Embed Size (px)
Citation preview

Retraction and Corrections
RETRACTION
PROFILERetraction for ‘‘Profile of Phillip Cohen,’’ by Carrie Arnold,which appeared in issue 12, March 24, 2009, of Proc Natl AcadSci USA (106:4581–4583; first published March 17, 2009;10.1073/pnas.0902168106).
The editors wish to note that because of several factual andtypographical errors we are retracting this article. A correctedversion will be published in a subsequent issue. The PNASeditors deeply regret the errors.
www.pnas.org�cgi�doi�10.1073�pnas.0903490106
CORRECTIONS
EVOLUTIONCorrection for ‘‘Origin, antiviral function and evidence forpositive selection of the gammaretrovirus restriction gene Fv1 inthe genus Mus,’’ by Yuhe Yan, Alicia Buckler-White, KurtWollenberg, and Christine A. Kozak, which appeared in issue 9,March 3, 2009, of Proc Natl Acad Sci USA (106:3259–3263; firstpublished February 12, 2009; 10.1073�pnas.0900181106).
The authors note that due to a printer’s error, the databaseaccession numbers in the footnote on page 3259 appearedincorrectly. The footnote ‘‘Data deposition: The sequencesreported in this paper have been deposited in the GenBankdatabase (accession nos. X97719, FJ603554, and X97720),’’should instead have appeared as: ‘‘Data deposition: The se-quences reported in this paper have been deposited in theGenBank database (accession nos. FJ603554–FJ603574).’’ Ad-ditionally, on page 3259, left column, the first line of the third fullparagraph, ‘‘Fv1 was cloned and identified as a coopted ERVsequence that is related to the gag gene of MuERV-L (3, 4), aClass III (spumavirus-related) ERV transposit family that istranspositionally active in mice but has no known infectious viruscounterparts,’’ should instead have appeared as ‘‘Fv1 was clonedand identified as a coopted ERV sequence that is related to thegag gene of MuERV-L (3, 4), a Class III (spumavirus-related)ERV family that is transpositionally active in mice but has noknown infectious virus counterparts.’’ These errors do not affectthe conclusions of the article.
www.pnas.org�cgi�doi�10.1073�pnas.0901885106
MEDICAL SCIENCESCorrection for ‘‘A genomic approach to colon cancer riskstratification yields biologic insights into therapeutic opportu-nities,’’ by Katherine S. Garman, Chaitanya R. Acharya, ElenaEdelman, Marian Grade, Jochen Gaedcke, Shivani Sud, WilliamBarry, Anna Mae Diehl, Dawn Provenzale, Geoffrey S. Gins-burg, B. Michael Ghadimi, Thomas Ried, Joseph R. Nevins,Sayan Mukherjee, David Hsu, and Anil Potti, which appeared inissue 49, December 9, 2008, of Proc Natl Acad Sci USA(105:19432–19437; first published December 2, 2008; 10.1073�pnas.0806674105).
The authors note that the term ‘‘prognostic score’’ should besubstituted for the term ‘‘Recurrence Score,’’ which is a regis-tered trademark of Genomic Health and is not associated in anyway with the authors or the article. The online version of thearticle has been corrected accordingly as of April 7, 2009.
www.pnas.org�cgi�doi�10.1073�pnas.0902004106
6878 � PNAS � April 21, 2009 � vol. 106 � no. 16 www.pnas.org

Origin, antiviral function and evidence for positiveselection of the gammaretrovirus restriction geneFv1 in the genus MusYuhe Yana, Alicia Buckler-Whitea, Kurt Wollenbergb, and Christine A. Kozaka,1
aLaboratory of Molecular Microbiology, bBioinformatics and Computational Biosciences Branch, Office of Cyber Infrastructure and Computational Biology,National Institute of Allergy and Infectious Diseases, Bethesda, MD 20892-0460
Communicated by Malcolm A. Martin, National Institutes of Health, Bethesda, MD, January 7, 2009 (received for review November 17, 2008)
The Fv1 virus resistance gene is a coopted endogenous retrovirus(ERV) sequence related to the gag gene of the MuERV-L ERV family.Three major Fv1 resistance alleles have been identified in labora-tory mice, and they target virus capsid genes to produce charac-teristic patterns of resistance to mouse leukemia viruses (MLVs).We identified Fv1 in 3 of the 4 Mus subgenera; its absence fromCoelomys and 1 of 3 species of Pyromys indicate Fv1 was acquiredshortly after the origin of the Mus genus. We sequenced Fv1 genesfrom 21 mice representative of the major taxonomic groups ofMus. Two lines of evidence indicate that Fv1 has had antiviralfunction for 7 million years of evolution. First, 2 species of Africanpygmy mice (subgenus Nannomys) show an Fv1-like MLV resis-tance, and transduced cells expressing the Nannomys Fv1 genereproduce this resistance pattern. Second, sequence comparisonssuggest that Fv1 has been involved in genetic conflicts throughoutMus evolution. We found evidence for strong positive selection ofFv1 and identified 6 codons that show evidence of positive selec-tion: 3 codons in the C-terminal region including 2 previouslyshown to contribute to Fv1 restriction in laboratory mice, and 3codons in a 10-codon segment overlapping the major homologyregion of Fv1; this segment is known to be involved in capsidmultimerization. This analysis suggests that Fv1 has had an anti-viral role throughout Mus evolution predating exposure of mice tothe MLVs restricted by laboratory mouse Fv1, and suggests amechanism for Fv1 restriction.
