Embed Size (px)
Citation preview

N e uron
A r t i c l e
E long ation F a ctor 2 a nd Fra gile X M e nta lR etard ation Prote in C ontrol th e Dyn a m icTra nslation of Arc / Arg3.1 E ss e ntia l for m G luR-LT DSung jin P ark ,1,7 J o o M in P ark ,1,7 S a ng m o k K im ,1 J in-A h K im ,1 J a s on D . Sh e p h erd ,4 C onsta n c e L. S m ith-H ic k s,2
Sho a ib C ho w d hury,1 W a lter K a ufm a nn,2 D ietm ar K uhl,5 A le x e y G . R y a z a no v,6 R ic h ard L. H ug a nir,1,3 D a vid J . Lin d e n,1
a n d P a ul F . W orle y1,2,*1D e p artm ent of N eurosc ienc e2D e p artm ent of N eurology3H ow ard Hughes M e d ic a l InstituteJohns H op kins University S chool of M e d ic ine , B a ltimore , M D 20205, U S A4The P ic ow er Institute for Le arning and M e mory, M assa chusetts Institute of T e chnology, C a m bridg e , M A 02139, U S A5M ole cular N eurob iology, D e p artm ent of B iology- C hemistry-Pharm acy, Fre ie Universit at B erlin, 14195 B erlin, G erm any6D e p artm ent of Pharm a c ology, University of M e d ic ine and D entistry of N e w J ersey-Rob ert Wood Johnson M e d ic a l S chool,P isc ata w ay, N Y 08854, U S A7These authors c ontribute d e qua lly to this work* C orrespond enc e: p [email protected] duD OI 10.1016/j.neuron.2008.05.023
S U M M A R Y
G rou p I m eta b o tro p ic g luta m a te re c e p tors (m G lu R)in d u c e long-t erm d e pre ssio n (LT D) th at re q uire sprote in synth e sis. H ere , w e d e m on strate th atArc / Arg3.1 is tra nslation a lly in d u c e d w ithin 5 m in ofm G lu R a ctiv ation, a n d this re s p on s e is e ss e ntia l form G lu R-d e p e n d e nt LT D . Th e in cre a s e in Arc / Arg3.1tra nslation re q u ire s e E F 2 K , a C a 2+ / c a lm o d ulin-d e-p e n d e nt k in a s e th at b in d s m G lu R a n d d iss o c iate su p on m G lu R a ctiv ation, w h ere u p on it p ho s p hory-late s e E F 2. P h o s p ho-e E F 2 a cts to slo w th e e long a-tion ste p of tra nslation a n d inhib its g e n era l prote insynth e sis b ut sim ulta n e ously in cre a s e s Arc / Arg3.1tra nslation. G e n etic d e letion of eEF2K re sults in a s e-le ctiv e d efic it of ra p id m G lu R -d e p e n d e nt Arc / Arg3.1tra nslation a n d m G lu R-LT D . This ra p id tra nslation a lm e c h a nis m is d isru p te d in th e fra g ile X d is e a s em ous e (Fmr1 K O) in w hic h m G lu R -LT D d o e s not re-q uire d e no v o prote in synth e sis b ut d o e s re q u ireArc / Arg3.1. W e pro p o s e a m o d e l in w h ic h e E F 2 K -e E F 2 a n d F M R P c o ord in ate ly c ontro l th e d yn a m ictra nslation of Arc / Arg3.1 m R N A in d e n drite s th at iscritic a l for syn a p s e-s p e c ific LT D .
IN T R O D U C T I O N
Long-lasting forms of syna ptic p lastic ity re quire d e novo prote insynthesis (K e lleher et a l., 2004; W ang and Tie dg e , 2004).N-m ethyl-D-asp artate (N M D A) re c e ptor-d e p end ent long-termpotentiation (LTP) of the S chaffer c ollatera l- C A1 syna pse lastinglong er than 60 min is b lo c k e d by a g ents that ha lt translation.Late-phase N M D A-d e p end ent LT D in hip po c a m p a l slic e cul-tures is a lso prote in synthesis d e p end ent (K aud erer and K and e l,
2000). D e novo prote in synthesis is a lso re quire d for forms oflong-term d e pression (LT D) that are induc e d by group I m eta bo-trop ic gluta m ate re c e ptor (m G luR) a ctivation or by p a ire d-pulselow-fre quency stimulation (P P-L F S) (Hub er et a l., 2000). In c on-trast to N M D A re c e ptor-d e p end ent LTP and LT D , where the re-quire m ent for prote in synthesis is d e laye d , m G luR-LT D re quiresd e novo prote in synthesis w ithin 5–10 min. (Hub er et a l., 2000).
The products of d e novo prote in synthesis are hypothesiz e d tob e ‘‘c a pture d’’ at a ctive syna pses (Frey and M orris, 1997), butthe ir id entity re m a ins unknown. Arc/Arg3.1 is an imm e d iate-e arlyg ene (IE G) that is induc e d by N M D A re c e ptor a ctivation in vivo(Link et a l., 1995; Lyford et a l., 1995) and m e d iates a postsyna pticendo cytic p athw ay by intera cting w ith endophilin 2/3 anddyna min that se le ctive ly traffics A M P A re c e ptors (A M P AR)( C how dhury et a l., 2006). The a ctivity-d e p end ent e x pression ofArc/Arg3.1 mR N A and prote in und erlie a hom e ostatic m e cha-nism that m a inta ins a pre c ise leve l of A M P AR-d e p end ent e x c it-a b ility in c ond itions of p ersistently incre ase d or d e cre ase dsyna ptic input (She pherd et a l., 2006). Stud ies presente d hered e monstrate that Arc/Arg3.1 is a lso re quire d for m G luR andP P-L F S LT D . In c ontrast to chang es in Arc/Arg3.1 e x pressionthat o c cur over hours to d ays in hom e ostatic p lastic ity, m G luRa ctivation results in incre ases in Arc/Arg3.1 prote in w ithin3–5 min. Using b io che mic a l and g enetic a p pro a ches, w e d e m-onstrate that m G luR evok es ra p id Arc/Arg3.1 translation viaa signa ling p athw ay that involves euk aryotic e long ation fa ctor2 kinase (e E F 2K) and euk aryotic e long ation fa ctor 2 (e E F 2).e E F 2 is re quire d for the e long ation ste p of translation (Rya z anovet a l., 1988), while phospho-e E F 2 a cts as a potent inhib itor of thee long ation ste p in a m anner that is similar to che mic a l prote insynthesis inhib itors, such as cyc lohe ximid e (B e gueret et a l.,1977; O brig et a l., 1971). O ur d ata sup port a mod e l in whichra p id translationa l upre gulation of Arc/Arg3.1 is re quire d form G luR-d e p end ent LT D , and this translationa l induction is g ener-ate d as a c onse quenc e of lo c a l and transient inhib ition of thetranslation of other mR N As via phospho-e E F 2.
70 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

O ur stud ies a lso provid e insights into the mole cular b asis of a l-tere d syna ptic p lastic ity in fra gile X m enta l retard ation syndrom e .Fra gile X syndrom e , which is the most c ommon inherite d c auseof m enta l retard ation and autism (O ’D onne ll and W arren, 2002),is c ause d by an e x p ansion of C G G in the 50 untranslate d re gion ofthe fra gile X m enta l retard ation prote in (F M RP) g ene (F mr1) thatre duc es its e x pression. F M RP b inds to G -quartet-c onta iningR N As through the R G G box (D arne ll et a l., 2001; S cha efferet a l., 2001). F M R P functions as a translationa l re pressor of sp e-c ific syna ptic mR N As by a propose d m e chanism that m ayinvolve B C 1 R N A , a nontranslata b le m essa g e a bund ant in d en-drites (Z a lfa et a l., 2003; but se e a lso Ia c o ang e li et a l., 2008).F M RP is a lso re porte d to function as p art of a RIS C nuc le asec om p le x that re presses translation by R N A interferenc e ( C audyet a l., 2002; Ishizuk a et a l., 2002; Jin et a l., 2004). In F mr1 kno c k-out (K O) mic e , Arc/Arg3.1, a - C a M KII, and M A P1 B prote ins arere porte d to b e e levate d both in tota l bra in and syna ptosom a lfra ctions (Z a lfa et a l., 2003). N ota b ly, an incre ase d asso c iationof Arc/Arg3.1 mR N A w ith polyribosom es sugg ests that a gre aterfra ction of Arc/Arg3.1 mR N A is b e ing a ctive ly translate d in F mr1K O . The notion that misre gulate d e x pression of these prote insmight und erlie c ognitive d efic its in fra gile X syndrom e is sup-porte d by physiologic a l stud ies that im p lic ate F M RP in a ltere dsyna ptic p lastic ity. F mr1 K O mic e show robust m G luR-LT D(Hub er et a l., 2002) that is insensitive to prote in synthesis inhib-itors (H ou et a l., 2006; N osyreva and Hub er, 2006), sugg estingthat, when translation is d ere presse d due to the a bsenc e ofF M RP , ‘‘LT D prote ins’’ that are norm a lly produc e d in a stimu-lus-d e p end ent m anner are now c ontinuously synthesiz e d andare suffic ient to susta in m G luR-LT D w ithout the re quire m entfor d e novo prote in synthesis. Insight into the mole cular b asisof m G luR and F M RP-d e p end ent translation is provid e d by theobservations that the effic a cy of F M RP to inhib it translation isre gulate d by phosphorylation ( C e m an et a l., 2003) and thatm G luR a ctivation results in d e phosphorylation of F M RP byP P2A w ithin 1 min that is link e d to ra p id translationa l upre gula-tion of a targ et prote in, S A P A P3 (N arayanan et a l., 2007). A sim-ilar ra p id m G luR- and F M RP-d e p end ent incre ase in a - C a M KII,P S D-95 (Mud d ashetty et a l., 2007), and M A P1 B (D avid kovaand C arroll, 2007) has b e en re porte d , and M A P1 B is sugg este dto p lay a role in gluta m ate re c e ptor traffic king in culture dneurons. In the present study, w e find that ra p id , d e novoArc/Arg3.1 e x pression is d isrupte d in F mr1 K O neurons, andArc/Arg3.1/ F mr1 doub le K O mic e show re duc e d m G luR-LT D .These observations sugg est that e E F 2K/e E F 2 and F M RP-d e p end ent translationa l p athw ays are c ofunctiona l in c ontrollinga ra p id and transient sw itch of the translationa l m a chineryto mR N As that are essentia l for m G luR-d e p end ent syna pticd e pression.
R E S U L T S
m G l u R - L T D a n d P P - L F S L T D R e q u ir e A r c / A r g 3 . 1To e x a mine the role of Arc/Arg3.1 in m G luR-LT D , w e monitore dS chaffer c ollatera l- C A1 syna pses in a cute hip po c a m p a l slic espre p are d from w ild-typ e (W T) and Arc/Arg3.1 kno c kout (K O)mic e . B ase line syna ptic prop erties, inc lud ing the fib er volley-fE P S P re lationship (an ind e x of b asa l syna ptic strength) and
p a ire d-pulse fa c ilitation ratio w ere norm a l in Arc/Arg3.1 K Omic e (se e F igures S1A and S1 B ava ila b le online), c onfirminga previous re port (P lath et a l., 2006). In W T slic es, tre atm entw ith the group I m G luR a gonist D H P G (50 mM) for 5 min follow e dby w ashout produc e d a sta b le re duction of syna ptic strength(72.8 % ± 2.0 % of b ase line , m e an ± S E M) (F igure 1A). Syna pticstimulation using the p a ire d-pulse low-fre quency stimulation(P P-L F S) proto c ol in the presenc e of the N M D A re c e ptor anta g-onist D-A PV (50 mM) resulte d in a similar sta b le re duction of syn-a ptic strength to 79.9 % ± 2.1 % of b ase line (F igure 1 B). In Arc/Arg3.1 K O slic es, tre atm ent w ith D H P G (92.1 % ± 3.7 % of b ase-line , p < 0.001 c om p are d to litterm ate W T c ontrols by unp a ire dtwo-ta ile d Stud ent’s t test) or P P-L F S (94.3 % ± 2.1 % of b ase line ,p < 0.0001) fa ile d to evok e robust LT D , a lb e it there is a slight re-sidua l LT D in Arc/Arg3.1 K O slic es (p = 0.09 for D H P G -LT D andp = 0.03 for P P-L F S LT D by p a ire d t test). The residua l LT D sug-g ests that an Arc/Arg3.1-ind e p end ent p athw ay a lso c ontributesto m G luR-LT D . The imm e d iate short-term syna ptic d e pressionduring the induction p eriod w ith D H P G and imm e d iate ly follow-ing the P P-L F S proto c ol w as not signific antly d ifferent b etw e enWT and K O mic e (F igures 1A and 1 B). F urthermore , m G luR-LT D induc e d by higher c onc entration of D H P G (100 mM) w asa lso im p a ire d in Arc/Arg3.1 K O slic es (88.5 % ± 7.4 % of b ase linefor Arc/Arg3.1 K O slic es; 64.4 % ± 2.6 % of b ase line for WTslic es, p < 0.01), ind ic ating that the re quire m ent for Arc/Arg3.1do es not d e p end on a sp e c ific rang e of m G luR a ctivation(F igure S1 C ).
