Embed Size (px)
Citation preview

THE JOURNAL 0 1991 by The American Society for Biochemistry and
OF BIOLOGICAL CHEMISTRY Molecular Biology, Inc.
Vol. 266. No . 17, Issue of June 15, PP. 10899-10905. 1991 Printed in U.S.A.
Nitric Oxide Reductase PURIFICATION FROM PARACOCCUS DENITRIFICANS WITH USE OF A SINGLE COLUMN AND SOME CHARACTERISTICS*
(Received for publication, December 20, 1990)
Marina DermastiaS, Tom Turks, and Thomas C. HollocherV From the Department of Biochemistgv, Brandeis University, Waltham, Massachusetts 02254
Nitric oxide reductase was purified from Paracoccus denitrificans very nearly to homogeneity by a simple method that involved the use of octyl glucoside to sol- ubilize the enzyme from membranes and required a single hydroxyapatite column. The enzyme had spe- cific activities of about 10 pmol NO reduced X min" X mg" at pH 6.5 in an amperometric assay system using phenazine methosulfate/ascorbate as the reducing agent and about 22 pmol NO reduced X min" X mg" at pH 5.0, which is the optimum pH. These values are based on average rates over kinetically complex pro- gress curves and would be about three times greater if based on maximum rate values. The enzyme appeared to be reversibly inhibited by NO,, and to have a K , too low (probably 51 p ~ ) to measure reliably by the am- perometric method. The effective second-order rate constant of the enzyme lay within 1 to 2 orders of magnitude of the diffusion controlled limit. The en- zyme was composed of a tight complex of two cyto- chromes: a cytochrome c (Mr = 17, 500) and a cyto- chrome b (Mr = 38,000). The mole ratios of cytochrome c to cytochrome b and M, 17,500 peptide to M, 38,000 peptide were both about 1.7, and the heme content was about 3 mo1/73,000 g (38,000 + 2(17, 500)). Each subunit therefore contained only one heme group. The M, 38,000 peptide aggregated when heated in the sam- ple buffer used for sodium dodecyl sulfate-polyacryl- amide gel electrophoresis. In addition to the ascorbate- based activity, the enzyme showed a little NADH-NO oxidoreductase activity which was not inhibited by antimycin A. The enzyme lost activity with a half-life of about 2 days at 4 "C but could be preserved at -20 "C and in liquid nitrogen. It seemed not to be inactivated by aerobic solutions.
These observations, and the recent ones by Carr and Ferguson (Carr, G . J., and Ferguson, S . J. (1990) Biochem. J. 269, 423-429) with a partially purified preparation of nitric oxide reductase, establish that the enzyme from Pa. denitrificans is a cytochrome bc com- plex which resembles that from Pseudomonas stutzeri (Heiss, B., Frunzke, K., and Zumft, W. G . (1989) J. Bacteriol. 171, 3288-3297). There would appear to
* This investigation was supported by Grant DCB 88-16273 from the National Science Foundation and fellowships (to M. D. and T. T.) from the Research Council of Slovenia, Yugoslavia. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "adver- tisement'' in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Inst. of Biochemistry, Medical Faculty, Univer- sity of Ljubljana, 61000 Ljubljana, Yugoslavia.
§ Present address: Dept. of Biology, Biotechnical Faculty, Univer- sity of Ljubljana, 61000 Ljubljana, Yugoslavia.
ll To whom correspondence should be addressed. Tel.: 617-736- 2345.
be no functional relationship between nitric oxide re- ductase and a M, = 34,000 peptide of Pa. denitrificans membranes reported previously to be present in puri- fied preparations of a nitric oxide reductase (Hoglen, J., and Hollocher, T. C. (1989) J. Biol. Chem. 264, 7556-7563).
Hoglen and Hollocher (1) described a procedure for enrich- ing nitric oxide reductase activity in vesicles from denitrifying Paracoccus denitrificans. Simultaneously a protein(s) with M , = 34,000 was enriched and finally comprised 90-95% of the total protein. A rough correlation between nitric oxide reduc- tase activity, and the abundance of the M , = 34,000 peptide led to the suggestion that this peptide represents the enzyme. It was recognized, however, that the specific activity of the preparation was on the order of 10 times lower than expected from the in vivo activity. The near absence of iron (0.05-0.1 iron/peptide of M , = 34,000) and the absence of other redox metals was unexpected. Nitric oxide reductase was purified recently from denitrifying Pseudomonas stutzeri strain ZoBell (form. sp. Pa. perfectomarina) by means of detergent (Triton X-100) solubilization to yield a preparation with high specific activity at pH 4.8-5, a substantial content of heme and non- heme iron, and peptides with M , = 17,000 and 38,000 (2). The former peptide was shown to be a cytochrome c and the latter, a cytochrome b.
In considering the remarkable differences between these two preparations, we set out to purify nitric oxide reductase from Pa. denitrificans following its solubilization by a deter- gent. Of the detergents tested, octyl glucoside seemed to offer the greatest advantage and a method was devised to purify the enzyme using this detergent. We now report on the method of purification and some characteristics of the en- zyme. The M , 34,000 peptide of Hoglen and Hollocher (1) is clearly not related to nitric oxide reductase.
