-1106 Lees DN, Henshilwood K, Dore WL (1994) Development of a method for detection of enteroviruses in shellfish by PCR with poliovirus as a model. Appl Environ Microbiol 60: 2999-3005 Pond, Kathy (2005) Water recreation and disease - Plausibility of associated infections: acute effects, sequelae and mortality. World Health Organization Puig M, Jofre J, Lucena F, Allard A, Wadell G et al (1994) Detection of adenoviruses and enteroviruses in polluted waters by nested PCR amplification. Appl Environ Microbiol 60: 2963-2970 Reynolds KA, Roll K, Fujioka RS, Gerba CP, Pepper IL (1998) Incidence of enteroviruses in Mamala Bay, Hawaii using cell culture and direct polymerase chain reaction methodologies. Can J Microbiol 44: 598-604 Tan EL, Chow VTK, Kumarasinghe G, Lin RTP, MacKay IM et al (2006) Specific detection of enterovirus 71 directly from clinical specimens using real-time RT-PCR hybridization probe assay. Mol Cell Probe 20: 135-140 Tong H, Lu Y (2011) Effective detection of human adenovirus in Hawaiian waters using enhanced PCR methods. Virol J 8: 57 Tong H, Connell C, Boehm AB, Lu Y (2011) Effective detection of human noroviruses in Hawaiian waters using enhanced RT-PCR methods. Water Research 45(18):5837-5848 U.S. Environmental Protection Agency (2007) EPA’s tentative decision on the renewal of CWA 301(h) variance for the Sand Island Wastewater Treatment Plant. Tentative Decision Document: Fact Sheet.

46
Zhang C, Wang X, Liu, Y, Peng D (2010) Simultaneous detection of enteroviruses from surface waters by real-time RT-PCR with universal primers. J Environ Sci 22: 1261-1266 Zoll GJ, Melchers JG, Kopecka H, Jambroes G, Van Der Poel HJ et al (1991) General primer-mediated polymerase chain reaction for detection of enteroviruses: application for diagnostic routine and persistent infections. J Clin Virol 30: 160-165

47
CHAPTER 3
HUMAN ENTEROVIRUS OCCURRENCE IN HAWAIIAN ENVIRONMENTAL WATERS
INTRODUCTION
In tropical regions such as Hawaii, human enteric
viruses, especially enteroviruses, are isolated throughout
the year (Fong and Lipp 2005). This is of significant
concern in a state where residents and tourists alike enjoy
year-round recreational activities in the local coastal
waters. Once a highly optimized assay for enhanced EnV
detection had been developed and validated (Chapter 2), it
was applied to an environmental surveillance study aimed at
evaluating the occurrence of EnV in Hawaiian waters. 22
recreational bodies of water were selected as sampling
sites. Coastal and freshwater sites were included in the
study, all of which receive, to varying degrees,
considerable human activity, including swimming,
snorkeling, diving, surfing, kayaking, canoeing, boating,
and fishing.
Because a diversity of viral strains are known to
exist in environmental samples (Loisy 2004), DNA sequencing
of PCR-positive EnV amplicons from several sampling
locations was conducted.

48
MATERIALS AND METHODS
Environmental water sample collection
Between June 2010 and October 2011, twenty-two surface
water samples were collected from various marine and
freshwater sites around the island of Oahu (Figure 2).
Marine sites include Sand Island State Recreational Area,
Kailua Bay, Waikiki Beach, Pokai Bay, Maunalua Bay, Kualoa
Regional Park, West Loch Community Shoreline Park, Kahala
Beach, and the beach parks of Ala Moana, Diamond Head,
Maili, Waialae, Kaiaka Bay, Kahana Bay, Ko Olina (Lagoons 3
and 4), Bellows Field, and Punalu’u. Freshwater sites
include Wahiawa Reservoir, Manoa Stream, and Kaelepulu
Stream. The sample collected from Ala Wai Canal was
brackish. 2-L samples were collected in sterile,
polypropylene containers and transported on ice to the
laboratory for immediate processing. A 2-L field blank
consisting of double-distilled H2O was prepared as a
negative control. A positive control was prepared by
spiking 2-L of seawater from Diamond Head Beach Park with
100 ml EnV-positive wastewater influent.

49
Figure 2. Geographic locations of environmental water sampling sites on Oahu, HI.

50
Sample concentration, nucleic acid extraction, and RT-PCR
Environmental samples were processed using the
filtration-based method described previously in Chapter 2.
Prior to filtration, MgCl2 solution was mixed into
freshwater samples to reach a final concentration of 25 mM.
2 L of environmental water samples were filtered through
0.45-µM pore size, type HA membranes (Millipore Corporation,
MA) on a filtration manifold under vacuum. For three water
samples with high sediment content, collected from Bellows
Field Beach Park, West Loch Community Shoreline Park, and
Kaelepulu Stream, filters became clogged before 2 L were
able to pass; therefore, smaller volumes of 0.80 L, 0.80 L,
and 0.50 L were passed, respectively. Nucleic acids were
extracted from the recovered membranes using the PowerWater
RNA Isolation Kit, supplied by MoBio Laboratories, CA,
according to a modified protocol designed for separate
extraction of both RNA and DNA. The DNase I digestion step
in the original protocol (steps 21-23) was skipped, and a
60 µL eluent containing both DNA and RNA was obtained. 15 µL
of the eluent mixture was aliquot to serve as the DNA
template for subsequent PCR. The remaining 45 µL eluent was
combined with 5 µL of 10X DNase I buffer (MoBio
laboratories, CA) and 3 µL of DNase I (MoBio laboratories,
CA) and incubated at room temperature for 20 min. DNase I

51
was heat-inactivated by incubating the reaction at 75°C for
5 min yielding pure RNA. The nucleic acid samples were
stored at -80°C until future usage.
Seven microliters of RNA extracted from each sample
was used as template for RT-PCR, performed with the DyNAmo
cDNA synthesis kit (New England Biolabs, NEB, MA) according
to the manufacturer’s instructions. Random hexamers were
used as primers.
Molecular amplification via PCR
Primer set EQ-1/EQ-2, selected as the optimal
candidate for surveillance of EnV presence in the
environment, was used to test the twenty-two environmental
water samples for EnV contamination. PCR was performed as
described in Chapter 2, using the newly-optimized PCR
conditions specified in Table 3. Results were visualized
via gel electrophoresis as previously described.
E. coli amplification as internal control
It is well known that environmental water samples
contain inhibitory compounds such as minerals, organic
matter, humic and fulvic acids, tannins, and biomass (Jofre
and Blanch 2010). If inefficiently removed during sampling
processing, the presence of these compounds can negatively

52
affect downstream molecular analysis (Shieh 1995,
Parshionikar 2004). In order to assess nucleic acid
extraction efficiency and inhibitor removal during sample
processing, DNA extracted from water samples was tested for
the presence of E. coli, which is known to grow naturally
in the Hawaiian environment and is expected to be readily
detectable in all samples (Byappanahalli 2004). In each 25
µL reaction, 3 µL sample DNA were added to 22 µL PCR mixture
containing 1X Taq (Mg2+ free) reaction buffer, 2.5 mM MgCl2
solution, 200 nM dNTP mixture, 0.1 μg/μL BSA, 400 nM of each
primer (URL301: TGTTACGTCCTGTAGAAAGCCC, URR-432:
AAAACTGCCTGGCACAGCAATT) (Bej 2001), and 2 units of Taq
polymerase. The amplification cycle consisted of an initial
5 min. denaturation at 94°C, followed by 35 cycles of 30
sec. denaturation at 94°C, 30 sec. annealing at 60°C, and 30
sec. extension at 72°C, completed by a final 5 min.
extension at 72°C. Results were visualized via gel
electrophoresis, performed as previously described.
PCR product sequencing and analysis
In order to confirm true EnV detection and identify
enteroviral strains present in the Hawaiian environment,
selected positive DNA fragments amplified by primer set EQ-
1/EQ-2 from environmental water samples were subjected to

53
DNA sequencing. DNA bands were excised, recovered, and
cloned into pCR®2.1-TOPO® vectors as described for EnV
amplicons detected in sewage (Chapter 2). 5 environmental
clones from 5 positive sampling sites (Manoa Stream, Pokai
Bay, Kaiaka Beach Park, Waikiki Beach, and Wahiawa
Reservoir) were submitted to ASGPB for DNA sequencing.
Resulting genomic sequences were aligned and compared with
available EnV sequences listed in the NCBI databank using
BLAST.