m G l u R - D e p e n d e n t A M P A R e c e p t o r E n d o c y t o s i sR e q u ir e s A r c / A r g 3 . 1m G luR1/5 a ctivation results in a ra p id and susta ine d loss of sur-fa c e A M P ARs that und erlies syna ptic d e pression (Snyd er et a l.,2001). S inc e Arc/Arg3.1 K O mic e have d efic ient m G luR-LT D , w ed ire ctly teste d whether Arc/Arg3.1 is re quire d for m G luR-d e p en-d ent A M P AR endo cytosis. D H P G (50 mM) w as a p p lie d to DIV 14–21 prim ary hip po c a m p a l neurons for 5 min follow e d by w ashout,and surfa c e and interna liz e d A M P ARs w ere m e asure d 15 or60 min after D H P G a p p lic ation. In WT cultures, D H P G resulte d ina signific ant loss of surfa c e G luR1 at 15 and 60 min as c om p are dw ith untre ate d cultures (F igures 1 C 1–1 C 3 and 1 C 7). H ow ever,G luR1 surfa c e leve ls w ere unchang e d after D H P G a p p lic ationin Arc/Arg3.1 K O neurons (F igure s 1 C 4–1 C 6 and 1 C 7). WT cul-tures e xhib ite d a signific ant incre ase in interna liz e d G luR1 at15 min (F igures 1D1–1D3 and 1D7). Arc/Arg3.1 K O neuronsd id not e xhib it an incre ase in interna liz e d re c e ptors after D H P Ga p p lic ation (F igures 1D4–1D6 and 1D7). Thus, w e c onc lud ethat Arc/Arg3.1 is re quire d for ra p id , m G luR-d e p end ent A M P ARendo cytosis.
m G l u R In d u c e s R a p i d T r a n s l a t i o n o f P r e e xi s t i n gA r c / A r g 3 . 1 m R N AIf Arc/Arg3.1 p lays a d ire ct role in m G luR-LT D , w e antic ip ate dthat its prote in leve l should b e a cute ly re gulate d in d endrites.Therefore , Arc/Arg3.1 prote in e x pression w as e x a mine d by im-muno cyto che mistry in DIV14 hip po c a m p a l cultures. The b asa lleve l of Arc/Arg3.1 prote in w as low but incre ase d signific antlyin both the som a and d endrites during the 5 min incub ationw ith D H P G (50 mM) (F igure 2A). The incre ase of Arc/Arg3.1
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 71
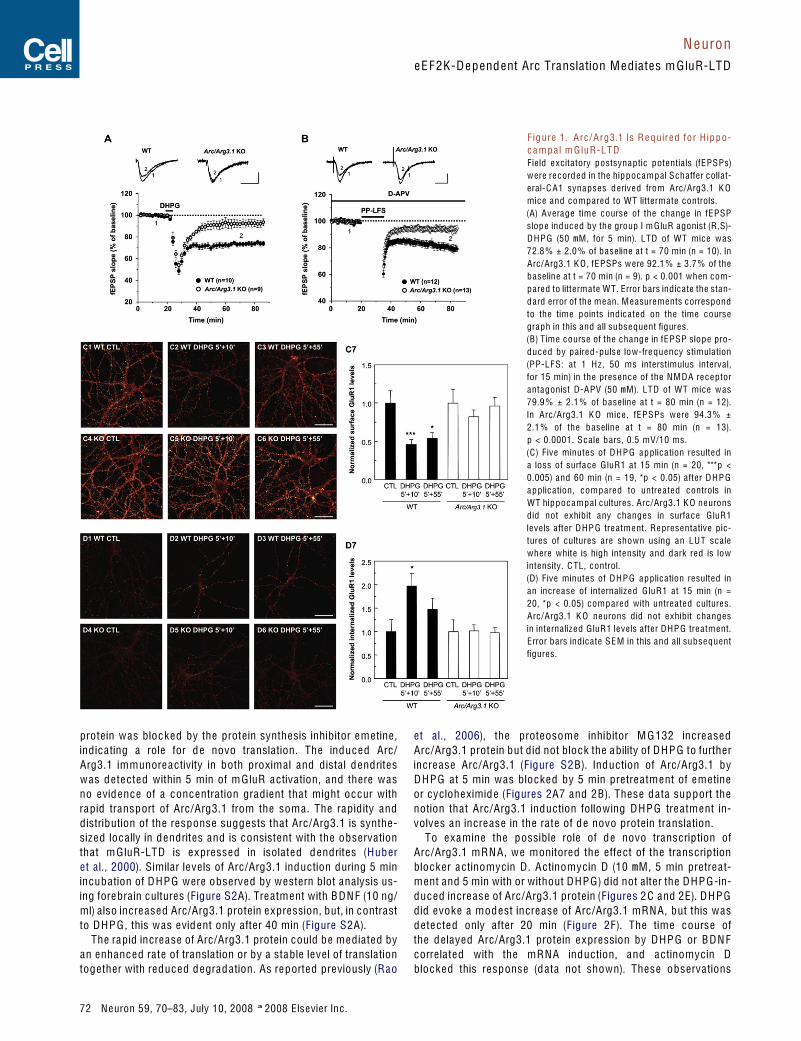
prote in w as b lo c k e d by the prote in synthesis inhib itor e m etine ,ind ic ating a role for d e novo translation. The induc e d Arc/Arg3.1 immunore a ctivity in both proxim a l and d ista l d endritesw as d ete cte d w ithin 5 min of m G luR a ctivation, and there w asno evid enc e of a c onc entration gra d ient that might o c cur w ithra p id transport of Arc/Arg3.1 from the som a . The ra p id ity andd istribution of the response sugg ests that Arc/Arg3.1 is synthe-siz e d lo c a lly in d endrites and is c onsistent w ith the observationthat m G luR-LT D is e x presse d in isolate d d endrites (Hub eret a l., 2000). S imilar leve ls of Arc/Arg3.1 induction during 5 minincub ation of D H P G w ere observe d by w estern b lot ana lysis us-ing fore bra in cultures (F igure S2A). Tre atm ent w ith B D N F (10 ng/ml) a lso incre ase d Arc/Arg3.1 prote in e x pression, but, in c ontrastto D H P G , this w as evid ent only after 40 min (F igure S2A).
The ra p id incre ase of Arc/Arg3.1 prote in c ould b e m e d iate d byan enhanc e d rate of translation or by a sta b le leve l of translationtog ether w ith re duc e d d e gra d ation. As re porte d previously (R a o
et a l., 2006), the prote osom e inhib itor M G 132 incre ase dArc/Arg3.1 prote in but d id not b lo c k the a b ility of D H P G to furtherincre ase Arc/Arg3.1 (F igure S2 B). Induction of Arc/Arg3.1 byD H P G at 5 min w as b lo c k e d by 5 min pretre atm ent of e m etineor cyc lohe ximid e (F igures 2A7 and 2 B). These d ata sup port thenotion that Arc/Arg3.1 induction follow ing D H P G tre atm ent in-volves an incre ase in the rate of d e novo prote in translation.
To e x a mine the possib le role of d e novo transcription ofArc/Arg3.1 mR N A , w e monitore d the effe ct of the transcriptionb lo c k er a ctinomyc in D . A ctinomyc in D (10 mM , 5 min pretre at-m ent and 5 min w ith or w ithout D H P G) d id not a lter the D H P G -in-duc e d incre ase of Arc/Arg3.1 prote in (F igures 2 C and 2 E). D H P Gd id evok e a mod est incre ase of Arc/Arg3.1 mR N A , but this w asd ete cte d only after 20 min (F igure 2 F). The tim e c ourse ofthe d e laye d Arc/Arg3.1 prote in e x pression by D H P G or B D N Fc orre late d w ith the mR N A induction, and a ctinomyc in Db lo c k e d this response (d ata not shown). These observations
F ig ur e 1. A rc / A r g3.1 Is R e q u ir e d f or H ip p o -c a m p a l m G lu R -L T DF ie ld e x c itatory postsyna ptic potentia ls (fE P S Ps)w ere re c ord e d in the hip po c a m p a l S ch affer c ollat-era l- C A1 syna pses d erive d from Arc/Arg3.1 K Omic e and c om p are d to WT litterm ate c ontrols.(A) Avera g e tim e c ourse of the chang e in fE P SPslop e induc e d by the group I m G luR a gonist (R,S)-D H P G (50 mM , for 5 min). LT D of W T mice w as72.8 % ± 2.0 % of b ase line at t = 70 min (n = 10). InArc/Arg3.1 K O , fE P SPs w ere 92.1 % ± 3.7 % of theb ase line at t = 70 min (n = 9). p < 0.001 when c om-p are d to litterm ate WT. Error b ars ind ic ate the stan-d ard error of the m e an. M e asure ments c orrespondto the time points ind ic ated on the time c oursegraph in this and a ll subse quent figures.(B) Tim e c ourse of the chang e in fE P S P slop e pro-duc e d by p a ire d- pulse low-fre quency stimulation(P P-L F S: at 1 H z , 50 ms interstimulus interva l,for 15 min) in the presenc e of the N M D A re c e ptoranta gonist D-A PV (50 mM). LT D of WT mic e w as79.9 % ± 2.1 % of b ase line at t = 80 min (n = 12).In Arc/Arg3.1 K O mic e , fE P S Ps w ere 94.3 % ±2.1 % of the b ase line at t = 80 min (n = 13).p < 0.0001. S c a le b ars, 0.5 mV/10 ms.( C ) F ive minute s of D H P G a p p lic a tion resulte d ina loss of surfa c e G luR1 at 15 min (n = 20, ***p <0.005) and 60 min (n = 19, *p < 0.05) after D H P Ga p p lic ation, c om p are d to untre ate d c ontrols inWT hip po c a m p a l cultures . Arc/Arg3.1 K O neuronsd id not e xhib it any chang es in surfa c e G luR1leve ls after D H P G tre atm e nt. R e presentative p ic-tures of cultures are shown using an LU T sc a lewhere whit e is high inte nsity and d ark re d is lowintensity. C TL, c ontrol.(D) F ive minute s of D H P G a p p lic ation resulte d inan incre a se of interna liz e d G luR1 at 15 min (n =20, *p < 0.05) c om p are d w ith untre ate d cultures.Arc/Arg3.1 K O neurons d id not e xhib it chang esin interna liz e d G luR1 leve ls aft er D H P G tre atm ent.Error b ars ind ic ate S E M in this and a ll subse quentfigures.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
72 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

sugg est that the ra p id incre ase in d e novo translation re quiresArc/Arg3.1 mR N A that is present in neurons prior to D H P Gstimulation, while the d e laye d Arc/Arg3.1 e x pression is c oup le dto d e novo transcription. W e note that Arc/Arg3.1 mR N A is d e-te cte d in d endrites of unstimulate d culture d neurons (G iorgiet a l., 2007), and w e d ete ct Arc/Arg3.1 mR N A in stratum ra d ia-tum of the hip po c a m p a l C A1 re gion from hom e-c a g e d mic e(F igure S3).
