Embed Size (px)
Citation preview

Neural coding underlying the cue preference forcelestial orientationBasil el Jundia,1, Eric J. Warranta, Marcus J. Byrneb, Lana Khaldya, Emily Bairda, Jochen Smolkaa, and Marie Dackea,b
aDepartment of Biology, Lund University, 223 62 Lund, Sweden; and bSchool of Animal, Plant, and Environmental Sciences, University of the Witwatersrand,Wits 2050, South Africa
Edited by John G. Hildebrand, University of Arizona, Tucson, AZ, and approved July 21, 2015 (received for review January 21, 2015)
Diurnal and nocturnal African dung beetles use celestial cues, suchas the sun, the moon, and the polarization pattern, to roll dungballs along straight paths across the savanna. Although nocturnalbeetles move in the same manner through the same environmentas their diurnal relatives, they do so when light conditions are atleast 1 million-fold dimmer. Here, we show, for the first time toour knowledge, that the celestial cue preference differs betweennocturnal and diurnal beetles in a manner that reflects theircontrasting visual ecologies. We also demonstrate how these cuepreferences are reflected in the activity of compass neurons in thebrain. At night, polarized skylight is the dominant orientation cuefor nocturnal beetles. However, if we coerce them to roll duringthe day, they instead use a celestial body (the sun) as their primaryorientation cue. Diurnal beetles, however, persist in using acelestial body for their compass, day or night. Compass neuronsin the central complex of diurnal beetles are tuned only to thesun, whereas the same neurons in the nocturnal species switchexclusively to polarized light at lunar light intensities. Thus, theseneurons encode the preferences for particular celestial cues and altertheir weighting according to ambient light conditions. This flexibleencoding of celestial cue preferences relative to the prevailing visualscenery provides a simple, yet effective, mechanism for enablingvisual orientation at any light intensity.
navigation | insect | vision | central complex | dim light
The blue sky is a rich source of visual cues that are used bymany animals during orientation or navigation (1, 2). Besides
the sun, celestial phenomena, such as the skylight intensity gra-dient or the more complex polarization pattern, can serve as ref-erences for spatial orientation (3–5). Polarized skylight is generatedby scattered sunlight in the atmosphere, and to a terrestrial ob-server, the resulting alignment of the electric field vectors extendsacross the entire sky, forming concentric circles around the positionof the sun (Fig. 1A). A similar distribution of brightness and po-larization pattern is also created around the moon (6). Althoughthis nocturnal pattern is 1 million-fold dimmer than the daylightpattern (6), some animals, such as South African ball-rolling dungbeetles, can use this lunar polarization pattern for orientation (7).To avoid competition for food at the dung pile, these beetles detacha piece of dung, shape it into a ball, and roll it away along a straight-line path. For this type of straight-line orientation, nocturnal beetlesseem to rely exclusively on celestial cues (8), such as the moon orpolarized light.As with all nocturnal animals, night-active beetles have to
overcome a major challenge: They need to maintain high ori-entation precision even under extremely dim light conditions.Indeed, recent experiments have shown that nocturnal dungbeetles orient at night with the same precision as their diurnalrelatives during the day (9), an ability partly due to the fact thattheir eyes are considerably more sensitive than the eyes of spe-cies that are active at brighter light levels (10–12). Nonetheless,for each species, orientation precision relies on being tuned tothe most reliable celestial compass cue that is available duringthe animal’s normal activity window. How salient are these cuesfor nocturnal and diurnal species? Do diurnal species have a
different celestial cue preference than nocturnal species? If so,how are these preferences represented neurally in the brain?In this study, we present a detailed picture of how the orientation
systems of two closely related nocturnal and diurnal animals havebeen adapted to the ambient light conditions, combining behavioralexperiments from the field with electrophysiological investigations ofthe underlying neural networks. Using behavioral experiments, weshow that nocturnal dung beetles switch from a compass that uses adiscrete celestial body (the sun) during the day to a celestial polari-zation compass for dim light orientation at night, whereas diurnalbeetles use a celestial body (the sun or moon) for orientation at alllight levels. In a second step, we simulated these skylight cues (thesun or moon and the polarization pattern) while electrophysiologi-cally recording responses from neurons in the dung beetle’s centralcomplex, a brain area that has been suggested to house the internalcompass for celestial orientation (13, 14). These neural data preciselymatched the cue preferences observed in behavioral field trials andshow how an animal’s visual ecology influences the neural activity ofits sky compass neurons. Our results also reveal, for the first time toour knowledge, how a weighting of celestial orientation cues could beneurally encoded in an animal brain.
Results and DiscussionNocturnal Dung Beetles Use the Polarization Pattern as a PrimaryOrientation Cue at Night. Dung beetles use a large repertoire ofcelestial cues for orientation (5, 7, 15–17), but their two maincompass cues are (i) the sun or moon and (ii) the celestial po-larization pattern (5). Here, we first sought to test if one compasscue is dominant over the other in the orientation systems of twoclosely related dung beetle species that are active at differenttimes of the day: Scarabaeus lamarcki (diurnal) and Scarabaeussatyrus (nocturnal). The robust straight-line orientation behavior of
Significance
Many animals use the sun or moon and the polarization pat-tern for navigation. We combined behavioral experiments withphysiological measurements of brain activity to reveal whichcelestial cue dominates the orientation compass of diurnal andnocturnal dung beetles. The preference found behaviorallyprecisely matches the preference encoded neurally and showshow the brain dynamically controls the cue preference fororientation at different levels: The sun or moon always domi-nates the orientation behavior and neural tuning of diurnalbeetles, whereas in nocturnal beetles, celestial bodies domi-nate tuning only in bright light, with a switch to polarized lightat night. This flexible neural tuning in the nocturnal speciesprovides a simple mechanism that allows it to use the mostreliable available orientation cue.
