Embed Size (px)
Citation preview

Orthopedic Reviews 2009; volume 1:exxxx
[Orthopedic Reviews 2009; 1:e11] [page 39]
Novel information on the non-neuronal cholinergicsystem in orthopedics providesnew possible treatmentstrategies for inflammatory anddegenerative diseasesSture Forsgren,1 Håkan Alfredson,2
Dennis Bjur,1 Solbritt Rantapää-Dahlqvist,3 Örjan Norrgård,4 Tore Dalén,5
Patrik Danielson1
1Dept. of Integrative Medical Biology,Anatomy, Umeå University, Umeå,Sweden; 2Dept. of Surgical and PerioperativeSciences, Sports Medicine, UmeåUniversity, Umeå, Sweden; 3Dept. of Public Health and ClinicalMedicine, Rheumatology, UmeåUniversity, Umeå, Sweden; 4Dept. of Surgical and PerioperativeSciences, Surgery, Umeå University,Umeå, Sweden; 5Dept. of Surgical and PerioperativeSciences, Orthopaedics, Umeå University,Umeå, Sweden
Abstract
Anti-cholinergic agents are used in thetreatment of several pathological conditions.Therapy regimens aimed at up-regulatingcholinergic functions, such as treatment withacetylcholinesterase inhibitors, are also cur-rently prescribed. It is now known that not onlyis there a neuronal cholinergic system but alsoa non-neuronal cholinergic system in variousparts of the body. Therefore, interference withthe effects of acetylcholine (ACh) broughtabout by the local production and release ofACh should also be considered. Locally pro-duced ACh may have proliferative, angiogenic,wound-healing, and immunomodulatory func-tions. Interestingly, cholinergic stimulationmay lead to anti-inflammatory effects. Withinthis review, new findings for the locomotorsystem of a more widespread non-neuronalcholinergic system than previously expectedwill be discussed in relation to possible newtreatment strategies. The conditions discussedare painful and degenerative tendon disease(tendinopathy/tendinosis), rheumatoid arthri-tis, and osteoarthritis.
New aspects on the usefulnessof interference with acetylcholine effects: basis forthe present review
Medications interfering with the effects ofacetylcholine (ACh) are frequently used today.Anti-cholinergic agents are widely used in themanagement of overactive urinary bladder andof obstructive lung disease. However, treat-ments leading to the upregulation of choliner-gic activity are increasingly applied. New pos-sibilities for interference with cholinergiceffects are currently discussed. This is relatedto the existence of a widespread non-neuronalcholinergic system.
The present review summarizes hithertoknown aspects of possible treatment strategiesconcerning interference with cholinergiceffects. That includes inflammation in general,cancer, and pain conditions. However, majorfocus is devoted to discussions concerningconditions afflicting the locomotor system andfor which a non-neuronal cholinergic systemhas been recently shown to exist. More specif-ically, the conditions discussed are rheumatoidarthritis (RA) and osteoarthritis (OA), andchronically painful tendons (tendinopathy)with degenerative-like tissue changes (tendi-nosis). Tendinopathy is a condition in whichthere is chronic pain in a tender portion of thetendon. The tendons most frequently affectedare the Achilles and patellar tendons. When, inaddition to chronic pain, there are structuraltissue changes of a degenerative-like nature,as seen by ultrasonography or magnetic reso-nance imaging (MRI), or by histological exam-ination, the condition is called tendinosis. Inthe following text, our recent observations con-cerning Achilles and patellar tendinosis will bediscussed. Nothing at all has been previouslyknown concerning the existence of a non-neu-ronal cholinergic system in tendinosis or thesynovial tissue of patients with RA or OA. Thecurrent review will, therefore, give new direc-tions concerning the cholinergic system inthese conditions.
Acetylcholine production andacetylcholine receptors
ACh was the first neurotransmitter to beidentified. It is synthesized from choline andacetyl-CoA via the activity of choline acetyl-transferase (ChAT).1,2 In addition to productionvia ChAT, ACh can be produced by carnitineacetyltransferase (CarAT). ACh is then trans-ferred to synpatic vesicles by vesicular acetyl-choline transporter (VAChT).3 ACh is degradedby cholinesterases.4 It is converted into the
inactive metabolites choline and acetate byacetylcholinesterase (AChE). It is well knownthat the effects of ACh are mediated via mus-carinic (G-protein coupled) and nicotinic (lig-and-gated ion channels) ACh receptors.5 Themuscarinic ACh receptors (mAChRs) aremetabotrophic, and are stimulated by mus-carine and ACh, whilst the nicotinic AChreceptors are ionotrophic receptors stimulatedby nicotine and ACh. Muscarine is an alkaloidextracted from certain mushrooms, and nico-tine is an alkaloid substance found in tobacco.Five subtypes of mAChRs have been identified(M1-M5), each with different functions andproperties.6 Certain features regarding theexpression patterns of the mAChRs in regionsoutside the central nervous system are wellknown, including the fact that the M2 receptorsubtype comprises more than 90% of themAChRs in the heart.7 The main ACh receptorin the smooth muscle of the gastrointestinaltract is the M2 receptor.8 The mAChRs associat-ed with smooth muscle cells are predominant-ly of the M2 and M3 subtypes. The nicotinicreceptors (nAChRs) are pentameric complexesconsisting of a large number of different alpha-and beta-subunits.9 The various nAChRs have
Orthopedic Reviews 2009; volume 1:e11
Correspondence: Sture Forsgren, Dept. of Integrative Medical Biology, Anatomy,Umeå University, SE-901 87 Umeå, SwedenE-mail: [email protected]
Key words: acetylcholine, tendinosis, tendinopa-thy; rheumatoid arthritis, osteoarthritis.
Acknowledgments: the authors thank Ms. U.Hedlund for excellent technical services, Drs. O.Grimsholm and M. Jönsson, and Mr. G.Andersson, for scientific co-operation on cholin-ergic research. Mr. Andersson is also acknowl-edged for the digital artwork of Figure 1. We final-ly thank Dr. A Scott for valuable collaboration ontendon research. The studies which are the basesfor this review have been financially supported bythe Faculty of Medicine at Umeå University, theSwedish National Centre for Research in Sports,the County Council of Västerbotten, the ArnerskaResearch Foundation, Lions Cancer ResearchFoundation, Dagmar Ferb’s MemorialFoundation, the J.C. Kempe MemorialFoundation Scholarship Fund, the Magn. BergvallFoundation, Muscle Fund North, and the J.C.Kempe and Seth M. Kempe MemorialFoundations, Örnsköldsvik.
Received for publication: 27 May 2009.Revision received: 23 June 2009.Accepted for publication: 26 June 2009.
