Embed Size (px)
Citation preview

574 RESEARCH REPORTS
Copyright Q 2004, SEPM (Society for Sedimentary Geology) 0883-1351/04/0019-0574/$3.00
Paleoenvironmental Recovery After the Cretaceous/Paleogene Boundary Crisis: Evidence From the Marine
Bidart Section (SW France)
LAIA ALEGRETDepartment of Earth Sciences, University College London, London WC1E 6BT, United Kingdom, E-mail: [email protected]
MICHAEL A. KAMINSKIDepartment of Earth Sciences, University College London, London WC1E 6BT, United Kingdom
EUSTOQUIO MOLINADepartamento de Ciencias de la Tierra, Facultad de Ciencias, Universidad de Zaragoza. 50009-Zaragoza, Spain
PALAIOS, 2004, V. 19, p. 574–586
The study of Upper Cretaceous and Lower Paleogene ben-thic foraminifera from the Bidart section (SW France) pro-vides detailed data on the paleobathymetry as well as pa-leoenvironmental conditions across the Cretaceous/Paleo-gene (K/Pg) boundary. A quantitative analysis of benthicforaminiferal assemblages from the Upper MaastrichtianAbathomphalus mayaroensis Biozone and the DanianGuembelitria cretacea, Parvularugoglobigerina eugubina,and Parasubbotina pseudobulloides Biozones was per-formed. Benthic foraminifera indicate that the upper Maas-trichtian and lower Danian sediments at Bidart were de-posited in the upper–middle part of the slope.
Benthic foraminiferal assemblages indicate mesotrophicconditions during the late Maastrichtian and a strong de-crease in the food supply to the sea floor coincident with theK/Pg boundary. This change in the trophic regime was re-lated to the collapse of the food web triggered by the massextinction of calcareous primary producers. Benthic assem-blages in the lower Danian are strongly dominated by fewspecies, and suggest that primary productivity was domi-nated by blooms of non-calcareous primary producers, cre-ating a stressful environment for the benthic fauna. Thefaunal turnover, together with the geochemical evidence, iscompatible with an asteroid impact scenario. Benthic fora-miniferal assemblages suggest that primary productivityhad not completely recovered more than 200 kyr after theK/Pg boundary event.
INTRODUCTION
The Cretaceous/Paleogene (K/Pg) boundary marks oneof the major mass extinctions in Earth’s history. Most au-thors now accept that a large meteorite impacted the Yu-catan Peninsula in Mexico (Alvarez et al., 1980; Smit andHertogen, 1980; Hildebrand et al., 1991) and triggered thecatastrophic mass extinction of calcareous plankton (e.g.,Luterbacher and Premoli-Silva, 1964; Smit, 1982, 1990;Molina et al., 1998), as well as an anomalous enrichmentin Ir and other extraterrestrial impact evidence, such asmicrospherules (interpreted as altered microtektites) andshocked quartz grains in the boundary sediments globally
(Alvarez et al., 1980; Smit and Hertogen, 1980; Smit andten Kate, 1982; Sharpton et al., 1992). Furthermore, d13Cand d18O values in bulk sediment and in tests of benthicand planktic foraminifera show abrupt changes just afterthe K/Pg extinction (Zachos and Arthur, 1986; Kaminskiand Malmgren, 1989; Kaiho et al., 1999), and reflect im-portant environmental changes that occurred immediate-ly after the impact. Nevertheless, there are still some re-searchers who do not support a catastrophic mass extinc-tion model (Keller, 1989 a, b; Keller et al., 1995; Abramov-ich and Keller, 2002), but an extended pattern ofextinction generated by multiple causes that may be dueto the Signor-Lipps effect and reworking processes affect-ing microfossils in the sediments immediately below andabove the K/Pg boundary (Huber et al., 2002; Soria et al.,2001, Molina et al., 2002).
Whereas planktic foraminifera suffered a catastrophicmass extinction coincident with the K/Pg boundary (e.g.,Molina et al., 1998), benthic foraminifera do not show sig-nificant extinctions, as reviewed by Culver (2003). Deep-sea benthic foraminifera show drastic, temporal changesin their community structure coincident with the K/Pgboundary, followed by a gradual, staggered pattern of re-covery across the early Danian (see review by Culver,2003). These temporary changes in benthic foraminiferalassemblages have been interpreted as resulting from thecollapse of the pelagic food web and the subsequent drop infood supply to the benthos (e.g., Kuhnt and Kaminski,1993; Thomas, 1990a, b; Alegret et al., 2001, 2003; Culver,2003).
