Embed Size (px)
Citation preview

ISSN 0032�9452, Journal of Ichthyology, 2010, Vol. 50, No. 11, pp. 977–984. © Pleiades Publishing, Ltd., 2010.
977
Rheoreaction of fish is an inalienable element oftheir behavior in the mobile environment. In the clas�sic understanding this is an inherent reactionexpressed as movement of fish against the current(Lyon, 1904; Arnold, 1974). Later, it was shown (Pav�lov, 1979) that this reaction is much more complexthan taxis (tropism) and the rheoreaction was under�stood as the reaction of fish to current generally—fishbehavior induced by the water flow. This approach ledto appearance of such parameter as rheoreaction typereflecting the direction of fish movement in the flow(Pavlov et al., 2010a, 2010b).
Several components of behavior are discerned inthe rheoreaction. The orientation capacity (thresholdflow rates) and locomotor capacity (swimming capac�ity, critical flow rates) are better known components ofrheoreaction. However, they allow for no possibility todetermine precisely the behavioral mechanisms ofmigration and fish distribution, especially in gradientflows. Therefore, preference of fish for different flowrates was investigated (Aslanova, 1952; Pavlov, 1979;Pavlov and Pakhorukov, 1983; Pavlov et al., 2007). Inthese studies, one component of the rheoreaction wasinvestigated, the motivational one, though the authorsdid not indicated this directly.
Recently (Pavlov et al., 1998, 2010a, 2010b; Hen�sleigh and Hendry, 1998), methods were elaboratedwhich allowed for the possibility to estimate selectionby fish of direction of movement in relation to currentdirection. We (Pavlov et al., 2007, 2020a, 2010b) intro�duced a new parameter of fish rheoreaction—the typeof rheoreaction—the preferred kind of movement of aspecimen in reaction to flow direction. It may be pos�itive (PTR), the fish moves against the flow; static(neutral) (STR), the fish resists drifting by the currentand retains its position in relation to relatively immov�able landmarks; and negative (NTR), the fish moves
downstream. PTR and NTR are dynamic rheoreac�tion type, in contrast to STR.
The purpose of the present study was elaboration ofmethodical approaches to determination of the rheo�reaction type manifested by fish. The tasks of the studycomprised elucidation of relationship of the rheoreac�tion type and the species of a specimen, its age, habitat(lotic and limnic), duration of starvation, illuminationand flow rate in the installation, as well as estimationof reproducibility of results.
MATERIAL AND METHODS
The study was performed at the Upper Volga, theIld River, the Rybinsk Reservoir, and at the fish farmAdler in 1994–2009. Fish of seven species belonging tothree families were studied: Salmonidae—the BlackSea trout (Salmo trutta labrax); Cyprinidae—roach(Rutilus rutilus), chub (Leuciscus cephalus), gudgeon(Gobio gobio), bream (Abramis brama), crucian carp(Carassius carassius); and Percidae—perch (Perca flu�viatilis).
The material was collected by means of a “wet” dipnet (with a watertight bottom) for larvae and a dragnetfor elder fish, as well as a basket trap and a lift net. Thefish were kept at the illumination 200–300 lx. Mostexperiments on cyprinids and percids were made at thesame illumination, except some experiments on theeffect of illumination. The Black Sea trout was keptunder natural illumination. The experiments on thisspecies were made in the daytime under illuminationof tens of thousands lx. Prior to experiments, thecaught fish were kept in aquaria during a day for thecontrol of manipulation stress during capture. If theloss of fish during this time exceeded 5%, such a batchof fish was not used in experiments.
The type of rheoreaction was determined in ahydrodynamic installation “fishway” according to the
On Methods of Determination of the Rheoreaction Type in FishD. S. Pavlov, V. V. Kostin, A. O. Zvezdin, and V. Yu. Ponomareva
Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences, Leninskii pr. 33, Moscow, 119071 Russia e�mail: [email protected] August 5, 2010
Abstract—Type of rheoreaction is investigated in some species of salmonids, cyprinids, and percids. Threetypes of rheoreaction are discerned: positive (the fish prefer to move against the current), static (the fish retaintheir position in the current), and negative (the fish prefer to move downstream). It is shown that the type ofrheoreaction is probabilistic and is determined by the motivational state of fish. Procedures used in investiga�tion of the rheoreaction type are discussed.