This work is licensed under a Creative CommonsAttribution 3.0 License (by-nc 3.0)
©Copyright S. Forsgren et al., 2009Licensee PAGEPress, ItalyOrthopedic Reviews 2009; 1:e11doi:10.4081/or.2009.e11

different physiological functions. The best-characterized nAChR, which, due to its rela-tionship to immune functions, is further dis-cussed below, is the α7nAChR.
There are both neuronal andnon-neuronal cholinergic systems
ACh is a neurotransmitter in the central, aswell as in the peripheral, nervous system. It isa major transmitter in the autonomic nervoussystem, the principal transmitter of theparasympathetic part of this system, and aneurotransmitter in all autonomic ganglia. Onthe other hand, nerve-related reactions forChAT or CarAT have, to the best of our knowl-edge, not been shown in synovial tissues ofjoints or for tendon tissue. It is well known thatACh synthesis is not only restricted to neu-rons; there is also a non-neuronal cholinergicsystem. Accordingly, ACh is produced in sur-face epithelia, such as those of the airways10
and the intestine.11,12 Furthermore, the ker-atinocytes of the skin13 and cells in the urothe-lium14,15 are also ACh-producing. Additionally,cells in blood vessel walls,16,17 as well as fibrob-lasts in various locations,18 show production of
ACh. An occurrence of expression of non-neu-ronal ACh and ACh-synthesizing activity hasalso previously been shown for a variety ofplants and organisms including fungi, algaeand bacteria.19,20 The occurrence of a non-neu-ronal ACh production includes the situation forcertain cancer cells and for inflammatory cells.It has been shown that ACh is synthesized bycells of small lung cell carcinoma,21,22 colon can-cer,23,24 and breast cancer.25 Furthermore,inflammatory cells have been shown to pro-duce ACh.26 The effects of ACh on the functionsof inflammatory cells do occur principally viaeffects on the α7nAChR. Recently we foundthat the tendon cells (tenocytes) in patellar27,28
and Achilles29 tendons showed evidence of AChproduction. This was observed by analysis ofChAT and VAChT reactions at both protein andmRNA level. We have furthermore observedthat mononuclear- and fibroblast-like cells ofthe synovium of patients with severe RA andOA show ChAT expression.30 We have alsomade original findings regarding the colon ofpatients with ulcerative colitis (UC). In thecolon from patients with UC, cells, includinginflammatory cells in the lamina propria, cellsof the blood vessel walls, and cells of theepithelial layer, were identified as being capa-ble of ACh synthesis.31 In the following text, theaspects of the non-cholinergic effects in thelocomotor system are focused upon. In our
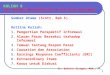
recent studies on the synovium of joints, andtendons of man we found a marked occurrenceof the M2 type of mAChRs. Immunoreactionsfor the M2 receptor were observed for the teno-cytes, nerves, and blood vessel walls of Achillesand patellar tendons,27-29 and were also noted inthe synovial tissue from patients with RA aswell as from those with OA. An interestingaspect noted is the fact that the evidence for anon-neuronal cholinergic system in tendonand synovial tissues is particularly apparent inpathological situations. Thus, the levels ofexpression of enzymes catalyzing ACh produc-tion, as well as the levels of expressions ofmAChRs, were much more evidently seen inderanged and chronically painful tendinosistendons than in normal tendons.27-29 A schemat-ic drawing summarizing the non-neuronalcholinergic system of human tendinosis ten-dons is shown in Figure 1. Furthermore,expression of enzyme related to ACh produc-tion was marked in specimens of the synovialtissue of knee joints of RA patients exhibitingpronounced invasion of mononuclear-likecells, as well as showing the occurrence ofnumerous fibroblast-like cells.30 Such anexpression was also noted for mononuclear-like and fibroblast-like cells in specimens ofthe synovial tissue of OA patients.30
Concerning the synovial specimens of both RAand OA patients, these corresponded to speci-mens of biopsies taken during prosthesis oper-ations.
The functions of the neuronaland non-neuronal cholinergicsystems
Effects in generalThe responses in the effector organs to
impulses leading to release and effects of AChfrom the autonomic nervous system are welldescribed in textbooks. Typical features are adecrease in heart rate, contraction of thesphincter muscle of the iris, contraction ofbronchial muscles, and increased exocrinesecretion from the pancreas. These are effectsof ACh released from postganglionic parasym-pathetic nerve fibres. In the intestine, forexample, it is well known that stimulation byACh leads to an increase in smooth musclecontraction and a stimulation of secretion.
Based on results of studies on experimentalanimals, it has been theorized that AChreleased from cholinergic nerves is involved invasoregulation in joints.32 However, asdescribed above, it has not yet been provenwhether cholinergic nerves are present in thesynovial tissues. There are no reports of effectsof neuronally-released ACh within tendons.
The function of non-neuronal ACh is in
Review
[page 40] [Orthopedic Reviews 2009; 1:e11]
Figure 1. Schematic drawing of human tendon tissue showing the occurrence of a non-neuronal cholinergic system in tendinosis. Violet dots represent acetylcholine (ACh) thatis locally produced in tenocytes of pathological appearance (unfilled arrow). Normallooking tenocytes (filled arrow) do not produce ACh.27-29 ACh can influence 1) nerves, 2)cells of blood vessel walls, and 3) the tenocytes themselves. These structures have beenshown to be supplied with muscarinic ACh receptors of subtype M2 (M2R).27-29 Image byGustav Andersson.

principle related to autocrine/paracrineactions.10,20 Thus, the cells types producing AChare also equipped with receptors for this sub-stance. That includes, for example, the situa-tion for the non-neuronal cholinergic systemof the granulose cells in the human ovary.33
More precisely, the functions of the non-neu-ronal cholinergic system are mainly related toeffects on differentiation and growth, secre-tion, barrier functions, and immuno-modula-tion (for reviews, see Kawashima and Fujii34
and Wessler and Kirkpatrick20). Important ACheffects are those on angiogenesis,35,36 prolifera-tion rates,13,37,38 and wound healing.13,39
It is frequently emphasized that the AChthat is produced by the inflammatory cells haseffects on those cells, suggesting that AChmodulates the activity of inflammatory cells viaautocrine and paracrine loops.40 The influ-ences of ACh on the inflammatory cells may berelated to the time scale with acute stimula-tion leading to proinflammatory effects whilstchronic stimulation, on the other hand, hasanti-inflammatory effects.40 The overwhelmingmajority of studies on ACh effects in relation toinflammation do, nevertheless, support theview that ACh has anti-inflammatory effects(see further below).