Author contributions: B.e.J., E.J.W., M.J.B., E.B., J.S., and M.D. designed research; B.e.J.,M.J.B., L.K., E.B., and M.D. performed research; B.e.J. and J.S. analyzed data; and B.e.J.wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1501272112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1501272112 PNAS | September 8, 2015 | vol. 112 | no. 36 | 11395–11400
PHYS
IOLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020

these beetles can be induced independent of the time of day,allowing us to examine the beetles’ choice of celestial cues even attimes when they are not usually active (i.e., the nocturnal speciesduring the day, the diurnal species at night). First, we measured thechange in the beetles’ directions between two consecutive rolls whenthey rolled their balls out of an arena (1-m diameter) on a sunlit dayor under a clear, moonlit sky (Fig. 1 B and C; control,●). We testedthe beetles’ orientation behavior at low sun elevations or (on full-moon nights) moon elevations (<30°). Under these conditions, acelestial body (sun or moon) and the celestial polarization patterncreated by it are both clearly visible to the animals (Fig. 1A). In allcases, the change of direction between the two repetitions wasclustered around 0° (P < 0.001 by V test, with an expected mean of0°); that is, the beetles did not change their bearing between con-secutive rolls (Fig. 1 and Figs. S1A and S2 A–D). This observationconfirmed that both species have a consistent orientation behavioreven at times when they are not naturally active.To investigate the relative importance of, or preference for,
the two main celestial cues in the beetles’ compass system (sun ormoon vs. polarized light), we also examined how beetles reactedwhen the two cues were set in conflict. Again, beetles were madeto roll out from the center of the arena on two consecutive oc-casions: once in the light of the sun or the moon and once withthe sun or full moon (celestial body elevations <30°) “displaced”to the opposite sky hemisphere using a mirror (at the start ofeither the first roll or the second roll), while simultaneouslyhiding the real sun or moon from the beetle’s view (Fig. 1 B andC; test, ○). If the beetle relies on polarized skylight as its maincue, it should adhere to its original rolling direction, which would
still be visible (and unaltered) across the remainder of the visiblesky. In contrast, if the beetle uses a celestial body (sun or moon)as its primary reference for orientation, it should use the reflectedmoon or sun as a directional cue and turn by about 180° withrespect to its original rolling direction. We found that the resultdepended on which species was tested. Diurnal beetles followed thereflected sun or moon (i.e., they changed their rolling directionsignificantly toward the opposite sky hemisphere), both during theday (P < 0.001 by V test, with an expected mean of 180°) and atnight (P = 0.002 by V test, with an expected mean of 180°) (Fig. 1 Band C, Left and Figs. S1A and S2 A and C). This result matches ourprevious findings (5, 17, 18) and suggests that diurnal dung beetlesrely primarily on a celestial body for orientation; that is, they useeither the sun or the moon as a compass. This cue preference wasalso true for the nocturnal species when orienting during the day[Fig. 1B, Right (P < 0.001 by V test, with an expected mean of 180°)and Figs. S1A and S2B]. However, when these nocturnal foragersrolled their balls in the light of the full moon, they displayed adifferent strategy: The beetles did not change their rolling direction(P = 0.996 by V test, with an expected mean of 180°), but keptrolling in their original direction of travel even when the moonwas reflected back at them from the opposite direction [Fig. 1C,Right (P < 0.001 by V test, with an expected mean of 0°) and Figs.S1A and S2D]. In contrast to the diurnal species, the nocturnalspecies thus primarily followed the direction given by the skylightpolarization pattern under dim light conditions. To confirm thisobservation, we also tested the nocturnal species rolling under apolarizing filter with a clear view of the full moon (elevation <30°)through the polarizer. The polarizer was placed on top of ascreened arena, with the filter’s E-vectors either aligned (Fig. S3A)or in conflict (Fig. S3B) with the lunar polarized skylight. In contrastto the diurnal species, which orients with respect to the sun even ifthe polarizer is turned by 90° (5), the nocturnal species significantlychanged its bearing in response to a change in the polarized lightdirection (Fig. S3). This experiment also shows that polarized lightis ranked higher than the disk of the moon in the internal compassof nocturnal dung beetles. Overall, we show that despite the taxo-nomic proximity of both species, they rely on different celestial cuesfor their dominant directional reference under dim light conditions.Because both species exhibit similar orientation behaviors but differ intheir activity window, our data suggest that nocturnal conditions(presumably the lower light intensity), or a circadian rhythm, cause theobserved switch to a polarization compass in the nocturnal species.
Central Complex Cells Are Sensitive to Polarized UV Light. Next, wesought to understand how the polarized light pattern and the po-sition of the sun or moon are processed neurally in the beetle’sbrain. To allow an accurate investigation of the neural substrate forpolarization-dependent orientation, we first had to define at whichwavelength of light the beetles perceive the skylight polarizationpattern. Dacke et al. (15) suggested that dung beetles detect po-larized light either in the green or UV wavelength. Therefore, weinvestigated the orientation performance of both of our modelspecies in an indoor arena lit by polarized light from above, with apeak either in the UV (365 nm) or green (530 nm) wavelength (Fig.2 A and B and Fig. S4). Each beetle was made to roll a ball twicefrom the center to the perimeter of a circular indoor arena. Be-tween each roll, the polarizer either remained in place (control) orwas turned by 90° (test). Under polarized UV light, both specieschanged their heading by 90° [Fig. 2B (in both species, P < 0.001 byV test, with an expected mean of 90°) and Figs. S1B and S2 E andF], whereas under polarized green light, the beetles were dis-oriented (Figs. S1B and S4). Thus, as in many other insects (19–22),both of our dung beetle species detect the skylight polarizationpattern in the UV range.To analyze the tuning of the polarization-sensitive neurons, we
presented the beetles with zenithal polarized UV light that hadsimilar light intensities as in the behavioral experiments. Beforestarting to roll, dung beetles always climb on top of their balls andperform “orientation dances” in which they rotate around theirown body axis (23). To simulate this vertical rotation underneath
0°
90°-90°
180°
0°
90°-90°
180°
B
C
controltest
D
0°
90°-90°
180°
N
A
sun/moon
controltest
Change of direction
Change of direction 0°
90°-90°
180°
ND
Fig. 1. Celestial cue preference in dung beetles under a natural sky. (A) Sche-matic illustration of the polarization pattern around a celestial body (sun ormoon). Change of direction in diurnal (D, Left) and nocturnal (N, Right)beetles rolling under a sun-lit (B) or moon-lit (C) sky. The change of directionwas calculated as the angular difference between two consecutive rolls, ei-ther without manipulation (control, ●) or when the sun or moon wasreflected to the opposite sky hemisphere between the two rolls (test,○). Themean directions (μ) are indicated by black (control) or red (test) lines, anderror bars indicate circular SDs. (B) Without manipulation, both species keptthe direction [P < 0.001 by V test; μdiurnal (±SD) = 2.6° ± 17.98°, n = 20;μnocturnal = −8.7° ± 38.34°, n = 20). When the sun was reflected to the op-posite sky hemisphere (and the real sun was shaded), both species respondedto this change (P < 0.001 by V-test; μdiurnal = 178.9° ± 54.6°, n = 20; μnocturnal =163.8° ± 46.58°, n = 20). (C) Under the moon in the control experiments,both species showed a constant rolling direction (P < 0.001 by V test; μdiurnal =3.1° ± 35.39°, n = 20; μnocturnal = −3.3° ± 37.87°, n = 20). When the moon wasreflected to the opposite sky hemisphere, the diurnal species followed theposition change of the moon (P = 0.002 by V test; μ = 179.5° ± 72.37°, n = 20),whereas the nocturnal species continued rolling in the original rolling direction(P < 0.001 by V test; μ = −13.4° ± 74.27°, n = 20).
