Embed Size (px)
Citation preview

TOXICOLOGICAL SCIENCES 99(2), 488–501 (2007)
doi:10.1093/toxsci/kfm178
Advance Access publication July 16, 2007
NF-jB Plays a Major Role in the Maturation of Human DendriticCells Induced by NiSO4 but not by DNCB
Nadege Ade,*,† Diane Antonios,*,† Saadia Kerdine-Romer,*,† Fanny Boisleve,*,† Francxoise Rousset,‡and Marc Pallardy*,†,1
*Univ Paris-Sud, †INSERM, 92296 Chatenay-Malabry, France; and ‡L’Oreal Recherche, 92117 Clichy-La Garenne, France
Received May 14, 2007; accepted July 3, 2007
Dendritic cell (DC) activation is a critical event for the
induction of an immune response to haptens. Although signal-
ing pathways such as mitogen-activated protein kinase (MAPK)
family members have been reported to play a role in DC
activation by haptens, little is known about the implication of
the nuclear factor kappa B (NF-kB) pathway. In this work, we
showed that NiSO4 induced the expression of HLA-DR, CD83,
CD86, and CD40 and the production of interleukin (IL)-8, IL-6,
and IL-12p40 in human DCs, whereas 1-chloro-2,4-dinitrobenzene
(DNCB) induced mainly the expression of CD83 and CD86 and
the production of IL-8. NiSO4 but not DNCB was able to activate
the degradation of IkB-a leading to the binding of the p65 subunit
of NF-kB on specific DNA probes. Inhibition of the NF-kB path-
way using BAY 11-7085 prevents both CD40 and HLA-DR expres-
sion and cytokine production induced by NiSO4. However, BAY
11-7085 only partially inhibited CD86 and CD83 expression in-
duced by NiSO4. In addition, p38 MAPK and NF-kB were
independently activated by NiSO4 since SB203580 did not inhibit
NF-kB activation by NiSO4. Interestingly, we also showed that
DNCB inhibited the degradation of IkB-a induced by tumor
necrosis factor-a leading to alteration of CD40, HLA-DR, and
CD83 expression but not of CD86 and CCR7. Extensive
modifications of DC phenotype by NiSO4 in comparison to
DNCB are probably the consequence of NF-kB activation by
NiSO4 but not by DNCB.
Key Words: contact hypersensitivity; NF-jB; hapten;
dendritic cells.
Allergic contact hypersensitivity is a T-cell immune
response resulting from dendritic cell (DC) activation in the
skin by small molecular weight compounds termed haptens.
Once DCs capture the hapten in the skin, they migrate to the
draining lymph nodes and present the hapten associated with
major histocompatibility complex (MHC) Class II molecules
to naıve T lymphocytes leading to the clonal expansion of
hapten-specific T lymphocytes (Bour et al., 1995; Rougier
et al., 2000).
Langerhans cells (LC) isolated from skin explants treated
with haptens have shown upregulation of CD86, CD54, and
HLA-DR expression (Tuschl and Kovac, 2001). However, the
use of LC from skin samples is limited in term of low LC yields
and sometimes spontaneous maturation occurs during the
extraction procedure. The discovery of methods for
in vitro generation of immature DC constituted a useful advance
to study molecular mechanisms involved in DC maturation. DC
can be generated either from monocytes (Sallusto and
Lanzavecchia, 1994) or CD34þ hematopoietic progenitors
obtained from neonatal cord blood (Caux et al., 1996). These
models offered new insight for studying DC response to contact
sensitizers (CS). Upon treatment with CS, DCs show upregu-
lation of markers such as MHC Class II molecules, CD54,
CCR7, CD86, CD83, CD80, and CD40 (Aiba et al., 1997, 2000;
Arrighi et al., 2001; Boisleve et al., 2004; De Smedt et al., 2001,
2005; Staquet et al., 2004; Tuschl et al., 2000). These
phenotypical changes are accompanied by cytokine production
such as interleukin (IL)-6, tumor necrosis factor (TNF)-a, IL-8,
and IL-1b (Aiba et al., 1997; De Smedt et al., 2001, 2005;
Tuschl et al., 2000; Verheyen et al., 2005). In addition, not only
cultured human LC but also in vitro generated human DC could
prime T cells to induce a proliferative response to strong
allergens (Rougier et al., 2000). All these results show that CS
are able to induce the activation of human DC in spite of the lack
of skin environment, suggesting that DCs act as a crucial
‘‘sensor’’ of danger signals including low molecular weight
compounds (Gallucci and Matzinger, 2001).
One purpose of our work is to elucidate if simple chemicals
are able to induce DC maturation in a manner similar to the one
observed with well described danger signals such as lipopoly-
saccharides (LPS) and proinflammatory cytokines (Arrighi
et al., 2001; Boisleve et al., 2004; Verhasselt et al., 1997). To
activate DC, danger signals use several common signal
transduction pathways with major roles for mitogen-activated
protein kinases (MAPKs) and nuclear factor kappa B (NF-jB).
However, signaling involved in CS-induced DC maturation
remains to be clarified. Kuhn et al. (1998) have demonstrated
1 To whom correspondence should be addressed at INSERM UMR-S 749
and Toxicology, Faculte de Pharmacie, 5 rue JB Clement, 92296 Chatenay-
Malabry Cedex, France. Fax: þ33-1-46-83-54-96. E-mail: marc.pallardy@
u-psud.fr.
� The Author 2007. Published by Oxford University Press on behalf of the Society of Toxicology. All rights reserved.For Permissions, please email: [email protected]
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

an increase in tyrosine-phosphorylated proteins after stimula-
tion with strong CS. Several authors have also reported the role
of members of the MAPK family, namely, extracellular
regulated kinases, jun kinases (JNK), and p38 MAPK in DC
maturation induced by CS (Aiba et al., 2003; Arrighi et al.,2001; Boisleve et al., 2004, 2005). Indeed, in monocyte-
derived DC (MoDC), p38 MAPK has been demonstrated to be
critical for the upregulation of CD80, CD83, and to a lesser
extent CD86 but not for CD1a, CD40, and HLA-DR
expression (Arrighi et al., 2001).
In contrast to the MAPK pathway, the involvement of the
NF-jB pathway in response to CS is still poorly understood.
Attar et al. (1998) have shown that mutation of the IjB-b gene
in mice impaired delayed type hypersensitivity in response to
fluorescence isothiocyanate (FITC). Nickel and cobalt, that are
well known sensitizers, have been shown to induce NF-jB
binding activity in human endothelial cells (Goebeler et al.,1995). Aiba et al. (2003) have also shown that NiSO4 can
activate NF-jB in MoDC. However, to date there is no report
evaluating the role of NF-jB in DC maturation upon hapten
exposure.
In unstimulated cells, NF-jB-family proteins exist as
heterodimers and homodimers that are sequestered in the
cytoplasm by members of the IjB family. Upon cell stim-
ulation, activation of IjB kinase (IKK) is observed leading to
phosphorylation of IjB proteins on serine residues. The
phosphorylation of IjB-a induced the dissociation of the
NF-jB/IjB complex and subsequent ubiquitination and pro-
teolysis of IjB-a by the proteasome. Free NF-jB heterodimers
(p65/p50) can then translocate to the nucleus where they bind
to NF-jB enhancer element of target genes. Signals such as
Toll-Like Receptors (TLR) agonists or proinflammatory cyto-
kines are known to activate NF-jB in DC leading to production
or expression of many inflammatory cytokines, chemokines,
immune receptors, and cell surface molecules (Verhasselt
et al., 1999).
NF-jB and MAPK pathways are the two major pathways
involved in DC maturation induced by danger signals such as
TLR agonists. Our hypothesis is that CS are perceived as danger
signals by DC inducing their maturation. In this study, we
investigated the role of NF-jB in phenotypic changes and
cytokine production induced in response to NiSO4 and DNCB.
We choose these two haptens as prototypes of metallic and
organic sensitizers. Our results show that NiSO4 but not DNCB
is able to induce NF-jB activation in CD34-derived DC (CD34-
DC). This observation may explain phenotypical differences
observed when DCs are treated with either NiSO4 or DNCB.
MATERIALS AND METHODS
Preparing DC from human cord blood. Normal human umbilical cord
blood was obtained from Biopredic (Rennes, France) and processed within 24 h.
Cord blood samples were diluted 1:6 in phosphate-buffered saline (PBS). After
Ficoll–Hypaque (Medium for Lymphocyte Isolation, Eurobio, Les Ulis,
France) centrifugation (300 g for 30 min at 20�C), mononuclear cells were
collected and washed three times in PBS supplemented with 2% of heat-
inactivated fetal bovine serum (FBS). Cord blood CD34þ hematopoietic cells
were isolated using MiniMACS separation columns (Miltenyi Biotec, Bergish,
Germany) through magnetic positive selection using the direct CD34
progenitor cell isolation kit (Miltenyi Biotec). After purification, the isolated
cells were 80–95% CD34þ cells.
CD34þ cells were adjusted to the concentration of 3 3 105 cells/ml and
cultured at 37�C in a humidified 5% CO2 atmosphere in RPMI 1640 Glutamax
I medium, 10% FBS, 100 U/ml penicillin, 100 lg/ml streptomycin, 1mM
sodium pyruvate (all from Gibco Invitrogen, Paisley, UK), and supplemented
with 200 U/ml granulocyte-macrophage-colony stimulating factor, (GM-CSF)
(Leucomax 400 a kind gift from Novartis, Rueil-Malmaison, France), 50 U/ml
rhTNF-a (kindly provided by Dr Schmidt, Mainz, Germany) and 50 ng/ml
Flt-3 Ligand (Flt-3L, PeproTech, Tebu, Le Perray-en-Yvelines, France). Cells
were then cultured for 7 days.
