Embed Size (px)
Citation preview

VIROLOGY 177,312-316 (1990)
Oligomerization of Friend Spleen Focus-Forming Virus (SFFV) env Glycoproteins
YANYAN YANG,*,’ ARINOBU TOJO,* NAOKO WATANABE,t AND HIROSHI AMANUMAt
*Department of Genetics, Institute of Medical Science, University of Tokyo, 4-6- 1 Shirokanedai Minato-ku, Tokyo 108, and tLaboratory of Gene Technology and Safety, Tsukuba Life Science Center, Institute of Physical and Chemical Research, Tsukuba, lbaraki 305, Japan
Received January 2, 1990; accepted March 15, 1990
The glycoprotein gp55 and its processed form gp65, which are encoded by the env gene of Friend spleen focus- forming virus (SFFV), have been implicated in the initiation of the murine acute erythroleukemia induced by Friend virus (FV). Analyses of these glycoproteins by chemical crosslinking and nonreducing/reducing two-dimensional electropho- resis showed that both gp55 and gp65 exist as monomer and disulfide-bonded dimer and trimer. These oligomers could be detected in various FV-infected etythroleukemia cell lines, as well as in the spleen cells of FV-induced ery-throleu- kemic mice, suggesting that oligomerization is an intrinsic feature of SFFV env glycoproteins. o 1990 Academic PWSS, IW.
INTRODUCTION
The env gene of SFFV has been identified as the only viral gene necessary for the initiation of FV-induced murine erythroleukemia (Ostertag et al., 1987, for re- view; Wolff and Ruscetti, 1988). Two env gene prod- ucts can be detected in SFFV-infected cells: the major product gp55 and its processed from gp65. Like other murine retroviral envelope glycoproteins, gp55 is syn- thesized in the rough endoplasmic reticulum (ER), where the first step of glycosylation occurs. Further processing follows during its transport to the plasma membrane via the Golgi apparatus. However, unlike the gp7Os, 5-l 0% of gp55 is expressed on the cell sur- face as gp65 which is not incorporated into virions (Ruscetti and Wolff, 1984, for review). The pathogenic mechanism of gp55 and gp65 in FV-induced erythro- leukemia is still unclear.
One possible mechanism for gp55 and gp65 to in- duce erythroblastosis is that they could interact with some cellular protein(s) involved in the proliferation and differentiation of erythroid progenitor cells. To investi- gate this possibility, we examined the possible com- plexformation of gp55 and gp65 by chemical crosslink- ing and nonreducing/reducing two-dimensional elec- trophoresis. We found that gp55 and gp65 form disulfide-bonded dimers and trimers, and that oligo- merization seems to be an intrinsic feature of SFFV env glycoproteins.
MATERIALS AND METHODS
Cell lines
Murine erythroleukemia cell lines T3Kl (Ikawa et al., 1978), F5-5 (Ikawa et a/., 1976), F4-6 (Anders et al.,
’ To whom requests for reprints should be addressed.
1985), and B8-3 (Dube et al., 1975) were induced by the polycythemia strain of FV (FVp), while TSA8 was induced by the anemia strain (FVa) (Shibuya and Mak, 1983). FVTCT originated from an FVp-induced trans- plantable reticulum cell sarcoma (Fieldsteel et al., 1969).
Mice and viral infection
The NB-tropic strain of FVp was maintained in qur laboratory by continuous passage in BALB/c mice. The 100/o (v/v) spleen cell homogenate in a-medium was prepared from mice intraperitoneally infected with FVp 10 days before, and stocked at -70”.
Male DDD mice (Fv-,?“), 4-8 weeks of age, were in- jected intraperitoneally with five-fold-diluted FV stock. Spteen cells were obtained 9 days later and prepared for protein analysis.
Antibodies
Rat anti-gp55 monoclonal antibody 7C-10 (Ab 7C- 10) was generously supplied by Dr. S. Ruscetti (Na- tional Cancer Institute, Bethesda, MD; Wolff et al., 1982; Li et al., 1987). Rabbit anti-rat IgG (Wako) was used as the second antibody for Ab 7C-10. Both of them were used at 1 :lOOO dilution.