11396 | www.pnas.org/cgi/doi/10.1073/pnas.1501272112 el Jundi et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020
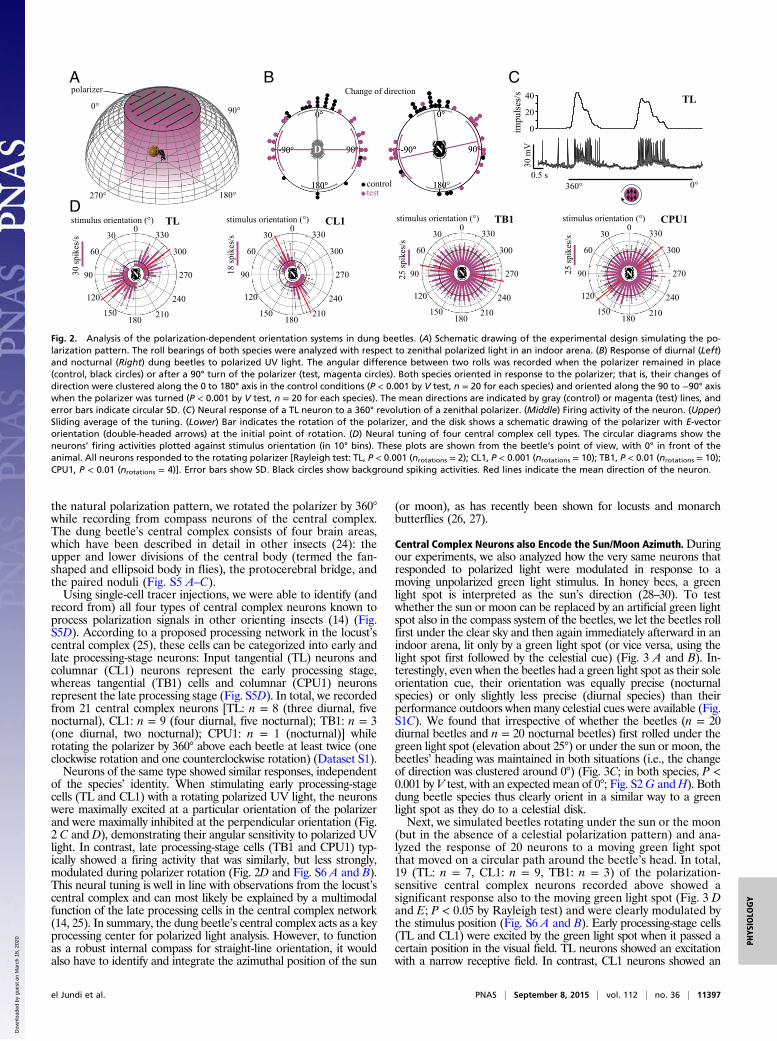
the natural polarization pattern, we rotated the polarizer by 360°while recording from compass neurons of the central complex.The dung beetle’s central complex consists of four brain areas,which have been described in detail in other insects (24): theupper and lower divisions of the central body (termed the fan-shaped and ellipsoid body in flies), the protocerebral bridge, andthe paired noduli (Fig. S5 A–C).Using single-cell tracer injections, we were able to identify (and
record from) all four types of central complex neurons known toprocess polarization signals in other orienting insects (14) (Fig.S5D). According to a proposed processing network in the locust’scentral complex (25), these cells can be categorized into early andlate processing-stage neurons: Input tangential (TL) neurons andcolumnar (CL1) neurons represent the early processing stage,whereas tangential (TB1) cells and columnar (CPU1) neuronsrepresent the late processing stage (Fig. S5D). In total, we recordedfrom 21 central complex neurons [TL: n = 8 (three diurnal, fivenocturnal), CL1: n = 9 (four diurnal, five nocturnal); TB1: n = 3(one diurnal, two nocturnal); CPU1: n = 1 (nocturnal)] whilerotating the polarizer by 360° above each beetle at least twice (oneclockwise rotation and one counterclockwise rotation) (Dataset S1).Neurons of the same type showed similar responses, independent
of the species’ identity. When stimulating early processing-stagecells (TL and CL1) with a rotating polarized UV light, the neuronswere maximally excited at a particular orientation of the polarizerand were maximally inhibited at the perpendicular orientation (Fig.2 C and D), demonstrating their angular sensitivity to polarized UVlight. In contrast, late processing-stage cells (TB1 and CPU1) typ-ically showed a firing activity that was similarly, but less strongly,modulated during polarizer rotation (Fig. 2D and Fig. S6 A and B).This neural tuning is well in line with observations from the locust’scentral complex and can most likely be explained by a multimodalfunction of the late processing cells in the central complex network(14, 25). In summary, the dung beetle’s central complex acts as a keyprocessing center for polarized light analysis. However, to functionas a robust internal compass for straight-line orientation, it wouldalso have to identify and integrate the azimuthal position of the sun
(or moon), as has recently been shown for locusts and monarchbutterflies (26, 27).