Chemical exposure of immature DCs. After a culture period of 7 days,
the cells were washed three times before treatment. Cells (1 3 106 cells/ml)
were exposed to either NiSO4 (Sigma, St Louis, MO) or DNCB (Sigma). After
performing dose–response experiments, optimal concentrations of the two
allergens were selected according to their impact on cell viability and on cell
surface marker determined after 24 h of treatment. For cell viability, the highest
concentration should not lead to a cell viability inferior to 75%. In the case of
DNCB (25lM dissolved in dimethyl sulfoxide [DMSO] at final concentration
of 0,05%), treatment was performed for only 30 min and cells were then
washed three times and reincubated for different periods of time. In the case
of NiSO4 cells were treated according to the different times mentioned for
each experiment. BAY 11-7085 (dissolved in DMSO, Calbiochem, AMD
Biosciences, Darmstadt, Germany) was added 1 h before any treatment.
SB203580 (dissolved in distilled water, Calbiochem) was added 30 min before
treatment with NiSO4.
Immunoblotting. Western blot analysis was performed according to stan-
dard procedure, as previously described (Boisleve et al., 2004). Briefly, cells
were washed with cold PBS 1X. Cell lysates were prepared by resuspending the
cell pellet containing 2 3 106 cells in lysis buffer (20mM Tris pH 7.4, 137mM
NaCl, 2mM ethylenediaminetetraacetic acid (EDTA), pH 7.4, 1% Triton X-
100, 25mM b-glycerophosphate, 1mM Na3VO4, 2mM sodium pyrophosphate,
10% glycerol, 1mM phenylmethanesulphonylfluoride (PMSF), 10 lg/ml
aprotinin, 10 lg/ml leupeptin, and 10 lg/ml pepstatin), incubated on ice for
20 min and followed by centrifugation at 17,600 g at 4�C for 20 min. Protein
concentration was determined using the bicinchoninic acid assay (Sigma).
Equal amount of proteins were subjected to 12.5% sodium dodecyl sulfate–
polyacrylamide electrophoresis (SDS-PAGE). The proteins were transferred
onto polyvinylidene difluoride membranes (Amersham Biosciences, Les Ulis,
France) and the membranes were probed with the rabbit anti-IjB-a poly-
clonal Ab (C-21, Santa Cruz Biotechnology, Santa Cruz, CA) or the rabbit
anti-phospho-p38 MAPK (Thr 180/Tyr 182) monoclonal Ab (3D7, Cell
Signaling Technology, Ozyme, St-Quentin-en-Yveline, France) followed by
goat anti-rabbit polyclonal Ab conjugated to horse radish peroxidase (Cell
Signaling Technology). The membranes were stripped for the primary Abs
and reprobed with Ab raised against total p38 MAPK as a loading control
(p38 N20, Santa Cruz Biotechnology). The immunoblots were visualized by
enhanced chemiluminescence (Amersham Biosciences). Densitometric analysis
of the blots was performed using the Quantity One Software (Bio-Rad
Laboratories, CA).
Flow cytometric analysis. Selected monoclonal antibodies (MoAb) were
used to phenotype the DC population. DCs were analyzed with dual-color flow
cytometry using PE-Cy5–labeled mouse anti-human CD86 MoAb (2331 (FUN-1)),
PE-labeled mouse anti-human CD83 MoAb (HB15e), Allophycocyanin (APC)-
labeled mouse anti-human CD40 MoAb (5C3), FITC-labeled mouse anti-
human HLA-DR (G46-6 (L243)), PE-Cy5-labeled mouse IgG1,j isotype
control, PE-labeled mouse IgG1,j isotype control, APC-labeled mouse IgG1,
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 489
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

j isotype control and FITC-labeled mouse IgG2a,j isotype control provided by
BD Biosciences Pharmingen (San Diego, CA). The PE-labeled mouse anti-
human CCR7 MoAb (150503) and the PE-labeled mouse IgG2a isotype control
was purchased from R&D Systems (Lille, France). The labeling procedure was
as follows: cells were washed in cold PBS (containing 0.5% bovine serum
albumin [BSA]) and stained with antibodies in the dark on ice for 30 min,
washed twice with cold PBS (containing 0.5% BSA), once with PBS without
BSA, and resuspended in PBS. Cells were then analyzed on a FACScalibur cell
analyzer (Becton Dickinson, San Jose, CA) using the CellQuest Software
(Becton Dickinson). Cellular debris were eliminated from the analysis using
a gate on the forward and side scatter. For each sample, 10,000 cells were
collected.
Cytokine measurements. The production of IL-8, IL-6, and IL-12p40 was
measured in supernatants collected 24 h after treatment. Levels of IL-8 and IL-6
in supernatants were measured using the BD CBA system and flow cytometry
using a FACSCalibur flow cytometer (BD Biosciences). Data analysis was
performed using the FCAP Array Software (BD Biosciences).
The production of IL-12p40 was measured using an enzyme-linked
immunosorbent assay (ELISA) kit from R&D Systems. The levels of IL-8,
IL-6, and IL-12p40 were calculated by using a standard curve obtained with
recombinant IL-8 (5–5000 pg/ml), recombinant IL-6 (20–5000 pg/ml), or
recombinant IL-12p40 (31.2–2000 pg/ml). The sensitivity of the methods was
5, 52, and 31 pg/ml for IL-8, IL-6, and IL-12p40, respectively.
Preparation of whole cell extract and DNA-binding assay. Following
treatment with NiSO4 or TNF-a, cells were lysed in Nonidet P-40 (NP-40)
hypertonic lysis buffer. In brief, cell pellets from 8 3 106 cells were
resuspended in buffer containing 0.2% (vol/vol) NP-40, 20% (vol/vol)
Glycerol, 20mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid-KOH,
pH 7.9, 420mM NaCl, 1mM dithiothreitol, 1mM sodium orthovanadate,
1mM sodium pyrophosphate, 0.125lM okadaic acid, 62.5mM EDTA, 40lM
ethyleneglycol-bis(2-aminoethylether)-N,N,N#,N#-tetraacetic acid, 0.5mM
PMSF, 1 lg/ml leupeptin, 1 lg/ml aprotinin, 1 lg/ml pepstatin, and incubated
at 4�C for 30 min. Cellular debris were removed by centrifugation at 4�C,
17,600 g for 20 min. The DNA-binding assay was carried out as described
previously (Wery-Zennaro et al., 1999). In brief, the following 5# biotin-
labeled single-stranded oligonucleotides were hybridized: 5#-TTG AGG GGA
CTT TCC CAG G-3#, and 5#-CCT GGG AAA GTC CCC TCA A-3# (MWG)
according to the human NF-jB promoter sequence. Mutated oligonucleotides
(5#-TTG AGG CGA CTT TCC CAG G-3# and 5#-CCT GGG AAA GTC GCC
TCA A-3#) were used for unspecific control binding. DNA-binding proteins
were isolated from 200 lg of whole cell extracts at 4�C for 90 min with 2 lg
double-stranded 5#-biotinylated oligonucleotides coupled to 50 ll of
streptavidin–agarose beads (Sigma). Complexes were washed in binding buffer
and eluted by boiling in reducing sample buffer. The binding proteins were
separated on 8% SDS-PAGE followed by Western blot analysis using an anti-
p65 mouse monoclonal Antibody (Santa Cruz Biotechnology).
Statistical analysis. Dunett’s multicomparison modification of the Stu-
dent’s t-test was used to assess the statistical significance of experimental data
for continuous variables. Experimental data were considered statistically
different from control at p < 0.05.
RESULTS
NiSO4 and DNCB Induce Phenotypical Changesin CD34-DC
Capacity of chemicals to induce the maturation of CD34-DC
was assessed by determining the expression of key membrane
proteins. CD34-DC generated from CD34þ hematopoietic
progenitors were cultured for 7 days with GM-CSF, TNF-a,
and Flt-3L (Boisleve et al., 2004) and incubated in the presence
of either the water-soluble metal NiSO4 or the DMSO-soluble
organic chemical DNCB. CD83, CD86, HLA-DR, and CD40
expression was measured after 24 h. We first performed dose–
response studies to determine the optimal concentration of
chemical for DC activation. The optimal concentrations could
be defined as a concentration that modify DC phenotype with
cell viability not under 75% viable cells. As seen in Figure 1A,
CD83, CD86, HLA-DR, and CD40 expression was induced
dose dependently after treatment with NiSO4. The maximal
activation level was reached when NiSO4 was tested at 500lM.
In this experiment, cell viability remained unchanged at all the
concentrations tested (data not shown).
DNCB was tested at 12.5, 25, and 50lM. The expression of
CD83, CD86, HLA-DR, and CD40 was increased since
12.5lM and reached a maximum at 25lM of DNCB (Fig.
1B). Although cell viability was not affected by the treatment
with DNCB tested at 12.5lM, it was slightly diminished with
25lM of DNCB (75%) and dramatically affected at 50lM
(47%), and corresponded to a loss of marker expression (Fig.