The cholinergic anti-inflammatorypathway
Based on findings by several groups, theexistence of a so-called ‘cholinergic anti-inflammatory pathway’ has been proposed.41-44
This implies that ACh, released in response toactivation of nerve fibres like those of thevagal nerve, has effects on local inflamma-tion.45 Electrical stimulation of the vagus nerveleads to inhibition in the synthesis of TNFα,and attenuates the release of different pro-inflammatory cytokines.41,42 The vagal anti-inflammatory effects are known to be mediat-ed by the Jak2/STAT3 signaling onmacrophages42 or via inhibition of the tran-scription factor NF-kappaB.46 It should beremembered that there are variations betweendifferent organs concerning the occurrence ofanti-inflammatory effects via stimulation ofthe vagus nerve. Such a stimulation has amarked anti-inflammatory effect within thegastrointestinal tract,42 but a limited anti-inflammatory effect in the lungs.41
The spleen has been shown to be importantin mediating the antiinflammatory effects thatoccur in response to electrical stimulation ofthe vagus nerve.47
The cholinergic antiinflammatory pathwayis shown to regulate the production of TNFα bymacrophages via effects on preganglionic neu-rons related to the vagus nerve and via effectson postganglionic neurons originating in theceliac-superior mesenteric plexus and project-ing within the splenic nerve.48 The findings
that muscarinic receptor antagonists mediateanti-inflammatory actions favor the suggestionthat there is a cholinergic anti-inflammatorypathway.49 Nevertheless, it is the α7nAChRthat is the ACh receptor, that is the essentialreceptor for the inhibition of cytokine synthe-sis via the cholinergic anti-inflammmatorypathway.50,51 One structure that may be a targetfor anti-inflammatory cholinergic mediators isthe endothelium, as the endothelium is a keyregulator of leukocyte trafficking duringinflammation.52
The occurrence of anti-inflammatory effectsof the parasympathetic nervous system hasbeen shown in different ways. The anti-inflam-matory effects that cholecystokinin is known tohave are thus mediated by the vagus nerve53
and activation of the vagus nerve preventsmanipulation-induced inflammation of thesmooth muscle layers.42 Interestingly, it is sug-gested that anti-inflammatory effects not onlyoccur via the neuronal but also the non-neu-ronal cholinergic system,40 and furthermore,that the release of non-neuronal ACh fromlocal cells, such as inflammatory cells, is trig-gered by neuronally-released ACh.20
Changes in magnitude of thenon-neuronal cholinergicsystem: occurrence of up- and downregulations
The new information obtained in our labora-tory shows that there is a more clearly markednon-neuronal cholinergic system in tendinosistendons than in normal tendons.27-29 Further-more, there is an extensive expression of M2
receptors in tendinosis tendons, but clearlyless so in normal tendons. It is also a fact thatthe non-neuronal cholinergic system is repre-sented to a large extent in the synovial tissuein RA if the tissue is heavily infiltrated withinflammatory cells and fibroblasts.30 Many ofthese cells show expression for the ACh-syn-thesizing enzyme ChAT. Upregulation of thecholinergic system also occurs in other situa-tions, for example such an upregulation hasbeen reported for cancer cells.22 It has alsobeen shown that the cholinergic system is up-regulated in the superficial skin in atopic der-matitis.54 However, the situation is apparentlydifferent in certain acute situations. Thus,there is a decrease in non-neuronal ACh syn-thesis in an acute model of allergic airwayinflammation.55 Whether upregulation of AChproduction in non-neuronal cells is of positiveor negative character for the tissue is onlypartly known. Obtaining a reduction in pul-monary ACh content in experimental studiesof lung injury was considered to be of positivecharacter, protecting the lung tissue.56 Further
aspects on this question are discussed below.For many conditions it seems as if anincreased ACh effect would be welcome.Cancer is a major exception (see furtherbelow).
Treatments leading to interfer-ence with cholinergic effects
General aspects Anti-cholinergics are useful in certain clini-
cal situations. However, treatments leading tocholinergic upregulation may also have a posi-tive outcome. Thus, as will be further dis-cussed below, treatment with AChE inhibitors,leading to increased ACh levels, has beenfound to be a treatment of choice inAlzheimer’s disease. Furthermore, the conceptof the cholinergic anti-inflammatory pathwayimplies that inducing increased ACh effectsmay be favorable for inflammatory situations.
There are also other reports suggesting thatincreased ACh effects may be helpful clinically.The results of recent studies on cystic fibrosissuggest that treatment with muscarinic recep-tor agonists may be beneficial.57 Furthermore,in a recent experimental study on acute aller-gic airway inflammation, the occurrence of adownregulations of non-neuronal ACh synthe-sis and release machinery was furthermoresuggested to contribute to epithelial sheddingand ciliated dysfunction.55
Treatment with AChE inhibitors Over recent decades, a number of therapies
aimed at up-regulating cholinergic functionhave been tested as treatment for Alzheimer’sdisease, with AChE inhibitors being the groupof substances that are best developed (for areview, see Shah et al.58). Thus, compoundsfrequently used nowadays for the treatment ofmild forms of this disease are blockers ofAChE activity59. The basis for this is the well-known fact that there is a loss of cholinergicfunction in Alzheimer’s disease and thatAChE is the enzyme that degrades ACh. AChEinhibitors are also used in myastheniagravis.60 Cholinergic agonism, via administra-tion of an AChE inhibitor, has also been test-ed for multiple sclerosis (MS) patients, andthis agonism was hereby shown to alter cog-nitive processing and to enhance brain func-tional connectivity.61 In studies on experimen-tal autoimmune encephalomyelitis, which isa model for the pathology of MS, it was foundthat treatment with an AChE inhibitor sup-pressed neuroinflammation and showedimmunomodulatory activity.62 A scheme of theanti-inflammatory outcome that is achievedusing AChE inhibitors during experimentalneuroinflammation has recently been depict-
Review
[Orthopedic Reviews 2009; 1:e11] [page 41]

ed by Brenner and collaborators.63 Theseauthors found that the AChE inhibitorsinduced cholinergic upregulation and effectson the neuroinflammation via α7nAChRexpressing cells.63
It has been shown that treatment of micewith AChE inhibitors attenuates the produc-tion of interleukin-1beta in both the hip-pocampus and blood, showing that choliner-gic enhancement produces both central andperipheral anti-inflammatory effects.64 Theresults of studies on endotoxemia also sug-gest that centrally acting cholinergicenhancers may have anti-inflammatory prop-erties.43 Inhibition of brain AChE via treat-ment with AChE inhibitors has recently beenshown to suppress systemic inflammation,suggesting that the brain’s cholinergic mus-carinic networks communicate with the anti-inflammatory pathway thereby suppressingperipheral inflammation.65
Obtaining of cholinergic anti-inflammatory effects via thenicotinic pathway
Nicotine is more active than ACh in inhibit-ing the production of pro-inflammatory media-tors by macrophages.51 The anti-inflammatoryeffects of nicotine on these cells can be coun-teracted by selective α7-antagonists.45
Influences on the nAChRs via nicotine havebeen tested in clinical situations, especially inUC. The therapeutic benefit is, however, limit-ed, and troublesome side-effects of nicotineoccur.66
The effects of ACh stimulation on theα7nAChR may, due to the known relationshipbetween this receptor and inflammation, be ofimportance in inflammatory conditions.Accordingly, it is suggested that α7nAChRsplay a critical role in the protection against thedevelopment of neurodegenerative diseases.67
The use of selective α7nAChR agonists has,furthermore, been shown to be of value inorder to diminish cytokine production bymacrophages and inflammation in several ani-mal models of inflammation (for a review, seede Jonge and Ulloa68).
