Embed Size (px)
Citation preview

Neuere Ergebnisse der vergleichenden Physiologie der Verdauung der Siiugetiere.
Von
W . LENKEIT- B e r l i n .
Mit 9 Abbildungen.
Inhal~sverzeichnis. seite
Literaturverzeichnis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 574
Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 577
A. Mundverdauung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 578
I. l~ahrungsaufnahme . . . . . . . . . . . . . . . . . . . . . . . . . . . 578
II. Kauen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 579
IIL Die Speichelsekretion . . . . . . . . . . . . . . . . . . . . . . . . . . 580
Die Speicheldriisen S. 580. - - Sekretion des Speichels S. 581. - - Die Zu- sammense tzung des Speichels S. 583.
IV. I)as Abschlucken der Nahrung . . . . . . . . . . . . . . . . . . . . . . 588 B. Allgemeines zur ~Flagen-Darmverdauung . . . . . . . . . . . . . . . . . . . . 589 C. Magenverdauung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 590
I. MagengrSsse. l~egenerationsf/~higkeit der Vorm/~gen der Wiederk/~uer . . . . . 590 II. Magensaftsekre~ion . . . . . . . . . . . . . . . . . . . . . . . . . . . . 591
Die Magendrfisen S. 591. - - Der Magensaft S. 593. - - Ablauf der Sekretion S. 596.
I I I . Mechanik des einhShligen )Sagens . . . . . . . . . . . . . . . . . . . . 598 IV. Mechanik des mehrhShligen Magens (Wiederk/~uer) . . . . . . . . . . . . . 599
Bewegung der Vorm/kgen und des Labmagens , einschl. Schlundrinne S. 599. - - Innerva t ion der Vorm~gen und des Labmagens S. 604. - - ])er Mechanismus des Wiederkauens S. 605. - - Wiederkauen und Erbrechen S. 610. - - I)er l~uctus S. 610. - - Uber t r i t t des Fu t t e r s aus d~n beiden ersten Vorm/igen in Psal ter und Haube S. 611. - - Fltissigkeitstranspor~ S. 612.
V. Ablauf der Magenverdauung . . . . . . . . . . . . . . . . . . . . . . . 612 EinhShlige 5~/~gen S. 612. - - Mehrh6hlige M/~gen S. 614. - - Panseninfusorien
S. 616. - - Resorpt ion in den Vom/~gen S. 617. D. Darmverdauung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 618
I. Morphologisches . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 618 Gr5ssenverh/~ltnisse S. 618. - - ])armdri isen S. 619.
II. Verdauungss/~fte des Darmes . . . . . . . . . . . . . . . . . . . . . . . 619
X)ankreassaft S. 619. - - Galle S. 620. - - Df inndarmsaf t S. 621. - - Dickdarm- saft S. 622.
I I I . 1V~echanik der Darmverdauung . . . . . . . . . . . . . . . . . . . . . . 622 IV. Ablauf der Darmverdauung . . . . . . . . . . . . . . . . . . . . . . . 624
Verdauung der pflanzliohen Nah rung im Di inndarm S. 624. - - Reak~ion des Darminhal tes S. 625. - - Resorpt ion S. 625.
E. Aufentha l t sdauer der Nahrung im Verdauungskanal und der quant i ta t ive Verlauf der Ausscheidung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 626

574 W. LENKEIT: Neuere Ergebnisse der verg]. Physiologie der Verdauung der Saugetiere.
L i t e r a t u r v e r z e i c h n i s . A. Zusammen[assende Darstellungen.
BA~KIN, B . P . : Die ~ussere Sekretion der Verdauungsdrfisen. Berlin- Julius Springer 1928. BIEDmZ~_~N, W. : Die Ern~hrung der h6heren Wirbcltiere. t tandbuch der vergleichenden Physiologic yon W I ~ s T ~ I ~ , Bd. 3, S. 1116--1492. 1911.
ELLE~BE~GE~-Sc~UZ~E~T: Vergleichende Physiologie der ~Luss~ugetiere, 3. Aufl. Berlin 1925.
DUKES, H. H. : 1%ecent advances in the Physiology of digestion. J. amer. vet. med. Assoo. 77, 225--228, 313--319 (1930); 78, 92--97, 235--239 (1931).
HESSE, 1%. u. F~. DOFLEI~: Tierbau und Tierleben, Bd. 1, 2. Leipzig 1910 u. 1914. K~zYvc~:~EK, F ~ . W . : (1) Die Verdauung des Schweines. I-Iar~dbuch der Ern~hrung
und des Stoffwechsels der landwirtschaftlichen Nutztiere, herausgeg, yon E. 1VIA~GOr.D, Bd. 2, S. 270--309. 1929. (2) Dos Wiederkauen. Handbueh der normalen und pathologisohen Physic- logic, Bd. 18, S. 36--44 (Nachtrag zu Bd. 3). 1932.
L]~lVK]~IT, W. : SpeiserShre und Magen. Anatomic und Pathologic der Spontanerkrankungen der kleinen Laboratoriumstiere yon 1%. JAFfa, 1931. S. 83--90.
M~GOI~]), E.: (1) Physiologic der Speiser6hre. I-Iandbuch fiir ~als-, Nascn- und Ohren- kra.nkheiten yon DE~xE~ u. KAOLin, Bd. 9, S. 19. 1929. (2) Die Verdauung der Wiederk~uer. I-Lndbuch der Ern~hrung und des Stoffwechsels der landwirtsohaftlichen Nutztiere, herausgeg. yon E. 1VIAduct,l), Bd. 2, S. 107--236. 1929. (3) Die Infusorien des Pansens und ihre Bedeutung fiir die Ern~hrung der Wiederk~tuer (Sammelreferat). Biedermanns Zbl. A~ N. F. 8, 161--187 (1933).
P~wK, It . : Der 1V[agen. ~andbuch der mikroskopischen Anatomic des ]~[enschen yon W. v. ]V[Srm~DO~F, : Bd. 5, Tell 2, S. 1--205. 1932.
1%os]~]~A~r t~. : Physikalisohe Eigenschaften und chemische Zusammensetzung der Ver- dauungss~fte unter normalen und abnormen Bedingungen. Handbueh fiir normale und patho- logische Physiologie, Bd. 3, S. 819--875. 1927.
Sc~mV]~l~% A. : (1) Verdauung der Wirbeltiere. Kandbuch der Bioehemie des iVIenschen und der Tiere yon O ~ ] ~ E I ~ , Bd. 5, S. 56--201. 1925. (2) Dos Wiederkauen. Handbuch ffir normMe und pathologische Physiologie, Bd. 3, S. 377--397. 1927. - - u. FE.. W. K_~zYwA~Ex: Die Verdauung des Pferdes. t tandbueh der Ern/~hrung und des Stoffwechsels der landwirt- sohaftliehen Nutztiere yon E. MAn,OLD, Bd. 2, S. 237--270. 1929.
W ~ s ~ , J. : Die Physiologic und Pathologie der Vormi~gen beim 1%inde. Berlin: 1%iohard Sehoetz 1926.
B. Spezielle Arbeiten.
A ] ~ o m z ~ z o ~ , E. u. E. W m z T ~ X ~ : Pflfigers Arch. 1~4, 168 (1922). - - u. 1%o~r Z. physiol. Chem. 47, 359 (1906). Ac]~A~, L . K . u . N . A . PODKOPAEW: Pflfigers Arch. 224, 539 (1930). Ao])xs, T.: Amer. J . Physiol. 99, 417 (1932). A_KSS]~OWA, ~ . J . : Arch. Tier- ern~thrg u. Tierzueht 7, 295 (1932). ASC~OFF, L.: Pfltigers Arch. 2Ol, 67 (1923).
B)_B~dEw, G., N. P~]~ST~W u. J. KUL]~S]~O: 1%ussk. fiziol. ~. 1~, 636 (1930). 1%el. Ber. Physiol. 61, 697. ]3~x~]~R, G. S ~ E w ~ : (1) Amer. J . Physiol. ~6, 343 (1931). 1%el. Ber. Physiol. 61, 89. (2) Trans roy. Soc. Canad~ V. Biol. Sci. I II . 24, 213 (1930). B]~c]~n~, E. 1%. and 1%. C. Ev]~R~.~T: Amer. J . Hbgg. 11, 362 (1930). - - and I-Is~uNo: Prec. nat. Acad. Sci. U. S. A. 1~, 684 (1929). BECK]~R, 1~. u. A. OP~]~N]~EI~R: Fortschr. 1%6ntgenstr. 42, 730 (1930). BEL- OOWS~a, J . : Pfliigers Arch. 148, 319 (1912). B~Nz, J . : ~ed . vet. Diss. Ziirich 1930. B~.Ro, B . N . : Prec. Soc. exper. Biol. a. l~[ed. 28, 724 (1931). B]~R~N~v, I-I.D. and t t . H . ] ) u K E s : J . amer. reed. Assoc. 69, 600 (1926). BICKEL, A. : Arch. Verdgskrkh. 46, 70 (1929). BIEN]~RT, G. : Pfliigers Arch. 213, 556 (1926). BIEST]~R, 1%.: Arch. Tierernahrg u. Tierzucht 7, 109 (1931). B~]~NK~rP: Beitr. path. Anat. 82, 475 (1929). BRVGSC~ u. tIoRsTER: Med. Klin. 1~24, Nr 20. Bv~s, A. : Diss. Tierarztl. I-Ioohseh. I-~annover 1931.
Cmsm~, P . P . : Diss. Petersburg 1894. C ~ m % ~ . : ])iss. Landw. ttochsch. Bonn 1930; Z. Zellforseh. 11, 342 (1930). Cm~z~szcz, T. u. Z. Scm~]~C~Tn6W~: Bioehem. Z. 219, 30 (1930). CI~CC~, ]).: An. Fac. IVied. e Chir. e Fac. 1Ked.-vet. Perugia 28, 143 (1926). CoL~N, G.:

Li~eraturverzeiehnis. 575
TrMt~ de physiol, eompar~e, 3. Aufl. Paris 1886. COLWtnvs, A. : Diss. Landw. Hoehsch. Berlin 1933. CzEI~A, A. u. 1%. STIOLE~: (1) 1)fliigers Arch. 212, 300 (1926). (2) Fortsehr. naturwiss. Forseh. 1929, H. 6. CZVBALS~: C. r. Soc. Biol. Paris 97, 964 (1927).
D A ~ G ~ , 1K, K. P r R A G ~ u. H. SCn~'L~ES: Pfliigers Arch. 220, 430 (1928). DIE- M~GER, H.: ivied. Diss. Wfirzburg 1898. DIEI~ST, C.: Z. exper. ~ed. 83, 718 (1932). DIoN~ssov: l%ussk, fiziol. Z. 9, 395 (1926). DOGI~L, V. u. W~OGRADOwA-FEDoRowA: Arch. Tiererni~hrg u. Tierzucht 3, 172 (1930). DYKes, H . H . : J. amer. vet. med. Assoc. 77, 225 (1930).
EASTMANN, I. and E. G.Y~LLE~ jr.: Prec. Soc. exper. Biol. a. Med. 30, 332 (1932). E1)~L)~AN~: Dtsch. Z. Tiermed. u. vergl. Path. 15, 165 (1889). E~RENBERr P. U. H. BRIES~: Biochem. Z. 257, 194: (1933). ELLENBEI~GEIr W. : I~andbuch der vergleichenden mikroskopisehen
.Anatomie, Bd. 3, S. 241. 1911. ~ u. I t . B~uM: Handbuch der vergleichenden Anatomic der ttaustiere. Berlin: Julius Springer 1933. - - u. V. I~OFME~S~: Dtsch. Z. Tiermed. u. vergl. Path. 11, 249 (1885). ER~EN, A.: Pflfigers Arch. 213, 571 (1926). E~Ss, G.: Frankf. Z. P~th. 36, 402 (1928).
F~RB~R, K. E.: Z. Tierzfichtg u. Zfichtgbiol. 12, 31 (1928); 15, 375 (1929). - - u. WI~O- GRADOw~-F~Do~ow~: Biol. Zbl. 49, 321 (1929). FERRA~, 1%.: Pflfigers Arch. 230, 215 (1932). FIscm~, E. u. 1%. ~i. K ~ N : Pfliigers Arch. 225, 694(1930). Fos C.: Pflfigers Arch. 133, 171 (1910). FoNs~,c~ et T~INCAO: C. r. Soc. Biol. P~ris 99, 1533 (1928). FR~ZZ~T~O, S.: Arch. itM. Anat. 28, 569 (1931). FR~sc~zB~]~R: Z. Vet.kde 45, 1 (1933).
GAL]~mr 0. : Fermentforsch. 9, 224 (1927). G~U~)ENZ~, P.: Ann. Fac. ~cd. e Chir. e Fae. Mcd.-vet. Pcrugia 28, 3 (1926). G~Y~)A, T. : Arch. di Fisiol. 24, 692 (1926). G~x~ovsKY, V.: Biol. Listy (tschech.) 14, 184 (1929). G~XssN~.R, K.: Biochem. Z. 127, 312 (1922). GO~D-
~ R G , S:L. : Arch. int. IVied. 49, 816 (1932). GO~)SC~IDT, I t . : Z. physiol. Chem. 19, 361 (1886). GOTTSCttALK, A.: Med.-vet. Diss. Ztirich 1910. G ~ E , K.: Z. Laryng. usw. 19, 405 (1930). G~YCE~, D. ~ . and E. G. M ~ L ~ : J. of biol. Chem. 76, 423 (1928). GROSSER, P.: Z. Physiol. 19, 265 (1905). GR~3ss: Abderhaldens/-Iandbuch dcr biologischen Arbeitsmethoden, Abt. 4, Tell 1, H. 1, S. 37. G~i)~z~E~: Pfltigers Arch. 16, 105 (1877). GUDZONSSON, V. SKU~X u. G~LY OPP~N~.~: Z. exper. Med. 71, 220 (1930).
~ E s ~ , K.: Z. Ziichtg, l~eihe B: Tierztichtg u. Zfichtgsbiol. 17, 339 (1930). / - I ~ - )IA~N, 0. : Landw. Jb. 20, 264 (1891). K~UC~CO~NE, FR.: Z. ~orph. u. Okol. Tiere 9, 439 (1927). H~vRow~z, F. u. W. Pv ,~o~ : t~oppe-Seylers Z. 144, 68 (1925). H~NRIC~S: Pflfigers Arch. 164, 303 (1916). K~CK~, /{.: Diss. Tierarztl. ttochseh. ]~nnover 1931. I~IEV~XE, W.: (1) Z. Ernahrg 2, 138 (1932). (2) Z. exper. IVied. 75, 83 (1931). H o ~ A ~ , W.: Arch. Tierheilk. 61, 373 (1930). ~ONCA~, F. u. ST. KONDE~: Z. Tierzfichtg u. Zfichtgsbiol. 10, 1 (1927). - - W. SC~RA~ u. H. STO~Z: Z. Tierzfichtg u. Zfichtgsbiol. 11, 433 (1928). I { U K V ~ A : Pfliigers Arch. 226, 518 (1931).
I~OKA: Pflfigers Arch. 204, 368 (1924). Jv~a, L.: C. r. Soc. Biol. P~ris 93, 526 (1925). KEST~ER, O.: Pflfigers Arch. 229, 206 (1932). KLEIN, W.: Biochem. Z. 62, 225 (1916);
117, 67 (1921). K~P~R: Zit. nach OPPENhEImeR. KNOTtt, IV[.: Z. Parasitenkde 1, 262 (1928). K O ~ A N ~ : Arch. Verdgskrkh. 35, 34 (1925). KoKhs, E . v . : Pfliigers Arch. 225, 416 (1930); 229, 486 (1932). - - u. G. v. LuDs Pfliigers Arch. 225, 421 (1930). KOSCHTOZANZ, Cm S.: (1) Pfliigers Arch. 220, 642 (1928). (2) Pflfigers Arch. 227, 359 (1931). - - I. I w ~ o ~ F , W. ~ U ~ F F , P. KO~N~EFF U. O~SC~t~KOWK~: Z. vergl. Physiol. 18, 112 (1932). KOST~AL, L.: Magy. err. Arch. 27, 276 (1926). Autoref. Ber. Physiol. 37, 345 (1926). K~A~OW, A. u. P. K ~ T ~ O W ~ : t~ussk, fiziol. ~. 15, 492 (1932). t~ef. Ber. Physiol. 71, 561 (!933). - - u. D. SKtr- LOW: l~ussk, fiziol. Z. 15, 484 (1933). Ref. Ber. Physiol. 71, 559 (1933). Y~x~OWA, P. u. A. K~ATI~OW: ]~ussk. fiziol. Z. 15, 502 (1932). ~ef. Ber. Physiol. 71, 561 (1933). K ~ . , It . : Diss. Kiel 1927. KRu~]~IE~E~, I.: Bio]ogie der Tiere Deutschlands yon P. SCHVLz]~, Lief. 3!, Teil 52, ~r I. I(~ZYW~N]~K, FR. W.: (1) Pflfigers Arch. 215, 481 (1927). (2) Arch. Tierheilk. 55, 523, 537; 56, 49, 57 (1927). (3) Pfliigers Arch. 222, 89 (]929). - - u. W. LA~x~E.: Dtsch. tierarztl. Wschr. 40, 289 (1932). KuLL: Z. mikrosk.-anat. Forseh. 2, 163 (1925).
L~FOND, M.: ~evue de Stomat. 32, 620 (1930). LAMPE, W.: Vet.-med. I)iss. Leipzig 1931. ~cL~NGHL~N, A~VA~ R.: Science (N.Y.) 1, 191 (1931). LENKE~T, W.: (1) Ratten- vormagen. Arch. Tierheflk. 64, 255 (1931). (2) Berl. ticrarztl. V~schr. 48, 17 (1932). (3)/qormMe

576 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologic der Verdauung der S/~ugetiere.
Anatomic und spontane Erkrankungen der kleinen Laboratoriumstiere. Herausgeg. yon JAFF]~, S. 87. Berlin 1931. (4) Arch. Tierern/~hrg u. Tierzucht 5, 376 (1931). (5) Arch. Tierern/~hrg u. Tierzucht 8, 631 (1930). (6) UnverOffentlicht. LENKEIT, W. u. A. COLUMBUS: Im Erscheinen.
u. HABECK: Arch. Tierern~hrg u. Tierzueht 2, 517 (1930). LEvI~, S.: Ter. Arch. (russ.) 8, 332 (1930). Autoref. ]3er. Physiol. 57., 204. LI~, R. K. S.: Quart. J . microsc. Sci. 66, 87 (1922). Lo~wr Miineh. reed. Wschr. 39, 474 (1892). L~3cx~zeG, W.: Dtsch. tier/~rztl. Wschr. 34, 257 (1926).
MAOE~, H. E.: J. of exper. Biol. 9, 409 (1932). 1V[~G~A~, A. : C. r. Acad. Sci. Paris 154, 129 (1912). iK~OOLD, E.: (1) Naturwiss. 16, 65 (1928). (2) 12. internat, physiol. Kongr. Stockholm. Skand. Arch. Physiol. (Berl. u. Lpz.) 1926. (3) Handbuch der biologischen Arbeits- methoden yon ABDERt~kLDE1% Abt. 4, Teil 6/2, S. 1775. 1928. (4) I-Iandbuch der Ern/~hrung und des Stoffwechsels der landwirtschaftlichen Nutztiere, ]3d. 2, S. 202. 1929. (5) Med. Klin. 2, 1336 (1932). (6) Ver/~nderurigen der Verdauungsmechanik durch k5rperliche Bewegung. Verh. 14. internat. Kongr. Physiol. 1932, 170. (7) Z. Hundeforsch. 2, 38 (1932). - - u. J. Dg- ]~ISKI: Arch. Tierern/~hrg u. Tierzucht 4, 507 (1931). - - u. W. KLEIN: ]3ewegungen und Inner- ra t ion des Wiederk/~uermagens. Leipzig: G. Thieme 1927. - - u. W. LENK~IT: Arch. Tier- ern/~hrg u. Tierzucht 5, 201 (1931). - - u. T. RA])EFF: Arch. Tierern/~hrg u. Tierzucht 4, 175 (1930). - - u. K. SCmVgTT-K~A~ER: Biochem. Z. 191, 411 (1927). - - u. F. USU~LLI: Arch. Tierern/~hrg u. Tierzucht 3, 189 (1930). MAt~KOFF, J . : ]3iochem. Z. 57, 1 (1913). MAI~TI~, P.: Anatomie der ttaustiere, ]3d. 1, 3. 1912 u. 1919. M~LLI et RADICI: Fol. olin. ehim. et microsc. (Bologna) 3, 169 (1928); ]3er. 49, 494. MEtqVILLE, L. I. AUE an(i S. N. ]3LACKBERG: Prec. See. expcr. Biol. a. Med. 27, 641 (1930). METZNER, R.: Pfliigers Arch. 201, 60 (1923). MEYER, W.: Z. vergl. Physiol. 6, 402 (1927). MIC~A]~LrS, E. u. PEC~STEIN: ]3iochem. Z. 59, 77 (1914). MOVALDI, M.: 1%iv. Biol. 13, l l 6 (1931). Mow~u H . A . and E. R. ]3ECKE~: Iowa State Coll. J . Sci. 5, 35 (1930). iKuss~r Ber. 4. russ. physiol. Kongr. Charkow, Mai 1930, 156.
1X~GL, F~.: Arch. Tierheilk. ~8, 198 (1928). NAKATSVKA, S.: Mitt. reed. Akad. Kioto 5, englische Zusammenfassung, 1931. S. 26. N ~ s o N , N.M. and K. F. MEYer.: J. inf. Dis. 28, 510 (1921). N ~ v ~ Y g , ]-I.: Sitzgsber. Ges. Morph. u. Physiol. Mfinchen l~, 139 (1899/1900). N~VnR, ]-I. E.: Pflfigers Arch. 225, 51 (1930).
OPPEL: Vergleichende mikroskopische Anatomic der Wirbeltiere, ]3d. 1. 1896. OPPeN- hEImEr, C.: Die Fermente und ihre Wirkungen, 5. Aufl., S. 1. Leipzig 1925. OST~OF, W.: Med.-vet. Diss. Giessen 1915.
PATZSCtIKE, A.: Med. Diss. Leipzig 1932. PELUFFO, A.: Rev. Soc. argent. Biol. 4, 1012 (1928). P~.RNKOPF, E.: ]3eitr/~ge zur vergleichenden Anatomie des Vertebratenmagens. Z. Anat. 91, 229 (1929). Po~LE, E. u. E. STI~mNO~R: Dtsch. iKsehr. Zahnheilk. 40, 306 (1922). PoPow, N . A . u . A . A . KUD~Z~WZEW (1) : Zur Frage fiber die Absonderung der ]3auchspeichel- drtise beim Schaf. Jber. Vet.-med. 59 I, 109 (1931). - - - ANu~ow (2) u .a . : Physiologic des Schafes. Moskau 1931. - - - - u. W. K. KRASOVSKu (3): Arch. Tiercrnahrg u. Tierzucht 9, 243 (1933). P~ECL, F,: Pflfigers Arch. 61, 359 (1895).
R A s ~ x o w , L. P.: (1) Pflfigers Arch. 219, 391 (1928). (2) Pfliigers Arch. 223, 156 (1930). (3) Russk. fiziol. 2. 9, 87 (1926); ]3er. Physiol. 44, 540 (1928). RASP, F. : Fermentforschg. 9, 50 (1926). R~UCH: Vet.-med. Diss. Giessen 1912. I~OCOZLXSKI u. STA~ZEWSKA: Acta ]3iol. exper. (Warzawa) 1, 1 (1928). 1%OSE~FEnD, E.: Diss. Dresden-Leipzig 1908. Ross, J . CLV~C~S: Austral. vet. J . 7, 122 (1931).
S ~ T S C ~ N ~ , K. S. : Pflfigers Arch. 230, 680 (1932). SC~AL~:, A. T. and R. S. A ~ D o N : North Dakota. Agricult. exper. State ]3ull. 216, 64 (1928). Sc~X~rr, E . : Magen und Darm des Wildschweines. Z. Anat. 95, 326 (1931). SCHEVN~T, A. : (1) ]3Av~-Festsohrift (Schaper- Hannover), 1929. S. 297. (2) Pfliigers Arch. 109, 145 (1905). (3) Pflfigers Arch. 139, 131 (1911). - - u. W. G R ~ E ~ : Internat. iVfschr. Anat. u. Physiol. 23, 335 (1906). ~ u. FR. W. KRZYWANEK: l~flfigers Arch. 223, 472 (1929). - - - - u. K. Z ~ r E ~ A ~ N : Pfliigers Arch. 223, 453 (1929). ~ u. A. SC]~T]~E: Der Ablauf der Magenverdauung des normal gefiitterten und getrankten Pferdes. Jena: Gustav Fischer 1913, - - u. A. T R A U T ~ t ~ : Pflfigers Arch. 192, 1, 33 (1921). SCm]~BT~C~, M.: Handbuch fiber Ern/ihrung und des Stoffweohsels der land- wirtschaftlichen Nutztiere, ]3d. 2, S. 321. 1929. S r ]3.: Arch. Tierern/~hrg 9, im Er- scheinen (1933). SC~6D]~ , W. : Z. Tierziiehtg 16, 291 (1929). SCn-WARTE, L. ~ . : Unver6ffentlicht.