Chemical crosslinking of proteins
Crosslinking reagent 3,3’-dithiobis (sulfosuccinimidyl propionate) (DTSSP, Pierce; Staros, 1982) was added to lo6 T3Kl cells suspended in 1 ml phosphate- buffered saline (PBS) at a final concentration of 0.5 mll/l. After incubation for 15 min on ice, the reaction was quenched by addition of 2 mM glycine (Kreis and Lo- dish, 1986). The extracts from these cells were analyzed
0042-6822/90 $3.00 CopyrIght 0 1990 by Academic Press, Inc. All rights of reproduction I” any form reserved.
312

OLIGOMERIZATION OF SFFV env GLYCOPROTEINS 313
by electrophoresis under nonreducing or reducing con- ditions
DTT ! 2 3 4 + - +
lmmunoprecipitation
Cells (1 O”/ml) were labeled with 50 $Cilml L-[35S]me- thionine (Amersham) for 2 or 18 hr and lysed with 0.1 o/o SDS buffer (0.1 o/o SDS, 1% sodium deoxycholate in 1% Triton X-100 buffer, see Electrophoresis) without any reducing reagent. The lysates were reacted with Ab 7C-10 and rabbit anti-rat IgG, and precipitated by pro- tein A-Sepharose CL-4B. The precipitated proteins were resuspended in the sample buffer with or without 50 mM reducing reagent dithiothreitol (DTT) and boiled
200KD-
116KD-
97KD-
67KD-
43KD-
FIG. 1. DTSSP-mediated crosslinking of SFFV eny glycoproteins. T3Kl cells (1 06) were treated with 0.5 mM DTSSP for 15 min at 4”.
for 5 min before electrophoresis.
Enzymatic digestions
L-[35S]Methionine-labeled gp55 monomers and complexes were recovered from the dried gel, digested with 10 pg Staphy/ococcus aureus \/8 protease per lane (Machida et al., 1984), and analyzed on a 15% acrylamide gel as described previously (Cleveland et al., 1977).
(Western blotting). The blot was then reacted with a rat anti-gp55 monoclonal antibody 7C-10, the secondary antibody rabbit anti-rat IgG, and visualized by ‘251-protein A. Lanes 1 and 2, DTSSP (-); lanes 3 and 4, DTSSP (+). Arrowheads indicate the 4 extra bands at 97, 116,170,and200kDa.
Then the extracts were treated with (+) or without DTT (-) before electrophoresis, followed by protein transfer onto a membrane
Electrophoresis
One- or two-dimensional gel electrophoresis under nonreducing or reducing conditions was performed ac- cording to the method of Phillips and Agin (1977). Pro- teins were extracted with 1% Triton X-l 00 buffer (1% Triton X-l 00, 50 mM Tris-HCI, pH 7.4, 150 mM NaCI, 20 mM EDTA, and 1 %Trasylol), treated with or without DlT before the first-dimensional electrophoresis on a 7.5% acrylamide gel. Gel strips containing the samples of interest were cut out, treated with or without DlT, and analyzed by second-dimensional electrophoresis. The L-[35S]methionine-labeled gel samples were pre- pared for Western blotting, dried, and exposed to film at -70”.
Western blotting
Western blotting was performed as previously de- scribed (Towbin et a/., 1979). The transferred mem- branes (blots) were reacted with Ab 7C-10 and rabbit anti-rat IgG, then with ‘251-protein A and exposed to film at -70”.
Quantitative analysis of gp55/gp65 monomers and oligomers
RESULTS
Since DTSSP-mediated crosslinking can be cleaved by reducing reagents such as DTT or 2-mercaptoetha- nol, the proteins of T3Kl cells were crosslinked with DTSSP and analyzed by electrophoresis under reduc- ing or nonreducing conditions, and then by Western blotting with Ab 7C-10 (Fig. 1). In addition to the bands at 55 and 65 kDa, four major extra bands at 97, 116, 170, and 200 kDa were generated by DTSSP-mediated crosslinking under nonreducing conditions and disap- peared under reducing conditions (Fig. 1, lanes 3 and 4). Surprisingly, under nonreducing conditions, the four extra bands also appeared when the proteins were not crosslinked (Fig. 1, lanes 3 and 4). This result sug- gested that gp55/gp65 may be in association with some protein(s) by disulfide bonds. Besides these, it can be seen from Fig. 1 that the nonreduced mono- meric gp55 and gp65 migrated faster than the reduced forms, suggesting that they contain at least one intra- molecular disulfide bond, which would affect their con- formations and change their mobility in electrophoresis (Pinter and Honnen, 1984). Also, the intensity of the radiolabeled bonds under nonreducing conditions was stronger than that under reducing conditions, probably due to the relatively higher affinity of Ab 7C-10 to the nonreduced SFFV env glycoproteins.