Possible usefulness of interference with cholinergiceffects in certain conditions
CancerIt is well-known that ACh receptors, includ-
ing both mAChRs and nAChRs, are functional-ly present on certain cancer cells (for reviews,see Paleari et al. 200869 and Song and Spindel200822). This includes the cancer cells in smallcell lung carcinoma (SCLC), in which the cells
have been shown to synthesize and secreteACh, and to be equipped with mAChRs andnAChRs.21 The M3 mAChR is over-expressed intumor cells of colon cancer.70 In recent studiesit was shown that cells of the human colon can-cer cell line HT-29 express the α7nAChR sub-type.24
It is known that the level of cholinergic sig-naling is up-regulated in squamous cell carci-noma,22 and that stimulation of cells of mam-mary adenocarcinoma cell lines with carbacolincreases their proliferation via M3 mAChR-mediated pathways.71 It is suggested thatmAChR antagonists may be useful adjuncts forSCLC treatment,21 and that blocking choliner-gic signaling can limit the growth of squamouslung carcinoma.22 In accordance with the lattersuggestion, α7nAChR antagonists are antici-pated to be a useful adjunct to the treatment ofsuch lung cancer.72 The potential implicationsof mAChRs in tumor progression and the pos-sible use of muscarinic ligands in cancer ther-apy have been recently reviewed.73 The factthat cholinergic signaling is increased in cer-tain cancers may reinforce the usefulness ofACh blockade in these situations. It should bestressed that stimulation of ACh leads to anincrease in cell proliferation13,38 and to angio-genesis.36 The fact that ACh stimulation leadsto these features indeed favors the proposalthat blocking the effects of ACh may be benefi-cial in cancers showing an upregulation ofcholinergic features. The fact that ACh inhibitslong-term hypoxia-induced apoptosis in mousestem cells,74 and that nicotine increases cellgrowth of a human colon cancer cell line, theeffect being depressed by an antagonist to theα7nAChR,24,75 also supports such a suggestion.
Pain conditionsInterference with cholinergic effects may
have some relevance in relation to pain.76,77
ACh has been shown to induce pain whenapplied to human skin.78 However, cholinergiceffects have mainly been found to be of ananalgesic nature. Administration of mucarinicagonists with effects on the central nervoussystem can thus induce pronounced analgesiceffects.77 Inflammatory joint pain can be partlymodulated via muscarinic cholinergic recep-tors as shown in animal model studies,79 e.g.,analgesia induced by an AChE inhibitor hasbeen shown in a rat model.80 The possible use-fulness of targeting the mAChRs in antinoci-ception has been recently discussed.73
Chronic pain is the major symptomatic fea-ture of tendinosis, and the pain mechanismsfor this disease are still largely unknown andfrequently discussed.81 The aspects of tendonpain in relation to the non-neuronal choliner-gic system in tendon tissue are discussedbelow.
TendinosisIn tendinosis, a non-inflammatory degener-
ative-like condition in which an increase intissue cells (tenocytes) and a hypervascularity/neovascularization are typical phenomena,82-84
it is likely that the proliferation of tenocytesand the angiogenesis are related to an initialtissue healing process in response to mechan-ically induced micro-trauma. The tenocytes arethe cells that produce not only the collagen butalso various signal substances that are likely tohave important roles in the turnover of theextracellular matrix.85 However, the blood ves-sel changes may in the long run be a drawback.Thus, hypervascularity and neovascularizationhave been correlated with the chronic painexperienced in tendinosis,86 and new treat-ment methods focusing on destroying theregion with hypervascularity/neovasculariza-tion by injection of the sclerosing substancepolidocanol have not only led to pain relief butin the long-term perspective also to tendonremodeling.87-89
Given the information that tenocytes oftendinosis tendons produce ACh, and thatthere are mAChRs on the tenocytes and on thecells of the blood vessel walls,27,29 it is possiblethat the non-neuronal ACh might have effectson cell proliferation, collagen production, andblood vessel regulation in tendinosis. It isalready known that ACh can increase the pro-liferation of myofibroblastic cells,90 and thatstimulation of ACh receptors on certain fibro-blasts may augment collagen accumulation.91
Also, as previously discussed, agonists of AChreceptors are known to promote angiogene-sis.36 It might be that treatments that lead toincreased cholinergic effects on the tenocytesand the blood vessels might be attractive inearly stages of tendinosis. At these stages, anincreased tenocyte population and anincreased vascularity are likely to be of valuefor the tendon. On the other hand, less cholin-ergic influences on the blood vessels may bedesirable in chronic stages. It may also be thatan excess of tenocytes is a drawback for ten-don function in the long run. The functionalimportance of the ACh production in the teno-cytes, and the marked existence of mAChRs onthese cells, as well as on blood vessel walls intendinosis tendons, should be furtherexplored.
One should bear in mind that signal sub-stances other than ACh, e.g., catecholamines,92-94
and glutamate,95 are produced by tenocytes intendinosis tendons and that the tenocytes areequipped with receptors for various signal sub-stances. Furthermore, there is a production ofneurotrophins in the tenocytes and the exis-tence of the p75 neurotrophin receptor inthese cells.96 This means that interactionsbetween a number of substances that are deliv-ered locally have to be considered. Some of thesubstances produced, such as glutamate, are
Review
[page 42] [Orthopedic Reviews 2009; 1:e11]

likely to be toxic rather than related to woundhealing. It may be that interference withcholinergic effects should be combined withinterference with the effects of other type(s)of signal substances.