Einleitung. 577
Zit. nach If. ill. DUKES. J. amer. vet. reed. Assoc. 77, 225 (1930). ScnwA~z, G.: Med.-vet. Oiss. Oiessen 1914. S0~WA~Z, C. n. GABRIEL: Pfliigers Arch. 213, 814 (1926). - - u. B. IfE~R- ~N~r t)fliigers Arch. 202, 475 (1924). - - u. If. KAPL~: Pflfigcrs Arch. 213, 592 (1926). - - u. H. ST~.I~TLEC~NER: Bioohem. Z. 156, 130 (1925). SEVFFERT, R. U. P. TmE~r Beitr. :Physiol. 2, 147 (1923). SI~r Dtsch. Arch. klin. ~ed. 51, 472 (1893). SO~ME~: Vet.-med. Diss. Giessen 1913. STXLFO~S: Arch. Tierheilk. ~4, 525 (1926). S~AMBX]~: Vet.-med. Oiss. Bern 1910. STA~, If. E.: J. of biol. Chem. 54, 43 (1922). STEI~CHA~rF: Diss. Tier~rztl. Ifochsch. Berlin 1921. STEIZ~ETZ~, X.: l%rmentforsch. 7, 229 (1924). - - u. K. CAIT~A~m: Pflfigers Arch. 213, 595 (1926). STIO~En, R. : (1) Arch. Tierern~hrg n. Tierzucht 4, 613 (1931). (2) Bildtmg 2~, It. 1 (1933). ST~ELKOW, A.: Zoo!. Ariz. 94, 37 (1931). Su~, T.P. : Anat. Rec. 34, 341 (1927). - - I{. BLV~E~CTHAL, E. If. SLIFE~, E. C. I I E ~ . ~ and C. C. WAhoo: Physiologic. Zool. ~, 191 (1932). McSw~EY and W~DGE: J. of Physiol. 6~, 350 (1928).
TA~CG~: Pfliigers Arch. 63~ 545 (1896). TA~CZE~, J . : Pfliigers Arch. 213, 563 (1926). TE~VEX, J.: Z. mikrosk.-anat. Forsch. 21, 462 (1930); 18, 71 (1929). TI~OFEJE~'F, N .W. : Arch. Verdgskrkh. 52, 304 (1932). ToMAto, R. V. : Tijdschr. ])iergeneesk. l~ef. Jber. Vet.reed. ~0 I, 109 (1930). ToVSSAI~r H. : Arch. Physiol. norm. et path. 7, 141 (1875). TSC~ASSOW~CI- KOW, N.: Anat. Anz. 61, 417 (1926); 65, 28 (1928). T~AU~A~r A.: (1) Pfiiigers Arch. 211, 440 (1926). (2) Arch. Tierern~hrg u. Tierzueht 7, 216 (1932). (3) Arch. Tierern~ihrg u. Tier- zucht 7, 400 (1932). (4) Arch. Tierern~hrg u. Tierzuoht 9, 19 (1933). (5) Arch. Tierern~hrg u. Tierzucht, im Erseheinen 1933. (6) Arch. Tierern~hrg u. Tierzucht 9, 178 (1933). - - u. If. ALBRECht: Arch. Tierheilk. 64, 93 (1931). - - u. J. SC~MI~T: (1) Arch. Tierern~hrg u. Tierzucht 7, 421 (1932). (2) Arch. Tierern~hrg u. Tierzucht 9, 1 (1933). (3) Arch. Tier- ern~hrg u. Tierzucht 9, 10 (1933). (4) Beobachtungen an einer vegetabiliseh normal sich er- n~hrenden, abet nicht wiederk~uenden Ziege. Arch. Tierheilk. 65, 559 (1932). TX~IE]r H. J. : Z. vergl. Physiol. 4, 305 (1926).
USTJA~CZEW: Bioohem. Z. 4, 154 (1907). USUE~.LI, F.: (1) Arch. Tierern~hrg u. Tierzucht 3, 4, 368 (1930). (2) Profilassi. l~iv. mens. ~Pat. com E. 6, 7 (1933).
VE~zs :F.: Klin. Wschr. 12, 489 (1933). - - u. E. v. KOKXS: Pflfigers Arch. 217, 397 (1927). VILS~I~, 1~.: C. r. Soc. Biol. 1)aris 83, 239 (1920). Vo~cx, If. J . : Z. vergl. Physiol. 9, 685 (1929). - - u. I{. t ). WO~.VEKAM~: Tijdschr. nederl, dierkd. Ver.igg 1, 144 (1929).
WE~DT, I{.: Erg. d. inn. IVied. 42, 213 (!932). WE~r If.: IVied. Diss. Giessen 1911. WES~E~, J. : (1) Die Physiologie und ~Pathologie der Vorm~gen beim ]~inde. Berlin: Richard Schoetz 1926. (2) Berl. tier~rztl. Wschr. 46, 895 (1930). (3) Berl. tier~rztl. Wsehr. 46, 397 (1930). (4) Berl. tier~rztl. Wsohr. 47, 337 (1931). WETZX~: t~OUX' Arch. 114, 65 (1928). WIESE, C. : Vet.-med. Diss. Giessen 1912. WIU, I~O, E.: Diss. Landw. Ifochseh. Berlin 1933. WILLSTXTTE~, t{., F. IfAVXOW~TZ U. F. ME~E~r Ifoppc-Seylers Z. 140, 203 (1924).
ZX~X~E~A~r162 A.: Term6szettudomanyi K6zl6ny Potfuzel 63, 1 (1931). ZI~E~MAZ;~, K . W . : Erg. Physiol. 24, 281 (1925).
Einleitung. I n d e r P h y s i o l o g i c de r V e r d a u u n g d e r S~uge t i e r e , ganz b e s o n d e r s d e r
H a u s t i e r e , s ind sei~ K r i e g s e n d e n n s e r e K e n n t n i s s e w e s e n t l i c h b e r e i e h e r t
w o r d e n . G a n z b e s o n d e r e s I n t e r e s s e i s t den Wiederki~uern z u g e w e n d e t w o r d e n .
D e r Mechanismus der Rumination (STIGLEa), z u m Tei l a u c h d ie lno~or i sehe
F u n k t i o n d e r e inze lnen M~gen (MANoOLD u n d KLnlN, STIG~,Ea, T~AUT~A~'N),
k a n n h e u t e a ls z i e m l i e h g e k l ~ r t ge l t en . D a g e g e n s ind t ibe r die c h e m i s c h e n
F u n k t i o n e n des L a b m a g e n s i m Verg l e i eh z u m M e n s c h e n u n d H u n d n o e h r e e h t
be t r~ ich t l i ehe U n k l a r h e i t e n v o r h a n d e n . P i e r d u n d S c h w e i n s ind f a s t v o l l k o m m e n
v e r n a c h l i i s s i g t w o r d e n . Die V e r d a u u n g d e r L a b o r a t o r i u m s n a g e r g e w i n n t ers~
n e u e r d i n g s e r h S h t e s In~eresse , da s K a n i n c h e n als P e l z t i e r m e h r aus p r a k t i s c h e n
Asher-Spiro, Ergebnisse der Physiologte und exper. Pharmakologie. 35. 37

578 W. LEI~KEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~ugetiere.
Grtinden, die tibrigen Nager, besonders die weisse l~atte, aus vergleichend physiologischen Griinden. Uber die Verdauung bei den wildlebenden Saugern des Landes und des Wassers wird naturgem~ss unser Wissen noch geringer sein und nicht viel Neues der umfangreiehen Darstellung yon BIEDERMAh'N im tIandbuch der vergleichenden Physiologie yon WINTEaSTmN zugefftgt werden kSnnen.
Damit ist auch gleich angegeben, welche Tiere in der vorliegenden Dar- stellung besonders berticksiehtigt werden k6nnen. Bezfiglich der ~ilteren Lite- ratur muss in sehr vielen F~illen auf die bereits vorliegenden umfangreiehen Zusammenstellungen in versehiedenen tIandbtiehern verwiesen werden.
A. Mundverdauung. L Nahrungsaufnahme.
Die 0rgane der Nahrungsaufnahme sind die Lippen, die SchneidezShne und die Zunge. Welches Organ in hervorragendem Masse gebraueht wird, ist nach der Tierart und nach dem Zustand der Nahrung verschieden.
Die Fleischf~resser nehmen das Fleisch vorwiegend mit den Schneide-, Eck- und bisweilen noch mit den vorderen Backenz~hnen auf, indem sie unter Festhalten der Nahrung mit den Vorderbeinen Stficke herausreissen bzw. abbeissen. Breiige Nahrung wird mit der Zunge ,,aufgeleckt". Auch der Maulwu'r[ (Talpa europaea), der sich ausnahmslos yon kleinen Tieren (l~egen- wtirmern, Arthropoden, seltener Mguse und V5gel) ernghrt (HAUCgnCORNE), eriasst seine Nahrung mit den Zghnen (HAuc~ECOR~). Das Schwein nimmt bei der Aufnahme vorgesetzter Nahrung mehr die Zunge zur Hilfe; dagegen beisst es au~ der Weide die Pflanzen mit den Schneidez~hnen ab oder reisst sie ab (ELLENBERGER und SCttEUNERT). Das Rind umsehlingt die Pilanzen zuni~chst mit tier vorgestreckten rauhen Zunge und ftihrt sie dann zwischen Schneidezi~hne und Dentalplatte, um sie abzubeissen oder abzureissen. Auch die kleinen HauswiederkSuer gebrauchen vorwiegend die Zunge. Bei der Gira//e spiett die Zunge bei der Nahrungsaufnahme eine hervorragende Rolle. Die Giraffe holt mit der kr~ftigen langen Zunge aus den Kronen der Akazien- und anderer Bi~ume der afrikanischen Steppe die BlOtter und kleinen Xstehen herab (DosLEI~). Das P/erd bedient sich mehr der Lippen und Sehneidez~hne beim Ergreiien der Nahrung. Der Ele/ant besitzt in seinem Rfissel ein aus- gezeichnetes Hilisorgan bei der Nahrungsauinahme.
Die Fltissigkeiten werden yon den Wiederkguern, Einhu/ern und Schwein durch Saugen aufgenommen. Wird noeh Luft neben dem Wasser eingesaugt, so entsteht das Schliirien, wie es beim Schwein bei nicht vollkommenem Ein- tauchen der Lippenspalte zu beobachten ist. Dieses kann ngmlich seine grosse Lippenspal~e beim 0ffnen vorne seitlich nicht lest schliessen. Der Hund 10ffelt das Wasser mit der dazu dureh Muskelwirkung 15ifelfOrmig geformten Zunge in die MundhShle hinein, w~hrend die Katze ihre mit Staeheln besetzte

Das Kauen. 579
Zunge in die Fltissigkeit eintaueht und beim Zurfiekziehen der Zunge die anhaftende Flfissigkeit in der Maulh6hle abstreift. In tthnlieher Weise erfolgt die Getrankeaufnahme bei den Nagern. Der Maulwur/sehleekt naeh HaVCHE- CO~Nn das Wasser wie der t tund.
IL Das Kauen.
Der Grad der Zerkleinerung der Nahrung, die Grfindlichkeit des Kauens zeigt sich yon manchen Faktoren, besonders yon der Art der Nahrung, yon der Art der Weiterverarbeitung der Nahrung im Verdauungskanal abh~ngig un4 ist natfirlich nach der Tierart versehieden. Mit INGo KRVMmEGEL kann man zwisehen ,,Reiss" oder -,,Beiss"tieren (Fleischfressern) und ,,Kau"- oder ,,Malm"-tieren (Pflanzeniressern) unterscheiden. Morphologiseh kommt dieser funktionelle Unterschied zum Ausdruck in der Ausbildung tier Kiefer, beson- ders tier Unterkie/er und in der Ausbildung der Molaren. Bei den reinen Fleisch- /ressern sind die Molaren, mit Ausnahme des unteren Reisszahnes, schwaeh ausgebildet; sehon bei den Allesfressern und in besonderem Masse be~ den Pflanzenfressern nehmen sie an Zahl und Gr6sse zu. Eine entspreehende An- passung zeigt der Unterkiefer in seiner L~nge, St~rke und in der Ausbildung des Gelenkes (HEssE, K~V~mE~EL). Je grSssere Rolle die Pflanzennahrung spielt, um so langer wird der Unterkiefer und um so spitzer die Winkelstellung seiner ~ste.
Auf den mehr oder weniger komplizierten Verlauf der Kaubewegungea bei den verschiedenen Pflanzenfressern ist bereits BIEOE~AN>r auf Grund der Untersuehungen yon LuBosc~ in seiner umfangreichen Darstellung aus- fithrlich eingegangen.
Der Grad der Zerkleinerung bei der Futteraufnahme ist bei den ver- sehiedenen Pilanzenfressern versehieden. Die wiederkauenden Pflanzenfresser kauen bei der Futteraufnahme ziemlieh oberfliiehlieh. Naeh SC~A*,K und AMADON kommen yon ganzen Haferk6rnern etwa 50%, yon ganzen Mais- kSrnern etwas mehr, zerkleinert in den Magen. Erst beim Wiederkauen (s. S. 605) der naeh einiger Zeit wieder in den Mund gebraehten Nahmng erfolgt ein grtindliehes Kauen. Die Niehtwiederk~uer maehen dagegen sehon bei der Futteraufnahme die Nahrung sozusagen ,,magenfertig" und kauen daher bedeutend sorgfiiltiger. Das Rind nimmt 21/4 kg Haler mit 23 Bissen in 10 Mi- nuten auf (DUKES), der Esel dagegen naeh GAUDENZI 2 kg Haler in 18 Minuten.
Die Zahl der Kaubewegungen soll sich nach GAUDENZI und CIHCClA umgekehrt wie das Gewieht der Tiere verhalten. Dadurch wfirde das versehie- dene Verhalten der grossen und ]deinen Wiederkiiuer erkli~rt werden. Die die Zahl der Kaubewegungen bestimmenden Faktoren sind mannigfaltig. Mit der Feuchtigkeit des Farters seheint die Zahl der Kaubewegungen im Durch- schnitt zuzunehmen (CIelCCih, G. Sc~wA~z), allerdings berichtete SCHWArtZ yon entgegengesetzten Beobachtungen beim Pferde. CiPiCClA maehte be[ seinen
37*

580 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S/iugetiere.
Untersuehungen an Tieren des Zoologisehen Gartens in Rom noeh die Beob- aehtungen, dass die Zahl der Kaubewegungen um so gr6sser ist, je behender die Tiere sind, je niedriger die Temperatur ist, und je diehter das Fell ist. Die anthropoiden Alien ftihren bei gleieher Nahrung zahlreiehere Kaubewe- gungen aus als der Mensch. Bei Tieren mit reiehlieher Speiehelabsonderung iand CIPICCIA eine geringere Zahl yon Kaubewegungen als bei solehen mit geringerer Speiehelabsondemng.
Bei den Herbivoren erfolgt das Kauen stets einseitig. Dieses ist dutch die anatomischen Verh~ltnisse bedingt. Infolge der Enge des Unterkiefers gegentiber dem Oberkiefer k6nnen sieh die Molaren auf beiden Seiten nieht gleiehzeitig gegentiberstehen. Daraus folgen aueh die Seitwiirtsbewegungen. Bei dem griSssten Teil der Pilanzenfresser, sowohl bei den wildlebenden als aueh bei unseren gesamten domestizierten I-Iaustieren (G. So.wAltZ) wird ein Bissen entweder mindestens mehrere Kiefersehliige oder sogar bis zum Absehlueken nut anf einer Seite gekaut. Naeh OST~OF soll yon Pferd, Ziege, Sehaf und Rind die linke Seite besonders bevorzugt werden. Alternierend, d. h. naeh jedem Kiefersehlage die Seite weehselnd, kauen nut die kamelartigen Wiederkiiuer.
Das Kauen beim Wiederkauen unterseheidet sieh yon dem Kauen bei der Futteraufnahme dutch die grosse Gleieh- und Regelmiissigkeit, dureh den taktm~ssigen Rhythmus (G. Sc~wAaz).
Zu erwghnen ist noeh die beim Kauen unterstfitzende golle der Zunge (HEssE). Ausser dass die Zunge mit Hilfe der Wangen die Bissen zwisehen die Backzghne schiebt, ist sie imstande dureh Druek gegen die harten Gaumen weiehere Nahrungsteile zu zerdriicken. Den besonders bei den Pflanzenfressern stark ausgebildeten transversalen Gaumenfalten mit den beim Rinde rachenwi~rts geriehteten Staeheln (MAgTIN) kommt vielleieht hierbei eine besondere funk- tionelle Bedeutung zu (LAFoND). Ausserdem spielt der harte Gaumen noeh eine Rolle bei der Nahrungsaufnahme und beim Sehluekakt.
III. Die Speichelsekretion. Die Speicheldriisen. Der Speichel ist das gemischte Sekret siimtlicher
Drtisen der Mundh~hle. Die gr6sseren Drtisen sind: die Glandula parotis, Glandula submaxilIar~s, s. mandibularis, Glandula sublingualis parv4canalaris und bei den Wiederkguern noeh die Glandula sublingualis grandicanalaris. Bei den einzelnen Tierarten ist das Sekret der einzelnen Drfisen versehieden. Die Ohrspeicheldriisen sondern fast bei allen Tieren ein rein ser(Sses Sekret ab, bei den Carnivoren aber naeh. METZNER ein muk6s-ser~Sses Sekret. Dass das ser6se Sekret bei der vegetabilisehen Nahrung besonders notwendig zu sein seheint, geht aus der betrttehtliehen Entwieklung der Parotis bei den Pflanzen- fressern hervor. Beim Biber ist die Parotis 20real grOsser als die Submaxillaris (K~v~mEGEL). Die Submaxillar4s ist bei den meisten Sgugern ser6s und mukSs, SO bei den herbivoren Haustieren, Fledermgusen und einzelnen Nagern (Muriden),

Die Speichelsekre~ion. 581
bei den Carnivoren ist sic muk6s, ser6s bei den meisten Nagern (K~UMBIEGEL). Die Sublingualis parvicanalaris liefert ein gemisch~es Sekret, die Sublingualis grandicanalaris ein vorwiegend mukSses. Im grossen und ganzen wird die Konsistenz des Speichels auch sehr yon der Art der Nahrung bestimmt.
Sekretion des Speichels. Die Sekretion des Speichels erfolgt in der Haupt= sache dureh mechanische oder chemische Reizung der Mundschleimh~ut durch die Nahrung (unbeding~e Reflex@ Bei den herbivoren Tieren and auch beim Schwein sind die mechanischen Reize die wichtigsten. Die psyehische Sekretion tritt jedoch gegenfiber der nach direkter l~eizung betri~chtlich zurtick. Deutlich vorhanden ist die psychische Sekretion bei Mensch, t Iund und nach den neueren Untersuchungen yon TRAUTMANN (b) noch beim Schwein. Bei den kleineren Wiederki~uern scheint eine geringgradige psychische Sekretion bisweilen m0g- lich zu sein (ScHEUNBRT und T~AUTMANN, TRAUTMAN?r und ALBRECHT); aller- dings neigen in sp~teren Untersuehungen SCHEU~ERT and Mitarbeiter mehr dazu, die geringe Steigerung der Sekretion bei Vorhalten der Nahrung nur auf Zungen- und Lippenbewegungen der Tiere zurfiekzuftihren. Beim Rind und beim P/erd ist eine psychisehe Speiehelsekretion sieher nieht vorhanden (COLIN, SCnEUNERT und TRAUT~ANN). Dieser Unterschied im Zus~andekommen der psyehisehen Speiehelsekretion (bedingter Reflex) weist auf eine versehiedene Bedeutung der Grosshirnrinde bei den Herbivoren einerseits und bei den 0mni- und Carnivoren andererseits hin. Die StiLrke der Sekretion haupts~tchlieh der Parotis s~eht in engem Zusammenhang mi~ dem Kauakt. Bei den einseitig kauenden Herbivoren zeigt sieh eine Abhangigkeit yon der Kausei~e. Bei den Wiederkguern ist auf der Kauseite die Sekretion der Parotis gegentiber der anderen Seite bedeutend gesteigert [ScHEVNERT (1), SCHEU~ERT und KRZYWANEK und ZIMMEn~XANN, TRAUTMANN und ALBRECHT]. Beim Pferde sezerniert bisweilen sogar nut die Parotis der Kauseite, wi~hrend die Sekretion der anderen Parotis vollkommen sistiert (ScHEV~ERT und TRAVTMAX~). Auch beim Schwein, das nicht ausgesprochen einseitig kaut (TRAuTMA_~N), wird nach den Untersuchungen yon TRAUT3IANN (2) mit permanenter Parotisfistel mit Untersuchung des Kauaktes die Sekretabgabe der Parotis entweder be- tr~chflich vermindert oder vollkommen eingestellt.
Die Menge des abgesonderten Speichels h~ngt nattirlich weitgehend yon der Beschaffenheit der Nahrung ab. Im Durchschnit~ betri~gt die pro Tag abgesonderte Gesamtmenge beim Menschen 500--1000 g, beim Pferd 40 kg, beim Rind 60 kg. Je troekener und rauher die Nahrung ist, um so grSsser ist die Speichelmenge, und um so geringer ist die Speiehelmenge, je wasserreicher oder dtinner die Nahrung ist.
Bei Aufnahme yon Fltissigkeiten wird im Mlgemeinen kein Sekret ab- gesondert. Doch scheint die Milch bei einigen Tieren eine Ausnahme zu machen. Bei Ziegen]i~mmern mit permanenten Parotisfisteln konnten TRAUT~ANN und ALBRECHT bei Milehnahrung etwa das 4--10fache der Ruhesekretion, fLir 1,0

582 W. LENKEIT: I~euere Ergebnisse der vergl. Physiologie der Verdauung der Saugetiere.
bis 1,5 Liter Milch 0,1--0,4 ccm Parotisspeichel feststellen. Bei Hunden konnten Ac~RAP und POD~OPAEW sowohl nach Verabfolgung yon Milch als auch yon Zuckerwasser eine Sekretion der Submaxillaris und der Parotis herbeiffihren, und zwar war die Speiehelmenge aus der Submaxillaris 3mal grSsser als aus tier Parotis. Beim Schwein bleibt wieder die Sekretion der Parotis naeh Milch- aufnahme aus (TRAuTMAN~). Die Speichelsekretion nach Milchaufnahme wird nicht durch das Fet~ und das Casein der Milch angeregt; auch sfissliche Molken ffihren n~imlich zur Sekretion (Ac~RAP und PODKOPAEW). Es scheint auch, dass die Schleimdrfisen und die gemischten Drfisen (Glandula submaxillaris) durch die Milch ganz besonders zur Sekretion angeregt werden, bei serSsen Drfisen (Glandula parotis) dagegen in geringerem Masse oder fiberhaupt nicht [T~AuT~IANN (2)7. Die mit schleimigem Speichel vermengte Milch koaguliert im Magen lockerer und wird daher leichter verdaut.
Die wShrend einer Mahlzeit abgesonderte Speichelmenge ist um so geringer, ]e schneUer die Nahrungsau/nahme erfolgt. Ein Versuch yon T~AUT~AN~ (2) am Schwein mit permanenter Parotisfistel zeigt dieses sehr deutlich. Nahm ein Schwein ein Futtergemisch (350 g Sehrot, 100 g Kartoifelfloeken, 50 g Kleie) in 80 Minuten auf, so sonderte die Parotis durch die Fistel in dieser Zeit 207,4 ccm Speichel ab, w~ihrend aber das gleiche Tier bei Aufnahme des gleichen Futters in nur der H~ilfte der Zeif nur 76,1 ccm Sekret entleerte. Die in der Zeiteinheit (5 Minuten) w~hrend der Mahlzeit abgesonderte Speichel- menge erreicht im Durchschnitt in den zweiten, oft schon in den ersten ffinf Minuten das Maximum, um dann allm~hlieh abzunehmen [Sc~EUN~I~T und T~AVT~A~, TI~AUT~A~N (2)J. Die Glandula mandibularis und Glandula sublingualis monostomatica des Sehweines sondert ihr Sekret yon Beginn bis zum Ende der • in den einzelnen Zeitabschnitten anschei- nend gleichm~issiger ab (HENcKE). Die Speichelabsonderung durch die grSsseren Drfisen erfolgt bei den Carnivoren, den Omnivoren und den Einhufern nut zur Zeit der I~ahrungsaufnahme; bei den Wiederk~iuern sezernieren dagegen die Ohrspeieheldrfisen und die grossen ventralen Baekendrfisen kontinuierlich, auch zwischen den Mahlzeiten.
Es sind daher bei den Wiederkiiuern neben der Speichelsekretion wi~hrend der Futterau/nahme die Sekretion wiihrend des Wiederkauens und bei den kon- tinuierlich sezernierenden Driisen (Parotis, ventrale Backendrfisen) die Ruhe- sekretion zu unterscheiden. An jeder Sekretionsart hat den gr5ssten Anteil
die Parotis. Die Ruhesekretion der Parotis kann recht betriichtlich sein. Beim Rinde
kann nach alten Un~ersuchungen yon COLIN die Sekretmenge 200--500 g in 15 Minuten betragen, beim Scha/erreicht nach SC~VNERT, Kl~ZYWANEK und Z I ~ ~ die Sekretmenge bei Berficksichtigung der KSrpergrSsse oft die beachtliche HShe yon 20, 30 und sogar 40 g in 10 Minuten. Die Parotis der Ziege sondert im Durchschnitt noch mehr Sekret ab als die des Schafes (T~vT-

Die Speichelsekretion. 583
~IANN und AI, BRECHT). Im grossen und ganzen sind die individnellen Schwan- kungen in der Ruhesekretion ziemlich gross [ScEEUN~RT (1)]. Bei Ziegen- ]gmmern ist w~hrend der Sgngeperiode die Ruhesekretion der Parotis sehr gering; erst mit ]Jbergang zum Rauhfutter steigt sie innerhalb 14 Tagen um das 101ache (TRAuTMA~N und AT,BRECET).
Es liegt nahe, an einen ursgchliehen Zusammenhang der Dauer- (Ruhe-) Sekretion mit der Pansentatigkeit zu denken. Auf diese Weise erhalten n~mlich die Vormggen zur Feuehterhaltung des Inhaltes die notwendige Fltissigkeit und zur Neutralisierung der sauren Ggreprodukte das notwendige Alkali. Die Versuche, die Danersekretion dureh kfinstliche Anderung der Reaktion des Panseninhaltes End dutch ErhShung des Panseninnendruekes zu beein- flussen, waren ergebnislos (ScEEUNE~T, KRZYWANEK, ZIM~ERMAI~N). Beob- achtnngen yon Tt~AUTMANN und ALBREClZT weisen auf eine BeziehEng zwisehen Bewegung des Pansens End der Speichelsekretion bin; bei Abnahme der Zahl der Pansenbewegungen seheint anch die Sekretion abzunehmen End bei Steige- rung der Pansenbewegungen zuzunehmen. Es ist nieht ausgeschlossen, class eine eingehende Prfifung dieser Beziehungen uns der Anfklgrung der Ursache der Dauersekretion n~her bringt.
Bei der Nahrungsau[nahme sezerniert yon den Parotiden nur die Parotis der Kauseite stgrker~ w~hrend die andere in Ruhesekretion verbleibt. Nur nach Aufnahme yon ~tafer und Rfiben sezernieren beide Parotiden vermehrt (Sc~uN~RT).
Beim Wiederkauen'sind nur die kontinuierlich sezernierenden Drtisen in Tatigkeit. Die Glandula mandibularis sezerniert dann nieht (ScHEUNERT und TRAUT~ANN). Von den 0hrspeieheldrtisen sonderi, aueh nur diejenige der Xauseite in erhShtem Masse ab.
Die Zusammensetzung des Speiehels. Der gemischte Mundspeichel ist bei allen Tieren das wasserreiehste Sekret (durehsehnittlich 99% Wasser). Naeh der Darlegung fiber die mukSsen und serSsen Speieheldrfisen wird der Speiehel der Fleisehfresser einen hSheren Gehalt an Mucin und einen geringeren an Eiweiss haben als der Speiehel der Pflanzenfresser.
Die Reakfion des gemisehten Speiehels ist gegen Laekmus alkalisch, und zwar am st~rksten bei den Wiederkguern. Die Menge des titrierbaren Alkalis im Speiehel, die ,,Alkaleseenz", entspricht
beim Menschen : nach DIs, MI~G~s~: 18,0--63,0 mg ~ a O g pro 100 ccm Speichel, nach Com~ (zitiert nach I~OSS,~ASN): ira Mittel 15,4rag pro 100ccm Speichel;
beim P/erd im Parotisspeichel: nach GO~SCHA~,K: 160 240 mg ~aOH pro 100 ecru Speichel, nach Sc~vNx~w und TaAVW~WA~r: 0,23---0,32% Na~CO~-L6sung;
beim t~ind im Gesamtspeichel: naeh MA~KOF~: 0,58----0,65 % ~a~COs-LSsung;
beim Scha] im Parotisspeicheh nach ScHv, v~v.~r und T~AUr 0,56~0,77% Na~COa-LSsung , nach S c ~ U ~ T , X~zYw~N~K und ZI~V~E~A~N: 1,26% ~attCOs,

58~ W. LEI~XEIT: Neuere Ergebnisse] der vergl. Physiologie der Verd~uung der 8augetiere.
~m kontinuierlich sezernierten Gesamtspeiohel (yon Parotis und Baokendrtisen durch Oesophagusfis~el gewonnen) naoh So~Eu~s.~ und IZ~zywA~]sK fiir 30 ecru Speichel 40 oom n/10 tt2SO 4 verbraueht;
bei tier Ziege im P~rotisspeichel: naoh T ~ A v ~ A ~ und ALmz~c~T: 0,54---0,77% NaeC03-L6sung.
In dem zuerst sezernierten Speichel fand GOTTSC~AL~ beim Pferde die Alkalitiit grSsser als in den zuletzt abgesonderten Portionen; desgleichen trat dureh Applikation yon S~ure in das Maul eine Zanahme der Alkaleseenz ein. Mit der Intensitat der Erregung nimmt die Gesehwindigkeit der Speichel- absonderung zu und damit im Durchsehnitt der Asehegehalt und so die Alkali- t~tt. Der Parotisspeiehel der Kauseite hat nach den Untersuehungen yon SCHEUNERT und ~]~I~AUTMANN am Pferde stets eine hShere Alkalit~it als der der anderen Seite.
Der Speiehel wirkt durch seinen Gehalt an Bicarbonaten, besonders yon iNaHCO~, freier C02 und Eiweiss wie eine PufferlSsung. Die Reaktion des Speichels wird vorwiegend dureh das Verhiiltnis yon C02 zu NaHC0~ bestimmt. Beim Entweiehen der Kohlens~ure (z. B. naeh zu langem SIehen) wird sich natfirlieh die Reaktion naeh der alkalischen Seite versehieben (Po~LE und ST~mNQE~).
Bei der Bestimmung der aktuellen Reaktion ist diese Tatsaehe sehr zu berfieksiehtigen.
Die aktuelle Reaktion weist nach der Tabelle 1 die gleiehe Reihenfolge wie vorhin auf. Der gemischte Speiehel des Mensehen ist mit der niedrigsten p~-Zahl sehwaeh sauer bis neutral, der der Wiederk~iuer mit der hSehsten p~-Zahl ausgesproehen alkaliseh.
Auffallend hoeh sind gegentiber den Befunden der anderen Autoren die Werte fiir Pferd und Rind yon Ci~ZASZCZ und SeeLECttTSbWNA. Es w~ire zuniiehst daran zu denken, dass dieser Untersehied durch eine hinsiehtlich der Nahrungsaufnahme zeitlieh versehiedene Entnahme des Speichels bedingt ist. W~ihrend C. Sc~wA~z und H E ~ A ~ stets den einige Stunden naeh der Fiitterung entnommenen Speiehel untersuehten, gewannen C~nz~szcz u~d S c ~ E e ~ b w ~ den Speiehel in den iiberwiegenden Fiillen w~hrend der Fiitterung; allerdings liegen nach den gleichen aueh die p~-Werte des w~ihrend der Naehtruhe entnommenen Speiehels sowohl beim Pferd als aueh beim Rind mit p~ 8,60 und 8,55 und 8,80 innerhalb der Variationsbreite der w~ihrend der Ffitterung yon ihnen gefundenen Werte. Leider sind die Untersuchungen w~thrend der l~aehtruhe beim Pferd nur einmal und beim Rind nur zweimal durehgefiihrt worden, und fehlt a~ch jede Angabe der Zeit der Speichelent- nahme naeh der F~itterung. Beim Mensehen ist n~mlieh die Reaktion des Speiehels in nii.chternem Zustand und vet der Mahlzeit sauer, nach der Mund- reinigung sehwach alkalisch.
Es ist aber wahrseheinlieh, dass die abweiehenden Werte yon C~InZASZCZ und S c ~ e ~ n 6 w ~ A auf Entweiehen yon CO~ zurtiekzuftihren sind.