As our report was nearing completion, a recent paper by Carr and Ferguson (3) came to our attention. That paper described the partial purification of nitric oxide reductase from Pa. denitrificans following its solubilization by dodecyl maltoside. Their preparation contained a cytochrome c (Mr 18,000 peptide) and a cytochrome b ( M , 37,000 peptide) and several other peptides including abundant ones with M , = 29,000 and 45,000. This variety of peptides precluded an unambiguous assignment between peptides and activity. The present work assigns activity to the cytochrome bc complex.
10899

10900 Nitric Oxide Reductase
PURIFICATION OF NITRIC OXIDE REDUCTASE EXPERIMENTAL PROCEDURES AND RESULTS’
DISCUSSION
The method described for the purification of nitric oxide reductase from Pa. denitrificans is simple, relatively rapid and yields very nearly pure enzyme with high specific activities. Preliminary experiments suggest that the procedure, perhaps with minor variations, may well prove useful for the purifi- cation of the enzyme from other denitrifying bacteria.
Based on data from Pa. stutzeri (2) and Pa. denitrificans (3, herein), it would appear that the enzyme is composed minimally of just two peptides, the one a cytochrome b and the other a cytochrome c. The preparation of Carr and Fer- guson (3) from Pa. denitrificans contained two additional major peptides (as well as minor ones). These two peptides appeared not to be heme proteins and, on the basis of our work and that of Heiss et al. (a), not to be essential for activity. The mole ratio of the M, 17,500 peptide to the M , 38,000 peptide in purified nitric oxide reductase may differ between P. stutzeri (ratio of 3.4 (2)) and Pa. denitrificans (ratio of 1.7 (this work)), possibly as the result of the different detergent and preparative methods used. Which of these two ratios may more closely correspond to the ratio i n vivo is unknown, as is the functional significance of the ratio.
The presence in the enzyme of non-heme iron is indicated for both organisms, but we find the amount to be somewhat smaller in the case of the enzyme from Pa. denitrificans than that reported by Heiss et al. (2) for the enzyme from Pa. stutzeri. The exact roles of the heme and non-heme iron are unclear. If there is a separate non-heme iron peptide (a third peptide), it might migrate by sodium dodecyl sulfate-poly- acrylamide gel electrophoresis with the M , 38,000 band which was always rather broad and diffuse. This would require however that the cytochrome b have more than 1 mol of heme/38,000 g.
Because different assay systems have been used at different pH values to assay purified nitric oxide reductase from Pa. stutzeri and Pa. denitrificans, it is difficult to make direct comparisons regarding specific activity among the prepara- tions reported to date. Nevertheless, they all present specific activities of 10-40 pmol NO x min-’ x mg”. Our data suggest, in addition, that the enzyme from Pa. denitrificans may be nearly diffusion controlled i n uitro. Given the amount of nitric oxide reductase in cells and the i n vivo specific activities recently estimated for the enzyme (17, 23), the enzyme also may be nearly diffusion controlled i n uiuo. This property was in fact predicted (23) as being essential for an enzyme obliged to process a gaseous intermediate present at low steady-state concentrations. Our spectral data suggest that the M , 17,500 and 38,000 peptides of nitric oxide reductase, taken together, probably contain relatively little tryptophan. This is not
Portions of this paper (including “Purification of Nitric Oxide Reductase,” “Experimental Procedures,” “Results,” Tables I and 11, and Figs. 1-7) are presented in miniprint at the end of this paper. The abbreviations used are: CHAPSO, 3-[(3-cholamidopro- pyl)dimethylammonio]-2-hydroxy-l-propanesulfonate; GC, gas chro- matography; NBS, N-bromosuccinimide; NO., and NO,, nitric oxide in the aqueous and gaseous phases, respectively (the same notation applies also to 0, and N20); OG, octyl glucoside (n-OCtyl-P-D-pyran- oside); PMS, 5-methylphenazonium cation (the methyl sulfate salt is commonly called “phenazine methosulfate”); SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; TMPD, N,N,N‘,N”tetramethyl-p-phenylenediamine. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
unusual for cytochromes (22). Spectral data also suggest that the ratio of cytochrome c to b in nitric oxide reductase is similar among the three preparations described to date, in spite of the fact that the enzyme was derived from two different bacteria, solubilized through the action of three different nonionic detergents, and subjected to a variety of techniques to separate one protein from another. The cyto- chrome bc complex of nitric oxide reductase would thus seem to be remarkably resistant to disaggregation in buffered so- lutions of nonionic detergents.
It is clear now that the procedures described by Hoglen and Hollocher (1) are without merit for the true purification of nitric oxide reductase. Two circumstances were chiefly re- sponsible for the error. First, an incomplete separation of the membrane fraction from soluble proteins assured an initially low specific activity and an inappropriate benchmark for assessing later increases in specific activity. Second, a false correlation developed between the abundance of a 34,000 peptide and the activity of nitric oxide reductase which, in fact, remained a minor constituent throughout. The enzyme and the 34,000 peptide both resisted extraction from the membranes, the latter somewhat more than the former. The cytochrome bc complex of nitric oxide reductase is also suffi- ciently well characterized now to be sure that it is distinctly different from the respiratory cytochrome co (2) and bcl (3) complexes of bacteria.