To further understand the pain mechanismsin tendinosis, the newly developed treatmentsfor this condition should be considered. Thesclerosing injection treatment, mentionedabove, targets the regions of tendinosisAchilles and patellar tendons showing patho-logically high blood flow.87,88 These regions con-form to outer parts of the tendons (the para-tendinous connective tissue). Our findings ofan abundance of mAChRs in blood vessel wallsin tendinosis tendons27,29 show that these ves-sels are under marked cholinergic influence.Nevertheless, it is probable that the pain-reliefis not primarily related to effects on the vascu-lature but to effects on the nerves located closeto the blood vessels. Consequently, it is inter-esting to note that we have also found mAChRson nerve fascicles in the tendinosis tendons.27
To what extent the ACh effects, and those ofother nerve signal substances such as sub-stance P in the sensory innervation, are relat-ed to the pain symptoms in tendinosis must,however, await further studies.
Rheumatoid arthritis It is evident that the infiltrating inflammato-
ry cells, as well as the proliferating fibroblast-like cells, in the synovium become part of anon-neuronal cholinergic system in advancedRA, as verified by the expression of enzymerelated to ACh production (ChAT). Such cellsin synovial specimens of OA patients also showthis feature. In both conditions, the existenceof ChAT production was noted in specimensobtained after prosthesis operations. It may bethat the existence of ACh production in thesecells is related to trophic/modulatory and anti-inflammatory, rather than damaging, effects inthe chronic stages of RA and OA. This could beseen as an attempt to balance the productionof pro-inflammatory substances.
A favorable outcome in arthritis via cholin-ergic agonism has recently been shown exper-imentally in mice. Based on evaluating theeffect of adding nicotine to the drinking waterof mice with collagen-induced arthritis (CIA),the effects of vagotomy and the effects of treat-ment with an α7nAChR agonist, the authorsconcluded that there exists a cholinergic anti-inflammatory pathway in the murine CIAmodel of RA.97 An existence of α7nAChRs onthe fibroblast-like synoviocytes in human syn-ovium has also been shown and ACh was foundto inhibit cytokine expression through a post-transcriptional mechanism in these cells.98 Inthe study by Waldburger and collaborators,98 itwas suggested that the α7nAChR is a potentialtherapeutic target for RA. In studies on a ratmodel of knee joint inflammation published as
early as 1998, it was shown that a centrallyadministered AChE inhibitor, neostigmine, ledto enhanced levels of endogenous ACh, whichin turn functioned as an analgesia-modulatingcompound at both central and peripheral sitesof inflammatory pain.80
It can be speculated that treatments thatcontribute to additional ACh effects could beuseful in order to prevent the establishment ofa marked derangement and damage of synovialtissues in advanced stages of RA and OA. Suchinterference would, thus, possibly lead to anti-inflammatory features. In accordance withsuch a suggestion are the recent findings byGoldstein and collaborators.99 In their studies itwas shown that the occurrence of a dimin-ished cholinergic anti-inflammatory pathwayactivity is associated with increased levels ofhigh mobility group box-1, a cytokine that isimplicated in the pathogenesis of arthritis,100
in patients with RA.99
Conclusions
In summary, the basis for the present reviewis the fact that new indications for interfer-ence with cholinergic effects should be consid-ered. The reason is that there exists a wide-spread local production of ACh in various tis-sues; a non-neuronal cholinergic system. AChreceptors are present in parallel. The effects ofnon-neuronal ACh are related to autocrine/paracrine effects and effects on angiogenesis,cell proliferation, wound-healing, and inflam-mation. Our recent observations on the situa-tion in tendinosis, RA, and OA show that thereis an unexpected presence of a non-neuronalcholinergic system in these conditions. Thismeans that new ideas for interference withcholinergic effects are indeed welcome forthese situations.
What should be the focus for further studiesevaluating the usefulness of interfering withcholinergic effects in various parts of thebody? One field that should be further exam-ined is the development of selective nicotinicagonists for man. The fact that selectiveα7nAChR agonists have been found to havepositive effects in a dextran sulphate-inducedcolitis model for mice101 supports a suggestionthat such agonists may be useful in inflamma-tory disorders. The idea that the α7nAChR is atarget of importance in RA is discussed in thepresent review.
Another field that should be further concen-trated on is research on the possible useful-ness of AChE inhibitors in peripheral inflam-matory disorders.64,65 Questions related toselectivity and specificity are, however, alsowarranted in this case.
In conclusion, knowledge of the occurrenceof a marked local signal substance production
within a diseased tissue, and the occurrence ofreceptors for the substances within the tissue,is on the whole of great importance for estab-lishing new possible treatment strategies. It isobvious that it is not necessary that choliner-gic nerves are present in order to make AChavailable for the tissue; the substances can belocally produced. This means that not onlyshould the neuronal but also the non-neuronalcholinergic system be taken into considerationwhen considering new treatments interferingwith cholinergic effects. This includes disor-ders related to orthopedics, namely tendinosis,RA and OA.
References
1. Tuček S. The synthesis of acetylcholine inskeletal muscles of the rat. J Physiol 1982;322:53-69.
2. Tuček S. Choline acetyltransferase andthe synthesis of acetylcholine. In:Whittaker VP, editor. Handbook ofExperimental Pharmacology, 86, TheCholinergic Synapse. Berlin: SpringerVerlag 1988. p. 125-65.
3. Eiden LE. The cholinergic gene locus. JNeurochem 1998;70:2227-40.
4. Hoffman BB, Taylor P. Neurotransmission:the autonomic and somatic motor systems.In: Hardman JG, Limbird LE, editors.Goodman & Gilmnan's The PharmalogicalBasis of Therapeutics, 10th ed. New York:McGraw-Hill; 2001. p. 115-53.
5. Miyazawa A, Fujiyoshi Y, Unwin N.Structure and gating mechanism of theacetylcholine receptor pore. Nature 2003;423:949-55.
6. Felder CC. Muscarinic acetylcholine recep-tors: signal transduction through multipleeffectors. FASEB J 1995;9:619-25.
7. Li M, Yasuda RP, Wall SJ, Wellstein A,Wolfe BB. Distribution of M2 muscarinicreceptors in rat brain using antisera selec-tive for M2 receptors. Mol Pharmacol 1991;40:28-35.
8. Iino S, Nojyo Y. Muscarinic M2 acetyl-choline receptor distribution in theguinea-pig gastrointestinal tract. Neuro-science 2006;138:549-59.
9. Lloyd GK, Williams M. Neuronal nicotinicacetylcholine receptors as novel drug tar-gets. J Pharmacol Exp Ther 2000;292:461-7.
10. Wessler I, Kilbinger H, Bittinger F,Kirkpatrick CJ. The biological role of non-neuronal acetylcholine in plants andhumans. Jpn J Pharmacol 2001;85:2-10.
