Embed Size (px)
Citation preview

Organization of genetic apparatus of prokaryotes.
Mechanisms of genetic recombinations.recombinations.
Prepared by Inzhevatkina S.M., Department
of Microbiology and Virology of Russian National Research
Medical University NI Pirogov

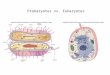
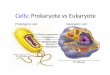
Comparison of genetic apparatus of eukaryotic and prokaryotic cells
• The most fundamental difference is that eukaryotes do have
"true" nuclei containing their DNA, whereas the genetic
material in prokaryotes is not membrane-bound.
• Prokaryotes also differ from eukaryotes in that they contain
only a single haploid chromosome in an area named the
nucleoid (exceptions – Vibrio cholerae and Brucella (exceptions
melitensis have 2 circular chromosomes), while eukaryotic
DNA is found on tightly bound and organized in several
(more than one) chromosomes.
• Consequently, any gene in prokaryotic cell is dominant.
Prokaryotic cell lacks recessive genes
• Bacterial DNA can be circular ( for most microorganisms) or
linear (rare exceptions - for Borrelia burgdorferi and several
Streptomyces species).

Comparison of genetic apparatus of
eukaryotic and prokaryotic cells
(continuation)
• Nucleoid lacks histone proteins (but histone-like proteins are
present in bacteria and presumably play the role similar to that
of histones of eukaryotes).
• Nucleolus is present in eukaryotic cell, but absent in prokaryotic
cell.
• Telomeres are present (linear DNA) in eukaryotic cell but
circular DNA of prokaryotes doesn't need telomeres.
• Plasmids are generally regarded as a prokaryotic feature and
many important genes in prokaryotes are stored on plasmids
(several eukaryotes can possess plasmids, but they are not
typical for Eucarya).
• Genetic recombination in eukaryotes is mitosis and fusion of
gametes, but genetic recombination in prokaryotes is partial,
undirectional transfers DNA.

Comparison of
eukaryotic and prokaryotic genesProkaryotes also differ from eukaryotes in the structure,
packing,density, and arrangement of their genes on the chromosome.
1. Prokaryotic genes lack introns and large non-coding regions betweeneach gene (nearly 98% of the human genome does not code forproteins or RNA or includes a gene promoter), nearly all of theprokaryote genome codes or controls something. 2. Prokaryotic genes are also expressed in groups, known as operons,instead of individually, as in eukaryotes. 3. In a prokaryotic cell, all genes in an operon (three in the case of thefamous lac operon) are transcribed on the same piece of RNA and thenmade into separate proteins, whereas if these genes were native toeukaryotes, they each would have their own promoter and be transcribedon their own strand of mRNA. This lesser degree of control over geneexpression contributes to the simplicity of the prokaryotes as comparedthe eukaryotes.

DNA StructureDNA Structure

DNA StructureDNA Structure

Expression of genetic information

Expression of genetic information

The Bacterial GenomeThe Bacterial Genome
The Bacterial Genome is
the total collection of genes
carried by a bacterium
both on its chromosome
and on its
extrachromosomal
genetic elements
(plasmids)

Structure of Bacterial Structure of Bacterial
ChromosomeChromosomeThe total length of E.coli chromosome is about 1 mm. The
bacterium itself is only several micrometers in length. The DNA
is about 1,000 times longer than bacterium and must be
condensed (supercoiling). Super coiling helps to pack the
circular molecule into a compact structure. Escherichia coli
nucleoid has 40 50 supercoiled loops but because of a nucleoid has 40 - 50 supercoiled loops but because of a
presence of a break in one segment it suffers from loss of super coiling. The protein component of the nucleoid includes
DNA gyrase and DNA topoisomerase I, which are the
enzymes responsible for maintaining supercoiled state.
The molecular weight of DNA molecule ranges from about 109 -
1010 Da. The typical bacterial chromosome contain about 4x106
base pairs, thus, it is considerably smaller than the size of
eukaryotic chromosomes.


DNA shapes
a) The DNA double helix
of most prokaryotes
has the shape of
a closed circle
b) The circular DNA strands,
already coiled in a double
helix, are twisted a second
time to produce supercoils

Enzymes in Bacterial Enzymes in Bacterial
ChromosomeChromosomeTopoisomerases are enzymes that alter
the supercoiling of double-stranded DNA.
The topoisomerases act by transiently cutting one or both strands of the DNA to relax the coil and extend the DNA molecule.relax the coil and extend the DNA molecule.
Because bacterial topoisomerases are essential and unique, they are targets of antibiotics ( DNA-gyrase or topoisomerase2 is target for fluoroquinolones).