L o w D o s e C y c l o h e x i m i d e C a n In c r e a s eA r c / A r g 3 . 1 P r o t e i n E x p r e s s i o nIn e x a mining the dose d e p end enc e of cyc lohe ximid e’s a ctions,w e note d that the leve l of Arc/Arg3.1 prote in ra p id ly incre ase dwhen neurons are tre ate d w ith low doses (F igure 2D). F or e x a m-p le , 100 nM cyc lohe ximid e incre ase d Arc/Arg3.1 prote in w ithin10 min. E ven at these low doses, cyc lohe ximid e effe ctive ly re-duc e d g enera l prote in synthesis. 100 nM cyc lohe ximid e re duc e dthe tota l inc orporation of 35S-la b e le d m ethionine and cyste ineinto T C A pre c ip itant to 60 % . Previous stud ies have note d the
p ara doxic a l a ction of low-dose cyc lohe ximid e to incre ase thesynthesis of sp e c ific prote ins and rationa liz e d this a ction by sug-g esting that glob a l re duction of e long ation c an incre ase theava ila b ility of fa ctors that are re quire d for translation initiationof sp e c ific transcripts that are poorly initiate d und er c ontrol c on-d itions (F ernand e z et a l., 2005; G upta and O no, 1997; P erlm anand F e ld m an, 1982; S che etz et a l., 2000; W a ld en and Tha ch,1986). This notion c ontribute d to our ana lysis of the e E F 2 p ath-w ay (b e low), sinc e a ctivate d e E F 2 inhib its e long ation and c anp ara doxic a lly incre ase translation of c erta in mR N As ( C hotineret a l., 2003; S che etz et a l., 2000).
e E F 2 K P h y s i c a lly A s s o c i a t e s w i t h H o m e ra n d G r o u p I m G l u R sH om er prote ins b in d group I m G luRs and p lay a role in th e irsigna ling b y a lso b ind ing signa ling p artners , in c lud ing IP3R(Tu e t a l., 1998). H o m er prote ins b ind tw o know n s e q ue nc emotifs: P Pxx F (typ e 1) and LP S S P S S (typ e 2) (Yua n e t a l.,2003). Wh en w e s e arch e d for c a nd id at e H o m er- b in d ing
F ig ur e 2. A r c / A r g3.1 Pro t e in Is R a p id ly Sy n-t h e s i z e d b y G r o u p I m G lu R A c tiv a tio n(A) Stimulation of hip po c a m p a l neurons w ithD H P G (50 mM) for 5 min incre a se d Arc/Arg3 .1 im-munore a ctivity in both c e ll body (1.34 ± 0.063 ofuntre ate d som a , n = 13) and d endrit es (1.58 ±0.095 of untre ate d d endrit es, n = 38). The ra p id in-cre ase of Arc/Arg3.1 w as b lo c k e d by the prote insynthesis inhib itor e m etine (10 ng/ml, 10 min).(B) H igh-dose cyc lohe ximid e ( C HX, 50 mM , tota l10 min: 5 min pretre atm ent and 5 min w ith or w ith-out D H P G) b lo c k e d the induction of Arc/Arg3 .1by D H P G (5 min).( C ) Transcription inhib itor a ctino myc in D (A ctD:10 mM , 5 min pretre atm ent and 5 min w ith orw ithout D H P G) d id not b lo c k the induction ofArc/Arg3.1 by D H P G (5 min).(D) Low-dose C HX incre ased the leve l of Arc/Arg3.1prote in. N eurons w ere treate d w ith vehic le or vari-ous doses of C HX for 10 min. Tota l prote in synthe-sis w as m e asured by c ounting the inc orporation of35S m ethionine and cyste ine in T C A pre cip itant.(E) Statistic a l ana lysis of w estern b lots. F iv eminute tre atm ent of D H P G signific antly incre ase dthe leve l of Arc/Arg3.1. Inhib ition of ne w prot e insynthesis by high dose of cyc lohe ximid e not onlyb lo c k e d the induction of Arc/Arg3.1 prot e in buta lso slightly d e cre ase d the leve l of Arc/Arg3 .1upon stimulation w ith D H P G . Inhib ition of tran-scription by a ctinomyc in D d id not affe ct the leve lof Arc/Arg3.1. Low-dose C HX (50–100 nM , 5 minpretre atm ent and 5 min w ith or w ithout D H P G)incre ase d the leve l of Arc/Arg3.1, which w as notfurther induc e d by D H P G . *p < 0.05, **p < 0.01.(F) The leve l of Arc/Arg3 .1 mR N A w as m e asure dusing re a l-tim e RT-P C R. Stimulation of neuronsw ith B D N F (10 ng/ml) and forskolin (50 mM) in-duc e d the leve l of Arc/Arg3.1 mR N A 40 min and20 min after stimulation, resp e ctive ly. D H P Gslightly incre ase d the leve l of Arc/Arg3.1 mR N Aat 20 and 40 min after stimulation.*p < 0.05, **p < 0.01, ***p < 0.005.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 73

m ole cule s (htt p://us .e x p a sy .org/c gi- b in/s c anprosite), w efoun d th at e E F 2 K p oss esse s a typ e 2 H om er-b ind ing motif(F igure 3A). e E F 2K is a high ly c ons erv e d enz ym e that phos-phorylat e s and re gulat es its only know n substrat e , e E F 2 (Ry a-z anov e t a l. , 1988). The N-term ina l h a lf of e E F 2K c ont a insa C a 2+ -c a lmod ulin ( C a M) b ind ing sit e , which is re quire d forits a ctiva tion , and an a kinas e do m a in. Th e C -termin a l ha lf ofe E F 2K func tions a s a targ e ting dom a in tha t is re quire d for itto phos phoryla te e E F 2. A link er re gion b etw e en th e N an d Cterminus inc lud es the puta tive H o m er-b in d ing sit e an d is phos-phorylat e d by multip le signa ling k ina s es , inc lud ing PI3 K/m T O R/S 6K , M A P K , and P K A (Brow ne and Prou d , 2002).
e E F 2K and H om er c oimmunopre c ip itate d (c o-IP) fromH E K293T c e lls (F igure 3 B). The E VH1 dom a in of H om er is re-quire d to b ind e E F 2K , and mutation of a critic a l b ind ing surfa c efor polyproline lig ands (H om er3 G 91N) d isrupte d b ind ing. As an-tic ip ate d by the c onservation of the ir E VH1 dom a ins, H om er 1,-2, and -3 b ind e E F 2K (d ata not shown).
F ig ur e 3. e E F 2 K B in d s H o m e r a n d m G lu R1/5(A) S che m atic d ia gra m of e E F 2K . The N terminusof e E F 2K c onta ins a C a 2+/c a lmo dulin ( C a M) b ind-ing motif and an a -kinase dom a in. The C -term ina le E F 2 targ eting dom a in, which re cruits the sub-strate , e E F 2, is link e d to the hyp erphosphorylate dinterna l re gion. Putative H om er b ind ing site isshown a bove the d ia gra m.(B) C oimmunopre c ip itation (c o-IP) of e E F 2K andH om er. H A-ta g g e d (H A-) e E F 2K w as c o e x presse dw ith myc-ta gg e d W T, W27A , or G 91N H om er3 inH E K293T c e lls, and IP w as p erform e d w ith anti-myc antibody. H A-e E F 2K c o-IP e d w ith WT orW27A H o m er 3 w as c o e x presse d , but not w ithG 91N H om er.( C ) m G luR5 incre ases the intera ction of e E F 2K andH om er. H A-e E F 2K w as transfe cte d w ith or w ithoutH A-m G luR5. IP w as p erform e d by anti-H om er2antibody, which IP e d endog enous H om er2 pro-te in. W estern b lot w as p erform e d using anti-H Aantibody. C o-IP of H A-e E F 2K w as incre a se dwhen m G luR5 w as c o e x presse d .(D) e E F 2K c o-IPs w ith m G luR5. H E K293T c ells w eretransfecte d w ith H A-e E F2K w ith or w ithout H A-m G luR5, and lysates w ere IP ed w ith anti-m G luR5antibody and blotte d w ith anti-H A antibody. e E F 2Kc o-IP ed only when m G luR w as c oe xpresse d. S am-ples w ere boiled b efore loa ding to a ggre gate andseparate m G luR5 monomer from e E F 2K .(E) m G luR1 c o-IP s w ith e E F 2K . H E K293T c e llsw ere transfe cte d w ith m G luR1 and e E F 2K , and ly-sates w ere IP e d w ith myc A b . S a m p le s w ere notboile d to show the mono m er of m G luRs.(F and G) m G luR2 and m G luR4 do not c o-IPw ith e E F 2K .
W e e x a mine d c ond itions that mightre gulate H om er-e E F 2K b ind ing andfound that c o e x pression of m G luR5strongly enhanc e d b ind ing (F igure 3 C ).M ore over, e E F 2K w as found to intera ctw ith group I m G luRs ind e p end ently of
H om er. The intera ction of e E F 2K and group I m G luRs w asobserve d even when H om er w as not c o e x presse d (F igure 3Dand 3 E), and e E F 2K bound to m G luR5 mutants that do notb ind H om er (F igures S4A and S4 B). e E F 2K a lso c o-IP e d w ithm G luR1 (F igure 3E), another m e m b er of group I m G luRs, butnot w ith other m G luRs, inc lud ing m G luR2 and m G luR4(F igure 3 F and 3 G).
To id entify re gions of e E F 2K that are critic a l for b ind ing H om erand m G luR, w e preform e d a d e letion ana lysis of e E F 2K (F iguresS4 C and S4D). The link er re gion of e E F 2K , which inc lud es thetyp e 2 H om er lig and , a p p e ars essentia l for b ind ing H om er, sinc eN-termina l fra gm ents that inc lud e this re gion b ind , while C -termi-na l fra gm ents or N-termina l fra gm ents that do not inc lud e thelink er re gion do not b ind . e E F 2K b ind ing to m G luR5 a p p e ars tob e more c om p le x, sinc e both N- and C -termina l fra gm ents ofe E F 2K b ind m G luR5 (F igure S4 C ). These d ata sugg est thatm G luR, H om er, and e E F 2K asse m b le by multip le intera ctionsinto a tertiary c om p le x.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
74 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

T h e In t e r a c t i o n o f e E F 2 K w i t h m G l u R Is D y n a m i ca n d Is M o d u l a t e d b y C a 2 + a n d m G l u R A c t ivi t yThe kinase a ctivity of e E F 2K is known to b e re gulate d by C a2+
via its C a2+- C a M-b ind ing dom a in (N a irn and P a lfrey, 1987;Rya z anov, 1987). To test whether C a2+ modulates the m G luR5-e E F 2K b ind ing, c o-IP e x p erim ents w ere p erform e d using lysatesfrom c otransfe cte d H E K293T c e lls in the presenc e of d efine dc onc entrations of fre e C a 2+ (F igure 4A). C o-IP w as robust at[ C a2+] less than 1 mM but m ark e d ly d e cre ase d at c onc entrations>10 mM . m G luR5 b ind ing to a C -termina l fra gm ent of e E F 2K thatla c ks the C a M-b ind ing dom a in but reta ins b ind ing to m G luR5w as not inhib ite d by [ C a 2+] (F igure S4 E). These results ind ic atethat [ C a2+] c an modulate the intera ction of group I m G luRsw ith e E F 2K and sugg est a role for C a M b ind ing to e E F 2K .
W e use d e E F 2K K O mic e in our ana lysis of m G luR-e E F 2Kb ind ing. e E F 2K K O mic e w ere via b le and fertile (Rya z anov,2002) and show e d the antic ip ate d a bsenc e of phosphorylate de E F 2 (Thr56) (F igure 4 B). The leve ls of severa l syna ptic prote insw ere not a ltere d in the hip po c a m pus of K O mic e (F igure 4 B).Syna ptoneurosom es from fore bra ins of W T and e E F 2K K Omic e w ere pre p are d and stimulate d w ith D H P G for 20 min.C o-IP e x p erim ents using anti-e E F 2K antibody c onfirm e d thatnative m G luR5 asso c iate d w ith e E F 2K (F igure 4 C ). The c o-IPof m G luR5 w as re duc e d when syna ptoneurosom es w ere stimu-late d w ith D H P G . Intera ction of endog enous m G luR5 ande E F 2K w as a lso re duc e d upon D H P G stimulation of culture dneurons (F igure S4 F). W e c onc lud e that m G luR and e E F 2Kasso c iate in vivo and that the ir intera ction is re duc e d bym G luR a ctivation.
G r o u p I m G l u R s D y n a m i c a lly R e g u l a t et h e P h o s p h o r yl a t i o n o f e E F 2A ctivate d e E F 2K se le ctive ly phosphorylates e E F 2 (Rya z anovet a l., 1988). To assess whether m G luR a ctivates this p athw ayin c ond itions that evok e LT D , w e monitore d the leve l of phos-pho-e E F 2 in hip po c a m p a l slic es of e ither WT or e E F 2K K Omic e using the sa m e stimulus p ara m eters that induc e m G luR-LT D . A ctivation of m G luR incre ase d the phosphorylation ofe E F 2 in the stratum pyra mid a l (s.p .), and stratum ra d iatum (s.r.)of the hip po c a m p a l C A1 re gion w ithin 5 min (F igure 5A). B y30 min after w ashout of D H P G , the leve l of phospho-e E F 2 w asre duc e d to prestimulation leve l. N o phosphorylation of e E F 2w as d ete cte d in e E F 2K K O slic es. The transient induction ofphospho-e E F 2 by D H P G w as c onfirm e d by w estern b lot ana lysisin hip po c a m p a l slic es (F igure S5A).