The Bidart section (southwestern France, Fig. 1) is con-sidered as one of the most complete K/Pg boundary sec-tions in southwestern Europe (Seyve, 1990; Haslett,1994). Nevertheless, the paleoenvironmental turnoveracross the K/Pg boundary in this area remains unclear,and benthic foraminifera from Bidart have not been stud-ied previously. Kuhnt and Kaminski (1993) analyzed theagglutinated benthic foraminiferal changes in the nearbyZumaya and Sopelana sections (Basque Basin, NorthernSpain), and concluded that deep-water agglutinated fora-miniferal turnover indicates a drastic decrease in surfaceproductivity and a collapse of the food-web coincident withthe K/Pg boundary. Apart from that study, no detailedanalyses on benthic foraminifera have been performed in

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 575
FIGURE 1—Paleogeographical reconstruction of the Pyrenean do-main during the Paleocene (modified from Pujalte et al., 1994), andlocation of the Bidart section.
this area, so the results presented here provide importantdata to infer environmental changes across the K/Pgboundary.
In addition to its completeness and excellent exposure,the Bidart section is particularly important because of itspaleogeographical location—between the Tethys and theNorth Atlantic—during the Late Cretaceous and EarlyPaleogene. According to Renard et al. (1982), the Maas-trichtian of Bidart was deposited under Tethyan influenc-es, whereas the Paleocene was influenced primarily by theNorth Atlantic. Consequently, the Bidart section has beenthe focus of various studies, such as stable isotope analysis(Romein and Smit, 1981; Renard et al., 1982; Nelson et al.,1991), iridium content (Smit and Ten Kate, 1982; Dela-cotte, 1982; Bonte et al., 1984), or biostratigraphy (Bonteet al., 1984; Delacotte et al., 1985), among others. Detailedbiostratigraphical studies across the K–Pg interval havebeen based on calcareous nannofossils (Perch-Nielsen,1979; Seyve, 1990) and planktic foraminifera (Haslett,1994; Fondecave-Wallez et al., 1995; Apellaniz et al.,1997). Nevertheless, benthic foraminifera from Bidarthave not been studied in detail, as pointed out by Haslett(1994).
The aim of this study is to perform a detailed, quantita-tive study of Upper Cretaceous and Lower Paleogene ben-thic foraminiferal assemblages from the Bidart section(SW France). Benthic foraminifera, which are closely re-lated to the sea-bottom conditions, provide detailed dataon the paleobathymetric setting of the uppermost Creta-ceous and lowermost Paleogene sediments at Bidart, aswell as on the paleoenvironmental changes across the K/Pg boundary.
MATERIAL AND METHODS
The Bidart section is situated between Hendaye andBiarritz (southwestern France), within the Basque-Can-tabric Basin. The outcrop is located behind the beach of Bi-dart, towards the north of this village (for a more preciselocation of the outcrop, see Peybernes et al., 1996). The up-per 4 m of the Maastrichtian and the lower 3.6 m of the Pa-
leogene were studied at Bidart. The Maastrichtian sedi-ments correspond to the ‘‘Bidart marls’’ (Seunes, 1891),which consist of gray marls and calcareous marls contain-ing abundant foraminifera and scarce macrofossils suchas echinoids. The uppermost two centimeters of the Maas-trichtian consist of gray clayey marls. The K/Pg boundaryis marked by a sharp contact between the Maastrichtianmarls and a 6-cm-thick layer of dark clays with a 2-mm-thick, red ferruginous level at its base (Fig. 2). The rest ofthis clay layer consists of brownish clays towards its lowerpart, and thinly laminated dark gray clays towards thetop. Delacotte (1982) and Bonte et al. (1984) identified aniridium anomaly in the dark clay layer, coinciding exactlywith a drop in the d13C curve and in the d18O values (Pey-bernes et al., 1996). Apellaniz et al. (1997) documented thepresence of microspherules with Ni-rich crystals in themiddle part of the clay layer. This layer is overlain by a;25-cm-thick level of gray to brown clays, which becomemore grayish and calcareous towards the top, reflecting anincrease in total bulk carbonate after a drastic reductionidentified just above the K/Pg boundary (i.e., in the darkclay layer; Renard et al., 1982). Fish teeth were observedtowards the upper part of this level, which is overlain by1.30 m of massive gray limestones. Higher in the section, a50-cm-thick, tabular breccia was identified (Fig. 2); thematrix has a mudstone texture, and the pebbles are cal-careous, poorly sorted (the maximum length ranges from 1mm to 10 cm), and subangular to rounded. This body isoverlain by two tabular, 60-to-90-cm thick levels of mas-sive gray limestones, each of which is strongly bioturbatedtowards the top.
