Embed Size (px)
Citation preview

NNMCB-Workshop on systems biology
K. Sriram
Indraprastha Institute of Information Technology
December 21, 2014
Chemical and biochemical reaction networks
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 1 / 33

Learning objectives
Three learning objectives from this lecture:
(i) To build dynamic ODE models from static chemical/biologicalnetworks using the laws of mass action (chemical/biochemical)kinetics.
(ii) To perform model reduction based on the time-scales of thereactions; quasi-steady-state approximation
To build and simulate ODE model of apoptosis network framed byEissing et al., and to identify the role of feedback loop(s).
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 2 / 33

References
John J Tyson, Katherine C Chen and Bela Novak, Sniffers, buzzers,toggles and blinkers: dynamics of regulatory and signaling pathwaysin the cell, Current Opinion in Cell Biology 2003, 15:221231.
Mathematical Modeling in Systems Biology - An Introduction, BrianP. Ingalls, MIT Press (2013).
Biochemical Oscillations and Cellular Rhythms: The Molecular Basesof Periodic and Chaotic Behaviour, Albert Goldbeter (1996).
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 3 / 33

Why model biological/chemical reaction networks?
With the advent of new biotechnological techniques it is possible todissect the molecular mechanisms that underlie the adaptive behaviorof living cells. Examples are cell-division cycle, lytic-lysogenic switchof virus etc.
The information of the molecular mechanisms are laid down in theform of graphical form. For example, http://www.biocarta.com/;http://discover.nci.nih.gov/mim/kohnk/kohnk.jsp
In these graphical forms, genes, proteins and metabolites are hookedtogether by chemical reactions and intermolecular interactions.
The temptation is irresistible to ask whether physiological regulatorysystems can be understood in mathematical terms.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 4 / 33
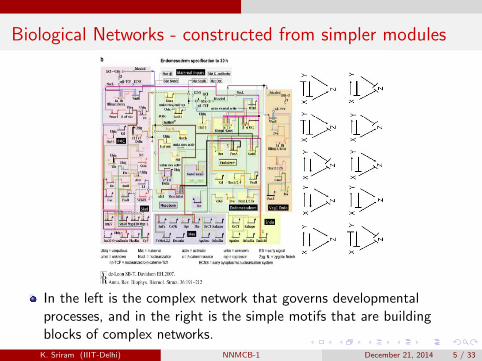
Biological Networks - constructed from simpler modules
In the left is the complex network that governs developmentalprocesses, and in the right is the simple motifs that are buildingblocks of complex networks.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 5 / 33
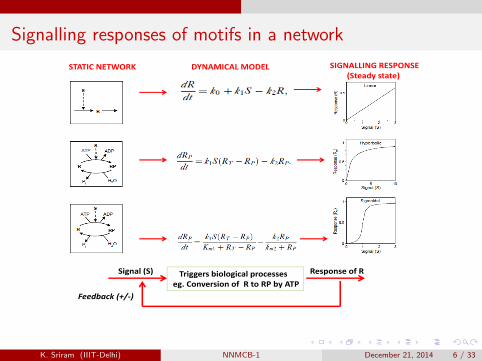
Signalling responses of motifs in a network
STATIC NETWORK DYNAMICAL MODEL SIGNALLING RESPONSE(Steady state)
Triggers biological processesSignal (S) Response of RTriggers biological processeseg. Conversion of R to RP by ATP
Feedback (+/‐)
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 6 / 33

Usefulness of mathematical models in analyzing biologicalnetworks
Account for experimental observations and to determine the validityof experimental conclusions.
Clarification of hypothesis.
Difficult to rely only on intuition. Mathematical equations provides astrong foundation for validating concepts and analyze complex datathat involves multiple coupled variables.
Models identify critical parameters for which certain phenomena canoccur. For eg., switch-like behavior, oscillations etc.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 7 / 33

Usefulness and cons of mathematical models
Different regulatory mechanisms can be explored through models fromwhich only plausible mechanisms can be identified, which otherwiseare expensive and time consuming to carry out through experiments.
Models can help to identify different dynamically important regimes,which may be hard or inaccessible to experimentalists.
Problems: When the numbers become small, differential equationdescriptions breaks down.
Estimating kinetic parameters becomes difficult if ODE models arehigh dimensional.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 8 / 33

Types of mathematical models frequently used to modelbiological/chemical networks
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 9 / 33