Fv1 gammaretrovirus restriction gene � mouse endogenous retrovirus �Mus evolution
W ild mouse species and inbred laboratory strains vary intheir susceptibility to gammaretrovirus infection, and
such resistance can be due to constitutively expressed antiviralfactors that target various stages of the retroviral life cycle. Theprototype for such virus resistance factors is the Fv1 gene,discovered 40 years ago in studies on resistance to Friend murineleukemia virus (MLV) (1).
There are 4 well characterized functional variants of Fv1 andadditional Fv1-like restrictions found in inbred strains and wildmouse species (2). Three of these alleles, termed Fv1n, Fv1b, andFv1nr, produce characteristic patterns of resistance to subgroupsof mouse-tropic viruses that are designated N-, B-, or NR-tropic.Cells with the Fv10 (null) allele restrict none of these virussubgroups, and NB-tropic viruses are not restricted by any ofthese Fv1 alleles.
Fv1 was cloned and identified as a coopted ERV sequence thatis related to the gag gene of MuERV-L (3, 4), a Class III(spumavirus-related) ERV transposit family that is transposi-tionally active in mice but has no known infectious virus coun-terparts. The major resistance variants of Fv1 differ from oneanother at 3 amino acid sites in its C-terminal region, and Fv1b
additionally differs from Fv1n and Fv1nr at its C terminus due toa 1.3-kb indel (3). Substitutions at the 3 sites and variation at theC terminus all contribute to resistance (5, 6). The mechanism ofresistance is unknown, but Fv1 typically blocks replication after
reverse transcription and before integration. Fv1 is known totarget the virus capsid gene; a single amino acid substitution atposition 110 distinguishes N- and B-tropic viruses (7), andsubstitutions at additional residues in the capsid N-terminaldomain are responsible for NR- and NB-tropism (5, 8).
Until recently, Fv1-type restriction had only been identified inlaboratory mice and wild mouse species closely related tolaboratory mice (2). Our examination of an African pygmymouse, subgenus Nannomys, identified an unusual postentryresistance to ecotropic MuLVs (9). Although this resistancetargets some of the same amino acid residues as the mouse Fv1gene, the pattern of virus resistance in the pygmy mouse cellsdoes not resemble that attributed to any of the laboratory mouseFv1 alleles.
We have now screened additional Mus species distantly relatedto laboratory strains for Fv1-like resistance phenotypes, andanalyzed the Fv1 sequence in wild mouse species of 3 Mussubgenera. We show here that the pygmy mouse Fv1 has antiviralactivity and demonstrate that Fv1 has been under strong positiveselection throughout 7 MY of Mus evolution. We identified 6codons under strong positive selection including 2 residuesimplicated in Fv1-mediated virus restriction, and 3 codons in asegment overlapping the Fv1 major homology region (MHR)region, a region that in retroviruses produces the interface forcapsid binding and dimerization.
ResultsAnalysis of the 4 Subgenera of Mus for Fv1. We examined repre-sentative species of all 4 Mus subgenera (Mus, Pyromys, Nanno-mys and Coelomys) for Fv1 sequences by Southern blot analysisusing a probe from the 5� end of Fv1 (Fig. 1A). One Fv1-reactiveband was detected in most species; Mus mus caroli has 2BglI-generated Fv1-related copies (Fig. 1B). Fv1 was identifiedin 3 of the 4 Mus subgenera; it was missing in Mus coelomys pahariand in Mus pyromys shortridgei, 1 of 3 Pyromys species tested.
PCR using primers designed to amplify the 3� end of Fv1 alongwith flanking sequences confirmed the absence of Fv1 fromthese 2 species (Fig. 1C). These primers also distinguish the 2major Fv1 variants found in laboratory mouse strains; Fv1n hasa 1.3-kb deletion at its 3� end relative to Fv1b. Both Fv1 variantswere identified in Mus species. Most mice carry the 1.3-kbsegment characteristic of Fv1b; the Fv1n deletion was found onlyin house mouse species, specifically all 4 Mus mus musculus
Author contributions: C.A.K. designed research; Y.Y. and A.B.-W. performed research;A.B.-W. and K.W. analyzed data; and C.A.K. wrote the paper.
The authors declare no conflict of interest.
Data deposition: The sequences reported in this paper have been deposited in the GenBankdatabase (accession nos. X97719, FJ603554, and X97720).
1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/cgi/content/full/0900181106/DCSupplemental.
www.pnas.org�cgi�doi�10.1073�pnas.0900181106 PNAS � March 3, 2009 � vol. 106 � no. 9 � 3259–3263
EVO
LUTI
ON

samples tested and some Mus mus domesticus and Mus musspretus mice.