The K/Pg boundary at Bidart, as well as in other K/Pgboundary sections from the Bay of Biscay, is includedwithin a single depositional sequence (Maastrichtian–Danian Depositional Sequence; e.g., Apellaniz et al.,1997). According to these authors, the base of this se-quence is located 10 m below the K/Pg boundary, whereasthe top of the sequence is erosive.
Flysch deposition persisted from Albian to Eocene timesin this area, which was situated in close proximity to a pa-leoslope to the northeast (Seyve, 1984). A distal deep-seafan environment with a 40-mm/kyr sedimentation rate(Nelson et al., 1991) prevailed in this area during theMaastrichtian, whereas an increase in the activity on theslope is recorded in the Paleocene (Haslett, 1994), there-fore, the Paleocene sediments are locally slumped (Seyve,1990). According to Peybernes et al. (1996), the brecciaidentified at Bidart was deposited under conditions of lowsea level, whereas the underlying limestones were depos-ited under high sea-level conditions. Alternatively, thisbreccia could be interpreted as a slump triggered by localtectonics during the early Danian. In spite of this geologi-cal context, the Paleocene foraminifera from Bidart areunlikely to have been affected by major reworking process-es, although the microfossils found in the breccia have notbeen taken into account in the paleoenvironmental inter-pretations.
The studies by Hasslet (1994), Peybernes et al. (1996),and Apellaniz et al. (1997) have been followed for bio-stratigraphic control. These authors identified the upperCretaceous Abathomphalus mayaroensis Biozone, as wellas the Danian Guembelitria cretacea, Parvularugoglobi-gerina eugubina and Parasubbotina pseudobulloides Bio-

576 ALEGRET ET AL.
FIGURE 2—Occurrence and relative abundance of infaunal morphogroups across the K–Pg transition at Bidart. (A) Most-characteristic infaunaltaxa. (B) Relative percentage of infaunal and epifaunal morphogroups. G. c.5 G. cretacea Biozone; Pv. eugu.5 Pv. eugubina Biozone.Biostratigraphy by Haslett (1994), Peybernes et al. (1996), and Apellaniz et al. (1997). See legend in Figure 4.
zones (Figs. 2–3). Foraminiferal assemblages from thelowermost 10 cm of the Danian are dominated by benthicforaminifera, and planktic foraminifera are almost absent.Haslett (1994) and Apellaniz et al. (1997) suggested thatthis level may correspond to the Guembelitria cretaceaBiozone of Smit (1982); the first appearance of Pv. eugubi-na 10 cm above the K/Pg boundary marks the Pv. eugubi-na Biozone, and the base of the Ps. pseudobulloides Biozo-ne is located ;1 m above the K/Pg boundary.
