Embed Size (px)
Citation preview

CYTOKININ AUTONOMY IN TISSUE CULTURES OF PHASEOLUS: A GENOTYPE-SPECIFIC AND HERITABLE TRAIT'
MACHTELD C. MOK,2 DAVID W. S. MOK,? DONALD J. ARMSTRONG,3 AIMPE RABAKOARIHANTA~ A N D SANG-GU KIM^
Department of Horticulture, Genetics Program and Department of Botany and Plant Pathology, Oregon State Uniuersity, Corvallis, Oregon 97331
Manuscript received March 1, 1979 Revised copy received November 8,1979
ABSTRACT
Intra- and interspecific differences in cytokinin requirement were detected in callus cultures of Phaseolus vulgaris L. and P. lunatus L. Of the ten geno- types of P. uulgaris tested in the present study, one required cytokinin for callus growth, six exhibited some to moderate growth on cytokinin-free medium, and the remaining three grew uniformly in the absence of cytokinin. In contrast, six of the P. lunatus genotypes were strictly cytokinin-dependent, while four genotypes displayed irregular amount of callus growth on cytokinin- free medium. The genotype-specific behavior of Phaseolus callus tissues was independent of the tissue of origin and the time in culture. The inheritance of the cytokinin requirement of Phareolus tissue cultures was studied in hybrid tissues resulting from crosses between a strictly cytokinin-dependent genotype (P.I. 2aO960) and two independent genotypes (cv. G 50 and P.I. 286303) of P . vulgaris. Fresh weights of hybrid tissues on cytokinin-free medium were intermediate between and significantly different from the parental tissues. No differences were found between reciprocal hybrids. These results suggest that cytokinin autonomy in tissue cultures of P. uulgaris is a genetic trait under nuclear control. Both parental and intermediate phenotypes were recovered in the F, progeny. The frequency distribution of cytokinin-dependent progeny in F, and backcross populations indicates that the cytokinin requirement of P. vulgaris callus tissue may be regulated by one set of alleles.
HE nature of the genetic regulatory mechanisms controlling hormonal Tmetabolism in plant tissues constitutes one of the major problems in plant hormone research. Derangements in the regulation of hormonal metabolism are known to occur in the tumor-prone hybrids that result from certain interspecific crosses (SMITH 1972), and a number of mutations affecting the biosynthesis of particular plant hormones have been described ( GELINAS and POSTLETHWAIT 1969; IMBER and TAL 1970; PHINNEY 1956; ZOBEL 1974). However, the devel- opment of strategies for the systematic identification and characterization of the
Technical paper No. 5103 of the Oregon Agricultural Experiment Station. Research was supported by the Science and Education Administration of the U.S. Deparbnent of Agriculture under Grant 5901-0410-8-0028-0 from the Com- petitive Research Grants Office, by the Research Council of Oregon State University (NIH Biomedical Research Support Grant RR07079) and by the Oregon Agricultural Experiment Station.
2 Depal-iment of Horticulture. Department of Botany and Plant Pathology.
Genetics 94: 675-686 March, 1980.

6 76 M. c. MOR et al.
genetic factors involved in the hormonal regulation of plant growth has received only limited attention to date.
The genetic regulation of cytokinin metabolism in Phaseolus and other food legumes is currently under investigation in our laboratories. Our initial studies have centered on the identification of intrinsic genetic differences in the cyto- kinin metabolism of particular Phaseolus genotypes. Phenotypic traits indicative of possible genetic variations in cytokinin metabolism are more easily defined for tissue-culture systems than for intact plants, and the criteria we have em- ployed for the selection of Phaseolus genotypes of interest have been based on the use of tissue-culture phenotypes. The detection of pronounced interspecific differ- ences in the ability of cytokinin-dependent callus tissues derived from P. vulgaris and P. lunatus to respond to cytokinim with saturated and unsaturated N6-side chains was reported in an earlier publication (MoK, MOK and ARMSTRONG 1978). The present paper reports the identification of genetically determined differences in the ability of Phaseolus callus cultures to exhibit cytokinin- autonomous growth (growth on cytokinin-free medium). The genetic variability in the cytokinin dependence of P. vulgaris and P. lunatus callus tissues, the influ- ence of tissue origin and time in culture on cytokinin-autonomous growth, and the results of studies on the inheritance of cytokinin autonomy in selected P. vulgaris genotypes are described.