The NADH-NO oxidoreductase activity detected with our nitric oxide reductase preparation probably has little physio- logical relevance, because antimycin A failed to inhibit NADH-NO oxidoreductase activity of the enzyme but is known to be a powerful inhibitor of this activity i n vivo through its action on the respiratory cytochrome bcl complex (3, 18, 23). Failure of antimycin A to inhibit also suggests that nitric oxide reductase is not contaminated with NADH de- hydrogenase and the cytochrome bc, complex of the electron transport system. The coupling of NADH to NO is interesting because it raises the possibility of one-electron reactions with NADH, and such reactions are rare. NAD radical, which can be formed by pulse radiolysis, is known to be a powerful reducing agent for heme proteins and compound I of peroxi- dases (26), but formation of NAD radical from NADH by chemical means is difficult and without precedence in enzy- mology, as far as we know.
We have no evidence at present that the nitric oxide reduc- tase preparation contains the NBR-reactive UV-chromo- phoric lipid which was found in vesicle preparations exhibiting nitric oxide reductase activity and which seemed to reactivate the enzyme after its inactivation by NBR (I). That lipid may not be essential, therefore, for enzyme activity, and the recon- stitution results may have depended on structural factors or other effects not directly related to intrinsic properties of nitric oxide reductase.
That nitric oxide reductase consists of cytochromes is some- what surprising in view of the great stability of ferrous- nitrosyl compounds. Such compounds do not readily disso- ciate to yield ferric complexes plus the reduced product, NzO. A classical example in biochemistry is nitrosyl hemoglobin (24). There may be ways to form N20 reductively from NO that avoid this problem. One approach might be to reduce ferrous-nitrosyl as a whole by one electron to yield nitroxyl (NO-) which would then rapidly dimerize, protonate, and dehydrate (27) to form NzO. During this cycle, the formal redox state of the ferrous center which coordinates NO need not change, but iron centers performing an electron transport function would undergo redox cycling. A reductive pathway involving nitroxyl as an intermediate is attractive in its sim-

Nitric Oxide Reductase 1090 1
plicity, but nitroxyl is a highly reactive species which can react with thiols (28), NO (29), and heme proteins (28, 30, 31), the latter via both inner and outer sphere mechanisms. Attempts to trap extracellular nitroxyl during denitrification using methemoglobin were unsuccessful (23). The release of nitroxyl as an intermediate might be avoided if nitric oxide reductase could bind two molecules of NO proximally, perhaps each at a heme center. In this way the events leading to N 2 0 may be sequestered within the active site. The binding of two substrate molecules might cause the substrate saturation curve to be sigmoidal. We have been unsuccessful so far in seeing sigmoidicity, at least in part because of the inhibitory effects that are significant at NO,, in excess of 10 pM. We plan to test this idea with use of NO/CO mixtures in the hope of observing nonlinear inhibition by CO.
It is interesting in regard to mechanism that the hexaheme c ammonia-producing nitrite reductase of bacteria can reduce NO to N20 in uitro with ascorbate/phenazine methosulfate as the reducing system (34). NO is not normally released in the reduction of nitrite to ammonia and it is problematical whether enzyme-bound NO is an intermediate. Nevertheless, the enzyme can serve as a N20-producing nitric oxide reduc- tase which contains only heme centers.
The work reported herein serves to reinforce recent results which show that nitric oxide reductase is a major and critically important enzyme of the denitrification pathway (2,3,17-19, 23, 32).
Acknowledgments-We acknowledge the technical assistance and cooperation of Chun-sheng Zhang and Carlton Soo-Hoo of this De- partment.
1.
2.
3. 4. 5.
6. 7.
REFERENCES
Hoglen, J., and Hollocher, T . C. (1989) J. Biol. Chem. 264,7556-
Heiss. B.. Frunzke. K., and Zumft, W . G. (1989) J. Bacteriol. 7563
. . 17i, 3288-3297
Carr. G. J.. and Fereuson. S. J. (1990) Biochem. J . 269,423-429 Laemmli, U. K. (1970) Nature 227, 680-685 Goodhew, C. F., Brown, K. R., and Pettigrew, G. W. (1986)
Weber, K., and Osborn, M. (1969) J. Biol. Chem. 244,4406-4412 Fuhrhop, J.-H., and Smith, K. M. (1985) in Porphyrins and
Metalloporphyrins (Smith, K. M., ed) pp. 757-869, Elsevier/ North-Holland Biomedical Press, Amsterdam
Biochim. Biophys. Acta 852,288-294
8. Laurent, T. C., and Killander, J. (1964) J. Chromatogr. 14, 317-
9. Strittmatter, P., and Velick, S. F. (1956) J. Bid. Chem. 221,
10. Strittmatter, P. (1963) The Enzymes (Boyer, P. D., ed) 2nd Ed.,
11. Massey, V. (1959) Biochim. Biophys. Acta 34, 255-256 12. Van Gelder, B. F., and Slater, E. C. (1962) Biochim. Biophys.
13. FIatmark (1964) Acta Chem. Scand. 18, 1517-1527 14. Peterson, G. L. (1985) Anal. Biochem. 100,201-220 15. Ji, X.-B., and Hollocher, T. C. (1988) Appl. Enuiron. Microbiol.