Electron micrograph of a DNA ofMicrococcus lysodeikticus

Replication of DNAReplication of DNA
DNA replication of bacteria is rapid and continues
throughout the life cycle of cell division, in contrast to replication of DNA by eukaryotic cells, which takes place during one distinct stage of cell cycle. The replication of DNA within the circular bacterial chromosome begins at a single specific site on the bacterial chromosome, termed oriC for the origin of replication. The DNA is termed oriC for the origin of replication. The DNA is unwound, and the double-stranded DNA is separated into two single-stranded templates, and synthesis proceeds from this point in the opposite directions. The bidirectional replication mechanism requires the two sites of presence of the new DNA. Each of these two sites, termed a replication fork, comprise a complex of proteins that incorporate complementary nucleotides, accurately copying the template of the parent cell.

DNA ReplicationDNA Replication
Semiconservative replicationof DNA
The replication fork of DNA
showing the synthesis of two
progeny strands. Each copy
contain one new and one old contain one new and one old
strand.
Replicon is a part of the genome
that contains an origin site and is
replicated as a whole unit. Closed
DNA circles contain necessary
information for their own replication.
Bacterial chromosome is called
replicon

Replication of DNAReplication of DNA
An autoradiograph of a replicating E.coli chromosome;
about one-third of the chromosome has been replicated

Replication of DNAReplication of DNA (continuation)
When the replication forks meet at approximately the
opposite side of the circular DNA duplex, each strand has been copied into double-stranded helix, which can be partitioned into daughter cells.
The replication of this enormous molecule is very rapid; replication forks proceed at a rate of 1,000 base pairs per second, resulting in replication of the total 1-mm-long second, resulting in replication of the total 1-mm-long chromosome in about 40 min. This fact doesn’t explain how certain rapidly growing bacteria can divide in 20 min (S.aureus and E.coli). To take advantage of optimal growing conditions, DNA replication is reinitiated before one round of replication is completed. The genomes partitioned into a new daughter cells will already replicated, enabling them to divide again more quickly. A concomitant effect is that in these rapidly dividing cells, genes near oriC will present in more than one copy.

Replication of circular DNAReplication of circular DNA


Replication of circular DNAReplication of circular DNA


Replication of circular DNAReplication of circular DNA
Mechanism used
by some bacterial
viruses, plasmids
and in chromosomal
transfer

Elements and functions of
prokaryotic genome
The primary function of the prokaryotic genome is to
store its hereditary information.
Elements of prokaryotic genome:
The bacterial chromosome • The bacterial chromosome
• Transposable elements
• Prophages
• Plasmids
In some prokaryotes genetic material can be
transformed from one m/o to another m/o.

Transposable elementsTransposable elementsTransposons (transposable elements or "jumping
genes" are small pieces of DNA that encode enzymes that transpose the transposon, that is, move it from one DNA location to another, either on the same molecule of DNA or on a different molecule. Transposons may be found as part of a bacterium's nucleoid (conjugative transposons) or in plasmids and are usually between one and twelve genes long. A transposon contains a number of and twelve genes long. A transposon contains a number of genes, coding for antibiotic resistance or other traits, flanked at both ends by insertion sequences coding for an enzyme called transposase. Transposase is the enzyme that catalyzes the cutting and resealing of the DNA during transposition. Thus, such transposons are able to cut themselves out of a bacterial nucleoid or a plasmid and insert themselves into another nucleoid or plasmid and contribute in the transmission of antibiotic resistance among a population of bacteria.

Transposable elementsTransposable elements
Insertion sequences are the smallest and simplest
“jumping genes”

Transposable elementsTransposable elements

Transposable elementsTransposable elements
Functions
1. Cause deletions and inversions of DNA sequences (internal or biological mutagenic agents)
2. Insert into genes and inactivate those genes2. Insert into genes and inactivate those genes
3. Spread of antibiotic resistance genes
4. Mobile genetic elements are responsible for the major part of genetic variability in natural bacterial populations
5. Transposons may enter other genera of bacteria during transfer of plasmids or via transducing phage

Insertion Sequences (Is)
The simplest transposons, insertion sequences (Is), are short sequences of DNA bounded at both ends by identical sequences of nucleotides in reverse orientation (inverted repeats). Insertion sequences can insert within a repeats). Insertion sequences can insert within a gene and cause a rearrangement mutation of the genetic material. If the sequence carries a stop codon, it may block transcription of the DNA during protein synthesis. It may also encourage the movement of drug-resistance genes between plasmids and chromosomes.