MATERIALS AND METHODS
Plant materials: Seeds of Plant Introduction lines (P.I. lines) of Phseolus uulgaris L. and P. Zunatus L. were obtained from the Regional Plant Introduction Station, Washington State University, Pullman, Washington, and from D. P. COYNE, University of Nebraska, Lincoln, Nebraska. Seeds of commercial varieties of P. uulgaris were prsvided by the Asgrow Seed Com- pany. Seeds of P. lunatus CY. Kingston were obtained locally. Ten genotypes of each of the two Phaseolus species were tested for tissue culture phenotype. The P. vulgaris genotypes were cv. Kentucky 191 (K 191) and Gallatin 50 (G 50), P.I. 165070, 169754, 181955,200960,286303, 306158 and 300671. The P. lunatus genotypes were cv. Kingston, P.I. 180324, 202830, u)876Ei, 256845, 257409, 257412, 257547, 257560 and 260415. P. vulgaris cv. G 50 and P.I. 200960 were used to study the influence of tissue origin and time in culture on cytokinin autonomy. Genetic studies were performed with P. uu2garis cv. G 50, P.I. 200960, and P.I.286303 as parental lines and F, hybrids obtained from P.I. 200960 x cv. G 50 (and reciprocal) and P.I. 200960 x P.I. 286303 (and reciprocal). A large number o€ F, progeny of P.I. 200960 x cv. G 50 (and recip- rocal) and P.I. 200960 x P.I. 286303 (and reciprocal) were sampled. In addition, backcross progeny of P.I. 200960 x P.I. 286303 to P.I. 200960 were examined.
Tissue-culture medium: In all experiments, the tissue-culture medium consisted of the mineral nutrients described by MURASHIGE and SKOCG (1962), with the following substances added: sucrose (30 g/l), myo-inositol (100 mg/l), thiamine.HC1 (1 mg/l), nicotinic acid (5 mg/l), pyridoxine.HC1 (0.5 mg/l) and picloram (2.5 p). Kinetin (5 ELM) was included in the medium used for callus initiation, stock cultures and control cultures. The pH of the medium was adjusted to 5.7, and DiEco Bacto-agar (IO g/l) was added. The medium was dispensed into 125 ml Erlenmeyer flasks (50 ml/flask) and autoclaved at 120" for 15 min.
Growth and harvest of Phaseolus callus cultures: Unless otherwise indicated, Phaseolus callus cultures were established from the hypocotyls of five-day-old seedlings as previously described (MOH and MOK 1977). The callus tissue that formed on the initial explants was transferred once (first passage) on medium containing 5 p ~ kinetin. Tests for cytokinin autonomy (growth on cytokinin-free medium) were performed on the second passage of the callus tissues, except

CYTOKININ AUTONOMY IN PHASEOLUS 677 as indicated. All cultures were grown in the dark at 27". Routinely, four seeds were germinated per genotype, and callus was tested on cytokinin-free medium, using two or four replicate flasks per seed for each harvest date. The replicate flasks contained callus with different lineage; i.e., the callus was derived from different hypocotyl explants. Four replicate flasks were used in tests to determine the genetic variability in cytokinin requirement and the inheritance of cytokinin autonomy. Two replicate flasks per seed were used for control cultures (5 PM kinetin). The results of all tests were expressed as mean fresh weights per flask. Standard deviations were calculated for mean fresh weights of callus derived €ram individual seeds and thus are measure- ments of the variation between seeds of the same line. In genetic studies, the parental and F, populations were compared in the Newman-Keuls test at the 5% level (SNEDECOR and C~CHRAN 1967). Chi-square values were used for goodness of fit tests (5% level) of F, and backcross population distributions.