Twenty samples were studied in the upper 4 m of theMaastrichtian and the lower 3.5 m of the Danian. Accord-ing to the biomagnetostratigraphic correlation and cali-bration recently proposed by Arenillas et al. (2004), thebase of the Ps. pseudobulloides Biozone has an estimatedage of 64.945 Ma (based on the calibration by Berggren etal., 1995, who placed the K/Pg boundary at 64.745 Ma) or
65.436 Ma (based on the astronomical calibration by Rohlet al., 2001, who placed the K/Pg boundary at 65.5 Ma).Arenillas et al. (2004) subdivided the Ps. pseudobulloidesBiozone into the Eoglobigerina trivialis Interval Subzone,which has a duration of 171.5–201.1 kyr (based on the cal-ibrations proposed by Berggren et al., 1995 and by Rohl etal., 2001, respectively) and the Subbotina triloculinoidesInterval Subzone. Although these subzones have not beenidentified by Hasslet (1994), Peybernes et al. (1996), orApellaniz et al. (1997) at Bidart, comparison of this sectionto the nearby Zumaya section, where the E. trivialis Sub-zone spans 2.75 m, suggests that at least this subzone isrepresented at Bidart. Further biostratigraphical studiesare needed to test the presence of the Subbotina triloculi-noides Subzone at Bidart. However, in the lack of a betterbiostratigraphical analysis, it may be tentatively inferred

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 577
FIGURE 3—Occurrence and relative abundance of the most-characteristic epifaunal benthic foraminiferal taxa across the K–Pg transition atBidart. G. c.5 G. cretacea Biozone; Pv. eugu.5 Pv. eugubina Biozone. Biostratigraphy by Haslett (1994), Peybernes et al. (1996), and Apellanizet al. (1997). See legend in Figure 4.
that, at least, the whole E. trivialis Subzone is representedat Bidart. These data suggest that the top of the studiedinterval corresponds, approximately, to the top of the E.trivialis Subzone, 225.6–264.5 kyr after the K/Pg bound-ary (according to the calibrations proposed by Berggren etal., 1995 and by Rohl et al., 2001, respectively); if the S.triloculinoides Subzone were partially represented at Bi-dart, the top of the studied section would be even younger.
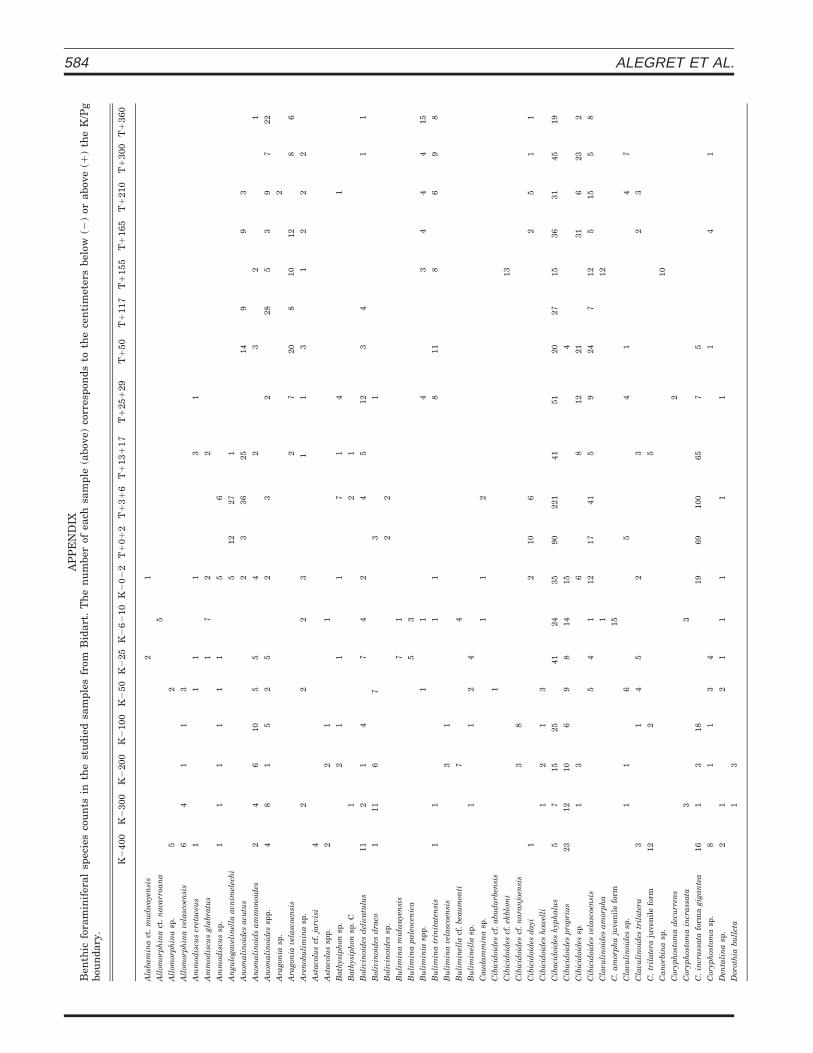
Samples were collected at one-meter intervals at thelower part of the section and at decimeter intervals to-wards the top, with closer sampling in the uppermostMaastrichtian and the lowermost Danian (Appendix 1).Only one micropaleontological sample from the basal 5 cmof the breccia was analyzed, but the results were not con-sidered for paleoenvironmental interpretations due to thepossible reworking processes affecting the microfossils inthis unit. Samples were disaggregated in water with dilut-ed H2O2, washed through a 63-mm sieve, and dried at508C. Species richness calculations and quantitative stud-ies were based on representative splits (using a modified