1B). Considering that a minimum of 75% of cell must be viable
to analyze the expression of cell surface markers and that
25lM seemed to be the concentration that induced the
strongest expression of cell surface markers, we choose
25lM for testing DNCB for all the following experiments.
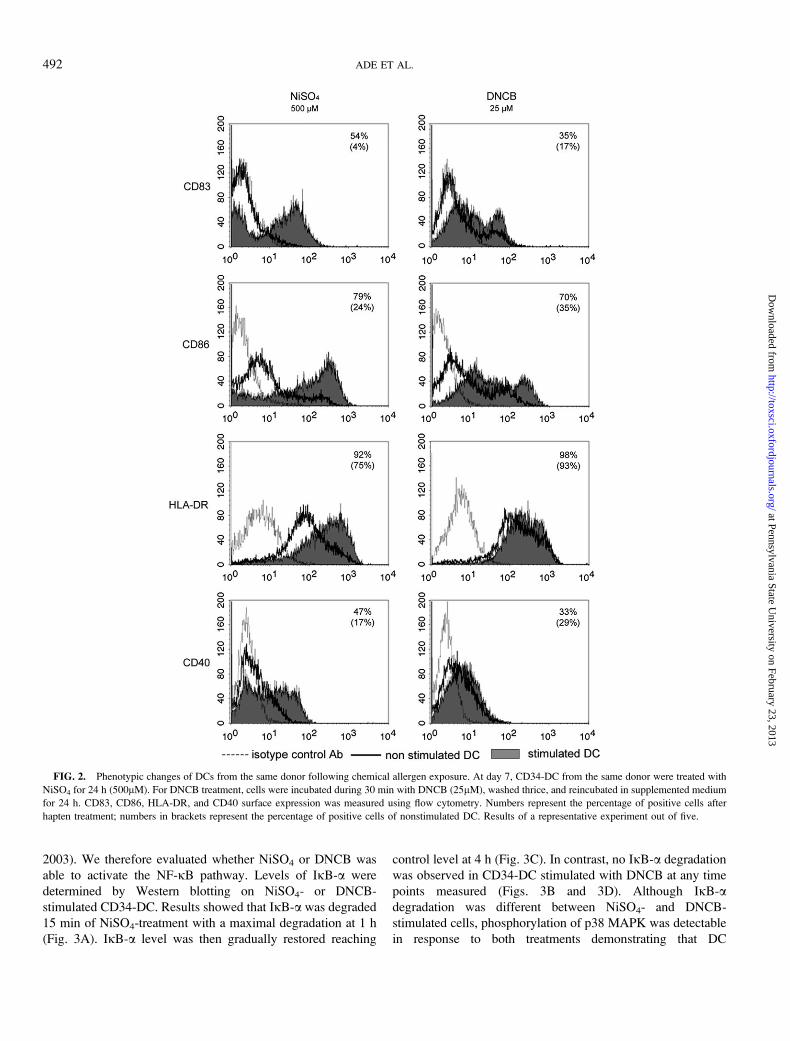
NiSO4 (500lM) and DNCB (25lM) were then tested on the
same donor. In Figure 2 and Table 1, NiSO4 displayed a major
effect on CD83 and CD86 but also upregulated significantly
HLA-DR and CD40. DNCB also induced the expression of
CD83 and CD86. However, HLA-DR and CD40 were only
slightly and not significantly affected in response to DNCB
(Table 2). Our results showed that NiSO4 had a strong effect on
CD83, CD86, HLA-DR, and CD40 expression, whereas
DNCB affected mainly CD83 and CD86 expression.
Cytokine Production after Treatment with NiSO4 or DNCB
Effect of various chemicals on cytokine production by
CD34-DC was assessed using the same protocol as for
phenotype analysis. Results showed that untreated DC did
not produce IL-6 nor IL-12p40 whereas a basal level of IL-8
was detected (Table 3). Upon NiSO4 addition, DC produced
large amounts of IL-6 and IL-12p40 and IL-8 secretion was
significantly increased (p < 0.05). However, in the presence of
DNCB, only IL-8 production was augmented (p < 0.01).
These results clearly showed that NiSO4 was able to induce the
production of cytokines involved in DC function, whereas
DNCB impact was limited to IL-8.
NiSO4 but not DNCB Activates the NF-jB Pathway
Our observations indicated that NiSO4 but not DNCB was
able to upregulate protein expression such as surface CD40 or
cytokine production such as IL-12p40, two events known to
depend strongly on the NF-jB pathway (Laderach et al.,
490 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
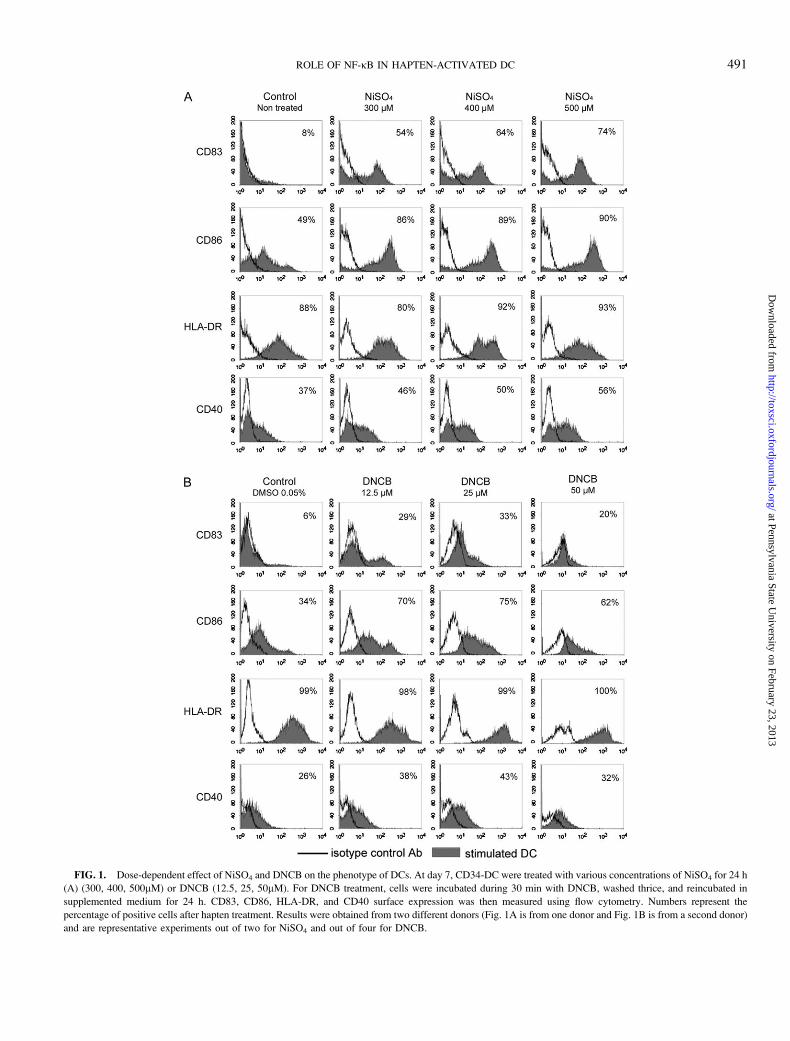
FIG. 1. Dose-dependent effect of NiSO4 and DNCB on the phenotype of DCs. At day 7, CD34-DC were treated with various concentrations of NiSO4 for 24 h
(A) (300, 400, 500lM) or DNCB (12.5, 25, 50lM). For DNCB treatment, cells were incubated during 30 min with DNCB, washed thrice, and reincubated in
supplemented medium for 24 h. CD83, CD86, HLA-DR, and CD40 surface expression was then measured using flow cytometry. Numbers represent the
percentage of positive cells after hapten treatment. Results were obtained from two different donors (Fig. 1A is from one donor and Fig. 1B is from a second donor)
and are representative experiments out of two for NiSO4 and out of four for DNCB.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 491
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
2003). We therefore evaluated whether NiSO4 or DNCB was
able to activate the NF-jB pathway. Levels of IjB-a were
determined by Western blotting on NiSO4- or DNCB-
stimulated CD34-DC. Results showed that IjB-a was degraded
15 min of NiSO4-treatment with a maximal degradation at 1 h
(Fig. 3A). IjB-a level was then gradually restored reaching
control level at 4 h (Fig. 3C). In contrast, no IjB-a degradation
was observed in CD34-DC stimulated with DNCB at any time
points measured (Figs. 3B and 3D). Although IjB-adegradation was different between NiSO4- and DNCB-
stimulated cells, phosphorylation of p38 MAPK was detectable
in response to both treatments demonstrating that DC
FIG. 2. Phenotypic changes of DCs from the same donor following chemical allergen exposure. At day 7, CD34-DC from the same donor were treated with
NiSO4 for 24 h (500lM). For DNCB treatment, cells were incubated during 30 min with DNCB (25lM), washed thrice, and reincubated in supplemented medium
for 24 h. CD83, CD86, HLA-DR, and CD40 surface expression was measured using flow cytometry. Numbers represent the percentage of positive cells after
hapten treatment; numbers in brackets represent the percentage of positive cells of nonstimulated DC. Results of a representative experiment out of five.