RESULTS
Genetic variability in the cytokinin requirement of Phasedus tissue cultures: Tissue culture lines derived from each of ten P. vulgaris and ten P. lunatus genotypes were tested for ability to grow on cytokinin-free medium. The results are illustrated in Figures 1 and 2, respectively. Of the ten P. vulgaris genotypes tested, three (P.I. 181955, P.I. 286303 and cv. G 50) yielded callus tissue that grew rapidly and uniformly in the absence of cytokinin; six genotypes exhibited moderate growth under these conditions; and only one (P.I. 200960) displayed little, if any, tendency to grow on cytokinin-free medium. In contrast, six of the P. lunatus genotypes gave callus lines that were strictly cytokinin-dependent and the four other genotypes exhibited very irregular growth on cytokinin-free medium. It should be noted that all tissues grew rapidly and uniformly on medium containing 5 PM kinetin. After five weeks growth on kinetin-containing medium, the fresh weights of callus ranged from 16.6 to 24.6 g/flask for the P. vulgaris genotypes and from 15.3 to 20.5 g/flask for the P. lunatus genotypes.
Further characterization of the callus tissues derived from selected P. vulgaris genotypes (P.Z. 200960 and cv. G 50): The growth responses of the cytokinin- requiring callus cultures derived from P . vulgaris P.I. 200960 and the cytokinin- autonomous cultures derived from P . vulgaris cv. G 50 were examined in detail. Growth curves established for tissues grown on medium with and without cyto- kinin are shown in Figure 3. Tissues of both genotypes grew rapidly on medium containing 5 JLM kinetin, and fresh weights of over 20 g/flask were attained in five weeks. On medium without cytokinin, tissues of cv. G 50 grew continuously, although the growth rate was slower than that on cytokinin-containing medium. The increase in fresh weight was linear from the third to the eighth week of the culture period and was followed by a stationary phase. In contrast, there was almost no increase in fresh weight of tissues of P.I. 200960 on cytokinin-free medium, and the callus rapidly turned brown.
To determine whether tissue origin might influence the cytokinin requirement of the callus tissues, callus cultures were established from hypocotyls, cotyledons and roots of five-day-old seedlings of P.I. 200960 and cv. G 50 and from petioles of mature plants of the same two genotypes. The fresh weights of tissues after growth periods of five and seven weeks on cytokinin-free medium and after five weeks on medium containing 5 PM kinetin are presented in Table 1. Regardless of

678 M. c. MOR et al.
PI. 200960
cv. K191
PI. 306158
PI. 300671
PI. 169754
PI. 310511
0.4
1.7
3.0
5.4
6.9
7.0
PI. 165070 9. I
cv. 650 12.5
12.9 PI. 286303
PI. 181955 19. I
FIGURE 1.-Representative callus cultures from ten P. vulgaris genotypes after a Culture period of seven weeks on cytokinin-free medium. Average fresh weights (g/flask) are given at the right of the figure.
the origin of explants, callus derived from P.I. 200960 grew very little in the ab- sence of cytokinin and callus of cv. G 50 was consistently cytokinin-autonomous.
The influence of time in culture on the cytokinin requirements of P.I. 200960 and cv. G 50 callus tissues was examined in the second, fourth and sixth passages (Table 2). The inability of P.I. 200960 callus to grow without cytokinin did not change in later passages, and callus tissues derived from cv. G 50 remained cytokinin-autonomous in all passages. In fact, the fresh weights of cv. G 50 cul- tures increased slightly with the number of passages.
Inheritance of tissue culture phenotypes in P. vulgaris: The inheritance of cytokinin autonomy was examined in crosses between the cytokinin-dependent genotype, P.I. 200960, and two cytokinin-autonomous genotypes, cv. G 50 and P.I. 286303. The fresh weights of callus of parents and hybrids on cytokinin-free

PI. 257547
PI, 208765
PI. 257560
PI, 256845
PI. 257412
P 1. 260415
CYTOKININ AUTONOMY I N PNASEOLUS 679
I 1 0.1
0.1
0.1
0.1
0. I
cv. Kingston
PI. 202830
P 1.257409
PI. 180324
0.1
0.4
2.5
2.7
4.1
FIGURE 2.-Representative callus cultures fram ten P. Iunatus genotypes after a culture period of seven weeks on cytokinin-free medium. Average fresh weights (g/flask) are given at the right of the figure.
medium were determined after five weeks (Table 3) and seven weeks (Figure 4). The fresh weights of hybrid callus were intermediate between and significantly different (5% level) from those of the parents. The fresh weights of reciprocal hybrids were not significantly different from each other.