Chemical/Biological reaction networks – What is it?
Species‐1
Species 3
Species‐4
Species‐3
Complex
Species‐2 Species‐5
Closed reaction network Open reaction network
For biochemical and geneticnetworks, the components inthese diagrams are molecularspecies like ions, small moleculesetc.
An interaction diagram depictsthe species in a system andindicates how they interact withone another.
The interactions or arrow in thediagram can represent a rangeof processes, such as chemicalbinding or unbinding, reactioncatalysis, or regulation ofactivity.
Rate of the process results inthe production, interconversion,transport, or consumption of thespecies within the network.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 10 / 33

Writing rate laws, stoichiometry and evolution ofconcentration in chemical reaction networks
Consider the following reactions (i) A + B → C + D, (ii) 3 A + B →A + C and (iii) A + 2 B → 3A + c.
Assume k to be the rate constant of the all the reactions.
What is the rate law, ’v’ for the reaction?
What are the stoichiometric coefficients for the reaction?
What is the integrated rate law (or) evolution of the concentration foreach of the reactions?
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 11 / 33

Solution - (i)
The rate of irreversible reaction is given as v = k [A][B].
The evolution equations or the concentration variations in time forthe reactant [A], [B], and the products [C] and [D] are given as d [A]
dt ,d [B]dt , d [C ]
dt , d [D]dt .
Stoichiometric coefficients for A, B, C, D are -1, -1, 1, and 1respectively.
The evolution /integrated rate laws of the species depends on the rate
– d [A]dt = –d [B]
dt = vd [C ]dt = d [D]
dt = v
Note the signs; -ve for reactant & +ve for product.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 12 / 33

Solution - (ii)
The rate of the reaction 3 A + B → A + C is given as v = k[A]3[B].
When we write the evolution of the concentration of A, we must takeinto consideration the fact that each time this reaction occurs, onlytwo molecules of A are transformed (one is conserved). So, thevariation of A is given by−1
2d [A]dt = v = k [A]3[B].
The coefficient 3 is the balance for the species A in reaction and thesign - stands because, globally, A is transformed (not produced).
Three things are vital to write a correct rate laws for a given reaction:(i) Sign, (ii) Stoichiometric coefficient, & (iii) Rate, v.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 13 / 33

Solution - (iii)
For the reaction A + 2 B → 3A + C, the stoichiometric coefficientsareηA = 3 – 1 = 2, ηB = 0 – 2 = -2, ηC = 1 – 0 = 1.
So – d [A]dt = ηA k[A][B]2 = 2k [A][B]2
–d [B]dt = ηBk [A][B]2 =2k [A][B]2
d [C ]dt = ηCk[A][B]2 = k[A][B]2
In general, for the reaction n X + ......... p X, the vectorial form of
the equation isd [X ]
dt= ηv ,with η = p − n .
η is called the stoichiometric coefficient. The coefficient is positive if,globally, the species is produced (p > n), and negative if the speciesis transformed (n > p).
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 14 / 33

Open and closed chemical reaction networks
Closed reaction network Open reaction network
Extra reactions added
(see the difference)
In closed reaction network,thereis no exchange of energy ormatter with the outsideenvironment. They are inthermal equilibrium.
In open reaction network,thereis an exchange of energy ormatter with the outsideenvironment. They are indynamic equilibrium.
These reactions in the networkproceed on a fixed volume. Thereaction volume is well stirred.
There are sufficiently greatnumber of species or molecules,so that the concentration variescontinuously.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 15 / 33
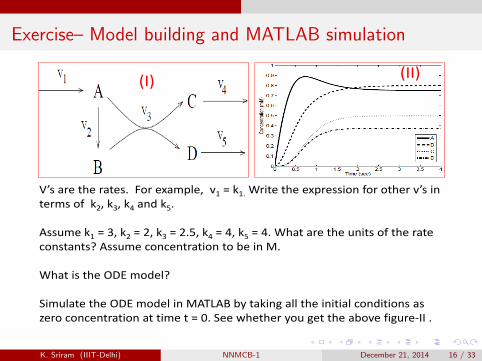
Exercise– Model building and MATLAB simulation
(I) (II)
V’s are the rates. For example, v1 = k1. Write the expression for other v’s in terms of k2, k3, k4 and k5.
Assume k1 = 3, k2 = 2, k3 = 2.5, k4 = 4, k5 = 4. What are the units of the rate constants? Assume concentration to be in M.
What is the ODE model?
Simulate the ODE model in MATLAB by taking all the initial conditions as zero concentration at time t = 0. See whether you get the above figure‐II .
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 16 / 33
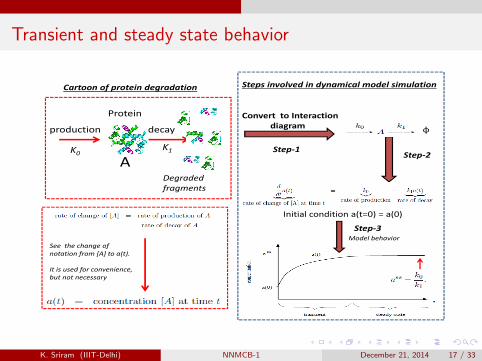
Transient and steady state behavior
Cartoon of protein degradation Steps involved in dynamical model simulation
Protein
decay φ
Convert to Interaction diagramproduction
AK1
Degraded
Step‐1Step‐2K0
Degraded fragments
Initial condition a(t=0) = a(0)
See the change of
Initial condition a(t=0) = a(0)
Step‐3Model behavior
notation from [A] to a(t).
It is used for convenience,but not necessary
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 17 / 33