To further e x a mine d endritic lo c a liz ation of e E F 2K a ctivity,DIV14 neurons w ere stimulate d w ith D H P G for 5 min andsta ine d w ith phospho-e E F 2 and P S D95, a m ark er for e x c itatorysyna pses (F igure 5 B). Phospho-e E F 2 w as present in d endriticshafts and the c e ll body. Phospho-e E F 2 a lso show e d a d istinctpuncta l d istribution in sp ines that c olo c a liz e d w ith P S D95.Sta ining w as a bsent in e E F 2K K O cultures (d ata not shown).This result is c onsistent w ith a previous re port that translationa lre gulators, inc lud ing e E F 2K , are enriche d in syna ptic fra ctions(Asa ki et a l., 2003).
Phosphoryla tion of e E F 2 is known to inhib it translationa le long ation. Therefore , w e e x a mine d the pre d iction that glob a lprote in translation might b e transiently re duc ed c o-inc ident w iththe transient incre ase of phospho-e E F 2. Stimulation of neuronsw ith D H P G for 5 min transiently d e cre ase d the inc orporation of35S a mino a c ids into T C A pre c ip itants, and this effe ct w as re-verse d 20 min after w ashout of D H P G (F igure S5 B). A previousstudy re porte d that D H P G ra p id ly incre ase d prote in synthesisin syna ptoneurosom es (W e iler et a l., 2004). W e observe d thatD H P G d id not induc e p-e E F 2 in syna ptoneurosom es (d ata notshown), and it is possib le that the e E F 2-d e p end ent translationa lm e chanism is not m a inta ine d in brok en c e ll pre p arations.
R a p i d D e N o v o A r c / A r g 3 . 1 T r a n s l a t i o n Is S e l e c t iv e lyA b s e n t i n e E F 2 K K O N e u r o n sArc/Arg3.1 e x pression w as e x a mine d in DIV14 fore bra in neuro-na l cultures pre p are d from W T and e E F 2K K O mic e . The
F ig ur e 4. D y n a m i c In t e r a c tio n o f e E F 2 K a n d m G lu R5(A) C a lc ium d isso c iates e E F 2K from m G luR5. H E K293T c e lls w ere transfe cte dw ith H A-e E F 2K w ith or w ithout myc-m G luR5, and c e lls w ere harveste d w ith ly-sis buffer w ithout c a lc ium or c onta ining various c onc entrations of fre e c a lc ium .C a lmodulin ( C a M) (25 mg/ml) w as a lso a d d e d to the lysis buffer as ind ic ate d .B ind ing w as d e cre ase d at [ C a2+] higher than 10 mM .(B) Phospho-e E F 2 w as not d ete cte d in the hip po c a m p us of e E F 2K K O , whilethe leve l of tota l e E F 2, G luR1, G lur2/3, m G luR5, a - C a M KII, Arc/Arg3.1, and a c-tin w as not a ltere d in e E F 2K K O mic e c om p are d to WT litterm ate c ontro ls.( C ) Syna ptone urosom es, pre p are d from the fore bra in of e E F 2K K O and W Tmic e , w ere stimulate d w ith vehic le or D H P G for 20 min. Syna ptoneurosom esw ere then lys e d and immunopre c ip itate d w ith anti-e E F 2K antib ody. m G luR5c o-IP e d w ith e E F 2K only in WT sa m p les. Stimulation of syna ptoneurosom esw ith D H P G d e cre ase d the c o-IP of m G luR5.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 75

ste a dy-state e x pression of Arc/Arg3.1 prote in w as id entic a lin WT and e E F 2K K O neurons; how ever, the incre ase inArc/Arg3.1 prote in 5 min after D H P G in WT neurons w as a bsentin e E F 2K K O neurons in both b io che mic a l (F igures 5 C and 5D)and immuno cyto che mic a l assays (F igure S6). B y c ontrast,Arc/Arg3.1 prote in w as induc e d to the sa m e e xtent in W T ande E F 2K K O neurons 60 min after D H P G stimulation. Arc/Arg3.1mR N A w as id entic a l in WT and e E F 2K K O neurons prior to a p p li-c ation of D H P G and incre ase d id entic a lly at 40 min after stimu-lation in both W T and e E F 2K K O neurons (F igure 5E). A c c ord-ingly, the la c k of ra p id induction of Arc/Arg3.1 prote in in thee E F 2K K O neurons is not due to re duc e d Arc/Arg3.1 mR N Ae x pression. W e a lso note that m G luR signa ling that is re quire dfor induction of Arc/Arg3.1 mR N A and the d e laye d incre ase ofArc/Arg3.1 prote in are inta ct in e E F 2K K O neurons. M ore over,
F ig ur e 5. R a p id In d u c tio n o f A r c / A r g3.1 b yG r o u p I m G lu R s Is D e p e n d e n t o n e E F 2 K(A) H ip po c a m p a l slic es w ere pre p are d from WTand e E F 2K K O mic e and w ere stimulate d w ithD H P G for 5 min. phos pho-e E F 2 (p-e E F 2, re d) inare a C A1 w as incre ase d by D H P G w ithin 5 minand d e c line d by 30 min follow ing w ashout. S p e c-ific ity of phospho-e E F 2 w as c onfirm e d by sta iningof e E F 2K K O slic es. s.p ., stratum pyra mid a l; s.r.,stratum ra d iatum.(B) C ulture d hip po c a m p a l neurons w ere tre ate dw ith D H P G for 5 min and sta ine d w ith phospho-e E F 2 (re d) and P S D95 (gre en) antibod ie s on DIV14.phospho-e E F 2 sho w e d puncta l d istribution in d en-dritic sp ines and d endritic shafts. phospho-e E F 2 insp ines c olo c a liz e d w ith P S D95 (arrows). (B 2), (B 3),and (B 4) are enlarg e d im a g es of the re ctangularre gion of (B 1).( C and D) m G luR-d e p end ent ra p id synthesis ofArc/Arg3.1 is a bsent in e E F 2K K O neurons. N eu-rons from the fore bra ins of W T or e E F 2K K O mic ew ere culture d for DIV14 and tre ate d w ith D H P G(50 mM , 5 min). Phosphorylation of e E F 2 w as und e-te cta b le in e E F 2K K O neurons. N o d iffere nc e in theleve l of m G luR5 w as observe d b etw e e n WT ande E F 2K K O neurons. An arrowhe a d ind ic ates a non-sp e c ific b and . p va lues w ere obta ine d by p a ire d ttest c om p aring b asa l and drug-tre ate d leve ls.p va lu es for c om p arison of WT and e E F 2K K Omic e w ere obta ine d by Stud ent’s t test. *p < 0.05,**p < 0.01, n = 8. Error b ars are S E M .(E) Arc/Arg3 .1 mR N A e x pression is not a lt ere d ine E F 2K K O neurons. The leve l of Arc/Arg3.1mR N A w as m e asure d in WT and e E F 2K K O neu-rons follow ing the stimulation w ith D H P G .(F) Low-dose cyc lohe xim id e ( C HX) incre a ses Arc/Arg3.1 prote in e x pression. C ulture d e E F 2K K Oneurons w ere tre ate d w ith ind ic ate d doses ofC HX for 10 min. *p < 0.05, n = 8.
Arc/Arg3.1 prote in e x pression is id entic a lin the hip po c a m pus of W T and e E F 2K K Omic e (F igure 4 B), ind ic ating that e E F 2Kis not re quire d for b asa l e x pression ofArc/Arg3.1 prote in in vivo.
If the fa ilure of D H P G to induc e ra p id synthesis of Arc/Arg3.1 prote in in the e E F 2K K O neurons is due to a se le ctiveinterruption of the a ction of phospho-e E F 2, w e pre d icte d thata low dose of cyc lohe ximid e , which do es not re quire e E F 2Kor phopho-e E F 2 to inhib it the e long ation ste p , should induc ethe synthesis of Arc/Arg3.1 prote in in e E F 2K K O neurons.Tre atm ent of DIV14 e E F 2K K O neurons w ith low-dose cyc lo-he ximid e (50 nM and 100 nM) incre ase d the leve l ofArc/Arg3.1 prote in in e E F 2K K O neurons (F igure 5 F), similarto W T neurons (F igure 2D). H igh-dose cyc lohe ximid e (>1 uM)d id not induc e Arc/Arg3.1 in e ither WT and e E F 2K K O neurons.The a b ility of low-dose cyc lohe ximid e to rescue ra p idArc/Arg3.1 induction ind ic ates that m e chanisms that m e d iatera p id Arc/Arg3.1 translation subse quent to inhib ition ofe long ation are inta ct in e E F 2K K O neurons.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
76 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

m G l u R - L T D a n d P P - L F S L T D A r e S e l e c t iv e ly A b s e n ti n e E F 2 K K O H i p p o c a m p a l S li c e sThe role of e E F 2K in p lastic ity of the S chaffer c ollatera l- C A1syna pse w as teste d using a cute hip po c a m p a l slic es. B ase linem e asures of syna ptic strength and presyna ptic function w erenot a ltere d in the e E F 2K K O slic es (F igure S7). H ow ever, LT Dinduc e d by e ither P P-L F S (97.5 % ± 2.4 % of b ase line) orD H P G (108.7 % ± 3.6 % of b ase line) w as im p a ire d in the e E F 2KK O slic es (F igures 6A and 6D). The imm e d iate short-term syna p-tic d e pression follow ing D H P G stimulation w as id entic a l in WTand e E F 2K K O slic es; how ever, syna ptic strength returne d tone ar b ase line leve ls in the e E F 2K K O slic es. S imilarly, syna ptictransmission returne d to ne ar b ase line leve ls w ithin 10 min ofc om p letion of the P P-L F S proto c ol. In c ontrast to the m ark e dd efic it of m G luR-d e p end ent LT D , N M D AR-d e p end ent LT D w asid entic a l in tim e c ourse and sta b ility in slic es d erive d from
e E F 2K K O mic e (72.7 % ± 2.2 % of b ase line) c om p are d to WTmic e (73.1 % ± 3.4 % of b ase line) (F igure 6 B). LTP w as a lso pre-serve d (F igure 6 C ). LTP of S chaffer c ollatera l- C A1 syna pses w asinduc ed by four trains of high-fre quency stimulation with an inter-tra in interva l of 3 s. In WT slices, fE PS P was incre ased to 171.5 % ±13.4 % of b ase line imm e d iate ly after stimulation (t = 30 min) andsusta ine d at the leve l of 138.4 % ± 7.7 % of b ase line at t = 175 min.These stimulus p ara m eters are re porte d to evok e a form ofsyna ptic p lastic ity that re quires d e novo prote in synthesis form a intenanc e long er than 60 min and is referre d to as lateLTP (L-LTP) (K e lleher et a l., 2004). In slic es pre p are d frome E F 2K K O mic e , the initia l induction w as 204.6 % ± 8.9 % ofb ase line at t = 30 min, and this w as susta ine d for 3 hr after stim-ulation (200.1 % ± 11.9 % of b ase line at t = 175 min) (F igure 6 C ).The m a gnitud e of LTP w as signific antly gre ater in e E F 2K K O thanW T after 30 min of induction (p < 0.005). These results ind ic at e
F ig ur e 6. m G lu R -L T D Is Im p a ir e d in H ip p o -c a m p a l S li c e s D e riv e d fr o m e E F 2 K K O M i c efE P S Ps w ere re c ord e d in the hip po c a m p a l C A1 re-gion of slic es d erive d from e E F 2K K O mic e andc om p are d to W T litterm at e c ontrols.(A) Tim e c ourse of the chang e in fE P S P slop e pro-duc e d by p a ire d-pulse low-fre quenc y stimulation(P P-L F S: at 1 H z , 50 ms interstimulus interva l, for15 min) in the prese nc e of D-A PV (50 mM). LT D ofWT mic e w as 77.0 % ± 2.1 % of b ase line at t = 75min (n = 13). In e E F 2K K O mic e , fE P S Ps w ere97.5 % ± 2.4 % of b ase line t = 75 min (n = 15)(p < 0.0001).(B) Tim e c ourse of the chang e in fE P S P slop e bylow-fre quenc y stimulation (L F S: 1 H z for 15 min).This form of N M D A R-d e p end e nt LT D w as not a l-tere d in e E F 2K K O hip po c a m p a l slic es (72.7 % ±2.2 % of b ase line at t = 75 min, n = 9) c om p are dto WT (73.1 % ± 3.4 % of b ase line at t = 75 min,n = 7) (p > 0.5).( C ) Late-ph ase of LTP w as induc e d by four stimu-lus tra ins (100 H z e a ch) w ith an intertra in interva l of3 s. In W T, fE P S Ps w ere incre ase d to 171.5 % ±13.4 % of b ase line imm e d iate ly after stimulation(t = 30 min) and w ere susta ine d at the leve l of138.4 % ± 7.7 % of b ase line at t = 175 min (n = 6).H ow ever, in e E F 2K K O , the initia l LTP (204.6 % ±8.9 % of b ase line at t = 30 min) w as m a int a ine dfor 3 hr after stimulation (200.1 % ± 11.9 % of b ase-line at t = 175 min, n = 5). LTP w as signific antlygre ater in slic es d erive d from e E F 2K K O mic ec om p are d to those from W T mic e at this tim e point(p < 0.005).(D) Avera g e tim e c ourse of the chang e in fE P S Pslop e ind uc e d by D H P G (50 mM , for 5 min). LT Dof WT mic e w as 64.7 % ± 5.2 % of b ase line at t =90 min (n = 7). In e E F 2K K O mic e , LT D w as signifi-c antly im p a ire d (108.7 % ± 3.6 % of b ase line at t =90 min, n = 8). Tre atm ent w ith low-dose cyc lohe x-imid e (LD- C HX, 50–75 nM) for 10 min starting from5 min prior to D H P G restore d D H P G -LT D in e E F 2KK O (75.7 % ± 7.4 % , n = 5). In WT mic e , tre atm e ntw ith LD- C HX d id not a lter the e x pression of LT D(69.0 % ± 2.6 % , n = 5). p < 0.001 when e E F 2K K OD H P G only w as c om p are d to e E F 2K K O LD- C HX +D H P G , WT D H P G only, or WT LD- C HX + D H P G .S c a le b ars, 0.5 mV/10 ms.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 77

that e E F 2K K O d isrupts m G luR-LT D but do es not a lter N M D AR-d e p end ent LT D or e arly LTP . The a p p arent enhanc e m ent of late-phase LTP d eserves further study.