Mus originated 7–8 MYA and quickly radiated into 4 subgen-era. The radiations leading to these subgenera are difficult toorder, but Coelomys is generally regarded as the most basalgroup in Mus (10). Our results indicate that Fv1 is absent fromspecies in 2 of the non-Mus subgenera, including Coelomys, andthat when Fv1 entered the Mus germ line it contained the 1.3-kbsegment found in the laboratory mouse Fv1b allele.
Restriction of Ecotropic MLVs in Cells of 2 Species of Nannomys. Todetermine if Fv1 sequences serve an antiviral function in speciesfrom non-Mus subgenera, we infected cells from these mice withvarious viruses known to be subject to restriction by laboratorymouse Fv1, and susceptibility was compared with NIH 3T3
(Fv1n), 129/J (Fv1nr) and M. dunni (Fv10) (Table 1). M. pyromysplatythrix was fully susceptible to all viruses tested indicating thatits Fv1 gene has no antiviral activity against this particular panelof MLVs. Cells of a second species, Mus nannomys minutoides,were completely resistant to all AKV MLVs, but susceptible toMoMLV and Friend MLVs, as reported (9). In contrast, cells ofanother Nannomys species, Mus nannomys setulosus, show low,but detectable susceptibility to AKV-B and AKV-NR, reducedsusceptibility to MoMLV, and differential susceptibility to the 2Friend MLVs. M. n. setulosus cells, like M. n. minutoides, arecompletely resistant to infection with AKV-N and to an MoMLVchimera, Mo-CA3, in which part of the Fv1 target region (codonpositions 99–129) of the capsid gene was replaced with that ofAKV-N (9). Thus, these 2 pygmy mice have similar butdistinctive patterns of susceptibility to MLVs, and their resis-tance to AKV-N is mediated by the region of capsid that istargeted by Fv1.
Fv1 gene of M. n. minutoides and M. n. setulosis. The 5� and 3� endsof the Fv1 gene of M. n. minutoides, Fv1m, were amplified fromgenomic DNA and sequenced (Fig. 2); this sequence included acomplete 1.3-kb Fv1-related ORF and flanking sequences onboth sides of the gene. The 24-bp segment flanking the 5� endis identical to that flanking Fv1b. The 803-bp flanking the 3� endof the ORF are homologous to the Fv1b f lanking segment, butalso contain a 170-bp B1 repeat inserted near the end of thecoding region. The 437-aa Fv1m ORF is highly homologous to the440-aa Fv1 gene of the laboratory mouse. The Fv1m predictedprotein sequence is 87% identical to Fv1n. The differencesinclude a 9-base in-frame deletion near the N terminus, and 9-aadifferences at the C terminus due to the B1 insertion. The restof the differences are scattered and notably include differencesat the 3 amino acid positions (352, 358, and 399) that distinguishFv1n, Fv1b, and Fv1nr and have been shown to modulate resis-tance (3, 5, 6). Residues at these positions for the knownrestrictive Fv1 alleles are: SKV (Fv1n), SER (Fv1b), and FKV(Fv1nr). The Fv1m sequence has a different combination at thesesites, FKS.
The predicted amino acid sequence of the Fv1 gene clonedfrom M. n. setulosus cells, Fv1s, is 98% identical to Fv1m, and hasa similar C terminus and the same 9-bp deletion in the Nterminus relative to Fv1n. For Fv1s, the amino acids at the 3 sitesthat distinguish the 3 known restrictive alleles are FKE, a uniquepattern.
Fig. 2. Comparison of predicted amino acid sequence of the M. n. minu-toides Fv1 gene and Fv1b. The position of MHR is indicated by a solid bar andshading indicates the 3 amino acids identified as critical for restriction basedon analysis of laboratory mouse Fv1 alleles (3, 5, 6).
Table 1. Virus titers of MLVs on cells of various Mus species andcells with known Fv1 restriction
Cells
Log10 Virus Titer*/Fv1 Tropism
AKV MLV MoMLV FBLV F-S MLV
N B NR NB NB NR
NIH 3T3 (Fv1n) 4.4 0.5 4.4 6.5 5.1 5.6129/J EF (Fv1nr) 1.8 0.9 4.2 5.8 ND 4.7M. dunni (Fv10) 4.1 4.5 4.4 2.5† 4.1 4.7M. n. minutoides �0 �0 �0 4.5 4.3 4.0M. n. setulosus �0 1.4 0.7 3.6 2.8 1.1M. p. platythrix 3.7 4.4 4.4 5.8 4.1 4.4
*Virus titers were determined by the XC overlay test in which the indicatedcells were infected with virus dilutions, irradiated 4 days later and overlaidwith XC cells to identify clusters of virus infected cells (9, 11). Titers representthe number of XC PFU in 0.2 ml in representative experiments. ND, not done.
†Restriction is mediated by receptor polymorphism (12), and is unrelated toFv1.