11. Klapproth H, Reinheimer T, Metzen J, et al.Non-neuronal acetylcholine, a signallingmolecule synthezised by surface cells ofrat and man. Naunyn Schmiedebergs Arch
Review
[Orthopedic Reviews 2009; 1:e11] [page 43]

Pharmacol 1997;355:515-23. 12. Ratcliffe EM, deSa DJ, Dixon MF, Stead
RH. Choline acetyltransferase (ChAT)immunoreactivity in paraffin sections ofnormal and diseased intestines. JHistochem Cytochem 1998;46:1223-31.
13. Grando SA, Pittelkow MR, Schallreuter KU.Adrenergic and cholinergic control in thebiology of epidermis: physiological andclinical significance. J Invest Dermatol2006;126:1948-65.
14. Lips KS, Wunsch J, Zarghooni S, et al.Acetylcholine and molecular componentsof its synthesis and release machinery inthe urothelium. Eur Urol 2007;51:1042-53.
15. Yoshida M, Masunaga K, Satoji Y, et al.Basic and clinical aspects of non-neuronalacetylcholine: expression of non-neuronalacetylcholine in urothelium and its clinicalsignificance. J Pharmacol Sci 2008;106:193-8.
16. Haberberger RV, Bodenbenner M, KummerW. Expression of the cholinergic genelocus in pulmonary arterial endothelialcells. Histochem Cell Biol 2000;113:379-87.
17. Ikeda C, Morita I, Mori A, et al. Phorbolester stimulates acetylcholine synthesis incultured endothelial cells isolated fromporcine cerebral microvessels. Brain Res1994;655:147-52.
18. Lips KS, Pfeil U, Reiners K, et al.Expression of the high-affinity cholinetransporter CHT1 in rat and human arter-ies. J Histochem Cytochem 2003;51:1645-54. 21
19. Horiuchi Y, Kimura R, Kato N, et al.Evolutional study on acetylcholine expres-sion. Life Sci 2003;72:1745-56.
20. Wessler I, Kirkpatrick CJ. Acetylcholinebeyond neurons: the non-neuronal cholin-ergic system in humans. Br J Pharmacol2008;154:1558-71.
21. Song P, Sekhon HS, Lu A, et al. M3 mus-carinic receptor antagonists inhibit smallcell lung carcinoma growth and mitogen-activated protein kinase phosphorylationinduced by acetylcholine secretion. CancerRes 2007;67:3936-44.
22. Song P, Spindel ER. Basic and clinicalaspects of non-neuronal acetylcholine:expression of non-neuronal acetylcholinein lung cancer provides a new target forcancer therapy. J Pharmacol Sci 2008;106:180-5.
23. Cheng K, Samimi R, Xie G, et al. Acetyl-choline release by human colon cancercells mediates autocrine stimulation ofcell proliferation. Am J PhysiolGastrointest Liver Physiol 2008;295:G591-7.
24. Pettersson A, Nilsson L, Nylund G, et al. Isacetylcholine an autocrine/paracrinegrowth factor via the nicotinic α7-receptorsubtype in the human colon cancer cell
line HT-29? Eur J Pharmacol 2009;609:27-33.
25. Espanol AJ, de la Torre E, Fiszman GL,Sales ME. Role of non-neuronal choliner-gic system in breast cancer progression.Life Sci 2007;80:2281-5.
26. Kawashima K, Fujii T. Expression of non-neuronal acetylcholine in lymphocytes andits contribution to the regulation ofimmune function. Front Biosci 2004;9:2063-85.
27. Danielson P, Alfredson H, Forsgren S.Immunohistochemical and histochemicalfindings favoring the occurrence ofautocrine/paracrine as well as nerve-relat-ed cholinergic effects in chronic painfulpatellar tendon tendinosis. Microsc ResTech 2006;69:808-19.
28. Danielson P, Andersson G, Alfredson H,Forsgren S. Extensive expression of mark-ers for acetylcholine synthesis and of M2
receptors in tenocytes in therapy-resistantchronic painful patellar tendon tendinosis- a pilot study. Life Sci 2007;80:2235-8.
29. Bjur D, Danielson P, Alfredson H, ForsgrenS. Presence of a non-neuronal cholinergicsystem and occurrence of up- and down-regulation in expression of M2 muscarinicacetylcholine receptors: new aspects ofimportance regarding Achilles tendontendinosis (tendinopathy). Cell Tissue Res2008;331:385-400.
30. Grimsholm O, Rantapaa-Dahlqvist S,Dalen T, Forsgren S. Unexpected finding ofa marked non-neuronal cholinergic sys-tem in human knee joint synovial tissue.Neurosci Lett 2008;442:128-33.
31. Jonsson M, Norrgard O, Forsgren S.Presence of a marked nonneuronal cholin-ergic system in human colon: study of nor-mal colon and colon in ulcerative colitis.Inflamm Bowel Dis 2007;13:1347-56.
32. McDougall JJ, Elenko RD, Bray RC.Cholinergic vasoregulation in normal andadjuvant monoarthritic rat knee joints. JAuton Nerv Syst 1998;72:55-60.
33. Fritz S, Fohr KJ, Boddien S, et al.Functional and molecular characterizationof a muscarinic receptor type and evidencefor expression of choline-acetyltransferaseand vesicular acetylcholine transporter inhuman granulosa-luteal cells. J ClinEndocrinol Metab 1999;84:1744-50.
34. Kawashima K, Fujii T. Basic and clinicalaspects of non-neuronal acetylcholine:overview of non-neuronal cholinergic sys-tems and their biological significance. JPharmacol Sci 2008;106:167-73.
35. Cooke JP. Angiogenesis and the role of theendothelial nicotinic acetylcholine recep-tor. Life Sci 2007;80:2347-51.
36. Jacobi J, Jang JJ, Sundram U, et al.Nicotine accelerates angiogenesis andwound healing in genetically diabetic
mice. Am J Pathol 2002;161:97-104.37. Grando SA, Crosby AM, Zelickson BD, Dahl
MV. Agarose gel keratinocyte outgrowthsystem as a model of skin re-epitheliza-tion: requirement of endogenous acetyl-choline for outgrowth initiation. J InvestDermatol 1993;101:804-10.
38. Metzen J, Bittinger F, Kirkpatrick CJ, et al.Proliferative effect of acetylcholine on rattrachea epithelial cells is mediated bynicotinic receptors and muscarinic recep-tors of the M1-subtype. Life Sci 2003;72:2075-80.
39. Tournier JM, Maouche K, Coraux C, et al.α3α5beta2-Nicotinic acetylcholine recep-tor contributes to the wound repair of therespiratory epithelium by modulatingintracellular calcium in migrating cells.Am J Pathol 2006;168:55-68.
