Embed Size (px)
Citation preview

OBIECTIVUL GENERAL 5. : Managementul durabil al resurselor genetice animaleObiectivul specific 5.1: Ameliorarea genetică a populaţiilor de animale (rase, linii) cu status normal
Numărul/codul proiectului : ADER 5.1.3Denumirea proiectului: CUANTIFICAREA STATISTICO-INFORMAŢIONALĂ A
BIODIVERSITĂŢII LA RASELE DE OVINE CRESCUTE ÎN ROMÂNIACU AJUTORUL MARKERILOR GENETICI
PARTENERIContractor: Staţiunea de Cercetare-Dezvoltare pentru Creşterea Ovinelor şi Caprinelor
Popăuţi-Botoşani – Conducător proiect (CP)Agent economic: Societatea Comercială AGRO IND COM SRL Botoşani – Partener 1 (P1)
Contract nr.: 5.1.3./22.10.2015
Anul începerii: 22.11.2015; Anul finalizării: 15.12.2018; Durata (luni): 38
Director de proiect: Dr. biolog Hrincă GheorgheDate de contactTel: 0742485444;E-mail: [email protected]

Proiect ADER 5.1.3. / Faza 2 = Raport anual 2016
Data începerii fazei 2: 16.12.2015;Data finalizării fazei 2: 30.06.2016
Buget: 93.750 lei (CP = 88.750 lei ; P1 = 5.000 lei)
Denumirea Fazei 2
IDENTIFICAREA ŞI ANALIZA UNOR MARKERI GENETICO-BIOCHIMICI ŞI GENETICO-MOLECULARI LA OVINE
Denumirea proiectului
CUANTIFICAREA STATISTICO-INFORMAŢIONALĂ A BIODIVERSITĂŢII LA RASELE DE OVINE CRESCUTE ÎN ROMÂNIA CU AJUTORUL MARKERILOR GENETICI

Obiectivele fazei: ● identificarea variantelor genetice de proteine polimorfe (hemoglobină, transferină, albumină,amilază) şi elemente chimice cu variabilitate discontinuă (potasiu sanguin) la ovinele dinRomânia;● evidenţierea unor markeri moleculari (calpastatina, -lactoglobulina, s1-cazeina) la ovineledin România;● descrierea structurii la locii determinanţi ai sistemelor genetico-biochimice şi genetico-moleculare la ovinele din România;;● studiu tehnico-ştiinţific privind însuşirile morfologice de exterior ale ovinelor şi descriereaarealelor meteo-geo-climatice şi floristice în care sunt crescute şi exploatate principalele rasede ovine româneşti;● diseminarea de cunoştinţe şi a rezultatelor experimentale realizate în faza a II-a aproiectului.;
Obiectivul proiectului:Principală ţintă a proiectului constă în identificarea unor markeri genetici pentru
realizarea unui management sustenabil al patrimoniului genetic şi de conservare abiodiversităţii la specia ovină, utilizând concepte de statistică informaţională.
Implementarea sa presupune realizarea unor cercetări complexe multidisciplinare şiinterdisciplinare prin interconectarea domeniilor biologic (genetică şi biochimie), zootehnic(creşterea şi ameliorarea animalelor), veterinar (asigurarea vigorii animalelor), matematic(fundamentarea metodologică a proiectului) şi informatic (aplicarea precisă şi promptă ametodologiei şi pentru extrapolare experimentală) în activităţi de parteneriat între o unitate decercetare-dezvoltare şi un agent economic din domeniul creşterii animalelor.

Rezultate preconizate pentru atingerea obiectivelor fazei:♦ Buletin de analize de laborator,;♦ Profil genetico-biochimic şi genetico-molecular la ovinele din România ;♦ Profil morfologic şi econogenic la ovinele din România,;♦ Articole ştiinţifice publicate în reviste de specialitate;♦ Comunicări susţinute la manifestări ştiinţifice naţionale şi internaţionale;♦ Raport de experimentare;♦ Raport de cercetare etapă.
Activităţi desfăşurate pentru atingerea obiectivelor fazei:Activitatea II.1 - Recoltarea probelor biologice de la ovine din ferme de elită, înmulţire şi
producţie şi prelucrarea lor primară (CP, P1);Activitatea II.2 - Determinarea şi analiza tipurilor genetice de hemoglobină, transferină,
albumină, amilază şi potasiu sanguin la ovine din ferme de elită, înmulţire şi producţie (CP);Activitatea II.3 - Determinarea şi analiza unor markeri moleculari la ovine din ferme de elită,
înmulţire şi producţie (P1);Activitatea II.4 - Urmărirea şi consemnarea unor însuşiri de morfologie externă şi de
adaptabilitate a diferitelor rase şi subpopuţii de ovine din ferme de elită, înmulţire şi producţie şicaracterizarea lor în context econogenic (CP, P1);
Activitatea II.5 - Susţinerea de comunicări şi publicarea de articole (CP).

Motivaţia cercetării
Creşterea animalelor pe baze ştiinţifice necesită din partea celor care activează în acest domeniucunoaşterea profundă şi detaliată a legilor şi fenomenelor care guvernează formarea, dezvoltarea şi evoluţiaontogenetică a organismelor animale, precum şi ale celor care le determină potenţialul de productivitate.Locul central în acest demers revine geneticii, ştiinţă care se ocupă cu studiul eredităţii, variabilităţii şireproducţiei organismelor. Multă vreme, genetica în domeniul creşterii animalelor a urmărit aspectelecantitative, biometrice, măsurabile, transpozabile într-o curbă de distribuţie normală, caracterizând populaţiilede animale în ansamblul lor. Recent însă, anumite tronsoane ale geneticii au abordat însuşirile ereditarecalitative prin care indivizii unei populaţii pot fi încadraţi în clase distincte, prin care se cuantifică cu mareexactitate patrimoniul genetic al taxonilor de interes economic și științific prin instrumente și concepte aleunor discipline de graniţă, precum genetica biochimică şi imunogenetica, dar mai ales genetica moleculară.
Investigaţiile genetico-biochimice şi genetico-moleculare au scos la iveală că un număr mare debiostructuri prezintă determinism genetic. Această fază a proiectului şi-a propus investigarea unor sistemepolimorfe din domeniul geneticii biochmice (sistemele hemoglobinic, transferinic, albuminic, amilazic, potasic)şi al geneticii moleculare (sistemele calpastatinic, -lactoglobulinic şi αs1-cazeinic) la principalele rase deovine din România (Merinos de Palas, Ţigaie, Ţurcană, karakul de Botoşani, Rasa de Carne Palas şi Rasa deLapte Palas) care, pe lângă aspectul lor corelaţional cu însuşirile de productivitate animală, au o însemnatărelevanţă pentru biodiversitatea speciei ovină. Totodată, pentru a vedea rolul factorilor tehnologici şi selectiviîn fixarea diferitelor gene în patrimoniul lor genetic, am descris rasele şi din punct de vedere econogenic(cadru natural, condiţii meteo-geo-climatice, habitat, context social şi cultural, specific productiv, tehnologiiaplicate, tradiţii din zonele de creştere a ovinelor etc.). Împreună cu alţi markeri (imunogenetici, microsateliţietc.) care vor fi investigaţi în următoarea fază de cercetare, aceste date vor constitui edificiul experimentalnecesar realizării managementului sustenabil a resurselor genetice la rasele de ovine din România, dar şi cuextensie la întreaga specie ovină de pe întreg mapamondul, graţie caracterului său de inter- şimultidisplinaritate dintre genetică, zootehnie, medicină veterinară, ecologie, etologie, matematică şiinformatică.

Metodologia cercetăriiMaterial biologic
Cercetările pentru atingerea obiectivelor acestei faze s-au efectuat pe populaţii de ovine dindiferite exploataţii zootehnice:
● Ferme de elită şi de înmulţire din staţiunile de cercetare-dezvoltare pentru creşterea ovinelor în funcţie despecificul lor rasial, precum:- Merinos de Palas, Rasa de Carne Palas, Rasa de Lapte Palas de la ICDCOC Palas-Constanţa în condiţiilegeografice din Dobrogea;- Ţigaie de la SCDCOC Secuieni-Bacău, dar şi de la SCDCOC Bilciureşti în condiţiile geografice din centrulMoldovei, respectiv din Dâmboviţa;- Țurcană de la SCDCOC Caransebeş, dar şi de la SCDCOC Popăuţi-Botoşani şi SCDCOC Reghin încondiţiile geografice din Caraş Severin, respectiv nord-estul Moldovei, respectiv centrul Transilvaniei;- Karakul de Botoşani de la SCDCOC Popăuţi Botoşani în condiţiile geografice din nord-estul Moldovei;
● Ferme de producţie din ferma de ovine a SC AGRO IND COM SRL Botoşani sau ferme particulare de tipindividual sau asociativ aflate în observaţia agentului economic pe ecotipuri de Karakul şi Ţurcană, în specialcele afiliate la ”Asociaţia Crescătorilor de Oi Karakul de Botoşani”.
Eşantionarea animalelor a fost făcută în funcţie de cerinţele experimentului pe criterii de vârstă,sex, varietate de culoare, linie zootehnică, tip morfo-productiv, ecotip, areal geografic, specific tehnologic etc.
Investigaţiile de laborator pentru decelarea structurilor genetico-biochimice şi genetico-moleculares-au realizat pe sânge din circulaţia periferică, prelevat prin venipuncţia jugulară a animalelor, pe diferite mediiîn funcţie de natura analizei: pe heparină (pentru determinarea fenotipurilor hemoglobinice şi potasice), peEDTANa2 (pentru determinarea calpastatinei, -lactoglobulinei şi s1-cazeinei) şi fără anticoagulant (pentrudeterminarea fenotipurilor transferinice, albuminice şi amilazice).

Metode de cercetare
Tipizarea genetică a ovinelor s-a realizat prin metode specifice de laborator.
Metode de cercetare a sistemelor genetico-biochimice▪ identificarea fenotipurilor de hemoglobină, transferină, albumină şi amilază –
prin metoda electroforezei cu specificitate pentru fiecărei tip de proteină;▪ decelarea fenotipurilor de potasiu din sânge integral - prin metoda
flamfotometrică;
Metode de cercetare a sistemelor genetico- moleculare▪ genotiparea ovinelor la locii determinanţi ai calpastatinei, .lactoglobulinei şi
s1-cazeinei - prin metoda PCR-RFLP.

Metode de testare şi interpretare statistică a rezultatelor experimentaleIndicatori statistici. Au fost calculate frecvenţele (f) alelice, fenotipice şi genotipice
ale structurilor sistemelor polimorfe:● În cazul sistemelor cu ereditare codominantă (hemoglobinic, transferininic, albuminic,
amilazic, calpastatinic, lactoglobulinic şi cazeinic)▪ frecvenţele alelice s-au calculat după formula:
- n = numărul alelelor de acelaşi fel;- 2N = numărul total de alele din acelaşi sistem.
▪ frecvenţele fenotipurilor şi genotipurilor (identice ca exprimare) s-au calculat după formula:
- n = numărul indivizilor cu acelaşi genotip (fenotip);- N = numărul total de indivizi dintr-o populaţie.
● În cazul sistemelor cu dominanță incompletă (kalemic)▪ se porneşte de la calcularea frecvenţelor fenotipurilor potasice conform formulei:
- n = numărul indivizilor cu acelaşi fenotip potasic- N = numărul total de indivizi dintr-o populaţie.

Testarea stării de echilibru genetic Hardy-Weinberg în populaţii - printestului Hi pătrat (2):
în care:
fi - reprezintă frecvenţa observată (practică, empirică);i – reprezintă frecvenţa aşteptată (teoretică, estimată).
▪ pentru alelele determinante ale fenotipurilor potasiului sanguin se pleacă de la stabilirea frecvenţei genei recesive:
în care:
- q = frecvenţa genei recesive;- Q= frecvenţa fenotipului determinat de alela recesivă.
.
▪ frecvenţa genei dominante (p) se obţine prin diferenţa frecvenţei genei recesive dintr-o unitate p = 1-q
▪ pentru calcularea frecvenţei genotipurilor potasiului sanguin se aplică o distribuţie binomială newtoniană:
(p+q)2 = p2+2pq+p2,
- p2 = frecvenţa homozigoţilor dominanţi;- 2pq = frecvenţa heterozigoţilor;- q2 = frecvenţa homozigoţilor recesivi.
în care:

Rezultate şi Discuţii
A. Caracterizarea genetică a raselor de ovine din România
A.1. Caracterizare genetico-biochimică a raselor de ovine din România
Sistemul hemoglobinic la rasele de ovine din România
Sistemul hemoglobinic la rasele clasice de ovine
Figura 1. Electroforegramă hemoglobinică la ovine