492 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
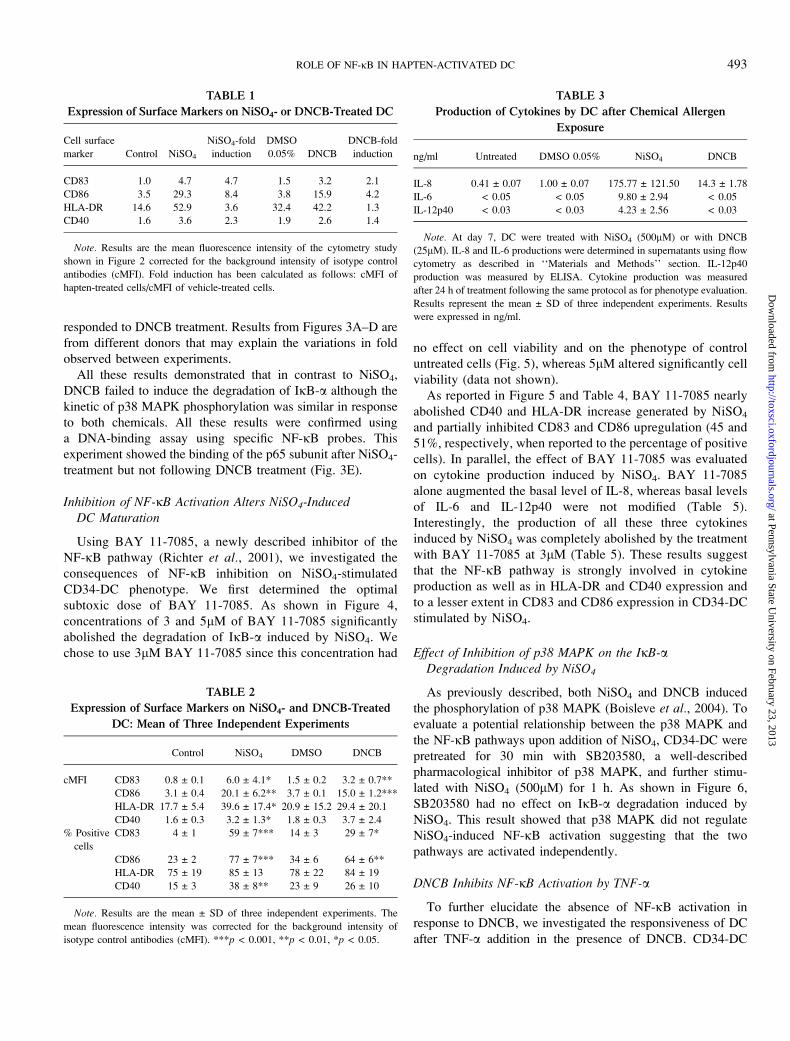
responded to DNCB treatment. Results from Figures 3A–D are
from different donors that may explain the variations in fold
observed between experiments.
All these results demonstrated that in contrast to NiSO4,
DNCB failed to induce the degradation of IjB-a although the
kinetic of p38 MAPK phosphorylation was similar in response
to both chemicals. All these results were confirmed using
a DNA-binding assay using specific NF-jB probes. This
experiment showed the binding of the p65 subunit after NiSO4-
treatment but not following DNCB treatment (Fig. 3E).
Inhibition of NF-jB Activation Alters NiSO4-InducedDC Maturation
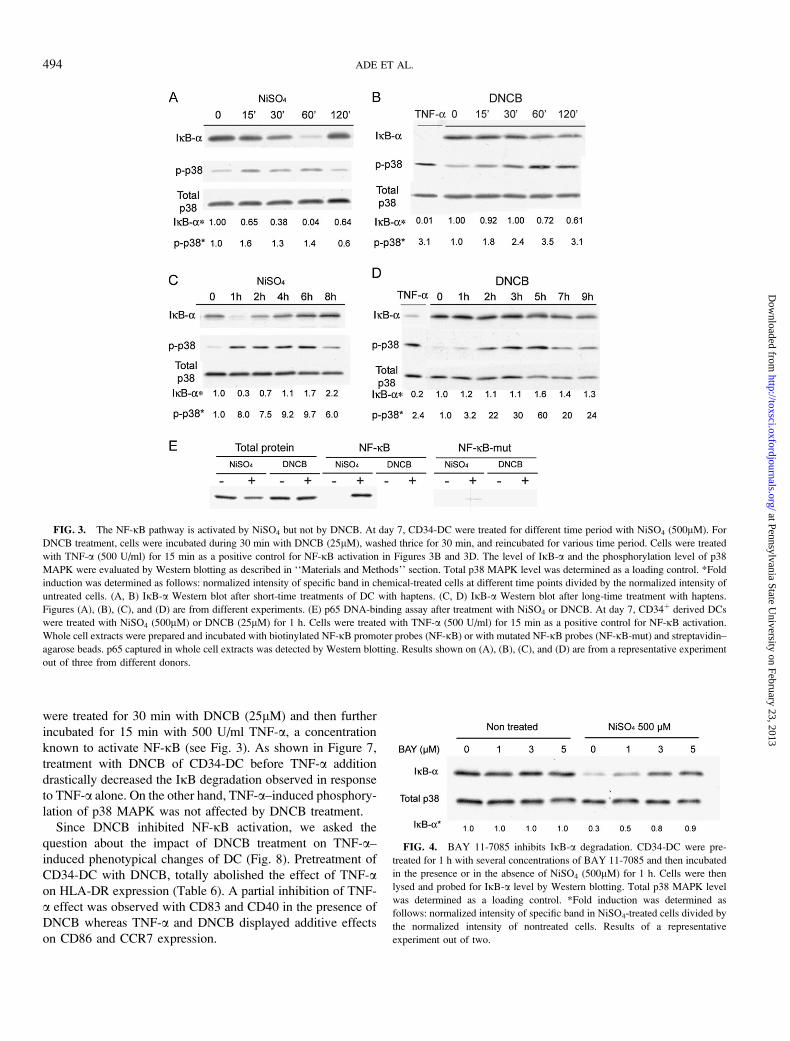
Using BAY 11-7085, a newly described inhibitor of the
NF-jB pathway (Richter et al., 2001), we investigated the
consequences of NF-jB inhibition on NiSO4-stimulated
CD34-DC phenotype. We first determined the optimal
subtoxic dose of BAY 11-7085. As shown in Figure 4,
concentrations of 3 and 5lM of BAY 11-7085 significantly
abolished the degradation of IjB-a induced by NiSO4. We
chose to use 3lM BAY 11-7085 since this concentration had
no effect on cell viability and on the phenotype of control
untreated cells (Fig. 5), whereas 5lM altered significantly cell
viability (data not shown).
As reported in Figure 5 and Table 4, BAY 11-7085 nearly
abolished CD40 and HLA-DR increase generated by NiSO4
and partially inhibited CD83 and CD86 upregulation (45 and
51%, respectively, when reported to the percentage of positive
cells). In parallel, the effect of BAY 11-7085 was evaluated
on cytokine production induced by NiSO4. BAY 11-7085
alone augmented the basal level of IL-8, whereas basal levels
of IL-6 and IL-12p40 were not modified (Table 5).
Interestingly, the production of all these three cytokines
induced by NiSO4 was completely abolished by the treatment
with BAY 11-7085 at 3lM (Table 5). These results suggest
that the NF-jB pathway is strongly involved in cytokine
production as well as in HLA-DR and CD40 expression and
to a lesser extent in CD83 and CD86 expression in CD34-DC
stimulated by NiSO4.
Effect of Inhibition of p38 MAPK on the IjB-aDegradation Induced by NiSO4
As previously described, both NiSO4 and DNCB induced
the phosphorylation of p38 MAPK (Boisleve et al., 2004). To
evaluate a potential relationship between the p38 MAPK and
the NF-jB pathways upon addition of NiSO4, CD34-DC were
pretreated for 30 min with SB203580, a well-described
pharmacological inhibitor of p38 MAPK, and further stimu-
lated with NiSO4 (500lM) for 1 h. As shown in Figure 6,
SB203580 had no effect on IjB-a degradation induced by
NiSO4. This result showed that p38 MAPK did not regulate
NiSO4-induced NF-jB activation suggesting that the two
pathways are activated independently.
DNCB Inhibits NF-jB Activation by TNF-a
To further elucidate the absence of NF-jB activation in
response to DNCB, we investigated the responsiveness of DC
after TNF-a addition in the presence of DNCB. CD34-DC
TABLE 1
Expression of Surface Markers on NiSO4- or DNCB-Treated DC
Cell surface
marker Control NiSO4
NiSO4-fold
induction
DMSO
0.05% DNCB
DNCB-fold
induction
CD83 1.0 4.7 4.7 1.5 3.2 2.1
CD86 3.5 29.3 8.4 3.8 15.9 4.2
HLA-DR 14.6 52.9 3.6 32.4 42.2 1.3
CD40 1.6 3.6 2.3 1.9 2.6 1.4
Note. Results are the mean fluorescence intensity of the cytometry study
shown in Figure 2 corrected for the background intensity of isotype control
antibodies (cMFI). Fold induction has been calculated as follows: cMFI of
hapten-treated cells/cMFI of vehicle-treated cells.