To investigate the properties of the four extra bands, proteins of T3Kl cells were analyzed by two-dimen- sional gel electrophoresis under nonreducing and re- ducing conditions, followed by Western blotting with Ab 7C-10. When both dimensions were under nonre- ducing conditions, gp55, gp65, and the four extra
The radioactivity of each gp55/gp65 band on an lz51- protein A-labeled Western blot was determined with the Ambis radioanalytic imaging system (Tack el a/., 1988) and the ratio of monomer, dimer, and trimer was calculated.
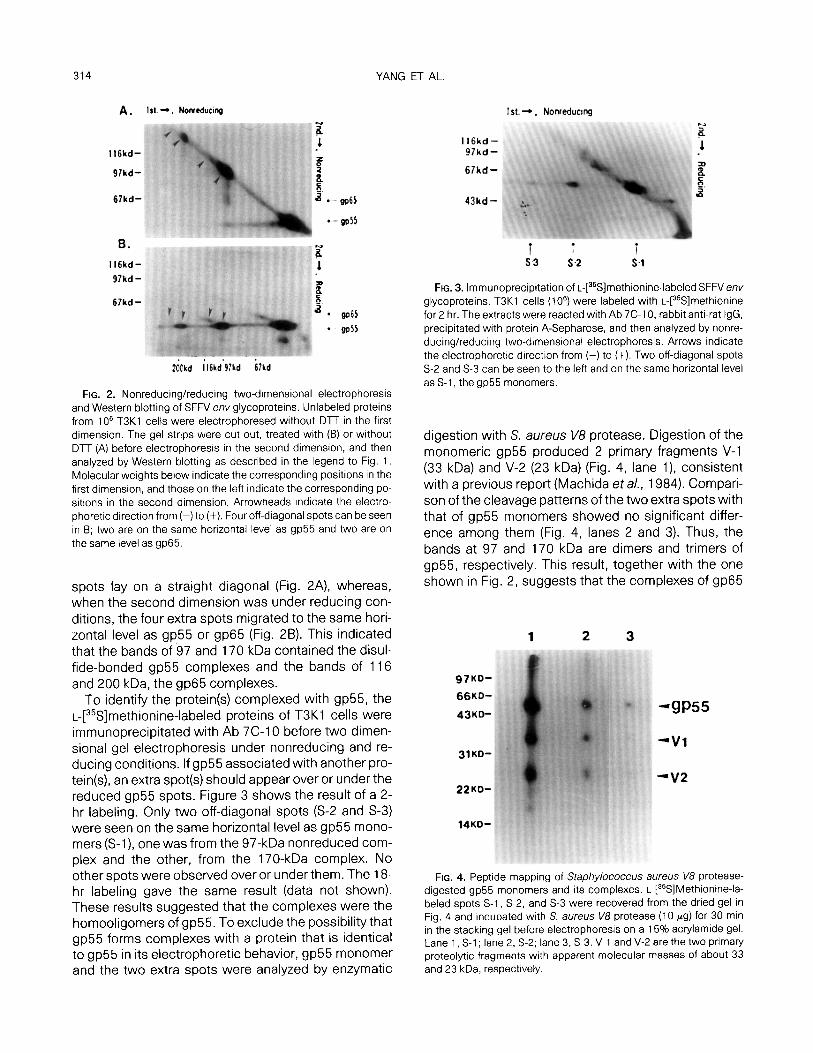
314 YANG ET AL.
A. 1st. +, Nonreducing
' SP65
- LIP55
1 st. + , Nonreducing
116kd-
97kd-
67kd-
?