54
RESULTS
Enterovirus detection in environmental water samples
Environmental screening indicated that eleven of the
twenty-two sample sites contained EnV contamination,
including Diamond Head Beach Park, Pokai Bay, Kailua Bay,
Waikiki Beach, Kaiaka Bay Beach Park, Wahiawa Reservoir,
Manoa Stream, Ala Moana Beach Park, Ko Olina Beach Park
Lagoon 3, Kahala Beach, and Punalu’u Beach Park (Table 4).

55
Table 4. Enterovirus detection in Hawaiian environmental waters
Map#a Site Condition EnV detection
1 Kaiaka Bay Beach Park Seawater +
2 Punalu’u Beach Park Seawater +
3 Wahiawa Reservoir Freshwater +
4 Kahana Bay Beach Park Seawater −
5 Kualoa Regional Park Seawater -
6 Kailua Bay Seawater +
7 Kaelepulu Stream Freshwater −
8 Bellows Field Beach Park Seawater −
9 Maunalua Bay Seawater −
10 Waialae Beach Park Seawater −
11 Kahala Beach Seawater +
12 Diamond Head Beach Park Seawater +
13 Manoa Stream Freshwater +
14 Ala Wai Canal Brackish −
15 Waikiki Beach Seawater +
16 Ala Moana Beach Park Seawater +
17 Sand Island State Recreational Area Seawater −
18 West Loch Shoreline Park Seawater −
19 Ko Olina Beach Park Lagoon 4 Seawater −
20 Ko Olina Beach Park Lagoon 3 Seawater +
21 Maili Beach Park Seawater −
22 Pokai Bay Seawater +
Field Blank ddH2O −
Spike control Seawater + sewage +
aSee Figure 2

56
E. coli detection as internal control
E. coli was detected in all water samples tested,
indicating efficient nucleic acid extraction and inhibitor
removal during sample processing. This finding supports the
notion that negative detection of EnV at several sample
sites is truly negative, as opposed to being due to
unsatisfactory nucleic extraction and/or inhibitor effects.
PCR product sequencing and analysis
Sequencing and BLAST analysis from selected EnV-
positive water samples revealed high sequence homology with
a variety of EnV strains listed in the NCBI database, as
expected when using a primer set broadly reactive for all
enterovirus types. Of the 13 sequenced EnV PCR products
(including amplicons from sewage and environmental
samples), 9 were identified as human coxsackie A/B viruses
(including human enterovirus 90), causative agents of
herpangina, meningitis, fever, respiratory disease, hand-
foot-and-mouth disease, myocarditis, heart anomalies,
thrush, pleurodynia, and diabetes (Bosch 1998). Also
detected were human enterovirus 68, associated with
respiratory illness (Oberste 2004) and 2 human echoviruses,
linked to meningitis, fever, respiratory disease, thrush,
gastroenteritis, and severe neonatal infections (Piraino

57
1982). EnV sequence alignment and BLAST analysis may be
further examined in Figure 3.
Figure 3. Nucleotide sequence analysis of EnV isolated from wastewater (multiple clones) and environmental water samples. (A) Sequence alignment of 142bp fragments amplified by primer set EQ-1/EQ-2. Dots indicate homology with sewage isolate #1. (B) Closest BLAST match (including E value and percentage identity) of sequenced PCR products with EnV strains listed in the NCBI database.
1 ACATGGTGTG AAGAGTCTAT TGAGCTAATT GGTAGTCCTC CGGCCCCTGA ATGCGGCTAA TCCTAACTGC 70 Sewage #1
.......... .......... .......C.. .AG....... .......... .......... .......CA. Sewage #2 .......... .......... .......... ....A..... .......... .......... .......... Sewage #3 .......... .......... .......CC. .AG....... .......... .......... ...C...CAC Sewage #4 .......... .......... .......G.. A......... .......... .......... .......... Sewage #5 .......... .......... .......CA. AAGA...... .......... .......... ...C...CAT Sewage #6 .......... .......... .......CC. .AG....... .......... .......... ...C...CA. Sewage #7 .......AGA .......... .......... .A..A..... .......... ........-. .......... Sewage #8 .......... .......... .......G.. A......... .......... .......... .......... H2O: Manoa Stream .......... .......... .......CAA .AG....... .......... .......... .......... H2O: Pokai Bay .......... .......... .......CA. AAG....... .......... .......... ...C...CAT H2O: Kaiaka Bay .......... .......... .......G.. .......... .......... .......... ...C...CAT H2O: Waikiki Beach .......... .......... .......G.. .......... .......... .......... .......... H2O: Wahiawa Reservoir
71 AGAGCGCGTA CCCTCAACCC AGGGGGCGGC GCGTCGTAAT GGGTAACTCT GCAGCGGAAC CGACTACTTT GG 142 Sewage #1 G....AG..G .T.A...A.. ..T...T... TT.......C .C.C..G... .TG....... .......... .. Sewage #2 G....ACAC. ...A..CA.. .......A.T .T.......C ...C...... .......... .......... .. Sewage #3 G....AA..G .T.A...A.. ..T...TA.. TT.......C .C.C..G... .TG....... .......... .. Sewage #4 .......... .......... .......... .......... .......... .......... .......... .. Sewage #5 G....AA..G A..A...T.. ..T..TTCT. TT.......C .C.C..G..C .TG....... .......... .. Sewage #6 G....AA..G .T.A...A.. ..T...TA.. TT.......C .C.C..G... .TG....... .......... .. Sewage #7 G....AGA.. ...A..CA.. ..T....A.T CT........ ...C...... .......... .......... .. Sewage #8 G......... .......... .......... .......... .......... .......... .......... .. H2O: Manoa Stream G....AGA.. ...A..CA.. ..T....A.T CT........ ...C...... .......... .......... .. H2O: Pokai Bay G....AA..G A.TA...T.. ..T..TTCT. TT.......C .C.C..G..C .TG....... .......... .. H2O: Kaiaka Bay G....AA..G A..A...T.. ..T..TTCT. TT.......C .C.C..G..C .TG....... .......... .. H2O: Waikiki Beach G....AGA.. ..TA.G.G.. ..T....A.T CT.......C ...C...... .......... .......... .. H2O: Wahiawa Reservoir
Sample Closest BLAST match (Accession #, Name) E value Max ident Sewage #1 GU236101.1 Human coxsackievirus A16 isolate 00.143.2668 5e-59 96% Sewage #2 DQ995634.1 Human coxsackievirus A11 strain BAN01-10589 1e-60 97% Sewage #3 GQ126860.1 Human coxsackievirus B5 1e-60 97% Sewage #4 AB192877.1 Human enterovirus 90 5e-59 96% Sewage #5 HQ423141.1 Human coxsackievirus A16 strain KMM/08 1e-60 97% Sewage #6 AF465511.1 Human coxsackievirus A13 strain Flores 5e-54 94% Sewage #7 AB192877.1 Human enterovirus 90 5e-59 96% Sewage #8 GU236215.1 Human echovirus 11 isolate 39351.82 8e-57 95% H2O: Manoa Stream HQ423141.1 Human coxsackievirus A16 strain KMM/08 2e-62 97% H2O: Pokai Bay AJ579633.1 Human echovirus 11 isolate JP-979/89 5e-59 96% H2O: Kaiaka Bay AF465511.1 Human coxsackievirus A13 strain Flores 2e-52 93% H2O: Waikiki Beach AY062274.1 Human enterovirus 68 strain VR-561 2e-47 91% H2O: Wahiawa Reservoir GU236263.1 Human echovirus 24 isolate 23927 2e-62 97%
A
B