TABLE 2
Expression of Surface Markers on NiSO4- and DNCB-Treated
DC: Mean of Three Independent Experiments
Control NiSO4 DMSO DNCB
cMFI CD83 0.8 ± 0.1 6.0 ± 4.1* 1.5 ± 0.2 3.2 ± 0.7**
CD86 3.1 ± 0.4 20.1 ± 6.2** 3.7 ± 0.1 15.0 ± 1.2***
HLA-DR 17.7 ± 5.4 39.6 ± 17.4* 20.9 ± 15.2 29.4 ± 20.1
CD40 1.6 ± 0.3 3.2 ± 1.3* 1.8 ± 0.3 3.7 ± 2.4
% Positive
cells
CD83 4 ± 1 59 ± 7*** 14 ± 3 29 ± 7*
CD86 23 ± 2 77 ± 7*** 34 ± 6 64 ± 6**
HLA-DR 75 ± 19 85 ± 13 78 ± 22 84 ± 19
CD40 15 ± 3 38 ± 8** 23 ± 9 26 ± 10
Note. Results are the mean ± SD of three independent experiments. The
mean fluorescence intensity was corrected for the background intensity of
isotype control antibodies (cMFI). ***p < 0.001, **p < 0.01, *p < 0.05.
TABLE 3
Production of Cytokines by DC after Chemical Allergen
Exposure
ng/ml Untreated DMSO 0.05% NiSO4 DNCB
IL-8 0.41 ± 0.07 1.00 ± 0.07 175.77 ± 121.50 14.3 ± 1.78
IL-6 < 0.05 < 0.05 9.80 ± 2.94 < 0.05
IL-12p40 < 0.03 < 0.03 4.23 ± 2.56 < 0.03
Note. At day 7, DC were treated with NiSO4 (500lM) or with DNCB
(25lM). IL-8 and IL-6 productions were determined in supernatants using flow
cytometry as described in ‘‘Materials and Methods’’ section. IL-12p40
production was measured by ELISA. Cytokine production was measured
after 24 h of treatment following the same protocol as for phenotype evaluation.
Results represent the mean ± SD of three independent experiments. Results
were expressed in ng/ml.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 493
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
were treated for 30 min with DNCB (25lM) and then further
incubated for 15 min with 500 U/ml TNF-a, a concentration
known to activate NF-jB (see Fig. 3). As shown in Figure 7,
treatment with DNCB of CD34-DC before TNF-a addition
drastically decreased the IjB degradation observed in response
to TNF-a alone. On the other hand, TNF-a–induced phosphory-
lation of p38 MAPK was not affected by DNCB treatment.
Since DNCB inhibited NF-jB activation, we asked the
question about the impact of DNCB treatment on TNF-a–
induced phenotypical changes of DC (Fig. 8). Pretreatment of
CD34-DC with DNCB, totally abolished the effect of TNF-aon HLA-DR expression (Table 6). A partial inhibition of TNF-
a effect was observed with CD83 and CD40 in the presence of
DNCB whereas TNF-a and DNCB displayed additive effects
on CD86 and CCR7 expression.
FIG. 3. The NF-jB pathway is activated by NiSO4 but not by DNCB. At day 7, CD34-DC were treated for different time period with NiSO4 (500lM). For
DNCB treatment, cells were incubated during 30 min with DNCB (25lM), washed thrice for 30 min, and reincubated for various time period. Cells were treated
with TNF-a (500 U/ml) for 15 min as a positive control for NF-jB activation in Figures 3B and 3D. The level of IjB-a and the phosphorylation level of p38
MAPK were evaluated by Western blotting as described in ‘‘Materials and Methods’’ section. Total p38 MAPK level was determined as a loading control. *Fold
induction was determined as follows: normalized intensity of specific band in chemical-treated cells at different time points divided by the normalized intensity of
untreated cells. (A, B) IjB-a Western blot after short-time treatments of DC with haptens. (C, D) IjB-a Western blot after long-time treatment with haptens.
Figures (A), (B), (C), and (D) are from different experiments. (E) p65 DNA-binding assay after treatment with NiSO4 or DNCB. At day 7, CD34þ derived DCs
were treated with NiSO4 (500lM) or DNCB (25lM) for 1 h. Cells were treated with TNF-a (500 U/ml) for 15 min as a positive control for NF-jB activation.
Whole cell extracts were prepared and incubated with biotinylated NF-jB promoter probes (NF-jB) or with mutated NF-jB probes (NF-jB-mut) and streptavidin–
agarose beads. p65 captured in whole cell extracts was detected by Western blotting. Results shown on (A), (B), (C), and (D) are from a representative experiment
out of three from different donors.
FIG. 4. BAY 11-7085 inhibits IjB-a degradation. CD34-DC were pre-
treated for 1 h with several concentrations of BAY 11-7085 and then incubated
in the presence or in the absence of NiSO4 (500lM) for 1 h. Cells were then
lysed and probed for IjB-a level by Western blotting. Total p38 MAPK level
was determined as a loading control. *Fold induction was determined as
follows: normalized intensity of specific band in NiSO4-treated cells divided by
the normalized intensity of nontreated cells. Results of a representative
experiment out of two.
494 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
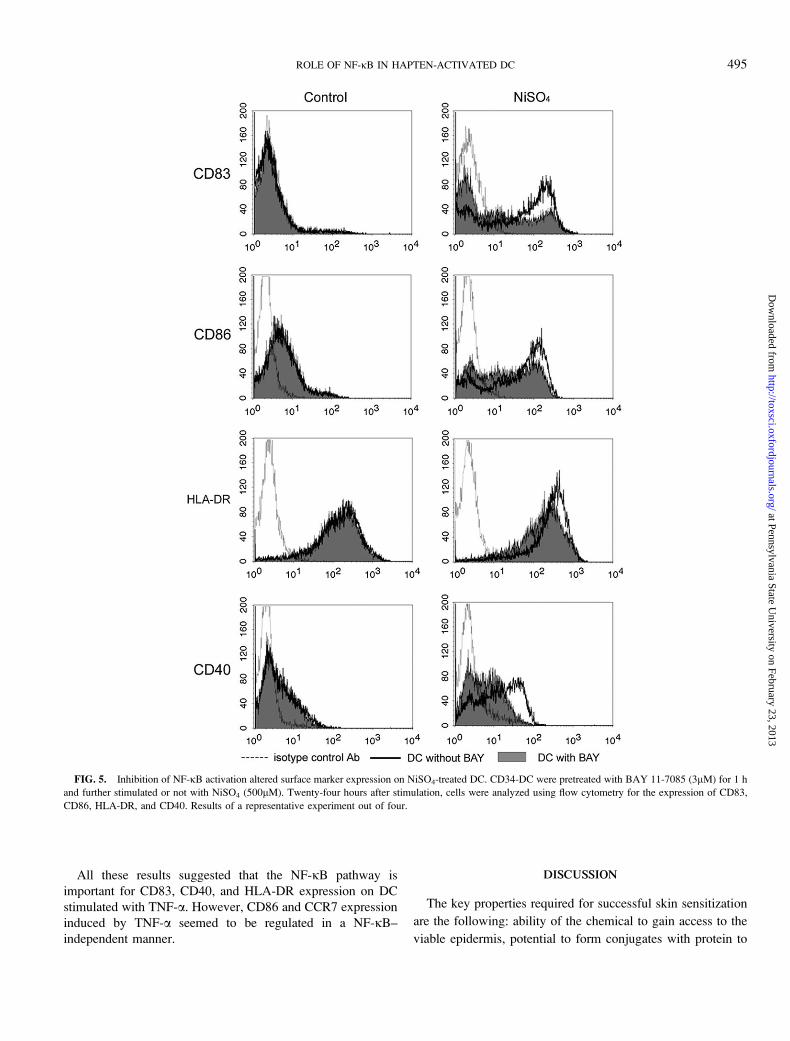
All these results suggested that the NF-jB pathway is
important for CD83, CD40, and HLA-DR expression on DC
stimulated with TNF-a. However, CD86 and CCR7 expression
induced by TNF-a seemed to be regulated in a NF-jB–
independent manner.
DISCUSSION
The key properties required for successful skin sensitization
are the following: ability of the chemical to gain access to the
viable epidermis, potential to form conjugates with protein to
FIG. 5. Inhibition of NF-jB activation altered surface marker expression on NiSO4-treated DC. CD34-DC were pretreated with BAY 11-7085 (3lM) for 1 h
and further stimulated or not with NiSO4 (500lM). Twenty-four hours after stimulation, cells were analyzed using flow cytometry for the expression of CD83,
CD86, HLA-DR, and CD40. Results of a representative experiment out of four.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 495
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

create an immunogen, dermal trauma, and expression of skin
cytokines involved in the induction of cutaneous immune
response and recognition by T lymphocytes of the allergen
complex displayed by immunostimulatory DCs.
DCs are activated by danger signals such as proinflammatory
cytokines, bacterial products, and viruses. Haptens have also
been demonstrated to induce the expression of markers related
to function and maturation of DC, the production of proin-
flammatory cytokines, and the phosphorylation of members of
the MAPK family. However, it is not clear if haptens are also
able to activate the NF-jB pathway like danger signals.
In this study, NiSO4 upregulated the expression of CD83,
CD86, HLA-DR, and CD40 in CD34-DC. However, the effect
of DNCB was limited to the expression of CD83 and CD86.
Using MoDC, Staquet et al. (2004) have also observed that, in
contrast to NiSO4, DNCB did not induce the expression of
CD40. Several authors have also reported that DNCB
compared to NiSO4 is a weak inducer of HLA-DR expression
(Aiba et al., 1997; De Smedt et al., 2005; Staquet et al., 2004).
Moreover, Aiba et al. (2000) showed that MoDC differentiated
in the presence of transforming growth factor-b responded to
DNCB by increasing the expression of CD86 and CD83 but
not of HLA-DR.