40. Kawashima K, Fujii T. The lymphocyticcholinergic system and its contribution tothe regulation of immune activity. Life Sci2003;74:675-96.
41. Borovikova LV, Ivanova S, Zhang M, et al.Vagus nerve stimulation attenuates thesystemic inflammatory response to endo-toxin. Nature 2000;405:458-62.
42. de Jonge WJ, van der Zanden EP, The FO,et al. Stimulation of the vagus nerve atten-uates macrophage activation by activatingthe Jak2-STAT3 signaling pathway. NatImmunol 2005;6:844-51.
43. Pavlov VA, Ochani M, Gallowitsch-PuertaM, et al. Central muscarinic cholinergicregulation of the systemic inflammatoryresponse during endotoxemia. Proc NatlAcad Sci USA 2006;103:5219-23.
44. Tracey KJ. Physiology and immunology ofthe cholinergic antiinflammatory pathway.J Clin Invest 2007;117:289-96.
45. Tracey KJ. The inflammatory reflex.Nature 2002;420:853-9.
46. Wang H, Liao H, Ochani M, et al.Cholinergic agonists inhibit HMGB1release and improve survival in experi-mental sepsis. Nat Med 2004;10:1216-21.
47. Huston JM, Ochani M, Rosas-Ballina M, etal. Splenectomy inactivates the choliner-gic antiinflammatory pathway duringlethal endotoxemia and polymicrobial sep-sis. J Exp Med 2006;203:1623-8.
48. Rosas-Ballina M, Ochani M, Parrish WR, etal. Splenic nerve is required for choliner-gic antiinflammatory pathway control ofTNF in endotoxemia. Proc Natl Acad SciUSA 2008;105:11008-13.
49. Profita M, Giorgi RD, Sala A, et al.Muscarinic receptors, leukotriene B4 pro-duction and neutrophilic inflammation inCOPD patients. Allergy 2005;60:1361-9.
50. Ben-Horin S, Chowers Y. Neuroimmuno-logy of the gut: physiology, pathology, andpharmacology. Curr Opin Pharmacol 2008;8:490-5.
Review
[page 44] [Orthopedic Reviews 2009; 1:e11]

51. Wang H, Yu M, Ochani M, et al. Nicotinicacetylcholine receptor α7 subunit is anessential regulator of inflammation.Nature 2003;421:384-8.
52. Saeed RW, Varma S, Peng-Nemeroff T, etal. Cholinergic stimulation blocks endo-thelial cell activation and leukocyterecruitment during inflammation. J ExpMed 2005;201:1113-23.
53. Luyer MD, Greve JW, Hadfoune M, et al.Nutritional stimulation of cholecystokininreceptors inhibits inflammation via thevagus nerve. J Exp Med 2005;202:1023-9.
54. Wessler I, Reinheimer T, Kilbinger H, et al.Increased acetylcholine levels in skin biop-sies of patients with atopic dermatitis. LifeSci 2003;72:2169-72.
55. Lips KS, Luhrmann A, Tschernig T, et al.Downregulation of the non-neuronalacetylcholine synthesis and releasemachinery in acute allergic airway inflam-mation of rat and mouse. Life Sci 2007;80:2263-9.
56. Grau V, Wilker S, Lips KS, et al. Admini-stration of keratinocyte growth factordown-regulates the pulmonary capacity ofacetylcholine production. Int J BiochemCell Biol 2007;39:1955-63.
57. Wessler I, Bittinger F, Kamin W, et al.Dysfunction of the non-neuronal choliner-gic system in the airways and blood cells ofpatients with cystic fibrosis. Life Sci2007;80:2253-8.
58. Shah RS, Lee HG, Xiongwei Z, et al.Current approaches in the treatment ofAlzheimer's disease. Biomed Pharmaco-ther 2008;62:199-207.
59. Desai AK, Grossberg GT. Diagnosis andtreatment of Alzheimer's disease.Neurology 2005;64:S34-9.
60. Conti-Fine BM, Milani M, Kaminski HJ.Myasthenia gravis: past, present, andfuture. J Clin Invest 2006;116:2843-54.
61. Cader S, Palace J, Matthews PM.Cholinergic agonism alters cognitive pro-cessing and enhances brain functionalconnectivity in patients with multiple scle-rosis. J Psychopharmacol 2008 E-pubahead of print.
62. Nizri E, Irony-Tur-Sinai M, Faranesh N, etal. Suppression of neuroinflammation andimmunomodulation by the acetyl-cholinesterase inhibitor rivastigmine. JNeuroimmunol 2008;203:12-22.
63. Brenner T, Nizri E, Irony-Tur-Sinai M, etal. Acetylcholinesterase inhibitors andcholinergic modulation in MyastheniaGravis and neuroinflammation. JNeuroimmunol 2008;201-202:121-7.
64. Pollak Y, Gilboa A, Ben-Menachem O, et al.Acetylcholinesterase inhibitors reducebrain and blood interleukin-1beta produc-tion. Ann Neurol 2005;57:741-5.
65. Pavlov VA, Parrish WR, Rosas-Ballina M, et
al. Brain acetylcholinesterase activity con-trols systemic cytokine levels through thecholinergic anti-inflammatory pathway.Brain Behav Immun 2009;23:41-5.
66. Thomas GA, Rhodes J, Ingram JR.Mechanisms of disease: nicotine--a reviewof its actions in the context of gastroin-testinal disease. Nat Clin Pract Gastro-enterol Hepatol 2005;2:536-44.
67. Streit WJ. Microglia as neuroprotective,immunocompetent cells of the CNS. Glia2002;40:133-9.
68. de Jonge WJ, Ulloa L. The α7 nicotinicacetylcholine receptor as a pharmacologi-cal target for inflammation. Br JPharmacol 2007;151:915-29.
69. Paleari L, Grozio A, Cesario A, Russo P. Thecholinergic system and cancer. SeminCancer Biol 2008;18:211-7.
70. Yang WL, Frucht H. Cholinergic receptorup-regulates COX-2 expression andprostaglandin E(2) production in coloncancer cells. Carcinogenesis 2000;21:1789-93.
71. Espanol AJ, Sales ME. Different muscarincreceptors are involved in the proliferationof murine mammary adenocarcinoma celllines. Int J Mol Med 2004;13:311-7.
72. Grozio A, Paleari L, Catassi A, et al. Naturalagents targeting the α7-nicotinic-receptorin NSCLC: a promising prospective in anti-cancer drug development. Int J Cancer2008;122:1911-5.
73. Tata AM. Muscarinic acetylcholine recep-tors: new potential therapeutic targets inantinociception and in cancer therapy.Recent Pat CNS Drug Discov 2008;3:94-103.