0
10
20
30
40
50
60
70
80
90
%
Merinos de Palas Ţigaie Ţurc ană Karakul de Botos ani
Fe n o tip h e m o g lo b in ic HbA HbA B HbB
Distribuţiile fenotipurilor hemoglobinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
100
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Alela Hb HbA HbB
Frecvenţele alelelor hemoglobinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
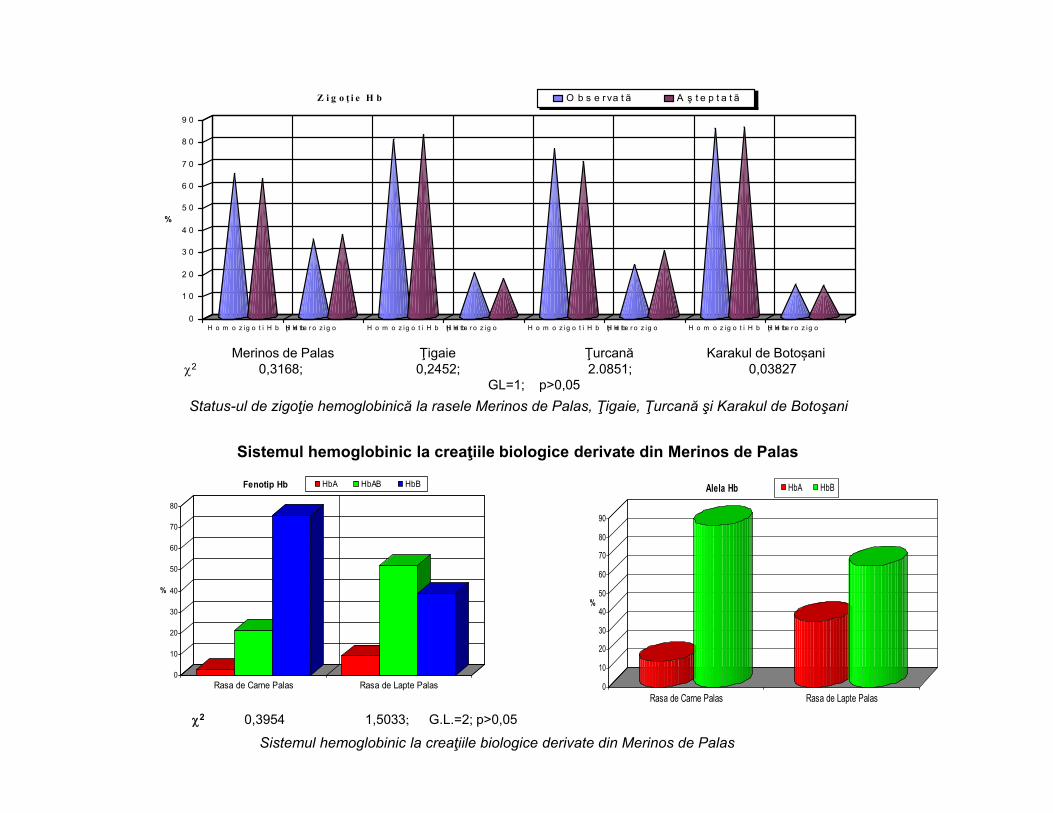
2 0,4962; 0,7388; 4,9418; 0,2893 G.L.=2; p>0,05
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
%
H o m o z ig o t i H b H e t e r o z ig oţi H b H o m o z ig o t i H b H e t e r o z ig oţi H b H o m o z ig o t i H b H e t e r o z ig oţi H b H o m o z ig o t i H b H e t e r o z ig oţi H b
Z i g o ţ i e H b O b s e r va t ã A ş t e p t a t ã
0
10
20
30
40
50
60
70
80
%
Rasa de Carne Palas Rasa de Lapte Palas
Fenotip Hb HbA HbAB HbB
0
10
20
30
40
50
60
70
80
90
%
Rasa de Carne Palas Rasa de Lapte Palas
Alela Hb HbA HbB
Merinos de Palas Ţigaie Ţurcană Karakul de Botoșani2 0,3168; 0,2452; 2.0851; 0,03827
GL=1; p>0,05 Status-ul de zigoţie hemoglobinică la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
Sistemul hemoglobinic la creaţiile biologice derivate din Merinos de Palas2 0,3954 1,5033; G.L.=2; p>0,05
Sistemul hemoglobinic la creaţiile biologice derivate din Merinos de Palas
0
10
20
30
40
50
60
70
80
90
%
Varietatea neagra Varietatea brumarie Varietati colorate
Fenotip hem oglobinic HbA HbAB HbB
0
10203040506070
8090
100
Varietatea neagra Varietatea brumarie Varietati colorate
Alela Hb HbA HbB
0102030405060708090
%
HbAHbA HbAHbB HbBHbB HbAHbA HbAHbB HbBHbB HbAHbA HbAHbB HbBHbB
Genotip Hb Frecventa observata Frecventa asteptata
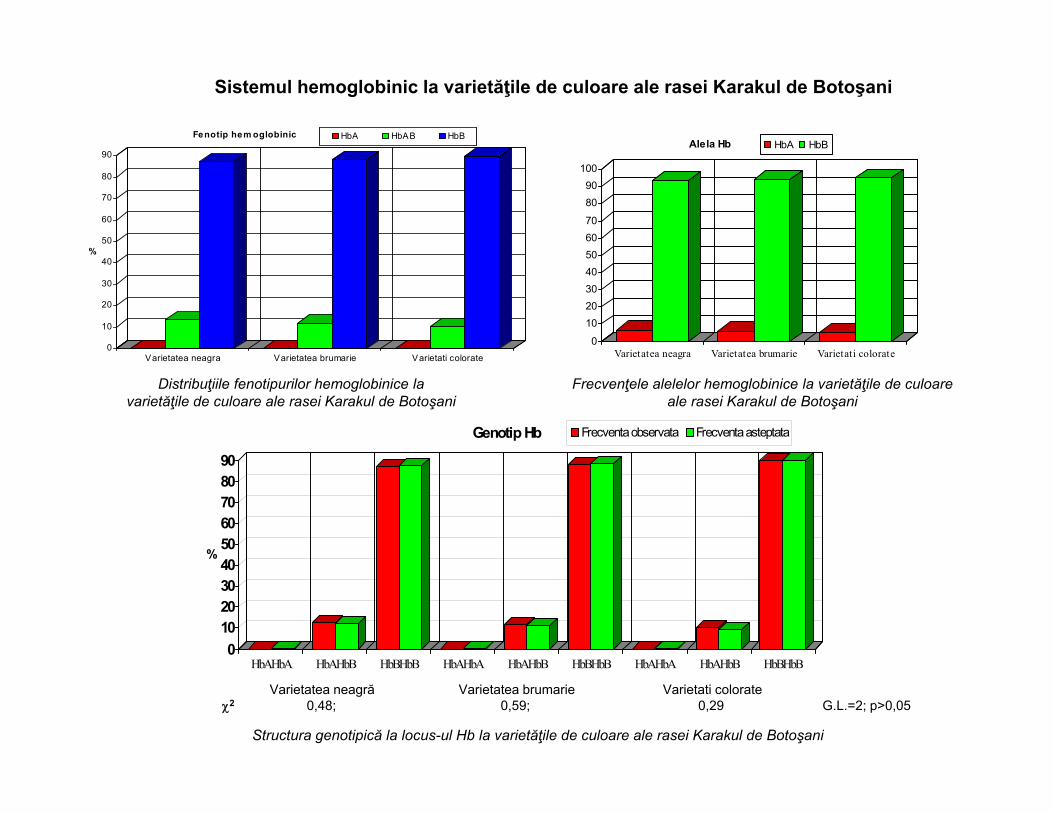
Distribuţiile fenotipurilor hemoglobinice la varietăţile de culoare ale rasei Karakul de Botoşani
Frecvenţele alelelor hemoglobinice la varietăţile de culoare ale rasei Karakul de Botoşani
Varietatea neagră Varietatea brumarie Varietati colorate2 0,48; 0,59; 0,29 G.L.=2; p>0,05
Structura genotipică la locus-ul Hb la varietăţile de culoare ale rasei Karakul de Botoşani
Sistemul hemoglobinic la varietăţile de culoare ale rasei Karakul de Botoşani
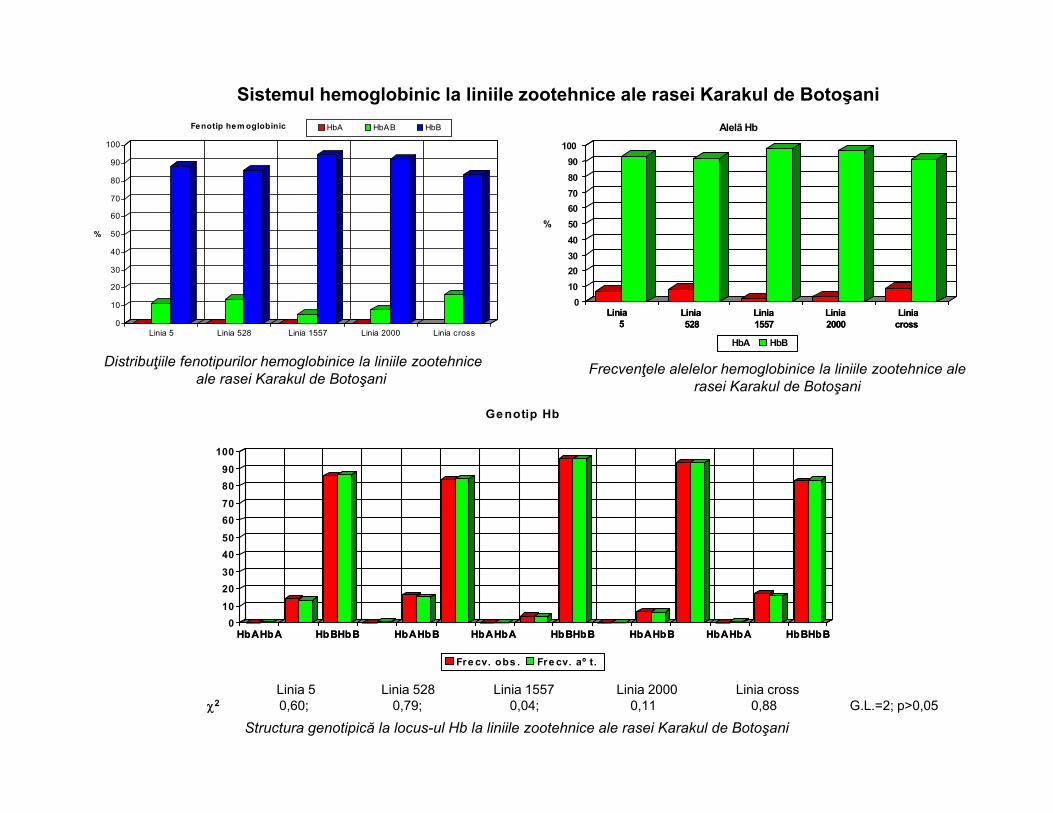
Sistemul hemoglobinic la liniile zootehnice ale rasei Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
100
%
Linia 5 Linia 528 Linia 1557 Linia 2000 Linia cross
Fenotip hem oglobinic HbA HbAB HbB
Linia 5
Linia 528
Linia 1557
Linia 2000
Linia cross
0 10 20 30 40 50 60 70 80 90
100
%
Linia 5
Linia 528
Linia 1557
Linia 2000
Linia cross
Alelã Hb
HbA HbB
HbAHbA HbBHbB HbAHbB HbAHbA HbBHbB HbAHbB HbAHbA HbBHbB0
102030405060708090
100
HbAHbA HbBHbB HbAHbB HbAHbA HbBHbB HbAHbB HbAHbA HbBHbB
Ge notip Hb
Fre cv. obs . Fre cv. aº t.
Distribuţiile fenotipurilor hemoglobinice la liniile zootehnice ale rasei Karakul de Botoşani
Frecvenţele alelelor hemoglobinice la liniile zootehnice ale rasei Karakul de Botoşani
Structura genotipică la locus-ul Hb la liniile zootehnice ale rasei Karakul de Botoşani
Linia 5 Linia 528 Linia 1557 Linia 2000 Linia cross2 0,60; 0,79; 0,04; 0,11 0,88 G.L.=2; p>0,05
0102030405060708090
Merinos(Romania)
Merinos(Germania)
MerinosAustralia)
Merinos(Africa de Sud)
Merinos(Tasmania)
Allele Hb HbA HbB
0102030405060708090
100
Tigaia de campie Tigaia de deal Tigaia premontana Tigaia de munte
Allele Hb HbA HbB
0102030405060708090
100
Turcana decampie
T surcana d edeal ( M o ro ieni)
Tsurcana ded eal ( Pop aut i)
T urcanap remont ana
Turnana demunt e
( A puseni)
Turcana demunt e
( M erid io nali)
Allele Hb HbA HbB
0102030405060708090
100
Ka ra kul(Ira n)
Ka ra kul(Uzbe kista n)
Ka ra kul(Polonia )
Ka ra kul(Unga ria )
Ka ra kul(Roma nia )
Ka ra kul(Ge rma nia )
Allele Hb HbA HbB
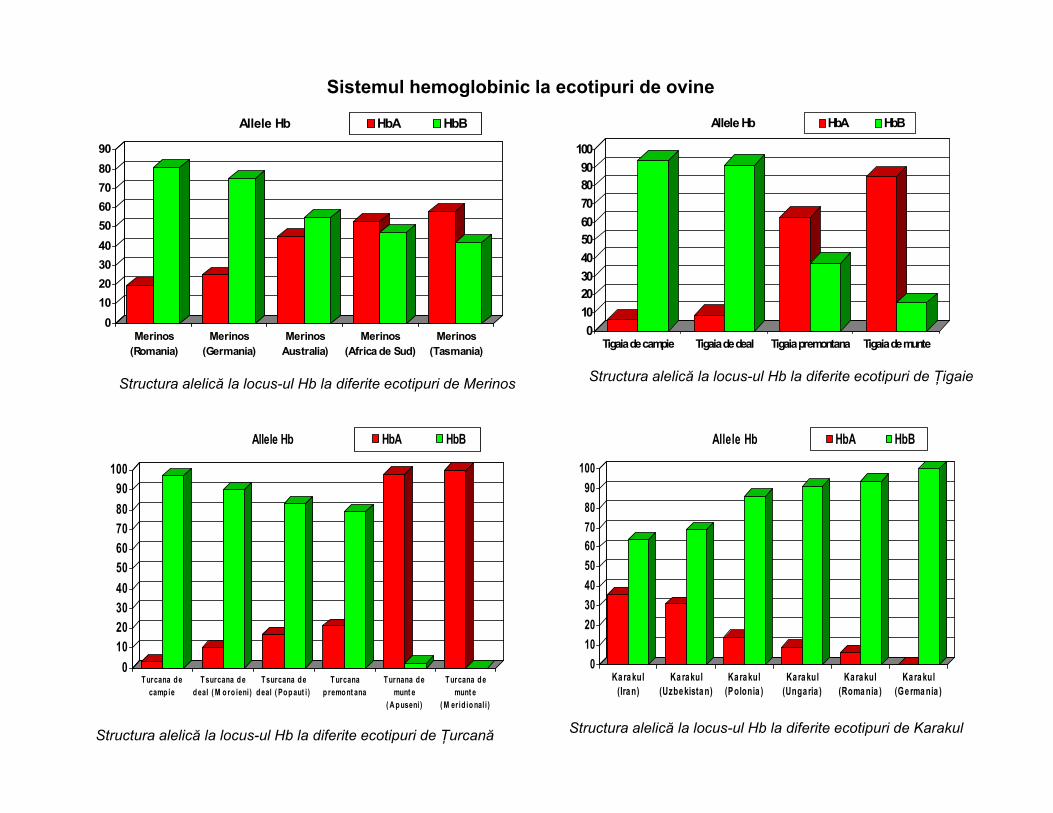
Sistemul hemoglobinic la ecotipuri de ovine
Structura alelică la locus-ul Hb la diferite ecotipuri de Merinos Structura alelică la locus-ul Hb la diferite ecotipuri de Țigaie
Structura alelică la locus-ul Hb la diferite ecotipuri de Țurcană Structura alelică la locus-ul Hb la diferite ecotipuri de Karakul