Die Speichelsekretion. 585
Tier~rt
Tabelle 1. A k t u e l l e R e a k t i o n des Spe iche l s (p~)~.
~ensoh ..........
~ I l l n d . . . . . . . . . . .
l{atze . . . . . . . . . . .
Schwein . . . . . . . . . .
1)ferd . . . . . . . . . . .
I~ind . . . . . . . . . . . .
[w~hrend des Wiederkauens]
Schaf . . . . . . . . . . .
P~
6,63 (5,93--6,86)
6,6 (5,75--7,05) 5,25--7,86
7,56 (7,34---7,80)
7,5
7,5
7,32 (7,15--7,44)
7,37
7,56 7,31--7,80
7,42 8,56
8,49--8,62
8,10 7,99--8,27
8,80 8,55---8,90
[7,90] 8,23
8,12--8,46
Autor
C. SCmYARZ und B. t IEl~l~gN
STA~
1)O}tL~ und ST~Br~(~E~
C. Scm~r~z und B. I-Iv.gI~ANN
L. tt. SC~WA~T~
L. It. SC~WART~
C. S c ~ z und B. I-I]~g~A~l~
L. It. S c m v ~ l ~
C. SCHWA~Z und B. :H:~r~MA~r
L. It. SC~w~T~ C~z~szcz und SCm~v, CH~L6W~A
C. Scn~c~z und B. H E r ~ r
Cm~zaszcz und SC~rLECI~T~6W~
[C~z~szcz und Sc~m]:cH~6w~] L. H. S c m ~ A ~
N. A. t)o~ow, A. Kvm~awz~w und Agmcow
Immerhin zeigen die unter gleiehen Bedingungen erhaltonen Befunde, dass die Art des anfgenommenen Futters (Trockenfutter, Grfinfutter) ohne Einfluss (C~zAszez und SeI~L~CgTn6WSA) is~. ~ber die Reaktion des Speichels bei verschiedenen Tieren vor, wiihrend and nach der Nahrnngsauf- nahme fehlen bisher systematisehe vergleiehend e Untersuehungen.
Das spezifische Gewisht des Speichels (Tabelle 2) unterliegt sowohl bei den einzelnen Tieren wie aueh bei den einzelnen Drtisen grossen Sehwankungen, entspreehend denen der einzelnen Bestandteile. Eine Regelmassigkeit weisen diese Schwankungen der Eigenschaf~en und in der Zusammensetzang des Speichels nieht auf. Im allgemeinen sind naeh den Untersuehungen yon SC}t~V~E~T nnd TRAUTMANN am Pferd and am Schaf, yon TRAUTMANN (2)
am Schwein, in dem zu Beginn der Nahrungsaufnahme abgesonderten Speichel die Werte ftir das spezifische Gewicht f~ir Trockensubstanz und Asche am hSch- sten und fallen dann allm~hlieh ab. Der zuletzt sezernierte Speichel ist arm an gel6sten Bestandteilen. Die Versuche yon BAS~E~ am Hand zeigen dasselbe. Der Speichel der Parotis der Kauseite ist, entsprechend der intensiven
1 STAIR und SO.WASTE bestimmten colorimetrisch, die fibrigen Autoren elektrometrisch.

586 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Sgugetiere.
Tabelle 2. Zusammensetzung des Speichels.
Tierart
Ziege . . . . .
Schaf . . . .
Rind bei Trockenfutter
Griinfutter
Pferd bei Trockenfutter
Griinfutter .
Schwein �9 . �9
Mensch . . . .
Speichelart
Parot~
Parot~
Mandibularis
Gemischter Speichel
Gemischter Speichel
Parotis
Parotis
NIandibularis Sublingualis
Gemischter Speichel
Spez. Gewicht
1,0019 bis 1,0634
1,0094 bls 1,0115
1,0028 bis 1,0088
1,0017 bis 1,0050
1,0020 bis 1,0080
1,0015 bis 1,00~5
i 1,005 bis 1,007
11,0019 bi,~ 1,0088
),9996 b~ i 1,0012
1,002 bis 1,008
Trocken- Asche substanz
% %
0,71 bis 0,62 bis 1,96 1,62
1,078 bis 1,228
1,1 bis 0,75bis 1,25 0,9
0,4 bis 0,05 bis 1,0 0,2
0,8 bis 2,2
0,4 bis 1,4
0,5 his 2,0
0,3 bis 1,3
0,446 bis 1,591
0,5 bis 1,3
0,3 bis 4,6 0,68
0,733
0,010 bis 0,026
0,014 bis 1,177
0,113 bisl0,020 bie 0,549 0,217
0,22 his 0,5
0,12 bis 0,36 I
0,164 ! 0,32
0,225 0,050
D,010 bis 0,046
~),064 bis 0,137
0,032 his 0,140
0,012 bis 0,278
0,016 bls 0,069
0,067
Untersucher
T~A~TMAWN und _An. BRECHT
N. A. PoPow, KVD~- AWZEW und ANu~ow
SCHEU~ERT und TRAUTMANN
8CamU~E~T und. TRAUTMANI~
CtmZASZCZ und SC~ECHTL6WlVA
CH~ZASZCZ und SCIILEC]:ITL6WNA
~OTTSC]YALI~
SOHEUNEI~T und TI%AUT~IKNN
T~XC~MA~ (2)
H. t{ENc~
CLiRK und SHELL, zit. n~ch BABKr~
Erregung, uach den Fests te l lungen am Pfe rd (ScRv, UNERT und TRAUTMANN)
reicher an Chloriden und an Asche, aber ~rmer an Eiweiss als der Speichel
der anderen Parot is . Im Dauerspeichel i r o n Parot is und sog. ven t ra len Backendrt isen, SGHEU-
NERT (1)] der Wiederkguer scheint die hohe Alkalit~it auf d e m Gehal t allein
yon Na (30, 25 % der Asche) und K (2,4 % der Asche) zu beruhen (ScHEUNEaT
und K~ZYWANEK) ; denn Ca und Mg waren beim 8chaf nicht vorhanden , l~ach-
zuweisen war noch COn, P20s und C1, dagegen nicht S0~. Der Dauerspe ichd des
Schafes ist darnach eine fast reine LSsung - - yon 1Na- und K-Bica rbona t bzw.
Carbonat mi t geringer Beimengung yon Chlorid und Phospha t . Zu erwghnen ist noch, dass das im Speichel des Menschen vo rkommende
Rhodankalium beim H u n d e sd ten , bei den fibrigen Haus t i e ren i iberhaupt
nicht gefunden wird.

Die Speichelsekretion. 587
Fermente. Der Speichel kann mehrere Enzyme enthalten: eine Diastase (Ptyalin), Maltase, Lipase und Oxydase. 0b nooh eine Peptase vorkomm~, wird sehr angezweilelt (BABKIN). Im Gehalt des Speiehels verschiedener Tiere an den einzelnen Fermenten, besonders an Ptyalin, sind be~r~ehtliehe Un~erschiede vorhanden. Bei den Speieheln ohne Amylase ist aber zu unter- scheiden, ob die Amylase tats~chlieh lehlt oder nur praktiseh fehlt, d. h. wegen ungtinstigen Bedingungen, z. B. PE nicht optimal, nieht wirken kann, oder ob sic nut zeitweise nicht vorhanden ist. Einen ziemlieh hohen Gehalt an vorhandener Diastase weist der Speiehel veto Menschen und vom A//en auf, und nach KNIP~R aueh der Speichel des Ele/anten. Endgfiltig naehgewiesen ist das Ferment auch im Parotisspeiehel des Schweines (STEINMETZER, T~AUT- ~tANN [2]), allerdings in geringeren Mengen als beim Mensehen (Lt)c~i~G), da- gegen nieht im Speiehel der Glandula mandibularis und Glandula sublingualis monostoma~iea (HENcKE). Beim Hamster konnte Se~EVNEaT aus der Parotis Diastase dureh Extraktion gewinnen.
Aueh dem Kaninchenspeichel kann wohl entgegen manchen ~lteren An- sichten (s. BAB~IN) nach den neueren Untersuchungen yon Jv~G, GAYDA, RASP und besonders yon F~sc~L und KA~N eine gewisse amylolytisehe Wirkung zugesproehen werden. Beim Meerschweinchen, Ratte und Maus konnte RAsr ein diastatiseh wirksames Ferment sieher feststellen, PATZSC~KE ebenfalls bei Ratte und Igel. Das Ferment der Parotis wurde bier starker gefunden als das der Submandibularis. Von BABKIN, 0P~N~E~MEa wird das Vorkommen der Diastase bei den l~agern als echtes Sekretionsprodukt der Speieheldrfisen ffir zweifelhaft gehalten.
Bei den fibrigen Tieren kann man sieher den Speiellel praktiseh als diastatiseh unwirksam ansehen. Bei den Carnivoren (Hund, Katze, Fuehs) wurde der Speiehel bereits yon G~TZNE~ U. a. und in neuerer Zeit yon S~Ex~- ~ETZER, BE~Z und L~CK~N~ amylolytisch unwirksam ge~unden. Aueh GA~E~ sah niemals beim Hund einen Abbau der St~rke bis zum Auftreten der Mal- rose; sondern die in wenigen Versuehen beobaehtete diastatisehe Wirkung ging nur bis zur Bildung yon Dextrinen, um dann stehen zu bleiben. Danaeh ist naeh GA~E~ zwar im I:Iundespeiehel die die St~rke veriliissigende" ,,Poly- amylase" ziemlieh inkonstant und in sehr geringer Menge vorhanden, w~ihrend die die Dextrine in Maltose tiberftihrende ,,Dextrinase" vollkommen fehlt. Allerdings konnte S A ~ Y T S C ~ bei lang dauernder Kohlehydraterniihrung im Speiehel des Hundes voriibergehend, veto 4. bis 7. Tage der Digit, das Auf- treten der Diastase nachweisen; dann nahm das Ferment allm~hlich ab und versehwand sehliesslieh vollkommen.
Es fehlt aber auch nicht in der neueren Zeit an deutliehen positiven Dias~ase-Befunden im Carnivorenspeiehel (SE~:~FE~T und TmEN, J~N6). Doch ist wahrseheinlieh der Speiehei in einem Tell der Versuehe auf nieht ganz einwandfreie Weise gewonnen worden, und es ist nieht ganz

588 W. LE~:E~r Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~uge~iere.
auszusehliessen, dass in diesen Fiillen das Ferment aus Leukocyten oder dem Blute stammt (GAYDA).
Beim P/erd konnte entgegen manehen ~lteren Befunden bei einer grSsseren Anzahl yon Untersuehungen eine diastatisehe Wirkung des Speiehels nieht festgestellt werden (Sc~EuNE~T und TaAUT~ANN, STEINMETZER, Lt~CmNG, B~z).
Zu gleichen negativen Ergebnissen liihrten die Untersuehungen bei den Wiederkauern. Als amylolytisch unwirksam erwies sich so der Speichel des Rindes (MAn.OFF, STEIN~ETZE~, LOCKING), des Scha[es (ScHEUNERT und TttAUTlgANN, LDCKING, RASP), der Ziege (LOCKING, RASP, TRAUTlVIANN und ALBttECtIT) und naeh der einzigen Untersuchung yon GROTZ~ER aaeh der Speiehel des Rehes.
Aus dem Fehlen der amylolytisehen Wirkung des Speichels der herbi- voren Haastiere kann nieht ohne weiteres auf Bin wirklieheS Fehlen tier Dia- stase gesehlossen werden. STEINM:ETZER stellte zwar lest, dass es nicht an einem Mangel der Aktivatoren (Co-Ferment, anorganiseher und organiseher Siiuren) des Fermentes liegt, sondern dass das Proferment (organische Komponente), der spezifisehe Faktor, fehlt. Es ist jedoeh mSg]ich, dass die im alkalischen Gebiet ]iegende aktuelle Reaktion des Speiehels eine Wirkung der Diastase unmSglich maehe, zumal das p~-Optimum ffir Amylase zwisehen neutralem and saurem Gebiet liegt. Allerdings is~ hier nach Vo~K and WOLVE~A~IP ZU betonen, dass die Verdauungsenzyme in natfirliehen Medien nich~ immer unter dem p~-Optimnm wirken. Under Bertieksiehtigung besonders der aktu- ellen Reak~ion haben in neuerer Zeit CHnASZCZ und SC~LEeETnbWNA das Vorhandensein einer Amylase im Speiehel des Pferdes and des Rindes naeh- weisen kSnnen. Sie fanden die optimale Wassersto//ionenkonzentration ffir die diastar Wirkung des Rinderspeiehels pE-6,5 bis 6,6 und des Pferdespeichels bei p~-6,2. Jedoeh wurde aueh bier nie eine vollstiindig verzuekernde Kraft, sondern nut eine Dextrinierung beobaehtet. Es diiri~e sich auch beim Pierd und Rind nut am die bis zum Dextrin abbauende ,,Polyamylase" handeln, analog den obigen Befunden yon GAr.nH~ im I-tandespeichel, wi~hrend die Dextrinase fehlt. Daraus geht immerhin hervor, dass das vorhandene dia- statisehe Teflferment infolge ungfinstiger Wirkung der alkalisehen Reaktion des Speiehels bei beiden Tierarten (s. oben) normalerweise nieht wirken kann. Es kommt damit entspreehend den fiberwiegenden nega~iven Befunden dem Speiehel der herbivoren tIaustiere praktiseh keine amylolytische Wirkung zu.
Von anderen Fermenten ist noeh eine Lipase im Speiehel yon t tund, Pferd, Rind und Ziege naehgewiesen worden (GEJNOVSKu PELUFFO).
IV. Das A b s c h l u c k e n der Nahrung
erfolgt bei allen Tieren ziemlich in der gleichen Weise. Es seien bier nur die Untersuehungen yon G~A~E fiber die Schluckstrasse beim Menschen mit

Allgemeines zur Nagen-Darmverdauung. 589
vergleiehenden Hinweisen auf Tiere erw~hnt. Uber die Beteiligung der Speise- r6hre am Sehluekakt ist auf die Darstellung yon MAsraOnD ZU verweisen. Auf
das Verhalten der SpeiserOhre bet der t~ejektion der Wiederkaumasse wird spi~• eingegangen.
B. Allgemeines zur Magen-Darmverdauung.
Bet allen Tieren ist bet der Gesamtverdauung zwisehen fermentativer, dm'ch die Magen-Darms~fte allein bedingter Verdauung und der bakteriellen Zersetzung zu nnterscheiden. J e pflanzenreicher die Nahrung ist, am so grSssere Rolle spielt die bakterielie Zersetzung, und um so grSsser sind die entspreehenden Absehnitte des Verdauungstraetus bet den einzelnen Tieren
ausgebildet. Bet den ausgesprochenen Pflanzenfressern kann man diese Ab- sehnitte sehon als , ,G~rkammern" bezeiehnen.
Die drei Ern~hrungstypen, Fleisch-, Pflanzen- und Allesfresser, zeigen einen dementspreehend versehiedenen Ausbau ihres Verdauungskanals. Das Ver- h~ltnis yon KSrperliinge zur Gesamtdarmliinge betri~gt beim Hund 1 : 6 (COLIN), bet der Katze 1 : 4 (COLIN), beim Schwein 1 : 14, beim P/erd 1 : 12, beim Rind 1 :20 und bet Scha/ und Ziege 1 :27 ; naeh der genaueren Messung Rumpf- li~nge: Gesamtdarmli~nge land ttAESLEI~ beim Sehwein das VerhNtnis 1 :25 bis 9,7,1. Auisehlussreieh ist die Beriieksiehtigung der inneren Oberfli~ehe des Magen-Darmkanals in Beziehung zur KOrperoberfli~ehe. Bet den Untersuehungen yon 280 Si~ngetieren verschiedener Arten land MAGNAN, dass die Fleisehfresser die kleinste und die Pflanzenfresser die grSsste innere Magen-Darmoberflgehe, bezogen auf die KSrperoberfli~ehe, haben. Naeh COLIN betri~gt das Verhi~ltnis der Hautoberfli~ehe zur Magen-Darmoberfli~ehe
beim t)ferd . . . . . . . . : 2,18 qm, beim Rind . . . . . . . 1 : 2,97 qm, beim Schwein . . . . . . 1 : 1,33 qm (die I-Iautoberfl/~che nach TA•GL), beim I-Iund . . . . . . . 1 : 0,59 qm, bet der I~2atze . . . . . 1 : 0,58 qm.
Der relative Anteil aueh derjenigen Absehnitte, in denen die fermentative Verdauung erfolgt, also des Drtisenmagens und des Diinndarmes, ist bet den tterbivoren etwas grSsser als bet den Carnivoren. Naeh den einzigen Zahlen yon COLIN bereehnet, verh~tlt sieh die Hautoberfli~ehe zur inneren Magen- Dtinndarmoberflgehe
wie I : 0,87 beim Pferde, wie 1 : 0,50 beim ttunde, ,, 1 : 1,17 beim l~inde, , 1 : 0,44 bet der Katze.
Ganz betri~ehtlieh ist der Untersehied zwisehen den Carnivoren nnd tterbivoren in der GrSsse der inneren Oberfl~tehe derjenigen Absehnitte des Verdauungskanals, in denen die bakteriellen Zersetzungen stattfinden. Beim Rinde sind zu diesen sog. , ,Ggrkammern" die Vorm~gen, der Blinddarm und der Diekdarm zu reehnen, beim Pferde der grosse Blinddarm und die beiden

590 W. LEI~KEIT: Neuere Ergebnisse der vergl. Physiologie der Verduuung der S~ugetiere.
gewaltigen Dickdarmlagen. Das so nach don Zahlen yon CoLin berechnete
Verhiiltnis der Hautoberfl~che zur ,,G~rkammer"-0berfl~iche betr~gt beim l%inde . . . . . . 1 : 1,80, beim t~ferde . . . . . . 1 : 1,30,
die entsprechenden Werte ffir den Blind- und Dickdarm betragen beim Hund . . . . . . 1 : 0,08, bet der Katze . . . . . 1 : 0,1.
Aus den gesamten Zahlen ist auch wetter zu ersehen, dass bet den Carni-
voren der Magen und Dtinndarm die gr5ssten Abschnitte des Verdauungs-
kanals sind, w~hrend bet Rind und Pferd die Giirkammern im Vordergrund
sr Anf besondere GrSssenverh~ltnisse einzelner Abschnitte des Verdau-
ungstractus und deren Beeinflussung dutch verschiedene Ern~hrung (I~AESLER~ MANGOLD-HAEsLER) wird sp~iter eingegangen.
C. Die Magenverdammg. I. Magengriisse.
Regenerationsf~higkeit der Vormiigen der Wiederk~iuer. Itinsichtlich der vergleichenden Anatomie des Magens seien bier die
neueren Arbeiten yon PERNKOPF, SCI-IXt)PI und die zusammenfassende Dar-
stellung yon PnE~'K erwiihnt, t t ier soll nur auf einige GrOssenverhgltnisse des
Magens, z. B. zu Darm und KOrper eingegangen werden.
Man kOnnte a priori annehmen, dass die Fermentabschnit te des Verdau-
ungskanals bei den Herbivoren, bei denen die grosse Ausbildung yon G~r-
kammern auf die Bedeutung der bakteriellen Zersetzung der Nahrung hinweist,
in ihrer relativen GrSsse entweder gleich gross oder eher etwas kleiner ws
als bei den Fleischfressern. Rechnet man die yon ConI~ ffir die Magenober- /15che angegebenen Werte auf I qm Hautoberflgche urn, so kommt man nicht
zu einem einheitlichen Ergebnis. Das Verhiiltnis yon Hautoberfl~che zur
Magenoberfl~che betriigt darnach
beim ttund . . . . . . . 1 : 0,13 qm, bei der Katze . . . . . 1 : 0,11 qm, beim Rind . . . . . . . 1 : 0,20 qm (Labmagen), beim l~ferde . . . . . . 1 : 0,07 qm.
Es ist anzunehmen, dass beim Rinde infolge der starken Faltung der
Schleimhau~ des Labmagens das Verhiiltnis etwas enger ist, beim Pferde
dagegen bet Ausschaltung des cutanen Vormagens etwas wetter. Die relative
0berfliiche der Vorm~igen des Rindes ist nattirlich betr~ich~lich, da sie ja auch
den grSss~en Tell der Gi~rkammern ausmachen. In quantita~iver Beziehung
zum Darm ist der Magen der Fleischfresser relativ am grSssten, der Magen (nur Fermentmagen) der Pflanzenfresser am kleinsten. Die Magenoberfl~che
verhMt sich zur Darmoberfliiche beim Pferd wie 1 :28 ,8 qm, beim Schwein wie 1 : 13,2 qm, beim t tund wie 1 : 1,36 qm. Das gleiche kommt natiirlich

Die Magensaftsekretion. 591
auch im Volumenverhaltnis zum Ausdruck. Das Verhiiltnis veto Magen- zum Darmvolumen betrggt beim IIunde 1:0,5, beim Menschen 1 :2 , beim Kaninchen 1 : 8 (NEu~AYER), beim Pferd 1:10,7, beim Sehwein 1:2,4, bet der Katze 1 : 1,4 (COLIN). Die Ursaehe dieser Unterschiede liegt in der starken Entwicklung besonders des Dickdarmes. Der Magen dient bet den reinen Pflanzenfressern in welt geringerem Masse zur Aufspeicherung der Nahrung als beim Fleischfresser; weil die tIauptzersetzung der Pflanzennahrung gerade im Diekdarm stattfindet; natfirlich gilt dieses mit Ausnahme des Rindes.
Zu erwiihnen ist noeh die interessante Eigenttimliehkeit der Vormagen, sieh naeh operativer Entfernung zu regenerieren [TRAvT~A~N und SC~MITT (1)]. Naeh Exstirpation yon Pansen und Haube bet Ziegenl~immern im Alter yon 2--4 Woehen fanden TaAUTMANN und SC~ITT (1) nach 4,6 und 8 Monaten Pansen und t taube neu gebildet, die im Laufe der Zeit makroskopiseh und mikroskopiseh immer mehr den normalen Organen ~hnelten. Je friiher naeh der Operation Rauhfutter gereicht wurde, um so vollkommener war die Aus- bildung besonders des Pansens. Aueh naeh der Entfernung des Psalters bildete sieh dieser zum Tell zurtick; nur die neugebildeten Bl~itter sind nieht so yell- kommen ausgebildet wie die normalen [T~AvTrIA~N (5)].
II. Die Magensaftsekretion.
1. Die Magendriisen. Zum leiehteren Verstiindnis des Ablaufes der Magen- verdauung sell zun~chst noch kurz aul einige morphologische Besonderheiten der Magensehleimhaut bet versehiedenen Tieren eingegangen werden. Folgende Schleimhautregionen kSnnen im Magen vorhanden seth: 1. die mit cutaner Schleimhaut ausgekleidete Region; 2. die Cardiadrfisenregion; 3. die Fundus- drfisenregion und 4. die Pylorusdrtisenregion. Die Ausdehnung der einzelnen Regionen, besonders der beiden ersten, ist bet den einzelnen Tierarten ver- schieden. Hier sollen nur die Haustiere und einige prinzipielle F~lle der iibrigen Siiugetiere berficksichtigt werden. Eine ausfiihrliche Darstellung des Aufbaues der Magenschleimhaut in der ganzen S~iugetierreihe haben bereits 0PPEL und BIEDERMANN gegeben.
Die cutane Schleimhaut kann aueh ganz allein den Magen auskleiden. Bet den Monotremen fehlen die Driisen im Magen vollkommen; der ganze Magen tr~gt nur eutane Schleimhant wie der Oesophagus (0PPEL). Bet den Einhu/ern und ltatte und Maus besitzt der eine Tell (1/3 bis 1/2 ) des Magens (der Vormagen) cutane Sehleimhaut, der andere Tell (der Driisenmagen) dagegen die drei Driisenregionen. Beim Schwein tritt der cutane oesophageale Abschnitt gegentiber den driisigen Absehnitten betr~ichtlich in den Hinter- grund (ELLENBEXGER). Bet stiirkerer Entwieklung des cutanen oesophagealen Magenabschnittes tritt dieser als selbst~indige HShle hervor, so beim Hamster [ S c ~ u N ~ (3)J und bet der Wasserratte (0r~EL). Bet den Wiederkiiuern hat der cutane Abschnitt mit der Ausbildung der komplizierten Vormiigen,

592 W. LI~NKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~uge~iere.
Pansen, Netzmagen, B]~ittermagen seine h6chste Differenzierung erreicht,
w~ihrend die Driisenregionen der Labmagen %r~igt. Die Cardiadrfisenregion feh]t vollkommen z. B. im Magen des IIasen, des Meersehweinehen, im Lab-
magen der Wiederk~iuer und im Magen der fleisehfressenden Cetaeeen. Beim
Mensehen, IIund und Kaninchen z. B. sind die Cardiadrfisen nur auf eine sehmale Zone an der Miindungsstelle des Oesophagus beschr~nkt. Beim
P/erd sind die Cardiadrfisen auf einem relativ schmalen Streifen zwischen Vor-
magen und Fundusdriisenzone vorhanden. Beim Hamster, bet Ratte und ~laus [OPPEL, LENKEIT (8)] ist die Cardiadrfisenzone schon wesentlich st~irker aus-
gebildet. Beim Schwein nehmen die Cardiadrfisen sogar ein reichliches Drittel der ganzen Magenschleimhaut ein. Nach OeeEn SO1] auch bei den ~aultieren die Cardiadrfisenregion eine enorme Ausdehnung zeigen.
Nach den neueren nmfangreichen Untersuchungen yon TIaAUT~ANN (I)
an Schweinen ~hneln die Cardiadrfisen nach ihrem strukturellen und tink- %oriellem Verhalten in hohem Masse den ser6sen Driisen.
Bet tIungertieren enthiilt der lumenhaltige Tell der Cardiadrtisenzellen zahlreiche, dicht beieinanderliegende, stark liehtbrechende Granula, die sogar fret im Lumen liegen kSnnen. W~ihrend der Verdauung nehmen die Granula betr~ichtlich ab. Manche Zellen sind vollkommen fret yon Granula. TRAUT- MAN~ (1) nimmt wetter an, dass die Granula ein Material mit fermentativen Eigenschaften liefern. Charakteristiseh ist auch ftir die Cardiadrtisenzone des Schweines der l~eichtum an cytoblastisehem Gewebe in Form grSsserer und kleinerer LymphknStchen. Diesem lymphatischen Apparat weist TI%AUTlVIANN eine bedeutende Rolle bet der Abt6tung und Wachstumshemmung won Mikro- organismen zu, die gerade das Schwein infolge seiner Wtihlt~itigkeit in reich- licher Menge aufnimmt.
Ftir die Fundusdri~senregion ist erwiihnenswert, dass bei den Chiropteren im Winter die Belegzellen fehlen (K~uMBIEGE~). Zwisehen tier Fundus- und der Pylorusdrfisenregion liegt die Intermediiirzone, die bet den Haustieren (ELLENBEgQn~), bet Mensch, Hund (Asc~OFF) und bet t~atte und Kanin- chen (BILLENICAMP) naehgewiesen ist. Sie ist fret von tIauptzellen und be- steht vorwiegend aus Zellen mit bestimmtem tinktoriellem VerhMten, den ,,Nebenzellen" (ELLENBE~GEa, K. W. ZIM~E~ANN), S. ,,Zwischendrtisenzellen" (Asc~OFF), S. ,,mucoide" Zellen (LI~). Einzelheiten sind in den Darstellungen yon ZIM~Ea~'~AN~, B~BK~ ZU linden. Das Ober/liichenepithel des Drtisen- magens besteht aus einer besonderen Art won Schteimzellen.
Ausserdem set noch auf die chromaffinen (KusL, TEI~Vna) und argent- affinen Zellen (Eni)s) hingewiesen. Diesen Zellen wird neuerdings eine inner- sekretorische Bedeutung zugewiesen (Kv~n, E~Os) ;sie sollen bet der Verdauung ~iberhaupt keine Rolle spielen (E~Ss).
Aus allem geht hervor, dass der drtisige Teil des Magens besonders vom physiologischen Gesiehtspunkt aus ein kompliziertes Gebilde ist. Die