DOI: 10.1134/S0032945210110020
Key words: fish, rheoreaction, rheoreaction type, motivation, illumination, starvation, current rate, methods.

978
JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
PAVLOV et al.
previously published method (Pavlov et al., 2010a,2010b). The installation is a labyrinth subdivided intosections (the start section in the center) connected byalternating slit�like passages (Fig. 1). In the prototypeof the installation (fishways for letting fish throughdams), such passages are usually called “windows.”Several modifications of the installation are used dif�fering in size, number, and arrangement of sections.Conditions in the installation “fishway” simulate nat�ural water courses: different flow rates and turbulencelevels over the length and width of the flow, “riffles” inwindows between sections, calm zones in their cor�ners. In each section, a specimen may select any ofthree types of behavior: to remain in the section, toleave it and move downstream, or to leave it and moveupstream, against the flow. The final position of thespecimen is determined by the sum of its choices dur�ing the experiment. Generally, the “fishway” allowsfor the possibility to reveal the ratio of rheoreactiontypes in the investigated fish group.
For experiments on juveniles up to 25 mm inlength, the installation was with 21 sections (Fig. 1a)and, for larger fish, fishways with 9–15 sections wereused; the size of each of them was 45–50 by 30–45 cm(Fig. 1b). The flow rate was regulated by changingwater delivery to the installation.
Prior to the experiment, the central section, thestart one, was preliminarily isolated with nets and agroup of ten fish was placed in it (fish longer than25 mm) or 20 fish (fish shorter the 25 mm). Theiradaptation without flow continued for the first 10 minand adaptation with flow was for the next 10 min.Then the nets were removed from the stat section, theexperiment was started, and continued for 20–30 min.After this period, the number of fish in each sectionwas usually counted (exceptions are indicated in thetext). The fish that moved upstream from the start sec�tion were counted as manifesting PTR, those thatremained in the start section were counted as mani�festing STR, and those that moved downstream werecounted as manifesting NTR.
Altogether 367 experiments on 4613 fish weremade. Statistical estimation of differences in fish dis�tribution by sections of the installation χ2�test wasused, comparison of the fractions of fish abundancewith different rheoreaction types—the student’s testfor fractions.
RESULTS
Behavior in installation. The juveniles of roach upto 25 mm long in calm zones of sections usually was ingroups (2–15 specimens) and moved from section tosection one by one. The fish which ascended againstthe current frequently were in movement during all20 min of the experiment, visited more than one sec�tion, and did not form groups. Most part of the juve�niles which moved to downstream sections kept inquite areas and did not change their position anymore. Only some specimens permanently movedwithin limits of lower sections.
During passing through windows of the installa�tion, the fish swam in darts (one dart or series of darts).Downstream, the fish moved predominantly head firstor tail first.
In larger cyprinids and percids, along with solitarymovement along the fishway, movement withinschools was also recorded. In roach and bream, insome experiments schools comprised up to 90% and inperch they comprised up to 70% of the experimentalspecimens.
The effect of flow rate in windows of the installationon parameters of fish movement was investigated onyearlings of river fish: chub (body length 57–68 mm,280 specimens), gudgeon (58–66 mm, 260 speci�mens), and perch (60–71 mm, 160 specimens), as wellas on crucian carps from a pond at the age 1+ andolder (47–64 mm, 130 specimens). The principalrecorded parameter was time use by the fish with PTRfor passing against the flow through seven sections ofthe fishway. The minimum necessary was 2.8 m. This
1 2 3 4
5678
9 10 11
121314
15 16 1817
19202110 cm
32 cm
a
a b (a)
(b)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15aa
b
Fig. 1 Basic schemes of the installations “fishway” for fish juveniles of length (a) up to 25 and (b) over 25 mm: (a) defense latticesand (b) windows. (1–21) Numbers of sections, (→) flow direction, and ( ) start sections.

JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
ON METHODS OF DETERMINATION OF THE RHEOREACTION TYPE IN FISH 979
was used in calculation of the minimum necessary dateof movement of fish through the fishway.