Sistemul transferinic la rasele de ovine din România
Sistemul transferinic la rasele clasice de ovine
Electroforegramă transferinică la ovine
0
2
4
6
8
10
12
14
16
18
20
TfA TfC TfM TfD TfAB TfAC TfAM TfAD TfAE TfAP TfGC TfBC TfBM TfBD TfCE TfME TfDE TfDP
Fenotip Tf
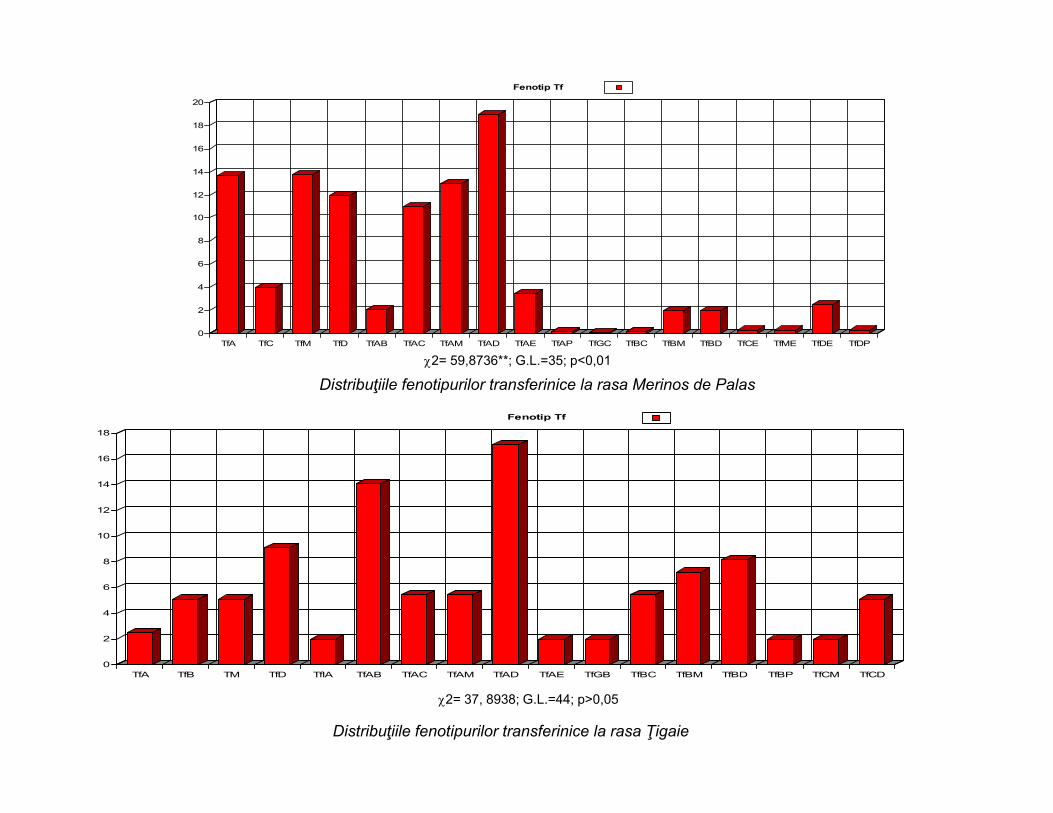
Distribuţiile fenotipurilor transferinice la rasa Merinos de Palas
Distribuţiile fenotipurilor transferinice la rasa Ţigaie
0
2
4
6
8
10
12
14
16
18
TfA TfB TM TfD TfIA TfAB TfAC TfAM TfAD TfAE TfGB TfBC TfBM TfBD TfBP TfCM TfCD
Fenotip Tf
2= 59,8736**; G.L.=35; p<0,01
2= 37, 8938; G.L.=44; p>0,05
0
2
4
6
8
10
12
14
16
18
20
TfA TfB TC TfM TfAB TfAC TfAM TfAD TfAE TfGC TfGM TfBC TfBM TfBD TfBE TfCM TfCD TfCE TfME
Fenotip Tf
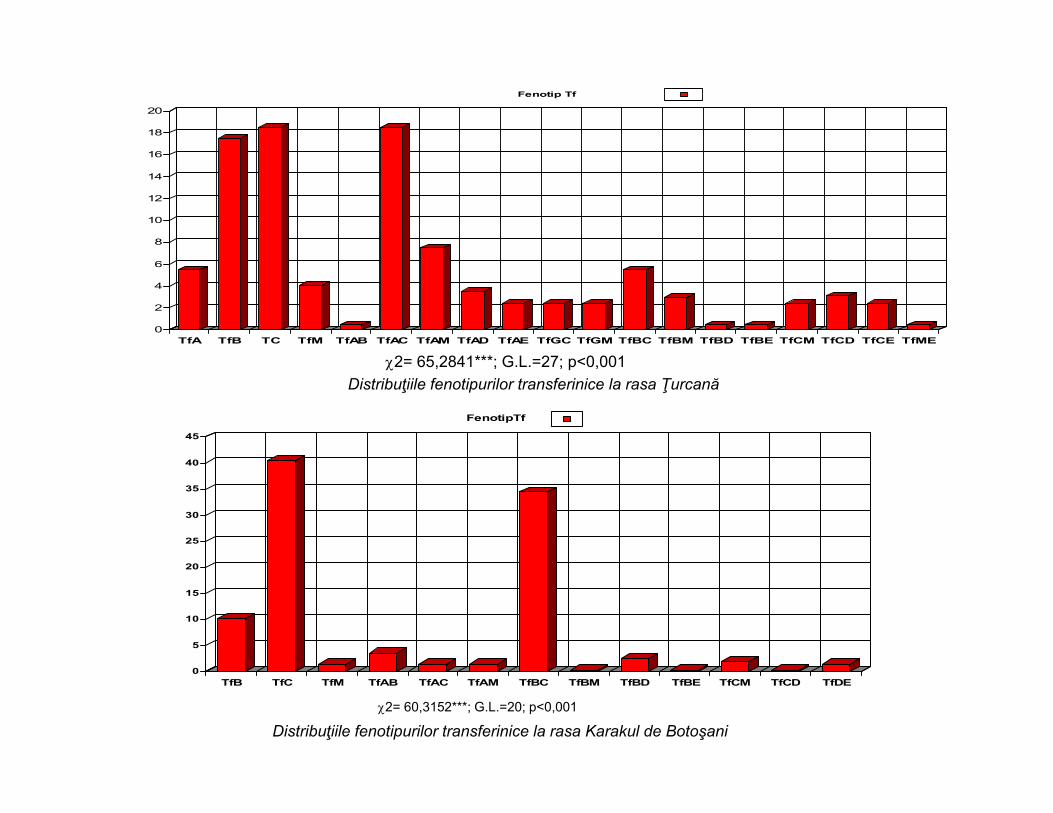
Distribuţiile fenotipurilor transferinice la rasa Ţurcană
0
5
10
15
20
25
30
35
40
45
TfB TfC TfM TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCM TfCD TfDE
FenotipTf
Distribuţiile fenotipurilor transferinice la rasa Karakul de Botoşani
2= 65,2841***; G.L.=27; p<0,001
2= 60,3152***; G.L.=20; p<0,001
0
10
20
30
40
50
60
70
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Alela Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
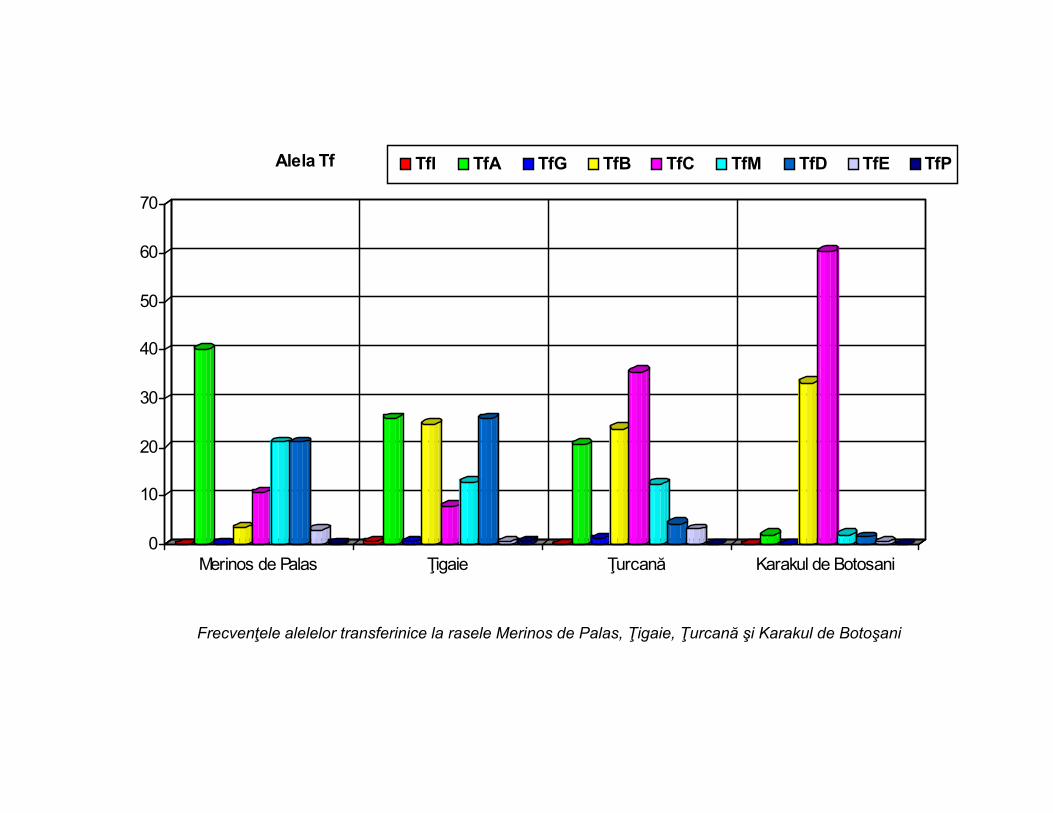
Frecvenţele alelelor transferinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf
Zigoţie Tf Observatã Aşteptatã
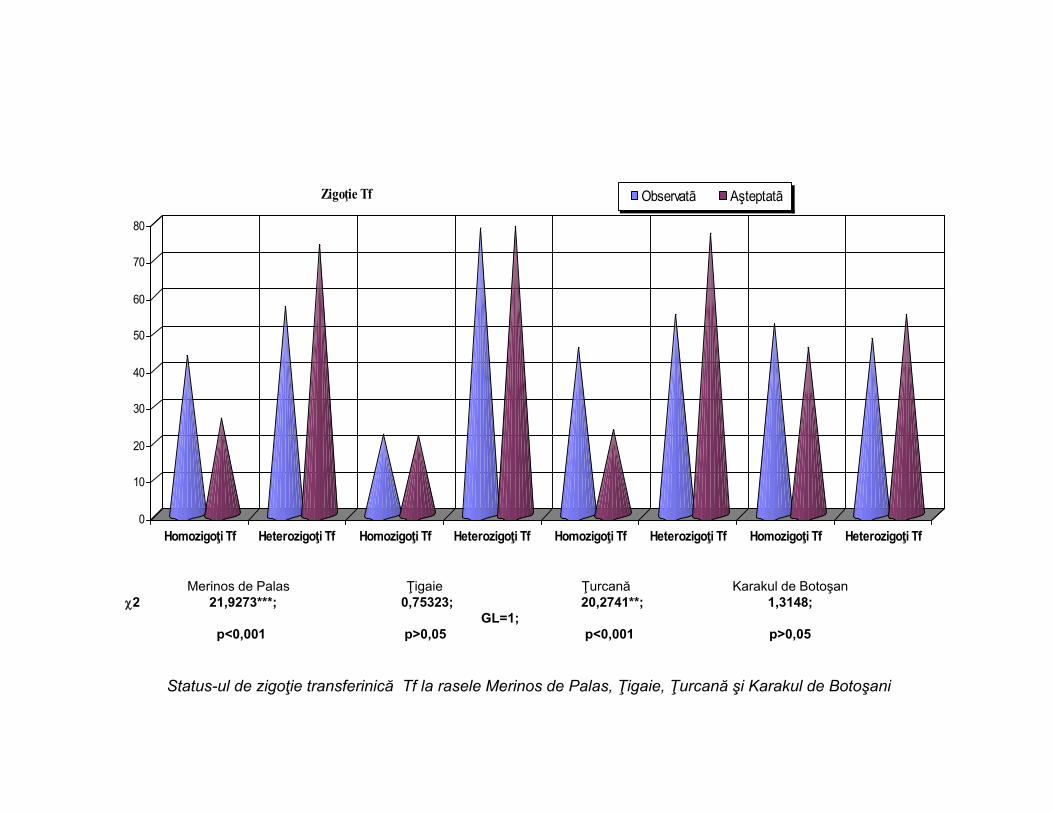
Merinos de Palas Ţigaie Ţurcană Karakul de Botoşan2 21,9273***; 0,75323; 20,2741**; 1,3148;
GL=1;p<0,001 p>0,05 p<0,001 p>0,05
Status-ul de zigoţie transferinică Tf la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
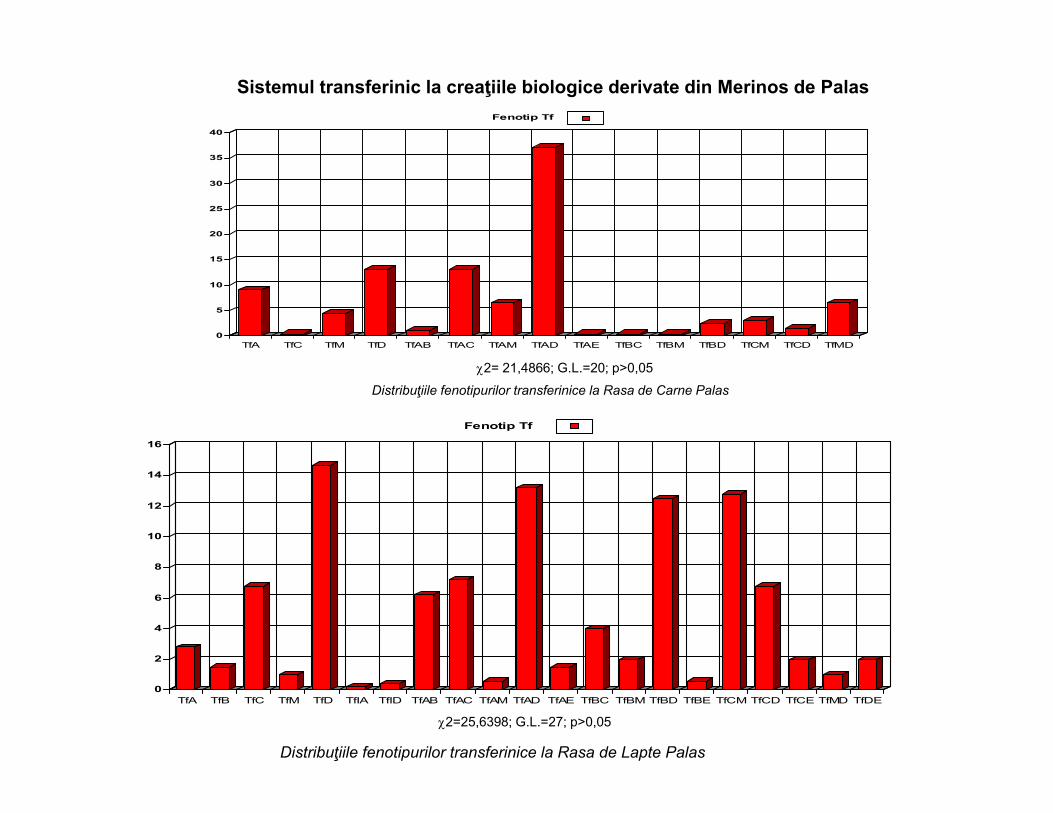
Sistemul transferinic la creaţiile biologice derivate din Merinos de Palas
0
5
10
15
20
25
30
35
40
TfA TfC TfM TfD TfAB TfAC TfAM TfAD TfAE TfBC TfBM TfBD TfCM TfCD TfMD
Fenotip Tf
Distribuţiile fenotipurilor transferinice la Rasa de Carne Palas
Distribuţiile fenotipurilor transferinice la Rasa de Lapte Palas
2= 21,4866; G.L.=20; p>0,05
2=25,6398; G.L.=27; p>0,05
0
2
4
6
8
10
12
14
16
TfA TfB TfC TfM TfD TfIA TfID TfAB TfAC TfAM TfAD TfAE TfBC TfBM TfBD TfBE TfCM TfCD TfCE TfMD TfDE
Fenotip Tf
0
5
10
15
20
25
30
35
40
Rasa de Carne Palas Rasa de Lapte Palas
Alela Tf TfI TfA TfB TfC TfM TfD TfE
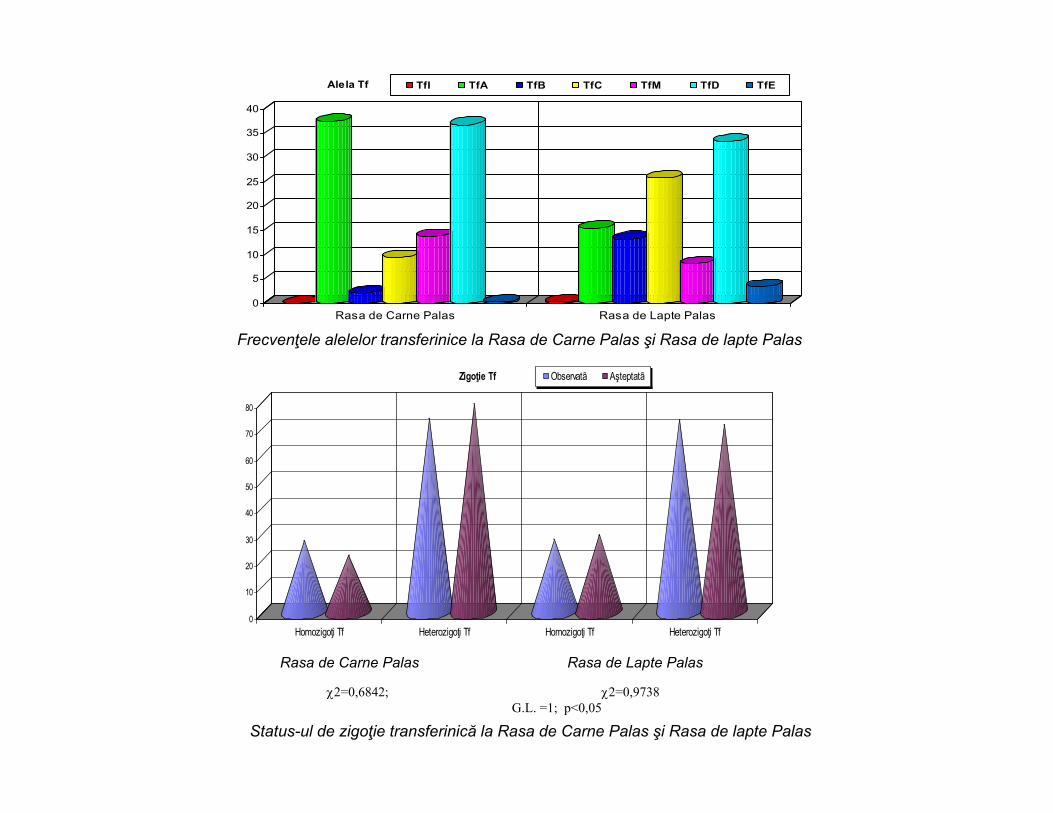
Frecvenţele alelelor transferinice la Rasa de Carne Palas şi Rasa de lapte Palas
0
10
20
30
40
50
60
70
80
Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf
Zigoţie Tf Observatã Aşteptatã
Rasa de Carne Palas Rasa de Lapte Palas
2=0,6842; 2=0,9738G.L. =1; p<0,05
Status-ul de zigoţie transferinică la Rasa de Carne Palas şi Rasa de lapte Palas
0
5
10
15
20
25
30
%
TfB TfC TfM TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCM TfCD TfDE
Fenotip Tf • Varietatea de culoare : Neagrã
0
5
10
15
20
25
30
%
TfB TfC TfM TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCD TfDE
Genotip Tf • Varietatea de culoare : Brumãrie
0
5
10
15
20
25
%
TfB TfC TfM TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCM TfCD TfDE
Genotip Tf • Varietatea de culoare : Coloratã
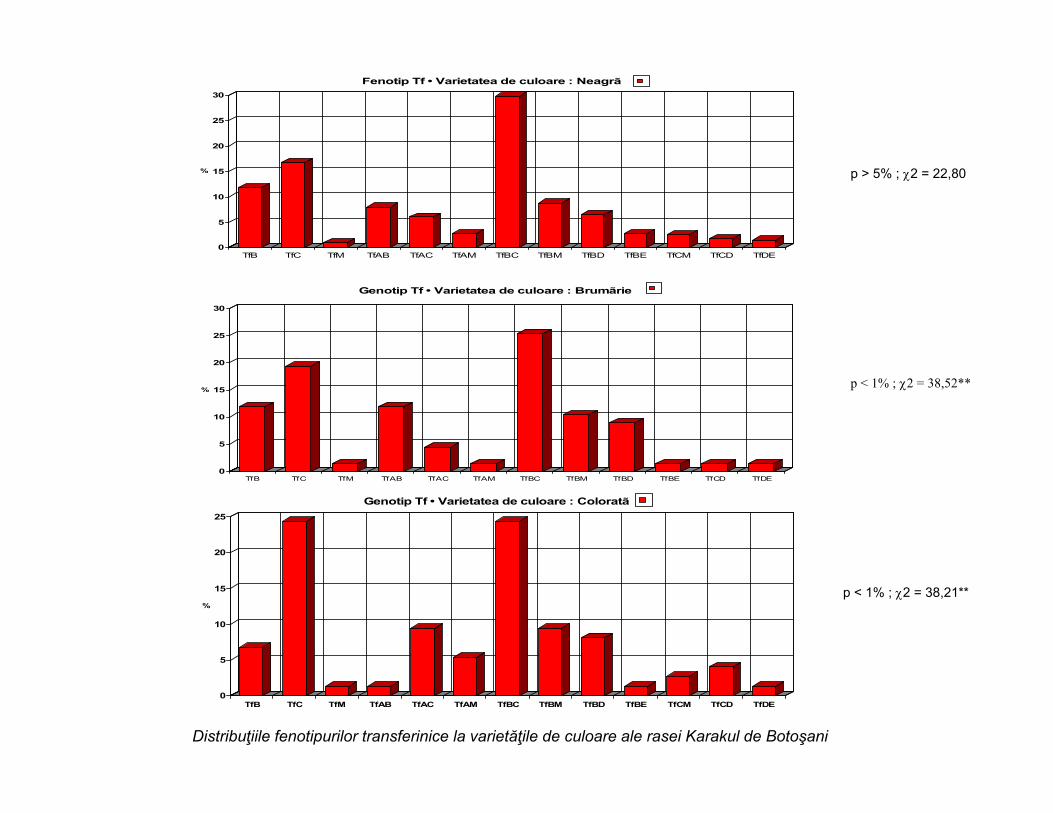
Distribuţiile fenotipurilor transferinice la varietăţile de culoare ale rasei Karakul de Botoşani
p > 5% ; 2 = 22,80
p < 1% ; 2 = 38,52**
p < 1% ; 2 = 38,21**
01020304050607080
Frecv. obs. Frecv. astept.
Varietatea de culoare : Neagrã
Homozigoþi Tf Heterozigoþi Tf
Frecv. obs. Frecv. astept.0
10
20
30
40
50
60
70
Frecv. obs. Frecv. astept.
Varietatea de culoare :Brumãrie
Homozigoþi T f Heterozigoþi T f
Frecv. obs. Frecv. astept.0
10
20
30
40
50
60
70
Frecv. obs. Frecv. astept.
Varietatea de culoare :Coloratã
Homozigoþi T f Heterozigoþi T f
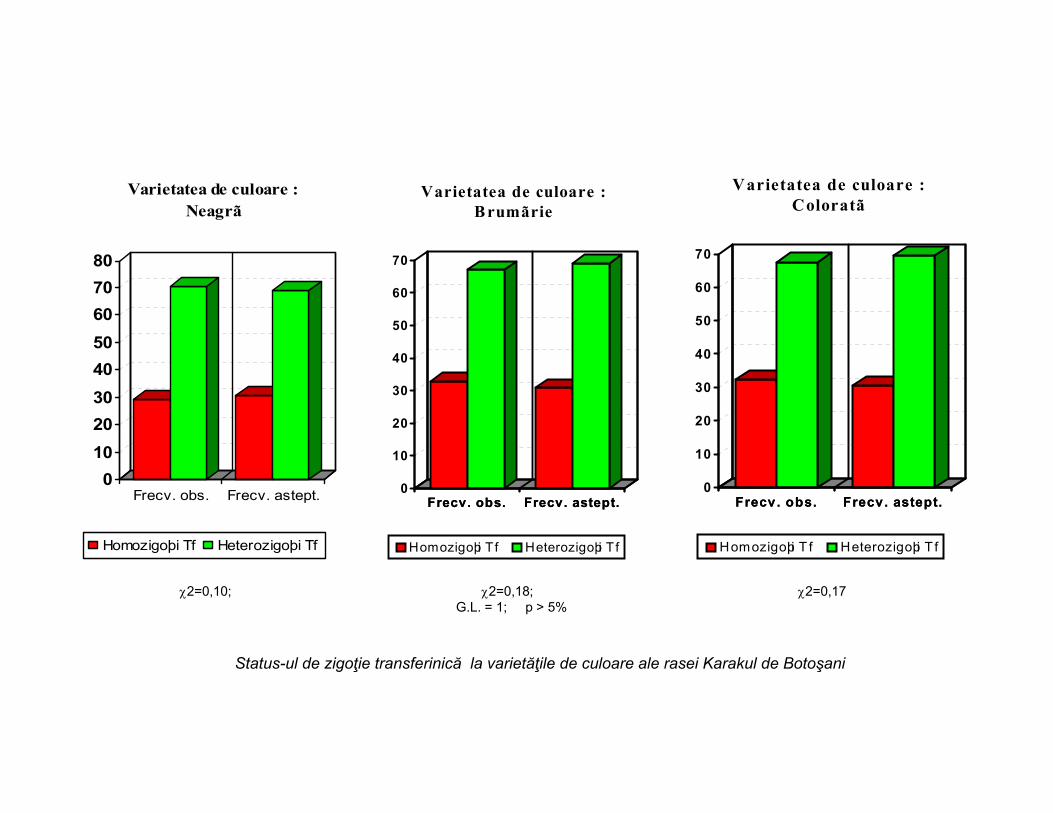
2=0,10; 2=0,18; 2=0,17G.L. = 1; p > 5%
Status-ul de zigoţie transferinică la varietăţile de culoare ale rasei Karakul de Botoşani