Die Magens~ftsekretion. 593
physiologisehe Bedeutung der Cardiadrfisenregion und der Intermedi~rzone ist noch reeht unklar, nieht weniger die lunktionelle Abh~ngigkeit der einzelnen Zonen voneinander. Gerade bei unseren Haustieren ist ifir die histophysio- logische Forschung noch ein weites Feld.
2. Der Magensaft. Am eingehendsten untersueht ist der Magensaft nur des Menschen und des Hundes. Auf die diese Magensiifie betreffenden auf- sehlussreichen Arbeiten (DIENS~: U. a.) ausffihrlieh einzugehen, ist hier nicht der Raum, zumal darfiber auch vor nieht allzulanger Zeit an anderen Stellen erschSpfend beriehtet worden ist (B~BKI~, I~OSEI~IANN). Bei weitem nieht so eingehend, man kann sagen, noch vollkommen unvollstiindig sind unsere Kenntnisse fiber den Magensaft unserer herbivoren und omnivoren Haustiere; bei den wildlebenden Tieren lehlen uns diese naturgemi~ss ganz. Die Ursachen liegen in erster Linie in den Schwierigkeiten der Gewinnung des Magensaftes bei unseren Haustieren.
Untersuehungen des Mageninhaltes sind durch seine sofortige Entnahme nach der T6tung der Tiere leicht m5glieh. Durch Ausheberung ist der Magen- inhalt evtl. nut beim Sehwein zu gewinnen, beim Rinde kann der Labmagen nieht erreicht werden infolge der Vorlagerung der Vorm~gen, beim Pferde nicht iniolge der festen Schliessung der Cardia. Der zur Gewinnung des wirklieh reinen Magensa[tes erforderliehe kleine Magen nach tt~n)EN~AI~, PAWLOW kann yon den ttaustieren nur beim Pferde nicht angelegt werden infolge der Entfernung des Magens yon den Bauchdeeken.
Der durch einen kle~nen Magen gewonnene reine Magensa[t, der wohl zu unterscheiden ist yore Mageninhalt , ist bei unseren Haustieren ausser beim Hund nur bei Schaf, Ziege und Kalb bisher untersueht worden.
l~ber die Zusammensetzung des Magensaftes sind nur ~ltere Angaben yon Kalb (BELGOWSKI) und Ziege (GROSSEIt) neben den zahlreichen Analysen yon I-Iund und Mensch vorhanden. In tier Tabelle 3 sind die angegebenen ana- lytisehen Werte angeffihrt und zum Vergleieh den Werten yon Hund nnd Mensehen gegenfibergestellt.
Weitere genauere analytisehe Daten des reinen Magensaftes der an- geffihrten Haustiere liegen nicht vor. Auffallend ist in der Zusammensgellung der Tabelle 3 bei der Ziege der hohe Wert ftir den Trockenrtiekstand und ffir die Asche; sonst sind zwisehen den einzelnen Gruppen keine Unterschiede vorhanden.
In der Acidit~t des Magensaftes sind dggegen zwischen den Wiederkguern und den Fleisehfressern deutliche Unterschiede festzustellen. Die Aciditgt des Magensaftes ist im Durchsehnitt bei den Fleisehlressern grSsser als bei den PIlanzenfressern. GesamtaciditSt betr~gt im Magensaft
der Z i e g e . . . . . . . . . 0,0438% HC1 (G~ossxR), des Schafes im I)urchschnitt 0,262--0,3% w~hrend der Fut teraufnahme
0,198~0,35% vor . . . . IN. A. PoPow (2)].
Asher-Spiro, Ergebnisse der Physiologffe und exper. Pharm~kologie. 35. 38

594 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologic der Verdauung der S/~ugetiere.
Tabelle 3. Z u s a m m e n s e t z u n g d e s M a g e n s a f t e s .
Tierart
Ziege . . .
Kalb . . .
Schaf . . .
Hund . . .
Mensch . .
Spez. Gewicht
1,006
1,0018 bis 1,0031
1,0058 bis 1,0084
1,002 bis 1,006
1,006 1,009
0,660
0,56 bis 0,640
10,470 bis 0,650
Troeken- substanz
%
1,142
0,353 bis 0,506
0,428 (0,360 bis
0,659)
0,556 (0,48 bis
0,606)
Orga- Asche nische N
Substanz
% % %
0,84
(,142 bis 0,238
0,133 (,),085 bi~
0,204) 0,126
(0,11 bis 0,14)
0,302 0,0526
0,294 0,036 (0,176 bis i 0,054
0,489)
0,430 0,060 (0,34 bis (0,051 bis
o,47) i o,o74)
Autor
GROSS~R
:BELGOWSKI
N. A. PoPow
ROSE~s
CARLS01% I-~AGER U. ROG]~R, zit. nach X~OSEMANlg
Beim t tunde ist dagegen im reinen Magensaft die gesamte Salzsgure als freie Sgure vorhanden (RosE,rAiN). Der Gohalt an/refer Salzs~ure betri~gt
beim Katbe . . . 0,1294--0,3560% HC1 (BELGOWSX_I), bei tier Ziege . . 0,0175% HC1 (G~ossE~), beim Schaf . . . 0,023--0,026% HC1 vor der Fut teraufnahme (N. A. PoPow).
0,029---0,04% ~C1 w~hrend ,, ,, beim I tund . . . 0,450~0,58% I-IC1 (RosEMA~r
Die Wasserstoffionenkonzentration des reinen Magensaftes ist bisher bei den herbivoren ttaustieren nur beim Schaf untersueht worden. N. A. PoPow (2) und Mitarbeiter fanden beim Schaf in dem aus einem HEII)ENtrAIN-Magen gewonnenen Magensait die aktuelle Reaktion yon p~-8,91--1,38 (--1,16); fiir den Hundemagensaft, durch Scheinffitterung gewonnen, gibt ROSE~rAN~ die Reaktion yon p~ 0,98--0,80 an.
Auch die Verdauungskraft des reinen Magensaftes scheint bei den Fleisch- fressern grSsser zu sein als bei den Pflanzenfressern. Leider sind die meisten Bestimmungen se!bst in neuerer Zeit (PoPow) nach der Methode yon METr durchgeffihrt worden trotz des Vorhandenseins besserer Methoden (GLXSSNER,
GR~-S8, VONK). BELGOWSKI land ffir den Magensaft des Kalbes nach METT nach 10--I2 Stunden eine Verdauungskraft yon 1,0--2,75 ram, and neuer- dings stellen N. A. Po~ow (2) und KVD~JAWZEW beim Schaf ftir den aus dem isolierten kleinen Magen gewonnenen Sal t mit der gleichen Methode eine ver- dauende Wirkung yon 1,2--2,4 ram nach 12 Stunden bei 380 fest. Zum Ver- gleich seien die fiir den Hund yon CmsmN (s. ROSE,ANN, BAnKIN) gefundenen Werte angeffihrt. CHISmN land eine verdauende Kraft des aus dem PAWLOW- Magen sezernierten Saftes nach METT bei Fleischffitterung im Durchsehnitt 3,65 ram, bei Brotftitterung 6,64 mm, nach Milchaufnahme 3,25 mm nach 10 Stunden bei 38 ~

D i e M a g e n s a f t s e k r e g i o n . 595
Die Pepsine der versehiedenen Tierarten seheinen naeh den Unter- suehungen yon gONE identiseh zu sein. Die p~-0ptima weiehen nur wenig voneinander ab; sic liegen z. B. beim Sehweinepepsin in der Sehwankungs- breite yon Pa 1,78--1,8. Das Vorkommen einer Lipase im reinen Magensaft wird aueh in neuerer Zeit sehr bestritten [RASENKOW (8), MELnI und RADICI). Eine praktiseh bedeutende lipolytisehe Wirkung kommt wohl dem Magensaft keines Tieres zu. Dagegen konnten in der Magensehleimhaut HAU~OWlTZ and P~T~ON erneut dieses Ferment naehweisen, und zwar in besonders grossen Mengen beim Mensehen, bei den Raubtieren und Nagetieren; in der Labmagen- sehleimhaut der Wiederki~ner ist es entweder fiberhaupt nieht zu linden oder nur in geringen Mengen vorhanden. Interessant ist, dass die Lage des p~-Opti- mums der Magenlipase ftir jede Tierspezies ziemlieh konstant ist. Ffir die Wirkung der rohen Magenlipase fanden WIr~LSrXT~ER, HAUROWITZ und ME~- MEN, ttAU~OWI~Z und P ~ o ~ folgendes p~-Optimum:
H u n d . . . . . . . . . 5 , 5 - - 6 , 3
K a t z e . . . . . . . . . 5,5
F u e h s . . . . . . . . . 5 , 5 - - 6 , 3
W i e s e ] . . . . . . . . . 5 , 5 - - 7 , 1
H a s e . . . . . . . . . 5 ,5
K a n i n e h e n . . . . . . . 6,3
~ a m s t e r . . . . . . . . 7,1
~ e e r s o h w e i n c h e n . . . . 7 , 1 - - 7 , 9
W e i s s e l ~ a t t e . . . . . . 7 , 1 - - 7 , 9
E i c h h 6 r n o h e n . . . . . 7 , 1 - - 7 , 9
S o h w e i n . . . . . . . . 7 , 1 - - 7 , 9
t~ferd . . . . . . . . . 7 , 1 - - 8 , 6
Uber den Sehleimgehalt des Magensaftes und fiber dessen Beziehung zu den anderen Konstanten sind wit bei unseren ttaustieren mit Ausnahme des I-tundes sehleeht unterriehtet, so dass man den zahlreiehen Untersuehungen am Magensaft des Mensehen und des tIundes (BaB~I~) keine vergleiehenden Daten gegenttberstellen kann. Eine genauere Kenntnis der Sehleimsekretion bei unseren ttaustieren wfirde wohl aueh zur Kl~rung der Aeidit~ttsunter- sehiede zwisehen Pflanzenfresser- and Fleisehfressermagensaft beitragen. Es is~ die Frage, ob die oft geringere Aeidit~tt bei den Pflanzenfressern auf einer festgelegten sekretorisehen Eigensehaft der Drtisenzellen beruht, oder eino An- passung an die vegetabilisehe Nahrung darstellt. Sehon die alten Versuehe yon Cmsm~ am tIunde ergaben n~mlieh naeh kurzdauernder Brotffitterung eine geringere Aeidit~t (0,47 % HC1) als naeh Fleisehffitterung (0,56% HC1). Dureh lang dauernde einseitige Ern~hrung konnte KOSCETOXANZ (1) in gleiehem Sinne eine Anpassungsreaktion der Magendrtisen an des entspreehende Nahrungs- regime feststellen. Die Resulta~ce yon NEVE~ spreehen allerdings gegen die Allgemeingfiltigkeit dieser Befunde. Aber manehe Tatsaehen lassen doeh noeh daran denken. Die bisweilen zu beobaehiende erhShte Ausnutzung der Roh- faser bei Zuffitterung yon tierisehem Eiweiss (FE~A~I) ist wahrseheinlieh aueh zum Teil auf die erhShte Aeidit~tt des Magensaftes zurfiekzuffihren, und zwar infolge einer besseren Aufquellung der Rohfaser. Endgtiltige Beweiso ffir diese Auffassung liegen noeh nieht vor.
Die verschiedenen Regionen des Dri~senmagens liefern, wie eingehend nur beim t Iund untersueht worden ist (BAB~IN, G O L D B E I ~ G , L E V I N ) , bei allen
38*

596 W. Lv.~K~IT: Neuere Ergeb~isse der verg]. Physiologie der Verdauung der S~uget, iere.
Tieren ein ganz verschiedenes Sekret. W~ihrend das Fundusdrtisensekret stets ausgesprochen saner ist (LEvIN), ist das Pylorusdriisensekret neutral oder gar alkalisch und mehr oder weniger stark sehleimhaltig (E~LENB~I~GEa und HOFMmST~, ROSEXFELD). Auf die Untersehiede des-Fundusdrtisensekretes zwischen den Pflanzen- und den Fleischfressern ist im vorhergehenden Ab- schnitt fiber den Magensait eingegangen worden. Der aus dem kleinen Magen gewonnene reine Magensaft ist fast durchweg nur retries Fundusdriisensekret. Im Sekret der Pylorusdritsen dtirfte yon besonderem Interesse der Schleim- und Pepsingehalt seth; leider stehen hier zum Vergleieh keine geniigenden Resultate zur Verftigung. Das Cardiadritsensekret ist neutraler oder alkalischer Reaktion und enthiilt kein Pepsin. Nach ~ilteren Angaben sollte es auch ein diastatisehes Ferment enthalten. EDEL~ANN land niimlieh Extrakte der Cardiadrtisenschleimhaut yon Hamster, Ratte, Pferd und Schwein diastatiseh wirksam. Jedoch kann aus dem Fermentgehalt des Sehleimhautextraktes nieht anf Bildnng des Fermentes in tier gleichen Sehleimhautregion gesehlossen werden. Aueh aus den anderen Schleimhautregionen des Magens z. B. des Sehweines konnten fermenthaltige Extrakte gewonnen werden (ELL~NBE~G~g). Bet diesen positiven Diastasebe/unden in der Magenschleimhaut handelt es sich nm adsorbierte Speicheldiastase (ELLENSERGE~). Dnrch neuere Unter- suchnngen mit Hilfe eines in der Cardiadrfisenregion angelegten kleinen Magens konnte tats~ehlieh in dem Sekret keine Diastase naehgewiesen werden [Sc~NE~T, T~A~T~AXN (1)].
3. Ablauf der Magensaftsekretion. Die Sekretion des Magensaftes ist bei den Pflanzenfressern, bei denen tier Magen nie leer wird, eine kontinuier- liche [PoPow (2)], im Gegensatz zu der vorrtibergehenden beim Mensehen un4 bei den Fleischfressern. Bei den letzteren sezernieren nur die Pylorusdrtisen und das sehleimproduzierende Oberfliichenepithel dauernd, und liefern das sog. Nfichternsekret, das sehwach alkaliseh ist nnd wenig Pepsin enthii!t. Die erste Phase der Sekretion, die reflektorische, tritt naeh N. A. PoPow (2) und Mitarbeitern bei den Wiederkiiuern bei weitem nicht so ausgepriigt in Erschei- nung wie bei den Fleischfressern und beim Menschen, und wird auch bei den fibrigen Pllanzenfressern und beim Sehwein, den Tieren mit dauernd geiiilltem Magen, kaum mehr hervortreten. Vollkommen zu fehlen seheint diese Phase jedoch auch hier nieht nach den Seheinftitterungsversnehen yon I~RATINOW und SI~uLow am I(alb nnd yon KRATINOWA am Sehwein. Die bei Hund und Menschen naehgewiesene zweite, die chemisehe oder naeh BABKIN die Pylorus- phase, ist bei den Pflanzeniressern wohl die wiehtigste. Welehe Bedeutung bei den iibrigen Tieren ausser Hund der dritten Sekretionsphase, der Darmphase (BABI~I~), zukommt, ist heute noeh nieht klar. Man muss sieh hiiten, die Ver- hgltnisse vom Fleisehiresser z. B. auf die Pflanzenfresser zn tibertragen; leider gesehieht dieses noeh zu oit.
Uber den Mechanismus der Wirkung bestimmter Stoiie yon der Pylorus-

I)ie Magensaftsekretion. 597
sehleimhaut aus sind viele Annahmen gemaeht worden (BA~IN, EDKINS).
Erw~ihnt seien hier nur die beweisenden Versuche yon RASE~KOW (1) ftir das Vorhandensein soleher die Fundusdrfisensekretion anregenden Stoffe im Blur auf der H6he der Magenverdauung. RASE~KOW konnte niimlieh mit dem Blut eines im Maximum der Magensekretion befindliehen Hundes bei einem anderen I-Iunde in nfiehternem Zustand dureh intraven6se Injektion gleiehfalls eine Magensekretion ausl6sen.
Es ist nun die Frage, ob bei den Fleisehfressern und bei den fibrigen Tieren dieselben Stoffe yon der Pylorussehleimhaut aus sekretionsanregend wirken. Beim Hunde kommen in erster Linie das Fleiseh und die Extraktions- stoffe des Fleisehes (Liebigs Fleisehextrakt) in Betraeht. Eine hervorragende Bedeutung als Reizstoffe haben hier naeh den Arbeiten yon TIMOFEJEFF zu- n~ehst die Eiweissabbauprodukte. Chemiseh reine Aminos~uren wirken jedoeh naeh den Untersuehungen yon BICKEL nur naeh intragastraler Zufuhr anregend auf die Magensekretion, yon den Aminen )2thyl-Methylamin-tIC1 und vor allen Dingen alas tIistamin sehon naeh subeutaner Injektion. Aueh die besonders naeh Aufnahme pflanzenreieher Nahrang auftretenden organisehen Siiuren wie Essigsiiure, Buttersiiure und Milchsiiure kSnnen eine sekretions- anregende Wirkung haben. Bei den Pflanzenfressern entstehen aber diese S~uren in betr~ichtliehem Masse bei den G~trungsprozessen in den Vormiigen der mehrh6hligen Miigen (Wiederk~uer, Hamster) und im Vormagen der ein- h6hligen M~gen (Pferd, Ratte, Maas), so dass sie hier als sekretionsanregende Stoffe ganz besonders in Frage k~men. N.A. Po•ow (2), N~STE~OW und UGA~ow konnten tats~ehlieh bei Sehafen mit Labmagenfistel and kleinen Magen dutch intragastrale Verabiolgung yon 2 % Essigsiiure, yon Essigsiiure und Milchsiiure und yon saarem Silosaft eine deutliehe Steigerang der Lab- magensekretion mit Zunahme der Aeidit~tt und des Pepsins hervorrufen. Naeh Zufuhr yon konzentriertem Futter wie Ha/er nahm die Sekretion unter Er- h6hang der Aeidit~t and der Fermentkonzentration bedeutend mehr zu als naeh Heuffitterung. Alkohol und Na2CO a regen die Labmagensekretion beson- ders intensiv an. Dagegen haben Pepton and Fleischbri~he beim Sehaf im Gegensatz zum Hund nur einen geringen steigernden Einfluss auf die Magen- saftsekretion. Hemmend wirkt wie beim Hund 0,5 % tIC1, desgleiehen NaC1 und MgC% and Fett (N. A. Porow [2] und NEST~OW).
Mit Hilfe der vorliegenden Tatsaehen lasst sieh aueh der yon GRossna am Labmagen einer Ziege mit tIilfe eines PAwLow-Magens beobaehtete Sekretionsverlauf erkl~ren. Bei einem hungernden Tier trat naeh der Futter- aufnahme eine Steigerung der Safte- and S~ureproduktion ein, die sieh im Laufe der folgenden 3 Stunden noeh betr~ehtlieh erh6hte und yon der 6. Stunde erst abnahm; mit der 11. Stunde versehwand die freie tIC1. Die Zunahme der Sekretion im Laufe der 3. Stande naeh der Ffitterung ist gerade als eine Wir- kung der 2., der ehemisehen Sekretionsphase anzusehen.

598 W. LE~J~EIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~ugetiere.
Beim Kalbe sah BELGOWSKI bei tiblicher Fiitterung die Sekretiou ziem- lich in gleieher Weise verlaufen; die Art des aufgenommenen Futters war yon geringem Einflnss; auch nach l~ngerem Hungern soll noeh freie HC1 zu linden sein.
Die kontinuierliche Sekretion bei den Pflanzenfressern ist nur auf die dauernde Fiillung des Magens zurtiekzuftihren, und wird infolgedessen allein auf dem Wege der 2. Sekretionsphase dureh Erregung der Magendrtisen yon der Pylorusschleimhaut aus bedingt. Dass aber auch eine kontinu@rliche Spontansekret~on aller Drtisenelemente, d .h . auch bei vollkommen leerem Magen, stattfindet, ~st aueh bei den/Pflanzenfressern nicht sehr wahrseheinlich und daftir ist auch bis ~jetzt nicht der experimentelle Beweis geliefert worden. Auch hier tritt wohl wie beim t tunde (B/~nKIN) eine Sekretion nur dann ein, wenn entsprechende Reize besonders der 2. Phase die Magendrfisen treffen. Der Labmagen wtirde sich danaeh nieht anders verhalten wie der Hundemagen bei fast vollsti~ndiger Ausschaltung der ersten Sekretionsphase.
Die yon ELLENBE~GEa, SC~rEUNERT fiir die Cardiadri~sen des Schweines angenommene kontinuierliehe Spontansekretion, auf Grund yon Unter- suchungen am kleinen Magen, ist wieder in Zweifel gezogen worden dureh die Untersuchung yon T~AUTSIA~N (1). TRAUTMANN land n~mlich, dass der sich aus dem kleinen Magen der Cardiadrfisenregion entleerende Salt stets mit dickflfissigen eiter~lmlichen Massen zunehmend durehsetzt wird. Die histo- logische Untersuehung ergab als Ursache eine starke Entwieklung des lym- lohoiden Gewebes, so dass das Sekret mit Exsudat durchsetzt war. Aueh die verschiedenen histologisehen Bilder w~hrend der Verdauung und w~hrend des Hungers schliessen eine kontinuierliche Spontansekretion aus. Am kleinen Magen kann im Hunger eine derartige Exsudation eine Sekretion vort~usehen.
HI. Mechanik des einhiihligen Magens. Dass die Nahrung im Magen sieh schichtet, muss besonders nach den
ausftihrlichen Darstellungen yon SCI~I]EUNEI~T als bekannt vorausgesetzt werden. Durch st~rkere Bewegung konnten jedoch ABDE]aIIALDEIV und WE~T~EI~E~ bei Meerschweinchen eine vollkommene Durchmischung des Mageninhaltes herbeiftihren.
Die Bewegung des einhShligen Magens ist bei allen Tieren ziemlich gleich- artig, und zwar am Pylorus betriichtlieh intensiver als am Fundus. Es sind wohl nur graduelle Unterschiede vorhanden, wie die vergleiehenden rOnt- genologisehen Untersuchungen von KRZYWANEK (1, 2) am Hund, Ratte, Meerschweinchen und Hamster zeigen. Beim Pferd sind neben der Konsistenz des Fntters auch wohl am Pylorusabschnitt weniger stark auftretende Bewe- gungen die Ursache dafiir, dass es selbst hier nicht zu einer vollkommenen Vermischung des Mageninhaltes bei normaler Mahlzeitfolge kommt. Dass keine sehr innige Durchmischung der aufeinanderfolgenden Mahlzeiten start-

Mechanik des einh6hligen M~gens. 599
finder, geht auch aus dem eigenartigen Verlauf der Gesamtausscheidung der Futterreste hervor [LENKEIT (2)]. Gcgentiber dem Schaf ist der Beginn der Ausscheidung mit 21--24 Stunden wesent]ieh sparer und die Dauer der Aus- scheidung mit 4--5 Tagen bedeutend kfirzer (s. S. 627). Vergleiehend inter- essant ist noch, dass beim Pferd ein Toil der aufgenommenen Mahlzeit sogleich in don Darm fibertritt; und zwar gesehieht dieses sowohl bei geftilltem als bei nahezu leerem Magen (Sc~EUNE~T und Se~ATT~E). Leider sind nach den fundamentalen Arbeiten ELLENBE~enRS und seiner Schiller in neuerer Zeit keine weiteren Untersuchungen fiber die motorischen Funktioaen des Magens weder beim Pferd noch beim Schwein durchgefiihrt worden. Es sei daher hier auf die Darstellung der Ergebnisse bis 1929 yon SC~EUNERT und KRZY- WANE~ im Handbuch der Erni~hrung und des Stoffweehsels der landwirtschaft- lichen Nutztiere yon E. MANGOr,D verwiesen. Beim Hunde sind dagegen im Hinblick auf den Menschen die neueren Arbeiten auch fiber die Magenbewe- gungen und -entleerungen recht zahlreich; jedoeh ist hier nicht der 0r t auf diese einzugehen.
IV. Mechanik des mehrhiihligen Magens.
1. Bewegungen der Vorm~igen und des Labmagens, einsehliesslieh Sehlund- rinne. Uber die Bewegungen der Wiederkiiuermiigen sind unsere Kenntnisse gerade in den letzten Jahren wesentlich bereichert worden.
Die Haube zeigt eigenartige Kontraktionsformen wie sie sonst an keinem Hohlorgan zu beobachten sind. Die Kontraktionen erfolgen in zwei aufeinander- folgenden Abs~tzen.
WESTER fasste zuni~chst die Bewegungen ,,als eine hin- (peristaltische) und rilcklanfende (antiperistaltische) Welle" auf. Spi~ter milderte er diesen Standpunkt. Wie aber die Untersuehungen yon CZEPA und STIGLER, Yon MANGOLD und KLEIN, yon SClZALK und A~ADON und yon TRAUT~ANN (4) jetzt endgilltig bewiesen haben, handelt es sieh bei den Spontanbewegungen der Haube um ruckartige zweizeitige konzentrische Totalkontraktionen des 0rganes [MANaOLO (1, 2, 4)]. In tier 1. Phase zieht sich die Haube auf die Hiilfte bis zwei Drittel ihrer GrSsse zusammen nnter'deutlicher Rnnzelung der 0berfl~che (MANGOLD und KLEIN). In der 2. sich anschliessenden Phase wird die Hanbe d u t c h eine stgrkere Kontraktion beim Schaf bis auf etwa Pflaumengriisse (MANGGLD nnd KLEIN) bzw. bei der Ziege 1/4 ihrer nrsprfing- lichen GrSsse [TaAvT~ASS (4)] verkleinert; die 0berfl~che ist jetzt besonders stark gernnzelt, und die Blutgefiisse sind stark geschlangelf [MANGOLD (4)].
In dem Ablauf der beiden Kontraktionen scheint es drei Typen zu geben (Abb. 2). Nach der ersten Kontraktion kann wieder eine teilweise Dilatation noch mit einem gewissen Kontraktionsrfickstand eintreten, so dass beide Kontraktionen in Form einer anakroten Kurve verlaufen (MA~CGOnD und KLEIN, Abb. 2a). Die zweite Kontraktion kann auch ohne jede Erschlaffung

600 W. L~KEIT: Neuere Ergebnisse der verg]. Physiologie der Verdauung der S~ugetiere.
der Haube sofort auf der HShe der ersten ansetzen (CzE~A und STIGLER, ~/fAN- ~OnD und KT,EIN, Abb. ~b). 1VIANGOLD und KLEIN konnten beide Arten des Kontraktionsablaufes bei demselben Tiere in unregelm~ssiger Folge beobaehten.
Oesophczg"us
Z. Pansen
Abb. I. Die s Abteil~en des ~iederk~uermagens. Ventrale Ansieht. Der Vorhof is t infolge der ]~[auben- kontrakt ion siehtbar. (Nach ~AIgGOLD and KLEIN.)
Von W~STEa (1, 2), 8CHaLK und AMADON wurde aber aueh eine vollstgndige Ersehlaffung der Haube naeh der ersten Kontrak- tion beobaehtet. Aueh T~A~ZT- MANN (4) sah beim noeh saugen- den Ziegenlamm, mit Hilfe der Bauehfenstermethode, die Haube naeh der 1. Kontraktion bis fast zur anfgngliehen GrSsse ersehlaf- fen. Eine einzeitige Kontraktion der Haube, die niemals zur maxi- malen Zusammenziehung ffihrt, s a h e n MANGOLD und KT,~,I~ an einigen Sehafen naeh zweitagi- gem Hunger, bei anderen Tieren
sogar abweehselnd mit der zweiphasigen Bewegung auftreten. In den letzteren Fallen seheint der einfaehe Kontraktionstyp mit dem Sehluekakt zusammen-
zuh~ngen. Unabh~ngig yon diesen Kontrakiionen treten als weitere Sponian-
bewegungen noeh pendelnde Kontraktionen der Haubenmuskulatur auf (MANGOLD und KLEIN) ; wie weir damit die vonCZEPA und
a. ~. c. bewegung der Haube, die dureh abweehselnde Verkleine- Abb. 2. Graphisehe Dar- steH~g aer sponta~en rung und VergrSsserung des Sehattens in kranio-eaudaler
lClaubenbewegungen" Richiung zum Ausdruek kommt, zusammenh~ngt, ist a, b , c siehe im Text. (~c~ MA~aoL, ~ a noeh nieht reeht klar.
~ ' ) Als dritte Bewegungsart beobaehtete MANGOLD und KLEIN an laparotomierten Sehafen noeh Rotationen der Haube um ihre Liingsachse.
Beim saugenden Ziegenlamm konnte dagegen TRAVTMANN (4) am Netz- magen ausser der zweiphasigen Zusammenziehung keine andere motorisehe T~tigkeit beobaehten. Diese Untersuehungen yon TRAUT~ANN w~hrend der Saugeperiode zeigen, dass die t tauptbewegungen der Haube bereits beim saugenden Wiederkauer, tier noeh nieht wiederkaut, in der gleiehen Weise auftreten wie beim erwaehsenen Tier.
Die Frequenz der Ha~bendoppelkontraktionen nimmt mit dem Alter ab, und zwar dadureh, dass die Pausen zwisehen den Doppelkontraktionen l~nger werden. Die Dauer der zweiphasigen Kontraktionen seheint in allen Lebens-