O ur propose d m e chanism for the m G luR-LT D d efic it inthe e E F 2K K O slic es is link e d to fa ilure to ra p id ly translateArc/Arg3.1. S inc e low-dose cyc lohe ximid e induc e d Arc/Arg3.1synthesis and d id not d e p end on phospho-e E F 2, w e e x a mine dthe possib ility that cyc lohe ximid e c ould rescue m G luR-LT D inslic es from e E F 2K K O mic e . W e found that a 10 min e x posureto 50–75 nM cyc lohe ximid e (low-dose C HX: LD- C HX) b e ginning5 min prior to a d d ition of D H P G rescue d m G luR-d e p end ent LT Din the e E F 2K K O slic e (75.7 % ± 7.4 % of b ase line , p < 0.001,c om p are d to D H P G only in e E F 2K K O slic es) (F igure 6D). Thesa m e tre atm ent of W T slic es d id not substantia lly a lter the tim ec ourse of m G luR-LT D (69.0 % ± 2.6 % of b ase line , p > 0.5, c om-p are d to D H P G only in WT slic es). Low-dose cyc lohe ximid e ha dno effe ct on b ase line syna ptic transmission in the a bsenc e ofm G luR stimulation (101.2 % ± 2.0 % for W T slic es; 100.4 % ±4.6 % for e E F 2K K O slic es). These observations c onfirm thatm G luR signa ling re quire d for m G luR-LT D is se le ctive ly im p a ire din e E F 2K K O in a m anner that c an b e rescue d by transient a p p li-c ation of low-dose cyc lohe ximid e .
m G l u R - L T D , b u t N o t H o m e o s t a t i c P l a s t i c i t y ,Is D i s r u p t e d i n e E F 2 K K O N e u r o n s i n C u l t u r eTo further assess the se le ctivity of the e E F 2K K O effe ct on neu-rona l function, w e e x a mine d two forms of neurona l p lastic ity thatc an b e assaye d in prim ary neurona l cultures. Tre atm ent of cul-tures w ith D H P G for 5 min to evok e m G luR-LT D re duc e d the ratioof surfa c e to tota l G luR2/3 by 30 % in W T neurons but d id notsignific antly re duc e this m e asure in e E F 2K K O neurons (F iguresS8A and S8 B). This result p ara lle ls the d efic it of m G luR-LT D se enin a cute slic es. C ultures w ere a lso assaye d for hom e ostatica d a ptations of surfa c e A M P A re c e ptors sinc e this responseis m ark e d ly a ltere d in Arc/Arg3.1 K O neurons (She pherd et a l.,2006). Tre atm ent of e E F 2K K O c ortic a l cultures for 2 d ays w ithe ither tetrodotoxin (TTX) or b icuculline evok e d hom e ostatic in-cre ases and d e cre ases of surfa c e G luR1 that w ere id entic a l toW T neurons (F igure S8 C ). Thus, e E F 2K K O results in a se le ctived isruption of m G luR-d e p end ent LT D .
F m r 1 K O D i s r u p t s R a p i d , b u t N o t D e l a y e d ,In d u c t i o n o f A r c / A r g 3 . 1 P r o t e i nW e e x a mine d the role of the e E F 2K/e E F 2/Arc m e chanism in thea b errant p lastic ity d escrib e d in F mr1 K O mic e . F M RP has b e enre porte d to b ind Arc/Arg3.1 mR N A (Ia c o ang e li et a l., 2008; Z a lfaet a l., 2003) and is hypothesiz e d to inhib it translation prior tom G luR stimulation (B e ar et a l., 2004; N arayanan et a l., 2007).To assess whether F M RP might b e critic a l for e ither ra p id or d e-laye d induction of Arc/Arg3.1 prote in follow ing m G luR stimula-tion, w e pre p are d prim ary neurona l cultures from F mr1 K Omic e and stimulate d w ith D H P G . Arc/Arg3.1 e x pression in unsti-mulate d cultures w as not c onsistently d ifferent b etw e en W T andF mr1 K O neurons. M ore over, Arc/Arg3.1 prote in incre ase d60 min after D H P G stimulation in F mr1 K O neurons id entic a llyas in W T neurons (F igure 7A). H ow ever, the ra p id incre ase ofArc/Arg3.1 prote in follow ing D H P G stimulation w as a bsent inF mr1 K O neurons (F igure 7A). D H P G a ctivate d m G luR/e E F 2K
signa ling in the F mr1 K O neurons sinc e phospho-e E F 2 w asid entic a lly induc e d as in W T neurons (F igure 7A). Assays ofArc/Arg3.1 prote in sta b ility and induction follow ing prote asom einhib ition w ith M G 132 d id not reve a l d ifferenc es b etw e en WTand F mr1 K O neurons (F igures S9 B and S9 C ). B io che mic a l e x-p erim ents to monitor Arc/Arg3.1 e x pression using a cute hip po-c a m p a l slic es reve a le d that b asa l Arc/Arg3.1 e x pression w ashighly varia b le even when norm a liz e d to tota l prote in or a ctin,ind ic ating a limitation of this pre p aration (T aub enfe ld et a l.,2002). When e x a mine d histo che mic a lly, b asa l Arc/Arg3.1 varie dthrough the thic kness of the slic e (d ata not shown). W e c on-c lud e that F mr1 K O neurons se le ctive ly la c k the a b ility to ra p id lyupre gulate Arc/Arg3.1 e x pression. The re porte d incre ase ofArc/Arg3.1 mR N A in polysom e fra ctions from F mr1 K O mic e(Z a lfa et a l., 2003) sugg ests that fa ilure to d ete ct a D H P G -evok e d ra p id incre ase of Arc/Arg3.1 prote in is link e d to e levate dc onstitutive e x pression.
A r c / A r g 3 . 1 Is R e q u ir e d f o r m G l u R - L T Da n d P P - L F S L T D i n F m r 1 K O M i c eIn antic ip ation of physiologic a l stud ies to assess the role ofArc/Arg3.1 in syna ptic p lastic ity of F mr1 K O mic e , w e e x a mine dArc/Arg3.1 prote in e x pression in hip po c a m pus. Arc/Arg3.1 pro-te in has previously b e en re porte d to b e mod estly upre gulate d inboth tota l bra in and syna ptosom a l fra ctions of F mr1 K O mic e(Z a lfa et a l., 2003). H ow ever, in our assays, Arc/Arg3.1 prote inw as not c onsistently d ifferent in hip po c a m pus (e ither in vivo orin a cute slic es) or c orte x when c are w as ta k en to sa crific emic e w ithout b ehaviora l a ctivation. W e g enerate d mic e in whichboth F mr1 (in F V B b a c kground) and Arc/Arg3.1 (in B 6 b a c k-ground) w ere d e lete d . D oub le Arc/Arg3.1/ F mr1 K O (D K O) mic eare via b le , fertile , and not d ifferent from W T mic e in siz e orpostnata l surviva l. Ind ic es of b asa l syna ptic transmission w erenorm a l in F mr1 K O and Arc/Arg3.1/ F mr1 D K O (F igures S9Dand S9 E). As re porte d previously (N osyreva and Hub er, 2006),D H P G evok e d a susta ine d re duction of syna ptic strength(68.2 % ± 2.6 % of b ase line for F mr1 single K O slic es; 73.0 % ±6.6 % of b ase line for F V B W T slic es; F igure 7 C ). J a c kson la bora-tory provid es F mr1 K O mic e in the F V B b a c kground , and them a gnitud e of LT D w as not signific antly d ifferent from F V B WTmic e . As re porte d previously in stud ies of F mr1 K O in theB 6 b a c kground , m G luR-LT D w as not inhib ite d by high-dosecyc lohe ximid e (60 mM) (F igure 7 B). In Arc/Arg3.1/ F mr1 D K O (inB 6/ F V B), D H P G evok e d an initia l re duction of syna ptic strengththat w as not d ifferent from W T, Arc/Arg3.1 K O , or F mr1 K O .H ow ever, e x pression of D H P G -evok e d LT D w as signific antlyim p a ire d in Arc/Arg3.1/ F mr1 D K O (85.9 % ± 4.1 % , p < 0.01c om p are d to F mr1 single K O or F V B W T). P P-L F S LT D w asa lso im p a ire d in Arc/Arg3.1/ F mr1 D K O (88.3 % ± 2.1 % of b ase-line for Arc/Arg3.1/ F mr1 D K O slic es; 75.5 % ± 3.7 % of b ase linefor F mr1 single K O ; 80.5 % ± 2.6 % of b ase line for F V B WT slic es,p < 0.05 when Arc/Arg3.1/ F mr1 D K O w as c om p are d to F mr1 sin-gle K O or F V B W T; F igure 7D). These results ind ic ate thatArc/Arg3.1 is re quire d for m G luR-LT D in both WT and F mr1K O neurons. D e letion of Arc/Arg3.1 do es not entire ly preventD H P G or P P-L F S LT D , sugg esting that a d d itiona l m e chanismsc ontribute to the a b errant LT D in F mr1 K O mic e .
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
78 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

D IS C U S SI O N
A r c / A r g 3 . 1 P l a y s a C e n t r a l R o l ei n m G l u R - D e p e n d e n t L T DThe present study id entifies Arc/Arg3.1 as a mole cule that is re-quire d for c erta in forms of LT D that are known to b e d e p end enton ra p id , d e novo prote in synthesis, inc lud ing m G luR-LT D andP P-L F S-LT D . U pon a ctivation of group I m G luRs, Arc/Arg3.1prote in is ra p id ly upre gulate d in a pro c ess that re quires d enovo translation. Arc/Arg3.1 prote in functions as a re gulatoryfa ctor for a postsyna ptic endo cytic signa ling p athw ay that in-c lud es endophilins 2 and -3 and dyna min, which tog ether m e d i-ate se le ctive endo cytosis of A M P A typ e gluta m ate re c e ptors( C how dhury et a l., 2006). A c c ord ingly, the lo c a l and ra p id in-cre ase of Arc/Arg3.1 prote in c an b e link e d to the se le ctive down-re gulation of A M P AR at sp e c ific syna pses during LT D (F igure 8).
While group I m G luR a ctivation results in a mod est rise ofArc/Arg3.1 mR N A , the translation that und erlies m G luR-LT D
utiliz es Arc/Arg3.1 mR N A that is present in neurons prior to stim-ulation. Arc/Arg3.1 is re gulate d as an imm e d iate-e arly g ene thatis strongly induc e d during le arning-re late d b ehaviors (G uzowskiet a l., 1999) and e x presse d in d iscrete populations of neuronsthat are p art of b ehaviora lly a ctivate d networks even whenrod ents are resting in hom e c a g e (M arrone et a l., 2008).Arc/Arg3.1 mR N A is present in d endritic re gions of C A1 neuronsof mic e sa crific e d imm e d iate ly from the ir hom e c a g e (F igure S3),ind ic ating that b asa l Arc/Arg3.1 mR N A c an b e e ither a ctivityind e p end ent or re late d to prior neurona l a ctivity. The ra p id trans-lation of d endritic Arc/Arg3.1 mR N A rationa liz es the knownd e p end enc e of m G luR-LT D on ra p id d e novo prote in synthesisand ind e p end enc e of d e novo R N A synthesis (Hub er et a l.,2000). It a lso sugg ests that prior neurona l a ctivity that drivesthe e x pression of Arc/Arg3.1 mR N A c ould mod ify subse quentm G luR-d e p end ent p lastic ity.