The minimum necessary rate in most cases washigher in the chub (Fig. 2). There was no relationshipof this rate to the flow rate in windows of the fishway.By the results of two�factor analysis of variance, thelevel of significance of this factor was much greaterthan 0.05.
Ascent to the upper section of the fishway of differ�ent fish species discontinued at different flow rates inwindows of the installation. Crucian carp did notascend to the upper section at the rate 30, perch did
not at 40, and gudgeon did not at 53 cm/s. Technicalreasons prevented determination of this rate in thechub.
Analogous results were obtained in analysis of thepart of fish that ascended to the upper section of the fish�way. In case of the chub, 100% of the maximally pos�sible in this experiment number of fish reached thissection (Fig. 3). Only at the rate 63 cm/s did this valuedecrease to 65%. In gudgeon, perch, and crucian carp,the maximum manifestation of PTR was recorded in25–56% of specimens. This part did not depend on theflow rate in windows over the whole range of flow rates
1
100 20 30 40 50 60 70
Minimum speed of fish, cm/s
Flow rate in windows of the installation, cm/s
10
0.1
Fig. 2. Minimum necessary movement rate of translocation of fish along the installation depending in flow rate in its windows.(�) Chub, (�) gudgeon, (�) crucian carp, and (×) perch.
50
100 20 30 40 50 60 70Flow rate in windows of the installation, cm/s
100
90
80
70
60
40
30
20
10
The portion of fish reached the upper sections, %
Fig. 3. The portion of fish with maximum manifestation of the positive rheoreaction type depending in the flow rate in windowsof the installation; Designations as in Fig. 2.

980
JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
PAVLOV et al.
at which this rheoreaction was observed. By the resultsof two�factor analysis of variance, the effect of the flowrate was not significant for all investigated fish species.
Generally, the results demonstrate that if the fishascend to the upper section of the fishway the speed ofthis ascent does not depend on the flow rate in thewindows of the installation.
The direct influence of the flow rate on the rheore�action type was investigated on fry of the Black Seatrout (length 32–69 mm). At increasing flow rate inwindows of the fishway from 0.5 to 0.9 of the criticalflow rate for experimental fish, the portion of fish withPTR decreases and, accordingly, the number of fishwith STR and NTR increases (Fig. 4). Distribution of
fish by sections of the fishway was significantly differ�ent (p < 0.05 by χ2 test).
The rheoreaction type in juveniles of roach in onto�genesis. Late larvae and fry at ontogenesis stages D2�Hcaught in a bay of the Rybinsk Reservoir and in the IldRiver (a tributary of the reservoir) were investigated.Altogether, 90 experiments were performed and 1641fish specimens were used. Simultaneously, observa�tions on fish movements in capture sites of juvenileswere observed. The larvae and fry from the bay werecharacterized by the static type of rheoreaction (Table 1).The ratio of specimens with different rheoreactiontypes in the larvae was almost identical. There was atendency to increase the portion of fish with SRT asthey grew. In fry at stage G, the fish with SRT becomesignificantly (p < 0.05) more numerous than in larvae.Generally, when the roach from the reservoir developsfrom stage D2 to G the rheoreaction type is qualitativeidentical, STR always prevailing. At the same stages, inthe river roach, the rheoreaction type changes bothqualitatively (at different time the different rheoreac�tion types prevail) and quantitatively.
In juveniles of roach from different habitats (river,reservoir) the rheoreaction type was different in caseswhen movement of fish in water bodies differed andidentical when it did not differ (Table 1). In the secondhalf of June 2009, the downstream migration of roachwas recorded in the river and NTR prevailed in theroach; at the same time, in the reservoir, STR pre�vailed in the absence of mass translocations of juve�niles. In 2008 in the river, the downstream migrationalready stopped in the middle of June; schools of juve�niles moving against flow were noted. This wandering
70
60
50
40
30
20
10
0
The portion of fish, %
Rheoreaction typePTR STR NTR
Fig. 4. The rheoreaction type of the Black Sea trout Salmotrutta labrax (160 specimens) depending on flow rated inwindows of the installation, The critical flow rate forexperimental fish: (�)—0.5 and ( )—0.9 cm/s.