Sistemul transferinic la liniile zootehnice ale rasei Karakul de Botoşani
0
5
10
15
20
25
30
35
%
TfB TfC TfM TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCM TfCD TfDE
Linia 5 Fenotip Tf
0
5
10
15
20
25
%
TfB TfC TfAB TfAC TfAM TfBC TfBM TfBD TfBE TfCM TfCD TfDE
Linia 528 Fenotip Tf
0
5
10
15
20
25
30
35
40
%
TfB TfC TfA B TfAC TfBC TfBM TfBD TfBE TfCM
Linia 1557 Fe notip T f
0
5
10
15
20
25
30
35
40
%
TfB TfC TfA B TfA C TfA M TfB C TfB M TfB D TfCM TfDE
Linia 2000 Fe notip T f
0
5
1 0
1 5
2 0
2 5
%
TfB TfC TfM TfA B TfA C TfA M TfB C TfB M TfB D TfB E TfC M TfC D TfD E
L in ia c r o s s
Distribuţiile fenotipurilor transferinice la liniile zootehnice ale rasei Karakul de Botoşani
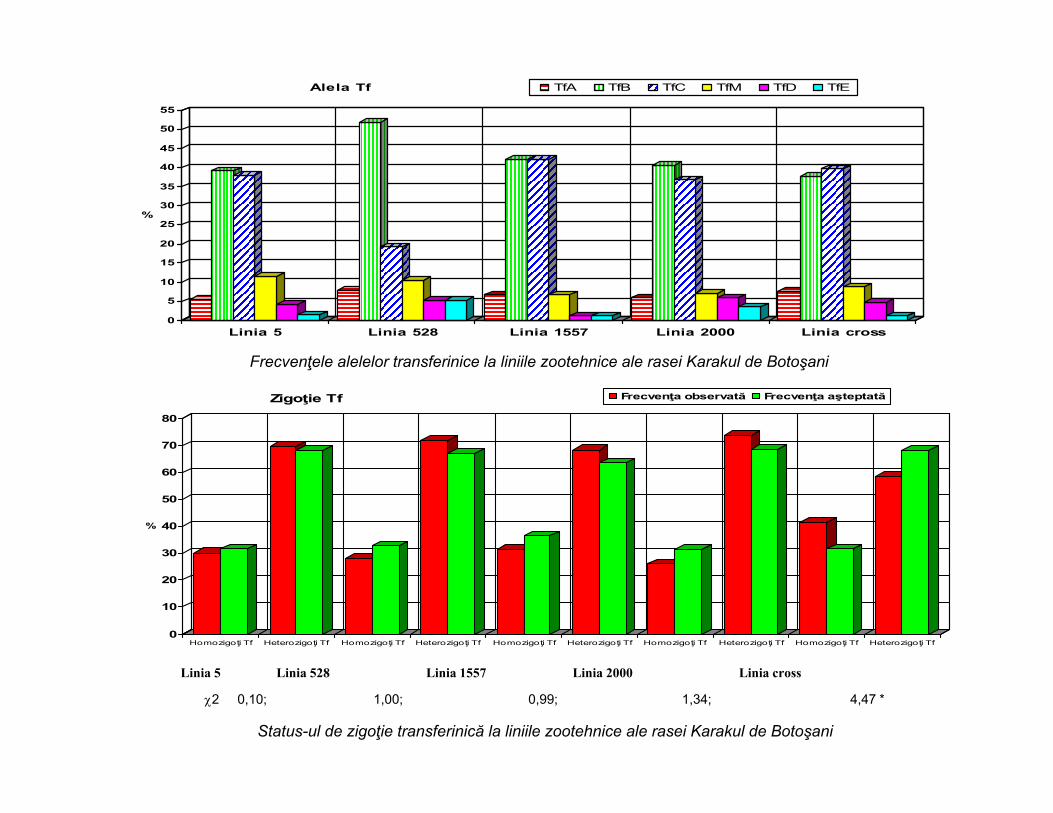
0
5
10
15
20
25
30
35
40
45
50
55
%
Linia 5 Linia 528 Linia 1557 Linia 2000 Linia cross
Alela Tf TfA TfB TfC TfM TfD TfE
Frecvenţele alelelor transferinice la liniile zootehnice ale rasei Karakul de Botoşani
0
10
20
30
40
50
60
70
80
%
Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf Homozigoţi Tf Heterozigoţi Tf
Zigoţie Tf Frecvenţa observată Frecvenţa aşteptată
2 0,10; 1,00; 0,99; 1,34; 4,47 *
Linia 5 Linia 528 Linia 1557 Linia 2000 Linia cross
Status-ul de zigoţie transferinică la liniile zootehnice ale rasei Karakul de Botoşani
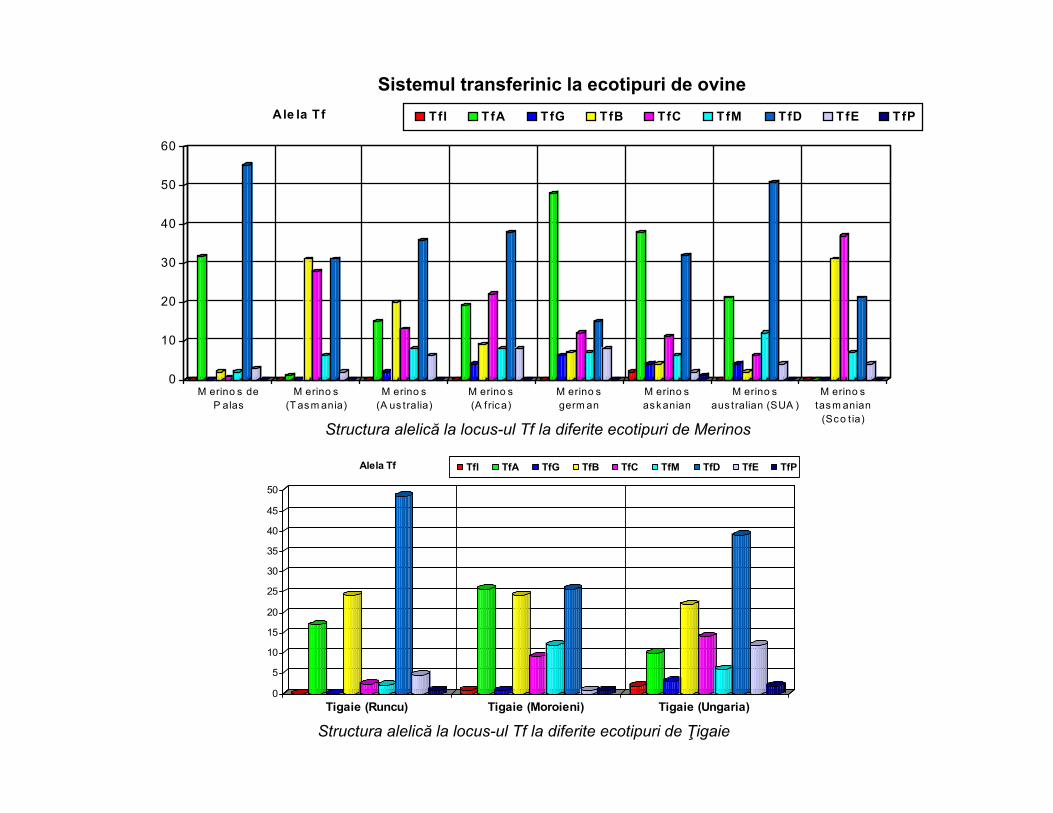
0
10
20
30
40
50
60
M erino s deP alas
M erino s(T asm ania)
M erino s(A us tralia)
M erino s(A frica)
M erino sgerm an
M erino saskanian
M erino saus t ralian (SUA )
M erino stasm anian
(Sco t ia)
Ale la Tf TfI T fA TfG TfB TfC TfM TfD TfE TfP
0
5
10
15
20
25
30
35
40
45
50
Tigaie (Runcu) Tigaie (Moroieni) Tigaie (Ungaria)
Alela Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
Sistemul transferinic la ecotipuri de ovine
Structura alelică la locus-ul Tf la diferite ecotipuri de Merinos
Structura alelică la locus-ul Tf la diferite ecotipuri de Ţigaie
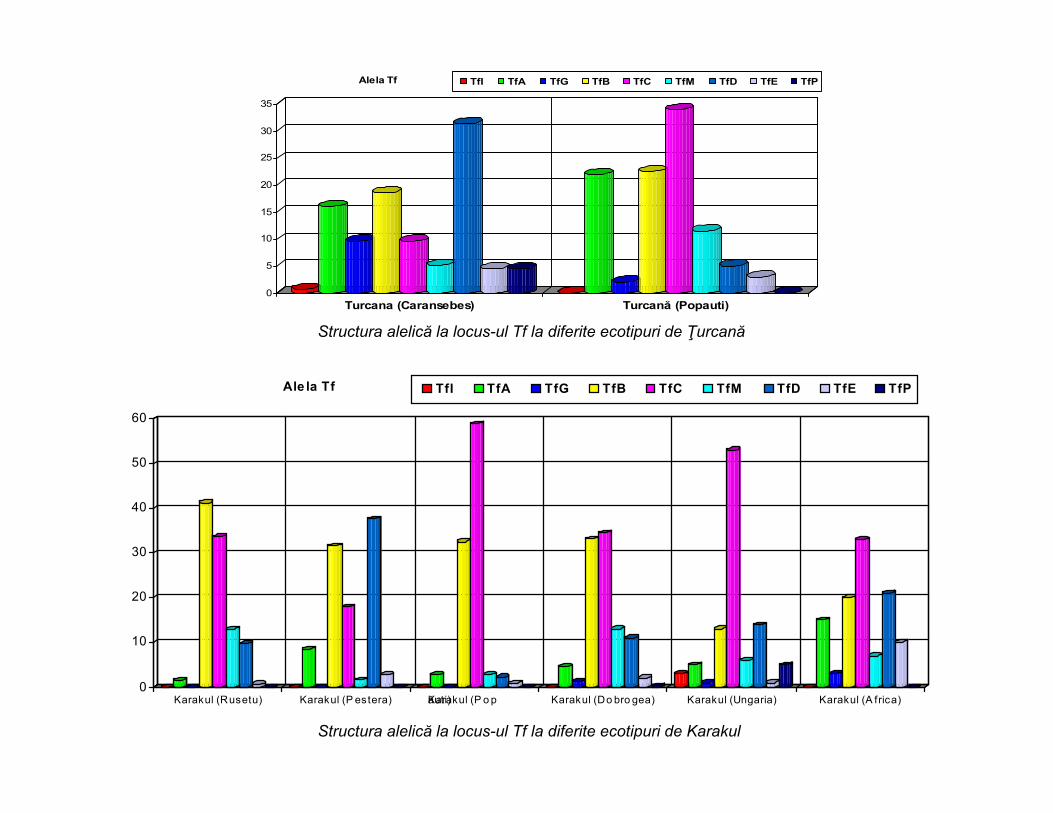
0
5
10
15
20
25
30
35
Turcana (Caransebes) Turcană (Popauti)
Alela Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
0
10
20
30
40
50
60
Karakul (Rusetu) Karakul (P estera) Karakul (P o păuti) Karakul (Do bro gea) Karakul (Ungaria) Karakul (A frica)
Ale la Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
Structura alelică la locus-ul Tf la diferite ecotipuri de Ţurcană
Structura alelică la locus-ul Tf la diferite ecotipuri de Karakul
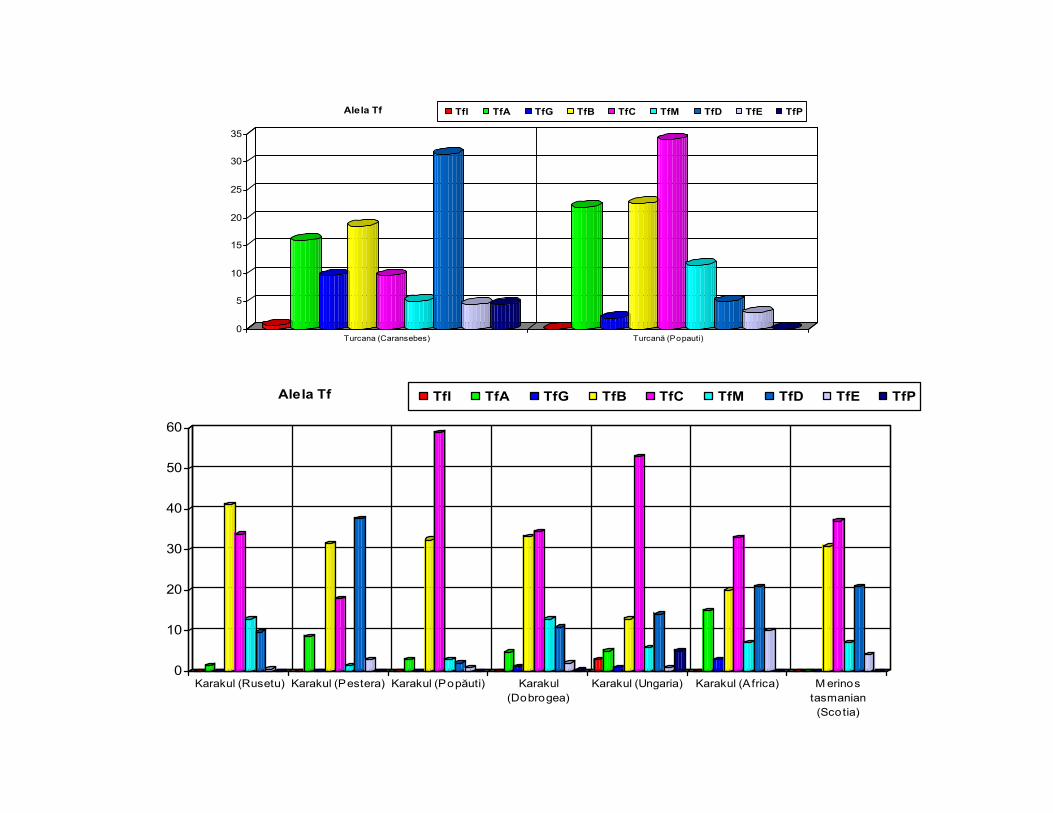
0
5
10
15
20
25
30
35
Turcana (Caransebes) Turcană (Popauti)
Alela Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
0
10
20
30
40
50
60
Karakul (Rusetu) Karakul (Pestera) Karakul (Popăuti) Karakul(Dobrogea)
Karakul (Ungaria) Karakul (Africa) M erinostasmanian(Scotia)
Alela Tf TfI TfA TfG TfB TfC TfM TfD TfE TfP
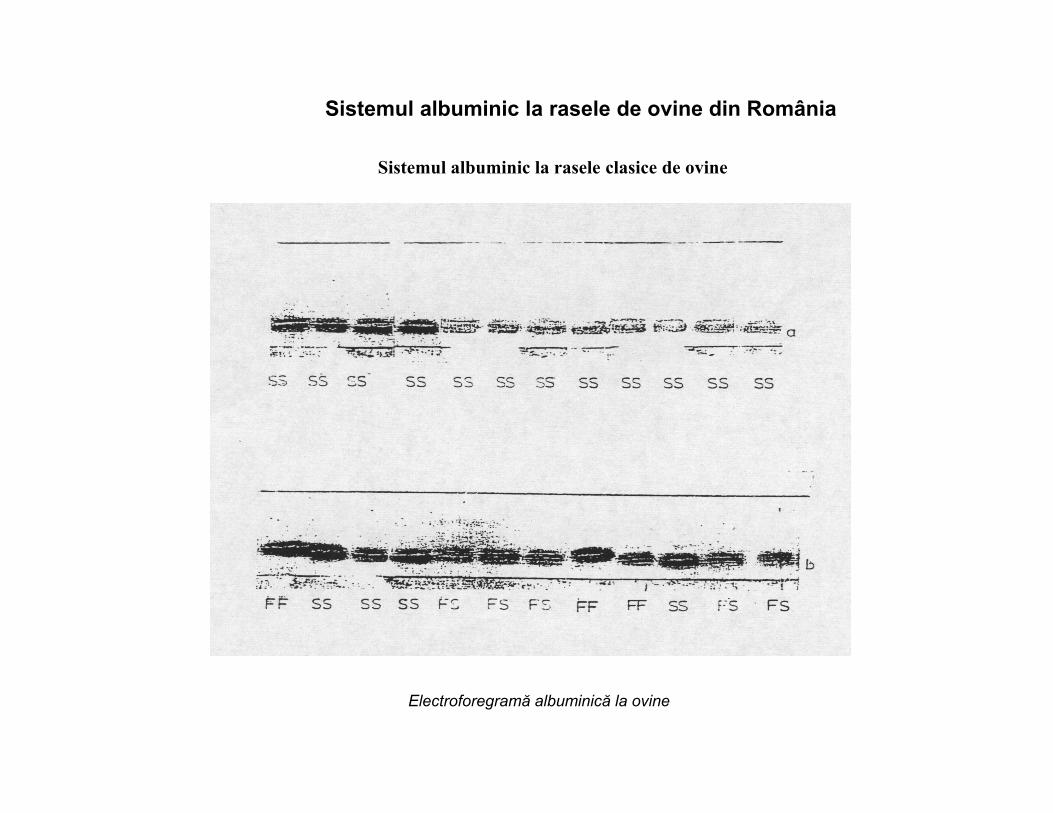
Sistemul albuminic la rasele de ovine din România
Sistemul albuminic la rasele clasice de ovine
Electroforegramă albuminică la ovine
0
10
20
30
40
50
60
70
80
90
100
Mer inos de Palas Ţiga ie Ţurc ană Karaku l de Botos an i
A lbF A lbF A lbS
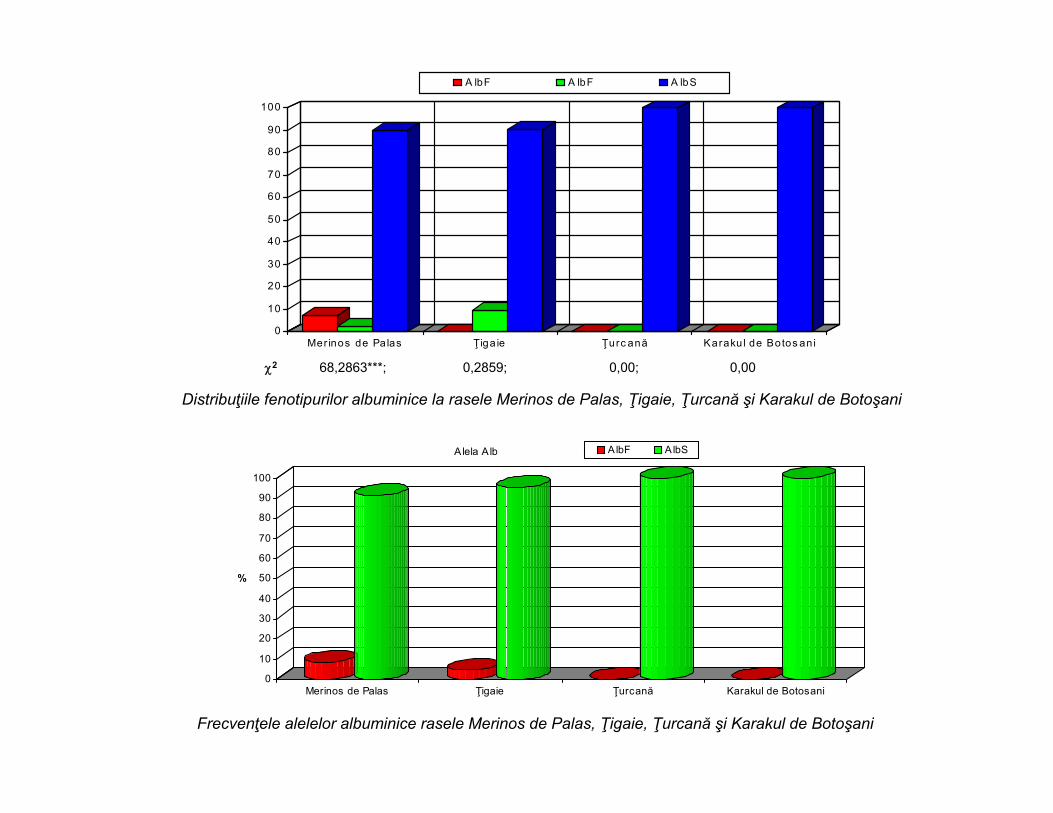
Distribuţiile fenotipurilor albuminice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
100
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
A lela A lb AlbF AlbS
Frecvenţele alelelor albuminice rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
2 68,2863***; 0,2859; 0,00; 0,00
0
10
20
30
40
50
60
70
80
90
100
Homozigoţi Alb Heterozigoţi Alb Homozigoţi Alb Heterozigoţi Alb Homozigoţi Alb Heterozigoţi Alb Homozigoţi Alb Heterozigoţi Alb
Zigoţie Alb Observatã Aşteptatã
Merinos de Palas Ţigaie Ţurcană Karakul de Botoşani
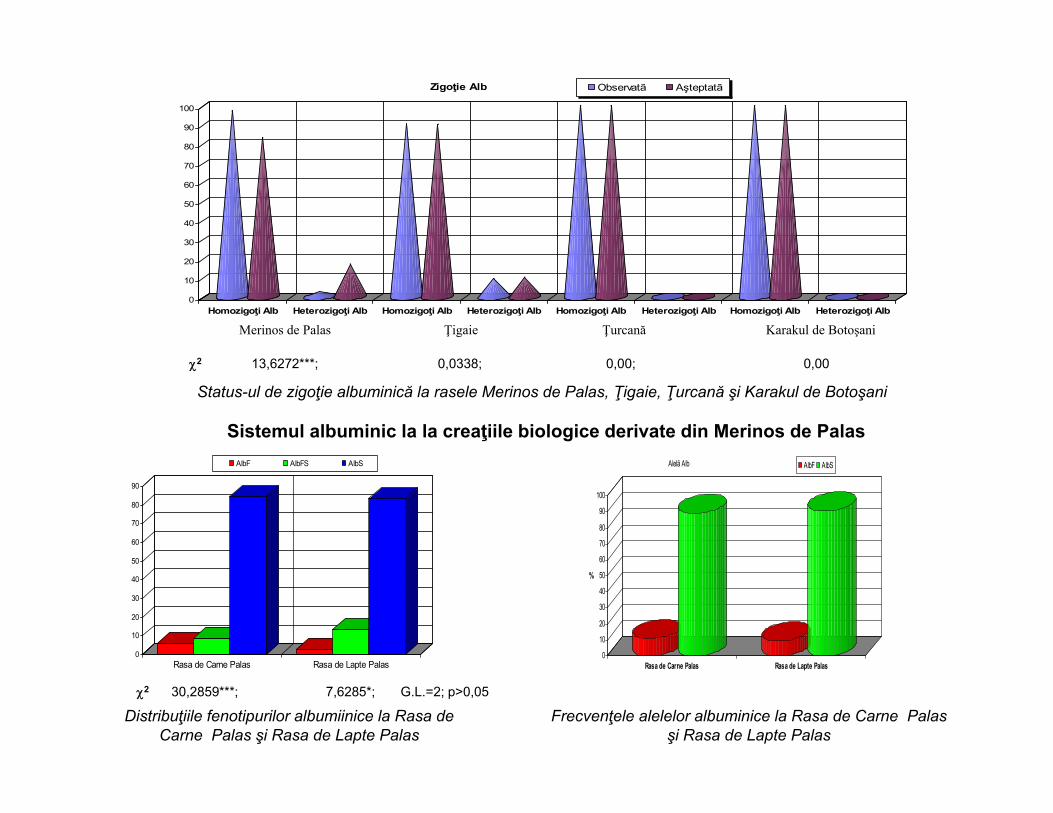
2 13,6272***; 0,0338; 0,00; 0,00
Status-ul de zigoţie albuminică la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
Sistemul albuminic la la creaţiile biologice derivate din Merinos de Palas
0
10
20
30
40
50
60
70
80
90
Rasa de Carne Palas Rasa de Lapte Palas
AlbF AlbFS AlbS
0
10
20
30
40
50
60
70
80
90
100
%
Rasa de Carne Palas Rasa de Lapte Palas
Alelã Alb AlbF AlbS
Distribuţiile fenotipurilor albumiinice la Rasa de Carne Palas şi Rasa de Lapte Palas
Frecvenţele alelelor albuminice la Rasa de Carne Palas şi Rasa de Lapte Palas
2 30,2859***; 7,6285*; G.L.=2; p>0,05
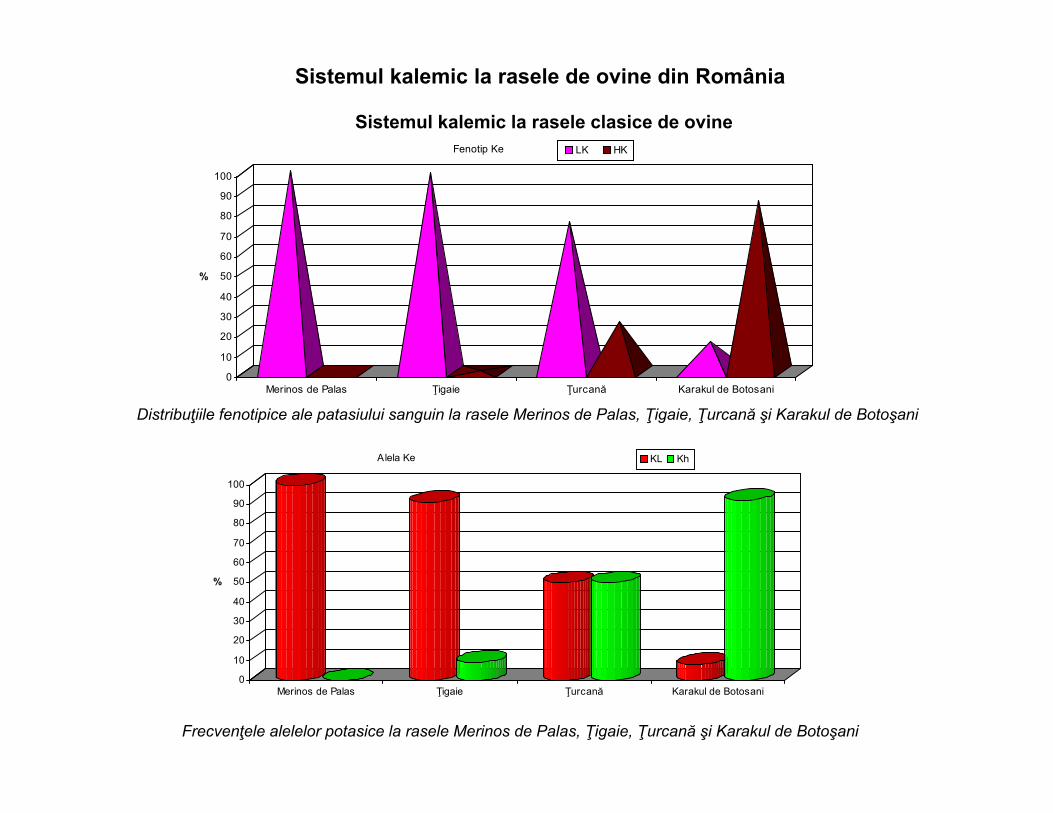
Sistemul kalemic la rasele de ovine din România
Sistemul kalemic la rasele clasice de ovine
0
10
20
30
40
50
60
70
80
90
100
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Fenotip Ke LK HK
0
10
20
30
40
50
60
70
80
90
100
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Alela Ke KL Kh
Distribuţiile fenotipice ale patasiului sanguin la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
Frecvenţele alelelor potasice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
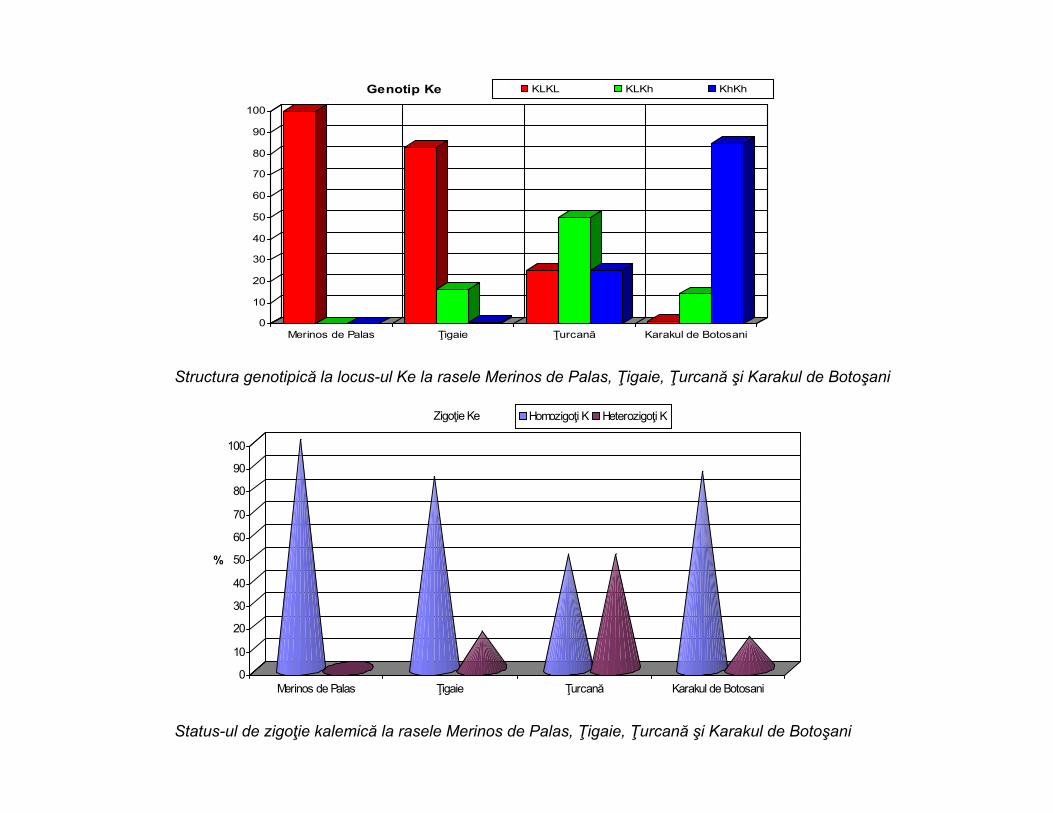
0
10
20
30
40
50
60
70
80
90
100
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Genotip Ke KLKL KLKh KhKh
Structura genotipică la locus-ul Ke la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
100
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Zigoţie Ke Homozigoţi K Heterozigoţi K
Status-ul de zigoţie kalemică la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
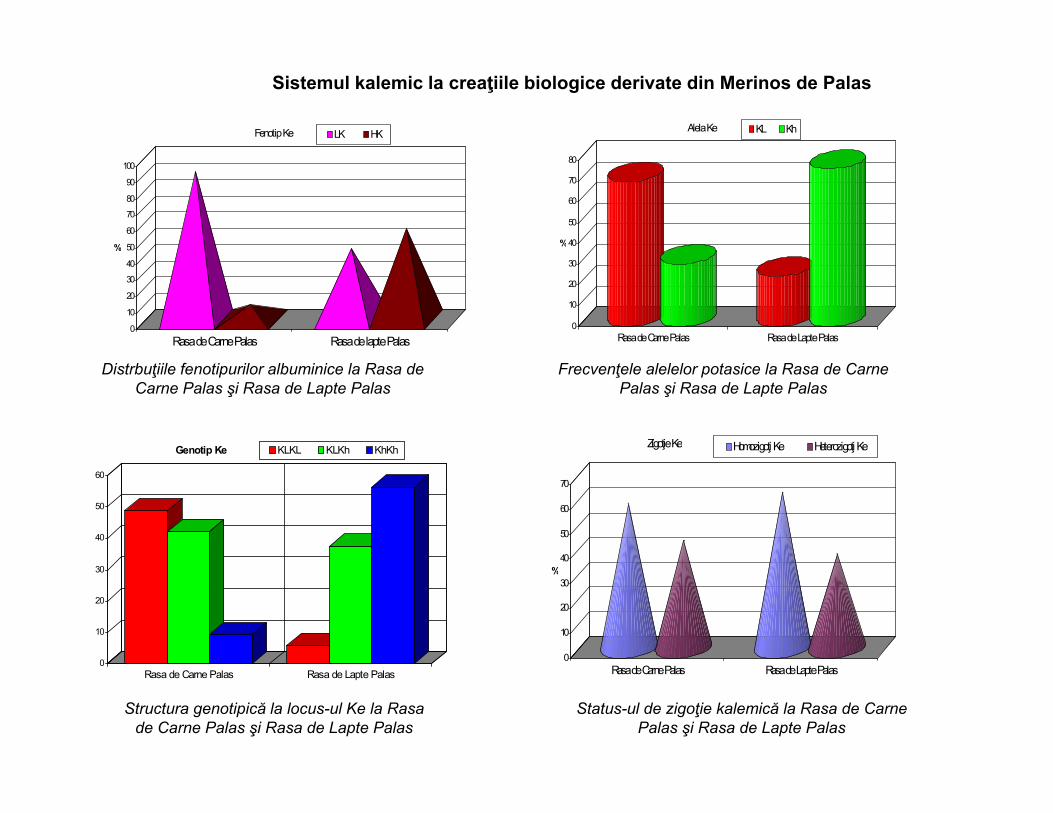
Sistemul kalemic la creaţiile biologice derivate din Merinos de Palas
0
10
20
30
40
50
60
70
80
90
100
%
Rasa de Carne Palas Rasa de lapte Palas
Fenotip Ke LK HK
0
10
20
30
40
50
60
70
80
%
Rasa de Carne Palas Rasa de Lapte Palas
Alela Ke KL Kh
Distrbuţiile fenotipurilor albuminice la Rasa de Carne Palas şi Rasa de Lapte Palas
Frecvenţele alelelor potasice la Rasa de Carne Palas şi Rasa de Lapte Palas
0
10
20
30
40
50
60
Rasa de Carne Palas Rasa de Lapte Palas
Genotip Ke KLKL KLKh KhKh
0
10
20
30
40
50
60
70
%
Rasa de Carne Palas Rasa de Lapte Palas
Zigoţie Ke Homozigoţi Ke Heterozigoţi Ke
Structura genotipică la locus-ul Ke la Rasa de Carne Palas şi Rasa de Lapte Palas
Status-ul de zigoţie kalemică la Rasa de Carne Palas şi Rasa de Lapte Palas
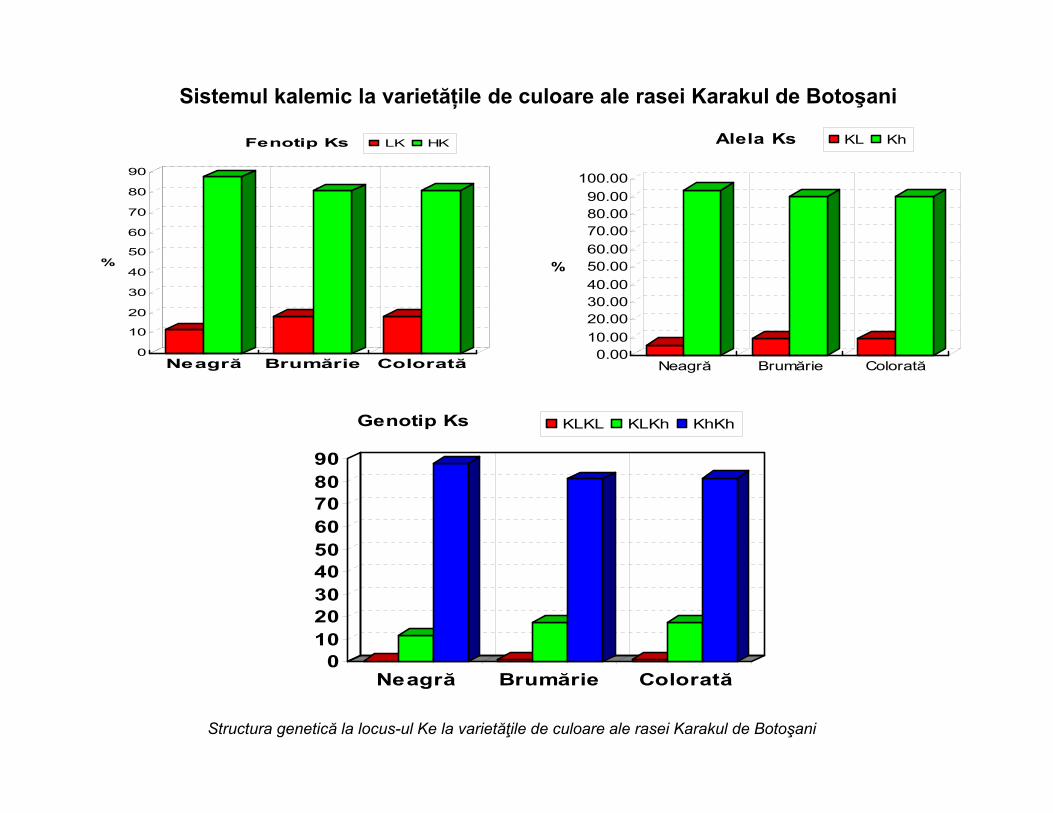
Sistemul kalemic la varietățile de culoare ale rasei Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
%
Neagră Brumărie Colorată
Fenotip Ks LK HK
0.0010.0020.0030.0040.0050.0060.0070.0080.0090.00
100.00
%
Neagră Brumărie Colorată
Alela Ks KL Kh
0102030405060708090
Neagră Brumărie Colorată
Genotip Ks KLKL KLKh KhKh
Structura genetică la locus-ul Ke la varietăţile de culoare ale rasei Karakul de Botoşani
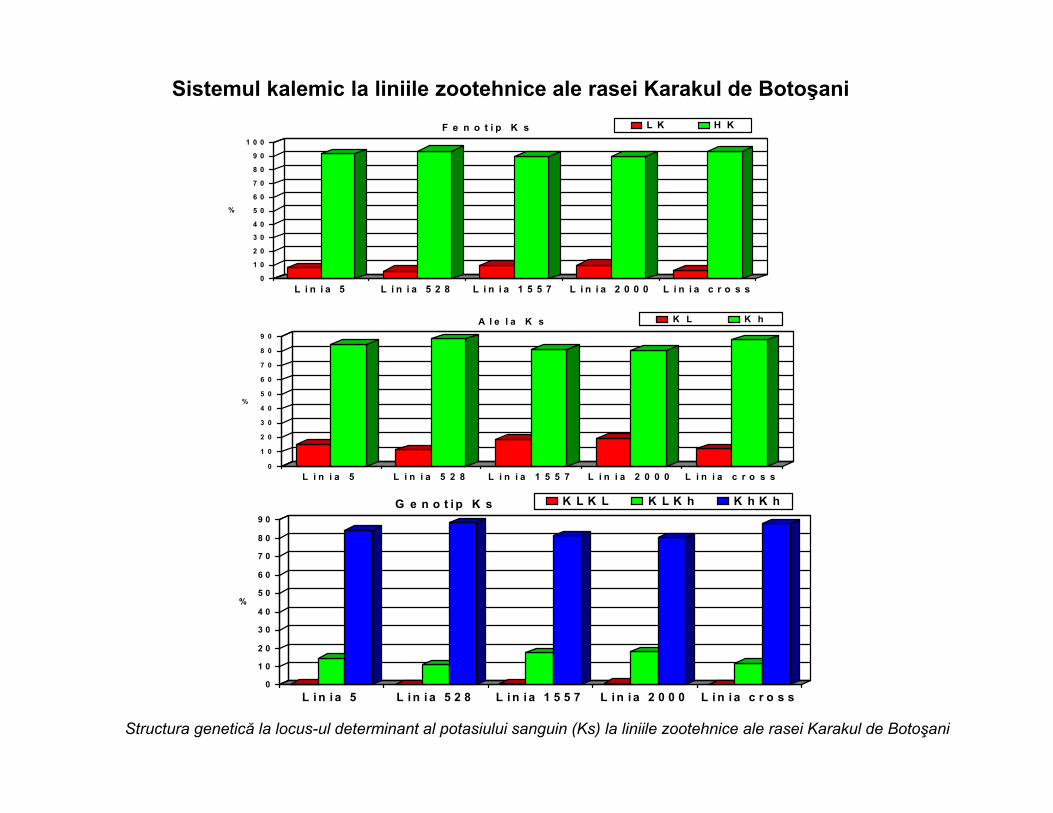
Sistemul kalemic la liniile zootehnice ale rasei Karakul de Botoşani
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
1 0 0
%
L i n i a 5 L i n i a 5 2 8 L i n i a 1 5 5 7 L i n i a 2 0 0 0 L i n i a c r o s s
F e n o t i p K s L K H K
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
%
L i n i a 5 L i n i a 5 2 8 L i n i a 1 5 5 7 L i n i a 2 0 0 0 L i n i a c r o s s
A l e l a K s K L K h
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
%
L i n i a 5 L i n i a 5 2 8 L i n i a 1 5 5 7 L i n i a 2 0 0 0 L i n i a c r o s s
G e n o t i p K s K L K L K L K h K h K h
Structura genetică la locus-ul determinant al potasiului sanguin (Ks) la liniile zootehnice ale rasei Karakul de Botoşani
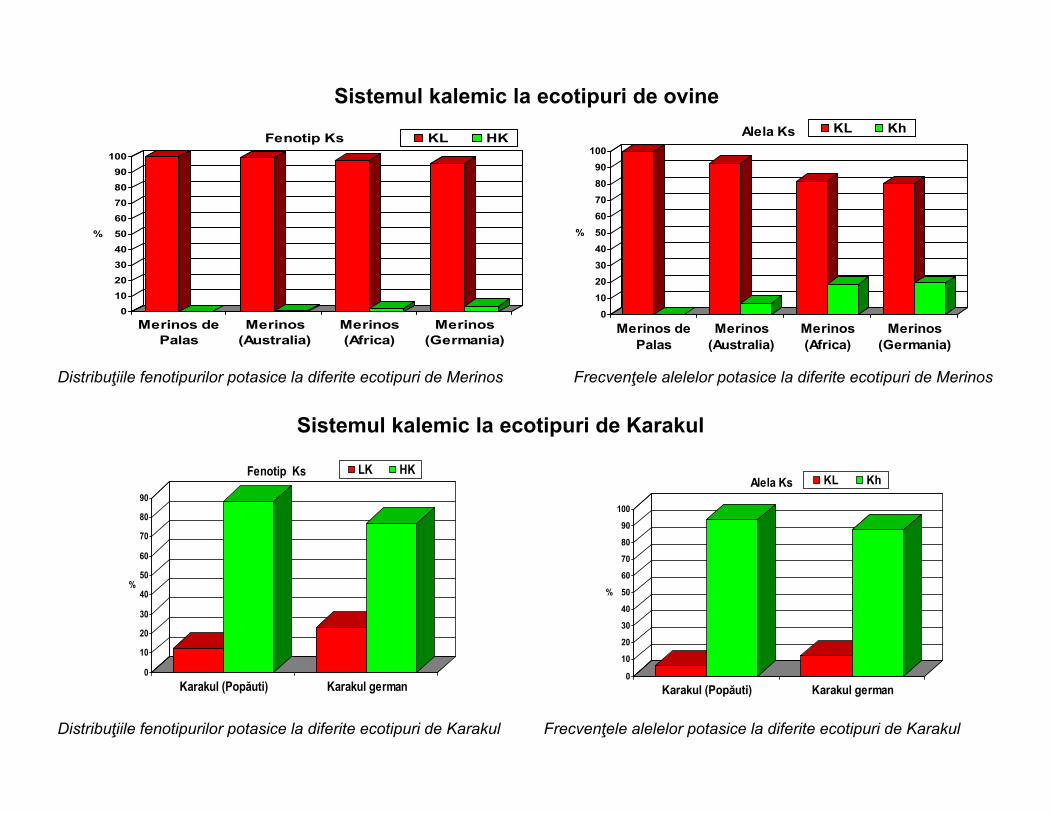
Sistemul kalemic la ecotipuri de ovine
0102030405060708090
100
%
Merinos dePalas
Merinos(Australia)
Merinos(Africa)
Merinos(Germania)
Fenotip Ks KL HK
0102030405060708090
100
%
Merinos dePalas
Merinos(Australia)
Merinos(Africa)
Merinos(Germania)
Alela Ks KL Kh
Distribuţiile fenotipurilor potasice la diferite ecotipuri de Merinos Frecvenţele alelelor potasice la diferite ecotipuri de Merinos
Sistemul kalemic la ecotipuri de Karakul
0
10
20
30
40
50
60
70
80
90
%
Karakul (Popăuti) Karakul german
Fenotip Ks LK HK
0
10
20
30
40
50
60
70
80
90
100
%
Karakul (Popăuti) Karakul german
Alela Ks KL Kh
Distribuţiile fenotipurilor potasice la diferite ecotipuri de Karakul Frecvenţele alelelor potasice la diferite ecotipuri de Karakul