Otto micro-splitter) of approximately 300–400 specimensof benthic foraminifera larger than 63-mm. All represen-tative specimens were mounted on microslides for a per-manent record and identification. Classification at the ge-neric level follows Loeblich and Tappan (1988); when pos-sible, benthic foraminifers were identified to the specificlevel largely following the taxonomy of Alegret and Thom-as (2001), since the Mexican faunas studied by these au-thors contain around 80% of the benthic foraminiferal spe-cies identified at Bidart. Moreover, 90% of the speciesidentified in this study were reported from the Upper Cre-taceous and lower Paleogene at the Agost section (south-eastern Spain) by Alegret et al. (2003). Benthic foraminif-eral counts are presented in Appendix 1.
The Fisher-a diversity index and the H(S) Shannon-Weaver information function, another index of diversity,were calculated in order to observe possible changes in di-versity across the K–Pg interval. High values of H(S) in-dicate an even distribution of specimens over species;

578 ALEGRET ET AL.
FIGURE 4—Characteristics of benthic foraminiferal assemblagesacross the K–Pg transition at Bidart. (A) Percentages of calcareousand agglutinated benthic foraminiferal tests. (B) Shannon-Weaver het-erogeneity index. (C) Fisher-a diversity index. G. c.5 G. cretacea Bio-zone; Pv. eugu.5 Pv. eugubina Biozone. Biostratigraphy by Haslett(1994), Peybernes et al. (1996), and Apellaniz et al. (1997).
Murray (1991) calls H(S) the index of heterogeneity, withhigh heterogeneity values indicating high diversity.
In order to infer probable microhabitat preferences andenvironmental parameters such as the nutrient supply tothe sea floor and seawater oxygenation (e.g., Bernhard,1986; Jorissen et al., 1995; Fontanier et al., 2002), all ben-thic foraminiferal taxa were allocated into morphogroupsfollowing Corliss (1985, 1991), Jones and Charnock (1985),and Corliss and Chen (1988). According to these authors,there is a general relationship between test morphologyand the microhabitat preferences of benthic foraminifera.Thus, benthic foraminifera with plano-convex, biconvex,and rounded trochospiral tests, as well as tubular andcoiled flattened tests, have an inferred epifaunal mode oflife, living at the sediment surface. Infaunal foraminifera,living in deeper layers of the sediment, have cylindrical orflattened tapered, spherical, rounded planispiral, flat-tened ovoid, globular unilocular, or elongate multiloculartests. Major changes in percentages of these morpho-groups (Fig. 2), as well as in the abundance of selectedtaxa (Figs. 2–3), and the comparison of fossil and recentcommunities of benthic foraminifera have been consideredin this study to infer the paleoenvironmental turnoveracross the K/Pg boundary. Nonetheless, one should becareful with these interpretations due to the complex andnot fully understood ecology of modern faunas, and theirquestionable comparison to Upper Cretaceous and lowerPaleogene faunas (Alegret et al., 2003).
PALEOBATHYMETRY
Benthic foraminiferal assemblages are dominated bycalcareous tests (;97%) through the whole section, exceptfor the upper 10 cm of the Maastrichtian, where theymake up 75% of the assemblages (Fig. 4). These values in-dicate deposition well above the calcite compensationdepth. The planktic/benthic ratio is high (85–90%) in allsamples except for those from the lowermost Danian,which are dominated by benthic foraminifera.