Importance of wiring diagram and modelling of transientactivation: Mitogen Activated Protein Kinase (MAPK)pathway as example.
MAPK, a signal transduction pathway that plays an important role incell decision process. i.e., either proliferation or differentiation.
In MAPK pathway, transient ERK activation is mediated byphosphorylated receptors, Ras small GTPase, SOS etc.
The signal(S) that activates this pathway is a growth factor whichcan be either EGF (Epidermal growth factor) or NGF (Nerve growthfactor), and the response is seen in the ERK activation.
Transient and sustained response of a ERK throws light on whethercell proliferation or cell differentiation takes place.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 18 / 33
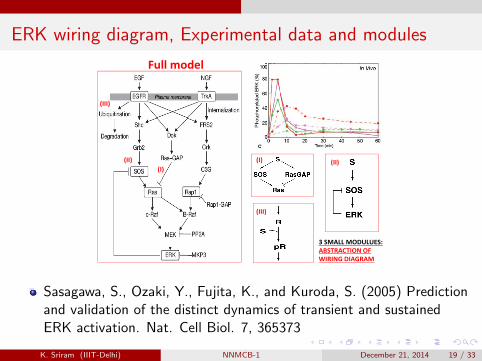
ERK wiring diagram, Experimental data and modules
Full model
(III)
(I) (II) (I)
(II)
(III)
3 SMALL MODULUES:ABSTRACTION OFWIRING DIAGRAM
Sasagawa, S., Ozaki, Y., Fujita, K., and Kuroda, S. (2005) Predictionand validation of the distinct dynamics of transient and sustainedERK activation. Nat. Cell Biol. 7, 365373
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 19 / 33

Continued
Based on the increasing knowledge of molecular mechanisms, severalkinetic models of the ERK signaling network have been developed,and the transient versus sustained nature of the ERK activation hasbeen analyzed.
What are the different molecular networks/wiring diagrams that mayproduce transient activation of ERK?.
What are the corresponding kinetic models?
Which among the 3 is the most robust model?.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 20 / 33
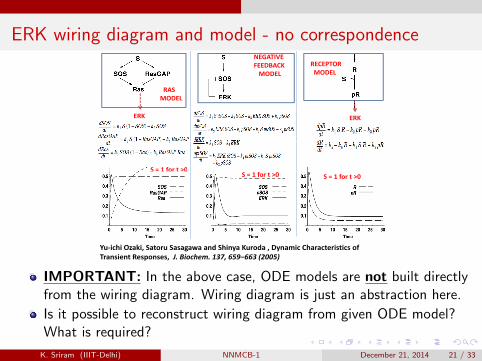
ERK wiring diagram and model - no correspondenceNEGATIVE FEEDBACK MODEL
RECEPTOR MODEL
RAS MODEL
ERK ERK
S = 1 for t >0S = 1 for t >0
S 1 for t 0S = 1 for t >0
Yu‐ichi Ozaki, Satoru Sasagawa and Shinya Kuroda , Dynamic Characteristics of Transient Responses, J. Biochem. 137, 659–663 (2005)
IMPORTANT: In the above case, ODE models are not built directlyfrom the wiring diagram. Wiring diagram is just an abstraction here.
Is it possible to reconstruct wiring diagram from given ODE model?What is required?
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 21 / 33
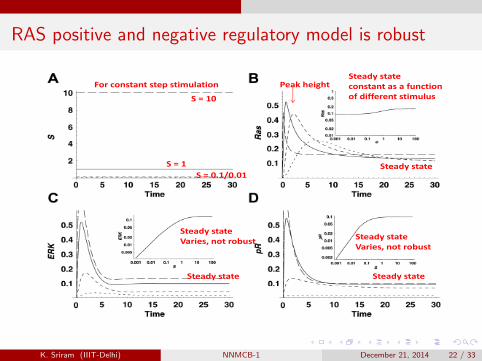
RAS positive and negative regulatory model is robust
Peak heightSteady stateconstant as a function of different stimulus
For constant step stimulation
S = 10
Steady stateS = 1S = 0.1/0.01S 0.1/0.01
Steady stateVaries, not robust Steady state
Varies, not robust
Steady stateSteady state
,
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 22 / 33