16. Zumft, W. G., and Frunzke, K. (1982) Biochim. Biophys. Acta
17. Goretski, J., and Hollocher, T. C. (1990) J. Biol. Chem. 265,
18. Carr, G. J., Page, M. D., and Ferguson, S. J. (1989) Eur. J .
19. Zafiriou, 0. C., Hanley, Q. S., and Snyder, G. (1989) J. Biol.
20. Gorbunoff, M. J. (1984) Anal. Biochem. 136, 425-445 21. Snyder, S. W., and Hollocher, T. C. (1987) J. Biol. Chem. 262,
22. Lemberg, R., and Barrett, J. (1973) Cytochromes, Academic Press,
23. Goretski, J., and Hollocher, T. C. (1988) J. Biol. Chem. 263,
24. Cassoly, R., and Gibson, Q. H. (1975) J. Mol. Biol. 91, 301-313 25. Yonetani, T. (1968) Structure and Function ofcytochromes (Oku-
nuki, K., Kamen, M. D., and Sekuzu, I., eds) pp. 289-292, University Park Press, Baltimore
26. Kobayoshi, K., Hayashi, K., and Swallow, A. J. (1990) Biochem- istry 29, 2080-2084, and references therein
27. Bazylinski, D. A,, and Hollocher, T. C. (1985) Inorg. Chem. 24, 4285-4288
28. Doyle, M. P., Mahapatro, S. N., Broene, R. D., and Guy, J . K. (1988) J. Am. Chem. Soc. 110, 593-599
29. Gratzel, M., Taniguchi, S., and Henglein, A. (1970) Ber. Bunsen- Ges. Phys. Chem. 74, 1003-1010
30. Bazylinski, D. A., and Hollocher, T. C. (1985) J . Am. Chem. Soc.