0
1020
304050
607080
90100
%
Merinos de Palas Tsigai Tsurcana Karakul de Botosani
Alela Am AmB AmC
Sistemul amilazic la rasele de ovine din România
Frecvenţele alelelor amilazice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
100
%
M e rin o s d e P a la s T ig a ie T u rc a n a K a ra k u l d e B o t o s a n i
Ge notip Am A mBA mB A mBA mC A mCA mC
2 0,3168; 0,2452; 2.0851; 0,03827GL=1; p>0,05
Structura genotipică la locus-ul Am la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
A2. Caracterizare genetico-moleculară a raselor de ovine din România
Sistemul calpastatinic la rasele de ovine din România
Sistemul calpastatinic la rasele clasice de ovine
Electroforegramă cu fenotipuri calpastatinice la ovine
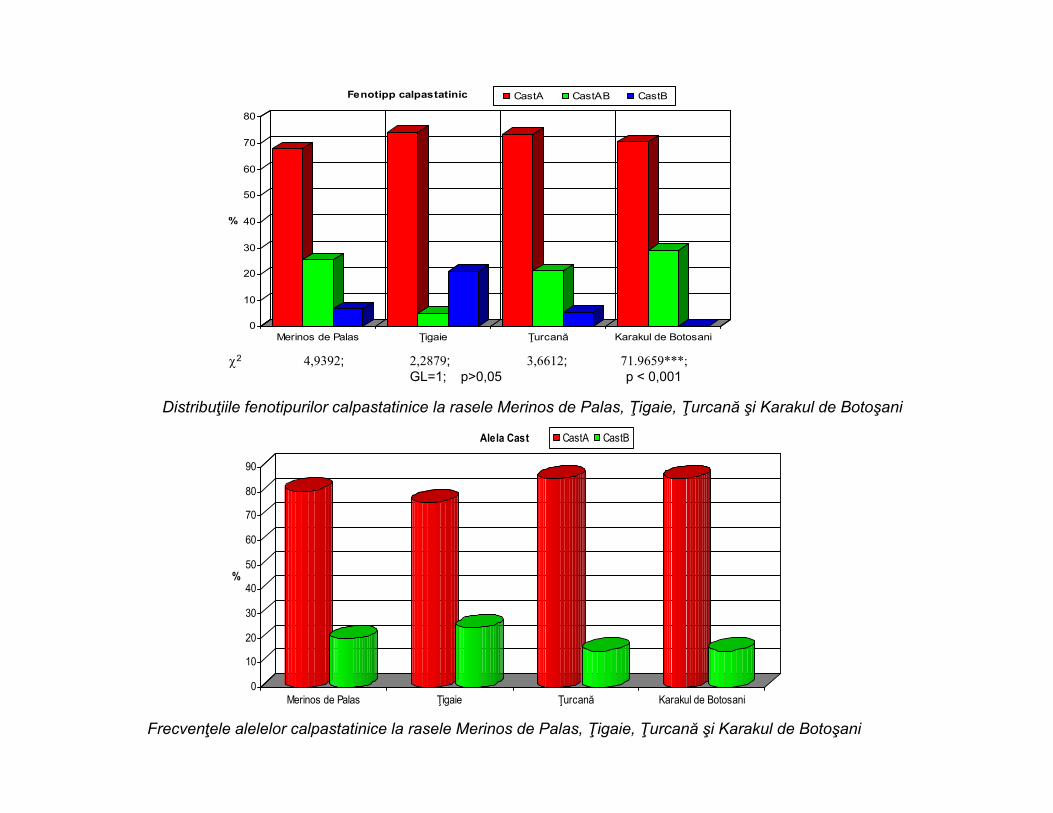
0
10
20
30
40
50
60
70
80
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Fenotipp calpastatinic CastA CastAB CastB
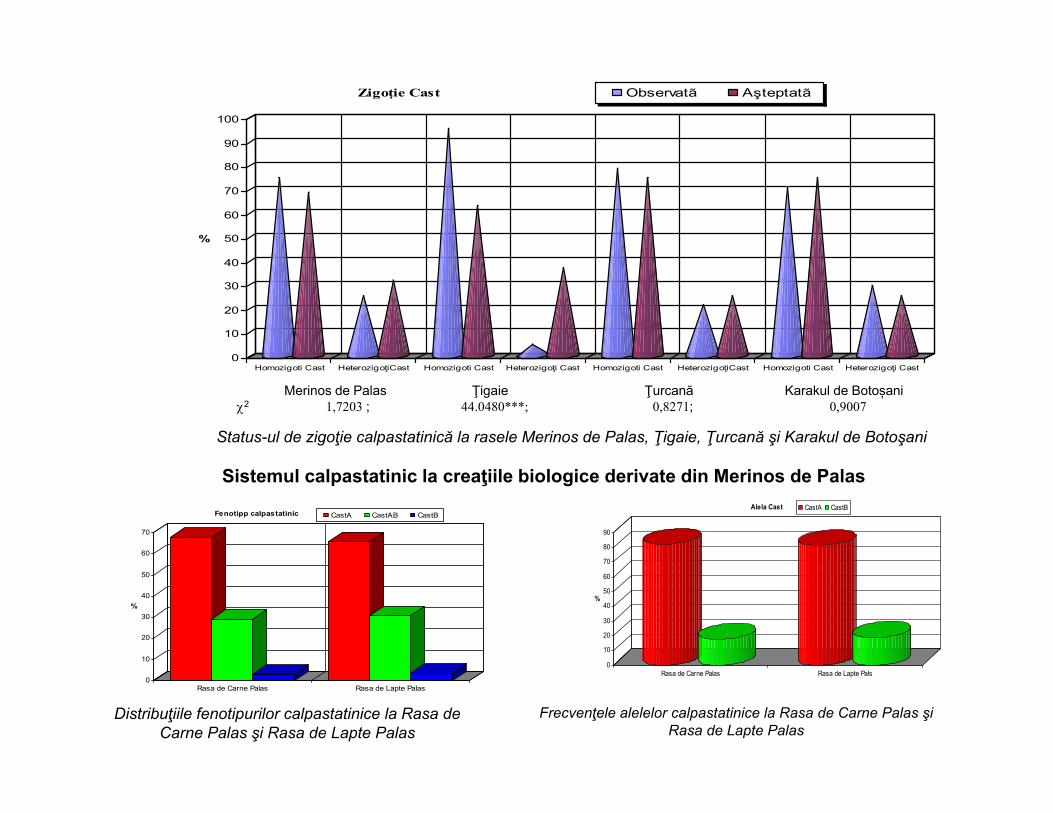
Distribuţiile fenotipurilor calpastatinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
90
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Alela Cast CastA CastB
Frecvenţele alelelor calpastatinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
2 4,9392; 2,2879; 3,6612; 71.9659***;GL=1; p>0,05 p < 0,001
0
10
20
30
40
50
60
70
80
90
100
%
Homozigoti Cast HeterozigoţiCast Homozigoti Cast Heterozigoţi Cast Homozigoti Cast HeterozigoţiCast Homozigoti Cast Heterozigoţi Cast
Zigoţie Cast Observatã Aşteptatã
Merinos de Palas Ţigaie Ţurcană Karakul de Botoșani2 1,7203 ; 44.0480***; 0,8271; 0,9007
0
10
20
30
40
50
60
70
%
Rasa de Carne Palas Rasa de Lapte Palas
Fenotipp calpastatinic CastA CastAB CastB
0
10
20
30
40
50
60
70
80
90
%
Rasa de Carne Palas Rasa de Lapte Pals
Alela Cast CastA CastB
Status-ul de zigoţie calpastatinică la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
Sistemul calpastatinic la creaţiile biologice derivate din Merinos de Palas
Distribuţiile fenotipurilor calpastatinice la Rasa de Carne Palas şi Rasa de Lapte Palas
Frecvenţele alelelor calpastatinice la Rasa de Carne Palas şi Rasa de Lapte Palas