Pathogenicity islands
Some bacterial species are efficient at causing disease in higher organisms because they possess specific genes for pathogenic determinants.
These genes are often clustered together in the DNA and are referred to as pathogenicity
islands. These genes encode adhesins, invasins, exotoxins…
Pathogenicity islands are a special class of
mobile genetic elements containing groups of coordinately controlled virulence genes, often with IS, direct repeat sequences at their ends.

Pathogenicity islands

PlasmidsPlasmids
Plasmids are a small extrachromosomal circular molecules, but some are linear. They are double-stranded DNA molecule. Plasmids are variable in size.Size ranges from 1 to 2 kbp up to 120 kbp or more ( Bacterial chromosome ranges from 580 kbp up to 5220 kbp).The number ranging from 1-1000 copies/cell. Plasmids do not carry genetic material which is essential to the growth of an organism, although they contain features that may enhance the survivability of the m/o in a particular environment.



Structure of PlasmidsStructure of Plasmids
The plasmids typically have three important elements:
1. An origin of replication
2. A selectable marker gene (e.g. resistance to ampicillin)
3. A cloning site (a place to insert foreign DNAs)
Plasmid is (by definition) an extrachromosomal element,
thus it cannot make use of any origin of DNA replication in a thus it cannot make use of any origin of DNA replication in a
chromosome. DNA synthesis within (i.e. copying of) a
plasmid depends on its having an origin of DNA synthesis of
its own.
If a plasmid couldn't be copied, it would be rapidly diluted out in a population of dividing cells because it
couldn't be passed on to daughter cells.

Classification of Plasmids by
Transfer Properties1. Conjugative plasmidsConjugative plasmids are those that mediated conjugation. These plasmids are usually large and have all the genes necessary for autonomous replication and for transfer of DNA to a recipient (e.g. genes for sex pilus).pilus).
2. Non-conjugative plasmidsNon-conjugative plasmids are those that cannot mediate conjugation. They are usually smaller than conjugative plasmids and they lack one or more of the genes needed for transfer of DNA. A non-conjugative plasmid can be transferred by conjugation if the cell also harbors a conjugative plasmid.

Classification of Plasmids by
Compatibility Properties
1. Compatible plasmids are categorized into different groups and can coexist in one bacterial cell simultaneously.
2. Incompatible plasmids are unable to 2. Incompatible plasmids are unable to coexist stably in the same bacterial cell because they are dependent on the same replication and maintenance functions.

Classification of Plasmids by
Phenotypic EffectsPlasmids made their presence by conferring
phenotypes of cell harboring them.
• F - fertility factor
• R - antibiotic resistance
• Col colicin production• Col - colicin production
• Virulence plasmids:
• Ent - enterotoxin production
• Hly - hemolysin production
• CFA-I; CFA-II - adhesin production



Plasmids can carry virulence genes
Plasmids may encode toxins and other proteins that increase the virulence of microorganisms:
in Shigella spp. adherence factors necessary for mucosal invasion;
in Bacillus anthracis both capsule (on one plasmid) and edema factor, lethal factor and protective antigen (on another plasmid);
the virulent enterotoxinogenic strains of E.coli that cause diarrhea produce one of two different types of plasmid-encoded enterotoxin (Ent plasmid);
in Staphylococcus aureus both an enterotoxin and a number of enzymes involved in bacterial virulence (hemolysin, fibrinolysin) are encoded by plasmid genes (Hly plasmid).

BacteriophagesBacteriophages• Bacteriophages are viruses that infect bacteria.
• Virulent bacteriophage is a bacteriophage which always causes the lytic cycle, resulting in a death of a cell and production of new phage particles
• Temperate (or lysogenic) bacteriophage is a bacteriophage whose DNA integrates into bacterial chromosome. chromosome.
• Replication of phages nucleic acid is linked to replication of host cell DNA.
• Temperate bacteriophage doesn’t cause the lyses of the host cell and production new phage particles.
• The integrative phage’s DNA is called prophage
• A bacterial cell with prophage is called lysogenic cell

Historical classification of
bacteriophages
History: Discovery and investigation of phages had been induced
by problems in food industry (milk products).
Phages cause lysis of lactic acid bacteria.