Table 1. The ratio of rheoreaction types in roach at different ontogenesis stages from the river and from the reservoir in 2008–2009
Habitat Rheoreaction type
Date of observations, age (phase/ontogenetic stage) and fish length
June 19, 2008,June 20, 2008;
D2, E13–14 mm
July 2, 2008,July 4, 2008;
E14–16 mm
June 17, 2009,June 18, 2009;
D2, E12.2–13.7 mm
July 14, 2009,July 12, 2009;
F, G17–23 mm
Sept. 13, 2009, Sept. 12, 2009;underyearlings
27–40 mm
Reservoir PTR, % 30 31 22 15 18
STR, % 55 57 67 77 28
NTR, % 15 12 11 8 54
No movements of fish in the water body –
River PTR, % 27 48 14 9 10
STR, % 59 39 30 82 11
NTR, % 14 13 56 9 79
Schools move against current Downstream migration noted
No movements of fish in the water body
–
Significance levels of differences of fish distribution over the installation by χ2 test
0.05 0.0002 0.000002 0.2 0.009
Note: In bold type are shown the values which differ significantly for juveniles from the river and from the reservoir by the Student’s test forfractions at p < 0.05; – no data on movements of fish in the water body.
JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
ON METHODS OF DETERMINATION OF THE RHEOREACTION TYPE IN FISH 981
became more intensive to the beginning of July andPRT became the prevailing rheoreaction type. In theabsence of movements, the rheoreaction type of roachfrom the reservoir did not change (STR prevailed). Inthe middle of July 2009, fish movements were not nei�ther recorded in the river nor in the bay, and the rheo�reaction type in roach was practically the same—STRprevailed. In autumn, when the juveniles usuallyretreat from the coast in the reservoir (Pavlov et al.,1999) due to abrupt decrease of water temperature inshallow places of water bodies and water courses, thedynamic rheoreaction (NTR) type prevailed both inriver roach and in the reservoir roach.
It is important that, in all cases, the direction ofmovements of fish (in relation to flow direction) in theriver (at constantly present current) and in the fishwaycoincide.
The material on the rheoreaction type of juvenilesof Atlantic salmon from the spawning river and fromits tributary after they left redds has been publishedpreviously (Pavlov et al., 2010b).
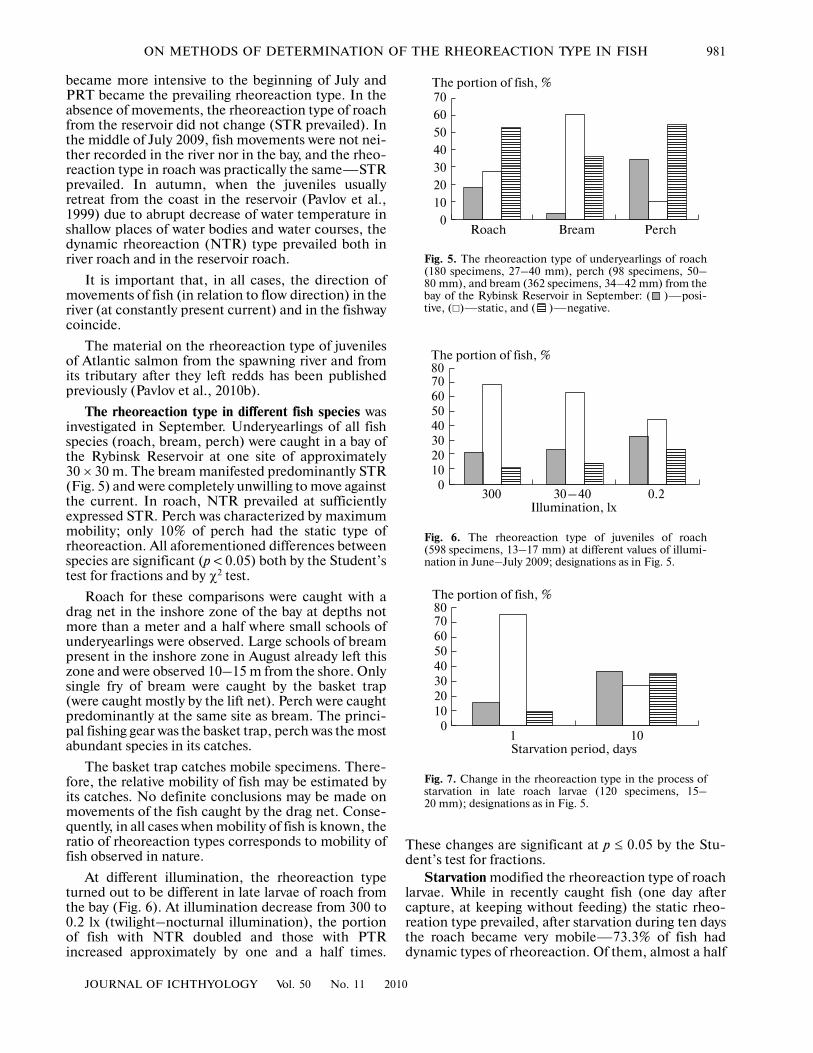
The rheoreaction type in different fish species wasinvestigated in September. Underyearlings of all fishspecies (roach, bream, perch) were caught in a bay ofthe Rybinsk Reservoir at one site of approximately30 × 30 m. The bream manifested predominantly STR(Fig. 5) and were completely unwilling to move againstthe current. In roach, NTR prevailed at sufficientlyexpressed STR. Perch was characterized by maximummobility; only 10% of perch had the static type ofrheoreaction. All aforementioned differences betweenspecies are significant (p < 0.05) both by the Student’stest for fractions and by χ2 test.