In parallel to phenotypical changes, NiSO4 showed a good
capacity to induce the production of cytokines such as IL-6,
IL-8, and IL-12p40 although DNCB only increased the produc-
tion of IL-8. Several authors have also reported discrepancies
in cytokine production between Nickel and DNCB. Aiba et al.(2003) have reported that NiCl2 induced the production of IL-8
FIG. 6. p38 MAPK does not regulate NiSO4-induced NF-jB activation.
CD34-DC were pretreated for 30 min with or without 20lM SB203580 and
further treated in the presence or in the absence of 500lM NiSO4 for 1 h. Cells
were then lysed and probed for IjB-a level by Western blotting. Total p38
MAPK level was determined as a loading control. *Fold induction was
determined as follows: normalized intensity of specific band in treated cells
divided by the normalized intensity of in nontreated cells. Blots are
representative of results obtained in three independent experiments.
FIG. 7. DNCB inhibits TNF-a–induced IjB-a degradation. CD34-DC
were pretreated in the presence or in the absence of DNCB (25lM) for 30 min
and then further stimulated or not with TNF-a (500 U/ml) for 15 min. Cells
were then lysed and probed for IjB-a level and phosphorylation of p38 MAPK
by Western blotting. Total p38 MAPK level was determined as a loading
control. *Fold induction was determined as follows: normalized intensity of
specific band in treated cells divided by the normalized intensity of non-
treated cells. Blots are representative of results obtained in three independent
experiments.
TABLE 4
Inhibition of the NF-kB Pathway Altered the Phenotype of
NiSO4-Stimultated CD34-DC
Control NiSO4
BAY
11-7085
NiSO4 þBAY 11-7085
%
Inhibition
cMFI CD83 1.0 13.1 1.2 4.3 74
CD86 2.3 16.7 2.7 8.1 62
HLA-DR 60.3 105.2 53.1 63.9 76
CD40 1.5 5.4 1.9 2.2 92
% Positive
cells
CD83 3.8 70.1 5.6 42.2 45
CD86 24.6 80.7 35.2 62.9 51
HLA-DR 95.6 98.1 96.1 95.2 0
CD40 11.0 43 10.3 12.7 92
Note. The results showed in this table are the cMFI and the percentage of
positive cells (% (þ) cells) from the cytometry study shown in Figure 5. Cells
were not treated (A) or treated with either NiSO4 (B), BAY 11-7085 (C), or
NiSO4 with BAY 11-7085 (D). % Inhibition was calculated using the following
formula: 100 � [(D � C)/(B � A)] 3 100.
TABLE 5
Inhibition of the NF-kB Pathway Altered Cytokine Production
Induced by NiSO4
ng/ml Untreated
BAY
11-7085 NiSO4
NiSO4 þBAY 11-7085
IL-8 0.3 42.2 112.8 46.3
IL-6 < 0.05 < 0.05 8.5 < 0.05
IL-12p40 < 0.03 < 0.03 1.8 < 0.03
Note. CD34-derived DCs were pretreated in the presence or in the absence of
BAY 11-7085 (3lM) for 1 h and further stimulated or not with NiSO4
(500lM). IL-8 and IL-6 productions were determined in supernatants using
flow cytometry as described in ‘‘Materials and Methods’’ section. IL-12p40
production was measured by ELISA. IL-8, IL-6, and IL-12p40 productions
were measured after 24 h of treatment. Results of a representative experiment
out of four. Results were expressed in ng/ml.
496 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from
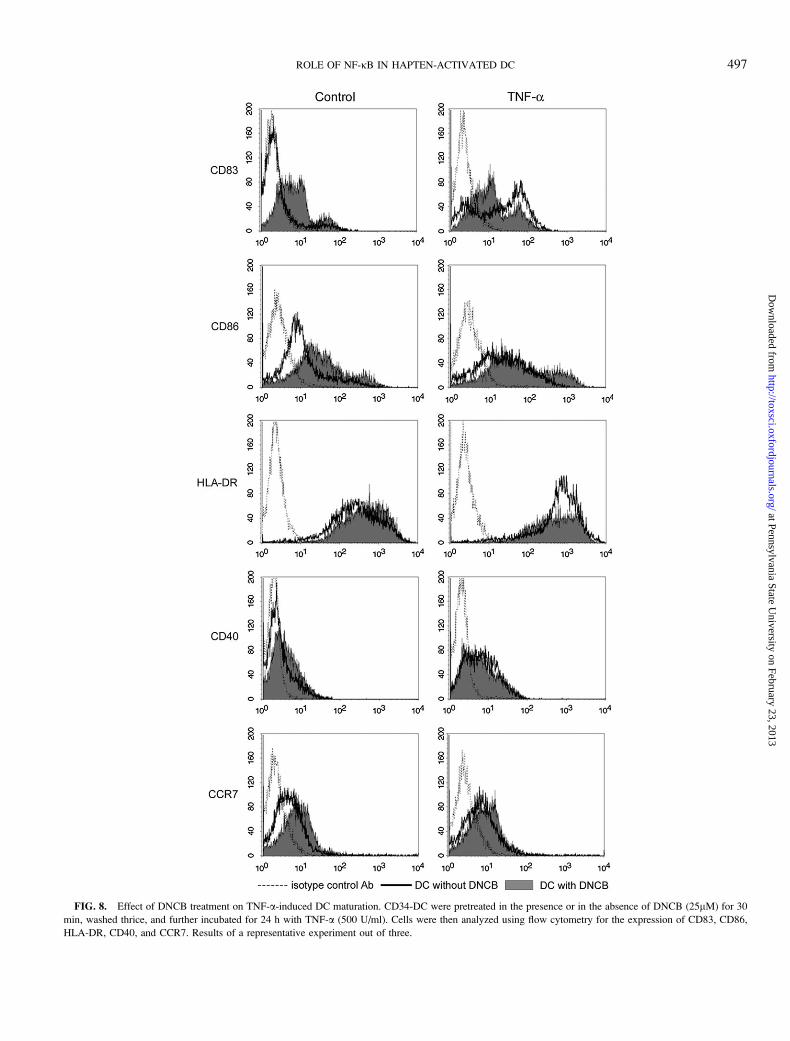
FIG. 8. Effect of DNCB treatment on TNF-a-induced DC maturation. CD34-DC were pretreated in the presence or in the absence of DNCB (25lM) for 30
min, washed thrice, and further incubated for 24 h with TNF-a (500 U/ml). Cells were then analyzed using flow cytometry for the expression of CD83, CD86,
HLA-DR, CD40, and CCR7. Results of a representative experiment out of three.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 497
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

and IL-12p40 in MoDC, whereas DNCB only affected the
production of IL-8. Furthermore, IL-6 has been also shown to
be produced by MoDC in response to NiSO4 but not to DNCB
(Tuschl et al., 2000). Recently, Verheyen et al. (2005) showed
similar results in CD34-DC based on the messenger RNA
expression of IL-8 and IL-6 in response to DNCB and NiCl2;
NiCl2 induced both IL-8 and IL-6 gene expressions, whereas
DNCB induced only IL-8 gene expression.
Based on these results, we hypothesized that differences
observed between NiSO4 and DNCB in DC maturation could
involve the NF-jB pathway. IL-8, IL-12p40, CD83, and CD86
promoters contain NF-jB consensus sequences that are
required for their expression (Berchtold et al. 2002; Hoffmann
et al., 2002; Li et al., 2000; Yoshimoto et al., 1996).
Furthermore, using adenoviral transfert or inhibitors of IjB-aor RNAi for p50, NF-jB has been shown to be essential for
HLA-DR, CD86, CD83, and CD40 expression as well as
cytokine production such as IL-6, IL-12, and IL-8 in MoDC in
response to LPS, TNF-a, and CD40L (Laderach et al., 2003;
O’Sullivan and Thomas, 2002; Yoshimura et al., 2001). Our
results showed that NiSO4 was able to induce the degradation
of IjB-a as well as the binding of the p65 subunit on specific
DNA probes. This effect was maximal after 1 h of stimulation
with NiSO4. However, DNCB neither induced the degradation
of IjB-a nor the binding of p65 on specific DNA probes. Aiba
et al. (2003) have also observed in MoDC that nickel but not
DNCB activated the phosphorylation of IjB-a with compara-
ble kinetics. These results suggested that the difference in
NF-jB activation observed between NiSO4 and DNCB
correlated to the differences in phenotype and in cytokine
production observed with these two chemicals.
To test whether activation of NF-jB was involved in hapten-
induced DC maturation, we used the inhibitor BAY 11-7085,
known to block the phosphorylation of IjB-a and to prevent its
degradation (Richter et al., 2001). Experiments were conducted
only on NiSO4-treated cells since DNCB does not activate NF-
jB in our model. We found that BAY 11-7085 effectively
blocked NiSO4-induced IjB-a degradation in our model. In
DC activated by NiSO4 and treated with BAY 11-7085,
increased expressions of HLA-DR and CD40 as well as IL-8,
IL-6, and IL-12p40 production were nearly completely
abrogated, but CD83 and CD86 potentialization was only
partially inhibited. Using pyrrolidinedithiocarbamate, another
inhibitor of NF-jB, Aiba et al. (2003) showed in MoDC
a slight inhibition of CD86 and no effect on CD83 expression
induced by nickel. To further elucidate the signaling pathway
involved in NF-jB activation, we studied the relationship
between p38 MAPK activation and NF-jB. It has previously
been reported that stimulators of p38 MAPK, such as TNF-acan augment the activity of the NF-jB pathway via cross-talk
between MKK6 and IKKb (Craig et al., 2000). Our results
showed that inhibition of p38 MAPK did not alter the
degradation of IjB-a induced by NiSO4 suggesting that p38
MAPK did not participate to this NF-jB activation by NiSO4
in CD34-DC. This result is in agreement with recent papers
reporting that p38 MAPK and NF-jB pathways are in-
dependent (Cloutier et al., in press).