A large number of Fz progeny derived from the crosses P.I. 200960 X cv. G 50, P.I. 200960 X P.I. 286303 and reciprocals have been examined. No reciprocal cross differences were apparent in either the F, progeny or the backcross progeny described below. Parental phenotypes were recovered in the F, progeny from each cross. Among the 170 F, progeny derived from the crosses P.I. 200960 X CV. G 50 and reciprocal, 46 exhibited the cytokinin-dependent phenotype and 24 were classified as completely cytokinin-autonomous. Among the 125 F, pro- geny derived from the crosses P.I. 200960 x P.I. 286303 and reciprocal, 36 exhib-

680 M. c. MOK et al.
5 L i - I A ’ : ~ ~ ~ ~ i ,/‘ ; _--
I 2 3 4 5 6 7 8 9 IO T I M E (WEEKS)
FIGURE 3.-Growth curves of P.I. 200960 ( 0 ) and cv. G 50 (A) callus tissues on medium without cytokinin ( ) and medium containing 5 kinetin (-----) .
ited the cytokinin-dependent phenotype and 15 were classified as completely cytokinin-autonomous. The remaining F, progeny from each cross were classi- fied as intermediate phenotypes, but the variation in growth habit of the inter- mediate F2 progeny was much greater than that in the corresponding F, pheno- type. Among the F, progeny classified as intermediate phenotypes, variations in the color of the callus tissue and in the amount and uniformity of growth were
TABLE 1 Influence of tissue origin on the cytokinin requirements of P. vulgaris callus tissues
Genotype Plant part
P.I. 200960 hypocotyl petiole cotyledon root
cv. G 50 hypocotyl petiole cotyledon root
5 w!a
0.3 0.2 0.6 0.4
8.2 8.3 7.7 6.4
Fresh weight (g/ilask) 0 5 PM kinetin
7 wks 5 wks
0.2 26.2 0.3 26.0 0.7 u1.7 0.2 24.3
14.1 25.9 14.3 27.7 15.3 20.1 12.9 16.1
Fresh weights of callus cultures derived from hypocotyl, petiole, cotyledon and root tissues of P.I. 200960 and cv. G 50 were compared after growth periods of five and seven weeks on cyto- kinin-free medium and five weeks on medium containing 5 PM kinetin.

CYTOKININ AUTONOMY I N PHASEOLUS 681
TABLE 2 Influence of time in culture on the cytokinin requirements of P. vulgaris callus tissues
Genotype Passage Fresh weight (g/flask) 5 NM kinetin 0
5 wks 7 wks 5 wks
P.I. 200960
cv. G 50
2 0.2 0.2 26.4 4 0.4 0.3 26.5 6 0.3 0.4 26.1
2 7.8 13.6 23.8 4 9.0 14.8 22.1 6 9.6 17.2 22.2
Fresh weights of callus cultures of P.I. 200960 and CY. G 50 were compared in the second, fourth and sixth passage after growth periods of five and seven weeks on cytokinin-free medium and five weeks on medium containing 5 PM kinetin.
evident. The F, progeny classified as parental phenotypes and examples of intermediate phenotypes are illustrated in Figures 5 and 6.
The inheritance of the cytokinin-dependent trait was further examined in backcrosses of the PI (P.I. 200960 X P.I. 286303 and reciprocal) to P.I. 200960. As expected, only cytokinin-dependent and intermediate phenotypes were observed in the backcross progeny. Among 44 backcross progeny examined, 17 were strictly cytokinin-dependent. The data for F, and backcross progeny are summarized in Table 4. Because it proved much easier to recognize the cytokinin- dependent phenotype than to distinguish between intermediate and fully cyto- kinin-autonomous phenotypes, only the cytokinin-dependent progeny are tabu- lated separately. On this basis, the proportion of cytokinin-dependent F, progeny was approximately 1/4 in all crosses examined, and the recovery of cytokinin- dependent progeny in the backcross was not significantly different from 1/2.
TABLE 3 Cytokinin requirements of tissue cultures derived from hybrids of selected P. vulgaris genotypes
Genatype Fresh weight (g/flask) 0 5 p kinetin
P.I. 200960 cv. G 50 P.I. 286303 P.I. 200960 x cv. G 50 cv. G 50 x P.I. 200960 P.I. 200960 X P.I. 286303 P.I. 286303 x P.I. 200960
0.2 * O.I** 24.0 7.0 +- 0.9c 18.6 7.2 f 1.1" 15.2 4.3 f 1.3b 25.2 3.1 +- 1.0b 24.9 2.9 zk 0.5b 22.2 3.4 f 1.4b 21.4
* Different superscripts indicate significant differences at the 5% level in the Newman-Keuls test.