Irreversible reaction and conservation relationship
Closed system, irreversible reactionsIf a(0) = A0 and b(0) = B0, then T = A0 + B0
Conservation from mathematical model
a(t) = concentration of [A] at time tb(t) = concentration of [B] at time t
We can also write b(t) = B0 + A0 – a(t)
The conservation of concentration is
a(t) + b(t) = T
Difficult to solve this ODE. Reduce to 1D system.
From the symmetry in the rates of change for A and B, we can write
Difficult to solve this ODE. Reduce to 1D system.
Assume, Total concentration of A and B areconserved .
i.e, Everytime a molecule B is produced, a molecule of A is consumed.
confirming that the total concentrationMathematically, It says that
confirming that the total concentration a(t) + b(t) does not change with time.
Exercise: Find the steady states ass and bss .
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 23 / 33

Biological time-scales
Biological processes take place over a wide range of time-scales.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 24 / 33
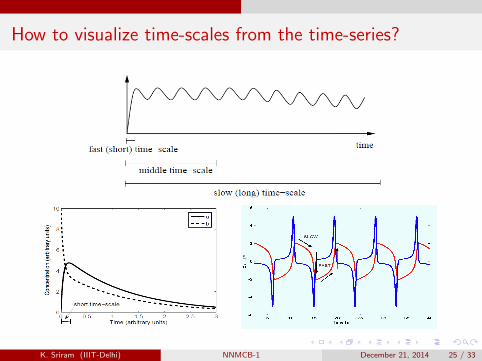
How to visualize time-scales from the time-series?
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 25 / 33

How to perform model reduction of biological system withmultiple time-scales?
To model a system that involves processes acting on differenttime-scales, a primary time-scale must be chosen. Other time-scalesare then treated as follows:
Processes occurring on slower time-scales are approximated asfrozen in time.
Elimination of slow time-scales is easy and it is done by assigning aconstant value or as a fixed parameter.
Processes occurring on faster time-scales are presumed to occurinstantaneously.
There are two ways to treat fast time-scales; (a) rapid equilibriumapproximation and (b) by Quasi-steady state approximation (QSSA).
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 26 / 33

Model reduction by quasi-steady-state approximation(QSSA)
The model-reduction method that focuses on individual species inQSSA.
Consider the reactionk0−→ A
k1←−→k−1
Bk2−→
The full network ODE’s areda
dt= k0 − k1a(t) + k−1b(t) and
db
dt= k1a(t)− k−1b(t)− k2b(t)
Let us assume that all dynamic reactions involving species A occur onthe fast time-scale, so that, compared to the dynamics of B, speciesA comes rapidly to its steady-state concentration.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 27 / 33

Quasi-steady-state approximation - continued
Mathematically, k1 + k−1 >> k2, and k0, the zero order reactionconstant do not contribute to the time-scales of [A].
Now the ODE for [A], which is dadt = k0 + k−1 b(t) - k1 a(t) becomes
an algebraic equation. i.e.,
dadt = 0 ⇒ aqss = k0+k−1b(t)
k1(algebraic equation)
The reduced model will be dbdt = k1 aqss(t) - (k−1 + k2) b(t).
The reduced model isdb̃
dt= k0 − k2b̃(t)
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 28 / 33
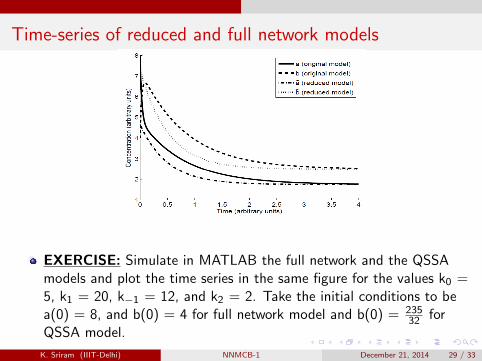
Time-series of reduced and full network models
EXERCISE: Simulate in MATLAB the full network and the QSSAmodels and plot the time series in the same figure for the values k0 =5, k1 = 20, k−1 = 12, and k2 = 2. Take the initial conditions to bea(0) = 8, and b(0) = 4 for full network model and b(0) = 235
32 forQSSA model.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 29 / 33