Our next question was to understand why DNCB did not
activate the NF-jB pathway, whereas p38 MAPK was fully
activated by this molecule. In the present study, we showed
that pretreatment with DNCB blocked the degradation of IjB-ainduced by TNF-a suggesting that DNCB acts as a potent
inhibitor of the NF-jB pathway. Brennan and O’Neill (1998)
have reported that DCNB, a structural analog of DNCB known
to bind on thiol functions, blocked the activation of NF-jB by
TNF-a in Jurkat cells. Other authors have also reported that
curcumin and nordihydroguaiaritic acid, two CS, inhibited the
degradation of IjB-a as we observed with DNCB (Brennan
and O’Neill, 1998). DNCB has also been previously described
to be an irreversible inhibitor of thioredoxin reductase, leading
to a strong activation of nicotinamide adenine dinucleotide
phosphate, reduced oxidase, production of reactive oxygen
intermediates (ROI), and suppression of NF-jB–dependent
transcription in HELA cells (Arner et al., 1995; Nordberg
et al., 1998). As previously described for high concentrations
of H2O2, strong production of ROI by DNCB could alter
NF-jB activation by inhibiting nuclear translocation of NF-jB
through preservation of cytoplasmic IjB-a levels (Korn et al.,2001).
Alteration of the NF-jB pathway induced by DNCB resulted
in a decrease in CD83, CD40, and HLA-DR expressions in DC
treated with TNF-a. These results confirmed that expressions
of CD83, CD40, and HLA-DR were dependent on NF-jB.
Interestingly, CD86 expression induced by TNF-a was further
augmented in the presence of DNCB. CD86 was the marker the
less affected by treatment with BAY 11-7085 in NiSO4-treated
DC suggesting that CD86 expression is less dependent on
NF-jB. We have previously described that CCR7 expression
was upregulated following exposure of CD34-DC to DNCB,
TABLE 6
Effect of DNCB on TNF-a–Induced Surface Marker Expression
Control
(DMSO) DNCB TNF-aDNCB þ
TNF-a
cMFI CD83 1.3 7.3 16.5 11.0
CD86 4.0 6.3 6.8 12.8
HLA-DR 87.5 103.8 221.8 91.6
CD40 1.5 1.7 3.1 2.6
CCR7 2.0 3.5 2.8 3.9
% Positive cells CD83 8.9 25.8 63.3 41.3
CD86 29.9 55.2 56.8 65.9
HLA-DR 98.4 98.5 98.3 98.1
CD40 19.4 16.6 49.3 33.4
CCR7 13.0 30.7 20.7 36.5
Note. The results showed in this table are the isotype control antibodies
(cMFI) and the percentage of positive cells from the cytometry study shown in
Figure 8.
498 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

and that an autocrine loop involving TNF-a was necessary
(Boisleve et al., 2004). Here we showed that DNCB did not
affect CCR7 expression induced by TNF-a confirming that
TNF-a plays a role in CCR7 expression in a NF-jB–
independent manner.
One important question remains if the effects we observed
could reflect that DNCB is a potent allergen and irritant,
whereas nickel is a moderate allergen and irritant. In human
DCs, results obtained with irritants such as SDS or benzalko-
nium chloride did not show any phenotype modification or
activation of signaling pathways such as MAPK or NF-jB
(Aiba et al., 1997; De Smedt et al., 2001; Mizuashi et al.,2005). In a recent paper, Toebak et al. (2005) have shown in
MoDC that allergens were able to increase IL-8 secretion,
whereas irritants rather decrease it. These results may suggest
that DCs can distinguish a chemical irritant from a chemical
sensitizer leading to the activation of specific signaling
pathways. However, the strong irritant potential of DNCB is
due to its capacity of inducing oxidative stress leading to the
activation of JNK and p38 MAPK and this event is directly
linked to DC phenotype alteration (Aiba et al., 2003; Boisleve
et al., 2004). Indeed, Aiba et al. have shown in MoDC that
DNCB is more potent than NiCl2 for reducing the GSH/GSSG
ratio (Mizuashi et al., 2005) and the authors suggested
a correlation with p38 MAPK activation. On the other hand,
Handley et al. (2005) have shown that H2O2 generates a strong
oxidative stress without altering DC phenotype. All these
results suggest that sensitizers probably need to produce some
oxidative stress to activate DC but this property is not sufficient
to induce full DC maturation.
In summary, our results suggested a causal relationship
between activation of NF-jB by chemical sensitizers and
alteration of DC phenotype. NiSO4 activates NF-jB and
inducing extensive modifications of DC phenotype. However,
DNCB induced the expression of CD86, CD83, CCR7, and the
production of IL-8 without activating NF-jB suggesting that
other signaling pathways such as MAPK are mainly involved.
The signaling pathways involved upon NiSO4 or DNCB
exposure provide an explanation for the phenotypical differ-
ences observed in DC maturation induced by these two CS.
Finally, a figure showing the signaling pathways already
identified as being involved in the activation of DC by CS, as
well as their possible interactions, is proposed in Figure 9.
FUNDING
Agence Francxaise de Securite Sanitaire des Produits de Sante
(AFSSAPS).
ACKNOWLEDGMENTS
The authors would like to acknowledge Silvia Martinozzi-Teissier,
Alexandre Larange, and Jacques Bertoglio for their technical advices and
helpful suggestions.
REFERENCES
Aiba, S., Manome, H., Nakagawa, S., Mollah, Z. U., Mizuashi, M., Ohtani, T.,
Yoshino, Y., and Tagami, H. (2003). p38 Mitogen-activated protein kinase
FIG. 9. Diagram representation of the pathways involved in CD34-DC activation following NiSO4 or DNCB treatment (from this work and from Boisleve
et al., 2004, 2005). NiSO4 induces the expression of CD83, CD86, HLA-DR, and CD40 and the production of IL-8, IL-6, and IL-12p40 through the activation of
both NF-jB and MAPK (p38 MAPK, JNK). CCR7 is induced by NiSO4 with MAPK playing an important role. The role of NF-jB is not known in this case.
DNCB treatment upregulates CD86, CD83, and CCR7 and increases the secretion of IL-8. An autocrine loop of TNF-a dependent on MAPK activation is
necessary for the expression of CCR7 and CD83.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 499
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

and extracellular signal-regulated kinases play distinct roles in the activa-
tion of dendritic cells by two representative haptens, NiCl2 and 2,4-
dinitrochlorobenzene. J. Invest. Dermatol. 120, 390–399.
Aiba, S., Manome, H., Yoshino, Y., and Tagami, H. (2000). In vitro treat-
ment of human transforming growth factor-beta1-treated monocyte-derived
dendritic cells with haptens can induce the phenotypic and functional
changes similar to epidermal Langerhans cells in the initiation phase of
allergic contact sensitivity reaction. Immunology 101, 68–75.
Aiba, S., Terunuma, A., Manome, H., and Tagami, H. (1997). Dendritic
cells differently respond to haptens and irritants by their production of
cytokines and expression of co-stimulatory molecules. Eur. J. Immunol. 27,
3031–3038.
Arner, E. S., Bjornstedt, M., and Holmgren, A. (1995). 1-Chloro-
2,4-dinitrobenzene is an irreversible inhibitor of human thioredoxin reduc-
tase. Loss of thioredoxin disulfide reductase activity is accompanied by a
large increase in NADPH oxidase activity. J. Biol. Chem. 270, 3479–3482.
Arrighi, J. F., Rebsamen, M., Rousset, F., Kindler, V., and Hauser, C. (2001).
A critical role for p38 mitogen-activated protein kinase in the maturation of
human blood-derived dendritic cells induced by lipopolysaccharide, TNF-
alpha, and contact sensitizers. J. Immunol. 166, 3837–3845.
Attar, R. M., Macdonald-Bravo, H., Raventos-Suarez, C., Durham, S. K., and
Bravo, R. (1998). Expression of constitutively active IkappaB beta in T cells
of transgenic mice: Persistent NF-kappaB activity is required for T-cell
immune responses. Mol. Cell. Biol. 18, 477–487.
Berchtold, S., Muhl-Zurbes, P., Maczek, E., Golka, A., Schuler, G., and
Steinkasserer, A. (2002). Cloning and characterization of the promoter
region of the human CD83 gene. Immunobiology 205, 231–246.
Boisleve, F., Kerdine-Romer, S., and Pallardy, M. (2005). Implication of the
MAPK pathways in the maturation of human dendritic cells induced by
nickel and TNF-alpha. Toxicology 206, 233–244.
Boisleve, F., Kerdine-Romer, S., Rougier-Larzat, N., and Pallardy, M. (2004).
Nickel and DNCB induce CCR7 expression on human dendritic cells
through different signalling pathways: Role of TNF-alpha and MAPK.