Fig. 1. Detection of Fv1 in DNAs of Mus species. (A) The structure of Fv1b isshown with a gray box marking the MHR, open boxes representing B2 repeatsand a dashed line representing the 1.3-kb segment deleted in Fv1n. The arrowsrepresent the PCR primers Fb3003 and Rb4831, and the black box representsthe segment used for blot hybridization. Fv1 is encoded by a single exon. (B)Southern blot analysis of BglI-digested Mus DNAs. All lanes taken from thesame exposure of a single blot; deleted lanes are indicated by vertical lines. (C)PCR products of Mus DNAs.
3260 � www.pnas.org�cgi�doi�10.1073�pnas.0900181106 Yan et al.
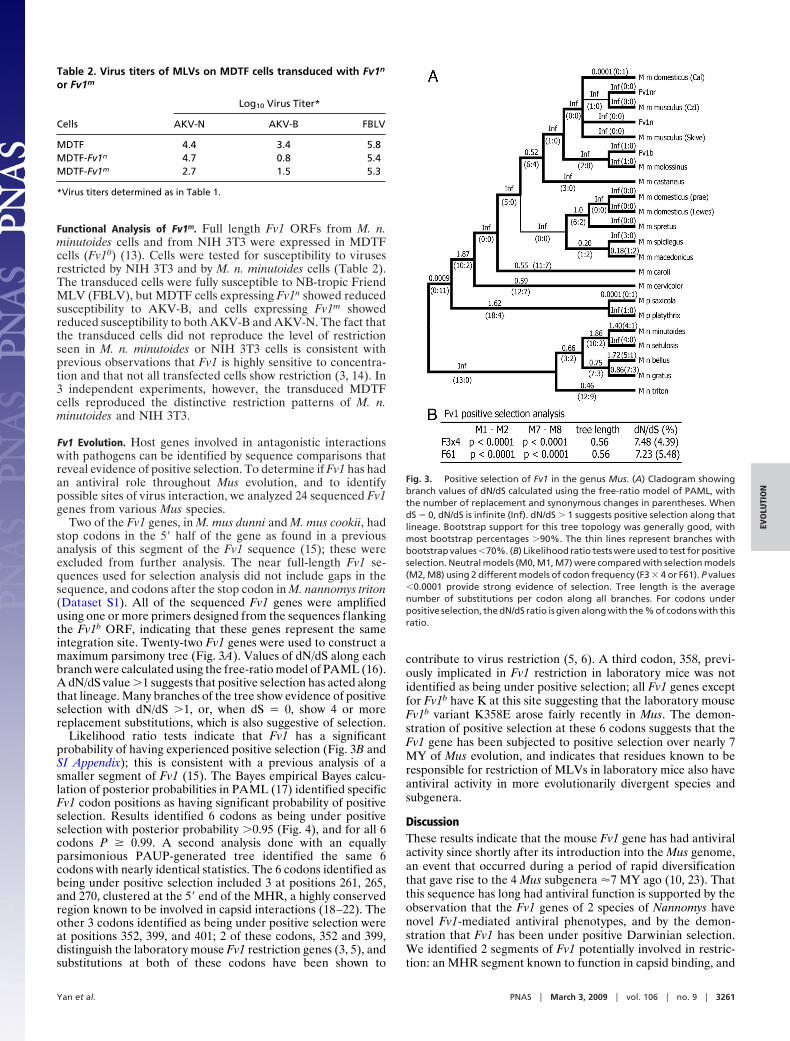
Functional Analysis of Fv1m. Full length Fv1 ORFs from M. n.minutoides cells and from NIH 3T3 were expressed in MDTFcells (Fv10) (13). Cells were tested for susceptibility to virusesrestricted by NIH 3T3 and by M. n. minutoides cells (Table 2).The transduced cells were fully susceptible to NB-tropic FriendMLV (FBLV), but MDTF cells expressing Fv1n showed reducedsusceptibility to AKV-B, and cells expressing Fv1m showedreduced susceptibility to both AKV-B and AKV-N. The fact thatthe transduced cells did not reproduce the level of restrictionseen in M. n. minutoides or NIH 3T3 cells is consistent withprevious observations that Fv1 is highly sensitive to concentra-tion and that not all transfected cells show restriction (3, 14). In3 independent experiments, however, the transduced MDTFcells reproduced the distinctive restriction patterns of M. n.minutoides and NIH 3T3.
Fv1 Evolution. Host genes involved in antagonistic interactionswith pathogens can be identified by sequence comparisons thatreveal evidence of positive selection. To determine if Fv1 has hadan antiviral role throughout Mus evolution, and to identifypossible sites of virus interaction, we analyzed 24 sequenced Fv1genes from various Mus species.
Two of the Fv1 genes, in M. mus dunni and M. mus cookii, hadstop codons in the 5� half of the gene as found in a previousanalysis of this segment of the Fv1 sequence (15); these wereexcluded from further analysis. The near full-length Fv1 se-quences used for selection analysis did not include gaps in thesequence, and codons after the stop codon in M. nannomys triton(Dataset S1). All of the sequenced Fv1 genes were amplifiedusing one or more primers designed from the sequences flankingthe Fv1b ORF, indicating that these genes represent the sameintegration site. Twenty-two Fv1 genes were used to construct amaximum parsimony tree (Fig. 3A). Values of dN/dS along eachbranch were calculated using the free-ratio model of PAML (16).A dN/dS value �1 suggests that positive selection has acted alongthat lineage. Many branches of the tree show evidence of positiveselection with dN/dS �1, or, when dS � 0, show 4 or morereplacement substitutions, which is also suggestive of selection.