Meehanik des mehrh6hllgen M~gens. 601
altern und bei allen kleinen Wiederkguern gleich zu sein; wahrend der Sgnge- zeig betrug sie beim Ziegenlamm naeh TRAUT~IAS~ 5 Sekunden und beim erwaehsenen Sehaf nach PoPow (2) 5--6 Sekunden, nnd die folgende Pause dauerte beim noeh sangenden Ziegenlamm 15 Sekunden [T~AuT~AN~ (4)]; nach der S~tugeperiode sehwankte die Pause bereits zwischen 25 und 40 Sekun- den und wurde im noeh spi~teren Lebensalter noeh l~tnger; sie kann sieh dann mitunter sogar mn das 3--4fache verlgngern (T~AuT~AN~'). Ftir erwaehsene Sehafe land N. A. PoPow Pausenwerte yon 80--35 Sekunden. In einer Minute erfolgen danaeh beim s~ugenden Wiederk~uer im Durehschnit~ 3 Doppel- kontraktionen, beim erwaehsenen Tier 1--2. Die gleiche Frequenz yon I his 2 Doppelkontraktionen pro Minute geben aueh WEST~ (1, 2), Buss ffir das erwaehsene Rind, MA~GOLD und KLEIN fiir das erwachsene Sehaf und CZEPA und STmLEa ffir die Ziege an. Man kann mit MANGOLD annehmen, dass die Zahl der Bewegungen yon versehiedenen Faktoren, so yon der Art des Futters und yore Ffitterungszustand nicht unwesenilieh beeinfiusst wird.
Sehr eng h~ngt mit der Bewegung der Haube die Bewegung des Atriums des Pansens zusammen. Anatomisch wird dieser Magenabsehnitt auf Grund des gleiehen Aufbaues zum Pansen gereehnet. Physiologiseh abet ist dieser Haubenpansenvorhof (MANaOSD und KLEIN) auf Grund der Untersuehungen yon MA~GOLD und KnmN als eine selbst~tndige Abteilung anzusehen. MAXaOr.D und KLEIN fanden ngmlieh, dass der Vorhof seine besondere Innervation hat. CZEPA und STmnER konnten r6ntgenologisch bei erwaehsenen Tieren ein regelreehtes Weehselspiel zwisehen Haube und Vorhof beobaehten, die sog. ,,Haubenatriumrevolution". Diese erfolgt in 3 Phasen:
1. Kontraktion der Haube und gleiehzeitige Ersehlaffung des Vorhofes. 2. Erweiterung der tIaube nnd gleiehzeitige Kontraktion des Vorhofes. 3. Pause, in der Haube und Vorhof ersehlafft sind.
Die beiden Abteilungen werfen sich also gegensei~ig den Inhalt zu, daher bezeichnen auch CZEPA und STmL~ den Vorhof als ,,Sehleudermagen". Bei noeh saugendem Wiederkiiuer ist aber etwa bis zur 3. Woehe diese tIauben- atriumrevolugion noeh nicht ausgebildet (Cz~PA und STmL~R); daffir bestehg in den ersten Woehen ein Weehselspiel zwisehen t taube und Pansen [CzEPA und S~IGLEn, T ~ a U ~ A ~ (4)], welches dagegen wieder bei den erwaehsenen Tieren fehlt. Und zwar sehliesst sieh naeh Cz~Pa und S~mLEa und aueh naeh den neueren Beobaehtungen yon T ~ U T ~ A ~ (4) an saugenden Sehaf- und Ziegenl~mmern an die Haubenkontraktion sofort die Kontraktion des dorsalen Pansensackes und folgend der fibrigen Pansenabsehnitte an. Gelegent- lieh sahen MANao~n und K~E~ aueh beim erwaehsenen Tier eine Koordination yon Hauben- und Pansenbewegung anftreten.
Am Pansen And sehwaehe .peristaltisehe Bewegungen vorhanden [Cz~A und S~G~Eg, T ~ A ~ A N N (4)]. Im Vordergrunde stehen abet die abweeh- selnden Totalkontrak~ionen des dorsalen und des ventralen Pansensaekes mit

602 W. LENXEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Saugegiere.
ihren Endblindsi~eken; der dorsale Pansenteil kontrahiert sieh stets vor dem ventralen, dann bleiben beide Pansens~eke kurze Zeit in ersehlafftem Zustande. Die Endblinds~eke ziehen sieh naeh TRA~:TMa~N (4) am st~rksten zusammen, und zwar oft bis zum Versehwinden des Lumens, dadureh wird eine kr~ftige Durehmisehung des Inhaltes gewi~hrleistet. Antiperistaltisehe Bewegungen des Pansens linden entgegen der Annahme yon WESrE~ (1, 2) nieht start [TnAuT- ~ANN (4)]. Der noeh saugende Wiederki~uer zeigt die gleiehen Bewegungs- formen auch des Pansens [CzErA und STmL~R, TRAUTMANN (4)].
Die Zahl der Pansenbewegungen betr~gt bei den erwaehsenen Wieder- ki~uern im Durehsehnitt 9,--3 in einer Minute. Bei noeh saugenden Ziegen- l~mmern fanden TRAVT~ANN (3) vorwiegend 3 Kontraktionen in einer Minute, CzEPa und STmLER 10 Kontraktionen in 3 Minuten. Eine Pansenrevolution dauerte naeh STmLE~ bei einem erwaehsenen Ziegenboek und einem erwaeh- senen Sehaf 9,7--9,7,8 Sekunden, bei einem 11 Tage alten Zieklein 19 Sekunden. WIESE beobaehtete dagegen beim Rinde eine Zunahme der Frequenz mit dem Alter.
Die die Zahl der Pansenbewegungen beeinflussenden Faktoren (Fiitte- rung, KSrperbewegung u. a.) sind besonders aus klinisehen Griinden eingehend untersueht worden und haben eine weitgehende Bertieksiehtigung in der Dar- stellung yon E. MANGOr,D (4) erfahren.
Die motorisehe Funktion des Psalters ist noeh reeht unklar. Vor dem RSntgensehirm konnte CZEPA und STIGSER bei Ziegen niemals eine Bewegung des Bl~ttermagens beobaehten. Bei elektriseher Reizung bekamen MANGOLD und KLEIN eine langsame tetanisehe Zusammenziehung, die sieh naeh Auf- hOren der Reizung ebenso langsam 15s~e. W~ST~R (1, 9,) kommt dagegen auf Grund seiner Un~ersuehungen mit der Ballonmethode dutch eine Pansen- fistel zu der Annahme, dass am Psalter peristaltisehe und antiperistaltisehe Kontrak~ionswellen ablaufen und auf dem HShepunkt der Kontraktion eine plgtzliehe Ersehla~fung eintrete und dadureh der Inhalt der ersten Vorm~tgen angesaugt werde. Aueh SC~ALK und ASIADON sehliessen aus ihren Registrie- rungen auf eine periodisehe Erweiterung des Psalters. Die Ansaugetheorie entbehrt einer physiologisehen Grundlage. Ein Hohlorgan mit schlaf/en Wiinden; wie es der Psalter ist, kann sich unm6glich aktiv, ohne Druckregulierung, erweitern. Wahrseheinlich hat der Psalter auf die WeiterbefOrderung des Futters keinen wesentliehen Einfluss (CzEPA und STmT,E~).
Auf eine Eigenbewegung der Blgtter weisen neben den Muskellagen die vorkommenden Ganglienzellen (ET,L~N~ERGE~) in den Bl~tttern bin. Die yon Czm, A und STmL~ fiber die Ansgtze der Blgt{er an der Psalteroberflgehe beobaeh{e~en hinweglaufenden Wellen deuten eine Versehiebung der Bli~tter aneinander an.
Eine gesonderte Bespreehung erfordert die Meehanik der Schlundrinne, jener eigenartigen rinnenfSrmigen Fortsetzung tier SpeiserOhre am Boden der

Mechanik des mehrhShligen Magens. 603
Haube zur HaubenpsalterSffnung hin und weRer zur PsaRerrinne in den Lab- magen. Durch Aneinanderlegen der begrenzenden wulstigen Lippen wird die Sehlundrinne zu einem gesehlossenen Rohr. Das Sehliessen der Sehlundrinne erfolgt reflektorisch bei Reizung bestimmter Gegenden des Raehens und wird daher, wie WESTEa (3) am Kalbe festgestellt hat, durch den Sehluekakt aus- gelSst (Sehlundrinnenreilex). Beim jungen noeh saugenden Wiederk~uer gelangt daher die getrunkene Milch dureh die geschlossene Schlundrinne und die PsaRerrinne direkt in den Labmagen. Wird dagegen die Raehensehleim- haut an~sthesiert [TRAuTMA~ und ScR~IT~ (2)] oder die FlfissigkeR unter AusschaRung des Rachens rnit der Sonde in den oberen Oesophagus eingeffihrt (CzEPA und STIGLE~), SO gelangt die Fl~issigkeit nieht direkt in den Labmagen, sondern in die Vorm~gen Haube und Pansen, weil die Schlundrinne nicht geschlossen ist. StSrungen der reflektorisehen Schliessung der Sehlundrinne treten naeh Beobaehtungen yon Tt~AUThIANN am saugenden Ziegenlamm bei Aufnahme kaRer FltissigkeRen und bei Erkrankungen des Verdauungskanals ein. MR der Aufnahme vegetabiliseher Nahrung erfolgt der reflek~orisehe Schluss der Schlundrinne nieht mehr regelm~ssig. TR~TZTMANN und SCHMITT (2) sahen dann n~mlich bei Ziegenl~mmern yon 3 Monaten, denen Fisteln am Pansen und am Labmagen oder am Netzmagen und am Labmagen angelegt waren, einmal aus den Vorm~gen, einmal aus dem Labmagen die aufgenom- menen FltissigkeRen, Milch oder Wasser, abfliessen. Selbst bei Ziegen fiber 1 Jahr [T~A~T~A~N und SCHMITT (2)] und erwaehsenen Schafen (LA~eE) kann bei Wasseraufnahme der Sehlundrinnenreflex bisweilen noeh vo]lkommen funk~ionieren. Das Rind zeigt anscheinend nach WESTER (3) gegenfiber den kleinen Wiederk~uern den Un~ersehied, dass beim Rinde der Sehlundrinnen- reflex ftir Milch naeh der Saugeperiode noch monatelang bestehen bleiben sell, dagegen fiir Wasser sehon einige Wochen nach der Geburt versehwinden sell (W~sTEa). Die in~eressanten Befunde yon W~STER, dass mR Natrium- chlorid versetztes Wasser beim ~Reren Rind wieder den Sehlundrinnenreflex auslSse, konnten weder beim Sehaf (Ross, LENKEIT und COLOMBO:S), noeh bei der Ziege (T~AvT~ANN) in vollem Umfange bestatigt werden. Vergleiehend physiologiseh ist dieser Unterschied nieht reeht verst~ndlich. Besteht die Annahme yon W~STE~ zu Reeht, dass neben den Proteinen vorwiegend die Natriumsalze den Schlundrinnenreflex auslSse, so mfisste dann der grSsste Tell des stark alkalisehen Speiehels des Rindes direkt in den Labmagen fliessen. Aueh naeh l~ngerem Dursten (48 Stunden) sahen LE~<E~ und ColumBUS unter Anwendung einer neuen nicht-operativen Methode (S. 630) entgegen den Angaben yon Ross keinen vollkommenen Sehluss der Sehlundrinne bei ~lteren Tieren. Die eintretende UnregelmassigkeR des Sehlundrinnen- reflexes mR Alterwerden der Tiere is~ zum Tell au~ Mlm~hliehe Rfiekbildung der Sehlundrinnenmuskulatur zurfickzuitihren (TRAuTMANN) und daher yon der Art der FltissigkeR unabh~ngig. Es is~ aber aueh nicht ausgesehlossen,

60r W. LENKiEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Sgugetiere.
dass die Reizempfindlichkeit far den Schlundrinnenreflex mit zunehmendem Alter abnimmt. Die lgeflexzeit ist anscheinend bei iilteren Tieren mit noch
funktionierendem Schluss der Schlundrinne v e r l a n g e r t [TRAuTMANN und SCItMITT (2)].
2. Die I n n e r v a t i o n der u u n d
des Labmagens. Eine endgtiltige Kt~rung des Einflusses des Nervus vagus auf die Bewegungen der einzelnen Magenabteilun- gen des Wiederkiiuers haben die Unter- suehungen yon MANGOLD und KLEIN ge- bracht. Eine der wichtigsten Feststellungen ist die, dass die Vagusversorgung des ge- samten Wiederkiiuermagens doppelt ge- sichert ist. Sowohl der rechte ale auch der ]inke Bauchvagus erhalten Fasern aus den beiden Halsvagusstiimmen. Innerhalb der Brust lindet niimlich zwischen d e n beiden Vagusstiimmen ein Faseraustausch start (Abb. 8). Der Verlanf der anastomo- sierenden Fasern ist wenig einheitlich, er zeigt betr~ichtliche individuelle Schwan- kungen (MANGOLD und KLEIn, C~IST).
Nach Durchschneiden des reehten oder des linken Halsvagus sahen MANGOnD und KnE~N keine nachhaltigen St(irungen auf- treten. Das kurz nach der Operation aus- gebliebene Wiederkauen nnd der erloschene
Abb. 3. Vagussystem des Ziegenmagens. Appetit stellten sieh naeh 3--5 Tagen wieder r . V . rechter, I.V. linker ttalsvagusstamm, ein. Aueh Atrophien einzelner auf der Seite fec. Nervus reeturrens vagi. Der Hnke Bauchvagusstamm geht hinter tier Haube der Vagotomie gelegenen Magenabsehnitte vorbei zur Raekseite des Pansen. Der konnten entgegen iilteren Feststellungen reehte verzweigg sieh in den ~anben- herren, den Psalter-Labmagennerv und (ELLENBERGER) n i c h t gefunden w e r d e n den an aer neber vorbeiziehenden langen (MANGOnD und KLEIN). Eine Ausscheidung Pylorusnerven. (Naeh NIANGOLD u. KLEIN.)
beider Nerven durch Abkiihlung hat naeh den r0ntgenologisehen Beobaehtungen yon PoPow, K~D~JAVCEW und K~A- sousKY ein Aufh6ren der spontanen Bewegungen der Haube zur Folge. Dutch elektrisehe Reizung eines Halsvagusstammes kSnnen Kontraktionen des Pansens, der I-taube und des Labmagens ausgel6st werden (MAcqSOLD und KLEIN, POPOW). Am Labmagen sollen die Kontraktionen jedoeh nur bei Abwesenheit der Spontanbewegungen auftreten, wiihrend vorhandene Bewegungen am Lab- magen durch Reizung des Halsvagus gehemmt werden [PoPow (2/3), KUDR-
JAWCEW und I~RASOUSKY); eine Verstiirkung der Bewegungen wurde seltener

Mech~nik des mehrh6hligen 5~gens. 605
beobachtet. Der Reizerfolg hgngt darnach wesentlich yon dem im Moment der Reiznng vorhandenen Tonus des Labmagens ab. Wie n~mlich MCSWINEY und ~VADOE am Katzenmagen beobachtet haben, ruR Vagusreizung bei ge- ringem Tonus des Magens Vermehrung der Kontraktionen, bei hohem Tonus dagegen Hemmung der Kontraktionen hervor. Funktionell gleichartig scheinen aber beide Halsvagi nicht zu rein. MANGOLD und KnE~N konnten eine Kontrak- tion der Haube yore rechten Vagus dutch weR schw~chere elektrische StrSrae erreichen als yore linken Vagus aus.
Die beiden Bauchvagusnerven versorgen bereRs verschiedene Gebiete. Der linke Bauchvagus ist der motorische Nerv fi~r den Pansen, der rechte far Haube, Psalter und Labmagen. Die Durchschneidung des linken Bauch- vagus ist ohne schwere Folgen ltir das Tier; selbst bei einem 8chaf, dem sowohl der rechte Halsvagus als auch der linke Bauchvagus durchschnRten war, konnten ~([ANGOLD und I~nEIN wahrend der sechsmonatigen Beobachtung im gesamten physiologischen Verhalten keine StSrungen feststellen. Dagegen ftihrt die Durchsehneidung des rechten Bauchvagus zu einem t6dlichen Aus- gang; im Vordergrund steht dabei das Bild der Pylorusstenose und der Lab- magenatonie. Demnach ist aueh beim Wiederk~tuer wie bei anderen Sgngern der Vagus der den Pylorusschluss hemmende Nerv.
Recht unklar seheint noeh die Pharmakologie des Wiederk~uermagens zu rein. PoPow (3), KUDRJAWCEW nnd I(~ASOUSXY konnten ngmlich nach PilocarpinvergiRnng durch Vagusreizung keine Kontraktion des Netzmagens hervorrufen; die Autoren nehmen danach an, dass das Pilocarpin auf die En- dungen des Vagus l~hmend einwirke. Dieses wtirde den bisherigen Er~ahrungen an anderen Tieren widerspreehen.
3. Der Meehanismus des Wiederkauens. Das HinaufbeiSrdern der Wieder- kaumasse aus den Vorm~tgen in das Maul (Re~ektion), der Wiederkauakt und das Abschluctsen der W~ederkaumassen rind die einzelnen zeRlich aufeinander- folgenden Phasen des Wiederkauens.
Der Wieclerkauakt hat naturgem~tss die grSsste Auimerksamkeit auf rich gezogen, wie aus der umfassenden Darstellung yon E. MANGOLD (4) hervor- geht. In neuerer Zeit rind keine weReren Beobach~ungen mehr ersehienen, weshalb im einzelnen auf die obige Zusammenstellung hingewiesen wird. hTach ELLE~BERGE~ kaut das Rind bei Hen- und Strohf~itterung tgglich in 6--8 Peri- oden yon 40--50 Minuten Dauer wieder. Schafe und Ziegen sollen hgufiger und auch schneller wiederkauen ( S o M ~ ) . Die Zahl und Dauer der Wieder- kauperioden wie die Zahl der Kaubewegungen liir jeden Wiederkaubissen kann natfirlich wesentlieh yon der Art und BeschaffenheR des Futters beein- flusst werden (MA~sOnD). Im Maule verweiR eine Wiederkauportion 20 his 75 Seknnden ( W ~ E ~ , RAue~) ; n a c h dem Absehlucken gelangt diese wieder in die Vorm~gen zurttck. Die rejizierte Futtermasse ist, wie die RSn~gen-

606 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Sgugetiere.
untersuehungen yon STmLER (1) siehsr ergsben habsn, keine bisssnartJg geformte Masse, sondern sin v011kommen formloser Brei.
Der Mechanismus des ttinaufbefOrderns des Vormageninhaltes in die MundhOhle, get Re]ektion, hat in neuerer Zeit seine endgCtltige Aufkl~rung eriahren. Ursprtinglich wurde sine aktive Beteiligung dsr Bauchpresse und der Vormagen und der SpeiserOhre an der Rejektion viel diskutiert. Auf Grund anatomiseher Tatsachen und rOntgenologiseher Beobaehtungen kam E. MANaOLD (4, S. 203) vor einiger Zeit zu dem entgegengesetzten Stand- punkt, na.mlieh dass an dsr BefOrderung der Wiederkaumasse vom Vor- magen in den Oesophagus iiberhaupt keins aktive Beteiligung irgendeines Organes notwendig sei. Die Mfindung der SpeiserOhre liegt stets fief unter dem Niveau des Inhaltes des Pansens, wovon man sieh leicht bei einem Tier mit Pansenfistel tiberzeugen kann. Diese hydrostatische Druckdifferenz, die bei der Ziege etwa 10--15 em Wasser betri~gt [STIGLE~ (1)], genfigt naeh E. MANGOnD, um den Vormageninhalt bei oftener Kardia in die SpeiserOhre tibertreten zu lassen. Dureh den ziemlieh horizontalen Verlauf der Speise- rOhre bei den Hauswiederk~uern wtirde eine weir hinaufreiehende Ftillung erleiehtert, so dass der Mageninhalt fiber das stwa in der Mitts tier SpeiserOhre gelegene Kontraktionszentrum [MANGOnD (4)] hinausreiehen kann. Naeh den Untersuehungen yon MANGOLD und INAOKA an Hunden und Kaninehen treten n~mlieh neben Totalkontraktionen am ganzen Oesophagus noch tetanische Kontraktionen auf, die sich yon einem pnnetum fixum, dem Kontratctions- zentrum, aus naeh dem Munde und hash dem Magen hin ausdehnen. Der fiber dem Kontraktionszentrum gelegene Vormageninhalt wtirde danaeh in das Maul und der darunter liegende Inhalt zurfick in den Vormagen bef/Jrderg werden (MANaOnD). Untersttitzt wird die Theorie yon MAN~OnD dureh die Tatsache, dass dureh Senken des Kopfes des Tieres alas Auftreten des Wieder- kauens gefOrdsrt wird (CZEPA und STIGnE~). Bei der Naehprtifung dieser Anschauung konnte STmLE~ (1) tatsiJ.chlieh durch kfinstliehe 0ffnung d er Kardia mittels sines Vaginalspeeulums eine Versehiebung des Panseninhaltes in den Sehlund hinein und manehmal sogar his in den Mund bewirken. Aber die Menge des /ibergetretenen Inhaltes war ziemlieh gering; und weiter kann durch dis hydrostatische Druckdif[erenz allein die betr~chtliche Gesehwindigkeit, mit der die Wiederkaumasse naeh STmLERs (1) Beobaehtungen im RSntgsn- bilde, naeh dem Eintritt in den Sehlund smporsehiesst, nieht bewirkt werden. Dazu sind Hilfsmechanismen notwendig.
Viel ist hierbei die Drucksteigerung dutch Kontraktion dsr Bauchmus- kulatur (FLOLII~ENS, ]~LLENBEttGEtr AGGAZZOTTI, zitiert naeh STIGLER) und der Vormiigen (ELr,ENBE~GER, FO3~ U. a.) diskutiert worden, durch die dsr Inha]t in den Schlund hineingepresst werden sollte. Diese Ansehauungen sind durch Untersuchungen yon WES~E~ (2), BEI~G3IANN und D~zKES und in neuerer Zeit besonders eingehend yon STIa~E~ (1) widerlegt worden. Als einwandfrei

~echanik des mehrhShligen Magens. 607
sicher hat sich dagegen die Ansicht yon CgAUVEAU erwiesen, dass der Vor- mageninhalt aus den Vormiigen in die Speiser6hre hineingesaugt wird. Die Saug- wirkung wird naeh der al%en Theorie yon CgAUVEAU und TOUSSAINT dadureh bedingt, dass durch ruckartige tiefe Inspirationsbewegung bei geschlossener Glottis ein Un~erdruck im Thorax und damit auch im Brustoesophagus entsteht. Dutch diese passive Erweiterung der SpeiserShre erfolgt dann die Ansaugung des Vormageninhaltes. WESTER (1, 2) will die Ansaugung durch aktive Ver- kfirzung der Speiser5hre und dadureh erfolgende trichterfSrmige Erweiterung des eaudalen Endes an der Kardia erkl~ren. Diese Annahme konnte STIGLE~ durch seine neuen umfangreichen Untersuehungen als vollkommen unhaltbar
naehweisen, die Richtigkeit der Theorie yon C]~AUVEAu-TousSAINT abet ebenso sicher beweisen. Dutch Anlegung eines Pneumothorax, wodnreh das Zustande- kommen eines Unterdruckes im Brustraum verhindert wird, konnte ngmlieh STIGL~n dem Tier das Wiederkauen unmSglich machen. Es ist fiberhaupt das Verdienst STIGLERs, dureh versehiedene Methoden den Vorgang der Rejek- tion endgiiRig geklgrt zu haben, so dass eine vollkommene Darstellung der Rejektion gegeben werden kann.
Zu Beginn der Rumination wird Speichel abgeschluekt [WEsTE~ (1, 2), STmLE~] und dann scheint meist eine Kontraktion im oralen AbschnRt des Brustoesophagus zu erfolgen, wodureh noeh in der SpeiserShre vorhandene LuR ausgepresst wird. Gleich darauf beghmt die inspiratorische Ansaugungs- phase der Re]ektion (STIGLER); es schliesst sich die Glottis und tri t t eine ruck- artige Inspiration ein, wobei zugleich Hals und Kopf gestreekt werden; der Brustoesophagus erweitert sich, und dutch den negativen Druck in seinem Innern wird durch die sich gleichzeRig 6ffnende Kardia Pansen- und Hauben- inhalt angesaugt und der Oesophagus bis zum Hals geftillt. Unmittelbar nach der Ansangung schliesst sich die Kardia und es beginnt die exspiratorische Auspressungsphase der Re]ektion, die Auspressung der Wiederkanmasse ans dem Brustoesophagus in den Halsoesophagus und in das Maul hinein. Bei noch immer gesehlossener Glottis erfolgt in dieser Phase eine Exspirationsbewegung, wodurch der intrathorakale Druck und damit aueh der Druek innerhalb des Brustoesophagus gesteigert wird. GefSrdert wird diese intrathorakale Druck- steigerung noch dureh Kontraktion der Banehmusknlatur, die den Bauch- inhaR gegen das erschlaffte Zwerchfell drfickt. W~hrend dieser exspiratorischen Drueksteigerung lguR als zweRer Hilfsmechanismus dieser Phase eine anti- peristaltisehe Welle am Oesophagus ab [STIGLE~ (1, 2)] und der angesaugte Vormageninhalt wird dadurch mit einer betr~chtlichen Gesehwindigkeit kon- tinuierlich his in das Maul getrieben. Ist die Antiperistaltik des Oesophagus aus irgendeinom Grunde nngent~gend, so wird die Bauchpresse besonders stark an der Rejektion beteiligt (STmLEn). Nach AnkunR des Mageninhaltes im Maul wird dessen fiberschfissiger fltissiger Anteil durch eine peristaltisehe Schluekwelle der SpeiserShre in die Vormggen zurfickbefSrdert. Damit ist die

608 W. LEI~VKEIT: Neuere Ergebnisse der vergh Physiologie der Verdauung der S~ugetiere.
gejektion beendet, die Glottis 5ffnet sich wieder und alas eigentliche Wieder- kauen beginnt (Tabelle 4).
Ausschlaggebend sind also ftir das Zustandekommen der Eejektion die Jmderungen des intrathorakalen Druckes. Dureh Tracheotomie, dureh Lgh- mung des Zwerchfelles, wird das Wiederkauen be~r~chtlich erschwert und aber nieht ganz und gar, wie dureh den Pnenmothorax, verhindert, wie schon erwahnt [STmL~.~ (1)]. Die Banehpresse ist yon untergeordneter Bedeutung; nur in der 2. Rejektionsphase, in der exspiratorisehen Auspressungsphase,
Tabelle 4. D ie l~e e k t i o n d e r W i e d e r k a u m a s s e .
Ansaugungs- Auspressungs- phase phase
Geschlossen 1. Glottis . . . . . . 2. Atmung . . . . . 3. Zwerehfell . . . . 4. Trachea . . . . . 5. Oesophagus . . . . 6. K~rdia . . . . . . 7. Bauchmuskula tur .
Inspi ra t ion Kont rah ie r t Unte rdruek
Often Unbetei l igt
Exspira t ion Ersehlafft Uberdruck
Gesehlossen Oft kont rah ie r t
spielt sie bisweilen eine unterstfitzende Rolle. Bei Tieren mit Pneumothorax ist die Bauehmuskulatur trotz st~rkster Anspannung nieht imstande, das Wieder- kauen wieder in Gang zu bringen.
Noch nieht so ein- gehend untersucht ist der Mechanismus der Kardia-
5Hnung beim Wiederkauen. In der Ruhe ist die Kardia locker geschlossen [AGGAZOTTI, STIGLER (1)] und 61fne~ sieh nur beim Schluckakt, bei Ankunft der peristaltisehen Welle der Speiser6hre an der Kardia, beim Wiederkauen (STmLER) und ausserdem beim Ructus.
Der Schluss der Kardia erfolgt anscheinend nieht sphincterartig nach den letzten Untersuehungen yon STmLE~, sondern es zieht sich wohl das kardiale 0esophagusende auf fast einer Fingerl~nge starker oder schw~cher zusammen. Eine mit dem Sehluss der Kardia gleiehzeitig erfo]gende Kon- traktion der Zwerehfellschleife (MANGOLD und K L E I N ) scheint nach STIGLER nicht stattzufinden.
Die Tatsache, dass es sieh bei dem Sehluss der Kardia nicht um einen richtigen Sphinctersehluss handelt, sondern um eine Zusammenziehung des caudalen 0esophagusendes, ]~sst auch die KardiaSffnung beim Wiederkauen etwas klarer erscheinen. Mit Beginn tier Ansaugungsphase wird wahrscheinlich dutch die intrathorakale Ansaugung der 0esopbaguswand eben dieser Schluss gelSst. Eine KardiaSffnung durch Ansaugung der benachbarten Magenwand [WEsTER (1)] dutch hSheren Fiillnngsdruck im Vormagen ist nach STm~ERs (1) Untersuchungen nicht anzunehmen.
Die AuslSsung der Rumination ist noch problematisch. Der F,gllungsdruck im Vormagen hat hierbei keine urs~chliche Bedeutung; durch Aufbliihen des Pansens ist es nicht m6glich, Wiederkauen einschliesslich Rejektion hervor- zurufen (STIGLER). Durch Berfihren tier Zitzen, dutch Melken, gelingt es fast regelm~ssig nach STmLER, TR~UT~A~-N und SCH~ITT (4), die Tiere zum Wieder-