%
$ f. Icl +gp65
T T T
2dOkd ilbkd’97kd k7kd
FIG. 2. Nonreducing/reducing two-dimensional electrophoresis and Western blotting of SFFV env glycoproteins. Unlabeled proteins from 1 O6 T3Kl cells were electrophoresed without Dn in the first dimension. The gel strips were cut out, treated with (B) or without DlT (A) before electrophoresis in the second dimension, and then analyzed by Western blotting as described in the legend to Fig. 1. Molecular weights below indicate the corresponding positions in the first dimension, and those on the left indicate the corresponding po- sitions in the second dimension. Arrowheads indicate the electro- phoretic direction from (-) to (+). Four off-diagonal spots can be seen in B; two are on the same horizontal level as gp55 and two are on the same level as gp65.
spots lay on a straight diagonal (Fig. 2A), whereas,
when the second dimension was under reducing con- ditions, the four extra spots migrated to the same hori- zontal level as gp55 or gp65 (Fig. 28). This indicated that the bands of 97 and 170 kDa contained the disul- fide-bonded gp55 complexes and the bands of 116 and 200 kDa, the gp65 complexes.
To identify the protein(s) complexed with gp55, the L-[35S]methionine-labeled proteins of T3Kl cells were immunoprecipitated with Ab 7C-10 before two-dimen- sional gel electrophoresis under nonreducing and re- ducing conditions. If gp55 associated with another pro- tein(s), an extra spot(s) should appear over or under the reduced gp55 spots. Figure 3 shows the result of a 2- hr labeling. Only two off-diagonal spots (S-2 and S-3) were seen on the same horizontal level as gp55 mono- mers (S-l), one was from the 97-kDa nonreduced com- plex and the other, from the 170-kDa complex. No other spots were observed over or under them. The 18- hr labeling gave the same result (data not shown). These results suggested that the complexes were the homooligomers of gp55. To exclude the possibility that gp55 forms complexes with a protein that is identical to gp55 in its electrophoretic behavior, gp55 monomer and the two extra spots were analyzed by enzymatic
s-3 s-2 S-l
FIG. 3. lmmunoprecipitation of L-[35S]methionine-labeled SFFV env glycoproteins. T3Kl cells (106) were labeled with L-[%]methionine for 2 hr. The extracts were reacted with Ab 7C-10. rabbit anti-rat IgG, precipitated with protein A-Sepharose, and then analyzed by nonre- ducing/reducing two-dimensional electrophoresis. Arrows indicate the electrophoretic direction from (-) to (+). Two off-diagonal spots S-2 and S-3 can be seen to the left and on the same horizontal level as S-l, the gp55 monomers.
digestion with S. aureus 1/8 protease. Digestion of the monomeric gp55 produced 2 primary fragments V-l (33 kDa) and V-2 (23 kDa) (Fig. 4, lane l), consistent with a previous report (Machida eta/., 1984). Compari- son of the cleavage patterns of the two extra spots with that of gp55 monomers showed no significant differ- ence among them (Fig. 4, lanes 2 and 3). Thus, the bands at 97 and 170 kDa are dimers and trimers of gp55, respectively. This result, together with the one shown in Fig. 2, suggests that the complexes of gp65
97KD-
66KD-
43KD-
31 KD-
22KD-
14KD-
+3P55
-VI
-v2
FIG. 4. Peptide mapping of Staphylococcus aureus V8 protease- digested gp55 monomers and its complexes. L@%]Methionine-la- beled spots S-l, S-2, and S-3 were recovered from the dried gel in Fig. 4 and incubated with S. aureus V8 protease (10 pg) for 30 min in the stacking gel before electrophoresis on a 15% acrylamide gel. Lane 1, S-l ; lane 2, S-2; lane 3, S-3. V-l and V-2 are the two primary proteolytic fragments with apparent molecular masses of about 33 and 23 kDa, respectively.

OLIGOMERIZATION OF SFFV em GLYCOPROTEINS 315
may also be its dimers and trimers. The ratio of mono- mer, dimer, and trimer of gp55 is 73:23:4 and that of gp65 is 57:37:6.
In addition to T3K1, the oligomeric gp55/gp65 could also be detected in several other FVp-infected cell lines such as F4-6, 68-3, FVTCT, and in the spleen cells of FV-induced erythroleukemia mice. In an FVa-infected erythroleukemia cell line TSA8, the gp55 of the anemia strain was also detected as oligomers. These results suggested that oligomerization may be an intrinsic fea- ture of SFFV env glycoproteins.