Main Families of Bacteriophages(more than 2000 species of phages)

Schemes and electron photograph
of bacteriophages


The life cycle of bacteriophages

BacteriophagesBacteriophagesLytic cycle
Lysogenic cycle

BacteriophagesBacteriophages• Lysogenic (prophage) conversion is a state
when bacterial cell exhibit new properties, that are coded by the prophage genes.
• Prophage presence may increase bacterial virulence
Examples: • Examples:
- Toxigenic (tox+) and nontoxigenic strains of Corynebacterium diphtheriae
- Production of erythrogenic toxin by Streptococcus pyogenes (scarlet fever)
- Production botulinum toxin of Clostridium botulinum

The mechanisms by which genetic information
is changed and exchanged among bacteria
• Change: Mutation
• Exchange: Genetic recombination
Exchange is most efficient between cells of the same species
These processes result in progeny with phenotypic characteristics that may differ from those of the parent, e.g., acquisition of new characteristics (antigens, toxins, antibiotic resistance, etc.).

Genetic VariationGenetic Variation
Mutations are stable hereditary changes in the coding sequence of DNAOccur spontaneously (result of errors in the DNA replication process) or are induced by different chemical or physical factors (mutagens)
Various kinds of mutations

Genetic VariationGenetic Variation
Genetic recombination at the molecular level is the process by which DNA from a donor cell and DNA from a recipient cell combine to yield a new genome containing information from both genome containing information from both sources
Molecular Mechanisms of Recombination:
1. Homologous (legitimate)
2. Nonhomologous (illegitimate)

Genetic VariationGenetic Variation• Homologous recombination occurs between
closely related DNA sequences and generally substitutes one sequence to another.
The process requires a set of enzymes produced by the group “rec” genes, and homology in 100-200 nucleotide pairs.
• Nonhomologous recombination occurs between dissimilar DNA sequences and generally produces insertions or deletions or both.
This process usually requires specialized (site-specific) recombination enzymes, such as those produced by many transposons and lysogenic bacteriophages.

Gene transfer. Integrons
Integrons are mobile genetic
elements that are able
to use site-specific
recombination to acquire to acquire
new genes in “cassette-like”
fashion (independent
open reading frames) and
express them in a coordinated
manner.

Gene TransferGene Transfer
• The transfer of genetic material between
prokaryotes is called horizontal (or lateral) gene transfer.
• It takes place by one of three ways:
1.Transformation
2.Conjugation
3.Transduction

Gene TransferGene Transfer
For prokaryotic cells recombination For prokaryotic cells recombination plays more important role than mutation in plays more important role than mutation in the process of changing in DNA.the process of changing in DNA.
Recombination give material for Recombination give material for evolution of the bacteria.evolution of the bacteria.
Gene transfer is oneGene transfer is one--way process:way process: a a piece of genetic material (the exogenote) piece of genetic material (the exogenote) is donated to the chromosome of a is donated to the chromosome of a recipient cell (the endogenote) and recipient cell (the endogenote) and integrated into it. Such forms temporarily integrated into it. Such forms temporarily diploid which called diploid which called merozygote.merozygote.

TransformationTransformation is gene transfer resulting from the uptake by a recipient cell of naked DNA from a donor
cell. Transformation is typical only for certain bacteria (e.g., Streptococcus pneumoniae, Haemophilus
influenzae, Bacillus spp., Neisseria spp.).

Gene TransferGene Transfer
Transformationwas the first mechanism of genetic transfer to be discovered in bacteria.
Was discovered by Griffits in 1928 during the experiments with Streptococcus pneumoniae. Later Avery, MacLeod and McCarty identified the DNA as the transforming agent and as the hereditary genetic material
Transformation is the process by which bacteria take up fragment of free DNA and incorporate this molecule into recipient chromosome in a heritable form.

Classical series of experiments by
Frederick Griffith

Factors affecting transformation
a. DNA size state Double stranded DNA of at least 5 X 105 daltons works best. Thus, transformation is sensitive to nucleases in the environment.
b. Competence of the recipient develops during the end of logarithmic growth. of logarithmic growth.
Some bacteria are able to take up DNA naturally. At this stage the bacteria are said to be competent. Other bacteria are not able to take up DNA naturally (E.coli). However, in these bacteria competence can be induced in vitro by electroporation ( the use of high-voltage pulses) or treatment with chemicals (e.g. CaCl2).

Steps in transformation
Uptake of DNA by Gram+ and Gram- bacteria differs. In Gram +
bacteria the DNA is taken up as a single stranded molecule and the complementary strand is made in the recipient. In contrast, Gram-bacteria take up double stranded DNA.
Legitimate/Homologous/General RecombinationAfter the donor DNA is taken up, a reciprocal recombination event occurs between the chromosome and the donor DNA. This recombination requires homology between the donor DNA and the chromosome and results in the substitution of DNA between the recipient and the donor as illustrated in scheme.