Mechanik des mehrhShligen Magens. 609
kauen zu bringen, selbst bei vollkommen leeren Vorm~gen. Efforderlich seheint ein gewisses Behaglichkeitsgeffihl zu sein, welches wahrseheinlieh aueh dutch das Melken hervorgerufen wird. Die Bedeutung dieses Zustandes sieht STIGLER in der Beseitigung eerebraler Reflexhemmungen. Von bedeu~endem Einiluss ist vor allen Dingen die Art der Nahrun 9. Naeh der Aufnahme yon Rauhfutter tritt das Wiederkauen wesentlich sehneller ein als nach der Anfnahme einer Breimahlzeit (Sc~EUNEnT, STIOLE~); Ziegenlgmmer, die fiber die Sgugezeit hinaus mit Milch erniihrt wurden, ruminierten bei dieser Nahrung fiberhaupt nicht [TRAuTMANN (3)].
Danach scheint es, dass das Rauhfutter dureh Reizung der Vormagen- sehleimhaut reflektorisch den Wiederkauakt auslSst. Bei diesem Reflex- vorgang beteiligt sich naeh der Annahme yon MAh'aOLD der Vagus nicht nut als motorischer Nerv, sondern auch als zentripetaler sensibler Nerv. Die sen- sible Leitung der Wiederkau- und Reflexbahn bildet sieh nach TRAUTMAN~ und SCSMITT (4) bei jungen Tieren wahrscheinlich erst allmiihlich aus; denn Saugl~tmmer ruminieren auch nach reichlicher Znfnhr yon Rauhfutter nicht (AKssE~-OWA, TRAUTMANN). Operative Unterbreehung der Reflexbahn, beiderseitige Durchschneidung des Vagus am Halse (MANGOLD und KLEIN), ffihrt zur Aufhebung des Wiederkauens. TRAVT~ANN und SCR~ITT (4) beob- achteten bei einer Ziege nach Haubenextirpation ebenfalls ein Ausbleiben des Wiederkauens, was auf eine ZerstSrung der sensiblen Leitung zurfickgeffihrt wird. Die Haube als solche ist n~mlich zum Wiederkanen nicht unbedingt notwendig; denn TRAVT~ANN sah einwandfrei Rumination bei Ziegen, denen die Haube bis auf ganz geringe Reste entfernt worden war. Man kSnnte aus diesen Versuehen aueh vermuten, dass in der tIanbe die Hauptzweigstelle ftir die AuslSsung des Wiederkaureflexvorganges liegt. Dutch meehanische Reizung der tIaubenschleimhaut, der Schlundrinne und des Psalterhalses gelingt es bisweilen, Wiederkauen hervorzurufen [WEsTE~ (1, 2)]. Derartige Versuehe sind nur beweisend, wenn nicht nur der Wiederkauakt sondern aueh der Rejektionsmeehanismus ausgel6st wird.
Die Bedeutung der Rumination liegt in der weitgehenden Zerkleinemng des Futters, was sowohl ffir die bakterielle Zersetzung im Pansen als aueh ffir den Weitertransport des Vormageninhaltes in den Psalter und Labmagen gfins~ig ist. Ansgesprochen lebenswiehtig seheint aber das Wiederkauen nicht zu sein naeh einer Beobaehtung yon TnAUT~AN~ U. Se~ITT (4). Die Ziege (s. oben), bei der seit einer ttaubenexstirpation das Wiederkauen ausblieb, blieb bei fiblieher Rauhffitterung auch ohne wiederzukauen li~ngere Zeit am Leben. Der stark angeftillte Pansen zeigte aber ausserordentlieh starke Bewegungen, wodurch wahrscheinlieh der Ausfall der Zerkleinerung des Futters durch das Wiederkauen bis zu einem gewissen Grade ersetzt wurde.
Die menschliche Rumination ist eine dem Wiederkauen der pflanzen- fressenden Tiere vollkommen analoge Erseheinung. Uber das Vorkommen des
Asher-Spiro, :Ergebnisse tier Physiologic lind expel l~ 35. 39

610 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologic der Verda,uung der Sgugetiere.
Wiederkauens beim Menschen, bei Shuglingen und Erwachsenen, ist wieder- holt such in neuerer Zeit berichtet worden (KoItL~AIvN, ausitihrliehe Literatur bei T~AUT)~ANN), und die Ursache wird in einer Reflexneurose des Magens gesucht. Der Mechanismus wird weitgehend dem des tierischen Wiederkauens ~thnlich sein. Naeh LoEwE, tier selber ruminierte, und besonders naoh S~NGEa beruht das Hinaufbringen der Speisen zum grOssten Teil auf einer 8augkraft in der 8peiserOhre, die durch die inspiratorische Luftverdtinnung im Brust- raum bei geschlossener Glottis bedingt ist. Genauere Untersubhungen beim Menschen liegen nieht vor.
4. Wiederkauen und Erbreehen. Wghrend das Wiederkauen ein yon Wohlbehagen begleiteter normaler Vorgang ist, ist das den ganzen Organismus in Mitleidenschaft ziehende Erbrechen eine pathologisehe Erscheinung. Der Meehanismus des Erbrechens hat aber naeh den neuesten Angaben [W~STn~ (2, 3, 4), 8TmLER (2), HOFfmAnn] vieles mit dem des Wiederkauens gemeinsam. Auch beim Erbrechen erfolgt als Hauptaktion bei geschlossener Glottis eine ruekartige kr~tftige Inspirationsbewegung und durch die dadurch ent- standene Druekverminderung im Brustraum eine Ansaugung des Magen- inhaltes durch die geOfinete Kardia in den Oesophagus hinein. Gleichzeitig mit dem Ansaugen erfolgt aber noeh im Gegensatz zum Wiederkauen eine krgftige Kontraktion der Bauehmuskulatur. Daraus lolgen als weitere Unter- schiede gegentiber dem Wiederkauen, class tier MageninhMt mit um so gr6sserer Krait mundwgrts befOrdert wird, und dass die Menge des Erbrochenen welt grSsser ist als die der rejizierten Wiederkaumasse. 0b auch beim Erbreehen, wie beim Wiederkauen, eine Antiperistaltik des Oesophagus bei der BefSrderung des Mageninhaltes aus der Speiser6hre in den Mund eine Rolle spielt, ist noeh nieht erwiesen, aber naeh STmL~Rs Angaben anzunehmen.
5. Der Ruetus. Bei einer Bespreehung des Wiederkauens ist aueh der Ructus oder das Ri~lpsen, zu erw~hnen, jener den Wiederki-iuern eigentiimliche normale Vorgang, die bei den Pansenggrungen entstehenden Gase dutch die SpeiserShre zu entleeren. Der Ablauf der Gasentleerung ist noch nicht ganz klar. Gegeniiber den glteren Untersuchungen yon WILDE kommt WnsTna zu dem Ergebnis, dass die Entleerungen regelmiissig, und zwar im Zusammen- hang mit den Pansenkontraktionen stattfinden. Der Gasdruek seheint dabei such eine Rolle zu spielen (8Ts Dutch den Zusammenhang des Ructus mit den Pansenbewegungen erklgrt sich vielleieht such der lJbertritt des Gases selbst bei der Abriegelung der Kardia6finung dutch den Panseninhalt, dessen .Niveau nach MAZVGoLn last stets fiber der gardia liegt (s. S. 606). Durch die Pansenbewegungen kommt es zu betriichtlichen Versehiebungen des Inhaltes und dsmit wohl such zum vorfibergehenden Freiwerden der Kardia, in welcher Zeit Gas in die SpeiserShre gelangen kann. Eine spezielle Tgtigkeit, wie sic K~zYwa>-ng in solchen Fgllen annimmt, ist sicher nicht notwendig. Schwieriger ist die Frage, welehe Kri~ite den Gasfibertritt in die SpeiserShre herbeiffihren~

Mechanik des mehrh6hligen Magens. 611
Hierbei spieR wahrseheinlieh auch die inspiratorisehe Druckverminderung im Brustraum eine Eolle; es ist hier wohl aueh eine weir geringere Druck- verminderung ausreichend als bei der Aufnahme der Wiederkaumasse. Es wiire sonst auch nieht das R(ilpsen bei Tieren mit Pansenfisteln (WES~E~) zu erkl~tren. Diese Annahme w~tre experimentell zu erhi~rten. Die 0ffnung der Kardia finder aueh beim Ruetus, wie aus den Versuehen yon MANGOr.D und KLm~ hervorgeht, unter Kontrolle des Vagus start.
6. Der (~bertrRt des Futters aus den beiden ersten Vormiigen in den Psalter und Labmagen. Die abgeschluckte feste Nahrung, einschliesslich Wiederkaumasse, gelangt sowohl bei jungen als auch bei alten Tieren in t taube und Pansen (CzEPA und STIGLER). Gelegentlieh kSnnen aueh kleinere und schmalere Bissen bei jungen Tieren direkt in die Psalterrinne und in den Lab- magen gelangen (Cz~rA und STmLEa).
Die Vorbedingungen ftir den UbertrRt des Futters aus Haube und Pansen in den Psalter und Labmagen sind naeh den Untersuchungen -con SCEEV~E~T und STEIN~AVF genfigende Menge, der Wassergehalt und vor allen Dingen tier Zerkleinerungsgrad des Panseninhaltes. Mangelhafte Zerkleinerung, wie es bei Ausbleiben des Wiederkauens der Fall ist, verl~ngert daher die Dauer des Auf- enthaltes des Futters im Pansen [TnAvT~ANN und SCE~ITT (4)]. BereRs zer- kleinert verabreichtes Futter kann aueh ohne Wiederkauen den Pansen vet- lassen; es bleibt daher ktirzere Zeit im Pansen als grobes FuRer; eine Mahlzeit aus feingesehrotenem Haler hag den Pansen des Sehafes naeh 5--6 Tagen ver- lassen, ganze Haferk6rner erst naeh 8--12 Tagen [LENKnIT (2, 5)]. B e i sehlechfer Ftillung des Pansens, z. B. infolge sehlechter Rauhiutteraufnahme, wahrscheinlich infolge schleehter Pansenbewegungen, kann erst der end: giiRige Ubertritt einer MahlzeR sogar naeh 3 Wochen erfolgt sein [LEN~mT (2, 5)].
Welche Kr~fte den/]ber t rRt des Fritters aus den Vormi~gen in den Lab- magen bewirken, ist noch nieht geniigend klar. Es erfolgt der IJbertritt nur periodiseh und nieht kontinuierlieh. Das den f]bertritt regulierende Organ ist zweifellos der Psalter. An der Ha~ben-Psalter6ffnung ist zwar ein Sehliess- muskel vorhanden, der yon der Muskulatur der Lippen der Schlundrinne gebildet wird (ELLENBEnGEn-BA~,lU). Diese 0ffnung ist aber naeh NESTER (1, 4) oft often oder nur locker gesehlossen, ohne dass jedoeh ein Absehluss erfolgt. 0b der an der Psalter-Labmagen6ffnung liegende Qnermuskelwulst des Psalterbodens sphineterartige Funktion hat, ist noeh strittig. Naeh den Beob- aehtungen yon CZEPA nnd STmSE~ ist diese 0ffnung versehlussfiihig; denn sie sahen manehmal bei alten Ziegen aufgenommene FltissigkeR zwar bis in den Psalter gelangen, aber nieht gleichzeitig in den Labmagen iibertreten. Naeh WnSTE~ (2, 3) kommt dagegen der Absehluss der Vorm~igen gegen den Lab- magen dadureh zustande; dass bei Kontraktion des Psalters der Psalterboden (Psalterbrtieke) sieh gegen den Inhalt des Bliittermagens kr~tRig anpresst und so den Durehgang zum Labmagen versperrt.
39*

612 W. LEI~KEIT: Iqeuere Ergebnisse der vergl. Physiologie der Verdauung der S~ugetiere.
Infolge pl6tzlieher Erschlaffung des Psalters sollte dann naeh WESTER (1, 2) der Panseninhalt angesaugt werden. Auf die Unhaltbarkeit dieser An- nahme ist bereits oben eingegangen (s. S. 602). Ein bei dem Ubertritt des Futters notwendiger Faktor ist sieher das Druckgef~lle yon Haube, Pansen zum Labmagen hin; der Spiegel des Hauben-Panseninhaltes steht namlieh in der Regel fiber der Hauben-PsalterSffnung. Eine Mitwirkung yon Haube und Pansen ist aueh w~hrseheinlich; in den bisherigen Untersuchungen ist dieses merkwfirdigerweise wenig berficksiehtigt worden.
Aufschlussreieh for die Mechanik des Futterfibertrittes in den Labmagen w~ren Untersuchungen an den Tylopoden, denen der Psalter fehlt.
Der Ubergang des Futters aus dem Labm~gen in den Darm erfolgt in der gleichen regulierenden Weise dutch den Pylorus wie bei den Tieren mit einhShligem Magen.
7. Der Fliissigkeitstransport. Auf die Bedeutung der Sehlundrinne and .des Sehlundrinnenreflexes ffir den Wassertransport wurde bereits hingewiesen. .Da der Sehluss der Schlundrinne bei erwachsenen Tieren im Durchsebnitt nieht vollkommen erfolgt, geht beim erwachsenen Wiederk~uer die spontan aufgenommene Flfissigkeit fast immer zuerst in Haube und Pansen (Ross, KRZYWANEE und LAMPE). Auch die Zugabe yon Mineralsa]zen zu den Flfissigkeiten begfinstigt nach den Untersuehungen yon Ross an Schafen enfgegen den Befunden yon WESTEn am Rinde keineswegs den direkten Ubergang der Flfissigkeit in den Labmagen durch die versehlossene Schlund- rinne. Ross land ein direktes Ubertreten in den Labmagen nur dann, wenn die Tiere mindestens 40 Stunden gehungert und gedurstet hatten, u n d zwar in 75 % der F~lle, nach LENKEIT und COLUMBUS ist aueh d~nn tier Ubertritt nicht vollkommen. Ist bereits reiehlich Flfissigkeit im Pansen vorhanden, so kann ebenfalls die neu aufgenommene Flfissigkeit direkt in den Labmagen gel~ngen (SoI~L~E~T, K~Z~WANEK).
Bei den jugendliehen Wiederk~tuern gelangt die Flfissigkeit infolge des prompt eintretenden Schlusses der Sehlundrinne (s. S. 608) regelm~tssig direkt in den Labmagen; aber es erfolgt bei den jangen Tieren aueh regelm~tssig ein Ri~ck[luss z. B. yon au[genommener Milch aus dem Labmagen in die Vormiigen [A~:ssE~owA, T~AVT~ANN und Sc~MImT (3)]. Der Rfickfluss ist sowohl kurz naeh der Aufnahme als auch noch l~ngere Zeit danaeh zu beobachten (T~v~- MANN llYld SCHMITT). Bei erwachsenen Ziegen sahen TRAUTMANN und SCl~I~T nur naeh Zufuhr grosser Meugen Milch in den Labm~gen Milch aus dem Labraagen in die Vormagen zurfickfliessen, und deuten daher hier den Rfick- fluss als ein ,,Uberlaufen" des gefiillten Labmagens.
V. Der Ablauf der Magenverdauung.
1. Einhiihlige M~igen. Der Ablauf der Verdauung in den einh6hligen Miigen ist durch amylolytische und proteolytisehe Prozesse gekennzeichnet,

Der Abl~uf der Magenverdauung. 613
Der Grad der amylolytischen Verdauung wird bedingt durch die neutralisierende und bei einigen Tieren noch diastatische Einwirkung des Speichels, durch die Einwirkung der Nahrungsfermente und der Bakterien, und begfinstigt durch die fehlende Durehmisehung des Mageninhaltes im Fundusteil (E~ENBESGER, SCnEU~ERT). Die diastatische Speiehelwirkung im Magen wird nach den neueren Untersuchungen yon NAKATSUKA beim Mensehen dutch Peptone und Salze wesentlich gefSrdert, so dass dadureh bisweilen die hemmende Wirkung der Salzs~ure fiberwunden wird. Mit dem Vordringen der Salzsaure in den Mageninhalt hinein und der zunehmenden Milehs~ureggrung im Inhalt geht die amylolytische Periode in eine amylolytisch-proteolytisehe und sehliesslich in eine reir; proteolytische fiber.
Bei den Fleisch/ressern tritt die St~rkeverdauung fast vollkommen in den Hintergrund, und anch beim Menschen ist sie nur yon kurzer Dauer. Am ausgepr~gtesten ist die amylolytische Periode in den M~gen mit grossen cntanen Vormggen oder mit grosser Kardiadriisenregion, also bei den Equiden, Muriden und bei den Sehweinen (s. E~LESBERGSR, SCrEWiNESS, BIEDER~ANN). Infolge der geringen Motilitgt der kardialen Abschnitte bleibt bier das mit Speichel durchsetzte Futter eine gewisse Zeit liegen; Bakterien, Nahrungs- mi*teldiastase (ELLENBEROER, MANGOLD-DUBISKI) und z .B. bei Ratte, Schwein noeh die Speieheldiastase kSnnen in diesem Milieu ungehindert ihre amylolytische Wirkung entfalten, die ganz betrgchtlich werden kann. Am gfinstigst~en sind diese Bedingnngen beim Pferd, well dessen Mageninhalt ziemlich lest ist und selbst ira Pylorusabschnitt nicht durchmischt wird. So sollen beim Pferde nach SC~IEUNE~T (2) 3 Stunden naeh der Futteraufnahme bei einer Haferfiitterung 1/s der gesamten Kohlehydrate des Mageninhal~es abgebaut sein. LSNKEIT (1) fand im Vormagen der Ratte nach 1--2 Stunden nach der Futteraufnahme einen Abbau yon 8--11% der aufgenommenen Stgrke, im Drfisenmagen yon durchschnittlieh 6 %. Trotz des nicht unerheb- lichen Abbaues der St~rke ira Vormagen blieb aber die Eesektion des Vor- magens auf die Gesam~ausnutztmg der Stgrke ohne Einfluss.
Bei Kaninchen und Meerschweinchen, denen der eutane Vormagen und die ausgedehnte Kardiadriisenregion fehlten, nghert sieh der Ablanf der Magen- verdauung der des Mensehen, wie auch die alten Untersuchungen yon GRt~TZNE~ (S. auch S c ~ u h - ~ , B~EDE~AS~) fiber die Verteilung des Pepsins ira Magen- inhalt zeigen.
Die Reaktionsverhiiltn~sse des Mageninhaltes werden durch Alkalit~tt des Speiehels, dureh die Magensalzsgure, und bei pflanzlieher Nahrung nicht wenig dutch den Grad der Milchs~uregarung bestimmt. Beim Pferd ist der Mageninhalt nach ROS~NFE~n bis ~/~ Stunde nach der Futteraufnahme infolge der hohen Alkalit~t des SpeicheIs durchweg alkalisch; naeh 1 Stunde wird nur noch im Vormagen alkalische Reaktion angetroffen, wghrend im Fundus- teil bereits eine Acidi~tt, anf HC1 bereehnet, his zu 0,05 %, im Pylorusteil bis

614 W. L~xmT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Sgugetiere.
0,075 % ,corhanden ist. Allmghlich steigt dann die Aoiditgt im gesamten Inhalt
bis auf 0,2--0,4% z. B. naeh 6 Stunden. Die zuerst auftretende Aciditgt ist
nur dureh die Milchs~ture bedingt, spgter tritt dann IICl auf bei gleiehzeitigem
Zurtiekgehen der Milchsgure. Im Vormagenabschnitt tiberwiegt stets die Milchsguro. Teilt man mit It. GOLDSC~IMIDT don Mageninhalt in Aciditi~ts- zonen ein, so liegt eine Milehsgurezone im Vormagen und ein3 im Pylorus- absehnitt, im Fundusabsehnitt die Milehs~uresalzsgurezone. Beim Sohwein (ELLEN~m~OEI~-HOF~EISTmQ ist kurz nach der Futteraufnahme der Magen-
inhalt nur in dor Kardiadrtisenregion alkalisoh in den iibrigen Abteilungen sehon sehwaeh sauer. 1 Stunde naeh der Mahlzeit kann aueh sehon in dem Kardiainhalt z. B. bei Haferfiitterung eine Aeiditgt, auf HC1 bereehnet, ,con 0,02--0,05% auitreten, bei Kartoffelf~tterung bisweilen sogar yon 0,08%. Die Sgurezunahme erfolgt im linken Magen langsamer als im reehten (Fundus und Pylorus), und sehliesslieh kann aueh hier eine Aeidit~t ,con 0,3 % erreieht werden. Aueh beim Sehwein ist der Anteil der Milehsiiure an dem Gesamt- sguregehalt ein reeht betrgehtlieher und kann bei kohlehydratreieher Nahrung sogar iiberwiegen (ELI~ENBEI%GEI%-HoFIVIEISTER). Bei den Fleisch- /ressern und beim Menschen ist am h~iufigsten Salzsiiure allein zu finden.
Die aktz~elle Realction wird bei der geringen Dissoziation dot Milehs~ure ,corwiegend ,con der Salzsiiure bestimmt. Im Mageninhalt dos Plerdes fanden 8TEINMETZER und CAITHAML in 75 Untorsuehungen bei Sehlaehtpferden p~-Werte yon 1,18--6,79, jedoeh in 50% der Untersuehungen 1,1--3,4. Die Werte sind aus dem Gesamtinhalt gewonnen. Im Mageninhalt, der 2 bis 3 Stunden naeh der F~tterung (Hafer, Heu) dureh Ausheberung gewonnen wurde, war die Wasserstoffionenkonzentration im Durehsehnitt goring, da wahrsehoinlieh tier Inhalt aus dem Vormagen stammte; nur in 8 yon 10 Fiillen wurden Werte yon p~ 3,49--3,96--3,71 festgestellt. Leider sind keine regio- niiren Bestimmungen durehgefiihrt worden, so dass die Werte wenig brauehbar sin& Ftir den Mageninhalt der gat te geben SUN und Mitarbeiter p~ 3,2--4,6 an, und zwar liegen hier die Werte bei den jungen Tieren h6her als bei den erwaehsenen (EAST~ANX und MILLEa); beim Kaninehen war im Mittel p~ 1,83 (LANaU~IN) gefunden worden. Beim ttunde betrggt bei Fleisehffitterung die p~ 1,35--4,95 (NAGs), be ider Katze ira Mittel 3,24 (LANoeLI~). Die Werte liegen bei Fleiseh- und Pflanzenfressern durehsehnittlieh fast in der gleiehen Variationsbreite, in der die Wirksamkeit dos Pepsins noeh m/Sglieh is t Nur darin ist ,cielleieht Bin Untersehied, dass die niedrigsten Werte hiiufiger bei den :Pflanzenfressern zu finden sind.
2. Mehrhghlige M~igen. In den gormiigen der Wiederkiiuer erreiehen die Giirnngsprozesse ein ausgesproehen hobos Mass. E s handelt sieh ,corwiegend um eine bakterielle Zersetzung der Cellulose; naeh den mikroskopisehen Unter- suehungen "con W. MEYEa kommt es zur Quellung, Aufquellung und sehliess- lieh zur Riss- und Spaltbildung der Cellulosemembranen. Diese Cellulose-

Der Abl~uf der Magenverdauung. 615
aufsehliessung geht bis zur Bildung yon C02, Ctt 4 und yon Fettsiiuren, haupt- saehlieh Essigs~ure, aueh Milehsaure (TAPPEINE~, ZUNTZ, KLEIN, S. MANGOr,D, Handbueh). In dem Gasgemisch sind noeh N 2 und H2S in geringen Mengen vorhanden; 02 kann tiberhaupt fehlen oder ist nur in ganz geringen Mengen vorhanden. Es herrsehen danach im Pansen doeh fast anaerobe Bedingungen. Eine starke Wasserstoffgarung kann naeh reiehlieher Aufnahme yon Mehl- tr~tnke auftre~en (KLEIN). C02 betr~gt 60--80% und steht zu OH 4 im Ver- h~ltnis wie 2 : 1 his 4 : 1 (KLEIN). Bei hungernden Tieren nimmt CH 4 betr~eht- lich zu. In intensiverem Masse wie die Cellulose verg~ren die Pansenbakterien die leieht angreifbaren Kohlehydrate, Stiirke und Zucker. Je mehr yon diesen Kohlehydraten vorhanden sind, um so weniger kommt es zur LSsung der Cellu- lose; mit der geringeren AuflSsung der Cellulose soll es naeh der alten Lehre (ZUNTZ) aueh zu einer geringeren Verdauung tier yon der Cellulose eingesehlos- senen N~hrstoffe kommen. Dadurch wird das Eintreten der ,,Verdauungs- depression" naeh reichlieher Verftitterung yon freien Kohlehydraten, z. B. yon Mehl, Kartoffeln erkl~rt. Naeh den neueren Untersuehungen (s. Ablauf der Darm- verdauung S. 624) am Mensehen(HEuPKE) und am Pferd (FuIsOHBIER) ist eine AuflSsung oder ErSffnung der Cellulosenmembranen jedoeh nieht unbedingt notwendig; HE~-~KE wie FRISCnBIER fanden n~tmlieh, dass die Verdauungs- fermente z. B. des Dtinndarmes die Zellmembran durehwandern und bei un- erSffneter Zellwand den Inhalt herausverdauen kSnnen. Wahrseheinlich ist hierzu ein bestiminter Quellungszustand notwendig. Dasselbe dtirfte auch ftir den Wiederk~uer gelten; es wird danaeh aueh aus den Pilanzenzellen, deren Membranen im Pansen nicht zerstSrt aber gequollen sind, der Inhalt im Lab- magen und Dtinndarm herausverdaut werden; die Ursaehe der Verdauungs- depression k6nnte darnaeh eine andere sein. Sie ist ~-ielleicht mehr in den Fermentabschnitten des Verdauungskanals zu suehen; ob die relativ ver- mehrte Zufuhr der Cellulose im Labmagen und Dtinndarm infolge der geringen Zersetzung im Pansen dabei eine Rolle spielt, ist noch ungewiss.
In geringem Umfange wird aueh Eiweiss im Pansen abgebaut, das geht bereits aus den alten Befunden yon Spaltprodukten hervor (TAPPEINER, S. MANOOLD, SCI~EU~EUT, SCHIEBLICH). Der Abbau erfolgt vorwiegend dureh proteolytisehe Bakterien; dass durch den bei jungen Wiederk~tuern beobaehteten Riiektritt des Labmageninhaltes in den Pansen (S. 612) das Pepsin etwa eine aueh nur geringgradige verdauende Wirkung noeh austibt, ist infolge des un- g~nstigen Milieus nicht wahrseheinlich. Interessant und yon Bedeutung ist d ie Frage der Synthese des N aus niehteiweissartiger Verbindung zu Eiweiss im Pansen. Dass im Wiederk~tuerorganismus gegentiber anderen Tieren der Stiekstoff nichteiweissartiger Natur in Eiweiss tibergeftihrt werden kann,
geh t wie aus ~Iteren Arbeiten aueh aus den neueren Untersuehungen yon E~UENBERG. und Mitarbeiter hervor. Bei Milchktihen konnte weitgehend alas Kraftfuttereiweiss dureh Ammoniumbicarbonat ersetzr werden, ohne dass

616 W. LI~NKIEIT: Neuere Ergebnisse der vergl. Physiologie der Verdanung der S/~ugetiere.
es zu einem R~tckgang in der Milchproduktion kam. Es sind abet keineswegs s~tmtliehe stickstoffhaltige Nichteiweissverbindungen als Eiweissersatz geeignet; so z. B. wohl nicht der Harnstoff; zusammen mit AmmoniaksMzen verfiittert kann allerdings aueh der Harnstoff eiweisssparend wirken (HoscAMP-Ko~'- D~LA). Diese auch f/ir die praktische Fiitterung wichtige Frage kann heute noeh nieht durchweg absehliessend beantwortet werden.
Der Auibau der nichteiweissartigen stiekstoffhaltigen Stoffe zu braueh- barem Eiweiss wird mit grosser Wahrscheinliehkeit dureh Bakterien des Pan- sens durchgeftihrt, und zwar dutch Aufban zu organisiertem Bakterieneiweiss
�9 (I-IAGEMANN). E8 ist nicht ausgeschlossen, dass auch die Infusorien des Pansens hierbei eine Rolle spielen. Es sprieht nieht dagegen, dass die Infusorienzahl (s. unten) bei VerftiRerung yon Ammoniumacetat und Harnstoff nieht zu- nimmt (F~aBE~). Das VerhaRen bei Ammoniumbiearbonat ist noch nicer naehgeprfiR. Der Anteil des Infusorien-N am Gesamt-N des Panseninhaltes betr~gt nfi,mlieh im DurehsehnRt 10--15 %, nut im Zustand erhShten Eiweiss- stofiwechsels (E. MANGOLI)) 15--20% (MA~GO~O und SC~MI~T, KI~AlV~E~, Sc~w~z) . ScHwA~z spricht sogar die Auffassung aus, dass der grSsste Tell des aufgenommenen Futtereiweisses im Pansen in Mikroorganismeneiweiss (Bakterien, Infusorien) umgewandelt und als solehes dem Wiederk~uerorganis- mus zugef~hrt wird.
]3ie In[usorien und Bakterien im Pansenint~lt w/~ren da~tuch gleiehwertige Symbiontea~ des Wiederk~uers, nur mit dem Untersehiede, dass die Pansenbakterien fiir das Leben des Tieres notwendig, die Panseninfusorien dagegen nicht ~nbedingt notwendig sind. l)ber die Bedeutung der P~nseninfusorien Nr die Ern~hrung des Wiederkguers sind besonders in den letzten Jahren zahlreiche Untersuchungen durchgefiihrt worden (s. das Sammelreferat yon E. MA~eo~ mit vollsti~ndiger Li~r~turfibersicht). N~eh tier heutigen Kenntnis kommen in den WiederkS, uer- m~gen welt fiber 100 Arten vet (MA~GOLD). Die normale ln]usorienzahl betr~gt beim Schaf und Ziege naeh F ~ E R im Durehsehnitt 1000 pro Kubikmillimeter bei normaler Ern~hrung; mit Versehleehterung des Ern~hrungszustandes sinkt aueh die Zahl. Dureh Hungern k6nnen die Infusorien in 3--4 Tagen zum Verschwinden gebraeht werden (M~GOLD und S c I ~ r r w - K ~ , T~m~), desgleichen dutch partielles ttungern mit ~lleiniger Zufuhr yon Milch (Doe~L und W]mqOOl~ADOWA); Mleinige Verffitterung yon Gerstensehrot-Leinkuehenmehl-Tr~nke ohne Zu- fiitterung yon Heu maeht die Tiere in 8 Tagen infusorienfrei. :Bei erneuter Zufuhr yon tIeu treten die typischen Panseninfusorien (s. M~NaOLD) nieht wieder auf; die eehten tteuinfusorien sind zw~r im P~nseninhalg in geringer Zahl vorhanden, zeigen ~ber keine Tendenz, sieh ~u ver- mehren (M~xoOL~ und L~m~[~). Wie jetzt endgfiltig feststeht, treten die spezifischen Pansen- infuso~ien erst wieder bei Zusammensein mit infusorienhMtigen Tieren auf (Kontak~hyTolhese: B~oK~, N~GOS~ und g ~ ) . Aueh das erste Auftreten der Panseninfusorien bei L~tmmern oder Iffi.lbern ist auf den Kontakt mit der Mutter und anderen Tieren zuriiekzufiihren. ])ass die Infusorien nieht sehon wghrend der Sgugeperiode auftreten, ist dureh die den Infusorien seh~dliehe Milchn~hrung bedingg, und zwar infolge der S~urebildung dutch Anderung der Wasser- stoffionenkonzentration (NAtgOOLD und USU~LLQ. G~genilber den einzelnen Ns zeigen die Panseninfusorien ein bestimmtes Verhalten. St~rke wird yon ihiien in um so gr6sserer Menge aufgenommen, je kleiner die Stgrkek6rner sind (M~aOnD und Usv~ns~); etwa sehon 3 Stunden naeh der S~/ixkeaufnahme is~ Glykogen im Infusorienleib naehzuweisen (TmEg, UsvELn][). Cellulose ~Jrd dutch die Infnsorien direkt nieht angegriffen; die Cellulosezersetzung im Innern tier Infusorien ist nur m0glieh dutch die mRaufgenommenen Bakteriem Der Eiweissbedarf der Infusorien ist deutlioh ~usgepragt. :Es besteht ngmlieh ein gewisser Pgr~llelismus zwisehen