31. Bazylinski, D. A,, Goretski, J., and Hollocher, T. C. (1985) J. A m . Chem. Soc. 107, 7986-7989
32. Goretski, J., Zafiriou, 0. C., and Hollocher, T. C. (1990) J. Biol. Chem. 265, 11535-11538
33. Canfield, R., and Liu, A. K. (1965) J. Biol. Chem. 240, 1997- 2002
34. Costa, C., Macedo, A., Moura, I., Moura, J . J . G., LeGall, J., Berlier, Y., Liu, M.-Y., and Payne, W . J. (1990) FEBS Lett.
330
253-264
Vol. 8, pp. 113-145, Academic Press, Orlando, FL
Acta 58,593-595
54,1791-1794
681,459-468
889-895
Biochem. 179,683-692
Chem. 264,5694-5699
6515-6525
New York
\ 2316-2323
107,7982-7986
276,67-70
Continued on next page.
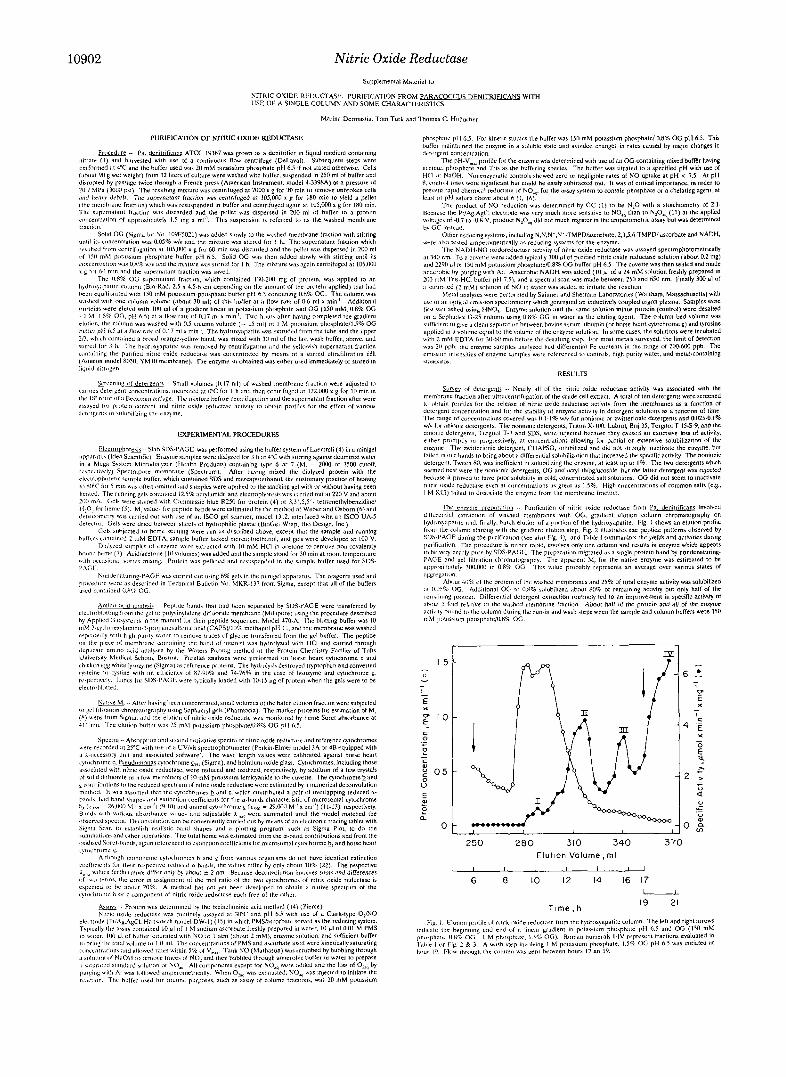
10902 Nitric Oxide Reductase Supplemental Material to
NITRIC OXIDE REDUCTASE. PURIFICATION FROM WITH USE OF A SINGLE COLUMN AND SOME CHARACXERISTICS
Mama Drrmama. Tom Turk and Thomas C. Hcllluchrr
1 5 - 0
- L 5 E" IO c 0 .- c
e +. c W
0 5 0 V c W - * e a
0
2 5 0 2 8 0 310 340 370 Elutlon Volume, m l
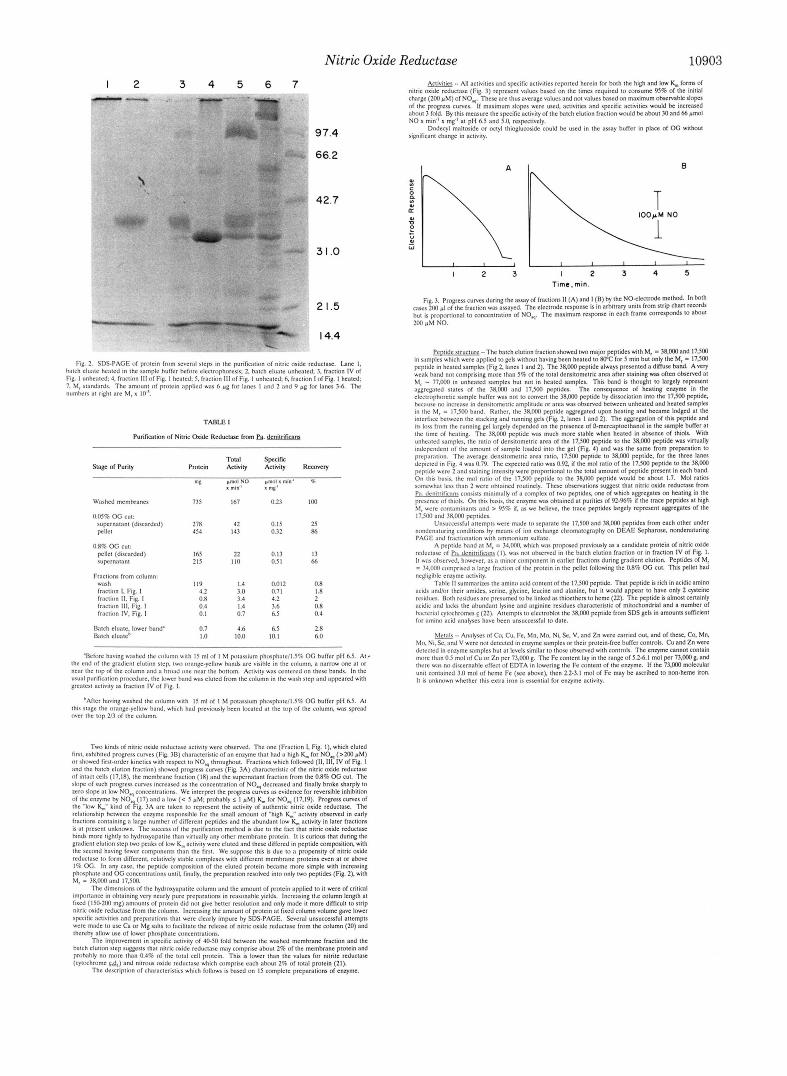
Nitric Oxide Reductase 10903 1 2 3 4 5 6
.. . .
w :;A
7
97.4
66.2
42.7
31.0
2 1.5
w- 14.4 .
TABLE I
1 2 3
\ IOOpM NO
I I 1 I I
1 2 3 4 5 Time, min.