In further sup port for the notion that Arc/Arg3.1 is an essentia lLT D mole cule , w e find that m G luR-LT D in F mr1 K O mic e is a lso
F ig ur e 7. L T D Is Im p a ir e d in H ip p o c a m p a lS li c e s D e riv e d fro m A rc / A r g3.1 / F m r1 D o u -b l e K O M i c e(A) DIV14 F mr1 K O neurons w ere tre ate d w ithD H P G as ind ic ate d in F igure 5 C . R a p id but not d e-laye d synthe sis of Arc/Arg3.1 w as a bsent in F mr1K O . The re gulation of phosp ho-e E F 2 w as inta ct inF mr1 K O neurons.(B) H igh-dose cyc lohe ximid e (60 mM: H D- C HX) d idnot b lo c k D H P G -LT D of F mr1 K O slic es. In thepresenc e of a high dose of cyc lohe ximid e ,D H P G -LT D of F mr1 K O w as 72.3 % ± 4.8 % ofb ase line at t = 105 min (n = 5), while D H P G -LT Din WT (F V B) slic es w as b lo c k e d (fE P S P w as95.5 % ± 2.9 % of b ase line at t = 105 min (n = 4);p < 0.01 when F mr1 K O w as c om p are d to F V BW T).( C ) Avera g e tim e c ourse of fE P S P slop e ofArc/Arg3.1/ F mr1 doub le K O (D K O) mic e . m G luR-LT D w as induc e d by D H P G (50 mM , for 5 min).D H P G-LT D of Arc/Arg3.1/ Fmr1 D K O w as 85.9 % ±4.1 % of b ase line at t = 75 min (n = 8). In F mr1K O , D H P G -LT D w as 68.2 % ± 2.6 % of b ase lineat t = 75 min (n = 6). In WT, D H P G -LT D w as73.0 % ± 6.6 % of b ase line at t = 75 min (n = 5).p < 0.01 when Arc/Arg3.1/ F mr1 D K O w as c om-p are d to e ither WT or F mr1 K O . fE P S Ps ofpost-D H P G in F mr1 K O w ere not signific antlyd ifferent from those in F V B WT.(D) Tim e c ourse of the chang e in fE P S P slop e byP P-L F S . P P-L F S LT D of Arc/Arg3.1/ F mr1 D K Ow as 88.3 % ± 2.1 % of b ase line at t = 65 min (n =6). In F mr1 K O , P P-L F S LT D w as 75.5 % ± 3.7 %of b ase line at t = 65 min (n = 8). In F V B WT, P P-L F S LT D w as 80.5 % ± 2.6 % of b ase line at t = 65min (n = 8). p < 0.05 when Arc/Arg3.1/ F mr1 D K Ow as c om p are d to e ither W T or F mr1 K O . fE P S Psof post-D H P G in F mr1 K O w ere not signific antlyd ifferent from those in F V B WT (p = 0.4).S c a le b ars, 0.5 mV/10 ms.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 79

d e p end ent on Arc/Arg3.1. This is nota b le b e c ause m G luR-LT Din F mr1 K O mic e is d istinctly d ifferent from m G luR-LT D in W Tmic e in severa l w ays, inc lud ing insensitivity to prote osom e inhib-itors, prote in synthesis inhib itors, and inhib itors of E R K signa ling(H ou et a l., 2006). D isruption of Arc/Arg3.1 function is the firste x p erim enta l m anipulation shown to re duc e m G luR-LT D inF mr1 K O slic es, but it is lik e ly that a d d itiona l mole cular m e cha-nisms c ontribute to the response sinc e m G luR-LT D is not entire lya bsent in the Arc/Arg3.1/ F mr1 D K O . S inc e F M RP functionsto re press translation of sp e c ific mR N As, the d e p end enc em G luR-LT D in the F mr1 K O mouse sugg ests that Arc/Arg3.1prote in that is present prior to m G luR stimulation c an m e d iatethis response and there by c onfers resistanc e to prote in synthe-sis inhib itors.
G r o u p I m G l u R s a n d R a p i d D e N o v o T r a n s l a t i o n ;C o n v e r g e n c e o f e E F 2 K / e E F 2 a n d F M R P P a t h w a y sm G luR a ctivation induc es the ra p id translation of Arc/Arg3.1 andre quires e E F 2K . e E F 2K b inds H om er and group I m G luRs andd isso c iates from this c om p le x in a C a2+-d e p end ent m anner.Phospho-e E F 2 is present at e x c itatory syna pses and d endritesand is ra p id ly upre gulate d in culture d neurons and hip po c a m p a lslic es by stimulation of group I m G luRs w ith D H P G . The a b ility ofphospho-e E F 2 to inhib it translation is thought to b e g enera l for
F ig ur e 8. e E F 2 K , F M R P , a n d R a p id D eN o v o T r a n s l a tio n o f A r c / A r g3.1 Pro t e in inm G lu R - L T DGroup I m G luRs a ctivate e E F 2K via c a lc ium-c a l-modulin ( C a M). e E F 2K phos phorylates e E F 2,which inhib its e long ation g enera lly but incre asesArc/Arg3.1 translation. Arc/Arg3 .1 forms a c om-p le x w ith endophilin2/3 (E ndo) and dyna min(Dyn) and induc es the interna liz ation of A M P AR( C how dhury et a l., 2006). F M RP inhib its the trans-lation of Arc/Arg3 .1 at the b asa l state . Arc/Arg3.1induction a lone is not suffic ient for m G luR-LT D ,ind ic ating that m G luR a ctivates another p athw aythat is re quire d to interna liz e A M P AR (C ho et a l.,2008). In Fmr1 K O mice, the synthesis of Arc/Arg3.1prote in is c onstitutive ly d ere pre sse d , and d enovo synthe sis of Arc/Arg3 .1 is not re quire d form G luR-LT D .
a ll mR N As and a p p e ars c entra l to itsrole in re gulating Arc/Arg3.1 prote intranslation sinc e low-dose cyc lohe xi-mid e , which mimics phospho-e E F 2 by in-hib iting translation e long ation, c an a lsoinduc e Arc/Arg3.1 e x pression. Low-dose cyc lohe ximid e has b e en re porte dto incre ase the translation of mR N Asthat are poorly initiate d in b asa l c ond i-tions, inc lud ing a - C a M KII (F ernand e zet a l., 2005; G upta and O no, 1997; P erl-m an and F e ld m an, 1982; S che etz et a l.,2000; W a ld en and Tha ch, 1986). B otha - C a M KII and Arc/Arg3.1 mR N As arelo c a liz e d to d endrites (M ori et a l., 2000;
Ste w ard et a l., 1998) and c onta in interna l ribosom e entry site(IR E S) se quenc es in the ir 50 untranslate d re gions (P inkstaffet a l., 2001), which m ay a llow the initiation of translation byboth C a p-d e p end ent and C a p-ind e p end ent m e chanisms (P es-tova et a l., 2001). C a p-d e p end ent initiation of translation is tightlyc ontrolle d by initiation fa ctors such as euk aryotic initiation fa ctor4 E (eIF 4E) and eIF 4E -b ind ing prote in (4 E B P). B y c ontrast, C a p-ind e p end ent initiation through IR E S se quenc e m ay a llow synthe-sis of prote ins when c a p-d e p end ent initiation fa ctors are not fullya ctive (P estova et a l., 2001). The se e mingly p ara d oxic a l effe ct ofphospho-e E F 2 and cyc lohe ximid e to incre ase Arc/Arg3.1 pro-te in translation is c onsistent w ith a mod e l in which inhib ition ofglob a l e long ation results in a ra p id incre ase in fa ctors that arerate limiting for translation initiation of mR N As that m ay b e inef-fic iently initiate d und er b asa l c ond itions (F ernand e z et a l., 2005;S che etz et a l., 2000; W a ld en and Tha ch, 1986). Whether theputative IR E S se quenc e of Arc/Arg3.1 mR N A is re quire d for thisresponse re m a ins to b e d etermine d .
m G luR a ctivity a lso ra p id ly re gulates F M RP-d e p end ent signa l-ing in a m anner that c an enhanc e ra p id , d e novo translation(Mud d ashetty et a l., 2007; N arayanan et a l., 2007; W estm arkand M a lter, 2007). m G luR a ctivation results in d e phosphoryla-tion of F M RP , and this re duc es its inhib itory a ction on translation(N arayanan et a l., 2007). D e phosphoryla tion of F M RP o c curs
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
80 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

w ithin 1 min of m G luR a ctivation and p ara lle ls the tim e c ourse ofm G luR-d e p end ent phosphorylation of e E F 2. Polysom e profilesfrom W T and F mr1 K O bra in ind ic ate that F M RP is asso c iate dw ith mR N As that are not in a ctive translation. In the a bsenc e ofF M RP , sp e c ific mR N As inc lud ing Arc/Arg3.1 shift to fra ctionsthat inc lud e a ctive ly translating polyribosom es (Z a lfa et a l.,2003). Group I m G luRs are re porte d to ra p id ly re duc e the histo-che mic a l c olo c a liz ation of F M RP and F mr1 mR N A at syna pses inasso c iation w ith incre ase d translation of d e novo F M RP (Antaret a l., 2004). While this is an ind ire ct assay of translationa l re gu-lation, it sup ports a mod e l in which m G luR a ctivity re duc esF M RP b ind ing to sp e c ific mR N As and there by provid es a dy-na mic incre ase in mR N As in c lose proximity to syna pses fortranslation. F M RP is not re quire d for m G luR-d e p end ent induc-tion of phospho e E F 2 (F igure 7A), sugg esting that m G luR-F M RP and m G luR-e E F 2K are p ara lle l p athw ays. A c c ord ingly,the ra p id , d e novo Arc/Arg3.1 translationa l response re quire dfor norm a l m G luR-LT D a p p e ars to re quire both a transient rever-sa l of F M RP’s re pressive effe ct on translation and the g enera linhib ition of the e long ation ste p by phospho-e E F 2 (F igure 8).
D e N o v o T r a n s l a t e d A r c / A r g 3 . 1 Is S p e c i fi c a llyR e q u ir e d f o r N o r m a l m G l u R - L T DArc/Arg3.1 prote in that m e d iates m G luR-LT D a p p e ars to b efunctiona lly d istinct from the tota l pool of Arc/Arg3.1 prote ine x presse d in neurons. F or e x a m p le , the b asa l e x pression ofArc/Arg3.1 prote in in e E F 2K K O neurons is identical to that in WTneurons, yet m G luR-LT D and P P-L F S LT D , which are d e p end enton Arc/Arg3.1, are a bsent in the e E F 2K K O . Arc/Arg3.1 prote in ine E F 2K K O neurons is functiona l, sinc e hom e ostatic sc a ling,which is a lso d e p end ent on Arc/Arg3.1 (She pherd et a l., 2006),is inta ct. This a p p arent p ara dox is resolve d w ith the observa-tions that e E F 2K K O neurons se le ctive ly la c k ra p id , d e novoArc/Arg3.1 induction, and this d efic it is rescue d by brief a p p lic a-tion of low-dose cyc lohe ximid e that a lso rescues m G luR-LT D .Thus, the e E F 2-d e p end ent pool of Arc/Arg3.1 prote in a p p e arsunique ly c a p a b le of m e d iating LT D . In another e x a m p le d e mon-strating the d istinct prop erties of Arc/Arg3.1 re late d to its mod eof translation, Arc/Arg3.1 prote in is e x presse d at ne ar norm a lleve ls in F mr1 K O mic e , and Arc/Arg3.1 prote in is induc e d after40 min of D H P G stimulation; how ever, the ra p id incre ase ofArc/Arg3.1 at 5 min is a bsent. In this re g ard , e x pression ofArc/Arg3.1 prote in in the F mr1 K O a p p e ars id entic a l to thee E F 2K K O , but w ith the te chnic a l c ave at that w e c annot d ete ctchang es in b asa l translation of Arc/Arg3.1 that are pre d icte dfrom other stud ies to b e incre ase d in F mr1 K O neurons (Z a lfaet a l., 2003). O ur d ata sugg est that an e levation of c onstitutiveArc/Arg3.1 translation in F mr1 K O und erlies m G luR LT D that isa b errantly ind e p end ent of d e novo translation but d e p end enton Arc/Arg3.1. This mod e l rationa liz es the d istinctly d ifferentm G luR-LT D phenotyp es in e E K2K and F mr1 K O s even thoughthey show a similar a bsenc e of a cute m G luR Arc/Arg3.1 induc-tion. The re lative a mount of this e E F 2K/ F M RP-d e p end ent poolof Arc/Arg3.1 prote in is typ ic a lly sm a ll c om p are d to theArc/Arg3.1 leve l of the d e laye d response to D H P G or B D N F inneurona l cultures, and w e c ould not d ete ct d ifferenc es in b asa land induc e d Arc/Arg3.1 in WT and e E F 2K K O neurons usingc onventiona l histo che mic a l te chniques. W e infer that prote ins
translate d by the e E F 2K/ F M RP-d e p end ent m e chanism areunique ly ava ila b le for m G luR-LT D and antic ip ate that ne w im a g-ining m ethods w ith high sp atia l and te m pora l resolution m ayshe d light on this pre d iction.