Der Ablauf der Magenverdauung. 617
der Zahl der Panseninfusorien und dem Eiweissgehalt des Putters; bei alleiniger tteufiitterung nimmt die Zahl ab, bei Kraftfutterzufuhr wieder zu (Fv.~B~u, Mow~Y und BECK~U). MANGOLD sieht daher eine Bedeutung der Infusorien darin, dass sie durch Umwandlung yon Pflanzen- eiweiss in leichter verdauliohes arteigenes Eiweiss dem Wiederk~uer als Eiweissquelle dienen (s. oben). Naah der yon F~UBE~ und WI~OC~ADOWA angegebenen Teilungsquote der Infusorien yon 7% wiirden bei der konstanten Zahl der Infusorien taglich 7% in den Magen und Darm gelangen. Da naoh !~r und SCmV~TT-K~A~ das Gesamteiweiss in 3 kg Panseninhalt 56,25 g betr~gt und die Infusorieneiweissmenge 8,44g (Infusorien-N-15% des Gesamt-5~ des Panseninhaltes), wiirden also nach ~A~OOLD (Sammelreferat) dem Schaf t~glich 0,600 g Infu- sorieneiweiss zugeffihrt werden, was etwa ] % der flit das Schaf zur ErhaItung t~glich not- wendigen Eiweissmenge entsprechen wiirde. Viel ist auch die mechanische Wi~kung der Pansen- infusorien erSrtert worden; infolge ihrer grossen Zahl und intensiven Bewegung kSnnen sie die Durchmischung, die Auflockerung und damit die Maceration des Panseninhaltes begtinstigen (MANOOLD, SCHEWN]~RT). Die Versuche, die Infusorien zu ziiohten, sind nicht gelungen (WILLING).
Die Reaktion des Pansen- und Haubeninhaltes wird allgemein als alkalisch bezeichnet. KREIPE gibt ffir das Rind p~-Werte yon p~ 7,5--8,0 an; F~BER fand beim Sehaf p~ 7,6--7,0 und S c n w ~ z und G ~ a ~ bei 32 Rindern sogar 8,61--9,68. Die ]etzten Werte sind sehr unwahrscheinlich, denn sie sind grSsser als die des kontinuierlieh zufliessenden Speiehels mit p~ 7,19--8,9, beim Rinde (s. Tabelle 1), d. h. der Panseninhalt w~re alkaliseher als der Speichel. Weiter ist noeh daran zu denken, dass die im Panseninhalt enthaltene Kohlen- s~ure bei der p~-Bestimmung entweieht und dadurch die Reaktion nach der alkalischen Seite versehoben wird. Die Befunde yon K~OTH zeigen dieses deutlich; in dem mit der Luft in Berfihrung gekommenen Inhalt waren eben- falls Werte yon p~ 7,46--7,82 festzustellen, in dem mit der Spritze entnom- menen Inhalt dagegen Werte yon p~ 6,78--6,98. Diese dtirften den tats~eh- lichen Reaktionsverh~ltnissen im Pansen am meisten entspreehen. Die Reak- tion im Panseninhalt ist danach schwach-sauer oder neutral.
Im Psalter ist der Inhalt anscheinend schon saurer. ST~I~E~ZE~ land hier im Mittelwert p~ 7,24 und FEa~R p~ 7,0. Im Labmageninhalt sehwanken die p~-Werte zwisehen 2,036 und 4,141 (C. S c ~ w ~ z und I(~A_~).
Die Resorption in den Vormiigen wird in neuester Zeit wieder diskutiert (WEsT~R). Neuerdings ist es Ta~UT~N~ (6) gelungen, experimentell zu zeigen, dass im Pansen, in der ttaube und im Psalter eine Resorption yon Wasser und yon im Wasser gelSsten Stoffen (Atropin, Piloearpin) stattfindet, und zwar scheint hier die Resorption sehneller zu effolgen als im Labmagen. Der Fliissigkeitsdurehtritt durch die stets feuehte Vormagenschleimhaut erfolgt nach Gesetzen der 0smose und ist der Resorption dutch die eutanen Mund- und Konjunktivalschleimh~ute an die Seite zu stellen.
Auch beim Hamster mit dem zweihShligen Magen finder nach den all- gemein bekannten Untersuchungen yon Sc~.u~Euw im cutanen Vor- magen durch die Speichel- und Nahrungsmitteldiastase ein weitgehender Kohlehydratabbau start, der schliesslieh dureh die MiIehs~ureg~rung eine Hem- mung effahrt. Im Dr~isenmagen l~uft die Verdauung in der gleichen Weiso wie bei den anderen Tieren ab.

618 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologic der Verdauung der S/~uge~iere.
D. Die Darmverdauung. I. Morphologisches.
1. Gr~issenverh~iltnisse. ])ber das Verh~ltnis der Gesamtdarmliinge zur K6rperliinge wurde bereits (S. 589) gesprochen und ist aueh schon weitgehend an anderen Stellen erOrtert worden (ScgEu~E~T, H A E S L E R , ELLENBERGER- BAUM). Physiologiseh wichtiger ist die Kenntnis des Verhal~ens der 0berfl~chen der Darmabschnitte zueinander und Zur Hautoberil~ehe. Als Ansgangs- material zu diesen Berechnungen liegen nur die Oberfl~chenzahlen yon COLIN vet. Das Verhaltnis der Hautoberflgche zur D~nndarmober/ldiche betr~gt nach diesen Berechnungen
beim Pferd . . . . . . . . . 1 : 0,79,
beim Rind . . . . . . . . . 1 : 0,965, beim I-Iund . . . . . . . . . 1 : 0,37,
bei der Xgtze . . . . . . . . 1 : 0,34.
Der Untersehied zwisehen den beiden extremen Pflanzenfressern und den beiden Fleisehfressern ist eindeutig; selbst die 0berfl~ehe des eigentliehen Fermentdarmes ist bei den Pflanzenfressern relativ grSsser. Dass der Diek- darm der Pflanzenfresser relativ welt gr6sser ist als bei den Fleisehfressern, ist naeh den bereits in Kapitel ,,Allgemeines zur Magen-Darmverdauung" angegebenen Bereehnungen verst~ndlieh. Aueh beim Rinde, bei dem der Diekdarm im Gegensatz zum Pferde nieht allein die G~rkammer bildet, son- dern nur den kleineren Tell derselben (Vormagen und Diekdarm) darstellt, ist immer noeh die Diekdarmoberfliiehe relativ zur Hautoberfli~ehe grSsser als bei den Fleisehfressern. Hautoberfl~ehe zur Diekdarmoberfl~ehe verhalten sieh n~mlieh
beim Pferd . . . . . . . 1 : 1,30 qm,
beim Rind . . . . . . . 1 : 0,42 q m (1 : 1,8 q m Vormagen und Diokdarm),
beim t I u n d . . . . . . . 1 : 0,08 qm,
bei der Katze . . . . . I : 0,10 qm.
Dasselbe kommt aueh in dem Verh~ltnis der Dtinndarmoberfl~ehe zur Diekdarmoberfl~ehe zum Ausdruek; dieses betragt
beim Pferd . . . . . . 1 : 1,61 qm, beim Rind . . . . . . 1 : 0,44 qm, beim Schwein . . . . . 1 : 0,50 qm, beim I l u n d . . . . . . 1 : 0,22 qm, bei tier Katze . . . . 1 : 0,38 qm.
Diese durehweg eindeutigen Untersehiede zwisehen den beiden Ern~h- rungsgruppen legen die Frage nahe, wie welt die DarmgrSsse dureh eine be- stimmte Ernghrung beeinflusst werden kann. Derartige Versuehe sind neuer- dings an Sehweinen (HAESLER, SCHRSDER)und Ratten (MANGOLD lind HAESLER, WnTZEL) durchgeftihrt worden, und zwar mlt rein vegetabiliseher und volu- minSser und mit animaliseher Nahrung, Beim Sehwein ( H A E S L E R ) , bei dem die Versuehe an neun Ferkeln eines Wurfes bis zur Erreiehung des gleiehen Gewiehtes (etwa 90 kg), durehgef~hrt wurden, zeigte die ~egetariseh erni~hrte Gruppe mit 21,96 m die geringste Darmliinge, die Animalier stehen mit 22,85 m

Die Verdauungss~fte des Dtinndarms. 619
den Normalen mit 23,20 m i m Durchsehnitt nahe. Bet den Animaliern war eine Tendenz zur Verl~ngerung des Dfinndarmes, bet den Vegetariern zur Verl~nge- rung des Diekdarmes gegenfiber den Normalen angedeutet. Im Volumen des Blind- und Diekdarmes waren noeh einige Untersehiede vorhanden; bet den Vegetariern war das Volumen etwas gr/Jsser und bet den Animaliern etwas kleiner Ms bet den Normalen, was den obigen Untersehieden zwisehen Pflanzen- und Fleisehfressern entsprieht. Immerhin ist der Einfluss der Nahrung auf die DarmgrSsse aber nieht derart, dass daraus ffir die Tierernahrung praktisehe Folgerungen gezogen werden kSnnten. Im gleiehen Sinne verliefen die Ver- suehe an weissen Ratten (MANGOLD und HAESLER, WETZEL, KESTNER, ADmS). WETZEn Rand ausserdem noeh, dass das Gewieht der Leber bet den Fleiseh- ratten grSsser ist als bet den Pflanzenratten.
2. Die Darmdriisen. Im Aufbau der Darmschleimhaut nehmen die Submueosa- (BRu~E~sehen) Drfisen des Duodenums eine besondere Stellung ein. Diese Submueosadrfisen sind am starksten bet den Pflanzenfressern ent- wiekelt, bet den Fleisehfressern dagegen gering und bet den Allesfressern mittel- m~ssig; beim Pferde sind sogar noeh im unteren Duodenumabsehnitt BRUNNE~- sehe Drfisen zu linden (VILr&,IIN). Naeh T~VE~ (2) ahneln die BRUNNER- sehen Drfisen nut bet Hund und Katze den Pylorusdrfisen, und zeigen aueh nur hier lJberg~nge zu letzteren. Bet Pferd und Rind z. B. kSnnen sie sehon naeh ihrer grossen Ausbreitung nnd dem Vorkommen yon PA~ET~sehen Zellen und Becherzellen nieht als vorgerfiekte Pylorusdrfisen angesehen werden, sondern ihre tIerkunft muss yore Darmepithel hergeleitet werden. Aueh bet I~aninehen und Feldhasen erwiesen sieh die B~uN~E~sehen und Pylorus- dr/isen genetiseh und anatomiseh als unabh~ngig voneinander (Tsc~Assow- NIKOW). Die Starke der Entwieklung der BRUNNERsehen Dr/Jsen seheint analog der Entwieklung der serSsen Speieheldrfisen mit der Pflanzennahrung zusammenzuh~ngen.
An den Darmzotten weisser M~use sah SuN im Hunger Degenerationen auftreten, die bet erneuter Futteraufnahme versehwinden. Dasselbe beob- aehtete FnAZZETO beim Meersehweinehen.
II. Die Verdauungss~ifte des Diinndarms.
Unsere Kenntnisse fiber die Verdauungss~fte (Pankreassaft, Gallo, Darmsaft) in vergleichender Beziehung sind nicht um~angreich.
Pankreassa[t. Das spezifische Gewieht betr~tgt beim Pferd 1008--1010 (COLIN) und beim Sehaf im Durehschnitt 10062 (N. A. PoPow nnd t~UDRAW- ZEW). Die Reaktion ist stets alkalisch und entspricht einer Alkalit~tt yon 0,28--0,65 % Na2COs. Die Wasserstoffionenkonzentration betr~gt beim Hund p~ 7,3--8,7 (s. BABKIN), beim Schaf p~ 8,12--8,33 [PoPow (2)]. Allerdings seheint die Art der Ern~hrUng auf die Wasserstolfionenkonzentration yon Einfluss zu sein; CZUBAnSKI fand beim Hund naeh Kohlehydraten und Eiweiss

620 W. LENXEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Si~ugetiere.
Werte yon p~ 8,23--8,64, naeh Milchaufnahme Px 7,06--8,45. Dio verdauende Kraft der 3 Fermen{e Trypsin, Ptyalin nnd Lipase zeigt bei den untersuehten Tieren keine wesentliehen Untersehiede.
Die Sekre~ion des Pankreassaftes erfolgt beim Sehaf (N. A. PoPow),
beim Kaninchen (BAxTEr) und hSehstwahrscheinlieh aueh bei Pferd und Rind kontinuierlieh im Gegensatz zu der vortibergehenden Absonderung bei Menseh und Hund. Beim Hunde wurde nur w~hrend des Hungers eine kontinuierliehe Sekretion beobaehtet (BERG). Eine psyehisehe Sekretion des Pankreas seheint naeh den Beobaehtungen yon FONSEC• und TRINCAO beim Mensehen vor- handen zu sein. Sonst erfolgt die Anregung des Pankreas zur T~tigkeit vor- wiegend auf ehemiseh-humoralem Wege. Die vorausgehende reflektorisehe Phase dauert beim Hunde naeh Zufuhr yon Brot und Fleiseh etwa 1/2 Stunde und naeh Milehaufnahme etwas li~nger (DIONESSOV). Bei den Pflanzen- fressern, wie beim Sehaf, steht anseheinend die humorale ganz und gar im Vordergmnd [N. A. Porow (2)]. Beim Sehaf konnte PoPow (1, 2) naeh In- jektion yon S~ure (0,5 % HC1) und naeh Injekr yon Sekretin eine deutliehe Steigerung der Pankreassekretion hervorrufen.
Das Sekretin, der spezffiseh auf die aussere Pankreassekretion wirkende Stoff (BAYLISS-STAnLI~G s. BaBKIN), wird naeh den vergleiehenden Unter- suehungen yon KOSe~TOJA~Z und Mitarbeiter bei allen Wirbeltieren, mit Ausnahme der VSgel; nur in einem bestimmten Darmabsehnitt, im Duodenum, unter Einfluss versehiedener Erreger gebildet. Bei den VSgeln kann das Sekretin aus dem gesamten Dfinndarm extrahiert werden (KoscI~TOJA~Z). Das Sekre~in ist artunspezifiseh. Aueh beim Embryo (S~ugetier) ist das Sekretin nut im Duodenum zu linden, jedoeh erst naeh der endgtiltigen Diffe- renzierung der Duodenalsehleimhaut (KosouTOJANZ). !Jber den Meehanis- mus der Sekretinbildung nimmt RAsE~ow (2) an, dass aus den Eiweiss- spaltungsprodukten des Magensaftes die Darmsehleimhaut das ,, Sekretin" bildet.
Galle. Auf die umfangreiehe Literatur der Chemie der Tiergalle, ins- besondere der Wiederk~uergalle und der Hundegalle hier einzugehen, kann bier nieht die Aufgabe sein, zumal sie aueh an anderen Stellen ausffihrlieh dargestellt ist (s. BABKIN). Nut die Hauptkonstanten sollen hier angeftihrt werden.
Es ist zweekmi~ssiger, nut die Lebergalle zu vergleiehen, da dann aueh die Tiere ohne Gallenblase (Pferd, Hirseh, Kamel) zum Vergleieh herangezogen werden kSnnen. In den meisten FMlen ist dann die Galle dureh Gallenfistel gewonnen. Das spezifisehe Gewieht der Ziegengalle sehwankt zwisehen 1004 his 1010 (ScHo~EGGE) und betr~gt beim Pferd im Durehsehnitt 1005. In der Gallenblase ist das spezffisehe Gewieht natfirlieh etwas hSher (1020 his 1026). Die Reaktion ist vorwiegend sehwaeh alkaliseh. Ffir die Lebergalle des Kaninehens fanden NmLSON und MEY~ Werte yon p~ 7,4--7,7 des Meersehweinehen Werte yon p~ 7,5 und ffir die dutch eine Fistel gewonnene Galle vom Mensehen p~ 8,0; naeh l~ngerem Stehen wird die Galle alkaliseher.

Die Verdauungss~Ite des Diinnd~rms. 621
wahrscheinlich dutch Entweichen yon C02, und erreicht dann pg Werte yon 9,2 beim Kaninchen und 8,6 beim Menschen. Die aktuelle Reaktion der dick- fl~issigen Blasengalle betr~gt beim t(aninchen p~ 7,3 im Durchschnitt, bei Rind, Schaf und Schwein pg 7,0--7,8 (1N-EILSON und MEYEr) ; nach den neueren Untersuchungen yon PoPow, KUDRJAWZEW und UGA~OW liegen die Mittel- werte fiir die Blasengalle beim Rind bei px 7,24 (7,10--7,38), beim Schwein bei p~ 7,00 (6,54--7,17) und beim Schaf bei p~ 6,62 (6,10--7,107). Auch bei Hund, Katze, Affe und Ziegen geht die Reaktion der Blasengalle mehr nach tier sauren Seite fiber als die Lebergalle (I~EILSO~ und MEYEa). Einen wesent- lichen Einfluss auf die Reaktion der Blasengalle hat die Ern~hrung; N~ILsoN und MEYER konnten beim Kaninchen durch ttungern, durch Verabfolgung yon sgurebildender Kost (Hafer)~ die Wasserstoffionenkonzentration steigern (PH 6,4--7,00), dutch Verabfolgung yon basischer Kost (Kohl) erniedrigen (p~ 7fl).
Der Galle der Haustiere kommt nach ELLENBERGER, HOFMEISTE~ auch eine diastatische Wirkung zu. Die lipolytische Kraft der Galle soll im Durch- schnitt bei den Rindern grSsser sein als bei Schaf und Schwein (MGRS~DE).
Die Sekretion der Galle ist durchweg eine kontinuierliche, wghrend des Hungers nur vermindert. Es besteht aber scheinbar der Unterschied, dass bei den Fleischfressern und sicher beim Menschen die Galle zun~tchst teilweise in der Gallenblase gesammelt wird und nut zur Zeit der Verdauuug in grSsseren Mengen in den Darm gclangt, class bei der Ziege aber z. B. die Lebergalle zuweilen mehrere Stunden hindurch direkt in den Darm fliesst (Sc~oREGGE). W~thrend des Hungers kommt allerdings auch bei der Ziege mehr Galle in die Gallenblase als w~hrend der Verdauung. Bei den Tieren ohne Gallenblase (Pferd, Hirsch, Kamel) ist die Zufuhr der Galle nach dem Darm nach den alten Angaben yon CoLI~ yore Pferd wohl weniger schwankend. Der Druck im Lebergallengang (Sekretionsdruck) betrggt bei der Ziege im Durchschnitt 270 mm Galle (ScttoREGG~), bei Meerschweinchen, Hund und Kaninchen 200 mm Wasser. In der Gallenblase steigt bei der Ziege der Druck nicht fiber 110 mm Gallensgule (Sc~o~EG~E). Die Menge der sezernierten Galle betrggt ~fir 24 Stunden ffir die Ziege pro Kilogramm KSrpergewicht 24--30 cem (Sc~oREGOE). Durch intraduodenale Zufuhr yon Galle and aueh yon Salz- s~ure wird die Gallensekretion gesteigert.
Darmsafl. In dem reinen Darmsaft, der dutch eine Darmfistel gewonnen ist, ist das Sekret der L~ESERKt~gNschen Drfisen und der BRUNNE~schen Drfisen enthalten. Das Mengenverh~ltnis der beiden Sekretarten wird bei den einzelnen Tieren entsprechend der verschiedenen Ausdehnung der BR~N- SERschen Drfisen verschieden sein. Nach ASDER~AnD~N und ROSA ist in dem Sekret der B~NSERschen Drfisen ein proteolytisches Ferment vorhanden; beim Schwein hat das Sekret keine proteolytische, sondern eine amylolytische Wirkung (Sc~Eu~RT und GH~E~) . Die endgfiltige Bedeutung des Sekretes der Submncosadrfisen ffir die Verdauung ist his heute noch nicht geklgrt.

622 W. LE~x~I~: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S/~ugetiere.
Naeh den iilteren Angaben yon SOHEUNERT und GRIM~EIa und naeh der beson- ders grossen Ausdehnung bei den Pflanzenfressern seheint Bin Zusammenhang mit der Pflanzennahrung gegeben zu sein. Den Hauptbestandteil bildet bei allen Tieren das Sekret der LIEBEaKi~I~Nsehen Drfisen, dieses enth~tlt die ver- danenden Fermente (Erepsin, Lipase, Maltase, Invertase, Laetase) bei allen Tieren in gleieher Weise. Das Oberfl~ehenepithel sondert als Hauptbestandteil Sehleim ab. Der gemisehte DarmsaR (Sekret der LIEBEnKtmNsehen Drfisen, der BRu~E~sehen Drtisen und des Oberfl~ehenepithels) ist mit Hilfe der TmaY-VELLAsehen Fistel leiehter der Untersuehung zug~nglieh. Das spezi- [ische Gewicht des Darmsaites des Diinndarmes hat beim Hund die GrSsse yon 1010 (ROSE~AN~), beim Schaf yon 10057--10202 [Po~ow (2) u. A~uaow und K~DRJAWCEW, PREGL). Die aktuelle Reat~tion ist alkaliseh; die Wasser- stoffionenkonzentration schwankt beim Sehafe yon p~ 7,69--8,89 (PoPow). Die Alkalit~it entsprieht beim Schaf naeh den iilteren Untersuehungen yon P~EGL einer 0,45%igen LSsung yon Natriumearbonat.
Die Sekretion des DarmsaRes ist eine kontinuierliehe. Sie erfiihrt naeh den Untersuehungen yon PoPow (2), A~v~ow and KUDRJAWCEW am Sehaf dutch den MagensaR, dutch Alkali, dureh S~uren, dutch MgSQ und dureh den Silagesaft eine ganz betri~ehtliehe Steigerung; Wasser und Sojamileh haben nut eine geringe Wirkung. Die angegebenen Sekretmengen sind nieht einwandfrei miteinander zu vergleiehen, da nieht immer die GrSsse des zur Tm~'-VEL~Asehen Fistel benutzten Darmes angegeben ist. Im Durehsehnitt seheint abet die Sekretabgabe bei den Pflanzenfressern grSsser zu sein als beim Fleisehfresser (PaEeL).
Die Sa/tabsonderung des Dickdarmes ist sehr gering; ttEUPKE (2) konnte bei seinen neueren Untersuehungen am Hund und an Kaninehen das Sekret nur dureh Abspfilen mit Wasser gewinnen; aueh dutch Physostigmin, Coffein, Adrenalin und Pituitrin konnte die Sekretmenge nieht vermehrt werden. Die Real~tion ist sowohl beim Kaninehen als auch beim ttunde stets alkalisch. Die Wasserstoffionenkonzentration liegt beim Kaninehen bei pg 7,4--8,4 und beim Hund bei p~ 7,6--8,4, also bei dem Pflanzen- und dem Fleisehfresser in demselben Gebiet. Der Hauptbesgandteil des Sekretes ist Sehleim; reiehlieh vorhanden sind aueh Darmepithelien, Leukocyten und Bakterien. An Fer- menten wurden Diastase, Lactase, Maltase und Lipase gefunden; allerdings sind die fermentativen Wirkungen sehr gering. Die Exkretion yon Mineralien geht deren Zu/uhr mit der Nahrung parallel. Ftir die Verdauung ist der Diek- darmsaR yon geringer Bedeutung.
IIL Meehanik der Darmverdauung. Der Ablaul der Diinndarmbewegungen ist im Prinzip bei allen S~ugetieren
gleich [KRZYWANEK (1, 2), I-IuKuHARA). Nach den umfangreichen Unter- suehungen yon I~[IaZYWANEI~ seheinen beim Meerschweinehen am Dtimldarm

Meohanik der ])armverdauung. 623
die peristaltischen Bewegungen vor den Pendel- und Misehbewegungen vor- zuherrsehen; denn der Transport durch den D/inndarm erfolgt naeh den rSnt- genologischen Beobachtungen besonders bei den erwaehsenen Tieren ziemlieh schnell. ]~eim Hund ist dagegen der Durchgang nur durch den proximalen Dfinndarmabschnitt ein schneller und dutch den distalen Abschnitt verlang- saint bis zum Stillstand im Ileumende; dieses fiihrt zu einer Stauung nnd sch!iesslich zu einer Auftreibung des distalen Ileum bis sehliesslich der Inhalt in den Dickdarm iibertritt. Dieses eigentfimliche Verhalten des Ileumendes konnte bei anderen Tieren (Hamster, Ratte, Meerschweinchen) nicht beob- aehtet werden (KazYwAxEK).
Der Blin&larm wird beim Hund nur in seltenen Fiillen gefiillt [K~zY- WANEK (1)]. Die relativ grossen Blinddttrme der Pflanzenfresser werden bei jedem lJbertritt des Darminhaltes aus dem DOnndarm in den Diekdarm gefifllt, so gelangt z. B. bei Ratte und Meerschweinehen [KRZYWA~EK (2)] hie- reals Dtinndarminhalt direkt ins Colon; beim Hamster ist dieses dagegen m6glieh. Die am Blinddarm ablaufende Bewegung ist vorwiegend peristal- tischer und antiperistaltischer Natur (CzErA und STmL~R, KeZYWA~E~).
Am Dickdarm sind motorisch funktionell zwei verschiedene Abschnitte sieher bei allen Siiugetieren zu unterscheiden, der orale, dutch lebhafte Misch- bcwegungen ausgezeiehnete noch resorbierende Absehnitt und der distale, bewegungstriige, mehr transportierende Abschnitt (BEct(E~ und OPPE~m~aE~). Uber die physiologische Grenze zwischen diesen beiden Dickdarmabschnitten, wie fiberhaupt fiber die Dickdarmbewegungen sind wir bei den Tieren noch wenig unterrichtet. Beim Menschen liegt nach den neueren r6ntgenologischen Untersuchungen yon B E c ~ und OP>EN~tEI~Ea die Grenze in der linken Flexur. Bei den Wiederkiiuern fgllt die Grenze nach CzErA und STmr~m~ mit dem 1Jbergang der Ansa proximalis in die Ansa spiralis (distalis) zusammen. Das Caecum und die Ansa proximalis sahen CzEPA und STm~ER am R{Jntgen- schirm niemals bewegungslos, die Ansa spiralis dagegen ,,immer in Ruhe verharren". Die Art der Dickdarmbewegungen ist bei den Pflanzen- und Fleischfressern die gleiche. Am proximalen Dickdarm laufen peristaltische und antiperistaltische Wellen ab, bei den Wiederk~uern in unregelm~ssiger Folge (CzEPA und STmLE~). Im distalen Dickdarm konnte B~OKER und O>PE~I~EI~R beim Mensehen nieht mit Sicherheit Peristaltik feststellen. Der distale Abschnitt ist bei den Pflanzenfressern gegentiber dem proximalen Abschnitt relativ liinger als beim Fleischfresser.
Die Grenze zwisehen diesen beiden funktionell verschiedenen Abschnitten ist aueh als die Stelle anzusehen, an der die Kotbeeren der Pflanzenfresser dureh segmentierte tonisehe Absehnfirungen entstehen. Beim Sehaf ist es da- naeh der Ubergang der Ansa proximalis in die Ansa spiralis (CzEPa und STmT,E~). Diese Stelle ist naeh Beobaehtung an Sehafen mit DarmstSrung wahrseheinlieh eher zum distalen Abschnitt als zum proximalen zu rechnen [LE~mT (2)].