Likelihood ratio tests indicate that Fv1 has a significantprobability of having experienced positive selection (Fig. 3B andSI Appendix); this is consistent with a previous analysis of asmaller segment of Fv1 (15). The Bayes empirical Bayes calcu-lation of posterior probabilities in PAML (17) identified specificFv1 codon positions as having significant probability of positiveselection. Results identified 6 codons as being under positiveselection with posterior probability �0.95 (Fig. 4), and for all 6codons P � 0.99. A second analysis done with an equallyparsimonious PAUP-generated tree identified the same 6codons with nearly identical statistics. The 6 codons identified asbeing under positive selection included 3 at positions 261, 265,and 270, clustered at the 5� end of the MHR, a highly conservedregion known to be involved in capsid interactions (18–22). Theother 3 codons identified as being under positive selection wereat positions 352, 399, and 401; 2 of these codons, 352 and 399,distinguish the laboratory mouse Fv1 restriction genes (3, 5), andsubstitutions at both of these codons have been shown to
contribute to virus restriction (5, 6). A third codon, 358, previ-ously implicated in Fv1 restriction in laboratory mice was notidentified as being under positive selection; all Fv1 genes exceptfor Fv1b have K at this site suggesting that the laboratory mouseFv1b variant K358E arose fairly recently in Mus. The demon-stration of positive selection at these 6 codons suggests that theFv1 gene has been subjected to positive selection over nearly 7MY of Mus evolution, and indicates that residues known to beresponsible for restriction of MLVs in laboratory mice also haveantiviral activity in more evolutionarily divergent species andsubgenera.
DiscussionThese results indicate that the mouse Fv1 gene has had antiviralactivity since shortly after its introduction into the Mus genome,an event that occurred during a period of rapid diversificationthat gave rise to the 4 Mus subgenera �7 MY ago (10, 23). Thatthis sequence has long had antiviral function is supported by theobservation that the Fv1 genes of 2 species of Nannomys havenovel Fv1-mediated antiviral phenotypes, and by the demon-stration that Fv1 has been under positive Darwinian selection.We identified 2 segments of Fv1 potentially involved in restric-tion: an MHR segment known to function in capsid binding, and
Table 2. Virus titers of MLVs on MDTF cells transduced with Fv1n
or Fv1m
Cells
Log10 Virus Titer*
AKV-N AKV-B FBLV
MDTF 4.4 3.4 5.8MDTF-Fv1n 4.7 0.8 5.4MDTF-Fv1m 2.7 1.5 5.3
*Virus titers determined as in Table 1.
Fig. 3. Positive selection of Fv1 in the genus Mus. (A) Cladogram showingbranch values of dN/dS calculated using the free-ratio model of PAML, withthe number of replacement and synonymous changes in parentheses. WhendS � 0, dN/dS is infinite (Inf). dN/dS � 1 suggests positive selection along thatlineage. Bootstrap support for this tree topology was generally good, withmost bootstrap percentages �90%. The thin lines represent branches withbootstrap values �70%. (B) Likelihood ratio tests were used to test for positiveselection. Neutral models (M0, M1, M7) were compared with selection models(M2, M8) using 2 different models of codon frequency (F3 � 4 or F61). P values�0.0001 provide strong evidence of selection. Tree length is the averagenumber of substitutions per codon along all branches. For codons underpositive selection, the dN/dS ratio is given along with the % of codons with thisratio.
Yan et al. PNAS � March 3, 2009 � vol. 106 � no. 9 � 3261
EVO
LUTI
ON

3 residues in the C-terminal domain (CTD) of which 2 are knownto contribute to the specificity of Fv1 restriction in laboratory mice.
The mechanism of Fv1 restriction has not been elucidatedafter 40 years of investigation, but the determination that Fv1encodes a retroviral capsid-like protein suggests that Fv1 maybind capsids of exogenous virus and interfere with capsiddisassembly and reverse transcription. Three of the positivelyselected codons identified in this study are in a 10-codonsegment associated with the Fv1 capsid-like MHR region. Mu-tations of codons in and around the MHR regions of variousretroviruses disrupt virus assembly, maturation and infectivity(18–20). Mutational analysis in and around the MHR ofMoMLV and HIV-1 has determined that the segment describedhere is critical for formation of the high-affinity capsid interface,and that this segment determines the specificity of heterodimericinteractions (20, 21). These interactions are important in theearly stages of replication as the majority of replication defectiveMoMLV CA mutants are blocked before reverse transcription(22). Mutations within this segment of the Fv1 MHR demon-strate this region is critical for Fv1 restriction (6), but whilestudies on Fv1 have not produced direct evidence for Fv1-capsidbinding, our results support an antiviral model based on Fv1-capsid interactions.