Steps in transformation (continuation)
Recombination requires the bacterial recombination genes (recA, B and C) and homology between the DNA's involved. This type of recombination is called legitimate or homologous or general recombination.Because of the requirement for homology between the donor and host DNA, only DNA from closely related bacteria would be expected from closely related bacteria would be expected to successfully transform, although in rare instances gene transfer between distantly related bacteria has been shown to occur.
Significance
Transformation occurs in nature and it can lead to increased virulence. In addition transformation is widely used in recombinant DNA technology.

Transduction
Transduction was discovered by Zinder and Lederberg
in 1951-1953.
Transduction is the transfer of genetic information from a donor to a recipient by way of a bacteriophage.The phage coat protects the DNA in the environment so that
transduction, unlike transformation, is not affected by transduction, unlike transformation, is not affected by
nucleases in the environment. Not all phages can mediate
transduction. In most cases gene transfer is between members
of the same bacterial species. However, if a particular phage
has a wide host range then transfer between species can
occur. The ability of a phage to mediated transduction is
related to the life cycle of the phage.

The ability of a phage to mediated transduction is related to the life cycle of the phage.

Types of transduction
• Generalized– any gene of the host bacterium can be transferred and doesn’t
require lysogeny (Salmonella enterica – P-22)
• Specialized (restricted)– only specific genes near the attachment sites of a lysogenic
phage in the host chromosome can be transferred (E.coli- λ)
• Abortive– the transferred DNA is not integrated but often is able to
temporary survive and express. The fragment inherits linearly and lost in progeny

Generalized transductionAny gene of the host bacterium can be transferred. Phages
that mediate generalized transduction generally breakdown host DNA into smaller pieces and package their DNA into the phage particle by a "head-full" mechanism. Occasionally one of the pieces of host DNA is randomly packaged into a phage coat. Thus, any donor gene can be potentially transferred but only enough DNA as can fit into a phage head can be transferred. If a recipient cell is infected by a phage that contains donor DNA, donor DNA enters the recipient. In the recipient a generalized recombination event can occur which substitutes the
•
donor DNA and recipient DNA.

Specialized transductionSpecialized (restricted) transduction is transduction
in which only certain donor genes can be transferred to the recipient. Different phages may transfer different genes but an individual phage can only transfer certain genes. Specialized transduction is mediated by lysogenic or temperate phage and the genes that get transferred will depend on where the prophage has inserted in the chromosome. chromosome.
During excision of the prophage, occasionally an error occurs where some of the host DNA is excised with the phage DNA. Only host DNA on either side of where the prophage has inserted can be transferred (i.e. specialized transduction). After replication and release of phage and infection of a recipient, lysogenization of recipient can occur resulting in the stable transfer of donor genes. The recipient will now have two copies of the gene(s) that were transferred. Legitimate recombination between the donor and recipient genes is also possible.

Specialized transduction
• only specific genes near the attachment sites of a lysogenic phage in the host
gal bio
galbio
gal bio
galbio
gal
bio
gal bio
gal bio
gal
bio
bio
phage in the host chromosome can be transferred
gal biogal bio

TransductionTransduction
The Bacterial Genome

ConjugationConjugation is the transfer of genetic material
between two living bacteria that are in physical
contact with one another. Was discovered in 1946 by
Lederberg and Tatum.
In 1952 Hayes demonstrated that the gene transfer
was polar. Polarity is mediated by plasmid, known as F-was polar. Polarity is mediated by plasmid, known as F
factor.

ConjugationDonorThe ability of a bacterium to be a donor is a consequence of the presence in the cell of an extra piece of DNA called the F factor or fertility factor or sex factor. The F factor is a circular piece of DNA that can replicate autonomously in the cell; it is an independent replicon. Extrachromosomal pieces of DNA that can replicate autonomously are given the general name of plasmids. The F factor has genes on it that general name of plasmids. The F factor has genes on it that are needed for its replication and for its ability to transfer DNA to a recipient. One of the things the F factor codes for is the ability to produce a sex pilus (F pilus) on the surface of the bacterium. This pilus is important in the conjugation process. The F factor is not the only plasmid that can mediated conjugation but it is generally used as the model.
Recipient The ability to act as a recipient is a consequence of the lack of the F factor.