J. Invest. Dermatol. 123, 494–502.
Brennan, P., and O’Neill, L. A. (1998). Inhibition of nuclear factor kappaB by
direct modification in whole cells—Mechanism of action of nordihydro-
guaiaritic acid, curcumin and thiol modifiers. Biochem. Pharmacol. 55,
965–973.
Bour, H., Peyron, E., Gaucherand, M., Garrigue, J. L., Desvignes, C.,
Kaiserlian, D., Revillard, J. P., and Nicolas, J. F. (1995). Major
histocompatibility complex class I-restricted CD8þ T cells and class
II-restricted CD4þ T cells, respectively, mediate and regulate contact
sensitivity to dinitrofluorobenzene. Eur. J. Immunol. 25, 3006–3010.
Caux, C., Vanbervliet, B., Massacrier, C., Dezutter-Dambuyant, C., de Saint-
Vis, B., Jacquet, C., Yoneda, K., Imamura, S., Schmitt, D., and
Banchereau, J. (1996). CD34þ hematopoietic progenitors from human cord
blood differentiate along two independent dendritic cell pathways in
response to GM-CSFþTNF alpha. J. Exp. Med. 184, 695–706.
Cloutier, A., Ear, T., Blais-Charron, E., Dubois, C. M., and McDonald, P. P.
Differential involvement of NF-{kappa}B and MAP kinase pathways in the
generation of inflammatory cytokines by human neutrophils. J. Leukoc. Biol.
81, 567–577.
Craig, R., Larkin, A., Mingo, A. M., Thuerauf, D. J., Andrews, C.,
McDonough, P. M., and Glembotski, C. C. (2000). p38 MAPK and NF-
kappa B collaborate to induce interleukin-6 gene expression and release.
Evidence for a cytoprotective autocrine signaling pathway in a cardiac
myocyte model system. J. Biol. Chem. 275, 23814–23824.
De Smedt, A. C., Van Den Heuvel, R. L., Van Tendeloo, V. F.,
Berneman, Z. N., and Schoeters, G. E. (2005). Capacity of CD34þprogenitor-derived dendritic cells to distinguish between sensitizers and
irritants. Toxicol. Lett. 156, 377–389.
De Smedt, A. C., Van Den Heuvel, R. L., Zwi Berneman, N., and
Schoeters, G. E. (2001). Modulation of phenotype, cytokine production
and stimulatory function of CD34þ-derived DC by NiCl(2) and SDS.
Toxicol. In Vitro 15, 319–325.
Gallucci, S., and Matzinger, P. (2001). Danger signals: SOS to the immune
system. Curr. Opin. Immunol. 13, 114–119.
Goebeler, M., Roth, J., Brocker, E. B., Sorg, C., and Schulze-Osthoff, K.
(1995). Activation of nuclear factor-kappa B and gene expression in human
endothelial cells by the common haptens nickel and cobalt. J. Immunol. 155,
2459–2467.
Handley, M. E., Thakker, M., Pollara, G., Chain, B. M., and Katz, D. R. (2005).
JNK activation limits dendritic cell maturation in response to reactive oxy-
gen species by the induction of apoptosis. Free Radic. Biol. Med. 38,
1637–1652.
Hoffmann, E., Dittrich-Breiholz, O., Holtmann, H., and Kracht, M. (2002).
Multiple control of interleukin-8 gene expression. J. Leukoc. Biol. 72,
847–855.
Korn, S. H., Wouters, E. F., Vos, N., and Janssen-Heininger, Y. M. (2001).
Cytokine-induced activation of nuclear factor-kappa B is inhibited by
hydrogen peroxide through oxidative inactivation of IkappaB kinase. J. Biol.
Chem. 276, 35693–35700.
Kuhn, U., Brand, P., Willemsen, J., Jonuleit, H., Enk, A. H., van Brandwijk-
Petershans, R., Saloga, J., Knop, J., and Becker, D. (1998). Induction of
tyrosine phosphorylation in human MHC class II-positive antigen-presenting
cells by stimulation with contact sensitizers. J. Immunol. 160, 667–673.
Laderach, D., Compagno, D., Danos, O., Vainchenker, W., and Galy, A.
(2003). RNA interference shows critical requirement for NF-kappa B p50 in
the production of IL-12 by human dendritic cells. J. Immunol. 171,
1750–1757.
Li, J., Colovai, A. I., Cortesini, R., and Suciu-Foca, N. (2000). Cloning and
functional characterization of the 5#-regulatory region of the human CD86
gene. Hum. Immunol. 61, 486–98.
Mizuashi, M., Ohtani, T., Nakagawa, S., and Aiba, S. (2005). Redox imbalance
induced by contact sensitizers triggers the maturation of dendritic cells.
J. Invest. Dermatol. 124, 579–586.
Nordberg, J., Zhong, L., Holmgren, A., and Arner, E. S. (1998). Mammalian
thioredoxin reductase is irreversibly inhibited by dinitrohalobenzenes by
alkylation of both the redox active selenocysteine and its neighboring
cysteine residue. J. Biol. Chem. 273, 10835–10842.
O’Sullivan, B. J., and Thomas, R. (2002). CD40 ligation conditions dendritic
cell antigen-presenting function through sustained activation of NF-kappaB.
J. Immunol. 168, 5491–5498.
Richter, G., Hayden-Ledbetter, M., Irgang, M., Ledbetter, J. A., Westermann, J.,
Korner, I., Daemen, K., Clark, E. A., Aicher, A., and Pezzuto, A. (2001).
Tumor necrosis factor-alpha regulates the expression of inducible costimulator
receptor ligand on CD34(þ) progenitor cells during differentiation into antigen
presenting cells. J. Biol. Chem. 276, 45686–45693.
Rougier, N., Redziniak, G., Mougin, D., Schmitt, D., and Vincent, C. (2000).
In vitro evaluation of the sensitization potential of weak contact allergens
using langerhans-like dendritic cells and autologous T cells. Toxicology 145,
73–82.
Sallusto, F., and Lanzavecchia, A. (1994). Efficient presentation of soluble
antigen by cultured human dendritic cells is maintained by granulocyte/
macrophage colony-stimulating factor plus interleukin 4 and downregulated
by tumor necrosis factor alpha. J. Exp. Med. 179, 1109–1118.
Staquet, M. J., Sportouch, M., Jacquet, C., Schmitt, D., Guesnet, J., and Peguet-
Navarro, J. (2004). Moderate skin sensitizers can induce phenotypic
changes on in vitro generated dendritic cells. Toxicol. In Vitro 18,
493–500.
Toebak, M. J., Pohlmann, P. R., Sampat-Sardjoepersad, S. C., von
Blomberg, B. M., Bruynzeel, D. P., Scheper, R. J., Rustemeyer, T., and
500 ADE ET AL.
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from

Gibbs, S. (2005). CXCL8 secretion by dendritic cells predicts contact
allergens from irritants. Toxicol. In Vitro 20, 117–124.
Tuschl, H., and Kovac, R. (2001). Langerhans cells and immature dendritic cells
as model systems for screening of skin sensitizers. Toxicol. In Vitro 15, 327–331.
Tuschl, H., Kovac, R., and Weber, E. (2000). The expression of surface
markers on dendritic cells as indicators for the sensitizing potential of
chemicals. Toxicol. In Vitro 14, 541–549.
Verhasselt, V., Buelens, C., Willems, F., De Groote, D., Haeffner-
Cavaillon, N., and Goldman, M. (1997). Bacterial lipopolysaccharide
stimulates the production of cytokines and the expression of cos-
timulatory molecules by human peripheral blood dendritic cells: Evidence
for a soluble CD14-dependent pathway. J. Immunol. 158, 2919–2925.
Verhasselt, V., Vanden Berghe, W., Vanderheyde, N., Willems, F.,
Haegeman, G., and Goldman, M. (1999). N-acetyl-L-cysteine inhibits
primary human T cell responses at the dendritic cell level: Association with
NF-kappaB inhibition. J. Immunol. 162, 2569–2574.
Verheyen, G. R., Schoeters, E., Nuijten, J. M., Van Den Heuvel, R. L.,
Nelissen, I., Witters, H., Van Tendeloo, V. F., Berneman, Z. N., and
Schoeters, G. E. (2005). Cytokine transcript profiling in CD34þ-progenitor
derived dendritic cells exposed to contact allergens and irritants. Toxicol.
Lett. 155, 187–194.
Wery-Zennaro, S., Letourneur, M., David, M., Bertoglio, J., and Pierre, J.
(1999). Binding of IL-4 to the IL-13Ralpha(1)/IL-4Ralpha receptor complex
leads to STAT3 phosphorylation but not to its nuclear translocation. FEBS
Lett. 464, 91–96.
Yoshimoto, T., Kojima, K., Funakoshi, T., Endo, Y., Fujita, T., and
Nariuchi, H. (1996). Molecular cloning and characterization of murine
IL-12 genes. J. Immunol. 156, 1082–1088.
Yoshimura, S., Bondeson, J., Foxwell, B. M., Brennan, F. M., and
Feldmann, M. (2001). Effective antigen presentation by dendritic cells is
NF-kappaB dependent: Coordinate regulation of MHC, co-stimulatory
molecules and cytokines. Int. Immunol. 13, 675–683.
ROLE OF NF-jB IN HAPTEN-ACTIVATED DC 501
at Pennsylvania State University on February 23, 2013
http://toxsci.oxfordjournals.org/D
ownloaded from





