Fresh weights and their standard deviations of callus cultures derived from three parental l i e s and reciprocal hybrids were determined after a culture period of five weeks on medium' without cytokinin and medium containing 5 p kinetin.

682 M. c. MOK et al.
cv. 650 i - x 12.1
cv. 650
PI. 200960 X 5.0
4.6
12.2
FIGURE 4.-Representative callus cultures derived from three P. uulgaris genotypes and their reciprocal hybrids after a culture period of seven weeks on cytokinin-free medium. Average fresh weights (g/flask) are given at the right of the figure.
DISCUSSION
Genetic analysis of the regulation of hormonal metabolism presents special problems because of the fundamental role of plant hormones in controlling development. Thus, cytokinin- and auxin-resistant mutants of the moss, Phys- comitrella patens, were reported to exhibit morphological abnormalities that resulted in sterility and necessitated the use of nonconventional methods of genetic analysis (ASHTON, GRIMSLEY and COVE 1979). In the dioecious angio-
TABLE 4
Frequency distribution of cytokinin-dependent progeny in F, and backcross populations
Progeny population Number of progeny Expected ratio Chi-square Cytokinin-dependerit 'I'otal -
F, (P.I. 200960-cv. G 50) 4 6 170 1 :4 0.38 n.s. F2 (P.I. 200960-P.I. 286303) 36 125 1:4 0.96 n.s. EC (P.I. 200960-P.I. 286303 17 44 1:2 2.27 n.s.
to P.I. 200960)
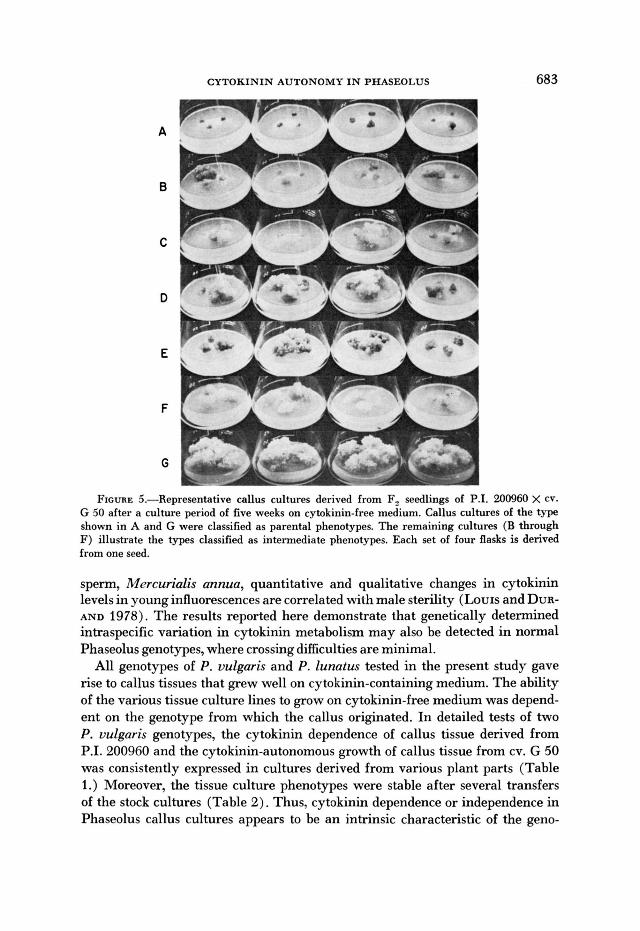
CYTOKININ AUTONOMT IN PHASEOLUS 683
A
B
C
D
E
F
FIGURE 5.-Representative callus cultures derived from Fz seedlings of P.I. '200960 X cv. G 50 after a culture period of five weeks on cytokinin-free medium. Callus cultures of the type shown in A and G were classified as parental phenotypes. The remaining cultures (B through F) illustrate the types classified as intermediate phenotypes. Each set of four flasks is derived from one seed.
sperm, Mercurialis annua, quantitative and qualitative changes in cytokinin levels in young influorescences are correlated with male sterility (LOUIS and DUR- AND 1978). The results reported here demonstrate that genetically determined intraspecific variation in cytokinin metabolism may also be detected in normal Phaseolus genotypes, where crossing difficulties are minimal.