Furthermore our examination of the products of two nonpathogenic SFFV env mutants (Amanuma et al., 1989; Watanabe eta/., in press) revealed that the oligo- mers were also formed, but in a less efficient manner apparently than those of the wild-type gp55 (data not shown). This result indicated that loss of pathogenicity cannot be linked to a deficiency in oligomerization of mutant env products, although further quantitative analysis would be needed to determine whether effi- cient oligomerization of the SFFV env glycoproteins is related to leukemogenesis or not.
DISCUSSION
Previous studies on wild-type and mutant oligomeric proteins, such as the hemagglutinin of influenza virus (Gething eta/., 1986) and the G protein of vesicular sto- matitis virus (VSV) (Kreis and Lodish, 1986) have sug- gested that oligomerization is a prerequisite for their intracellular transport. As for SFFV env glycoproteins, Kilpatrick et al. (1988) have investigated the folding of SFFV envglycoproteins using sucrose density gradient centrifugation and demonstrated that gp65 formed oligomers and was transported to the cell surface, whereas gp55 was defective in oligomerization and re- mained as a monomer in the ER. On the other hand, studies of Gliniak and Kabat (1989) using nonreducing/ reducing two-dimensional electrophoresis showed that gp55 exists as both monomer and disulfide- bonded dimer. Using ‘251-mediated cell surface label- ing, they also showed that only the dimer of gp55p (the processed form of gp55) could be detected on the cell surface. However, our studies clearly showed that both gp55 and gp65 could be detected as disulfide- bonded monomer, dimer, and trimer in the whole cell lysate. This was obtained not only from the results of nonreducing/reducing two-dimensional electrophore- sis, in which iodoacematide (50 mM) was used to pre- vent the formation of any spurious disulfides, but also from the results of chemical crosslinking with DTSSP and an uncleavable crosslinking reagent DSS (disuc- cinimidyl suberate; Sayer eta/., 1987.) which crosslink proteins before lysis of cells (data not shown).
These observations, together with the fact that only gp65 but not gp55 can be detected on the cell surface, indicate that gp55 does oligomerize but is defective in transport to the cell surface. Gliniak and Kabat (1989) interpreted the retention of gp55 in ER as due to the formation of heterogeneous dimers and suggested that only one “properly folded” homodimer is compe- tent for export from the ER, since on their nonreducing/ reducing two-dimensional electrophoresis gel, the mo- bilities of dimeric gp55 seem different. However, this may result from the large amount of gp55 or the experi- mental conditions. In our opinion, oligomerization is prerequisite, but it alone may not be sufficient for the intracellular transport. Previous studies on the G pro- teins of VSV have shown that some mutant G proteins, which were identical to the wild-type G proteins in fold- ing and trimerizing, were either transported to the cell surface slowly or not at all (Doms et al., 1988). Also, the proteolysis or early glycosylation that occurs in ER may be involved in the transport of some proteins such as the gp70 of F-MuLV, whose transport is temporally linked with the proteolysis of its precursor PrENV (Srini- vas et a/., 1982). Thus it seems likely that besides oligo- merization, another additional transport signal(s) could be involved in governing the retention of gp55 in the ER and the transport of SFFV env glycoproteins. An argu- ment in favor with this view is the observation that enzy- matic analysis of gp55p and gp55”(secreted gp65) with S. aureus V8 protease showed that the C-termini of gp55p and gp55’ have undergone some modifications (Gliniak and Kabat, 1989). Investigating whether a pro- teolytic event occurs before gp55 is exported from the ER may be of interest.
It has been suggested that the gp65 on the cell sur- face is essential for SFFV-induced pathogenesis. Gli- niak and Kabat (1989) found that only dimeric gp65 could be detected on the cell surface, and the present studies showed that besides the dimer, both monomer and trimer of gp65 were detected in the whole cell ly- sates. It seems likely that after the oligomeric gp55 was exported from ER, the oligomerfell apart and gp55 was glycosylated into gp65 separately in the Golgi appara- tus. Then the gp65 was reoligomerized and the dimeric one was transported to cell surface.