58
DISCUSSION
Establishment of highly sensitive EnV-detection assay
allowed a survey study of 22 recreational water sites
around the island of Oahu, 11 of which tested positive for
enterovirus, indicating fecal pollution in a significant
portion of Hawaii’s surface water. It is worthy to note
that this is the first report of using an effective
molecular detection method to demonstrate a relatively high
occurrence of enterovirus in Hawaiian recreational waters.
The utilization of this optimized protocol to rapidly and
reliably screen multiple water samples from across the
island of Oahu represents a powerful research tool of
important public health significance.
It should be noted that public health implications are
limited based solely on PCR-positive results (Choi 2005,
Fout 2003). While molecular detection assays can be useful
for indicating fecal contamination in an area, they do not
distinguish between the presence of truncated genomic
fragments or complete, viable, and infectious virus
particles (Boehm 2003, Rodriguez 2009). Therefore,
infectivity assays based on the observance of viral-induced
CPE in cell culture are important in order to make valid
determinations of health risks (Fong 2005). Current
research in this laboratory is directed at assessing the

59
infectivity (defined as of the ability of a virus to enter
a host cell and utilize its resources to produce infectious
virus particles, Rodriguez 2009) of the enteric viruses for
which permissive cell lines are available (AdV + EnV).
Nevertheless, whether or not the viruses are infectious,
their positive molecular detection is a useful indicator of
fecal contamination in Hawaii’s waters. Even if the viruses
themselves are non-infectious, they indicate a current or
recent source of fecal pollution that has potential to
introduce other pathogens of concern into local
recreational waters.
Of notable practical significance is that comparable
optimization studies in our laboratory have produced
similar protocols for the efficient environmental detection
of other human enteric viruses, including adenovirus (Tong
and Lu 2011), norovirus genogroups I and II (Tong et al
2011), and F-specific RNA coliphage (Tong 2011). When
combined with the EnV detection protocol established here,
these procedures comprise a powerful array for monitoring
and comparing fecal pollution levels. The ability to
reliably screen environmental waters for the presence of
multiple strains of enteric viruses is a highly desirable
research tool, facilitating a thorough investigation of
potentially contaminated recreational waters. The

60
relatively simple protocols using well-established,
conventional RT-PCR procedures are adoptable by a broad
range of environmental health agencies, for which more
advanced equipment and techniques (e.g. real-time PCR) may
be unavailable.
Although the described methods are powerful
supplements to aid microbial water quality monitoring,
public health implications are limited without conclusive
infectivity data. Risk assessment at any particular
recreational site cannot be based solely on PCR-detected
EnV presence or absence from a single sample collection.
Additionally, the present study is limited to the detection
of EnV strains present in Hawaii, which may not be a
complete representation of the EnV composition present
elsewhere. For serious consideration as a valid and
established alternative monitoring system, broader large-
scale trials, including additional sampling sites and
replicate samples from each site, will be necessary. Also,
comparisons with standardized bacterial surveillance
systems will contribute to a more thorough understanding of
water quality assessment.

61
REFERENCES
Bej AK, DiCesare JL, Haff L, Atlas RM (2001) Detection of Escherichia coli and Shigella spp. in water by using the polymerase chain reaction and gene probes for uid. Appl Environ Microbiol 57: 1013-1017 Boehm AB, Fuhrman JA, Mrse RD, Grant SB (2003) Tiered approach for identification of a human fecal pollution source at a recreational beach: Case study at Avalon Bay, Catalina Island, California. Environ Sci Technol 37: 673-680 Bosch A, Guix S, Sano D, Pinto RM (1998) New tools for the study and direct surveillance of viral pathogens in water. Curr Opin Biotechnol 19: 295-301 Byappanahalli M, Fujioka R (2004) Indigenous soil bacteria and low moisture may limit but allow faecal bacteria to multiply and become a minor population in tropical soils. Water Sci Technol 50: 27-32 Choi S, Jiang SC (2005) Real-time PCR quantification of human adenoviruses in urban rivers indicates genome prevalence but low infectivity. Appl Environ Microbiol 71: 7426-7433 Fong T, Lipp EK (2005) Enteric viruses of humans and animals in aquatic environments: health risks, detection, and potential water quality assessment tools. Microbiol Mol Bio. Rev 69: 357-371 Fout GS, Martinson BC, Moyer MWN, Dahling DR (2003) A multiplex reverse transcription-PCR method for detection of human enteric viruses in groundwater. Appl Environ Microbi 69: 3158-3164 Jofre J and Blanch AR (2010) Feasibility of methods based on nucleic acid amplification techniques to fulfill the requirements for microbiological analysis of water quality. J of Appl Microbiol 109: 1853-1867 Loisy F, Atmar RL, Guillon P, Cann PL, Pommepuy M, Le Guyader FS (2004) Real-time RT-PCR for norovirus screening in shellfish. J Virological Methods 123:1-7 Oberste MS, Maher K, Schnurr D, Flemister MR, Lovchik JC et al (2004) Enterovirus 68 is associated with respiratory

62
illness and shares biological features with both the enteroviruses and the rhinoviruses. J Gen Virol 85: 2577-2584 Parshionikar SU, Cashdollar J, Fout GS (2004) Development of homologous viral internal controls for use in RT-PCR assays of waterborne enteric viruses. J Virol Methods: 121:39-48 Piraino FF, Sedmak G, Raab K (1982) Echovirus 11 infections of newborns with mortality during the 1979 enterovirus season in Milwaukee, Wis. Public Health Rep 97: 346-353 Rodriguez RA, Pepper IL, Gerba CP (2009) Applicaton of PCR-based methods to assess the infectivity of enteric viruses in environmental samples. Appl Environ Microbiol 75: 297-307 Shieh YSC, Wait D, Tai L, Sobsey MD (1995) Methods to remove inhibitors in sewage and other fecal wastes for enterovirus detection by the polymerase chain reaction. J Virol Methods: 54:51-66 Tong H (2011) Development of effective detection of human enteric viruses in Hawaiian recreation waters. MS thesis, University of Hawaii at Manoa, Honolulu. 85 p. Tong H, Connell C, Boehm AB, Lu Y (2011) Effective detection of human noroviruses in Hawaiian waters using enhanced RT-PCR methods. Water Res 45: 5837-5848 Tong H, Lu Y (2011) Effective detection of human adenovirus in Hawaiian waters using enhanced PCR methods. Virol J 8: 57

63
CHAPTER 4
SHELLFISH-MEDIATED BIOACCUMULATION OF HUMAN ENTERIC VIRUSES IN OAHU’S MARINE ENVIRONMENT
INTRODUCTION
As addressed previously, the low concentration of
enteric viruses in environmental water samples can present
a major barrier to their effective utilization as
alternative water quality indicators. In addition to highly
optimized molecular detection assays (discussed in previous
chapters), efficient viral concentration techniques can
also help overcome this obstacle. The natural
bioconcentration phenomenon of marine bivalve shellfish
endemic to Oahu is explored here as a novel means of
assessing microbial water quality.
Bivalve molluscs are shellfish that have two shell
halves hinged together (Lees 2000). These stationary
animals obtain food by filter-feeding, meaning that they
process large volumes of water daily in order to capture
algae and other particulate matter for food (Hernroth 2002,
Karamoko 2005, Vilarino 2009, Bosch 2010). Research has
documented that the filter-feeding behavior of shellfish
can be affected by various environmental factors, including
turbidity, particulate chemical composition, pH,
temperature, and salinity (Asahina 2009). Other factors are

64
shellfish size and species, as larger animals are capable
of filtering more water and are thus potentially exposed to
greater numbers of pathogens in polluted waters (Nappier
2008).
As a result of filter-feeding, microbes present in
surrounding water will bioaccumulate within the internal
digestive tissues of the shellfish to a level higher than
that in the surrounding water (Kingsley 2001, Formiga-Cruz
2003, Jamieson 2005, Schultz 2007, Umesha 2008, Asahina
2009). According to Goblick et al, shellfish are able to
concentrate enteric viruses to levels up to 100-fold
greater than what exists in the nearby water (Goblick
2011). Following a contamination event, shellfish will
typically bioaccumulate viruses to detectable levels within
24-48 hours. In a study conducted by Asahina et al,
shellfish were placed into a tank containing seawater
seeded with a 10-6 dilution of a clinically positive
norovirus stool sample. Even at this high dilution, the
shellfish were capable of bioaccumulating the virus to a
readily detectable level (Asahina 2009). Additionally,
marine sediments, which are known to harbor enteric
viruses, may increase viral accumulation rates in shellfish
by acting as contamination sources in addition to the
overlying water (Landry 1983).