O ur stud ies a lso ind ic at e that m G luR-LT D re quires a se c ondsigna l in a d d ition to ra p id , d e novo Arc/Arg3.1 prote in. E x p eri-m ents that use cyc lohe ximid e to rescue m G luR-LT D in thee E F 2K K O d e monstrate that cyc lohe ximid e a lone is suffic ientto induc e Arc/Arg3.1 prote in but do es not induc e LT D unless a c-c om p anie d by m G luR a ctivation. S imilarly, low-dose cyc lohe xi-mid e induc es Arc/Arg3.1 in W T neurons but do es not induc eLT D w ithout c oncurrent m G luR a ctivation. These observationsrationa liz e why misre gulate d e x pression of Arc/Arg3.1 prote inin the F mr1 K O , and presum a b ly other prote ins, do es not o c-c lud e m G luR-LT D . The dua l d e p end enc e of Arc/Arg3.1 prote inand m G luR a ctivity for a b errant LT D in F mr1 K O mic e provid esa sup porting rationa le for tre atm ent re gim ens of Fra gile X syn-drom e that inc lud e group I m G luR anta gonists (B e ar et a l.,2004; M c Brid e et a l., 2005). In other stud ies, w e have re porte dthat m G luR-LT D re quires the ra p id c le ava g e of the neurona lp entra xin re c e ptor prote in (N PR) in a pro c ess that involves thee xtra c e llular m eta lloprote ase tumor ne crosis fa ctor- a c onvertingenzym e (TA C E /A D A M17) ( C ho et a l., 2008). The p entra xin do-m a in, re le ase d of its transm e m brane tether, a p p e ars to c a ptureA M P A re c e ptors at the site of endo cytosis, and this m e chanismis re quire d for m G luR-LT D in both the hip po c a m pus and c ere-b e llum. It is possib le that this N PR p athw ay functions in c on-junction w ith the Arc/Arg3.1 endosom a l p athw ay and offers apotentia l ne w targ et for a g ents to mod ify m G luR-LT D .
E X P E RI M E N T A L P R O C E D U R E S
C onstructs, c e ll culture , re a l-tim e RT-P C R, m eta bolic la b e ling, and fluores-c enc e in situ hybrid iz a tion assay are inc lud e d in Sup p le m enta l E x p erim e nta lPro c e dure s.
A n t i b o d i e sAnti-phospho-e E F 2 (Thr56: ra b b it poly c lona l) and tota l-e E F 2 (ra b b it poly-c lona l) from C e ll S igna ling; e E F 2K (ra b b it poly c lona l) and m G luR1 (mousemono c lona l) from B D B iosc ienc e s; m G luR5, m G luR2, and P S D-95 from U p-state; m G luR4 from Z ym e d; horse ra d ish p eroxid ase (H RP) c onjug ate d H A an-tibody, H RP-c onjug ate d myc antibody , myc (mouse mono c lona l), and a ctin(mous e mono c lona l) from S anta Cruz; Arc/Arg3.1 (Lyford et a l., 1995);N- G luR1 antibody (She ph erd et a l., 2006).
A M P A R e c e p t o r T r a f fi c k i n g E x p e ri m e n t s a n d I m m u n o s t a i n i n gLa b e ling of surfa c e or interna liz e d pool of A M P A re c e ptor w as p erform e d asd escrib e d w ith minor mod ific ations (She pherd et a l., 2006). Briefly, surfa c eG luR1-c onta ining A M P A re c e ptors w ere la b e le d by a d d ing 2.5 mg of G luR1-N J H1816 p A b to the neurona l growth m e d ia and subse quently incub ate d at37 C for 15 or 60 min after 5 min D H P G a p p lic ation. To visua liz e surfa c e andinterna liz e d G luR1, A le x a 555 se c ond ary w as a d d e d in e x c e ss live at 10 C .N eurons w ere fix e d , p erm e a b iliz e d , and subse quently e x pose d to A le x a 488se c ond ary to sta in interna liz e d re c e ptors (b a c kground in the nonp erm e a b i-liz e d c ontro l w as ne gligib le). Immuno cyto che mistry of culture d neurons w asp erform e d as d escrib e d (She pherd et a l., 2006). Immunohisto che mistry ofphosp ho-e E F 2 in WT and e E F 2K mic e w as p erform e d as d escrib e d (R a m-ire z-Am aya et a l., 2005) w ith slight mod ific ations. S e e Sup p le m enta l E x p eri-m enta l Pro c e dures for d eta ils.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 81

E l e c t r o p h y s i o l o g yF ie ld re c ord ing of e x c itatory postsyna ptic potentia l (fE P S P) of hip po c a m p a lC A1 neurons of postna ta l d ay (P)21–30 m a le mic e as d escrib e d w ith minormod ific a tions (Hub er et a l., 2000). m G luR-LT D w as induc e d by an m G luR1/5a gonist, (R,S)-3,5-D H P G , for 5 min (To cris, 50 mM , unless otherw ise ind ic ate d),or by p a ire d- pulse low-fre quency stimulation (P P-L F S: 50 ms interstimulusinterva l, 1 H z , for 15 min) in the prese nc e of D-A PV (To cris, 50 mM). N M D A R-d e p end e nt LT D w as ind uc e d by using 900 single pulse s d e livere d at 1 H z(Hub er et a l., 2000).
LTP w as m e asure d in S ch affer c ollatera l- C A1 syna pse s in hip po c a m p a l sli-c es d erive d from 8- to 10-w e e k-o ld m a le mic e . Late-phase LTP (L-LTP) w asindu c e d by four tra ins of high-fre que ncy stimulation (H F S) (100 H z , 1 s) w ith3 s of intertra in interva l. S e e Sup p le m enta l E x p erim e nta l Pro c e dure s ford eta ils.
W e s t e r n B l o t t i n g , I m m u n o p r e c i p i t a t i o n A s s a y ,a n d S u r f a c e B i o t i n y l a t i o n A s s a yW estern b lotting, IP assay , and surfa c e b iotinylation assay w ere p erform e d aspreviously d escrib e d (C ho w dhury et a l., 2006; C ho et a l., 2008). S e e Sup p le-m enta l E x p erim e nta l Pro c e dures for d eta ils.
S U P P L E M E N T A L D A T A
The Sup p le m enta l D ata inc lud e Sup p le m enta l E x p erim e nta l Pro c e dures andfigures and c an b e found w ith this artic le online at http://w w w .neuron.org/c gi/c ont ent/full/59/1/70/D C 1/.
A C K N O W L E D G M E N T S
This work w as sup porte d by NIM H grants M H053608 (P . F .W .), M H068830(R.L. H .), M H51106 (D .J.L.), NID A grant D A00266 (P . F .W .), NIA grantA G 019890 (A . G .R.), G M057300 (A . G .R .), and F RAXA (W . K .). W e tha nk M arlinD ehoff and G lory H arris for anim a l husb a ndry and D avid Lie b erm a n for he lpfuld iscussions.
R e c e ive d: August 21, 2007R evise d: D e c e m b er 22, 2007A c c e pt e d: M ay 17, 2008Pub lishe d: July 9, 2008
R E F E R E N C E S
Antar, L. N ., Afroz , R., D ictenb erg , J. B ., C arroll, R. C ., and B asse ll, G .J. (2004).M eta botrop ic gluta m ate re c e ptor a ctiva tion re gulates fra gile x m enta l retard a-tion prot e in and F M R1 mR N A lo c a liz ation d iffere ntia lly in d endrites and atsyn a pses. J. N eurosc i. 24, 2648–2655.
As a ki, C ., Usud a , N ., N a k a z a w a , A ., K a m etani, K ., and Suzuki, T. (2003). Lo c a l-iz ation of translationa l c om ponents at the ultra microsc op ic leve l at postsyna p-tic sites of the rat bra in. Bra in R es. 972, 168–176.
B e ar, M . F ., Hub er, K .M . , and W arren, S .T. (2004). The m G luR the ory of fra g ile Xm enta l retard ation. Trends N eurosc i. 27, 370–377.
B e guere t, J., P errot, M ., and Crouz et, M . (1977). Ribosom a l prote ins in the fun-gus Podos pora anserina: evid enc e for an e le ctrop horetic a lly a ltere d 60Sprot e in in a cyc lohe ximid e resist ant mutant. M ol. G en. G enet. 156, 141–144.
Browne , G .J., and Proud , C . G . (2002). R e gulation of p e ptid e-c ha in e long ationin m a mm a lia n c e lls. E ur. J. B io che m. 269, 5360–5368.
C audy, A .A ., Myers, M ., H annon, G .J., and H a mmond , S .M . (2002). Fra gileX-re lat e d prote in and VIG asso c iate w ith the R N A interferen c e m a chinery.G ene s D ev. 16, 2491–2496.
C e m a n, S ., O ’D onn e ll, W .T., R e e d , M ., P atton, S ., Pohl, J., and W arren, S .T.(2003). Phosphoryla tion influenc es the translation state of F M RP-asso c iate dpolyribosom es. Hum. M ol. G enet. 12, 3295–3305.
C ho , R.W ., P ark, J.M ., Wolff, S . B . E ., Xu, D ., H o pf, C ., K im, J.-a ., R e d dy , R. C .,P etra lia , R.S ., P erin, M .S ., Lind en, D .J., and Worley, P . F . (2008). m G luR1/5-
d e p end ent long-term d e pression re quires the re gulat e d e ctodo m a in c le ava g eof neurona l p entra x in N PR by TA C E . N euron 57, 858–871.
C hotiner, J. K ., Khorasani, H ., N a irn, A . C ., O ’D e ll, T.J., and W atson, J. B . (2003).A d enylyl cyc lase- d e p end ent form of che mic a l long-term pot entiation trigg erstranslationa l re gulation at the e long ation ste p . N eurosc ienc e 116, 743–752.
C how dhury, S ., She pherd , J.D ., O kuno , H ., Lyford , G ., P etra lia , R.S ., P lath,N ., Kuhl, D ., Hug anir, R.L., and Worley, P . F . (2006). Arc/Arg3.1 intera ctsw ith the endo c ytic m a chinery to re gulat e A M P A re c e ptor traffic king. N euron52, 445–459.
D arne ll, J. C ., J ensen, K . B ., Jin, P ., Brown, V., W arren, S .T., and D arne ll, R. B .(2001). Fra gile X m enta l retard ation prote in targ ets G quartet mR N As im portantfor neurona l function. C e ll 107, 489–499.
D avid kova , G ., and C arroll, R. C . (2007). C hara cteriz ation of the role of micro-tubule-asso c iate d prote in 1 B in m eta botrop ic gluta m ate re c e ptor-m e d iate dendo cytosis of A M P A re c e ptors in hip po c a m pus. J. N eurosc i. 27, 13273–13278.
F ernand e z , J., Y a m an, I., Huang, C ., Liu, H ., Lop e z , A . B ., K om ar, A .A ., C a p-rara , M . G ., M erric k , W . C ., Snid er, M .D . , K aufm an, R.J. , et a l. (2005). Ribosom esta lling re gulates IR E S-m e d iate d translation in euk aryotes, a p ara lle l to pro-k aryotic attenuation. M ol. C e ll 17, 405–416.
Frey, U ., and M orris, R. G . (1997). Syna ptic ta gging and long-term pot entiation.N ature 385, 533–536.
G iorgi, C ., Y e o, G .W ., Stone , M . E ., K atz , D . B ., B urg e , C ., Turrigiano , G ., andM oore , M .J. (2007). The E J C fa ctor eIF 4AIII modulates syna ptic strength andneurona l prot e in e x pression. C e ll 130, 179–191.
G upta , K . C ., and O no , E . (1997). Stimulation of S end a i virus C ’ prote in synthe-sis by cyc lohe ximid e . B io che m. J. 321, 811–818.
G uzowsk i, J. F ., M c N aughton, B .L., B arnes, C .A ., and Worley, P . F . (1999).Environm ent-sp e c ific e x pression of the im m e d iate-e arly g ene Arc in hip po-c a m p a l neurona l ense m b les. N at. N euros c i. 2, 1120–1124.
H ou, L., Antion, M .D ., Hu, D ., S p enc er, C .M ., P aylor, R., and K lann, E . (2006).Dyna mic translationa l and prote asom a l re gulation of fra gile X m enta l retard a-tion prote in c ontro ls m G luR-d e p e nd ent long-term d e pression. N euron 51,441–454.