Although no cellular protein associated with gp55/ gp65 was found in this study, it is still possible that the interaction of gp55/gp65 with some cellular proteins might be detected by a method other than that which we used. A recent study by Li et a/. (1990) using double immunoprecipitation with anti-gp70 Ab and anti-EPO- receptor Ab has shown that gp55 binds to the EPO re- ceptor in fibroblasts and hemopoietic cells. In the pres- ent study, several high-molecular-weight bands were detected when [35S]methionine-labeled proteins from

316 YANG ET AL.
T3Kl cells were immunoprecipited with 7C-10 Ab (Fig. 3). However further studies have not been performed to characterize them. Since each T3Kl cell expresses less than 1 X 1 O3 EPO receptors, it would be difficult to detect the association of gp55 with the EPO receptor in this cell line. It is of interest to investigate whether this association also occurs in EPO receptor-positive, FV- induced erythroleukemia cells, or in the erythroblasts of FV-infected mice.
ACKNOWLEDGMENTS
We thank Dr. S. Ruscetti for the generous gifts of anti-gp55 mono- clonal antibody 7C-10 and Dr. Takeshi Odaka for discussions and reading of the manuscript. This work was supported in part by Grants-in-Aid for Cancer Research from the Ministry of Education, Science and Culture of Japan.
REFERENCES
AMANUMA, H., WATANABE, N., MASAHIRO, N., and IKAWA, Y. (1989). Requirement of the single base insertion at the 3’ end of the env- related gene of Friend spleen focus-forming virus for pathogenic activity and its effect on localization of the glycoprotein product (gp55). /. Viral. 63(1 l), 4824-4833.
ANDERS, C. U., VON-KREY. B. T., and KOCH, G. (1985). Inhibition of proliferation and differentiation of mouse erythroleukemia cells by hydroxamic acids. Leuk. Res. 9(4), 463-469.
CLEVELAND, D. W., FISCHER, S. G., KIRSCHNER, M. W., and ~AEMMLI, U. K. (1977). Peptides mapping by limited proteolysis in sodium dodecyl sulfate and analysis by gel electrophoresis. J. Biol. Chem. 252,1102-l 106.
DOMS, R. W., RUUSALA, A., MACHAMER, C., HELENIUS, J., HELENIUS, A., and ROSE, J. K. (1988). Differential effects of mutations in three do- mains on folding, quaternary structure, and intracellular transport of vesicular stomatitis virus G protein. J. Cell Biol. 107, 89-99.
DUBE, S. K., PRAGNELL. I. B., KLUGE, N., GAEDICKE, G., STEINHEIDER, G., and OSTERTAG, W. (1975). Induction of endogenous and of spleen focus-forming viruses during dimethylsulfoxide-induced differen- tiation of mouse erythroleukemia cells transformed by spleen fo- cus-forming virus. Proc. Nat. Acacf. SC;. USA 72, 1863-l 867.
FIELDSTEEL. A. H., KURAHARA, C., and DAWSON, P. (1969). Moloney leukemia virus as a helper in retrieving Friend virus from a noninfec- tious reticulum cell sarcoma. Nature (London) 223, 1274.
GETHING, M.-J., MCCAMMON, K.. and SAMBROOK. J. (1986). Expression of wild-type and mutant forms of influenza hemagglutinin: The role of folding in intracellular transport. Cell46, 939-950.
GLINIAK, B. C., and KABAT, D. (1989). Leukemia membrane glycopro- tein encoded by Friend spleen focus-forming virus: Transport to cell surface and shedding are controlled by disulfide-bonded di- merization and by cleavage of a hydrophobic membrane anchor. J. Viral. 63(g), 3561-3568.
IKAWA, Y., INOUE. Y., AIDA, M., KAMEJI, R., SHIBADA, C., and SUGANO, H. (1976). Phenotypic variants of differentiation-inducible Friend leukemia lines: Isolation and correlation between inducibility and virus release. Comp. Leuk. Res. Proc. lnt. Symp. 7th 1975; Bibl. Haematal. (Basel) 43, 37-47.
IKAWA, Y., YOSHIDA, M., and YOSHIKURA, H. (1978). Identification of protein specific to Friend strain of spleen focus-forming virus (SFFV). Proc. Japan. Acad., Ser. 8 54, 651-656.
KILPATRICK, D. R., SRINIVAS, R. V., and COMPANY, R. W. (1988). The
spleen focus-forming virus envelope glycoprotein is defective in oligomerization. J. Biol. Chem. 264(18), 10,732-l 0,737.