65
Shellfish are known to retain viruses longer than they
retain bacterial indicators commonly used for water quality
assessment (Dore 1995, Kingsley 2011, Goblick 2011).
Depuration, a natural process utilized by filter-feeders,
acts to expel substances that are inessential to the
animals. Enteric viruses are purged more slowly than most
bacteria, and research has shown that depuration is an
ineffective method for the removal of viruses from
shellfish tissue (Richards 2010). In fact, viral retention
mechanisms may exist (Le Guyader 2006, Hansman 2007). In a
study conducted by Asahina et al, enterovirus was detected
in shellfish tissue via RT-PCR 12 days after a seeded tank
exposure event (Asahina 2009). Similarly, Nappier et al
allowed oysters exposed to various enteric viruses to
depurate and found that viruses persisted even after 29
days (Nappier 2008). Also, it has been suggested that
phagocytic processes in shellfish hemocytes, involving
lysosomal enzymes, toxic oxygen intermediates, and
antimicrobial peptides, are utilized to kill bacteria.
However, these processes have not been shown to affect
viruses; enteric viruses have actually been shown to
persist within phagocytic hemocytes of live bivalves
(Richards 2010, Kingsley 2010).

66
As a result of this natural concentration phenomenon,
bivalve shellfish may act as bioindicators of microbial
contamination in the waters they inhabit (Manso 2010). In
this way, these animals represent “a snapshot” of water
quality over the past days to weeks (Miller 2005).
Shellfish have been used for decades as bioindicators of
heavy metals, pesticides, and other toxins in the aquatic
environment, and they have been recognized in recent years
as potential bioindicators of bacteria, viruses, and
parasites of fecal origin (Miller 2005, Ismail 2006,
Sombrito 2007, Connell et al 2012). By subjecting nucleic
acids extracted from shellfish internal tissue to optimized
PCR assays developed by our laboratory, greatly
enhancedenteric virus detection efficiency has been
achieved. This novel shellfish-mediated bioaccumulation
phenomenon shows promising potential to aid in effective
surveillance of Hawaiian environmental waters.
Commonly found inhabiting Hawaii’s ocean environment
is the marine bivalve Isognomon spp. Ranging in size from
40-55 x 15-30 mm, this mollusk is usually found 1-2 meters
below the ocean surface, living underneath rocks and in
reef crevices of intertidal and subtidal habitats.
Isognomon spp. are typically short-lived (2 years or less)
and reach sexual maturity within their first year. They

67
have no commercial value and have received little attention
in the literature (Harper 1994, Asahina 2009).
For this study, Isognomon spp. are collected from
various beaches around the island of Oahu and nucleic acids
are extracted from internal digestive tissues. Highly
optimized RT-PCR/PCR protocols are then applied to test for
the presence of enterovirus, norovirus genogroups I and II,
adenovirus, and F-specific RNA coliphage. Resulting
information regarding the occurrence of various enteric
viruses in local marine waters is an extremely useful
resource to enhance water quality surveillance measures.

68
MATERIALS AND METHODS
Sample collection
Between July and December 2011, marine bivalves
Isognomon spp. were collected from reef crevices and from
underneath rocks at twelve various beaches around the
island of Oahu (Figure 4). Sampling site selections were
based on shellfish availability, recreational activity
levels, and potential sources of fecal pollution (including
both point and non-point sources). Several other sampling
sites were visited, including Kuilima Cove, Hukilau Beach
Park, Swanzy Beach Park, Ka’a’awa Beach Park, and various
sites along the Waianae coastline; however, either no
shellfish were found, or sampling conditions were too rough
at these locations. Whenever possible, samples were
harvested from a wide collection area in order to ensure
assessment of overall water quality at each particular site
(as opposed to collecting all samples from underneath a
single rock or ledge). Between 15 and 55 specimens were
collected from each site, depending on size and
availability.

69
Figure 4. Geographic locations of twelve sample collection sites included in shellfish-mediated bioaccumulation study.

70
Shellfish dissection + nucleic acid extraction
Following transport to the laboratory on ice,
shellfish were counted, measured, weighed, and shucked
using sterile No. 4 scalpels + No. 20 blades. Internal
digestive tissues were removed using sterile forceps and
separated into 1.0 – 2.0 g aliquots, from which nucleic
acids were extracted using the MoBio PowerSoil RNA
Isolation Kit + DNA Elution Accessory Kit (MoBio
Laboratories, CA), according to the manufacturer’s
instructions. This kit, designed for the isolation of
nucleic acids from soil samples, was selected for its
powerful removal of inhibitory substances, yielding high
quality, biologically intact, RT-PCR amplifiable RNA (MoBio
Laboratories, PowerSoil instruction manual). Extracted RNA
was subject to DNase treatment using the RTS DNase Kit
(MoBio Laboratories, CA), according to the manufacturer’s
instructions. DNA and DNase-treated RNA were stored at -80°C
until future usage. Also, during animal dissection, 0.5 –
1.0 g of tissue from each animal was aliquoted into 1.5 ml
Eppendorf tubes and stored at -80°C to be utilized in future
infectivity studies.

71
Simultaneous water sample processing
At each of the 12 sites where shellfish were
collected, a 2 L water sample was also collected in order
to compare the sensitivity between these two viral
detection methods. Water samples were collected and
processed using the same membrane filtration and nucleic
acid extraction methods described in previous chapters. The
entire volume of 2 liters was filtered from each sample
site. Extracted nucleic acids were stored at -80°C until
future usage.
RT-PCR/PCR
In order to generate cDNA to test for the presence of
RNA viruses EnV, NoVI, NoVII, and F+, RT-PCR was performed
(as described in previous chapters) using RNA extracted
from shellfish and corresponding water samples as template.
To test for the presence of AdV (DNA virus), extracted DNA
was utilized directly as the PCR template. PCR for all five
enteric virus subsets was performed according to their
individual optimized conditions (Tables 5 and 6). Details
of the primer sets employed may be found in Table 7.
Results were visualized by performing gel electrophoresis
as previously described.

72
Table 5. Optimized PCR reaction components for enteric virus detection
Virus Primer [Mg] (mM)
[Primer] (µM)
BSA (0.01 µg/µL)
AdV ADV-F
ADV-R 1.5 0.6 +
NoV GI QNIF4
NV1LCR 2.0 0.4 +
NoV GII COG2F
COG2R 2.0 0.8 +
EnV EQ-1
EQ-2 1.5 0.6 +
F+ OgorzalyIII F
OgorzalyIII R 2.0 0.8 +

73
Table 6. Optimized PCR cycle conditions for enteric virus detection
Step Temperature
Time
Initial denaturation 94°C 5 min
Denaturation 94°C 30 sec
Anneal 54°C (AdV)
58°C (EnV, NoV, F+) 30 sec
Extension 72°C 30 sec
GO TO: 2 40 cycles ---
Final extension 72°C 10 min
Hold 4°C ---

74
Table 7. Primer sets employed in bioaccumulation study
Virus Primer Sequence (5' -> 3') Size
NoV GI QNIF4 CGCTGGATGCGNTTCCAT 86 bp
NV1LCR CCTTAGACGCCATCATCATTTAC
NoV GII COG2F CARGARBCNATGTTYAGRTGGATGAG 97 bp
COG2R TCGACGCCATCTTCATTCACA
EnV EQ-1 ACATGGTGTGAAGAGTCTATTGAGCT 142 bp
EQ-2 CCAAAGTAGTCGGTTCCGC
F+ Ogorzaly III F CCGCGTGGGGTAAATCC 115 bp
Ogorzaly III R TTCTTACGATTCCGAGAAGGCT
AdV ADV-F GCCACGGTGGGGTTTCTAAACTT 131 bp
ADV-R GCCCCAGTGGTCTTACATGCACATC

75
E. coli amplification as internal control
As mentioned previously, it is well known that
environmental samples contain high levels of inhibitory
compounds. Shellfish are no exception; in fact, the
inhibitors present in shellfish tissue are often especially
problematic due to humic substances, glycogen, and other
properties of shellfish extracts (Lees 1994, Atmar 1995,
Schwab 2001, Loisy 2004, Kingsley 2011). In order to
efficiently detect viruses via PCR, nucleic acids must be
as pure as possible. This is especially important when
dealing with RNA viruses, as the additional reverse
transcriptase step often a major limiting factor due to the
enzyme’s susceptibility to interfering and inhibitory
substances (Arnal 1999). Therefore, just as E. coli was
used as an internal amplification control for the
environmental samples discussed in Chapter 3, DNA extracted
from shellfish tissue was also tested for the presence E.
coli. PCR conditions and gel electrophoresis were performed
as described in the previous chapter.
PCR product sequencing and analysis
In order to confirm true enteric virus detection and
identify specific strains present in the Hawaiian
environment, selected positive amplicons detected from

76
shellfish and corresponding water samples were subjected to
DNA sequencing. DNA bands were excised, recovered, and
submitted to ASGPB, as described previously, for direct DNA
sequencing. Resulting genomic sequences were compared with
available enteric virus sequences listed in the NCBI
databank using BLAST.