Central Complex Neurons also Encode the Sun/Moon Azimuth.Duringour experiments, we also analyzed how the very same neurons thatresponded to polarized light were modulated in response to amoving unpolarized green light stimulus. In honey bees, a greenlight spot is interpreted as the sun’s direction (28–30). To testwhether the sun or moon can be replaced by an artificial green lightspot also in the compass system of the beetles, we let the beetles rollfirst under the clear sky and then again immediately afterward in anindoor arena, lit only by a green light spot (or vice versa, using thelight spot first followed by the celestial cue) (Fig. 3 A and B). In-terestingly, even when the beetles had a green light spot as their soleorientation cue, their orientation was equally precise (nocturnalspecies) or only slightly less precise (diurnal species) than theirperformance outdoors when many celestial cues were available (Fig.S1C). We found that irrespective of whether the beetles (n = 20diurnal beetles and n = 20 nocturnal beetles) first rolled under thegreen light spot (elevation about 25°) or under the sun or moon, thebeetles’ heading was maintained in both situations (i.e., the changeof direction was clustered around 0°) (Fig. 3C; in both species, P <0.001 by V test, with an expected mean of 0°; Fig. S2G andH). Bothdung beetle species thus clearly orient in a similar way to a greenlight spot as they do to a celestial disk.Next, we simulated beetles rotating under the sun or the moon
(but in the absence of a celestial polarization pattern) and ana-lyzed the response of 20 neurons to a moving green light spotthat moved on a circular path around the beetle’s head. In total,19 (TL: n = 7, CL1: n = 9, TB1: n = 3) of the polarization-sensitive central complex neurons recorded above showed asignificant response also to the moving green light spot (Fig. 3 Dand E; P < 0.05 by Rayleigh test) and were clearly modulated bythe stimulus position (Fig. S6 A and B). Early processing-stage cells(TL and CL1) were excited by the green light spot when it passed acertain position in the visual field. TL neurons showed an excitationwith a narrow receptive field. In contrast, CL1 neurons showed an
A B C
D
Fig. 2. Analysis of the polarization-dependent orientation systems in dung beetles. (A) Schematic drawing of the experimental design simulating the po-larization pattern. The roll bearings of both species were analyzed with respect to zenithal polarized light in an indoor arena. (B) Response of diurnal (Left)and nocturnal (Right) dung beetles to polarized UV light. The angular difference between two rolls was recorded when the polarizer remained in place(control, black circles) or after a 90° turn of the polarizer (test, magenta circles). Both species oriented in response to the polarizer; that is, their changes ofdirection were clustered along the 0 to 180° axis in the control conditions (P < 0.001 by V test, n = 20 for each species) and oriented along the 90 to −90° axiswhen the polarizer was turned (P < 0.001 by V test, n = 20 for each species). The mean directions are indicated by gray (control) or magenta (test) lines, anderror bars indicate circular SD. (C) Neural response of a TL neuron to a 360° revolution of a zenithal polarizer. (Middle) Firing activity of the neuron. (Upper)Sliding average of the tuning. (Lower) Bar indicates the rotation of the polarizer, and the disk shows a schematic drawing of the polarizer with E-vectororientation (double-headed arrows) at the initial point of rotation. (D) Neural tuning of four central complex cell types. The circular diagrams show theneurons’ firing activities plotted against stimulus orientation (in 10° bins). These plots are shown from the beetle’s point of view, with 0° in front of theanimal. All neurons responded to the rotating polarizer [Rayleigh test: TL, P < 0.001 (nrotations = 2); CL1, P < 0.001 (nrotations = 10); TB1, P < 0.01 (nrotations = 10);CPU1, P < 0.01 (nrotations = 4)]. Error bars show SD. Black circles show background spiking activities. Red lines indicate the mean direction of the neuron.
el Jundi et al. PNAS | September 8, 2015 | vol. 112 | no. 36 | 11397
PHYS
IOLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020
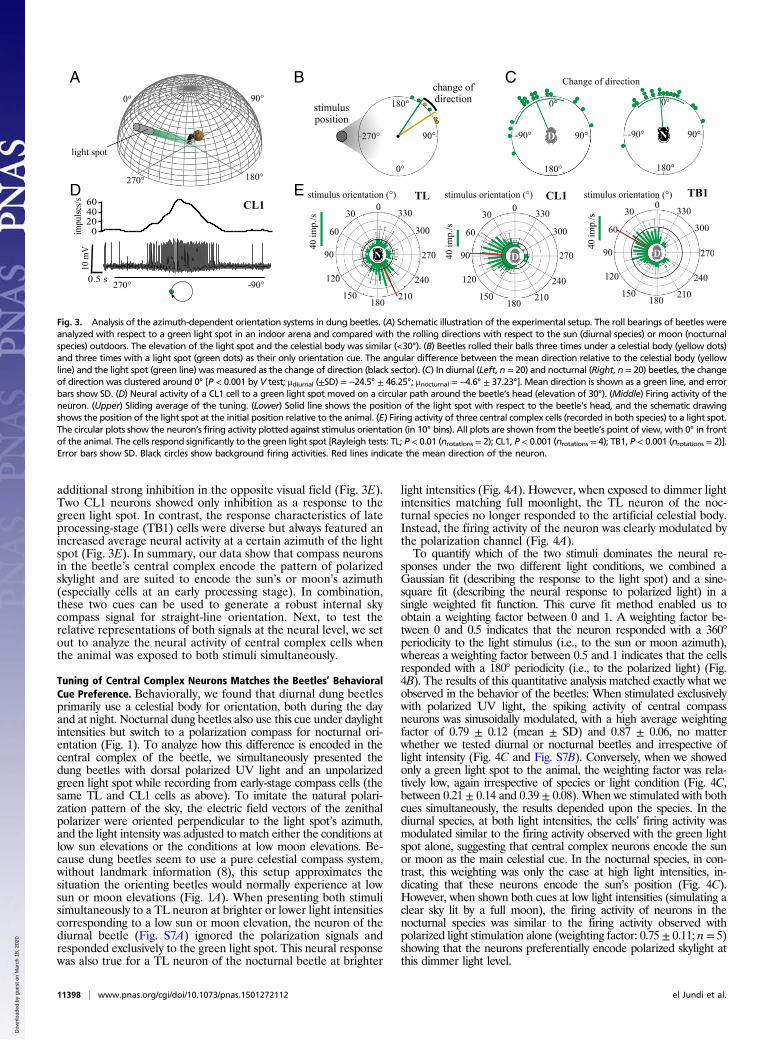
additional strong inhibition in the opposite visual field (Fig. 3E).Two CL1 neurons showed only inhibition as a response to thegreen light spot. In contrast, the response characteristics of lateprocessing-stage (TB1) cells were diverse but always featured anincreased average neural activity at a certain azimuth of the lightspot (Fig. 3E). In summary, our data show that compass neuronsin the beetle’s central complex encode the pattern of polarizedskylight and are suited to encode the sun’s or moon’s azimuth(especially cells at an early processing stage). In combination,these two cues can be used to generate a robust internal skycompass signal for straight-line orientation. Next, to test therelative representations of both signals at the neural level, we setout to analyze the neural activity of central complex cells whenthe animal was exposed to both stimuli simultaneously.