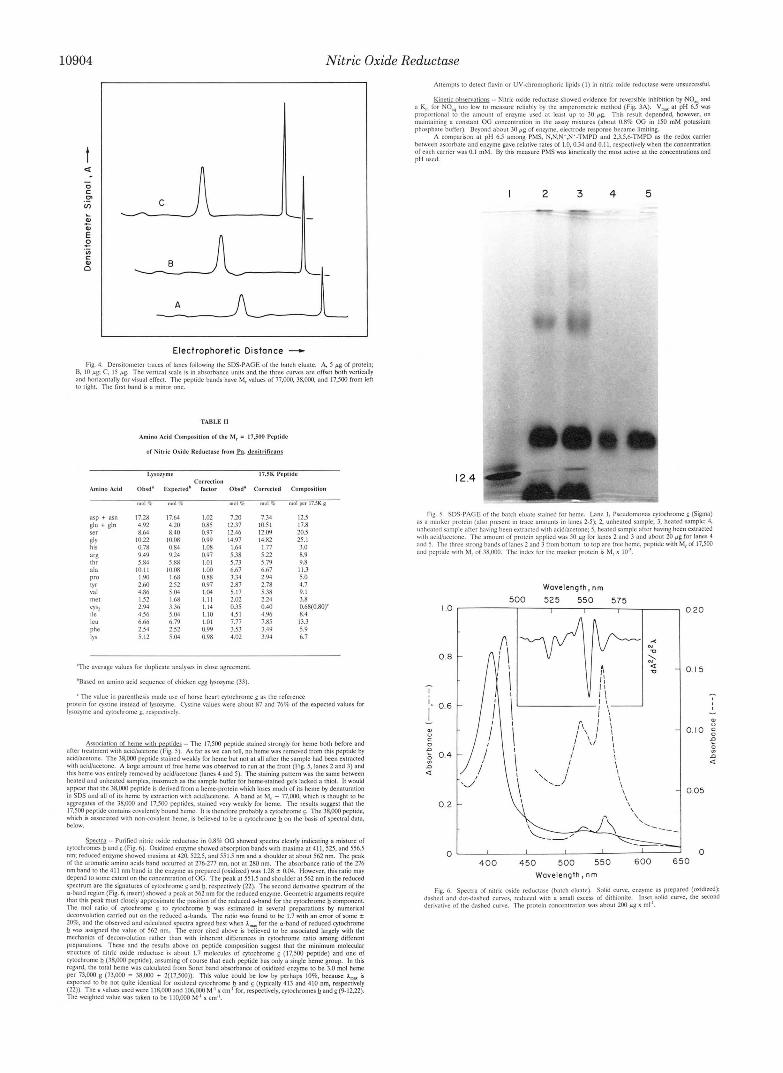
Nitric Oxide Reductase
A A L
Electrophoretic Distance - Fig. 4. Dcnritometer tritces of hnes following the SDS-PAGE of the hatch eluate. A 5 r g of prolein:
and hortrontally for visual effect. The peptide hands have M, values nf 77,000. 38.000, and 17.500 from left B. 10 pg: C. I5 fig. The vertical scale is in rhrorhanee units andthe three curves are offset both venlcally
t o right. The Brst hand is B minor one.
TABLE II
Amino Acid Camporition d t h e M, = 17.SW Peptide
or h'itrir Oxide Reductase rmm m.denitri l irnnr
Lyrozrme 17.SK Peptide Corn t i on
Amino Acid Obrd' Expectedb factor Obsd' Corrmtd Composition
m01 ?r "?"I % mol ?a mol % mol F? 17.SK 8
17.64 4.20
m u 8.40
ILX4 '1.24
II1.lIX 5.XX
2 52 1.0s
5N 1.6R S.36 5.W 6.79
5.11 2.52
0x5 12.37 10.51 17.x 1.02 7.211 7.34 12.5
0.97 12.46 m 9 20.5 0.99 14.97 I w 25.1
0.97 5 . 3 ~ 5.22 x.9 1.01 s.73 5 . 7 ~ 9.x
nnx 3.34 2.94 5.0 0.97 2.87 2 . 7 ~ 4.7
1.11 2.02 2.24 3.x 1.14 0.35 0.40 ~.~R(o.RII)~ 1.10 4.51 4.96 n.4
0.99 3.53 3.49 5.9 1.01 7.77 7 x 5 13.3
0 . 9 ~ 4.02 3.04 6.7
I.IIR 1.64 1.77 3.0
1.00 6.67 6.67 11.3
1.N 5.17 5.3X 9.1
Asrociatm of heme with nemides - -The 17.500 pepudc stained strongly for heme hath hefnre and after trmlment wlth acidlacetone (Ftg. 5 ) . As for as WE can tell, no heme was removed from this peptide hy acidlacetone. The 3R,MHI peptide stmned wcakly for heme hut not at all nfter the sample had heen extracted with scidiacelonr. A krge amotm of free hemc war ohserved to run at the front (Fig. 5. lanes 2 and 3) and I B I S hcme was entirely rsmaved hy acidlacetone (Inner 4 and 5) . T h e staining pattern was the Same hetween
a p p r that the 3 X . W peptide I? derived from P heme-protein which loses much of its heme hy denaturation heated and unhealed ramplrs. inasmuch as !he sample huffer for hems-stained gels lacked a thiol. I1 would
agregmcs of the 3 X . W and 17.500 peptides. rt;$ined very weakly for heme. The results suggest that the ~n SDS and 811 of i l l heme hy rx~raclmn w!!h ac~Ilaectane. A hand ill M, - 77.000. which is thought to he
17.51Hl ppt idc c ~ n l a m l eovalenlly hound hrmc. I! is lhrrefore prohahlya cytochrome e. The 3 X . W peptide. which is sseociated with "on-covalm! heme. IS believed t o he a cytochrome h on the hasir of spectral dam. helow.