Hub er, K .M ., K ayser, M .S ., and B e ar, M . F . (2000). Role for ra p id d endritic pro-te in synthesis in hip po c a m p a l m G luR-d e p e nd ent long-term d e pression.S c ienc e 288, 1254–1257.
Hub er, K .M ., G a lla gher, S .M ., W arren, S .T., and B e ar, M . F . (2002). A ltere d syn-a ptic p lastic ity in a mouse mod e l of fra gile X m enta l retard ation. Pro c . N atl.A c a d . S c i. U S A 99, 7746–7750.
Ia c o ang e li, A ., Rozhd estvensky , T.S ., D olzhansk aya , N ., Tournier, B ., S chutt,J., Brosius, J., D enm an, R. B ., Kh and jian, E .W . , K ind ler, S ., and Tie dg e , H .(2008). O n B C 1 R N A and the fra gile X m enta l retard ation prote in. Pro c . N atl.A c a d . S c i. U S A 105, 734–739.
Ishizuk a , A ., S iom i, M . C ., and S iomi, H . (2002). A Drosop hila fra gile X prote inintera cts w ith c om ponents of R N A i and ribosom a l prote ins . G ene s D ev. 16,2497–2508.
Jin, P ., Z arnesc u, D . C ., C e m a n, S ., N a k a moto, M ., M owrey , J., Jong ens , T.A .,N e lson, D .L., M oses , K ., and W arren, S .T. (2004). B io che mic a l and g enetic in-tera ction b etw e e n the fra g ile X m enta l retard ation prote in and the microR N Ap athw ay. N at. N eurosc i. 7, 113–117.
K aud erer, B .S ., and K and e l, E .R. (2000). C a pture of a prote in synthesis-d e p en-d ent c om ponent of long-term d e pression. Pro c . N atl. A c a d . S c i. U S A 97,13342–13347.
K e lleher, R.J., 3rd , G ovind ara jan, A ., and Tone g a w a , S . (2004). Translationa lre gulatory m e chanisms in p ersiste nt forms of syna ptic p lastic ity. N euron 44,59–73.
Link, W ., K onietz ko, U ., K ause lm ann, G ., Krug, M ., S chw ank e , B ., Frey, U ., andKuhl, D . (1995). Som atod endritic e x pression of an imm e d iate e arly g ene is re g-ulate d by syn a ptic a ctivity . Pro c . N atl. A c a d . S c i. U S A 92, 5734–5738.
Lyford , G .L., Y a m a g a ta , K ., K aufm ann, W . E ., B arne s, C .A ., S and ers, L. K ., C o-p e land , N . G ., G ilb ert, D .J., J enkins, N .A ., Lanahan, A .A ., and Worley , P . F .(1995). Arc , a gro wth fa ctor and a ctivity-re gula te d g ene , enc od es a nov e l
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
82 N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc .

cytosk e leton-a sso c iate d prote in that is enriche d in neurona l d endrites. N euron14, 433–445.
M arrone , D . F ., S chaner, M .J., M c N aughton, B .L., Worley, P . F ., and B arne s,C .A . (2008). Imm e d iat e-e arly g ene e x pression at rest re c a p itulates re c ent e x-p erienc e . J. N euros c i. 28, 1030–1033.
M c Brid e , S .M ., C hoi, C . H ., W ang, Y ., Lie b e lt, D ., Braunst e in, E ., F erre iro, D .,S ehg a l, A ., S iw ic ki, K . K ., D o c k e ndorff, T. C ., N guyen, H .T., et a l. (2005).Pharm a c ologic a l rescue of syna ptic p lastic ity, c ourtship b ehavior, andmushroom body d efe cts in a Drosophila mod e l of fra gile X syndrom e . N eu-ron 45, 753–764.
M ori, Y ., Im a izumi, K ., K ataya m a , T., Yone d a , T., and Tohya m a , M . (2000). Twoc is-a cting e le m e nts in the 30 untra nslate d re gion of a lpha- C a M KII re gulate itsd endritic targ eting . N at. N euros c i. 3, 1079–1084.
Mud d ashetty, R.S ., K e lic , S ., Gross, C ., Xu, M ., and B asse ll, G .J. (2007). Dys-re gulate d m eta botrop ic gluta m at e re c e p tor-d e p end ent translation of A M P Are c e ptor and postsyna ptic d ensity-95 mR N A s at syn a pses in a mouse mod e lof fra gile X syndrom e . J. N eurosc i. 27, 5338–5348.
N a irn, A . C ., and P a lfrey, H . C . (1987). Id entific ation of the m a jor Mr 100,000substrate for c a lmodulin- d e p end ent prote in kinase III in m a mm a lian c e lls ase long ation fa ctor-2. J. B iol. C he m. 262, 17299–17303.
N arayanan, U ., N a lava d i, V., N a k a m oto, M ., P a llas, D . C ., C e m an, S ., B asse ll,G .J., and W arren, S .T. (2007). F M RP phosphory lation reve a ls an imm e d iat e-e arly signa ling p athw ay trigg ere d by group I m G luR and m e d iate d by P P2A .J. N eurosc i. 27, 14349–14357.
N osyreva , E .D . , and Hub er, K .M . (2006). M eta botrop ic re c e ptor-d e p end entlong-term d e pression p ersists in the a bsenc e of prote in synthe sis in the mousemod e l of fra gile X syndrom e . J. N europhysiol. 95, 3291–3295.
O brig, T. G ., C ulp , W .J., M c K e ehan, W .L., and H ard esty, B . (1971). The m e ch-anism by which cyc lohe xim id e and re late d glutarimid e antib iotics inhib itp e ptid e synthesis on reticulo cyte ribosom es . J. B iol. C he m. 246, 174–181.
O ’D onne ll, W .T., and W arren, S .T. (2002). A d e c a d e of mole cular stud ies offra gile X syndrom e . Annu. R ev. N euros c i. 25, 315–338.
P erlm an, J., and F e ld m an, J. F . (1982). C yc lohe ximid e and he at sho c k in-duc e ne w polyp e ptid e synthesis in N eurospora crassa . M ol. C e ll. B iol. 2,1167–1173.
P estova , T.V., K olup a eva , V. G ., Lom a kin, I. B ., P ilip enk o, E .V., Sha tsky, I. N .,Agol, V.I., and H e llen, C . U . (2001). M ole cular m e chanisms of translation initia-tion in euk aryotes. Pro c . N atl. A c a d . S c i. U S A 98, 7029–7036.
P inkstaff, J. K ., C ha p p e ll, S .A ., M auro, V.P ., E d e lm an, G .M ., and Krushe l, L.A .(2001). Interna l initiation of translation of five d endritic a lly lo c a liz e d neurona lmR N As. Pro c . N atl. A c a d . S c i. U S A 98, 2770–2775.
P lath, N ., O han a , O ., D a mm erm ann, B ., Errington, M .L., S chmitz , D ., Gross, C .,M a o, X., E ng e lsb erg, A ., M ahlk e , C ., W e lz l, H ., et a l. (2006). Arc/Arg3 .1 isessentia l for the c onsolid ation of syna ptic p lastic ity and m e m ories. N euron52, 437–444.
R a mire z-Am ay a , V., Va z d arjanova , A ., M ikha e l, D ., Rosi, S ., Worley , P . F ., andB arnes, C .A . (2005). S p atia l e x p loration- induc e d Arc mR N A and prote in e x-pression: evid enc e for se le ctive , network-sp e c ific re a ctivation. J. N euros c i.25, 1761–1768.
R a o, V.R., P intchovski, S .A ., C hin, J., P e e b les, C .L., M itra , S ., and F ink b e iner,S . (2006). A M P A re c e ptors re gulate transcription of the p lastic ity-re late dimm e d iate-e arly g ene Arc . N at. N eurosc i. 9, 887–895.
Rya z anov, A . G . (1987). C a2+/c a lmo dulin-d e p end ent phosp horylation of e lon-g ation fa ctor 2. F E B S Lett. 214, 331–334.
Rya z anov, A . G . (2002). E long ation fa ctor-2 kinase and its ne w ly d isc overe dre latives. F E B S Lett. 514, 26–29.
Rya z anov, A . G ., Shesta kova , E .A ., and N ata pov, P . G . (1988). Phosphoryla tionof e long ation fa ctor 2 by E F -2 kinase affe cts rate of translation. N ature 334,170–173.
S cha effer, C ., B ardoni, B ., M and e l, J.L., E hresm ann, B ., Ehresm ann, C ., andM oine , H . (2001). The fra gile X m enta l retard ation prote in b inds sp e c ifi c a llyto its mR N A via a purine quartet motif. E M B O J. 20, 4803–4813.
S che etz , A .J., N a irn, A . C ., and C onstantine-P aton, M . (2000). N M D A re c e ptor-m e d iate d c ontrol of prote in synthesis at d eve lop ing syn a pses. N at. N eurosc i.3, 211–216.
She ph erd , J.D ., Rum b augh, G ., Wu, J., C how dhury , S ., P lath, N ., Kuhl, D ., Hu-g anir, R.L., and W orley, P . F . (2006). Arc/Arg3.1 m e d iates hom e ostatic syna pticsc a ling of A M P A re c e ptors. N euron 52, 475–484.
Snyd er, E .M ., Philpot, B .D ., Hub er, K .M ., D ong, X., F a llon, J.R., and B e ar, M . F .(2001). Intern a liz ation of ionotrop ic gluta m ate re c e ptors in response to m G luRa ctivation. N at. N eurosc i. 4, 1079–1085.
Ste w ard , O ., W a lla c e , C .S ., Lyford , G .L., and Worley, P . F . (1998). Syna ptic a c-tivation c auses the mR N A for the IE G Arc to lo c a liz e se le ctive ly ne ar a ctivate dpostsyna ptic sites on d endrites. N euron 21, 741–751.
T aub enfe ld , S .M ., Stevens, K .A ., Pollonini, G ., Ruggiero, J., and A lb erini, C .M .(2002). Profound mole c ular chang es follow ing hip p o c a m p a l slic e pre p ara tion:loss of A M P A re c e ptor subunits and unc oup le d mR N A/ prote in e x pression. J.N euro ch e m. 81, 1348–1360.
Tu, J. C ., Xia o, B ., Yuan, J.P ., Lanahan, A .A ., Le offert, K ., Li, M ., Lind en, D .J.,and W orley, P . F . (1998). H om er b inds a nove l proline-rich motif and links group1 m eta botrop ic gluta m ate re c e ptors w ith IP3 re c e ptors. N euron 21, 717–726.
W a ld en, W . E . , and Tha ch, R. E . (1986). Translationa l c ontro l of g ene e x pressionin a norm a l fibrob last. C hara cteriz a tion of a sub c lass of mR N As w ith unusua lkinetic prop erties. B io che mistry 25, 2033–2041.
W ang, H ., and Tie dg e , H . (2004). Translat iona l c ontro l at the syna pse . N euro-sc ientist 10, 456–466.
W e iler, I.J., S p angler, C . C ., K lintsova , A .Y ., Grossm a n, A .W ., K im, S . H ., B er-ta ina-Ang la d e , V., Kha liq , H ., d e Vries, F . E ., La m b ers , F .A ., H atia , F ., et a l.(2004). Fra gile X m enta l retard ation prote in is ne c essary for neurotransmit-ter-a ctivat e d prot e in translation at syn a pses. Pro c . N atl. A c a d . S c i. U S A101, 17504–17509.
W estm ark, C .J., and M a lter, J.S . (2007). F M RP m e d iates m G luR5-d e p end enttranslation of a myloid pre cursor prote in. PLoS B io l. 5, e52.
Yuan, J.P ., K ise lyov, K ., Shin, D .M . , C he n, J., Shcheynikov , N ., K ang, S . H .,D ehoff, M . H ., S chw arz , M . K ., S e e burg, P . H ., Mua lle m, S ., and Worley, P . F .(2003). H om er b ind s TRP C fa mily chann e ls and is re quire d for g ating ofTRP C 1 by IP3 re c e p tors. C e ll 114, 777–789.
Z a lfa , F ., G iorgi, M ., Prim era no, B ., M oro, A ., D i P enta , A ., R e is, S ., O ostra , B .,and B a gni, C . (2003). The fra gile X syndrom e prote in F M RP asso c iates w ithB C 1 R N A and re gulates the translation of sp e c ific mR N As at syna pses. C e ll112, 317–327.
N e urone E F 2K-D e p end ent Arc Translation M e d iates m G luR-LT D
N euron 59, 70–83, July 10, 2008 ª 2008 E lsevier Inc . 83





