Tuning of Central Complex Neurons Matches the Beetles’ BehavioralCue Preference. Behaviorally, we found that diurnal dung beetlesprimarily use a celestial body for orientation, both during the dayand at night. Nocturnal dung beetles also use this cue under daylightintensities but switch to a polarization compass for nocturnal ori-entation (Fig. 1). To analyze how this difference is encoded in thecentral complex of the beetle, we simultaneously presented thedung beetles with dorsal polarized UV light and an unpolarizedgreen light spot while recording from early-stage compass cells (thesame TL and CL1 cells as above). To imitate the natural polari-zation pattern of the sky, the electric field vectors of the zenithalpolarizer were oriented perpendicular to the light spot’s azimuth,and the light intensity was adjusted to match either the conditions atlow sun elevations or the conditions at low moon elevations. Be-cause dung beetles seem to use a pure celestial compass system,without landmark information (8), this setup approximates thesituation the orienting beetles would normally experience at lowsun or moon elevations (Fig. 1A). When presenting both stimulisimultaneously to a TL neuron at brighter or lower light intensitiescorresponding to a low sun or moon elevation, the neuron of thediurnal beetle (Fig. S7A) ignored the polarization signals andresponded exclusively to the green light spot. This neural responsewas also true for a TL neuron of the nocturnal beetle at brighter
light intensities (Fig. 4A). However, when exposed to dimmer lightintensities matching full moonlight, the TL neuron of the noc-turnal species no longer responded to the artificial celestial body.Instead, the firing activity of the neuron was clearly modulated bythe polarization channel (Fig. 4A).To quantify which of the two stimuli dominates the neural re-
sponses under the two different light conditions, we combined aGaussian fit (describing the response to the light spot) and a sine-square fit (describing the neural response to polarized light) in asingle weighted fit function. This curve fit method enabled us toobtain a weighting factor between 0 and 1. A weighting factor be-tween 0 and 0.5 indicates that the neuron responded with a 360°periodicity to the light stimulus (i.e., to the sun or moon azimuth),whereas a weighting factor between 0.5 and 1 indicates that the cellsresponded with a 180° periodicity (i.e., to the polarized light) (Fig.4B). The results of this quantitative analysis matched exactly what weobserved in the behavior of the beetles: When stimulated exclusivelywith polarized UV light, the spiking activity of central compassneurons was sinusoidally modulated, with a high average weightingfactor of 0.79 ± 0.12 (mean ± SD) and 0.87 ± 0.06, no matterwhether we tested diurnal or nocturnal beetles and irrespective oflight intensity (Fig. 4C and Fig. S7B). Conversely, when we showedonly a green light spot to the animal, the weighting factor was rela-tively low, again irrespective of species or light condition (Fig. 4C,between 0.21 ± 0.14 and 0.39 ± 0.08). When we stimulated with bothcues simultaneously, the results depended upon the species. In thediurnal species, at both light intensities, the cells’ firing activity wasmodulated similar to the firing activity observed with the green lightspot alone, suggesting that central complex neurons encode the sunor moon as the main celestial cue. In the nocturnal species, in con-trast, this weighting was only the case at high light intensities, in-dicating that these neurons encode the sun’s position (Fig. 4C).However, when shown both cues at low light intensities (simulating aclear sky lit by a full moon), the firing activity of neurons in thenocturnal species was similar to the firing activity observed withpolarized light stimulation alone (weighting factor: 0.75± 0.11; n= 5)showing that the neurons preferentially encode polarized skylight atthis dimmer light level.