All genotypes of P . vulgaris and P. lunatus tested in the present study gave rise to callus tissues that grew well on cytokinin-containing medium. The ability of the various tissue culture lines to grow on cytokinin-free medium was depend- ent on the genotype from which the callus originated. In detailed tests of two P . vulgaris genotypes, the cytokinin dependence of callus tissue derived from P.I. 200960 and the cytokinin-autonomous growth of callus tissue from cv. G 50 was consistently expressed in cultures derived from various plant parts (Table I.) Moreover, the tissue culture phenotypes were stable after several transfers of the stock cultures (Table 2). Thus, cytokinin dependence or independence in Phaseolus callus cultures appears to be an intrinsic characteristic of the geno-

684 M. C. MOK d.
A
D
E
F
G
FIGURE 6.-Representative callus cultures derived from F, seedlings of P.I. U)O960 X P.I. 286303 after a culture period of five weeks on cytokinin-free medium. Callus cultures of the type shown in A and G were classified as parental phenotypes. The remaining cultures (B through F) illustrate the types classified as intermediate phenotypes. Each set of four flasks is derived from one seed.
type, detectable in cultures regardless of the age and the origin of the tissue. Although it has been possible to obtain fully cytokinin-autonomous tissue culture lines from cytokinin-dependent Phaseolus genotypes, this has required careful selection and numerous transfers of the slow-growing callus pieces that occa- sionally arise in the cytokinin-dependent lines. The strong auxin, 2,4-dichlorophenoxyacetic acid, has been reported to stimu-
late some proliferation of soybean and tobacco callus tissue in the absence of cytokinin ( FOSKET and TORREY 1969; WITHAM 1968). Picloram has been used as an auxin source in our work with Phaseolus tissue cultures, because optimal growth of Phaseolus callus tissue is observed over a wide range of concentrations of this auxin (MOK and MOK 1977). Preliminary studies (unpublished) suggest that picloram (a rather potent auxin) may enhance the amount of callus growth obtained with cytokinin-independent Phaseolus genotypes, but qualitatively similar results may be obtained using other auxins. Callus tissues derived from Phaseolus genotypes such as P.I. 200960 were cytokinin-dependent regardless of the type of auxin used in the tissue culture medium.

CYTOKININ AUTONOMY IN PHASEOLUS 685
The difference between the two Phaseolus species with regard to the frequency of occurrence of cytokinin-autonomous genotypes was striking. The majority of the P . vulgaris genotypes tested displayed at least some tendency to be cyto- kinin-independent in culture, whereas most of the P . Zunatus genotypes gave rise to cytokinin-dependent culture lines. In an earlier study (MoK, MOK and ARM- STRONG 1978), cytokinins with an unsaturated N6-isoprenoid side chain [zeatin and N6- ( A2-isopentenyl) adenine] were found to be much less active in supporting growth of cytokinin-dependent P . vulgaris callus than the carresponding satur- ated analogs (dihydrozeatin and N5sopentyladenine). In P. Zunatus cultures, the two types of cytokinins were equally active. These observations, together with the results presented in this paper, suggest that there are important differences in the cytokinin metabolism of P . vulgaris and P . Zunatus.
The inheritance of cytokinin autonomy has been examined in crosses of selected P . vulgaris genotypes. The intermediate and similar growth of callus tissues derived from the reciprocal F, populations obtained by crossing cytokinin- dependent (P.I. 200960) and cytokinin-autonomous (cv G 50 and P.I. 286303) genotypes indicates that this trait is controlled by nuclear genes. Furthermore, the recovery of the strictly cytokinin-dependent phenotype at frequencies of approximately 1/4 of the F, progeny and approximately 1/2 in backcrosses to P.I. 200960 suggests that cytokinin dependence in these genotypes is determined by one pair of alleles. Certainly, the frequency of recovery of parental pheno- types in the F, populations indicates that the number of genes involved in the determination of the cytokinin requirement of P. vulgaris callus cultures must be small. The relatively low recovery of the completely cytokinin-autonomous phenotype in the F, populations is probably a product of the stringent criteria used in the classification of parental phenotypes and the variation in the growth rate of F, intermediate phenotypes. The latter phenomenon is presumably the result of the use of nonisogenic lines and may reflect either the presence of addi- tional genes influencing the level of cytokinin autonomy or genetic variation in other factors that affect the rate of growth of the callus tissue.