Paleobathymetric assignments were obtained mainlyfollowing the depth distributions and upper depth limits ofbenthic foraminiferal species, as reviewed by Alegret et al.(2003). Benthic foraminiferal assemblages contain abun-dant representatives of the bathyal and abyssal Velasco-type fauna (Berggren and Aubert, 1975), namely Aragoniavelascoensis, Cibicidoides velascoensis, Nuttallides truem-pyi, or Stensioeina beccariiformis, as well as other deep-water species such as Bulimina trinitatensis, Cibicidoideshyphalus, or Marssonella oxycona. The upper depth limitof some of the most abundant species at Bidart (e.g., Buli-mina trinitatensis, Nuttallides truempyi, Stensioeina bec-cariiformis) is located at 500–700 m depth (Alegret et al.,2003). These data suggest that the upper Maastrichtianand lower Danian sediments at Bidart were deposited inthe upper–middle part of the slope.
An increase in the percentage of Loxostomum eleyi hasbeen observed in the uppermost meter of the Maastrich-tian (Fig. 2). The preservation of these specimens is verysimilar to that of other species in the same samples, andthere is no apparent size sorting. These data indicate thatthe abundant specimens of Loxostomum eleyi identified inthe uppermost Maastrichtian sediments are unlikely tohave been reworked. Although Loxostomum eleyi is typical
of outer-shelf environments (MacNeil and Caldwell, 1981;Nyong and Olsson, 1984), its depth occurrence may in-crease under a high food supply, as suggested by Alegretet al. (2003) at the Agost section. The high relative abun-dance of infaunal morphogroups in the Maastrichtian atBidart (Fig. 2) indicates an abundant food supply to thesea-bottom floor, as in Agost. Under these conditions, thedepth of occurrence of L. eleyi may have been somewhatgreater than at other locations.

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 579
FIGURE 5—Scanning electron micrographs of two opportunistic ben-thic foraminiferal species. Scale bar 5 50mm. (A) Glomospira serpenscalcareous isomorph. (B) Spirillina.
BENTHIC FORAMINIFERAL TURNOVER
Upper Maastrichtian assemblages are diverse and het-erogeneous (Fisher-a index ;21, Shannon-Weaver indexH(S) ;3.5; Fig. 4B, C), and they consist of a mixture of in-faunal (;58%) and epifaunal morphogroups (Fig. 2B).Among the infaunal morphogroups, species with cylindri-cal-tapered (e.g., Praebulimina cf. P. reussi, laevidentalin-ids), flattened-tapered (e.g., Loxostomum eleyi, Bolivino-ides draco), and spherical tests dominate (Fig. 2), whereastaxa with trochospiral-planoconvex (e.g., Cibicidoides hy-phalus, Cibicidoides proprius) and biconvex (Nuttallidestruempyi) tests dominate among the epifaunal morpho-groups (Fig. 3). These data suggest mesotrophic conditionsduring the uppermost Maastrichtian, and a nutrient sup-ply to the sea-bottom floor enough to sustain both infaunaland epifaunal morphogroups.
Only three species (Bolivinoides draco draco, Loxosto-mum eleyi, and Praebulimina cf. P. reussi) disappear at Bi-dart in coincidence with the K/Pg boundary (Fig. 2). Twospecimens of Bolivinoides draco draco were found two cen-timeters above the K/Pg boundary, but the different colorof the sediment inside their shells as compared to the oth-er foraminifera in the same sample suggests that they arereworked. Although benthic foraminiferal assemblageswere not significantly affected by the K/Pg extinctions, thepercentage of infaunal morphogroups (Fig. 2) and the di-versity of the assemblages (Fig. 4) drastically decreasedjust after the K/Pg boundary. Lowermost Danian assem-blages (dark clay layer) contain up to 74% of epifaunalmorphogroups, which are dominated mainly by the tro-chospiral planoconvex Stensioeina beccariiformis and Cib-icidoides hyphalus (Fig. 3). Such a decrease in the abun-dance of infaunal taxa and the presence of peaks in theabundance of two epifaunal species suggest a drastic de-crease in the food supply to the sea floor related to the cat-astrophic mass extinction of calcareous plankton at theK/Pg boundary (Zachos and Arthur, 1986; Thomas 1990a,b; Widmark and Malmgren, 1992; Kuhnt and Kaminski,1993; d’Hondt et al., 1998; Widmark, 2000; Alegret et al.,2001, 2002a, b, 2003). Under oligotrophic conditions, theepifaunal Stensioeina beccariiformis and Cibicidoides hy-phalus, living at the sediment surface, bloomed togetherwith the infaunal species Coryphostoma incrassata formagigantea.