IS2tra
F
IS3
IS3
γδ
F-plasmid
FF--plasmidplasmid
The Bacterial Genome
oriT
oriV
F
100 kb
plasmid

Physiological states of the F factora. Autonomous (F+) In this state the F factor carries only those genes necessary for its replication and for DNA transfer. There are no chromosomal genes associated with the F factor in F+ strains. In crosses of the type F+ x F- the F- becomes F+ in 100% of cases, while F+ remains F+. Thus, the F factor is infectious. No bacterial genes are transferred.
b. Integrated (High frequency of recombination (Hfr) In this state the F factor has integrated into the bacterial chromosome via a recombination event. In crosses of the type Hfr x F- the F- rarely becomes Hfr and Hfr remains Hfr. In addition, there is a high frequency of transfer of donor remains Hfr. In addition, there is a high frequency of transfer of donor chromosomal genes.
c. Autonomous with chromosomal genes (F') In this state the F factor is autonomous but it now carries some chromosomal genes. F' (F prime) factors are produced by excision of the F factor from an Hfr. Occasionally, when the F factor is excising from the Hfr chromosome, donor genes on either side of the F factor can be excised with the F factor generating an F'. F' factors are named depending on the chromosomal genes that they carry. In crosses of the type F' X F- the F- becomes F' while F' remains F'. In addition there is high frequency of transfer of those chromosomal genes on the F‘.

Donor types
1. F+ cells with free F-plasmid:
F+ x F- F+
2. Hfr cells with integrated plasmid:
Hfr x F- : mating leads to
acquisition of a few new genes by acquisition of a few new genes by
recipient , which usually doesn’t
become Hfr
3. F’ free F-plasmid with a portion of
chromosomal genes
F’ x F- F’

F+ x F- crosses
• 1) Pair formation The tip of the sex pilus comes in contact with the recipient and a conjugation bridge is formed between the two cells. This bridge as a tube ensures that DNA transfer from the donor to the recipient. Thus, the DNA is protected from environmental nucleases. The mating pairs can be separated by shear forces and conjugation can be interrupted. Consequently, the mating pairs remain interrupted. Consequently, the mating pairs remain associated for only a short time.
• 2) DNA transfer The plasmid DNA is nicked at a specific site called the origin of transfer and is replicated by a rolling circle mechanism. A single strand of DNA passes through the conjugation bridge and enters the recipient where the second strand is replicated.
• 3) This process explains the characteristics of F+ x F-crosses. The recipient becomes F+, the donor remains F+. Indeed, there is no transfer of donor chromosomal genes.


ConjugationF+ x F-
F+ F- F+ F-
F+ F+F+ F+

Formation of Hfr
F+ F+ Hfr
Hfr F’

Hfr x F- crosses• 1) Pair Formation• 2) DNA transfer
The DNA is nicked at the origin of transfer and is replicated by a rolling circle mechanism. But the DNA that is transferred first is the chromosome. Depending upon where in the chromosome the F factor has integrated and in what orientation, different chromosomal genes will be transferred at different times. However, the relative order and distances of the genes will always remain the same. Only when the entire chromosome is transferred will the F factor be transferred.Since shearing forces separate the mating pairs it is rare that the entire Since shearing forces separate the mating pairs it is rare that the entire chromosome will be transferred. Thus, usually the recipient does not receive the F factor in a Hfr x F- cross.
• 3) Legitimate recombination Recombination between the transferred DNA and the chromosome results in the exchange of genetic material between the donor and recipient.
• 4) This mechanism explains the characteristics of Hfr x F- crosses. The recipient remains F-, the donor remains Hfr and there is a high frequency of transfer of donor chromosomal genes.

Conjugation
Hfr x F-
Hfr F- Hfr F-
Hfr F-Hfr F-

F' x F- crosses
• 1) Pair formation
• 2) DNA transfer This process is similar to F+ x F- crosses. However, since the F' has some chromosomal genes on it these will also be transferred.genes on it these will also be transferred.
• 3) Homologous recombination is not necessary although it may occur.
• 4) This mechanism explains the characteristics of F' x F- crosses. The F- becomes F', the F' remains F' and the is high frequency transfer of donor genes on the F’.

Conjugation
F’ x F-
F’ F’F’ F’
F’ F- F’ F-

Gene TransferGene TransferCell-to-cell transfer of a conjugative plasmid
Integration of conjugative plasmid into
The Bacterial Genome
plasmid into the bacterial chromosome an Hfr
The order of genes on the bacterial chromosome can be determined by the time of entry of the genes into a recipient cell

Significance of conjugationAmong the G- bacteria this is the major way
that bacterial genes are transferred. Transfer can occur between different species of bacteria. Transfer of multiple antibiotic resistance by conjugation has become a major problem in the treatment of certain bacterial diseases. Since the recipient cell becomes a donor after transfer of a recipient cell becomes a donor after transfer of a plasmid it is easy to see why an antibiotic resistance gene carried on a plasmid can quickly convert a sensitive population of cells to a resistant one.
G+ bacteria also have plasmids that carry multiple antibiotic resistance genes, in some cases these plasmids are transferred by conjugation while in others they are transferred by transduction.