Roach for these comparisons were caught with adrag net in the inshore zone of the bay at depths notmore than a meter and a half where small schools ofunderyearlings were observed. Large schools of breampresent in the inshore zone in August already left thiszone and were observed 10–15 m from the shore. Onlysingle fry of bream were caught by the basket trap(were caught mostly by the lift net). Perch were caughtpredominantly at the same site as bream. The princi�pal fishing gear was the basket trap, perch was the mostabundant species in its catches.
The basket trap catches mobile specimens. There�fore, the relative mobility of fish may be estimated byits catches. No definite conclusions may be made onmovements of the fish caught by the drag net. Conse�quently, in all cases when mobility of fish is known, theratio of rheoreaction types corresponds to mobility offish observed in nature.
At different illumination, the rheoreaction typeturned out to be different in late larvae of roach fromthe bay (Fig. 6). At illumination decrease from 300 to0.2 lx (twilight–nocturnal illumination), the portionof fish with NTR doubled and those with PTRincreased approximately by one and a half times.
These changes are significant at p ≤ 0.05 by the Stu�dent’s test for fractions.
Starvation modified the rheoreaction type of roachlarvae. While in recently caught fish (one day aftercapture, at keeping without feeding) the static rheo�reation type prevailed, after starvation during ten daysthe roach became very mobile—73.3% of fish haddynamic types of rheoreaction. Of them, almost a half
Roach Bream Perch
70
60
50
40
30
20
10
0
The portion of fish, %
Fig. 5. The rheoreaction type of underyearlings of roach(180 specimens, 27–40 mm), perch (98 specimens, 50–80 mm), and bream (362 specimens, 34–42 mm) from thebay of the Rybinsk Reservoir in September: ( )—posi�tive, (�)—static, and ( )—negative.
70605040302010
0
80
300 30–40 0.2Illumination, lx
The portion of fish, %
Fig. 6. The rheoreaction type of juveniles of roach(598 specimens, 13–17 mm) at different values of illumi�nation in June–July 2009; designations as in Fig. 5.
70605040302010
0
80The portion of fish, %
1 10Starvation period, days
Fig. 7. Change in the rheoreaction type in the process ofstarvation in late roach larvae (120 specimens, 15–20 mm); designations as in Fig. 5.

982
JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
PAVLOV et al.
of the fish manifested PTR and the second half mani�fested NTR (Fig. 7). The differences are highly signif�icant (p < 0.001) both by the Student’s test for frac�tions and by the χ2 test.
The results on change of rheoreaction type in starv�ing Black Sea trout were published previously (Pavlovet al., 2010a).