74. Kim MH, Kim MO, Heo JS, et al.Acetylcholine inhibits long-term hypoxia-induced apoptosis by suppressing theoxidative stress-mediated MAPKs activa-tion as well as regulation of Bcl-2, c-IAPs,and caspase-3 in mouse embryonic stemcells. Apoptosis 2008;13:295-304.
75. Wong HP, Yu L, Lam EK, et al. Nicotine pro-motes cell proliferation via α7-nicotinicacetylcholine receptor and catecholami-nesynthesizing enzymes-mediated path-way in human colon adenocarcinoma HT-29 cells. Toxicol Appl Pharmacol2007;221:261-7.
76. Dussor GO, Helesic G, Hargreaves KM,Flores CM. Cholinergic modulation ofnociceptive responses in vivo and neu-ropeptide release in vitro at the level of theprimary sensory neuron. Pain 2004;107:22-32.
77. Wess J, Duttaroy A, Gomeza J, et al.Muscarinic receptor subtypes mediatingcentral and peripheral antinociceptionstudied with muscarinic receptor knock-out mice: a review. Life Sci 2003;72:2047-54.
78. Vogelsang M, Heyer G, Hornstein OP.Acetylcholine induces different cutaneoussensations in atopic and non-atopic sub-jects. Acta Derm Venereol 1995;75:434-6.
79. Baek YH, Choi DY, Yang HI, Park DS.Analgesic effect of electroacupuncture oninflammatory pain in the rat model of col-lagen-induced arthritis: mediation bycholinergic and serotonergic receptors.Brain Res 2005;1057:181-5.
80. Buerkle H, Boschin M, Marcus MA, et al.Central and peripheral analgesia mediatedby the acetylcholinesterase-inhibitorneostigmine in the rat inflamed knee jointmodel. Anesth Analg 1998;86:1027-32.
81. Khan KM, Cook JL, Maffulli N, Kannus P.Where is the pain coming from intendinopathy? It may be biochemical, notonly structural, in origin. Br J Sports Med2000;34:81-3.
82. Cook JL, Malliaras P, De Luca J, et al.Neovascularization and pain in abnormalpatellar tendons of active jumping ath-letes. Clin J Sport Med 2004;14:296-9.
83. Khan KM, Cook JL, Bonar F, et al.Histopathology of common tendino-pathies. Update and implications for clini-cal management. Sports Med 1999;27:393-408.
84. Movin T, Gad A, Reinholt FP, Rolf C. Tendonpathology in long-standing achillodynia.Biopsy findings in 40 patients. Acta OrthopScand 1997;68:170-5.
85. Riley G. Chronic tendon pathology: molec-ular basis and therapeutic implications.Expert Rev Mol Med 2005;7:1-25.
86. Cook JL, Malliaras P, De Luca J, PtasznikR, Morris M. Vascularity and pain in thepatellar tendon of adult jumping athletes: a5 month longitudinal study. Br J SportsMed 2005;39:458-61.
87. Alfredson H, Ohberg L. Sclerosing injec-tions to areas of neo-vascularisationreduce pain in chronic Achilles tendinopa-thy: a double-blind randomised controlledtrial. Knee Surg Sports Traumatol Arthrosc2005;13:338-44.
88. Hoksrud A, Ohberg L, Alfredson H, Bahr R.Ultrasound-guided sclerosis of neovesselsin painful chronic patellar tendinopathy: arandomized controlled trial. Am J SportsMed 2006;34:1738-46.
89. Willberg L, Sunding K, Ohberg L, et al.Treatment of Jumper's knee: promisingshort-term results in a pilot study using anew arthroscopic approach based on imag-ing findings. Knee Surg Sports TraumatolArthrosc 2007;15:676-81.
90. Oben JA, Yang S, Lin H, et al. Acetylcholinepromotes the proliferation and collagengene expression of myofibroblastic hepat-ic stellate cells. Biochem Biophys ResCommun 2003;300:172-7.
91. Sekhon HS, Keller JA, Proskocil BJ, et al.
Review
[Orthopedic Reviews 2009; 1:e11] [page 45]

Review
[page 46] [Orthopedic Reviews 2009; 1:e11]
Maternal nicotine exposure upregulatescollagen gene expression in fetal monkeylung. Association with α7 nicotinic acetyl-choline receptors. Am J Respir Cell MolBiol 2002;26:31-41.
92. Bjur D, Danielson P, Alfredson H, ForsgrenS. Immunohistochemical and in situhybridization observations favor a localcatecholamine production in the humanAchilles tendon. Histol Histopathol 2008;23:197-208.
93. Danielson P, Alfredson H, Forsgren S.Studies on the importance of sympatheticinnervation, adrenergic receptors, and apossible local catecholamine production inthe development of patellar tendinopathy(tendinosis) in man. Microsc Res Tech2007;70:310-24.
94. Danielson P, Alfredson H, Forsgren S. Insitu hybridization studies confirmingrecent findings of the existence of a local
nonneuronal catecholamine production inhuman patellar tendinosis. Microsc ResTech 2007;70:908-11.
95. Scott A, Alfredson H, Forsgren S. VGluT2expression in painful Achilles and patellartendinosis: evidence of local glutamaterelease by tenocytes. J Orthop Res 2008;26:685-92.
96. Bagge J, Lorentzon R, Alfredson H,Forsgren S. Unexpected presence of theneurotrophins NGF and BDNF and theneurotrophin receptor p75 in the tendoncells of the human Achilles tendon. HistolHistopathol. 2009;24:839-48.
97. van Maanen MA, Lebre MC, van der Poll T,et al. Stimulation of nicotinic acetyl-choline receptors attenuates collagen-induced arthritis in mice. Arthritis Rheum2009;60:114-22.
98. Waldburger JM, Boyle DL, Pavlov VA, et al.Acetylcholine regulation of synoviocyte
cytokine expression by the α7 nicotinicreceptor. Arthritis Rheum 2008; 58:3439-49.
99. Goldstein RS, Bruchfeld A, Yang L, et al.Cholinergic anti-inflammatory pathwayactivity and High Mobility Group Box-1(HMGB1) serum levels in patients withrheumatoid arthritis. Mol Med 2007;13:210-5.
100.Pullerits R, Jonsson IM, Verdrengh M, etal. High mobility group box chromosomalprotein 1, a DNA binding cytokine, inducesarthritis. Arthritis Rheum 2003;48:1693-700.
101.Ghia JE, Blennerhassett P, Kumar-Ondiveeran H, et al. The vagus nerve: atonic inhibitory influence associated withinflammatory bowel disease in a murinemodel. Gastroenterology. 2006;131:1122-30.