Mapping of bacteria
• In genetic map
distances
between loci are
measured in
Duration of period before
interruption of mating
measured in
number of
minutes
required for
transfer in
conjugation

Gene TransferGene Transfer
Conjugation is very useful for
genetic mapping of bacteria
The Bacterial Genome
A circular genetic map of E.coli K12
with the location of selected genes.
The map is divided into 100 minutes

Conjugation
The Bacterial Genome

Gene Transfer

Genetic EngineeringGenetic Engineering
• Transformation, conjugation and transduction
are natural recombination processes.
• The artificial, laboratory-controlled DNA
recombination of bacteria is known as gene
manipulation, also called genetic engineering or
recombinant DNA technology.
• Specified DNA fragments can be isolated and
amplified, and their genes can be expressed at
high levels.

Genetic Engineering – a combination of methods which allows
to conduct artificial recombination of DNA and produce chimerical
molecules, non-typical for nature
Genetic EngineeringGenetic Engineering

1. The production of hormones (insulin, human
growth hormone, etc.)
2. The production of cytokines (interferons,
Medical Significance of Medical Significance of
Genetic EngineeringGenetic Engineering
interleukin-2, tumor necrosis factors, etc.)
3. Recombinant vaccines (hepatitis B vaccine,
papillomavirus vaccine, vaccine against Lyme
disease)
4. Genetically engineered antigens which are
used in diagnostic purposes.

The basic tools of Genetic
Engineering:
1. Restriction enzymes which
are used to cleave DNA at
defined sequences
2. Cloning vectors which can be
used to deliver the DNA
Genetic EngineeringGenetic Engineering
used to deliver the DNA
sequences into receptive
bacteria
3. DNA – ligase – the enzyme
that links the fragment to the
cloning vector

Types of vectors:
1. Plasmids (pUC, pBR322, pGEM) are used for
DNA fragments up to 20Kb
2. Bacteriophages (λ, T7) are used for larger
Genetic EngineeringGenetic Engineering
2. Bacteriophages (λ, T7) are used for larger
fragments up to 25Kb
3. Cosmid (combination of plasmid and phage
genes, pJC720) for fragments up to 45Kb
4. Viruses (smallpox vaccine)

The specific properties of plasmid cloning vectors:
1. Small size to be easy inserted into bacteria
2. High number of copies to be easily purified in sufficient
quantities
3. Ability to replicate within a host cell
4. Selectable traits (resistance to an antibiotic)
5. One or few sites for restriction endonucleases which cut
Genetic EngineeringGenetic Engineering
5. One or few sites for restriction endonucleases which cut
DNA and allow the insertion of foreign DNA

Steps in Cloning a Gene:
• Isolation a DNA to be cloned:
a. short genes can be synthesized
chemically, or
b. use a restriction enzyme to
generate necessary fragments of
DNA from prokaryotic genome, or
c. use mRNA of eukaryotes and
Genetic EngineeringGenetic Engineering
c. use mRNA of eukaryotes and
reverse transcriptase for synthesis of
gene sequence (elimination of introns
of eukaryotic DNA)
• Generate a recombinant molecule by
inserting DNA fragments into a
cloning vector
• Introduce recombinant molecule into
new host
• Select bacterial clones carrying
specific genes

Principles of laboratory diagnosis
of infectious diseases

Polymerase Chain Reaction
(PCR)
Polymerase chain reaction (PCR) is an in vitromethod for enzymatic synthesis of specific DNA sequences, using two oligonucleotide primers that hybridize to opposite strands and flanks the region of interest in the target DNA. Following a series of repetitive interest in the target DNA. Following a series of repetitive cycle that involve template denaturation, primer annealing, and the extension of annealed primers by Taqpolymerase, there is an exponential accumulation of the specific target DNA.
Taq polymerase is thermostable enzyme, which is isolated from the bacteria Thermus aquaticus. Due the stability of the enzyme, fresh enzyme no longer had to be added after each amplification cycle.