624 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~ugetiere.
Bei anderen Tieren hat KRZYWANEK (9,) diese Stelle dureh rSntgenologisehe Beobaehtungen genauer fixiert. Bei der Ratte finder die Kotiormung am Ubergang des Colon ascendens in das Colon transversum statt, beim Meer- sehweinehen am Ende der Doppelspirale des Colon, beim Hamster in der ersten eaudalen Sehleife. Darnaeh diirfte man aueh bei diesen Tieren annehmen, dass nur bis zur Stelle der Kotformung der intensiv bewegliehe und noeh resorbierende Diekdarmabsehnitt reieht, zumal J~RzYwANEK auch bei der Ratte im distalen Tell niemals Antiperistaltik beobaehtet hat.
Vol]kommen ungeklart ist noeh die Frage, ob diese iunktionell versehie- denen Absehnitte tiberall aueh versehieden innerviert sind, ob etwa der Vagus nur den proximalen Tell bis zur funktionellen Grenze und der Pelvieus den distalen Tefl versorgt.
IV. Ablauf der Darmverdauung. Die fermentative Verdauung im Diinndarm wird zwar im Prinzip bei
allen Tieren die gleiche sein (s. SCHEUNERT, BIEDERMANN); aber lfir die pflanz- ]iehe Nahrung war his vor kurzem noch unklar, ob wirklieh naeh der alten Annahme eine Verdauung der Niihrstoffe der Pflanze (GetreidekSrner) erst naeh ZerstSrung der Cellulosehfille mSglieh ist. Bei der Pansenverdauung ist bereits darauf hingewiesen. Vergleiehende Ausnutzungsversuche z. B. mit ganzen HaferkSrnern und mit gesehrotenen HaferkSrnern lassen jedoeh diese Annahme nieht als allgemeingfiltig erseheinen. HO~CAMP, Se~RAM~ und STOTZ fanden bei derartigen Versuehen an Sehafen im Durehsehnitt keinen praktisehen Untersehied zwisehen der Verdauung der ganzen HaferkSrner und des Hafers in grobgesehrotener Form. Aueh beim Pferd, bei dem im Kot unversehrte Haferk6rner in nieht geringer Menge wiedererseheinen, ergab die mikrosko- pisehe Untersuehung derartiger KSrner z. B. deutliehe Verdauungsvorggnge an der Aleuronsehieht, was weiter dureh den Verlust tier Keimf~higkeit bewiesen wurde (FRISCItBIER). Der Menseh verwertet die Nghrstoffe gewisser Pflanzen ebenfMls gut, ohne dass die Zellen erSffnet sind [HEUPKE (1)]. Nach diesen vor- liegenden Untersuehungen kSnnen die Fermente in die unerSffneten Zellen eindringen und den Inhalt herausverdauen. Es seheint aber Bin bestimmter Quellungszustand der Cellulosemembran fiir das Eindringen der Fermente notwendig zu sein, der naeh l~ngerem Aufenthalt, besonders im Magen, sieher erreieht wird, denn in vitro durehgefiahrte Verdauungsversuehe mit ganzen Haferk6rnern verliefen negativ (FRISCItBIER).
Im Diekdarm stehen bei den Pflanzen- und Allesfressern die bakteriellen Zersetzungen der Cellulose im Vordergrund." Wie im Pansen geht hier, beson- ders im Blinddarm, die Zersetzung der Rohfaser bis zu den Fetts~uren (To,AN, USTJANZEW und ZU~TZ). Aueh die hier reiehlieh vorkommenden Infusorien (ST~ELKOW) spielen anseheinend bei den im:Diekdarm ablaufenden Prozessen wie im Pansen eine Rolle (BIENERT, TANZEtt, ERBEN).

Ablauf der D a r m v e r d a u u n g . 625
Die Reaktionsverh~iltnisse des Darminhaltes sind auch bei den Tieren mit gleicher Ern~hrung etwas versehieden. Im oberen Diinndarm ist die Reaktion fast iiberall sehwach-sauer und dann sinkt nach dem Diekdarm zu die Wasser- stoffionenkonzentration, ohne aber mit wenigen Ausnahmen welt fiber die Konzentration yon ph 7 zu gehen (KosTYA~, Tabelle 5).
Tabelle 5. W a s s e r s t o f f i o n e n k o n z e n t r a t i o n d e s D a r m i n h a l t e s .
Tier
H u n d . . .
Katze . . .
K a n i n c h e n .
Rathe . . .
R ind . . .
Pferd . . .
D u o d e n u m
5,91 6,51
7,35
6,5
6,678
6,722
J e j u n u m
5,99--6,27
6,905
8,416
I l eum
6,36
6,79
7,99 7,2
8,208
7,09
Coeeum
6,57
6,26
5,5--7,2
8,224
8 , 1 2 5
Colon
6,84 5,25
5,6--7,1
Au~or
GI%AYCEL u n d IV[ILLER M c L A N G ~ r s
P~ SUIt', ]~LUMENTHAL, SLI-
~El% HERBER U. WANG DAN~TINGER, I)FRAGNER
u n d SCH~LT~S
F fir die Verdauungsenzyme, besonders fiir das Trypsin, ist im Darm keineswegs fiberall (Tabelle 5) das p~-0ptimum, denn das p~-0pt imum f(ir Trypsin z. B. liegt bei p~ 7--8 (VoNK und WOLVEKA~P). Die gallensauren Salze wirken aber in solehen F~illen naeh den Untersuehungen yon VONK und WOLVEKAMP als kompensierendes Moment ffir die ungiinstige Wasserstoff- ionenkonzentration und ermSglichen so die tryptische Verdauung. Im alkalischen Milieu hemmen dagegen die gallensauren Salze die enzymatisehe Wirkung. Danaeh mfisste beim Rinde mit der alkalischen Reaktion im Dfinn- darm die feinere Verdauung etwas anders verlaufen, und auch z. B. der Ablauf der Fettresorption (VERzi~), die gerade im Jejunum und Ileum (s. Tabelle) am lebhaftesten ist (W~.~DT), ein anderer sein wie bei den anderen Tieren. Eine eingehende Naehprfifung der vorliegenden Ergebnisse unter weitgehender Ber~ieksichtigung der Zus~mmenh~inge mit der Verdauung und Resorption w~ire notwendig.
Bei den t~esorptionsvorg'~ngen sind auch die Zottenbewegungen yon Bedeutung; und zwar haben diese wohl weniger aufsaugende Wirkung als vielmehr einen beschleunigenden Einfluss auf die Entleerung des Zotten- lymphramnes (VERzAR). Es sind zwei verschiedene Meehanismen der Ent- leerung des Zottenlymphraumes naeh den Untersuchungen yon v. KoKs zu unterscheiden. Die langen, /inger/Srmigen Zotten des tIundes, der Katze, des Fuehses entleeren den Chylusraum dutch Verkfirzung, der die VerI~ingerung folgt. Die platten, blattfSrmigen Zotten, die bei Schwein, Kaninchen, Ratte, Maus, Ziesel zu finden sind, pressen den Lymphraum nieht durch Verkfirzung, sondern dutch Kontraktionen der Museularis mucosae in der Querrichtung aus. Auch beim Maulwurf, d e r n u r Falten besitzt, sind Kontraktionen der Mnscularis nmcosa vorhanden. Beim Igel sind beide Zottenarten vorhanden und dementsprechend auch beide Entleerungsmechanismen zu beobachten
Asher-Spiro, Ergebnisse der Physiologie un4 expcr. Pharmakologie. 35. 40

626 W. LENKEIT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der S~ugetiere.
(v. Ko~As). Als alas Zentrum tier Zottenbewegungen wurde der Plexus sub- mueosae Meissneri gefunden (v. Ko~As und v. L v ~ Y ) .
E. Die Aufenthaltsdauer der Nahrung im Verdauungskanal und der quantitative Verlauf der Ausscheidung.
Ftir die Enfleerungszeiten des Futters aus den einzelnen Absehnitten des Verdauungskanals sind nattirlich die Tiere, bei denen der Magen zwischen den Mahlzeiten leer wird, die Fleisehfresser, yon den Pflanzenfressern zu unter- scheiden, bei denen der Magen bei der tibliehen Mahlzeitfolge niemals leer wird. Die Entleerungszeit des Magens wird bei den Pflanzenfressern aus- gedehnter sein als bei den Fleisehfressern. Beim Fleisehfresser (K~zYWANnK) beginnt die Entleerung selten vor 1/2 Stunde, ist am gr6ssten nach 3--4 Stunden (HEILE) und ist im Durchsehnitt nach 6 Stunden beendet. Bei der Ratte und der Maus liegt der Beginn des Austrittes einer KOrnermahlzeit zwisehen 1/4 und 1/2 Stunde [KRzYWANEK, LENXEIT (3)] und das Ende bei der 1%atte naeh un- gef~hr 7 Stunden, bei der Maus nach ungefKhr 6--7 Stunden (LENKEIT). Beim Meerschweinehen ist der ~bertr i t t in den Darm nach 1/2--1 Stunde zu beob- aehten; naeh 7 bis 8 Stunden sind nut noeh Spuren oder niehts mehr yon der Probemahlzeit im Magen vorhanden [LE~KEIT (3)]. Beim Kaninehen hat eine Mahlzeit den Magen naeh ungefi~hr 9 Stunden verlassen (HE~nICHS). Aueh beim Pferd und beim Sehwein erfolgt die Magenentleerung naeh den bekannten i~lteren Untersuehungen aus dem ELLENBERQE~sehen Institut reeht langsam (s. SCHEUNEaT). Dureh Bewegung wird die Magenentleerung gehemmt (SC~E~:N~aT, TA~, STA~BKE, BIEST~R).
Aus dem Pansen der Wiederk~uer kann das aufgenommene Futter, wenn es hinreiehend zerkleinert ist, sofort austreten; abet deutlieh merkbar ist die Entleerung einer Mahlzeit aus dem Pansen erst naeh 4--5 Stunden (CoLv~BVS). Nach durchsehnittlieh 8 Tagen ist yon einer K/Srnermahlzeit niehts mehr im Pansen zu finden; rein gesehrotener Hafer kann den Pansen sehon nach 6 Tagen endgfiltig verlassen haben [LEN~EIT (5), USUnLLI]. Dureh sehleehte Rauhfutteraufnahme, wie bei Itungertieren, kann die Aufenthalts- dauer des Futters im Pansen wesentlich verli~ngert werden (LE~(EIT, COLUMBUS).
l~ber die Durchgangszeiten dutch den gesamten Verdauungskanal liegen eine Reihe neuerer Untersuehungen vor (Ln~EIT, Usvnn~, B~ns~E~). Und zwar ist hier nieht allein, wie in i~lteren Untersuchungen, der Beginn und das Ende, sondern aueh der quantitative Verlauf der Ausscheidung der Futter- reste festgestellt worden. Es ist zweekmi~ssiger und aueh einwandfreier, den Verlau~ der Ausseheidung stets vom Beginn der Ausseheidung an, zu rechnen, und nieht yore Zeitpunkt der Ftitterung. In den meisten Versuehen ist mit Fuehsindiamant stabil gefi~rbtes Versuchsfutter (LENKE~ und H~B~C~) benutzt worden. Der Beginn, das Maximum und das Ende der Ausscheidung
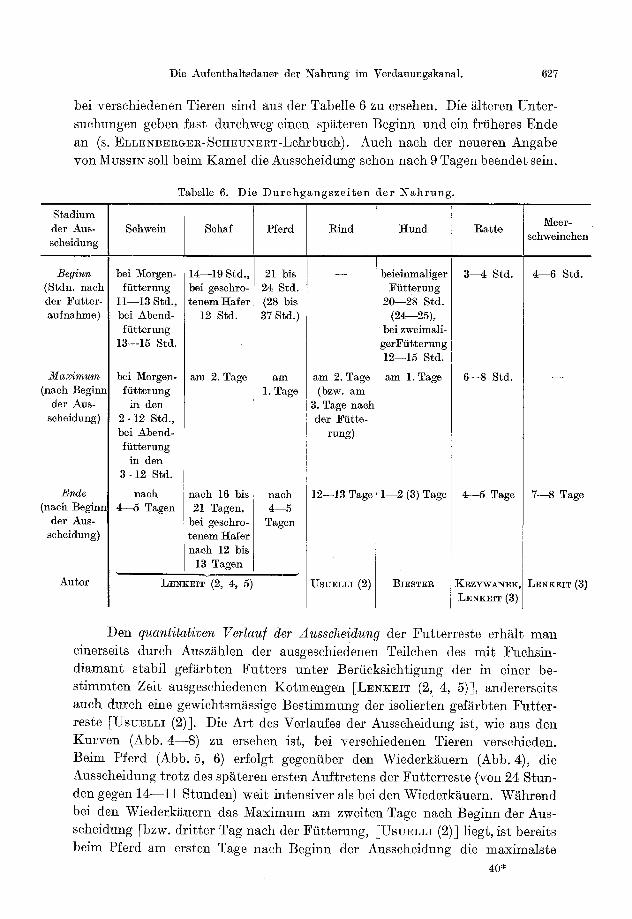
Die Aufenthaltsdauer der Nahrung im VerdauungskanM. 627
bei verschiedenen Tieren sind aus der Tabelle 6 zu ersehen. Die i~lteren Unter- suehungen geben fast durehweg einen sp~teren Beginn und ein fftiheres Ende an (s. ELLnNBE~GE~-SCHEUNERT-Lehrbueh). Auch naeh der neueren Angabe yon MussIN soll beim Kamel die Ausseheidung sehon naeh 9 Tagen beendet sein.
Tabelle 6. Die D u r c h g a n g s z e i t e n d e r N a h r u n g .
Stadium der Aus- scheidung
Beginn (Stdn. nach der Futter- aufnahme)
Ma ximum (nach Beginn
der Aus- scheidung)
Ends (nachBeginn
der Aus- scheidung)
Autor
Schwein
bei Morgen- fiitterung
11--13 Std., bei Abend- fiitterung
13--15 Std.
bei Morgen- ffitterung
in den 2 .12 Std., bei Abend- ffitterung
in den 3-12 Std.
naoh 4--5 Tagen
Schaf
14--19 Std., bei geschro- teneml:fafer
12 Std.
am 2. Tage
naoh 16 bis 21 Tagen,
bei geschro- tenem I~a~fer nach 12 bis
13 Tagen
LENKEIT~2, 4, 5)
Pferd
21 bis 24 Std. (28 bis 37 Std.)
&m
1. Tage
n~oh 4--5
Tagen
Rind
am 2. Tage (bzw. am
3. Tage nach der Ffitte-
! rung)
1 12--13 Tage
U s ~ LI (2)
l-Iund
beieinmaliger Fiitterung I
20--28 Std. ]
(2~-25), I bei zweimali-
gerFtitterung 12--15 Std.
am 1. Tage
1--2 (3) Tage
BIESTER
Ratte
3--4 Std.
6--8 Std.
4 - - 5 Tage
K~ZYWANEK, LE~XEIT (3)
Meer- schweinchen
4--6 Std.
7--8 Tage
LE~K~Ir (3)
Den quantitativen Verlauf der Ausscheidung der Futterreste erh~tlt man einerseits durch Auszi~h]en der ausgeschiedenen Teilchen des mit Fuchsin- diamant stabfl gefarbten Futters unter Berficksichtigung der in einer be- stimmten Zeit ausgesehiedenen Kotmengen [LENKEIT (9,, 4, 5)], andererseits auch dutch eine gewichtsm~ssige Bestimmung der isolierten gef~rbten Futter- reste [Us~JELLI (9,)]. Die Art des Verlaufes der Ausseheidung ist, wie aus den Kurven (Abb. 4--8) zu ersehen ist, bei verschiedenen Tieren verschieden. Beim Pferd (Abb. 5, 6) erfolgt gegenfiber den Wiederk~tuern (Abb. 4), die Ausscheidung trotz des spateren ersten Auftretens der Futterreste (yon 24 Stun- den gegen 14--11 Stunden) weir intensiver als bei den Wiederkauern. Wahrend bei den Wiederk~uern das Maximum am zweiten Tage naeh Beginn der Aus- scheidung [bzw. drifter Tag nach der Ffitterung, [UsuELLI (2)] liegt, ist bereits beim Pferd am ersten Tage nach Beginn der Ausscheidung die maximalste
40*

628 W. L~KEzm: Neuere Ergebnisse der verg]. Phys[ologie der Verdauung der S~uge~iere.
Ausscheidung, und ausserdem ist das Maxhm~m behl~ Fferd nooh hSher als bei den Wiederk~tuern. Dieses spricht dafiir, dass beim Plerd eine auf-
%
,, " . . . ............... 3.c !~
s
--."~ .~.~_..__:----- --,
7~ N del'XsussChelYUi'lj Abb. 4. Sehaf. KOrnerfutter. Verlauf der Ausscheidungszahl der Futterreste in Prozenten tier
ausgeschiedenen Gesamtmenge. (Nach LENKEIT.)
. ~ y _ _
\
I \
\
\ \
\ Z 3. q. s T~q der4usscim/u'ung
Abb. 5. P~ercL H~fer ganz. Verlauf tier Ausscheidung. (Nach LE~KEIT.)
%
I
l/ersuch 2,~) - . ,Zu.~. CaOOg
\
3. ~. ~? 7kj7 #er Ausx~e/~'un~7
Abb. 6. Pferd. l:Iaier geschroten. (Nach LEI,'gEIT.)
genommene Mahlzeit mit den vorangegangenen und mit den folgenden Mahl- zeiten wenig durchmiseht wird, was ja auch im Magen der Fall ist (s. S. 598). Beim tIunde, bei dem die Gesamtausseheidung hSehstens 2 Tage, selten
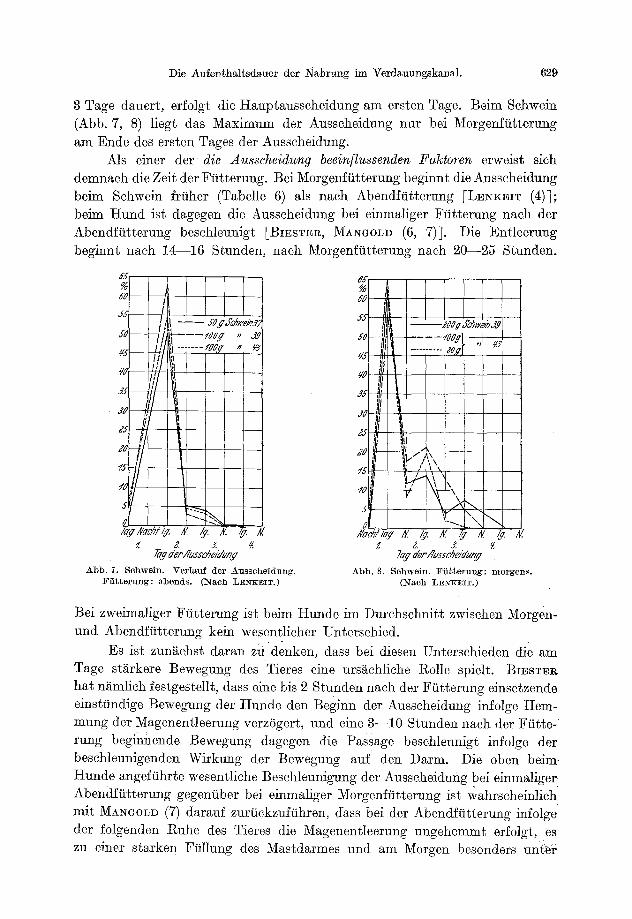
Die Aufenthaltsda.uer der Nalorung im Verdauungskana]. 629
3 Tage dauert, eriolgt die tIauptausscheidung am ersten Tage. Beim Sehwein (Abb. 7, 8) liegt das Maximum der Ausseheidung nur bei Morgenftitterung am Ende des ersten Tages der Ausscheidung.
Als einer der die Ausscheidung beein/lussenden Faktoren erweist sieh demnach die Zeit der Ftitterung. Bei Morgenffitterung beginnt die Ausseheidung beim Sehwein frtiher (Tabelle 6) als naeh Abendftitterung [LENKEIT ( 4 ) ] ,
beim Hund ist dagegen die Ausscheidung bei einmaliger Ftitterung naeh der Abendifitterung besehleunigt [BIESTER, MANGOLD (6, 7)]. Die Entleerung beginnt naoh 14--16 Stunden, naeh Morgenfiitterung naeh 20--25 Stunden.
$5 % /
- D E L
--/I Y3
-I Y
o ~g AJachl ~ ~ ~ i
4, 2 3. 4. 7~j der iTus s ck ei gcn y
Abb. 7. Schwein. Verlauf tier A1]sscheidung. Fiitterung: abends. (Nach LENKEIT.)
8Y
Y"'- d ~ 2 o o y , . ; ' ~ # , ~ z Y~
�9 " ' . . . . . . . . . . #U~J ~/J i i /x 0 ,,!
,]5 !g
30 II
z s ~ zoO- :': % 15 I'~ Io ~ \k
X 2. 3. g 7aJ #gf tTalssCkel~tliz/A7
Abb. 8. Sohwein. Fiigterung- morgens. (Naoh LE~<K~IT.)
Bei zweimMiger Ffitterung ist beim IIunde im Durchschnitt zwischen Morgen- und Abendfiitterung kein wesentlieher Unterschied.
Es ist zun~chst daran zu donken, dass bei diesen Unterschieden die am Tage stiirkere Bewegung des Tieres eine ursiiehliehe Rolle spielt. B~ESTER hat n~tmlieh festgestellt, dass eine his 2 Stunden nach der Fiitterung einsetzende einsttindige Bewegung der Hunde den Beginn der Ausseheidung in~olge ttem- mung der Magenentleerung verzSgert, und eine 3--10 Stunden naeh der Fiitte- rung beginnende Bewegung dagegen die Passage beschleunigt infolge der beschleunigenden Wirkung der Bewegung auf den Darm. Die oben beim: Hunde angeftihrte wesentliehe Besehleunigung der Ausscheidung bei einmalige~ Abendfiitterung gegentiber bei einmaliger Morgenfiitterung ist Wahrschein]ich mit MANGOLD (7) darauf zurtickzuftihren, class bei der Abendftitterung infolge: der folgenden Ruhe des Tieres die Magenentleerung ungehemrat edolgt, es zu einer starken Ftillung des Mastdarmes und am Morgen besonders un%r
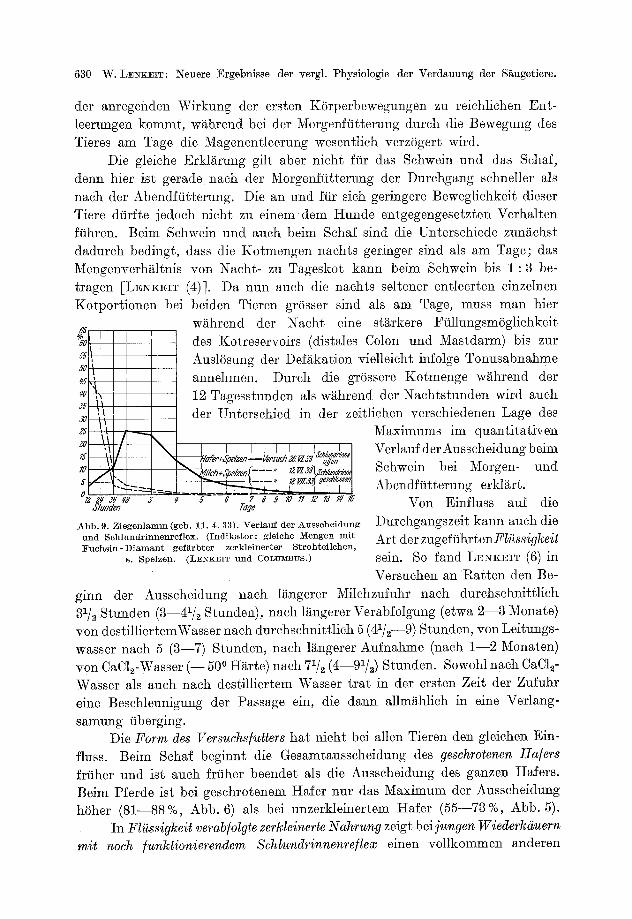
630 W. LENKI~IT: Neuere Ergebnisse der vergl. Physiologie der Verdauung der Saugetiere.
der anregenden Wirkung der ersten KSrperbewegungen zu reiehliehen Ent- leerungen k0mmt, wiihrend bei der Morgenftitterung dureh die Bewegung des Tieres am Tage die Magenentleerung Wesentlieh verzSgert wird.
Die gleiehe Erkl~irung gil~ aber nieh~ fiir das Sehwein und das Sehaf, denn bier ist gerade naeh der Morgenftitterung der Durehgang sehneller als naeh der Abendftit~erung. Die an und for sieh geringere Bewegliehkeit dieser Tiere dOrfte jedoeh nieht zu einem d e m Hunde entgegengesetzten Verhalten fOhren. Beim Sehwein und aueh beim Sehai sind die Unterschiede zun~ehst dadureh beding~, dass die Kotmengen naehts geringer sind als am Tage; das Mengenverh~ltnis yon Naeht- zu Tageskot kann beim Sehwein bis 1 : 3 be- tragen [LENKEIT (4)]. Da nun aueh die naehts seltener entleerten einzelnen Kotpor~ionen bei beiden Tieren grSsser sind als am Tage, muss man hier
wahrend der Nach~ eine starkere Ftillungsm6gliehkeit }o! des Ko~reservoirs (distales Colon und Mastdarm) bis zur ss! AuslSsung der Def~ikation vielleieht infolge Tonusabnahme se ~s annehmen. Dureh die gr6ssere Kotmenge w~ihrend tier ~a 12 Tagesstunden als w~hrend der Naehtstunden wird aueh ~s der Untersehied in der zeitl~ehen versehiedenen Lage des
~ I I t I I t r r ' I I I ~ : [ - - - - - , 18 ~..33j .r176
,5'/unden Tage
Abb. 9. Ziegenlamm (geb. 11. t . 33). Verl~iE der Ausseheidung und Sehlnndrinnenreflex. ( Ind ik~ tor : g]eiehe ~[engen m i t F u c h s i n - D i a m a n t geff~rbter ze rk le iner te r S t rohte i lchen,
s. Spelzen. (LENKEIT u n d COLUMBUS.)
Maximums im quantitativen Verlauf der Ausseheidung beim Sehwein bei Morgen- und Abendftitterung erkl~rt.
Von Einfluss auf die Durehgangszeit kann aueh die Art der zugeftihrten Fli~ssggkeit sein. So fand LENKEIT (6) in Versuehen an Ra t t en den Be-
ginn der Ausseheidung naeh l~ingerer Milchzufuhr naeh durchsehnittlieh 31/2 Stunden (3--45/2 Stunden), nach l~tngerer Verabfolgung (etwa 2--3 Monate) yon des%illiertemWasser naeh durehsehnit%lich 5 (41/2--9) Stunden, yon Leitungs- wasser naeh 5 (3--7) Stunden, naeh langerer Aufnahme (nach 1--2 Monaten) yon CaC12-Wasser ( - - 500 H~irte) naeh 75/2 (4--95/2) Stunden. SowohI naeh CaC12- Wasser als auch naeh destilliertem Wasser %rat in der ers%en Zeit der Zufuhr eine Besehleunigung der Passage ein, die dann allms in eine Verlang-
samung Oberging. Die Form des Versuchs/utters ha% nicht bei allen Tieren den gleiehen Ein-
fluss. Beim Schaf beginnt die Gesamtausseheidung des geschrotenen Ha/ers frfiher und ist auch frfiher beendet als die Ausseheidung des ganzen Itafers. Beim Pferde is% bei gesehrotenem Hafer nur das Maximum der Ausseheidung h0her (81--88%, Abb. 6) als bei unzerkleinertem IIafer (55--73%, Abb. 5).
In Flgssigkeit verab/olgte zerkleinerte Nahrung zeigt bei ]ungen Wiederkguern mit noch /unktionierendem Schlundrinnenreflex einen vollkommen anderen

Die Aufenthaltsdauer der Natu'ung im Verdauungskanal. 631
Verlauf der Ausscheidung, als er bei Trockeniutter (Abb. 9) und bei er- waehsenen Wiederkguern (Abb. 4) gefunden wurde. Wghrend bei Troeken- futter aueh bei jungen Wiederk~nern das Maximum naeh 48 Stnnden (Abb. 9) zu finden ist, liegt dagegen naeh Anfnahme yon Milch mit gefgrbten Futter- teilchen bei jungen Wiederkguern das Maximum sehon in den ersten 12 Stunden (Abb. 9 LENI<EIT U. COLU~BT:S). Dieser Untersehied ist stets so eindeutig, dass die Untersuehung des Verlaufes der Ausscheidung znr Prfifung des Schlund- rinnenreflexes benntzt werden kann.
Durch ungeniigende Futteraufnahme, bei den Wiederk~uern und Pferd allein durch mangelhafte Rauhiutteraufnahme, kann die endgfiltige Ent- leerung betr~chtlich verzSgert werden [ELLENBERGEa, KnZYWAN~K (2), LENEmr (2)1.





