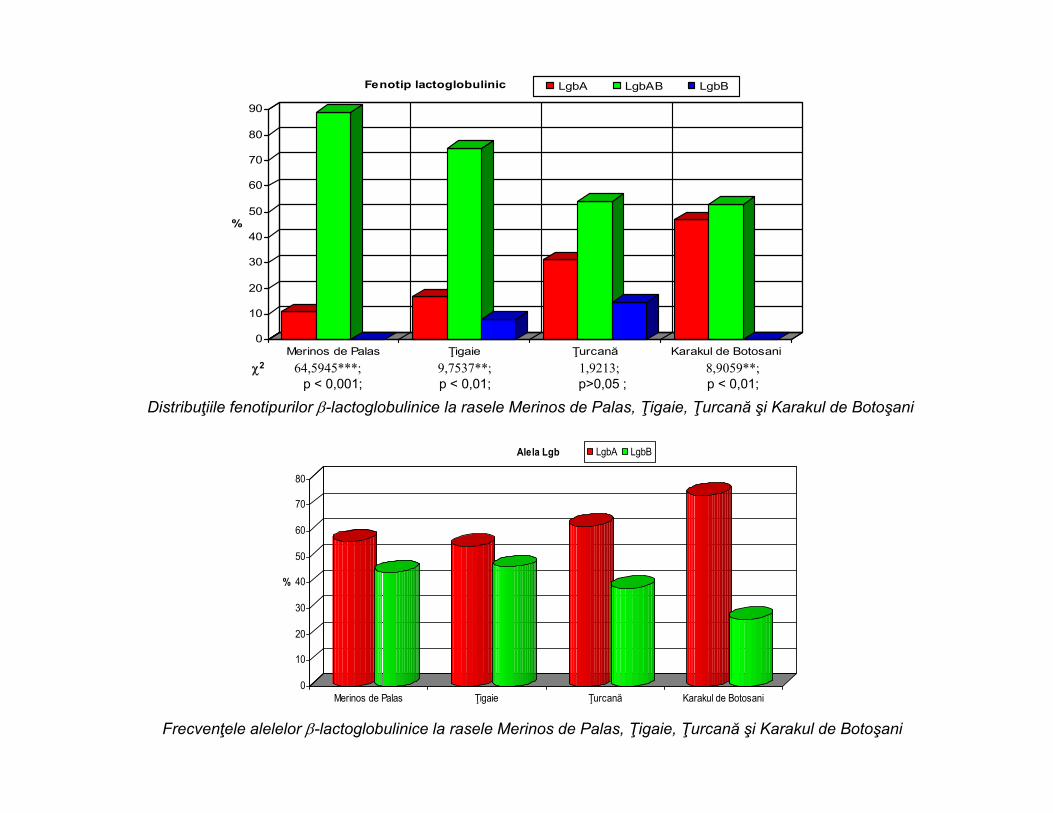
Sistemul -lactoglobulinic la rasele de ovine din România
Sistemul -lactoglobulinic la rasele clasice de ovine
Electroforegramă cu profilele de restricţie pentru genotiparea locus-ului -lactoglobulină; 1, 2, 3, 5: genotip AB (103bp, 66bp, 37 bp şi 17bp); 4, 6: genotip AA (66bp, 37bp, şi 17 bp); 7: fragment netăiat; 8: marker de masă moleculară 50bp
0
10
20
30
40
50
60
70
80
90
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Fenotip lactoglobulinic LgbA LgbAB LgbB
Distribuţiile fenotipurilor -lactoglobulinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
0
10
20
30
40
50
60
70
80
%
Merinos de Palas Ţigaie Ţurcană Karakul de Botosani
Alela Lgb LgbA LgbB
Frecvenţele alelelor -lactoglobulinice la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
2 64,5945***; 9,7537**; 1,9213; 8,9059**; p < 0,001; p < 0,01; p>0,05 ; p < 0,01;
0
10
20
30
40
50
60
70
80
90
%
Homozigot i Lgb Heterozigoţi Lgb Homozigot i Lgb Heterozigoţi Lgb Homozigot i Lgb Heterozigoţi Lgb Homozigot i Lgb Heterozigoţi Lgb
Zigoţie Lgb Observatã Aşteptatã
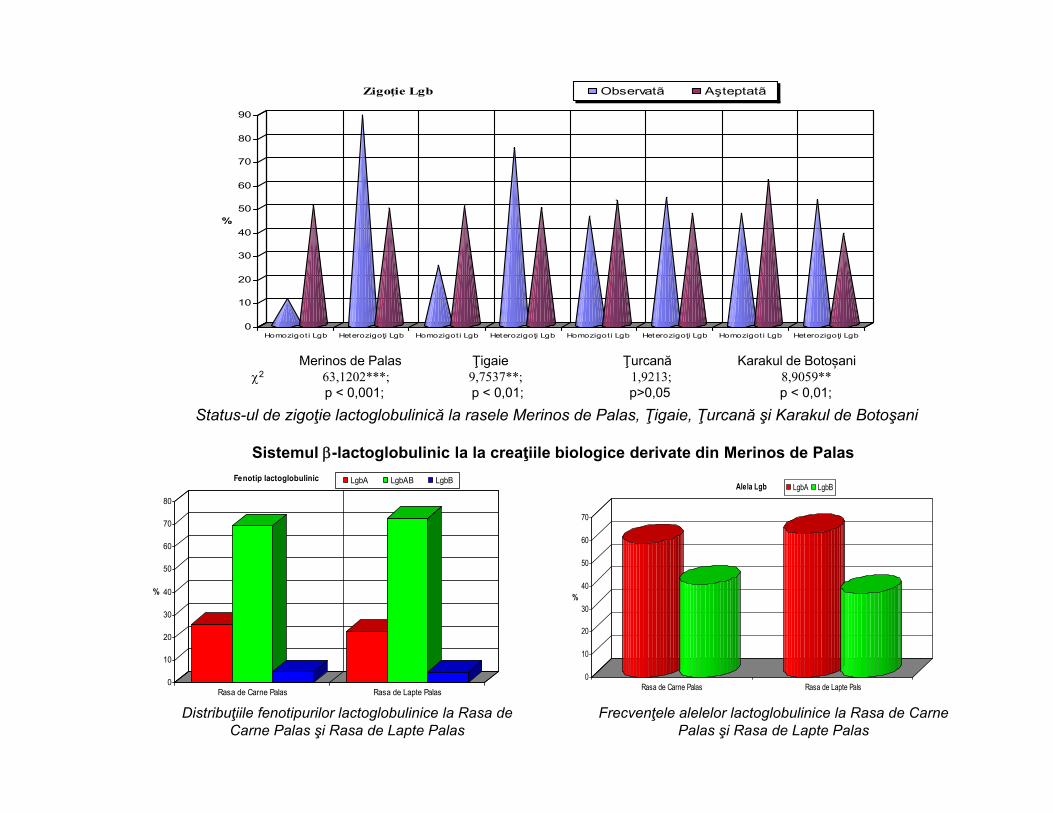
Status-ul de zigoţie lactoglobulinică la rasele Merinos de Palas, Ţigaie, Ţurcană şi Karakul de Botoşani
Merinos de Palas Ţigaie Ţurcană Karakul de Botoșani2 63,1202***; 9,7537**; 1,9213; 8,9059**
p < 0,001; p < 0,01; p>0,05 p < 0,01;
Sistemul -lactoglobulinic la la creaţiile biologice derivate din Merinos de Palas
0
10
20
30
40
50
60
70
80
%
Rasa de Carne Palas Rasa de Lapte Palas
Fenotip lactoglobulinic LgbA LgbAB LgbB
0
10
20
30
40
50
60
70
%
Rasa de Carne Palas Rasa de Lapte Pals
Alela Lgb LgbA LgbB
Distribuţiile fenotipurilor lactoglobulinice la Rasa de Carne Palas şi Rasa de Lapte Palas
Frecvenţele alelelor lactoglobulinice la Rasa de Carne Palas şi Rasa de Lapte Palas
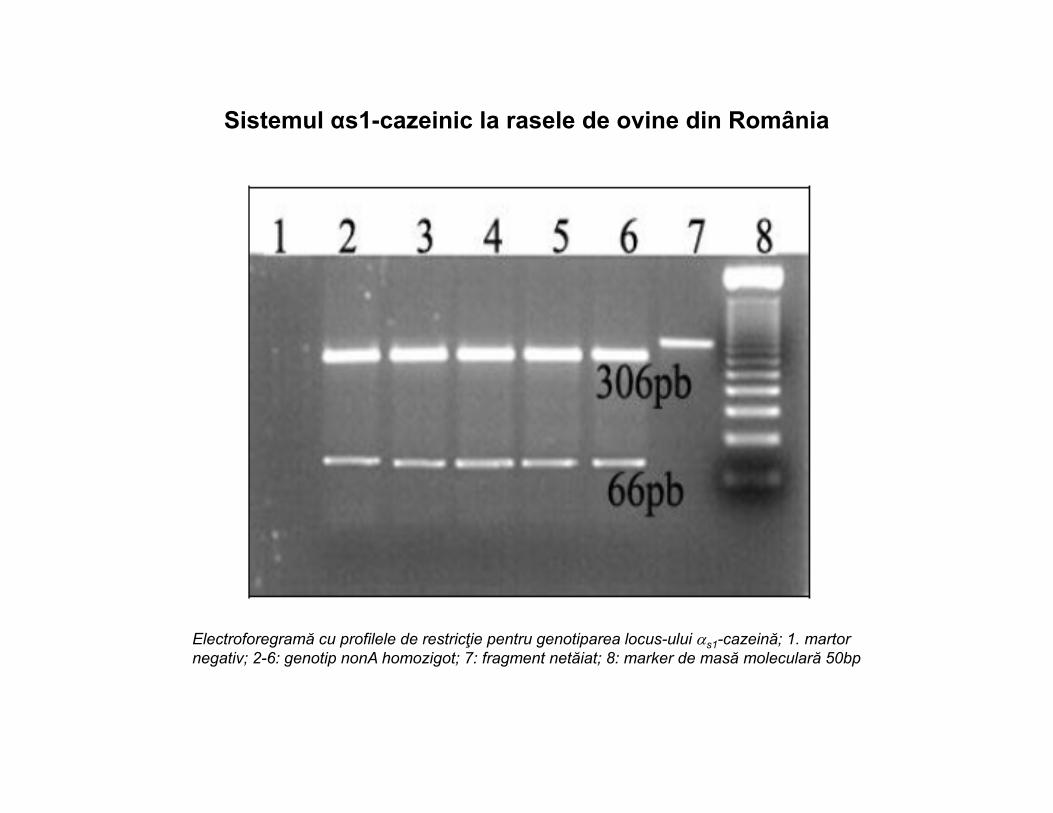
Sistemul αs1-cazeinic la rasele de ovine din România
Electroforegramă cu profilele de restricţie pentru genotiparea locus-ului s1-cazeină; 1. martor negativ; 2-6: genotip nonA homozigot; 7: fragment netăiat; 8: marker de masă moleculară 50bp