Reproducibility of the results. The group estimationof this parameter was made both in the course of indi�vidual reproducibility and in special experiments atminimum influence of manipulation on fish. For this,after the first experiment, the larvae were carefullydriven to the start section and the second experimentwas started. Altogether, five pairs of experiments on100 specimens of roach were carried out (D2�F; length12.2–15.2 mm) from the bay of the Rybinsk Reservoir(Fig. 8). The measure of recurrency (reproducibility)of the results was the coefficient of correlation of thenumber of fish in fishway sections. The value of thesecoefficients in particular experiments fluctuatedwithin 0.930–0.976 (P < 0.001) and the coefficient ofcorrelation for all experiments was 0.99 (p < 0.001).
Such values indicate to high reproducibility of theresults obtained on one fish group.
For estimations of individual reproducibility of theresults, the fish groups in the repeated experimentswere formed of the specimens with one rheoreactiontype. The fish after primary experiments were placedinto three aquaria (for specimens with PTR, STR, andNTR) and in 1–6 h the experiments were repeated. OnBlack Sea trout, 25 initial and repeated experimentswere carried out; on larvae and fry of roach, 15 exper�iments were carried out.
The same rheoreaction type was manifested in22.2–85.7% of fish (Table 2) indicating to a low repro�ducibility of this parameter. In addition, the coeffi�cients of correlation of fish distribution by all sectionsof the installation were calculated for primary experi�ments and for distribution of specimens with eachrheoreaction type in the repeated experiments (Table 2).Analysis of these data demonstrated that, irrespectiveof the rheoreaction type of the investigated fish, theirinitial distribution is well reproduced. The coefficientsof correlation of the sum fish distribution in the initialand in all repeated experiments turned out to be iden�tical for juveniles of trout and roach—0.98 at the sig�nificance level less than 0.001, i.e., the reproducibilityof experiments on the fish group almost did notdecrease in comparison with experiments at the mini�mal interference of manipulation.
Generally, at any method of formation of thegroups for the repeated experiment reproducibility ofthe results is very high. Frequently, particular speci�mens do not repeat their selection of a section of theinstallation recorded in the previous experiment.
DISCUSSION
The rheoreaction type turned out to be anextremely labile parameter. It was both different andidentical in fish at different ontogenesis stages caughtin different water bodies. These differences and coin�
908070605040302010
0
The portion of fish, %
1 3 5 7 9 11 13 15 17 19 21Sections of the installation
Fig. 8. Distribution of juveniles of roach (100 specimens,body length 12.2–15.2 mm) by sections of the installationsin the (–) initial and in (���) repeated experiments.
Table 2. Reproducibility of the results on determination of the rheoreaction type
Primary experiments Repeated experiments, % of fish with the rheoreaction type
Correlation of fish distribution by sections of the installation in primary and in repeated experiments
rheoreaction type specimens % PTR STR NTR coefficient significance level
Black Sea trout, age 5 months, 250 specimens, 32–69 mm
PTR 45 18 22.2 26.7 51.1 0.92 <0.001
STR 70 28 5.7 64.3 30 0.92 <0.001
NTR 135 54 3.8 10.5 85.7 0.98 <0.001
Roach, late larvae, 274 specimens, 13–17 mm
PTR 67 24.5 38.8 52.2 9 0.98 <0.001
STR 175 63.9 12.6 80.5 6.9 0.99 <0.001
NTR 32 11.7 18.8 18.8 62.5 0.80 <0.001

JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
ON METHODS OF DETERMINATION OF THE RHEOREACTION TYPE IN FISH 983
cidences may be presence or absence of mass move�ments of fish—migrations and wanderings. In the riv�ers with constant current, the prevailing type of rheo�reaction type of roach at stages D2�G coincided withthe direction of movement of fish under natural condi�tions. In reservoirs where there is no constant flow, theportion of fish with STR significantly decreases duringwanderings and the portion of fish with dynamic rheo�reaction type (PTR and NTR) increases. Here, thisparameter reflects the mobility of fish. This is shownboth in the seasonal aspect (retreat of roach of thecoast in September) and in a diurnal aspect. It isknown (Pavlov et al., 1999) that the distribution of fishjuveniles in the inshore zone is different in the daytimeand at night in lakes and reservoirs. We demonstrated(Pavlov et al., 1998) that redistribution of fish in thecoastal zone occurs at illumination decreased to unitsor decimal fractions of lux. It is at such illuminationthe rheoreaction type of fish changed, the part of fishwith dynamic types increased.