A B C
D E
Fig. 3. Analysis of the azimuth-dependent orientation systems in dung beetles. (A) Schematic illustration of the experimental setup. The roll bearings of beetles wereanalyzed with respect to a green light spot in an indoor arena and compared with the rolling directions with respect to the sun (diurnal species) or moon (nocturnalspecies) outdoors. The elevation of the light spot and the celestial body was similar (<30°). (B) Beetles rolled their balls three times under a celestial body (yellow dots)and three times with a light spot (green dots) as their only orientation cue. The angular difference between the mean direction relative to the celestial body (yellowline) and the light spot (green line) was measured as the change of direction (black sector). (C) In diurnal (Left, n = 20) and nocturnal (Right, n = 20) beetles, the changeof direction was clustered around 0° [P < 0.001 by V test; μdiurnal (±SD) = −24.5° ± 46.25°; μnocturnal = −4.6° ± 37.23°]. Mean direction is shown as a green line, and errorbars show SD. (D) Neural activity of a CL1 cell to a green light spot moved on a circular path around the beetle’s head (elevation of 30°). (Middle) Firing activity of theneuron. (Upper) Sliding average of the tuning. (Lower) Solid line shows the position of the light spot with respect to the beetle’s head, and the schematic drawingshows the position of the light spot at the initial position relative to the animal. (E) Firing activity of three central complex cells (recorded in both species) to a light spot.The circular plots show the neuron’s firing activity plotted against stimulus orientation (in 10° bins). All plots are shown from the beetle’s point of view, with 0° in frontof the animal. The cells respond significantly to the green light spot [Rayleigh tests: TL; P < 0.01 (nrotations = 2); CL1, P < 0.001 (nrotations = 4); TB1, P < 0.001 (nrotations = 2)].Error bars show SD. Black circles show background firing activities. Red lines indicate the mean direction of the neuron.
11398 | www.pnas.org/cgi/doi/10.1073/pnas.1501272112 el Jundi et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020

In summary, the neural encoding of celestial orientation cues inbright and dim light conditions closely matches the observed cuepreference of these cues during the natural orientation behavior ofour model species in the field (Fig. 5): Nocturnal dung beetles usethe celestial polarization pattern as their main compass cue at night,whereas diurnal beetles use the sun as their main compass cueduring the day.
Celestial Polarization Pattern: A Reliable Orientation Cue forNocturnal Animals? What then are the mechanisms that generatethe switch to polarized light under dim light conditions in thenocturnal species? Studies in locusts and crickets have shown thatthe tuning of compass neurons to polarized light is intensity-independent above a particular threshold level of polarized lightintensity (31, 32), whereas responses to unpolarized light stronglydepend on light intensity (32). Consequently, when both stimuli aredimmed to lunar light intensities, only the response in the channelthat is processed in an intensity-dependent manner should be di-minished (i.e., the input from the light spot in these experiments).At lunar light intensities, the inputs should thus be modified in favorof the polarization channel for as long as the polarization analyzerof the eye is sensitive enough to detect a signal. In the nocturnaldung beetle, we found that the neurons responded reliably to brightas well as dim polarized light. The neuronal response in the diurnalspecies, on the other hand, was weak or absent at dim light in-tensities compared with the neuronal response at high light in-tensities (Fig. S6C). These neural responses might, at least partially,be a result of the size ratio between the dorsal rim area and mainretina. The dorsal rim areas of crepuscular beetles are enormous(33), and our first preliminary observations suggest that this is alsotrue for the nocturnal species tested here. In contrast, the dorsal rimareas of the diurnal species seem to be relatively small. The highersensitivity to polarized light, and thus the weighting of differentcelestial cues in central complex neurons, might therefore be de-termined at an earlier stage in the brain. This difference in sensi-tivity to the polarization of light is also reflected in the orientationbehavior of our two model species: Under dim light intensities,nocturnal navigators will adhere to the direction given by the po-larization pattern rather than to the direction indicated by themoon’s position, whereas the diurnal species follows the directionderived from the moon (Fig. 5). This cue preference raises an im-portant question regarding the advantages that a sky-wide polari-zation pattern offers over a bright point source (in the form of amoon) for nocturnal orientation. Because this difference in cuepreference is a phenomenon observed only under dim light in-tensities, the answer is most likely a gain in sensitivity. One excellent
strategy to gain visual sensitivity is to pool signals from many pho-toreceptors in one second-order neuron, a mechanism known asspatial summation (34, 35). The polarization pattern extends overthe entire sky, and therefore allows the receiver to integrate signalsover a wide visual field. At low light intensities, the polarizationpattern should be a more reliable orientation cue than a moon,appearing as a dimly illuminated disk in the sky that may also beblocked from view by clouds. Moreover, even under the low light ofa quarter or crescent moon, S. satyrus can consistently roll a straight-line path when the moon itself is blocked from view, almost cer-tainly using the lunar polarization pattern for orientation (9). Sim-ilarly, crickets are able to detect polarized light even at lightintensities equivalent to the light intensities of a moonless night sky(36). Under all these extremely dim conditions, spatial summationmight be even more relevant for the orientation compass of noc-turnal insects, as hypothesized by Labhart et al. (31); thus, the po-larization pattern might offer the most reliable celestial referencefor nocturnal navigation. Even though the moon, in any phase, is
A
0° 360°
010
20
30
-90° 270°
sulumits
dim
bright N B
5
15
25weight: 0.34
weight: 0.87
5
15
25
360°270°
0°-90°
0.0
0.2
0.4
0.6
0.8
1.0
thgiew
LOP
tops thgiL
dim
10 9 7 5 5 5 7 7 6 6 6 6
N Dbright
sulumits
Cdimbright)s/p
mi(ytivitca
n aem
360°270°
0°-90°
)s/pmi(
ytivitcanae
m
010
20
30
Fig. 4. Decoding the celestial cue preference in the dung beetle’s central complex. (A, Upper) Sliding average of the response of a TL neuron to a rotatingpolarizer (Left, magenta), a light spot (Middle, green), and when both stimuli were shown simultaneously (Right, black) at light intensities that simulatedconditions at low sun elevations. Gray curves show individual rotations. (A, Lower) Sliding average of the response of the same neuron, with light intensitiesthat simulated conditions of a full-moon–lit night. Solid lines show stimulus timing, and disks and circles indicate stimulus type (Figs. 2 and 3). (B) Mean spikingactivity (bin size of 10°) of the same neuron when both stimuli were shown simultaneously at high light intensity (Upper) and low light intensity (Lower) (sameresponses as in A, Right, black). The solid curve shows the combined Gaussian fit and sine-square fit. The weighting factor indicates how well the neuraltuning was described by a Gaussian fit (between 0 and 0.5, response to the moving light spot) or a sine-square fit (between 0.5 and 1, response to rotatingpolarizer) function. Error bars show SD. (C) Weights of the neural response to different light stimuli. (Left) Neural responses of central complex neurons(TL and CL1) of the nocturnal species to polarized (POL) light (Left; TL, n = 5; CL1, n = 5), a light spot (Middle; TL, n = 4; CL1, n = 5), or both stimuli presentedsimultaneously (Right, red; TL, n = 4; CL1, n = 3). A white background denotes high light intensities, and a gray background denotes full-moon light conditions(TL, n = 3; CL1, n = 2). Numbers represent cells analyzed. Error bars show SD. (Right) Same, but for cells analyzed in the diurnal species (TL, n = 3; CL1, n = 4).