Three of the 6 positively selected residues include 2 in the CTDthat have already been identified as critical for the specificity ofFv1 restriction. F/S352 distinguishes Fv1n and Fv1nr restrictiontypes, and substitutions at codon 399 alter restriction specificity(5, 6). Our analysis also identified positive selection at a positionin this region not previously known to affect restriction, 401; itsrole in restriction may be defined by further mutational analysis.A 4th codon in this region of the CTD, 358, is known to affectMLV-N/B/NB restriction specificity (6), but we found this site tobe highly invariant in Mus species. Lack of sequence variation atthis site suggests that the only known substitution at the 358codon identified to date, in the Fv1b laboratory mouse strains,likely emerged very recently in Mus evolution, perhaps inresponse to the emergence of the N-tropic MLV it restricts.
Fv1 is not found in all mice. Fv1 entered the mouse germ lineshortly after the origination of Mus during a 1 MY period of
diversification �7 MYA; this is supported by the observationthat 2 species from 2 of the 4 subgenera that originated at thattime lack Fv1. Evidence of positive selection of the Fv1 gene, theidentification of 2 different but related Fv1-like restriction genesin Nannomys and the demonstration that one of these cloned Fv1genes, Fv1m, reproduces this resistance phenotype in transducedcells suggests Fv1 has had an antiviral role throughout Musevolution.
The exposure of mice to MLVs is marked by the appearanceof related ERVs, but the appearance of Fv1 in Mus significantlypredates the acquisition of ERVs of MLVs, the viruses restrictedby Fv1. We demonstrated that MLV ERVs are largely found onlyin house mouse species that appeared only �1 MYA, that is, M.m. domesticus, castaneus and musculus (24). Our evolutionaryand functional analysis of Fv1 in other species and subgenerauncovered evidence of antiviral activity in MLV-free species,which suggest that Fv1 may have broader antiretroviral activitythan previously appreciated.
Materials and MethodsCells and Viruses. Ecotropic (mouse-tropic) MLV isolates were obtained fromJ. W. Hartley (National Institute of Allergy and Infectious Diseases, Bethesda,MD) and included AKV MLV (AKV-N), WN1802B (AKV-B), AKR-L1 (AKV-NR),Moloney MLV (MoMLV), and 2 Friend MLV (FrMLV) isolates, NR-tropic F-S MLVand NB-tropic FBLV (9). Mo-CA3 is a chimera of MoMLV with a segment of AKVMLV capsid containing the Fv1 target region between residues 99 and 129 (9).Virus stocks were made by collecting culture fluids from virus infected cells.
The XC overlay test (9, 11) was used to test for susceptibility to virusinfection in various cell lines including 2 lines derived from the Asian speciesMus m. dunni, MDTF and M. dunni (13), NIH 3T3 and cell lines derived from tailfibroblasts of the wild mouse species M. n. minutoides, M. n. setulosus, and M.p. platythrix obtained from J. Rodgers (Baylor College of Medicine, Houston,TX) (25). Embryo fibroblasts were prepared from strain 129/J from The JacksonLaboratory.
Wild Mouse Genomic DNA. DNA was isolated from the wild mouse-derived celllines listed above, from individual NFS/N mice from our colony and from wildmice from various sources (Table S1). Most mice were originally obtained fromM. Potter (National Cancer Institute, Bethesda, MD). A set of African pygmymouse DNA samples was obtained from Y. Cole and P. D’Eustachio (New YorkUniversity, New York); these mice had been classed into 4 species of Nannomysmice on the basis of skeletal features by J. T. Marshall (Smithsonian NaturalHistory Museum, Washington, DC). A sample of M. m. macedonicus DNA wasprovided by R. Elliott (Roswell Park, Buffalo, NY), and M. m. domesticus micetrapped in California were provided by S. Rasheed (University of SouthernCalifornia, Los Angeles). M. m. spretus (SPRET/EiJ), M. m. castaneus (CAST/EiJ),M. m. molossinus, BALB/cJ, and C57BL/6J were obtained from The JacksonLaboratory. All animal protocols were reviewed and approved (Proposal no.LMM1) by the National Institute of Allergy and Infectious Diseases AnimalCare and Use Committee.
All wild-derived mice are named using genus, subgenus, and species.
Cloning and Functional Analysis of M. n. minutoides Fv1. The 5� end of the Fv1gene of M. n. minutoides (Fv1m) was PCR amplified and sequenced usingprimers based on Fv1b and numbered according to GenBank accession no.X97719: Fb2136-AGCCGAGTTCTAGGGAAACAA and Rb3133-TCAGCCAAC-CAATCAAACAAACT. The 3� end of Fv1m was then amplified from NcoI di-gested DNA using inverse PCR (26) with Fv1m-based primers from GenBankaccession no. FJ603554: Fm568-ATAGATTCTGATGGGACTGAG and Rm184-TACACAGAGTCATTAAGTTCCTTACC. The PCR product was sequenced.