Benthic foraminifera from the lower Pv. eugubina Bio-zone are still dominated by epifaunal morphogroups (Fig.2B), especially by trochospiral biconvex (Osangulariaspp.) and planoconvex (Globorotalites spp.) tests (Fig. 3). Itmay be argued that this interval is dominated by epifau-nal taxa not because of the paleoecological characteristicsat that time, but only due to the fact that they correspondto the solution-resistant forms. Nevertheless, both infau-nal and epifaunal species show evidence of dissolution, notonly in these samples but also in the upper two centime-ters of the Maastrichtian and through the whole Danianpart of the section. Moreover, in spite of the fact that epi-faunal species are more susceptible to dissolution in themodern oceans, a peak in the abundance of epifaunal mor-phogroups is observed in the lower Danian, thus suggest-ing that the percentage of epifaunal morphogroups mayhave been even greater and that dissolution was presentbut not overwhelming. In addition, a wide range of sizes
may be observed in all the studied samples, which containboth thick-walled, solution-resistant forms, and thin-walled benthic foraminiferal tests. Two taxa, namely Glo-mospira serpens calcareous isomorph and Spirillina (Fig.5), first appear during this interval. Both taxa morpholog-ically are very similar to Glomospira serpens and Ammo-discus, respectively, which have been documented to bemobile epifaunal forms that feed on organic detritus (Lyand Kuhnt, 1994) and tolerate environmental instability,thus blooming whenever other taxa cannot compete (Ka-minski et al., 1996). The Glomospira serpens calcareousisomorph and Spirillina were not the only taxa in the Pv.eugubina Biozone and in the lower part of the Ps. pseudob-ulloides Biozone, but their appearance and increase inabundance, together with the low diversity and low per-centage of infaunal morphogroups, possibly indicate envi-ronmental instability at the sea floor (Figs. 2–3) An in-crease in the relative abundance of the infaunal Spiroplec-tammina israelskyi and Aragonia velascoensis also is re-corded in the Pv. eugubina Biozone (Fig. 2). Thepercentages of Spirillina and Paralabamina hillebrandtisignificantly increase towards the lower part of the Ps.pseudobulloides Biozone, at which point, their abundancerapidly starts to decrease (Fig. 3).
Higher in the section, benthic foraminiferal assemblag-es from the rest of the Ps. pseudobulloides Biozone aredominated by the epifaunal Stensioeina beccariiformis,Cibicidoides hyphalus, Nuttallides truempyi, and Nuttalli-nella spp. (Fig. 3). The percentage of infaunal morpho-groups increases slightly, but remains low (Fig. 2), as dothe diversity and heterogeneity indices (Fig. 4B, C). Theabundance of typically bathyal, oligotrophic species, to-gether with the dominance of epifaunal morphogroups to-wards the top of the studied interval, suggest that the foodsupply to the sea floor had not completely recovered atthat time, at least more than 200 kyr after the K/Pgboundary.
DISCUSSION
The dominance of calcareous benthic foraminiferathrough the whole section (Fig. 4A) may indicate a smallsupply of terrigenous sediment at Bidart as compared tothe coeval north Spanish Sopelana and Zumaya sections

580 ALEGRET ET AL.
(Kuhnt and Kaminski, 1993), where agglutinated forami-nifera make up two-thirds of the benthic foraminiferal as-semblages. Upper Cretaceous benthic foraminiferal as-semblages are diverse at Bidart, and consist of a mixtureof both infaunal and epifaunal morphogroups, suggestingmesotrophic conditions during the late Maastrichtian.
The drastic decrease in the relative abundance of infau-nal taxa and in the diversity and heterogeneity of the as-semblages coincident with the K/Pg boundary indicates ol-igotrophic conditions, which are related to the decrease inthe organic flux from surface waters as the result of themass extinction of primary producers (e.g., Kuhnt and Ka-minski, 1993; d’Hondt et al., 1998; Alegret et al., 2001,2003). Under conditions of very limited food supply to thesea floor, food particles were consumed at the sedimentsurface by epifaunal species, whereas the percentage of in-faunal species decreased due to the lack of organic matterin the underlying sediment. The drastic reorganization ofbenthic foraminiferal assemblages, together with the geo-chemical evidence at Bidart (e.g., Delacotte, 1982; Bonte etal., 1984; Peybernes et al., 1996), are compatible with anasteroid impact at the K/Pg boundary.