Fig. 5. Summary of results. The illustration of the ball-rolling dung beetlerepresents the behavioral experiments. The schematic drawing of the cell withthe recording trace represents the physiological experiments. Diurnal species(Left) and nocturnal species (Right) are shown. Both were analyzed at low sunelevation (behavior) or with light conditions simulating low sun elevation(physiology) (Upper) and at low moon elevations (behavior) or with lightsimulating low moon elevation (physiology) (Lower). The diurnal species usesthe celestial body as its primary orientation cue during both day and night, butthe nocturnal species switches to the polarization pattern as its primary cue whenrolling balls at night. The behavioral demonstration of the compass cue prefer-ence accurately matches the neural encoding of the celestial cue weightingin the compass neurons of the brain. In each panel, pale gray cues indicatethat this cue is not used as the main reference in this condition.
el Jundi et al. PNAS | September 8, 2015 | vol. 112 | no. 36 | 11399
PHYS
IOLO
GY
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020

only available for about half of the month, it can still polarize thenight sky when it is up to 15° below the horizon (37), extendingthe period when S. satyrus can reliably forage. Nevertheless, whenthe sun and moon are further below the horizon than 15°, and thenight sky has no overall pattern of polarization, S. satyrus can fallback on the stars of the Milky Way to roll dung balls through theAfrican bush (16), a cue that presumably ranks lowest in the cuepreference of orientation signals.In summary, our study clearly demonstrates how the visual
ecology of an orienting animal is reflected in its orientation compasssystem. Here, the dynamic activity of neurons in the central complexprovides the nocturnal orienting animal with a simple yet efficienttool to lock onto the most reliable signal available. This dynamictuning generates a robust internal compass that can ensure preciseorientation behavior with the cues available.
Materials and MethodsGeneral. Beetles were collected in South Africa. Details are provided in SIMaterials and Methods.
Investigation of the Celestial Cue Preference. To test which of the cues (sun/moon or polarization pattern) dominates the sky compass of dung beetles,we analyzed the rolling behavior with the celestial cues aligned or set inconflict (SI Materials and Methods).
Indoor Experiments with Polarized Light. To analyze the wavelength rangeover which the beetles perceive polarized light, we presented zenithal po-larized light in the UV or green range to the beetles in an indoor arena (SIMaterials and Methods).
Orientation Under the Sun and a Green Light Spot. To test whether beetlesinterpret a green light spot as the sun or moon, we compared the bearingstaken with respect to the sun or moon with the bearings taken withrespect to a green light spot indoors. Details are provided in SI Materialsand Methods.
Physiology. To analyze neural activity, we used a standard electrophysiologymethod to record responses of neurons intracellularly. Details are provided inSI Materials and Methods.
Visual Stimulations. To test whether central complex neurons are sensitive topolarized UV light and/or to a green light spot, we designed a stimulus similarto the one described by Pfeiffer and Homberg (38). Details are provided in SIMaterials and Methods.
Anatomy. Immunohistochemistry, image acquisition, and 3D reconstructionprocedures for neurons and neuropils have been described in detail by elJundi et al. (39). Details are provided in SI Materials and Methods.
Data Analysis. All experiments were analyzed with custom-written programsin MATLAB (MathWorks). Details are provided in SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Drs. James Foster, Stanley Heinze, OctavianKnoll, Keram Pfeiffer, and Christine Scholtyssek for their helpful discussionsand comments. We are grateful to Ted and Winnie Harvey for their invaluablehelp in the field. We also thank Drs. Erich Buchner and Christian Wegener forproviding the antisynapsin antibody. Funding for this project was provided byVetenskapsrådet, the Wallenberg Foundation, the Wenner-Gren-Foundation,the Royal Physiographic Society in Lund, the Lars-Hierta Memorial Foundation,and the South African National Research Foundation.
1. Wehner R (1984) Astronavigation in insects. Annu Rev Entomol 29:277–298.2. Wiltschko W, Wiltschko R (1991) Magnetic orientation and celestial cues in migratory
orientation. Orientation in Birds, ed Berthold P (Birkhäuser, Basel), pp 16–37.3. Frisch KV (1949) Die Polarisation des Himmelslichtes als orientierender Faktor bei den
Tänzen der Bienen. Experientia 5(4):142–148. German.4. Horváth G, Varjú D (2004) Polarized Light in Animal Vision: Polarization Patterns in
Nature (Springer, Heidelberg).5. el Jundi B, Smolka J, Baird E, Byrne MJ, Dacke M (2014) Diurnal dung beetles use the
intensity gradient and the polarization pattern of the sky for orientation. J Exp Biol217(Pt 13):2422–2429.
6. Gál J, Horváth G, Barta A, Wehner R (2001) Polarization of the moonlit clear night skymeasured by full-sky imaging polarimetry at full Moon: Comparison of the polari-zation of the moonlit and sunlit skies. J Geophys Res 106(D19):22647–22653.
7. Dacke M, Nilsson DE, Scholtz CH, Byrne M, Warrant EJ (2003) Animal behaviour: Insectorientation to polarized moonlight. Nature 424(6944):33.
8. Dacke M, Byrne M, Smolka J, Warrant E, Baird E (2013) Dung beetles ignore land-marks for straight-line orientation. J Comp Physiol A Neuroethol Sens Neural BehavPhysiol 199(1):17–23.
9. Dacke M, Byrne MJ, Baird E, Scholtz CH, Warrant EJ (2011) How dim is dim? Precisionof the celestial compass in moonlight and sunlight. Philos Trans R Soc Lond B Biol Sci366(1565):697–702.