KREIS. T. E., and LODISH, H. F. (1986). Oligomerization is essential for transport of vesicular stomatitis viral glycoprotein to the cell surface. Gel/46,929-937.
LI, J.-P., D’ANDREA, A. D., LODISH, H. F., and BALTIMORE, D. (1990). Activation of cell growth by binding of Friend spleen focus-forming virus gp55 glycoprotein to the erythropoietin receptor. Nature (London) 343,762-764.
LI, J.-P., BESTWICK, R. K., SPIRO, C., and KABAT, D. (1987). The mem- brane glycoprotein of Friend spleen focus-forming virus: Evidence that the cell surface component is required for pathogenesis and that it binds to a receptor. J. Viral. 61(g), 2782-2792.
MACHIDA, C. A., BESIWICK, R. K., and KABAT, D. (1984). Reduced leu- kemogenicity caused by mutations in the membrane glycoprotein gene of Rauscher spleen focus-forming virus. J. Viral. 49(2), 394- 402.
OSTERTAG, W., STOCKING, C., JOHNSON, G. R., KLUGE, N., KOLLEK, R., FRANZ, T., and HESS, N. (1987). Transforming genes and target cells of murine spleen focus-forming virus. Adv. Cancer Res. 48, 193-354.
PHILLIPS, D. R., and AGIN, P. P. (1977). Platelet plasma membrane glycoproteins. J. Biol. Chem. 252(6), 212 l-2 126.
PINTER, A., and HONNEN, W. J. (1984). Characterization of structural and immunological properties of specific domains of Friend eco- tropic and dual-tropic murine leukemia virus gp7Os. J. Viral. 49(2), 452-458.
RUSCET~, S., and WOLFF, L. (1984). Spleen focus-forming virus: Rela- tionship of an altered envelope gene to the development of a rapid erythroleukemia. Curr. Top. Microbial. Immunol. 112, 2 l-44.
SAWYER, S. T., KRANTZ. S. B., and LUNA, J. (1987). Identification of the receptor for erythropoietin by cross-linking to Friend virus-infected erythroid cells. Proc. Nat. Acad. Sci. USA 84, 3690-3694.
SHIBUYA, T., and MAK, T. W. (1983). Isolation and induction of erythro- leukemic cell lines with properties of erythroid progenitor burst- forming cell (BFU-E) and erythroid precursor cell (CFU-E). Proc. Nat. Acad. Sci. USA 80,3721-3725.
SRINIVAS, R. V., MELSEN. L. R., and COMPANS, R. W. (1982). Effects of Monensin on morphogenesis and infectivity of Friend murine leukemia virus. J. Viral. 42(3), 1067-l 075.
STAROS, I. V. (1982). N-Hydroxysulfosuccinimide active esters: bis (N-hydroxysulfosuccinimide) esters of two dicarboxylic acids are hydrophilic, membrane-impermeant, protein cross-linkers. Bio- chemistry 21,3950-3955.
TACK, L. C., WRIGHT, J. H., and GURNEY, E. G. (1988). Characterization of Simian virus 40 large T antigen by using different monoclonal antibodies: T-p53 complexes are preferentially ATPase active and adenylylated. J. Viral. 62(3), 1028-l 037.
TOWBIN, H., STAEHELIN, T., and GORDON, J. (1979). Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some application. Proc. Nat/. Acad. Sci. USA 76,4350-4354.
WATANABE, N., NISHI, M., IKAWA, Y., and AMANUMA, H. (1990). A dele- tion in the Friend spleen focus-forming virus envgene is necessary for its product (gp55) to be leukemogenic. J. Viral. in press.
WOLFF, L., KOLLER, R., and RUSCET~, S. (1982). Monoclonal antibody to spleen focus-forming virus-encoded gp52 provides a probe for the amino-terminal region of retroviral envelope proteins that con- fers dual tropism and xenotropism. 1. Viral. 43(2), 472-481.
WOLFF, L., and RUSCE~I, S. (1988). The spleen focus-forming virus (SFFV) envelope gene, when introduced into mice in the absence of other SFFV genes, induces acute erythroleukemia. J. Viral. 62(6), 2158-2163.





