77
RESULTS
Enteric virus detection in shellfish and corresponding
water samples
Detection results of this bioaccumulation study are
summarized in Tables 8 (shellfish), 9 (corresponding
coastal water samples), and 10 (correlation between EnV
detection in shellfish and water samples). The most
commonly detected virus in shellfish was human norovirus
genogroup I, for which 8 of 12 beach sites and 33 of 64
total sample groups (51.6%). Human enterovirus was also
quite prevalent, detected in shellfish from 8 beach sites
and in 30 of the 64 total sample groups (46.9%).
Contamination levels of the remaining three viruses were
lower but still quite significant. Adenovirus was detected
in shellfish from 7 beach sites and in 21 of the total
sample groups (32.8%). Norovirus II was detected in
shellfish from 4 beach sites and in 15 of the total sample
groups (16.3%). F-specific RNA coliphage was detected at 6
beach sites and in 11 of the total sample groups (17.2%).
Based on detection results from the twelve
corresponding water samples, NoVI was most prevalent
(positive detection at 7 of 12 sites). EnV was detected at
4 of 12 sites, followed by AdV and NoVII (3 and 2 positive

78
sites, respectively). F+ was not detected in any of the
twelve water samples.
E. coli as internal control
All sample groups tested positive for E. coli,
indicating efficient nucleic acid extraction and inhibitor
removal.
PCR product sequencing
A variety of enteric virus strains were detected from
both shellfish and coastal water samples. See Table 11 for
details.

79
Table 8. Enteric virus detection in shellfish
Site # Collected
# Sample Groups
# (+) Groups
AdV EnV NoVI NoVII F+
Ala Moana 15 3 3 3 3 - -
Kahala 30 5 - 3 2 2 1
Kahana Bay 40 4 - - 4 2 -
Kawaikui 21 4 1 - 4 - 1
Ko Olina Lagoon 3 18 5 3 5 5 3 2
Ko Olina Lagoon 4 48 4 - - 4 - -
Kualoa Park 55 5 3 4 5 - 1
Magic Island 28 7 - 3 - - 4
Punalu’u 48 6 5 5 - - 2
Sand Island 26 6 1 - 6 - -
Waialae (Resort) 36 9 - 2 - 8 -
Waialae Beach Park 20 6 5 5 - - -
TOTAL 385 64 21 30 33 15 11

80
Table 9. Enteric virus detection in corresponding water samples
Site AdV EnV NoVI NoVII F+
Ala Moana + - + - -
Kahala - + - - -
Kahana Bay - - + + -
Kawaikui - - + - -
Ko Olina Lagoon 3 + + + + -
Ko Olina Lagoon 4 - - + - -
Kualoa Park - - + - -
Magic Island - + - - -
Punalu’u - + - - -
Sand Island - - + - -
Waialae (Resort) - - - - -
Waialae Beach Park + - - - -
12 SITES TOTAL 3 4 7 2 0

81
Table 10. Enterovirus detection correlation between shellfish and water samples
Site EnV detection
Water Shellfish
# (+) of Total # Tested
Ala Moana Beach Park - 15 of 15
Kahala Beach + 18 of 30
Kahana Bay Beach Park − 0 of 40
Kawaikui Beach Park - 0 of 21
Ko Olina Beach Park Lagoon 3 + 18 of 18
Ko Olina Beach Park Lagoon 4 − 0 of 48
Kualoa Regional Park - 44 of 55
Magic Island + 12 of 28
Punalu’u Beach Park + 40 of 48
Sand Island State Recreational Area - 0 of 26
Waialae (Resort) - 8 of 36
Waialae Beach Park − 17 of 20

82
Selected gel images are displayed below (Figures 5-8).
Figure 5. Agarose gel depicting adenovirus detection from shellfish collected at Kualoa Park. Amplified with primer set ADV-F/ADV-R. (+) = TOPO plasmid containing fragment of interest confirmed via DNA sequencing. (-) = no template control.
Figure 6. Agarose gel depicting enterovirus detection from samples collected at Ko Olina Lagoon 3. Amplified with primer set EQ-1/EQ-2. (+) = TOPO plasmid containing fragment of interest confirmed via DNA sequencing. (-) = no template control.

83
Figure 7. Agarose gel depicting norovirus genogroup I detection from samples collected at Ko Olina Lagoon 3. Amplified with primer set QNIF4/NV1LCR. (+) = TOPO plasmid containing fragment of interest confirmed via DNA sequencing. (-) = no template control.
Figure 8. Agarose gel depicting enterovirus detection from samples collected at Kahala Beach. Amplified with primer set EQ-1/EQ-2. (+) = TOPO plasmid containing fragment of interest confirmed via DNA sequencing. (-) = no template control.

84
Table 11. Enteric virus strains detected in Hawaiian marine environment
a) Enterovirus
Strain Site of Detection
(if from H2O: italics)
Human coxsachievirus A16 Kahala Resort
Ko Olina Lagoon 3
Human coxsachievirus A22
Punalu’u
Kualoa
Kahala Beach
Kahala Beach
Human echovirus 11 Magic Island
Human echovirus 30 Ko Olina Lagoon 3
Human enterovirus C Kahala Beach
Human enterovirus 71 Ko Olina Lagoon 3
b) Norovirus genogroup I
Strain Site of Detection
(if from H2O: italics)
Human norovirus Saitama Ko Olina Lagoon 4
Norovirus Hu/GI/2005/8338/Chelyabinsk Kahana Bay
Norovirus genogroup 1 strain 1336 Kahana Bay
Norovirus Hu/GI.1/P7587/2007/Stromstad Sand Island
Norovirus Hu/GI.1/P7-587 Kualoa
Norovirus Hu/GI/Guangxi/DL131-2/2010 Kualoa Ko Olina Lagoon 4
Norovirus Hu/GI.3/C13/Bonaberi/2009 Ko Olina Lagoon 3

85
c) Norovirus genogroup II
Strain Site of Detection
(if from H2O: italics)
Norovirus Hu/GII/Maizuru/7734/2007/JPN Kahala Beach
Norovirus Hu/GII/Beijing/589/2008 Ko Olina Lagoon 3 Ko Olina Lagoon 3
d) F-specific RNA coliphage
Strain Site of Detection (if from H2O: italics)
Enterobacteria phage Qbeta
Magic Island Kualoa Park Kahala Beach
Ko Olina Lagoon 3
e) Adenovirus
Strain Site of Detection (if from H2O: italics)
Human adenovirus 41
Punalu’u
Sand Island
Kualoa
Ko Olina Lagoon 3
Ko Olina Lagoon 3
Human adenovirus B isolate DA0630 Ala Moana

86
DISCUSSION
The novel enteric virus detection method employed here
has interesting implications for enhanced environmental
surveillance. The substantial number and wide variety of
enteric viruses detected in shellfish collected from Oahu’s
coastal waters suggests that these animals are indeed
useful biosentinels of microbial water quality.
Shellfish dissection, nucleic acid extraction, and
subsequent PCR analysis revealed positive detection of at
least one enteric virus subset at every beach site tested,
suggesting fecal pollution in a significant portion of
Hawaii’s marine environment. It is interesting to note that
NoVI was the most commonly detected enteric virus in
shellfish, followed by EnV and AdV. Corresponding water
samples revealed a similar hierarchy of enteric virus
prevalence in the Hawaiian marine environment. These
results suggest widespread occurrence of these two enteric
virus subtypes in our local waters. NoVII was detected at a
lesser extent, due perhaps to lower general occurrence in
the state of Hawaii, or decreased environmental persistence
as compared to other enteric viruses. It is also possible
that, compared to other types of enteric virus, NoVII is
not preferentially bioaccumulated within shellfish tissue.
F+ was also detected at a low level, most likely due to the