10. McIntyre P, Caveney S (1998) Superposition optics and the time of flight in onitinedung beetles. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 183:45–60.
11. Greiner B, et al. (2007) Eye structure correlates with distinct foraging-bout timing inprimitive ants. Curr Biol 17(20):R879–R880.
12. Warrant EJ (2008) Seeing in the dark: Vision and visual behaviour in nocturnal beesand wasps. J Exp Biol 211(Pt 11):1737–1746.
13. Heinze S, Homberg U (2007) Maplike representation of celestial E-vector orientationsin the brain of an insect. Science 315(5814):995–997.
14. Homberg U, Heinze S, Pfeiffer K, Kinoshita M, el Jundi B (2011) Central neural codingof sky polarization in insects. Philos Trans R Soc Lond B Biol Sci 366(1565):680–687.
15. Dacke M, Byrne MJ, Scholtz CH, Warrant EJ (2004) Lunar orientation in a beetle. ProcBiol Sci 271(1537):361–365.
16. Dacke M, Baird E, Byrne M, Scholtz CH, Warrant EJ (2013) Dung beetles use the MilkyWay for orientation. Curr Biol 23(4):298–300.
17. Dacke M, el Jundi B, Smolka J, Byrne M, Baird E (2014) The role of the sun in thecelestial compass of dung beetles. Philos Trans R Soc Lond B Biol Sci 369(1636):20130036.
18. Byrne M, Dacke M, Nordström P, Scholtz C, Warrant E (2003) Visual cues used by ball-rolling dung beetles for orientation. J Comp Physiol A Neuroethol Sens Neural BehavPhysiol 189(6):411–418.
19. Labhart T, Meyer EP (1999) Detectors for polarized skylight in insects: A survey ofommatidial specializations in the dorsal rim area of the compound eye. Microsc ResTech 47(6):368–379.
20. Sauman I, et al. (2005) Connecting the navigational clock to sun compass input inmonarch butterfly brain. Neuron 46(3):457–467.
21. Weir PT, Dickinson MH (2012) Flying Drosophila orient to sky polarization. Curr Biol22(1):21–27.
22. Wernet MF, et al. (2012) Genetic dissection reveals two separate retinal substrates for
polarization vision in Drosophila. Curr Biol 22(1):12–20.23. Baird E, Byrne MJ, Smolka J, Warrant EJ, Dacke M (2012) The dung beetle dance: An
orientation behaviour? PLoS One 7(1):e30211.24. Ito K, et al.; Insect Brain Name Working Group (2014) A systematic nomenclature for
the insect brain. Neuron 81(4):755–765.25. Heinze S, Gotthardt S, Homberg U (2009) Transformation of polarized light in-
formation in the central complex of the locust. J Neurosci 29(38):11783–11793.26. Heinze S, Reppert SM (2011) Sun compass integration of skylight cues in migratory
monarch butterflies. Neuron 69(2):345–358.27. el Jundi B, Pfeiffer K, Heinze S, Homberg U (2014) Integration of polarization and
chromatic cues in the insect sky compass. J Comp Physiol A Neuroethol Sens Neural
Behav Physiol 200(6):575–589.28. Brines ML, Gould JL (1979) Bees have rules. Science 206(4418):571–573.29. Edrich W, Neumeyer C, von Heiversen O (1979) “Anti-sun orientation” of bees with
regard to a field of ultraviolet light. J Comp Physiol A Neuroethol Sens Neural Behav
Physiol 134(2):151–157.30. Rossel S, Wehner R (1984) Celestial orientation in bees: the use of spectral cues.
J Comp Physiol A Neuroethol Sens Neural Behav Physiol 155(5):605–613.31. Labhart T, Petzold J, Helbling H (2001) Spatial integration in polarization-sensitive
interneurones of crickets: A survey of evidence, mechanisms and benefits. J Exp Biol
204(Pt 14):2423–2430.32. Kinoshita M, Pfeiffer K, Homberg U (2007) Spectral properties of identified polarized-
light sensitive interneurons in the brain of the desert locust Schistocerca gregaria.
J Exp Biol 210(Pt 8):1350–1361.33. Warrant E (2004) Vision in the dimmest habitats on earth. J Comp Physiol A Neuroethol
Sens Neural Behav Physiol 190(10):765–789.34. Dacke M, Nordström P, Scholtz CH (2003) Twilight orientation to polarised light in the
crepuscular dung beetle Scarabaeus zambesianus. J Exp Biol 206(Pt 9):1535–1543.35. Warrant EJ (1999) Seeing better at night: Life style, eye design and the optimum
strategy of spatial and temporal summation. Vision Res 39(9):1611–1630.36. Herzmann D, Labhart T (1989) Spectral sensitivity and absolute threshold of polari-
zation vision in crickets: A behavioral study. J Comp Physiol A Neuroethol Sens Neural
Behav Physiol 165(3):315–319.37. Cronin TW, Warrant EJ, Greiner B (2006) Celestial polarization patterns during twi-
light. Appl Opt 45(22):5582–5589.38. Pfeiffer K, Homberg U (2007) Coding of azimuthal directions via time-compensated
combination of celestial compass cues. Curr Biol 17(11):960–965.39. el Jundi B, et al. (2009) The locust standard brain: A 3D standard of the central
complex as a platform for neural network analysis. Front Syst Neurosci 3:21.40. Johnsen S, et al. (2006) Crepuscular and nocturnal illumination and its effects on color
perception by the nocturnal hawkmoth Deilephila elpenor. J Exp Biol 209(Pt 5):
789–800.41. Batschelet E (1981) Circular Statistics in Biology (Academic, London).42. Labhart T (1996) How polarization-sensitive interneurones of crickets perform at low
degrees of polarization. J Exp Biol 199(Pt 7):1467–1475.
11400 | www.pnas.org/cgi/doi/10.1073/pnas.1501272112 el Jundi et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 1
8, 2
020





