B. Caracterizarea econogenică a raselor de ovine din România
Rasa Merinos de Palas
Principalele dimensiuni corporale la rasa Merinos de Palas
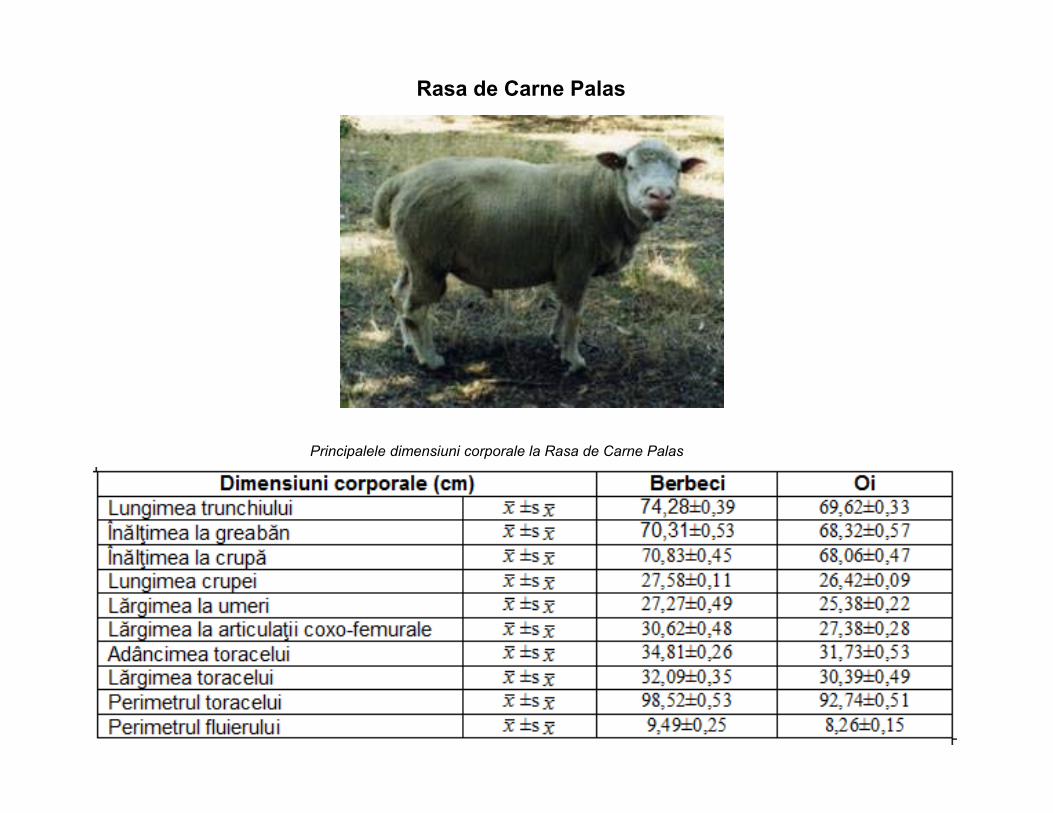
Rasa de Carne Palas
Principalele dimensiuni corporale la Rasa de Carne Palas
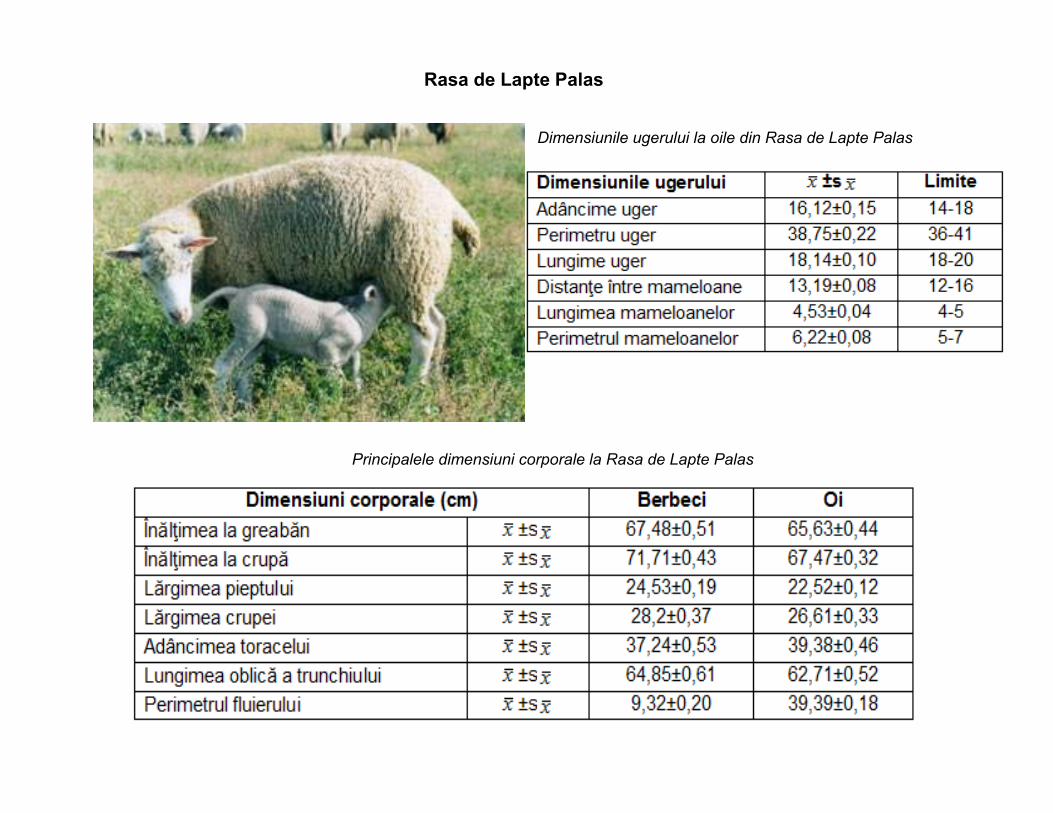
Rasa de Lapte Palas
Principalele dimensiuni corporale la Rasa de Lapte Palas
Dimensiunile ugerului la oile din Rasa de Lapte Palas
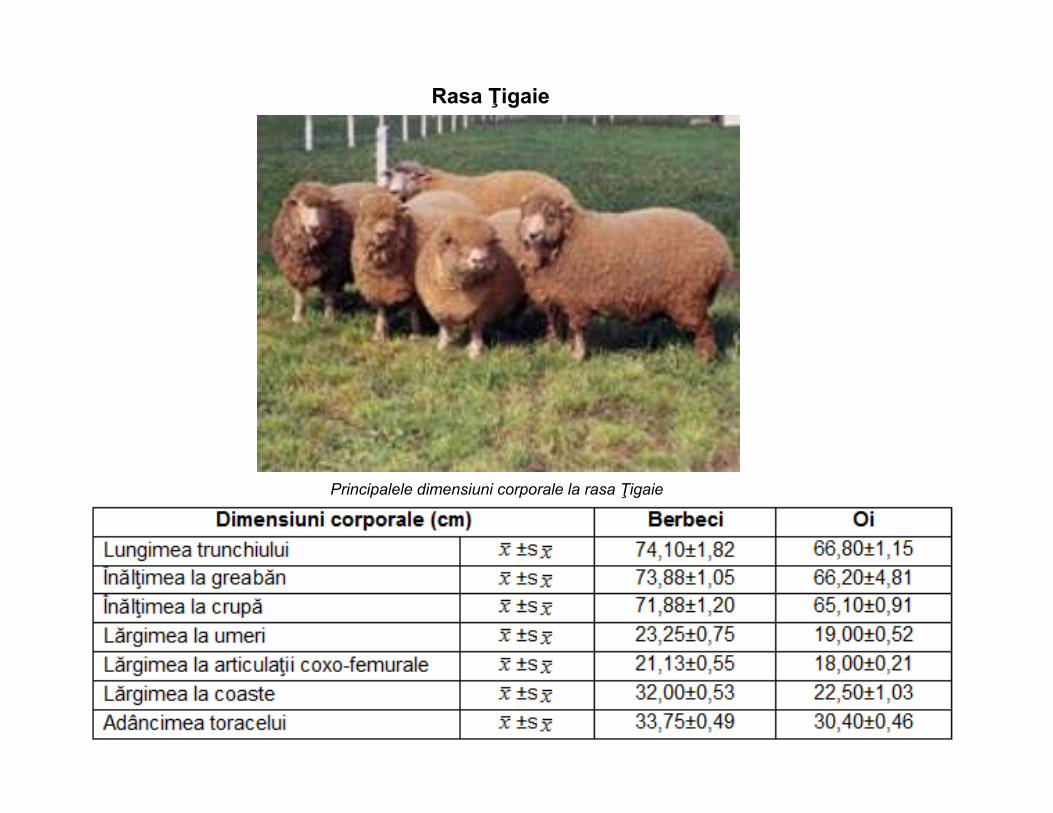
Rasa Ţigaie
Principalele dimensiuni corporale la rasa Ţigaie