Life and death of a cell – Apoptosis
The process of programmed cell death cellular suicide – is calledapoptosis. When cells are no longer needed, apoptosis is a tidyprocess.
Apoptosis is invoked by caspase proteins, which are always present inthe cell, but lie dormant until activated.
Two types of caspase proteins (i) Initiator that responds to inputstimuli, like stress triggered signals from mitochondria, and (ii)Executioner caspases that are activated by initiator. One role is todegrade cell’s DNA.
To guarantee that the decision to undergo apoptosis is irreversible, afeedback mechanism is in place: active caspase-3 activates caspase-8.This positive feedback ensures that caspase activity isself-perpetuating. The network is given below.K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 30 / 33
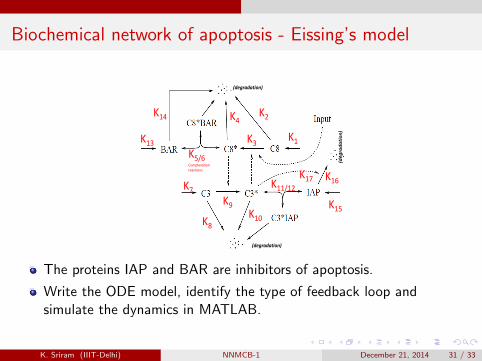
Biochemical network of apoptosis - Eissing’s model
(degradation)
K2K4K14
K1K3K
K13
grad
ation)
K5/6Complexationreactions K17
(deg
KK7
KK11/12
K
K17 K16
K8
K9K10
K158
(degradation)
The proteins IAP and BAR are inhibitors of apoptosis.
Write the ODE model, identify the type of feedback loop andsimulate the dynamics in MATLAB.
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 31 / 33

Feedback loops – Simple representations
Negative feedback loops(N b t t ffi i t diti f ill ti )(Necessary but not sufficient conditions for oscillations)
oop M
edba
ck lo
Mutual An
itive fe
e ntagonisPo
s sm
Necessary and sufficient conditions for multi‐stability(nonlinearity + choice of parameters)
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 32 / 33
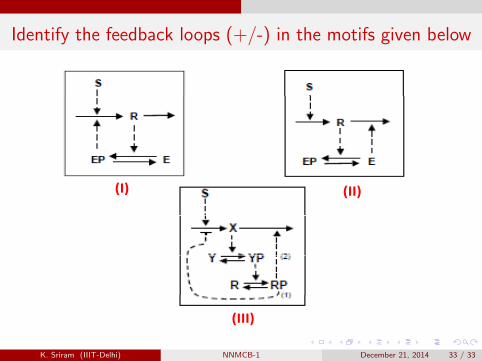
Identify the feedback loops (+/-) in the motifs given below
(I) (II)
(III)
K. Sriram (IIIT-Delhi) NNMCB-1 December 21, 2014 33 / 33


![Abstract arXiv:1901.09237v1 [cs.CV] 26 Jan 2019Anubhav Jain, Richa Singh, Mayank Vatsa IIIT Delhi, India fanubhav15129, rsingh, mayankg@iiitd.ac.in Abstract Digitally retouching images](https://img.pdfslide.net/doc/110x75/5f785e1733d37a1d7d2d60e0/abstract-arxiv190109237v1-cscv-26-jan-2019-anubhav-jain-richa-singh-mayank.jpg)



![arXiv:2006.08696v1 [cs.CV] 15 Jun 2020 · Prashant Pandey?, Aayush Kumar Tyagi , Sameer Ambekar, and Prathosh AP Indian Institute of Technology Delhi getprashant57@gmail.com, aayush16081@iiitd.ac.in,](https://img.pdfslide.net/doc/110x75/5f6b2417cc24456ac85cde46/arxiv200608696v1-cscv-15-jun-2020-prashant-pandey-aayush-kumar-tyagi-sameer.jpg)