The biochemical differences between cytokinin-dependent and cytokinin- autonomous plant tissue cultures are not certain. Although production of endo- genous cytokinins by cytokinin-autonomous strains of soybean and tobacco tis- sue cultures has been demonstrated (DYSON and HALL 1972; EINSET and SKOOG 1973; MIURA and MILLER 1969), this result in itself does not establish whether cytokinin autonomy is associated with an increase in cytokinin biosynthesis or a decrease in the rate of conversion of endogenous cytokinin to inactive metabo- lites. I t is, of course, conceivable that the biochemical and genetic mechanisms involved in determining the cytokinin requirements of plant tissue cultures may differ in different systems. In any event, the identification of genetic differences in the cytokinin requirements of Phaseolus tissue cultures should provide useful tools for biochemical investigations of the regulation of cytokinin biosynthesis and metabolism.
We wish to thank L. E. WARREN of Doiv Chemical U.S.A. for the gift of picloram.

686 M. c. MOK et al.
LITERATURE CITED
ASHTON, N. W., N. H. GRIMSLEY and D. J. COVE, 1979 Analysis of gametophytic development in the moss, Physcomitrella patens, using auxin and cytokinin resistant mutants. Planta 144: 427435.
DYSON, W. H. and R. H. HALL, 1972 N6-(A2-Isopentenyl)adenine: Its occurrence as a free nucleoside in an autonomous strain of tobacco tissue. Plant Physiol. 50: 616-621.
EINSET, J. W. and F. SKOOG, 1973 Biosynthesis of cytokinins in cytokinin-autotrophic tobacco callus. Proc. Natl. Acad. Sci. U.S. 70: 658-660.
FOSKET, D. E. and 5. G. TORREY, 1969 Hormonal control of cell proliferation and xylem dif- ferentiation in cultured tissues of Glycine maz var. Biloxi. Plant Physiol. 44: 871-880.
GELINAS, D. and S. N. POSTLETHWAIT, 1969 IAA oxidase inhibitors from normal and mutant maize plants. Plant physiol. 44: 1553-1559.
IMBER, D. and M. TAL, 1970 Phenotypic reversion of flacca, a wilty mutant of tomato, by abscisic acid. Science 169: 592-593.
LOUIS, J. P. and B. DURAND, 1979 Studies with the dioecious angiosperm, Mercurialis annud L. (2n = 16): Correlation between genic and cytoplasmic male sterility, sex segregation, and feminizing hormones (cytokinins). Molec. gen. Genet. 165: 309-322.
MIURA, G. A. and C. 0. MILLER, 1969 Cytokinins from a variant strain of cultured soybean cells. Plant Physiol. 44: 1035-1039.
MOK, M. C. and D. W. S. MOK, 1977 Genotypic responses to auxins in tissue cultures of Phaseolus. Physiol. Plant. 40: 261-264.
MOH, M. C., D. W. S. MOK and D. J. ARMSTRONG, 1978 Differential cytokinin structure- activity relationships in Phaseolus. Plant Physiol. 61 : 72-75.
MURASKIGE, T. and F. SKOOG, 1962 A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 15 : 473-497.
PIIINNEY, B. O., 1956 Growth response of single-gene dwarf mutants in maize to gibberellic acid. Proc. Natl. Acad. Sci. U.S. 42: 185-189.
SMITH, H. H., 1972
SNEDECOR, G. W. and W. G. COCHRAN, 1967
WITHAM, F. H., 1968
ZOBEI, R. W.. 1974
Plant genetic tumors. Progr. exp. Tumor Res. 15: 138-164. Statistical Methods. Iowa State University Press,
Effect of 2,4-dichlorophenoxyacetic acid on the cytokinin requirement
Control 3f morphogenesis in the ethylene-requiring tomato mutant, dia-
Correcponding editor: R. L. PHILLIPS
Ames.
of soybean cotyledon and tobacco stem pith callus tissues Plant Physiol. 43: 1455-1457.
geotropica. Can. J. Bot. 52: 735-741.





