87
fact that this virus is thought to restrictively indicate
point source contamination (direct sewage), but not other
various non-point sources. Another possibility is that,
despite precise PCR condition optimization for individual
primer sets, certain detection protocols are simply more or
less sensitive than others.
Due to the bioconcentration phenomenon of shellfish,
it is expected that the utilization of these filter-feeders
proves to be a more sensitive method for viral detection
than testing water samples directly. Findings from this
study confirm this expectation. Whenever a coastal water
sample tested positive for a particular virus, shellfish
collected from the same location also tested positive.
Conversely, not all viruses detected in shellfish were
detected in water samples tested directly from the same
site. In fact, for each of the 5 of the viruses included in
this study, the number of contaminated sites determined
through shellfish processing exceeds that of direct water
sampling.
To compare enteric virus detection in shellfish versus
corresponding water samples more deeply, detection
correlation results are compared site-by-site for EnV
specifically, as shown in Table 10. Shellfish dissection,
nucleic acid extraction, and subsequent PCR analysis

88
revealed positive EnV detection in specimens from eight of
twelve coastal locations. From the four sites where EnV was
not detected in shellfish, corresponding water samples also
tested negative; this correlation suggests that these sites
are free of EnV contamination. Four of the eight sites
where EnV was detected in shellfish, including Kahala
Beach, Magic Island, Ko Olina Lagoon 3, and Punalu’u Beach
Park, were also shown to contain EnV through membrane
filtration of water samples. This positive correlation
strongly suggests fecal pollution at these four beaches. It
is interesting to note that from the remaining four sites,
where shellfish were shown to contain EnV (Ala Moana Beach
Park, Kualoa Regional Park, Waialae-Resort, and Waialae
Beach Park), water tested EnV-negative. Detection analysis
for all other viral types included in the study yields
similar correlation results. These findings strongly
contribute to the notion that using shellfish as sentinels
of water quality is a more sensitive monitoring method than
testing water directly.
However, although using shellfish as sentinels of
water quality yields enhanced detection sensitivity, this
method does require additional processing time and effort.
Prior to nucleic acid extraction, an adequate number of
shellfish must be acquired and dissected, which is a much

89
lengthier process than the simple membrane filtration
procedure utilized when testing direct water samples.
Therefore, shellfish processing may be suitable for more
in-depth water quality studies, while the ease and
simplicity of direct water sample collection may be more
practical for routine recreational water monitoring.
PCR product sequencing is a useful research component
for effective water quality monitoring, as it provides
valuable insight into the various enteric viral strains
that contaminate our local waters. The diverse composition
of strains identified here confirms that our optimized
primer sets are broad enough to include a wide range of
members within each viral subset. Similar viral strains
were detected from shellfish and direct water samples,
indicating shellfish-mediated uptake of whichever viral
strains are present in the surrounding water. A wide
variety of EnV and NoVI were detected from multiple sites
around the island of Oahu. The most commonly detected AdV
was adenovirus 41, a critical etiological agent of viral
gastroenteritis in children (Xagoraraki 2007). The
seemingly lower prevalence of NoVII and F+ in Hawaiian
waters is reflected in corresponding DNA sequencing
results.

90
An additional research area of interest is the
identification of the origins of fecal matter in
recreational waters. It is known that major pollution
sources of concern include sewage discharge, storm water
runoff from streams and storm drains, and contamination
from bathers themselves (Pond 2005). All swimmers release
fecal organisms upon entering the water in a process called
bather shedding. Fecal accidents and diaper-aged children
are also of concern (Devine 2011). It has been demonstrated
that bathers particularly impact water quality in confined
waters with limited flow (Fattal 1991). High levels of
bacterial indicators have been shown to correlate with a
high number of visitors during periods of high recreational
use, such as weekends and the summertime (McDonald 2008).
Other potential contamination sources include input from
groundwater discharge, beach sand, and boats (Bosch 2010,
Viau 2011). Marinas are usually located in sheltered areas
with reduced water circulation, resulting in the
accumulation of boating waste. Also, boats in use may
dispose of waste improperly into recreational waters
(Devine 2011). In developed countries, because most point
sources of wastewater are well regulated, non-point sources
such as urban runoff during storms become main contributors
to surface water pollution (Viau 2011). In fact, the most

91
common route of fecal contamination in recreational waters
is following heavy rainfall, when increased runoff and
overflow exacerbate pre-existing causes of recreational
water pollution (Bosch 2010). 96% of Hawaii’s beach
advisories in 2010 were due to “brown water advisories”, or
rain (Dorfman 2011). Also, flooding over saturated soils
can result in larger sediment flow, resulting in increased
microbial counts in recreational waters (Devine 2011).
In the waters of Hawaii specifically, pollution source
tracking is a difficult task. Because water flow is
unidirectional in streams, fecal contamination will most
likely originate upstream. However, in the marine
environment, source tracking is more difficult due to the
continuous mixing of water (Miller 2005). Also, several
recreational bodies of water receive multiple inputs from
land sources, making accurate source targeting a
challenging task. For instance, Kailua Bay, receives inputs
from the Mokapu wastewater outfall, Kawainui Canal, and
Ka’elepulu Stream, all of which are potential sources of
contamination (Krock 1993). Several sites routinely test
positive for various fecal contaminants, yet the sources of
pollution remain unknown and ambiguous (Hunt, USGS).
Coastal waters receiving discharge from streams are
particularly susceptible to fecal contamination (Viau

92
2011), and numerous recreational beach sites in Hawaii
receive freshwater runoff. It is estimated that Kahana Bay,
located on the windward side of Oahu, receives 30 million
gallons of freshwater runoff per day (C&C of Honolulu).
Septic wastewater is suspected as another major influence,
but convincing data is currently unavailable.
Malfunctioning septic systems at just a few near-shore
properties can result in beachwater contamination
significant enough to require beach closures, and pathogens
from faulty inland septic systems may enter streams that
flow into recreational waters (Devine 2011). In 2007,
Kualoa Beach Park was closed for several months due to
bacterial levels seven times the acceptable standard, an
event most likely attributable to leaching from the park
restrooms (Hunt, USGS). Another likely contamination source
is open defecation of homeless populations inhabiting areas
around environmental waters. In a 2011 study conducted by
Viau et al, four streams with large homeless encampments
nearby tested positive for several types of enteric
viruses, including adenovirus, enterovirus, norovirus I,
and norovirus II (Viau 2011). The identification of fecal
pollution sources is important if preventative action is to
be taken in hopes of minimizing future contamination events
in our precious tropical waters.

93
It was originally hypothesized that the degree of
viral contamination at various Hawaiian beaches (as
detected by the bioaccumulation study) may be correlated
with various impact factors, such as nearby wastewater
effluent discharge, recreational activity levels, or
freshwater input. This categorical approach was aimed at
beginning to understand the sources of fecal pollution in
the Hawaiian environment. However, because many marine
sites are exposed to a combination of impact factors (e.g.
high recreational activity + freshwater input), it is
difficult at this point to accurately pinpoint direct
sources of fecal contamination. Despite the lack of
particularly clear-cut correlations, results of the
bioaccumulation study do provide an opportunity to explore
various possibilities regarding enteric virus distribution
in the marine environment. Such suppositions are discussed
below.
Ala Moana Beach Park (including the Magic Island
sampling site) is an extremely popular bathing beach with
exceptionally high recreational activity levels. In 1994,
this beach accommodated an estimated 1.5 million people
(Paul 1997). Therefore, bather shedding may be largely
responsible for the fact that 4 of the 5 enteric virus
subgroups included in the study were detected in this