The Fv1m coding region was then amplified from total RNA of M. n.minutoides cells using primers with introduced HindIII and ClaI sites (Fm22-TCAAGCTTAGGATGAATTTCCTACGTGCGCTTGCTG and Rm1313-TTATCGATT-TAAAGGCCACCACGCCCGGCTTTTG) and cloned into the retroviral vectorpLNCX2 (Clontech). The full length Fv1n gene was similarly amplified fromtotal RNA extracted from NIH 3T3 cells with primers based on GenBankaccession no. X97720: Fn35-TCAAGCTTAGGATGAATTTCCCACGTGCGC-TTGCTG and Rn1332-TTATCGATTCAGAGTTTTGTAGCTGCTGTTGGCT andcloned into pLNCX2. The Fv1 genes were packaged by cotransfecting GP2–293cells (Clontech) with the Fv1 clones or empty vector and pVSV-G (Clontech).Culture fluids were collected and used to infect MDTF cells, and stably trans-duced cells were selected for virus infection.
Fig. 4. Fv1 sites that have been subject to positive selection. At the top aresequence alignments for 22 Mus genes in the regions containing the 6 posi-tively selected codons; all changes relative to Fv1b are shaded. MHR is boxed.At the bottom is a diagram of the Fv1 coding region showing the locations ofthe MHR (black box), the 6 positively selected codons and the 1 additionalcodon (358) implicated in restriction (3, 6).
3262 � www.pnas.org�cgi�doi�10.1073�pnas.0900181106 Yan et al.

Fv1 Sequences of Other Wild Mouse Species. BglI digested DNAs were screenedfor the presence of Fv1 by Southern blot analysis using as probe a 358-bpsegment of the 5� end of Fv1b amplified from C57BL/6 DNA using primersFb2165-AGATGAATTTCCCACGTGC and Rb2497-AGGACACACTTAGAAGC-CTTTAGATC. This segment shows 27% identity to the MuERV-L family ofwhich Fv1 is a member (4).
Fv1 was amplified from various mouse genomic DNAs using primers de-signed from coding and flanking sequences of Fv1m and Fv1b. Flanking se-quence primers included Fb2136-AGCCGAGTTCTAGGGAAACAA, Rb3703-TTTGCAACCAACCAGTGGCA, Rb4179-TCATAGCATATGTGAACAATCA, andRb4831-CATCTATACTATCTTGGTGAG. From Fv1 coding sequences, we usedprimers Fb2165, F/Rb3003-TTTAAGGGTGTGGGATAATGGT and F/Rb3133-AGTTTGTTTGATTGGTTGGCTGA. Most Fv1 genes were sequenced as 2 over-lapping PCR products, some were first cloned into pCR2.1-TOPO (Invitrogen,Carlsbad, CA) before sequencing (Dataset S1).
Selection Analysis of Lineages and Codons. DNA sequences were aligned usingMUSCLE (27) and improved manually. The phylogeny used was 1 of 4 equallyparsimonious phylogenies returned by PAUP* (version 4.0b10) (28), and waschosen by correspondence to consensus Mus phylogeny (10, 23). Two of the 4equally parsimonious trees differed only in the arrangement of zero-lengthbranches, leaving only 2 different tree topologies. The codeml program of thePAML4 package (16) was used for maximum likelihood analysis of codonevolution (29). The free ratio model (codon model � 1) was used to calculate
branch-specific rates of dN/dS. In this model each branch is assumed to have aspecific dN/dS ratio. The likelihood of the phylogeny under this model wastested against the likelihood of the phylogeny under the model of oneuniform dN/dS ratio across all branches (codon model 0) using a likelihoodratio test (LRT). The significance of the LRT value was assessed using a �2
distribution with 36° of freedom.Selection acting on Fv1 codons was analyzed using 2 models of equilibrium
codon frequencies and 4 models of codon selection. The 2 codon frequencymodels used were the F3 � 4 model (codon frequencies estimated from thenucleotide frequencies in the data at each codon site) and the F61 codon tablemodel (frequencies of each of the 61 non-stop codons estimated from thedata). The codon selection models were 2 neutral/negative selection models(M1 and M7), which were compared against corresponding models includinga category for dN/dS �1 (M2 and M8, respectively). The significance of thisadditional codon selection category was assessed using LRTs of the phylogenylikelihoods under the neutral and positive selection models. Significance ofthe test statistics was calculated using a �2 distribution with 2 degrees offreedom. The Bayes empirical Bayes algorithm (17) was used to calculate theposterior probability of individual codons experiencing dN/dS �1.
ACKNOWLEDGMENTS. We thank Esther Shaffer and Qingping Liu for experttechnical assistance. This work was supported by the Intramural ResearchProgram of the National Institutes of Health, National Institute of Allergy andInfectious Diseases.
1. Lilly F (1967) Susceptibility to two strains of Friend leukemia virus in mice. Science155:461–462.
2. Kozak CA (1985) Analysis of wild-derived mice for the Fv-1 and Fv-2 murine leukemiavirus restriction loci: A novel wild mouse Fv-1 allele responsible for lack of host rangerestriction. J Virol 55:281–285.