The rheoreaction type was different in different fishspecies. When estimation of movements of these spe�cies in nature could be estimated (underyearlings ofperch and bream), the cause of the species differencesof the parameter of rheoreaction was the difference infish mobility.
Is it possible to state that the rheoreaction typereflects exactly motivation of fish for migration andwandering? Let us consider the results of experimentson starving fish. It is known that the motor activity offish increases (Pavlov et al., 2007) during starvation. Inaddition, prolonged starvation (ten days) is a signal ofshortage of food in a particular habitat and indicatesthat it should be changed. Thus, such prolonged star�vation creates a motivation for migration or wandering(Pavlov et al., 2010a). In the fish existing in the riverduring all their life (roach and specimens of the BlackSea trout with resident life strategy) (Pavlov et al.,2010a), the number of specimens with the static rheo�reaction type drastically decreases and the positive andnegative types manifest themselves in almost equalparts. Underyearlings of trout that selected anadro�mous life strategy mostly (75%) prefer downstreammovement, which, under natural conditions, contrib�utes to their downstream migration to the sea.
The fact that the rheoreaction type reflects themotivation for motion is also demonstrated by theexperiments on juvenile of Atlantic salmon in theperiod of their initial distribution from redds (Pavlovet al., 2010b). Having made the same migration route,the specimens from the same redds subdivide at theplace of inflow of the tributary: the fish possessing pre�dominantly PTR move against rapid current and getinto the tributary, and the fish with STR remain in theinshore zone of a slower spawning river.
Thus, the relationship of the rheoreaction typewith such factors as fish species, age, hydraulic condi�tions in the habitat, and illumination are realized viamotivation of fish.
Rheoreaction type is a stochastic parameter. Aspecimen in a certain state may manifest any of threetypes of rheoreaction but at a different probability.This is confirmed by experiments on recurrence of therheoreaction type. If the same fish group is tested, thecoefficient of correlation of primary and repeatedmeasurements was at the level 0.98–0.99. At the sametime, not all specimens manifesting this rheoreactiontype show it in the repeated testing. But distribution offish is well reproduced irrespective of their rheoreac�tion type determined in the initial experiments. Thisindicates the stochastic nature of the investigatedparameter. At first approximation, the probability ofmanifestation of a certain rheoreaction type in a spec�imen may be estimated by the ratio of the number offish with these types. For example, any of the investi�gated underyearlings of the Black Sea trout (both withPTR and with STR and NTR) (Table 3) in multipletests would demonstrate PTR in 18% of the cases, STRin 28% of the cases, and NTR in 54% of the cases.
All components of rheoreaction are intercon�nected. They all participate in any of its manifestationsas shown by investigation of dependence of the rheo�reaction type on the current rate in windows of thefishway. With the increase of flow rate, the portion offish demonstrating positive rheoreaction typedecreased. If our data are compared with the data oncritical flow rate for the investigated fish species indi�cated by Pavlov (1979), discontinuation or consider�able retardation of movement rate against current isobserved within the range 0.7–0.9 of the critical flowrate for these fish. Minimum values are recorded forthe fish preferring slowly running waters (crucial carpand perch) and maximum values are recorded ininhabitants of rapid water courses (chub and gud�geon). That is, at flow rated in windows of the fishwayover 0.5–0.6 of the critical flow rate for the investi�gated fish the results of the applied method reflect notonly the motivational component of the rheoreactionbut locomotor capacity of fish begins to play a signifi�cant role.
Estimation of motivation of fish movement in thecurrent allows for the possibility to verify behavioralmechanisms of such phenomena as fish migrations.For example, the rheoreaction type is one of thebehavioral mechanisms of fish migrations and wan�dering. The data on negative type of rheoreaction inroach during its downstream migration supplementbehavioral mechanisms of “motivated” and “exter�nally controlled” downstream migration (Pavlov et al.,2007). Due to specific behavioral reactions, fish juve�niles reach the boundary of the riverbed flow. Then thespecimens with expressed negative type of rheoreac�tion select movement downstream and, thus, realizedownstream migration. That is, realization of thedownstream migrations may take place not only dueloss of orientation or inhibition of rheoreaction bystronger behavioral reaction but also as a result ofmanifestation in the negative type of rheoreaction.