94
region. The reef-protected waters of Ala Moana Beach Park
remain quite calm throughout the year, resulting in
decreased water circulation and, therefore, increased viral
persistence. Input from the nearby Ala Wai Canal may also
contribute to pollution levels, particularly during storm
events, as this body of water receives a significant
fraction of urban runoff (Connolly 1999). There has been
concern that migrating effluent from the Sand Island
Wastewater Treatment Plant outfall deteriorates water
quality at shoreline beaches such as Ala Moana; however,
findings of Paul et al suggest that the offshore outfall is
not a major contaminating influence (Paul 1997). Waste
discharge from boats an adjacent harbor is yet another
potential source of contamination at Ala Moana Beach Park.
At the Sand Island State Recreational Area, all
shellfish tested positive for NoVI, and a very small number
of samples contained AdV. It was originally thought that
this area may be more heavily polluted due to effluent
drifting back to shore from the Mamala Bay wastewater
output. However, although enteric viruses are frequently
detected near the output itself, it is believed that they
do not commonly return to nearshore waters (Paul 1997).
Also, wave action was very high during the period of sample
collection; this dramatic mixing of water probably had a

95
major dilution effect on any enteric viruses that may have
present in the surrounding water.
The beaches of Kahala, Kawaikui, and Waialae may be
grouped similarly in that all receive moderate levels of
recreational activity and non-point source input. Pipes
connected to storm drainage systems may release runoff into
these waters following periods of heavy rainfall.
Additionally, Waialae Beach is impacted by discharge from
Kapakahi Stream (Vijayavel 2010). The detection of several
types of enteric viruses in shellfish from these locations
may be attributable to these reasons. Also, the waters of
Waialae and Kawaikui were especially turbid during the time
of sample collection. Because enteric virus survival is
enhanced through association with particles within the
water column (as previously discussed), this may have been
another contributing factor to their positive detection.
Kahana Bay and Punalu’u Beach Park receive discharge
from Kahana Stream and Punalu’u Stream, respectively. Both
sites receive low to moderate recreational activity. AdV,
EnV, and F+ were detected at Punalu’u. At Kahana Bay, NoVI
and NoVII were detected. Often observed near these two
areas are a number of homeless individuals, whose open
defecation may be discharged into the ocean. It should be
noted that, due to extremely high turbidity in the main

96
section of the bay, shellfish tested from Kahana Bay were
collected from a relatively small sampling quadrant quite
remote from the rivermouth. The highly diluted nature of
the water in this region may explain why norovirus was the
only virus detected.
At Kualoa Park, all viruses except NoVII were
detected. As previously mentioned, the park’s faulty septic
system has been a major pollution problem in the past. The
beach park restrooms are now closed with caution tape.
However, camping is quite popular at Kualoa Park, and
campers may either attempt to use the restrooms anyway, or
choose to defecate freely on park grounds. Both
possibilities will clearly negatively impact the water
quality at Kualoa. Additionally, shellfish were collected
from a calm region protected from the open ocean by cement
barricades; decreased wave action and circulation in this
area may boost viral persistence.
Findings from Isognomon spp. inhabiting the lagoons of
Ko Olina are particularly interesting. While shellfish from
Lagoon 3 tested positive for all 5 viral subtypes,
shellfish from Lagoon 4 were only shown to contain NoVI.
Conditions at these two man-made lagoons are extremely
similar; both are extremely well protected by rock
barricades and very popular bathing beaches. While the

97
differences in enteric virus prevalence at these sites may
seem contradictory, two possible explanations are
hypothesized. First is the difference in sampling periods.
Because these lagoons are part of a resort, tourists are
the major beachgoers. Lagoon 3 was sampled during the
summertime, the peak vacation season for tourist and
swimmer activity. In contrast, Lagoon 4 was sampled in
October, a less popular month for recreational water
activity at Ko Olina. This may partially explain the
opposing results of the bioaccumulation study in this
region. Another contributing factor may be the precise
sampling location. Shellfish from Lagoon 3 were collected
in the shallow waters nearest the beach, where bathers tend
to congregate and where circulation is lowest. In contrast,
shellfish from Lagoon 4 were harvested from the offshore
region opening out past the protective rock barrier, where
few swimmers travel and where the diluting effect of wave
action is stronger than within the cove. It might be
expected that shellfish inhabiting this area would
bioconcentrate enteric viruses to a lesser extent than
their counterparts closer to the shore. Because of these
interesting factors and because Isognomon spp. are
extremely prevalent and easily harvestable at Ko Olina,
this site is ideal for future exploratory studies aimed at

98
deciphering the complex dynamics of shellfish-mediated
bioaccumulation of viral pathogens.
Although the high presence of enteric viruses reported
in Hawaii’s marine environment may be alarming, caution
should be made when interpreting results. PCR detects both
infectious and damaged viruses, so without correlating
infectivity data, public health risks are difficult to
assess. However, regardless of their infectivity, enteric
viruses may be used as indices of human fecal pollution.
Their positive detection indicates the potential presence
of other human pathogens of concern in aquatic
environments.
This pilot study utilizing indigenous Hawaiian
mollusks as competent bioindicators of water quality is an
innovative approach to screening environmental waters for
fecal pollution. Because enteric viruses often persist in
environmental waters at such low concentrations, their
effective detection is greatly aided by the natural
bioconcentration phenomenon of shellfish. Research findings
using the novel protocol suggest a high prevalence of viral
pathogens in Hawaiian recreational waters. Future research,
including infectivity assays and laboratory-controlled
spike studies to more directly measure bioaccumulation and
inhibition levels, will further investigate the practical

99
feasibility of using shellfish as natural bioindicators of
microbial water quality.

100
REFERENCES
Arnal C, Ferre-Aubineau V, Besse B, Mignotte B, Schwartzbrod L, Billaudel S (1998) Comparison of seven RNA extraction methods on stool and shellfish samples prior to hepatitis A virus amplification. J Virological Methods 77: 17-26 Asahina AY, Lu Y, Wu C, Fujioka RS, Loh PC (2009) Potential biosentinels of human waste in marine coastal waters: bioaccumulation of human noroviruses and enteroviruses from sewage-polluted waters by indigenous mollusks. J Virol Methods 158: 46-50 Atmar RL, Neill FH, Romalde JL, Le Guyader F, Woodley CM, Metcalf TG, Estes MK (1995) Detection of Norwalk virus and hepatitis A virus in shellfish tissues with the PCR. Appl Environ Microbiol 61: 3014-3018 Bosch A and Le Guyader SF (2010) Introduction: Viruses in shellfish. Food Environ Microbiol 2: 115-116 City & County of Honolulu. Hawaii’s implementation plan for polluted runoff control, Appendix F: Detailed descriptions of Hawaii’s 18 water quality limited segments Connell C, Tong H, Wang Z, Allmann E, Lu Y (2012) New approaches for enhanced detection of enteroviruses from Hawaiian environmental waters. PLoS ONE (in production) Connolly JP, Blumberg AF, Quadrini JD (1999) Modeling fate of pathogenic organisms in coastal waters of Oahu, Hawaii. J Environ Engineering 398-406 Devine J (2011) Sources of beachwater pollution. Natural Resources Defense Council Dore WJ, Lees DN (1995) Behavior of Escherichia coli and male-specific bacteriophage in environmentall contaminated bivalve molluscs before and after depuration. Appl and Environ Microbiol 61: 2830-2834 Dorfman M and Rosselot KS (2011) Testing the waters: A guide to water quality at vacation beaches. Twenty-first annual report. Natural Resources Defense Council Fattal B, Olevsky E, Cabelli VJ (1991) Bathers as a possible source of contamination for swimming associated