3. Best S, LeTossier P, Towers G, Stoye JP (1996) Positional cloning of the mouse retrovirusrestriction gene Fv1. Nature 382:826–829.
4. Benit L, et al. (1997) Cloning of a new murine endogenous retrovirus, MuERV-L, withstrong similarity to the human HERV-L element and with a gag coding sequence closelyrelated to the Fv1 restriction gene. J Virol 71:5652–5657.
5. Stevens A, et al. (2004) Retroviral capsid determinants of Fv1 NB and NR tropism. J Virol78:9592–9598.
6. Bishop KN, Bock M, Towers G, Stoye JP (2001) Identification of the regions of Fv1necessary for murine leukemia virus restriction. J Virol 75:5182–5188.
7. Kozak CA, Chakraborti A (1996) Single amino acid changes in the murine leukemiavirus capsid protein gene define the target of Fv1 resistance. Virology 225:300–305.
8. Jung YT, Kozak CA (2000) A single amino acid change in the murine leukemia viruscapsid gene responsible for the Fv1nr phenotype. J Virol 74:5385–5387.
9. Yan Y, Kozak CA (2008) Novel post-entry resistance to AKV ecotropic mouse gamma-retroviruses in the African pygmy mouse, Mus minutoides. J Virol 82:6120–6129.
10. Veyrunes F, et al. (2006) Phylogenomics of the genus Mus (Rodentia: Muridae):Extensive genome repatterning is not restricted to the house mouse. Proc R Soc B273:2925–2934.
11. Rowe WP, Pugh WE, Hartley JW (1970) Plaque assay techniques for murine leukemiaviruses. Virology 42:1136–1139.
12. Eiden MV, Farrell K, Warsowe J, Mahan LC, Wilson CA (1993) Characterization of anaturally occurring ecotropic receptor that does not facilitate entry of all ecotropicmurine retroviruses. J Virol 67:4056–4061.
13. Lander MR, Chattopadhyay SK (1984) A Mus dunni cell line that lacks sequences closelyrelated to endogenous murine leukemia viruses and can be infected by ecotropic,amphotropic, xenotropic, and mink cell focus-forming viruses. J Virol 52:695–698.
14. Bock M, Bishop KN, towers G, Stoye JP (2000) Use of a transient assay for studying thegenetic determinants of Fv1 restriction. J Virol 74:7422–7430.
15. Qi C-F, et al. (1998) Molecular phylogeny of Fv1. Mamm Genome 9:1049–1055.16. Yang Z (1997) PAML: a program package for phylogenetic analysis by maximum
likelihood. Comput Appl Biosci 13:555–556.17. Yang Z, et al. (2005) Bayes empirical Bayes inference of amino acid sites under positive
selection. Mo Bio Evol 22:1107–1118.18. Strambio-de-Castilla C, Hunter E (1992) Mutational analysis of the major homology
region of Mason-Pfizer monkey virus by use of saturation mutagenesis. J Virol 66:7021–7032.
19. Orlinsky KJ, Gu J, Hoyt M, Sandmeyer S, Menees TM (1996) Mutations in the Ty3 majorhomology region affect multiple steps in Ty3 retrotransposition. J Virol 70:3440–3448.
20. Gamble TR, et al. (1997) Structure of the carboxyl-terminal dimerization domain of theHIV-1 capsid protein. Science 278:849–853.
21. Alin K, Goff SP (1996) Mutational analysis of interactions between the Gag precursorproteins of murine leukemia viruses. Virology 216:418–424.
22. Alin K, Goff SP (1996) Amino acid substitutions in the CA protein of Moloney murineleukemia virus that block early events in infection. Virology 222:339–351.
23. Lundrigan BL, Jansa SA, Tucker PK (2002) Phylogenetic relationships in the genus Mus,based on paternally, maternally, and biparentally inherited characters. Syst Biol51:410–431.
24. Kozak CA, O’Neill RR (1987) Diverse wild mouse origins of xenotropic, mink cellfocus-forming, and two types of ecotropic proviral genes. J Virol 61:3082–3088.
25. Davis BK, Cook RG, Rich RR, Rodgers JR (2002) Hyperconservation of the putativeantigen recognition site of the MHC Class I-b molecule TL in the subfamily Murinae:Evidence that thymus leukemia antigen is an ancient mammalian gene. J Immunol169:6890–6899.
26. Silver J, Keerikatte V (1989) Novel use of polymerase chain reaction to amplify cellularDNA adjacent to an integrated provirus. J Virol 63:1924–1928.
27. Edgar RC (2004) MUSCLE: Multiple sequence alignment with high accuracy and highthroughput. Nucl Acids Res 32:1792–1797.
28. Swofford DL (2003) PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Meth-ods). Version 4. Sinauer Associates, Sunderland, Massachusetts.
29. Bielawski JP, Yang Z (2005) in Statistical Methods in Molecular Evolution, ed NielsenR (Springer, New York), pp 103–124.
Yan et al. PNAS � March 3, 2009 � vol. 106 � no. 9 � 3263
EVO
LUTI
ON


