cytochmmer h and e_ (Pig. 6) . Oxldizrd enzyme showed absorption hands wlth maxima at 41 I. 525. and 556.5 Srrectrn .. Purified nitric oxidc reduclare in 0.8% OG showed spectra clearly indicating a mixture of
nm: rcduced enzyme showed m w m a a! 420.522.5. and 551.5 nm and a shoulder a! ahnut562 nm. T h e peak nf the aromatic ammo acids hand occurred at 27c277 mn. nut at 2x0 nm. The ahrorhance ratio of the 276 nm hand In the 41 I nm hand in tile enzyme as prepared (omhzed) was 1.211 ? 0.04. However. this ratio may depend to Some extent nn the concmtrat~on of OC. The peak at 551.5 and rhouldcr at 562 nm in the reduced spectrum are the signatures of oytachrome E and h, respectively (22). The second derivawe spectrum of the 0-hand region (Fig. 6. mrert) showed it peak at 562 nm for Ihc reduced enzyme. Geometric arguments rcqulre that this peak must closely approximate (he poritmn of the reduced *hand for the cytochrome h companent. The mol ratio o f cytochrome E to cytochrome b was estimated in several preparations hy numeric81 drconvolutlnn carried out on the reduced o-hands. The ratio was found to he 1.7 with an error of Some
h will assigned the value of 562 nm. The error cited ahave is hrlievrd 10 he assmiated largely with the 20%. and the ohsewed and calculated spectra agrccd b e s t when A, for the e-hand of reduced eytochromc
mechsnicr of deconwlution rather than with inherent differences in cytochrome ratio among different preparauonr. These and the results ahovr on peptide composition sugest that the minimum molecular slructure of nitric oxide reductase is about 1.7 molrc~ l r~ of cytochrome e (17.500 pepttde) and one of cytochrome !? (3R.000 pcptide). ilssumlng o f course that each peptide has only a single heme group. In this regard. Ihe lotill heme was cvlculnted f r o m Soret hand ilhrrrhance of oxldlzed enzyme to be 3.0 mol heme per 73.000 g (73.000 = 38.000 + 2(17.500)). This vaiur could be low hy perhaps 10%. because 1, is expected IO he no1 qulle idcnlical for Omhred cytochrome h a n d (typlcaiiy 413 and 410 nm, rerpectively (22)). Thetv~luesusedwercl lX.000andIOh.MH1M'xcm'far .respect ively.Eylochroml2.22) . The wclghled value was taken to be I10,000 M I x cm".
I 2 3 4 5
12.4
Wavelength.nm
500 525 550 575 1.0 , I I I I 0 20
0.1 5
- 1
0) " 0.10 g
D s B 4
0.05
0
Nitric Oxide Reductase 10905 It was observed that NADH could also ~ e r v e at pH 6.5 as a kinetically slow electron donor to purified
n8Irlc cnxlde reducrase. The reaction was f o l l w e d spectrnphotametrically and amperometrically in parallcl and showed a stoichiometry of 2 h r t w e n NO uptake and NADH oxidation. Although antimycin A is a pawerful inhibitor of NO uptake activtty and in the membrane fraction of Pa. denitrificans (3,1&23), i t had no Inhibitory effect up to 50 uM an the NADH-NO or PMSiascorbale-NO oxidoreductase activities of the purlfled enzyme. When concentrations of NADH and NO were initially 83 and 200 uM, rerpectively, the mitis1 specific rate was a b u t 0.05 pmol NADH x min ' x mg' with the hatch eluate. and the rate decreased monotonically as the reaction progressed. The 1, for the Sore1 band was 420 nm in presence of NADH, shifted to 414-416 nm on addition of NO, and became 420 or 410 nm in the end. depending on whether
hecnme partially oxidized (or formed NO complexes) on addition of NO, and returned finally Lo the reduced NADH or NO, respectR.ely. war in CXCCS. The pattern fuggcrts that the cytochromes arc initially reduced,
or oxldired slate depending on which substrate was in CXECSI.
activity at maximum slopc of 30 "mol NO x mi".' x mg.' at pH 6.5 and a minimal M, of 73.MHl. Our An enzymatic turnover number (kc#,) of 36 I ' IS calculated for nitrlc Oxide reductase based on a specific
amprrometric data indicate that of the purtfied enzyme for NO is probably i 1 uM and thh value is cnmmrnsurate with the in vivo d u e for NO uptake hy P. slutze? Strain ZoBell of 0.4 uM (19) and by several other denitrifyingbactcria oi i l y M (3). It is m u c h t h a n the value of fQ uM prapased hy
ayltem. Because the uptake of NO can easily become &ifusion lhmited in G C assay systems at low panial Helsr et al. (2) for the purified nitne oxide reductase from LCgtzcj ZoBell on the hasis of a G C assay
pressures of NO (17) i f is plrsihle :hat the 60 uM v d ~ e is much greater than the tmc value. l ime accepts K, 5 I pM. th& the effective second-order rstr constant for purified nitric Oxide reductase from P, drn#tr#ficanr hecomes k,,/K, L 4 x IO' M I x I ? ; ~t hrwxnrs 9 x 10' M' x d if Km = 0.4 pM. There values are wnhm one to two orders of magnitude o f the dtffunon-conlrolled limlf and cxcred the rate con~tant for ampler formation hetween hrmaglohln and NO (24).