Procedure of PCRThe PCR requires two synthetic oligonucleotide primers that are
complementary to the region on opposite strands of a target piece of DNA, a target sequence in a DNA sample that occurs between the pair of primers, a termostable DNA polymerase and the four deoxyribonucleotides. The procedure, performed in termocycle, usually consists of three basic steps and is completed within minutes.
The first step is denaturation, which increases the temperature within the sample vial to about 94-96°C, causing the double-stranded d DNA within the sample to separate into two pieces.
The second step is renaturation (annealing). The step is completed by dropping the temperature within the thermocycler to about 50-55°C, which allows primers to anneal with the complementary sequences in the source of DNA.
The third (final) step (extension, elongation) is the synthesis portion on the reaction, wherein the temperature is raised to about 72-74°C, the optimum temperature for the catalytic functioning of Taq DNA polymerase. Target DNA is extended, replicating to the form additional copies of the target DNA.
Within 30 cycles, over a million copies of the original target DNA can be reproduced.

Three Basic Steps of the PCR Cycle



Significance of PCR 1. PCR allows to obtain the results faster in comparison with
classical methods of cultivation and identification of m/o.
2. Application of PCR to infectious diseases focused on m/o that were impossible or slow to grow in culture, were difficult to cultivate, or posed significant health hazards using standard recovery techniques (Mycobacterium tuberculosis, Helicobacter pylori, Chlamydia trachomatis, and HIV).
3. Clinicians use the PCR method to monitor the effect of 3. Clinicians use the PCR method to monitor the effect of antimicrobial therapy.
4. PCR is used for retrospective analysis of samples from outbreaks or deaths caused by previously unidentified etiological agent. Sometimes the diagnosis is absolutely changed retrospectively.
5. Several diagnostic techniques needed very rare and expensive reagents for the ability to perform the methods. PCR has provided a solution to this problem.
6. PCR is less stressful to the patient, because serial serum samples are not required and results can be obtained in less than a day.

Reverse Transcriplase
Polymerase Chain Reaction
(RT-PCR)
A technique similar to conventional PCR, except that the starting material is RNA rather than DNA. Because of this, a RNA rather than DNA. Because of this, a DNA copy first must be made from RNA, using an enzyme known as reverse transcriptase. Once a copy of DNA is made, the PCR proceeds as usual.

Scheme of RT-PCR

Quantitative, or Real-Time PCRQuantitative, or real-time, PCR is standard PCR with the
advantage of detecting the amount of DNA formed after each
cycle with either dyes or fluorescently-tagged oligonucleotide
probes. Real-time PCR results can be obtained faster and with
less variability than standard PCR due to sensitive fluorescent
chemistry and elimination of post-PCR detection procedures.
All real-time PCR instruments combine a thermal cycler to All real-time PCR instruments combine a thermal cycler to
drive DNA amplification, an optical system to excite
fluorophores and capture emitted fluorescence from the
detection chemistry, and specialized software to collect and
analyze the quantitative data generated. The only difference
between real-time and regular PCR machines is the
fluorescence detection system.
Real-time PCR is a useful for the investigation of gene
expression, viral load, pathogen detection, etc.


SYBR Green ChemistrySYBR Green, a non-specific chemistry, is the
simplest and least costly approaches to real-time PCR
that utilizes DNA-binding fluorophores for nonspecific
detection of target DNA sequences. In this type of
chemistry, the sensitivity and specificity of the assay is
determined only by the primers. SYBR Green is a determined only by the primers. SYBR Green is a
naturally fluorescing dye that can intercalate between
DNA bases. SYBR Green exhibits low fluorescence
when unbound in solution, but starts to fluoresce
brightly when associated with double stranded DNA
(dsDNA) and exposed to a suitable wavelength of
light: the fluorescence increases exponentially as the
DNA target sequence is amplified.

A) DNA is denatured and SYBR Green molecules are free in the reaction mix. B) Primers anneal and SYBR Green molecules bind to the dsDNA. C) DNA
polymerase elongates the template and more SYBR Green molecules bind to the product formed resulting in exponential increase in the fluorescence level.

Evaluation of Real-time PCR

Multiplex polymerase chain reactionMultiplex-PCR consists of multiple primer sets
within a single PCR mixture to produce amplicons of varying sizes that are specific to different DNA sequences. By targeting multiple sequences at once, additional information may be gained from a single test run that otherwise would require several times the reagents and more time to perform. Annealing temperatures for each of the primer sets Annealing temperatures for each of the primer sets must be optimized to work correctly within a single reaction, and amplicon sizes, i.e., their base pair length, should be different enough to form distinct bands when visualized by gel electrophoresis. Alternatively, if amplicon sizes overlap, the different amplicons may be differentiated and visualised using primers that have been dyed with different colour fluorescent dyes.

Scheme of Multiplex Polymerase
Chain Reaction


Scheme of Multiplex Polymerase
Chain Reaction

Distinct bands are visualized by gel
electrophoresis