Rasa Ţurcană
Principalele dimensiuni corporale la rasa Ţurcană
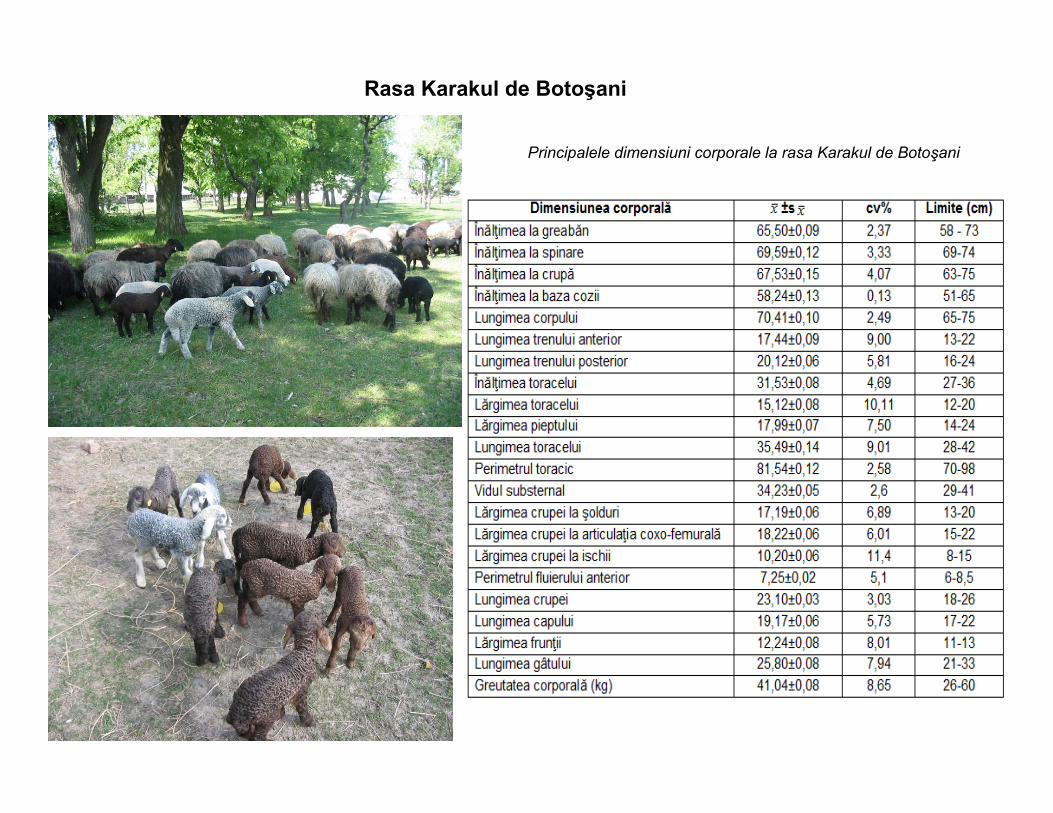
Rasa Karakul de Botoşani
Principalele dimensiuni corporale la rasa Karakul de Botoşani

Concluzii1. Principalul obiectiv al celei de a doua faze a proiectului este caracterizarea
genetico-biochimică, genetico-moleculară şi econogenică a raselor de ovine crescute şi exploatate în România (Merinos de Palas, Ţigaie, Ţurcană, Karakul de Botoşani, Rasa de Carne Palas şi Rasa de Lapte Palas).
2. Determinarea structurilor genetice ale raselor de ovine din România s-a realizat prin metode specifice de laborator: electroforeza pentru identificarea fenotipurilor de hemoglobină, transferină, albumină şi amilază, flamfotometria pentru decelarea fenotipurilor potasice şi PCR-RFLP pentru genotiparea ovinelor la locii determinanţi ai calpastatinei, .lactoglobulinei şi s1-cazeinei.
3. Rasele de ovine din România se caracterizează printr-un polimorfism mai mult sau mai puţin pronunţat la locii determinanţi ai diferitelor sisteme genetico-biochimice şi genetico-moleculare, amplitudinea acestui polimorfism depinzând de sistemul genetic analizat şi de rasa de ovine.
4. Tipizarea genetică la locii determinanţi ai unor proteine sau elemente minerale cu forme moleculare multiple au sos în evidenţă polimorfismul gradual al acestor structuri, de la monotipism (albumina la Ţurcană şi Karakul de Botoşani, amilaza la toate rasele cu excepţia Ţigaiei, potasiul sanguin la Merinos de Palas sau s1-cazeina la toată rasele), la polimorfism de tip binar (hemoglobina, potasiul sanguin, calpastatina, -lactoglobulina la majoritatea raselor) sau la polimorfism polialelic foarte pronunţat (transferina la toate rasele de ovine); la specia ovină nu există monotipism la locus-ul Tf.

5. Sistemul hemoglobinic la ovine se caracterizează printr-un polimorfism mediude tip binar, concretizat prin existenţa a două alele: HbB mult mai des întâlnită decâtcodominanta sa alela HbA, ceea ce determină exprimarea a trei genotipuri: homozigoţiiHbBHbB sunt preponderenţi, însă homozigoţii pentru alela HbA au o incidenţă scăzută;prezenţa heterozigoţilor HbAHbB este moderată. Nici o rasă românească de ovine nu estemonotipică, rasa Karakul de Botoşani având două genotipuri, iar la celelalte cinci rase segăsesc toate cele trei genotipuri.
6. Sistemul transferinic este cel mai polimorf sistem cu nouă alele Tf codominante(TfI, TfA, TfG, TfB, TfC, TfM, TfD, TfE, TfP), răspândite în toate rasele,. Cele nouă alele segăsesc doar la Ţigaie,la celelalte rase numărul acestor alele este mai mic, scăzând la şasela Karakul de Botoşani. cele mai frecvente sunt alelele: TfA, TfD, TfC, TfB şi TfM. Alelele TfI,TfG, TfE şi TfP au o răspândire mai slabă. În funcţie de numărul alelelor şi de capacitatea lorcombinativă se defineşte tabloul genotipic transferinic caracteristic pentru fiecare rasă
7. Sistemul albuminic se caracterizează printr-un polimorfism restrâns, de tipbinar, ca la Merinos de Palas, Ţigaie, Rasa de Carne Palas şi Rasa de Lapte Palas, sau prinmonotipism ca la rasele Ţurcană şi Karakul de Botoşani. La rasele politipice, predominanteîn panelul albuminic sunt alela AlbS şi genotipul homozigot al acesteia AlbSAlbS.
8. Sistemul potasiului sanguin are un polimorfism mediu. La rasele Merinos dePalas, Ţigaie, Ţurcană şi Rasa de Carne Palas predomină fenotipul LK (cu potasemiescăzută), iar la Karakul de Botoşani covârşitor este fenotipul HK (cu potasemie ridicată). LaRasa de Lapte Palas, deşi fenotipul HK este mai frecvent decât fenotipul LK, totuşi raportuldintre cele două fenotipuri este ceva mai echilibrat.
9. Sistemul amilazic este cel mai puţin polimorf sistem proteic, singura rasăromânească politipică fiind Ţigaia la care alela AmB foarte frecventă în comparaţie cu alelaAmC. Acest fapt determină ca tabloul amilazic să fie ocupat de homozigoţii AmBAmB. Raselemonotipice sunt pe alela AmB.

10. La rasele autohtone din România se găsesc toate cele trei variante fenotipicede calpastatină. Fenotipul CAST A este foarte frecvent la toate rasele, iar heterozigoţiiCASTACASTB ocupă între 25% şi 35% din panelul calpastatinic. Prezenţa homoserotipuluiCast B este destul de redusă. Cea mai evidentă asemănare în privinţa acestui markermolecular este consemnată între Rasa de Carne Palas şi Rasa de Lapte Palas.
11. Cele două alele identificate în sistemul -lactoglobulinic, LGBA şi LGBB, cu ofrecvenţă mai mare a primei alele, determină expresia a trei tipuri genetice a acesteiproteine. Cel mai bine reprezentat este heterozigotul LGBALGBB, în special la rasa Merinosde Palas. Homozigoţii LGBBLGBB sunt foarte puţin răspândiţi la majoritatea raselor sauabsenţi, precum la Merinos de Palas sau Karakul de Botoşani. Homozigoţii pentru alelaLGBA înregistrează o incidenţă apreciabilă la Ţurcană şi mai ales la Karakul de Botoşani,însă la celelalte rase se întâlnesc destul de rară.
12. La locus-ul determinant al s1-cazeinei nu există polimorfism, raseleromâneşti de ovine fiind monotipice sub acest aspect, la toate fiind evidenţiată doarprezenţa genotpului homozigot CnCCnC.
13. Gradul de polimorfism sistemic în cadrul fiecărei rase de ovine estecondiţionat, uneori fundamental, de numărul de alele pe care-l posedă fiecare rasă, dar maiales capacitatea combinatorie a alelelor pentru exprimarea genotipurilor specifice la nivelulfiecărui locus proteic sau mineral, numărul de genotipuri fenotipizate, distribuţiile observateale acestora, aşteptările apariţiei acestora conform legii Hardy-Weinberg, modul lor degrupare, marja de variabilitate, gradul de dispersare etc.
14. Polimorfismul genetic al proteinelor şi elementelor minerale contribuie în moddecisiv la stratificări chiar în cadrul aceleiaşi rase prin crearea de entităţi infrarasiale cum arfi varietăţile, liniile zootehnice, ecotipurile, tipurile morfo-productive etc., contribuind lasporirea diversităţii genetice la specia ovină.

15. Un rol foarte important în caracterizarea genetică a raselor de ovine dinRomânia este testarea echilibrului genetic Hardy-Weinberg la nivelul diverşilor loci careexprimă gradul de homeostazie fiziologică şi genetică a raselor respective şi a entităţilorlor infrarasiale.
16. Diferenţierea genetico-biochimică şi genetico-moleculară a raselor de ovinedin ţara noastră la locii sistemelor polimorfe este datorată acţiunii combinate şi complexea presiunii de selecţie la care sunt supuse populaţiile de animale în proceseletehnologice de ameliorare, creştere şi reproducţie, dar şi condiţiilor meteo-geo-climatice(temperatură, altitudine, umiditate, regim de păşunare), precum şi particularităţiloreconogenice (istoric, tradiţii, obiceiuri, motivaţie, specific cultural şi social în creştereaovinelor) din arealele de existenţă specifice raselor respective.
17 Polimorfismul sistemelor genetice poate avea caracter tranzitoriu. Acţiuneasupralicitantă a unor factori de mediu sau tehnologici poate avea ca rezultat eliminareaunor alele de la nivelul anumitor loci, ducând la restrângerea polimorfismului acestorasau chiar, în anumite cazuri extreme, la apariţia formelor monotipice.
18. Existenţa polimorfismului genetic proteic şi mineral este condiţia decisivăde diferenţiere a raselor între ele şi de sporire a biodiversităţii în cadrul speciei ovină.
19. Graţie caracterului foarte heterogen al sistemelor genetico-biochimice şigenetico-moleculare, biostructurile cu forme moleculare multiple prezintă un imensinteres economic şi ştiinţific putând fi folosite cu succes ca markeri genetici în lucrările deselecţie timpurie pentru ameliorarea efectivelor de ovine în direcţia dictată deimperativele economice, de creare a noi entităţi infrarasiale mai productive, dar şi laelaborarea unor programe sustenabile pentru protecţia potenţialului genetic, conservareasa şi sporirea biodivesităţii folosind informaţii furnizate de investigaţiile biochimice şimoleculare în consonanţă cu imperativele politicilor Comunităţii Europene în domeniulagriculturii şi mai ales a ştiinţei animalelor.





