101
illness at marine bathing beaches. Int J Environ Health Res 1: 204–214 Formiga-Cruz M, Allard AK, Conden-Hansson AC, Henshilwood K, Hernroth B E et al (2003) Evaluation of potential indicators of viral contamination in shellfish and their applicability to diverse geographical areas. Appl Environ Microbi 69: 1556-1563 Goblick GN, Anbarchian JM, Woods J, Burkhardt W, Calci K (2011) Evaluating the dilution of wastewater treatment plant effluent and viral impacts on shellfish growing areas in Mobile Bay, Alabama. J of Shellfish Research 30: 979-987 Hansman GS, Oka T, Okamoto R, Nishida T, Toda S, Noda M, Sano D et al (2007) Human sapovirus in clams, Japan. Emerg Infect Dis 13: 620-622 Harper E and Morton B (1994) The biology of Isognomon legume (bivalvia: pterioida) at Cape D’Aguilar, Hong Kong, with special reference to predation by muricids. The Malacofauna of Hong Kong and Southern China III: Proceedings of the Third International Workshop on the Malacofauna of Hong Kong and Southern China, Hong Kong 13 April – 1 May 1992. Hong Kong: Hong Kong University Press Hernroth BE, Conden-Hansson ACC, Rehnstam-Holm AS, Girones R, Allard AK (2002) Environmental factors influencing human viral pathogens and their potential indicator organisms in the blue mussel, Mytilus edulis: the first Scandinavian report. Appl Environ Microbiol 68: 4523-4533 Hunt C, USGS. Wastewater and nutrient source tracking – results of reconnaissance chemical mapping at Kualoa and Kahana, Oahu. HI DOH/USGS Cooperative Program, Pacific Islands Water Science Center Ismail A (2006) The use of intertidal molluscs in the monitoring of heavy metals and organotin compounds in the west coast of Peninsular Malaysia. Coastal Marine Science 30: 401-406 Jamieson RC, Joy DM, Lee H, Kostaschuk R, Gordon RJ (2005) Resuspension of sediment-associated Escherichia coli in a natural stream. J Environ Qual 34: 581-589 Karamoko Y, Ibenyassine K, Aitmhand R, Idaomar M, Ennaji M M (2005) Adenovirus detection in shellfish and urban sewage

102
in Morocco (Casablanca region) by the polymerase chain reaction. J Virological Methods 126:135-137 Kingsley DH, Richards GP (2001) Rapid and efficient extraction method for reverse transcription-PCR detection of hepatitis A and Norwalk-like viruses in shellfish. Appl Environ Microbiol 67: 4152-4157 Kingsley D, Provost K (2010) A potential mechanism of virus persistence within bivalve shellfish [abstract]. American Society of Virology. Bozeman, Montana, p. 1 Krock HJ, Rujioka RS (1993) Kailua Bay bacteriological water quality and circulation assessment report (KB-6) Water Resources Research Center, University of Hawaii at Manoa, Honolulu, HI Landry EF, Vaughn JM, Vicale TJ, Mann R (1983) Accumulation of sediment-associated viruses in shellfish. Appl Environ Microbiol 45: 238-247 Le Guyader FS, Loisy F, Atmar RL, Hutson AM, Estes MK, Ruvoen-Clouet N, Pommepuy M et al (2006) Norwalk virus-specific binding to oyster digestive tissues. Emerg Infect Dis 12: 931-936 Lees DN, Henshilwood K, Dore WL (1994) Development of a method for detection of enteroviruses in shellfish by PCR with poliovirus as a model. Appl Environ Microbiol 60: 2999-3005 Lees D (2000) Viruses and bivalve shellfish. International J Food Microbiol 59: 81-116 Loisy F, Atmar RL, Guillon P, Cann PL, Pommepuy M, Le Guyader FS (2004) Real-time RT-PCR for norovirus screening in shellfish. J Virological Methods 123:1-7 Manso C F, Polo D, Vilarino M L, Romalde J L (2010) Genotyping of hepatitis A virus detected in bivalve shellfish in Falicia (NW Spain). Water Sci Tech 61:15-24 McDonald AT, Chapman PJ, Fukasawa K (2008) The microbial status of natural waters in a protected wilderness area. J Environ Management 87: 600-608 Miller WA, Atwill ER, Gardner IA, Miller MA, Fritz HM, Hedrick RP, Melli AC, Barnes NM, Conrad PA (2005) Clams

103
(Corbicula fluminea) as bioindicators of fecal contamination with Cryptosporidium and Giardia spp. in reshwater ecosystems in California. International J Parasitology 35: 673-684 Nappier SP, Graczyk TK, Schwab KJ (2008) Bioaccumulation, retention, and depuration of enteric viruses by Crassostrea virginica and Crassostrea ariakensis oysters. Appl Environ Microbiol 74: 6825-6831 Paul JH, Rose JB, Jiang SC, London P, Xhou X, Kellogg C (1997) Coliphage and indigenous phage in Mamala Bay, Oahu, Hawaii. Appl Environ Microbiol 63: 133-138 Pond, Kathy (2005) Water recreation and disease - Plausibility of associated infections: acute effects, sequelae and mortality. World Health Organization Richards GP, McLeod C, Le Guyader FS (2010) Processing strategies to inactivate enteric viruses in shellfish. Food Environ Virology 2: 183-193 RNA PowerSoil Total RNA Isolation Kit Instruction Manual. Version 02162010. MoBio Laboratories, Inc. Schultz AC, Saadbye P, Hoorfar J, Norrung B (2007) Comparison of methods for detection of norovirus in oysters. International J of Food Microbiol 114: 352-356 Schwab KJ, Neill FH, Le Guyader F, Estes MK, Atmar RL (2001) Development of a reverse transcription-PCR-DNA enzyme immunoassay for detection of Norwalk-like viruses and hepatitis A virus in stool and shellfish. Appl Environ Microbiol 67:742-749 Sombrito EZ, Honrado MCV, de Vera A, Tabbada RS, Ranada ML, Relox J, Tangonan MC (2007) Use of Perna viridis as a bioindicator of paralytic shellfish toxins at low Pyrodinium bahamense var compressum density using a radioreceptor assay. Environmental Bioindicators 2: 264-272 Umesha KR, Bhavani NC, Venugopal MN, Karunasagar I, Krohne G, Karunasagar I (2008) Prevalence of human pathogenic enteric viruses in bivalve molluscan shellfish and cultured shrimp in south west coast of India. International J Food Microbiology 122: 279-286

104
Viau EJ, Lee D, Boehm AB (2011) Swimmer risk of gastrointestinal illness from exposure to tropical coastal waters impacted by terrestrial dry-weather runoff. Environmental Sci Technol 45: 7158-7165 Vijayavel K, Rujioka R, Ebdon J, Taylor H (2010) Isolation and characterization of Bacteroides host strain HB-73 used to detect sewage specific phages in Hawaii. Water Research 44: 3714-3724 Vilarino ML, Le Guyader SL, Polo D, Schaeffer J, Krol J, Romalde JL (2009) Assessment of human enteric viruses in cultured and wild bivalve molluscs. International Microbiology 12: 145-151 Xagoraraki I, Kuo DHW, Wong K, Wong M, Rose JB (2007) Occurrence of human adenoviruses at two recreational beaches of the Great Lakes. Appl Environ Microbiol 73: 7874-7881

105
CHAPTER 5
CONCLUSIONS
Described here are the development and application of
novel protocols for the effective detection of human
enteric viruses in Hawaiian environmental waters. First, a
highly optimized and sensitive protocol for enterovirus
detection was established. Once confirmed through testing
urban wastewater, this protocol was utilized in a
surveillance study of 22 recreational bodies of water
located on Oahu, 11 of which tested enterovirus-positive.
Next, an innovative approach to increased enteric
virus detection sensitivity was explored, utilizing the
bioconcentration phenomenon of filter-feeding shellfish.
Specimens were collected from 12 marine sites and tested
for the presence of an array of important viruses,
including adenovirus, enterovirus, norovirus genogroups I
and II, and F-specific RNA coliphage. Multiple virus types
were detected in sampling groups from all around the
island, indicating fecal contamination in a significant
portion of Oahu’s costal waters. Enteric virus detection
from corresponding water samples revealed that the
utilization of Isognomon spp. dramatically enhances
detection sensitivity.

106
The conveyed methods provide practical means of
utilizing enteric viruses as alternative indicators, with
potential to enhance accurate assessment of microbial water
quality and minimize risks associated with polluted
recreational waters. Reliable microbial water quality
monitoring is vital to ensure the safety of swimmers,
surfers, divers, and other recreational users of waters in
Hawaii and worldwide. Research findings provide meaningful
insight into the degree of fecal contamination here in the
Hawaiian environment. The revealed polluted regions are
sites of potential health risk and should instigate
additional investigation of our beloved recreational
waters.
















![Biological and water and sediment quality surveys in Mānoa … · AECOS Inc. [FILE: 1463] Page | 1 Biological and water and sediment quality surveys in Mānoa Stream, Honolulu, Hawaiʻi](https://img.pdfslide.net/doc/110x75/60180488105cd656d734b2f5/biological-and-water-and-sediment-quality-surveys-in-mnoa-aecos-inc-file-1463.jpg)