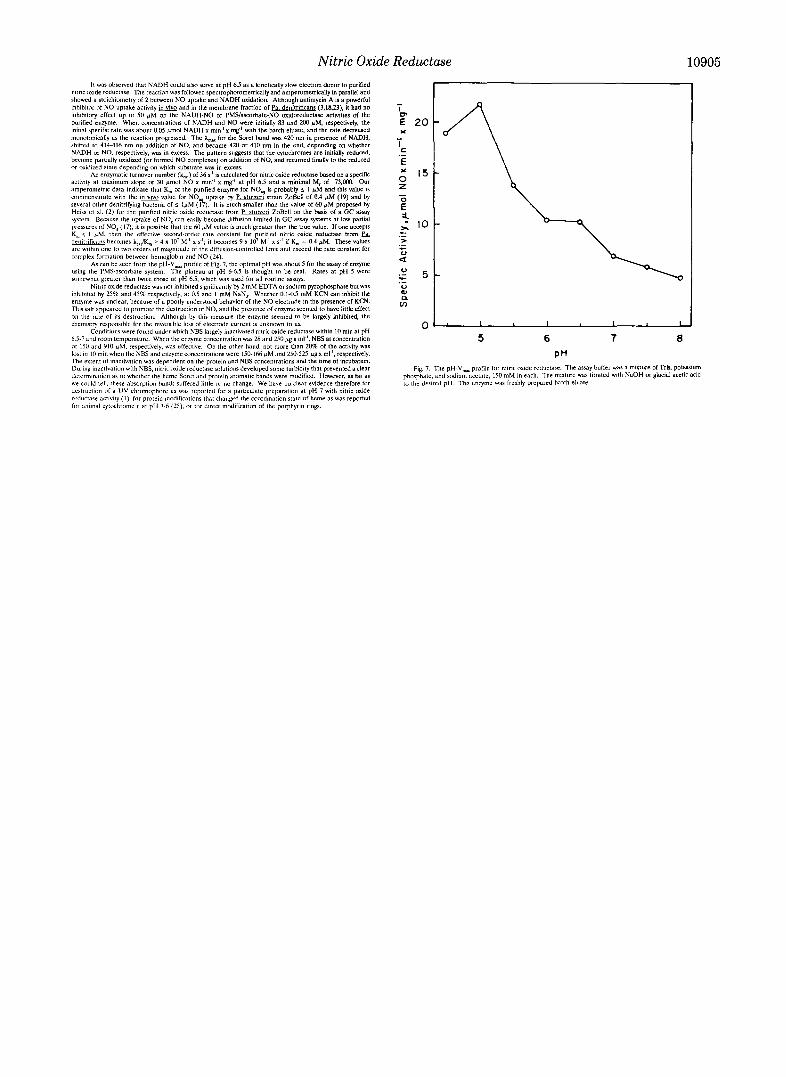
As can he seen from the pH-Vm,, profile of Fig. 7, the optimal pH was ahaut5 for the array of enzyme using Ihr PMSbscorhate SYSIE~. The plateilu at pH 6-6.5 is thought !n he real. Rater at pH 5 were somewhat greater than twice those et pH 6.5. which was used for all routine assays.
Nitrlc omde reductase was not inhibited sigmf~cmtly hy 2 mM EDTA or sodium pyrophosphate hut was inhiblted by 25% and 45% respectwely, at 0.5 and I m M NaN,. Whether 0.1-0.5 mM KCN can inhibit the enzyme was unclear, hecause of a poorly understood behavior of the NO-electrode in the presence of KCN. Thi? salt appeared to promote the destructton of NO, and the presence of enzyme seemed to have little effect on the rate of 11s destruction. Although hy thn measure the enzyme seemrd to be largely Inhibited, the chrmirtry responshle for the reversible I u s of electrode currenf is unknown to US.
Conditions were found under whlch NBS largely innclivaled nllric oxlde reductase withm 10 mm at pH 6.5-7 and mom temperature. When the enzyme con~enfratmn war 28 and 250 pg I ml'. NBS at concentration of 150 and 910 pM, rrspectwely. was sffemvr. On the other hand. not mare than 20% of the activity was Ihs1 m 10 mm when the NBS and enzyme c ~ n c e n t r a l m ~ were 150-1% p M m d 250-525 ug x ml I. respeetwely. The rxtrnt of inactivatmn was dependent on the protein and NBS concentrations and the time of incubation. Durmg mactiwfion with NBS, nimc oxide reductase soluticm~ developed some turhidlly that prevented a clear drrrrminalicm ar to whether the heme Sorrt and p w e m aromatic hands wcre mdificd. However, 8s far as we could fell. there absorption hands suffered l81tlr or no change. We have no clear evidence therefore for dertruut8m of a U V chromophore as was reported for a particulate prrpvrat~on at pH 7 w t h nitric onde reductase vcl8vily ( I ) , fnr protein modifications t h a t changed the coordmarian slate of heme ar was reponed lor animal cytochrome y at pH 3-6 ( 2 5 ) . or fur dlrect modificatiun of the porphyrm rings.
;;" 5 .- u al a m
0 I I I I
5 6 7 8 pH
phusphutr, and sadvm acetate, 150 m M in each. The mixture war titrated wlfh NaOH or glacial acetic acid Fig. 7. The pH-Vm, profile for nitric oxide reductase. The asray buffer was a mixture aiTr i r . patPasium
1 0 the desired pH. The rnzymr WBS freshly prepared hatch eluate.





