984
JOURNAL OF ICHTHYOLOGY Vol. 50 No. 11 2010
PAVLOV et al.
Thus, the negative rheoreaction type is one of “thesecond�order mechanisms of downstream migration.”
CONCLUSIONS
The present study demonstrated that the rheoreac�tion type is probabilistic; each specimen may manifestany rheoreaction type but at a different probability.This parameter is connected not so much with fishspecies, development stage, illumination, and habitatas with their behavior in water bodies—presence orabsence of movements (migrations and wanderings)significantly depending on the listed factors. Therheoreaction type may be determined in a wide rangeof flow rates in windows of the installation, from 0.1 to0.6–0.7 of critical flow rates. At higher flow rates, theresults are influenced by locomotor capacity of fish.On the whole, the method adequately reflects the pre�vailing selection by specimens of the direction ofmovement in relation to flow direction in juveniles ofvarious fish species living under different conditions ininvestigations of migratory and wandering behavior offish when the specimens select the direction of move�ment depending on the flow direction.
ACKNOWLEDGMENTS
The study is supported by the Russian Foundationfor Basic Research (08�04�00927�a), the “Biodiver�sity, Inventory, Functions, Conservation” Program ofFundamental Investigations of the Presidium of theRussian Academy of Sciences, the Federal Agency onScience and Innovations (Goskontracts02.740.11.0280, 14.740.11.0165; 16.740.11.0174), andthe “Leading Scientific Schools” Program of Presi�dent of the Russian Federation (NSh�3231.2010.4).
REFERENCES
1. N. E. Aslanova, “Experimental Study of Fish Behaviorin a Water Stream,” Dokl. VNIRO, No. 1, 13–18(1952).
2. G. P. Arnold, “Rheotropism in Fishes,” Biol. Rev.,No. 49, 515–576 (1974).
3. J. E. Hensleigh and A. P. Hendry, “RehotacticResponse of Fry from Beach�Spawning Populations ofSockeye Salmon: Evolution after Selection Is Relaxed,”Can. J. Zool., No. 76, 2186–2193 (1998).
4. E. P. Lyon, “On Rheotropism. 1. Rheotropism inFishes,” Am. J. Physiol. 12 (2), 149–161 (1904).
5. D. S. Pavlov, Biological Basis of Control of Fish Behaviorin a Water Stream (Nauka, Moscow, 1979) [in Russian].
6. D. S. Pavlov, V. V. Kostin, and V. Yu. Ponomareva,“Behavioral Differentiation of Underyerlings of theBlack Sea Salmon Salmo trutta labrax: Rheoreaction ina Year Preceding Smoltification,” Vopr. Ikhtiol. 50 (2),251–261 (2010a) [J. Ichthyol. 50 (3), 270–280(2001a)].
7. D. S. Pavlov, A. I. Lupandin, and V. V. Kostin, Migra�tions of Fish through Dams of Hydroelectric Power Plant(Nauka, Moscow, 1999) [in Russian].
8. D. S. Pavlov, A. I. Lupandin, and V. V. Kostin, Mecha�nisms of Downstream Migration of Juvenile River Fish(Nauka, Moscow, 1999) [in Russian].
9. D. S. Pavlov, I. V. Nechaev, V. V. Kostin, et al., “Hor�monal Mechanisms of Downstream Migration of Lar�val Roach Rutilus rutilus,” Vopr. Ikhtiol. 38 (2), 257–266 (1998).
10. D. S. Pavlov and A. M. Pakhorukov, Biological Basis ofFish Defense from Getting into Intake Structures (Leg. iPishch. Prom�t’, Moscow, 1983) [in Russian].
11. D. S. Pavlov, V. Yu. Ponomareva, A. E. Veselov, andV. V. Kostin, “Rheoreaction as a Mechanism of Forma�tion of Phenotypic Groups of Underyerlings of theAtlantic Salmon Salmo salar,” Vopr. Ikhtiol. 50 (4),548–553 (2010b) [J. Ichthyol. 50 (6), 483–488(2010b)].





