Peybernes et al. (1996) documented the rapid increasein the abundance of the dinoflagellates Thoracosphaeraoperculata, Cyclagelosphaera reinhardii, and Braaduros-phaera bigelowii in the dark clay layer (G. cretacea Biozoneand lower part of the Pv. eugubina Biozone) at Bidart.Such blooms would have provided large amounts of foodthat reached the sea floor even in the absence of pellet-producing zooplankton, which suffered a mass extinction(e.g., Ittekot, 1993; Verity et al., 1996). According to thesedata, the following questions arise: why do benthic fora-minifera indicate oligotrophic conditions just after the K/Pg boundary, when phytoplankton blooms suggest animportant food supply to the sea floor? Under these high-productivity conditions driven by non-calcareous phyto-plankton blooms, why are lowermost Danian benthic fo-raminiferal assemblages apparently stressed, of low diver-sity, and dominated by few species? Speculatively, the ob-served peaks of the epifaunal species Stensioeinabeccariiformis and Cibicidoides hyphalus, and the increasein the percentage of the infaunal Coryphostoma incrassataforma gigantea just after the K/Pg boundary (Figs. 2–3),may be related to the drastic increase in the abundance ofdinoflagellates documented in the lowermost Danian atBidart. Although the species composition of the assem-blages and the low percentage of infaunal morphogroups(Fig. 2), as well as the drop in the d13C curve and in totalbulk carbonate (Renard et al., 1982; Bonte et al., 1984), in-dicate a drastic decrease in primary productivity just afterthe K/Pg boundary, some of the benthic foraminiferal spe-cies that bloomed just after the K/Pg boundary might havebeen able to feed on short-lived, local phytoplanktonblooms that occurred even in the absence of carbonate-richsediment contributed by nannoplankton, as described byAlegret et al. (2003) in the Agost section (southeasternSpain). In the Modern oceans, many organisms do not con-sume dinoflagellates, and the unusually low-diversity fau-nas live under a stressful environment because of thischange in the composition of food directly supplied fromthe photic zone (Peryt et al., 2002). Therefore, most of thebenthic foraminiferal species could not easily use the foodnutrients supplied by the abundant dinoflagellates, thus
accounting for the low diversity and heterogeneity of thelowermost Danian benthic foraminiferal assemblages. Itmay be concluded that it was not only the drastic decreasein primary productivity, but also the changes in phyto-plankton type (from calcareous to organic-walled phyto-plankton) just after the K/Pg boundary that caused thelow-diversity, high-dominance benthic foraminiferal as-semblages from the lower part of the dark clay layer.
The appearance of the opportunistic taxa Spirillina andGlomospira serpens calcareous isomorph, together withthe high percentages of epifaunal taxa such as Globorotal-ites or Paralabamina hillebrandti during the Pv. eugubinaBiochron and the early Ps. pseudobulloides Biochron (Fig.3), suggest that the food supply was still low, and probablydominated by non-calcareous phytoplankton.
The percentage of infaunal taxa increases slightlythrough the studied part of the Ps. pseudobulloides Bio-zone, although it does not reach the high values of the lateMaastrichtian (Fig. 2). Benthic foraminiferal assemblagesfrom this interval are dominated by the deep-water spe-cies Nuttallides truempyi, Stensioeina beccariiformis, andCibicidoides hyphalus (Fig. 3), which are common inbathyal settings with a scarce nutrient supply to the seafloor (Van Morkhoven et al., 1986; Widmark and Malm-gren, 1992; Speijer and Van der Zwaan, 1996; Alegret etal., 2001). The species composition, together with the lowdiversity and heterogeneity of the assemblages (Fig. 4 B–C), suggests that primary productivity had not completelyrecovered by that time—more than 200 kyr after the K/Pgboundary.
Kuhnt and Kaminski (1993) reported a similar faunalturnover in the northern Spanish Zumaya and Sopelanasections, including Gavelinella and Stensioeina-dominatedcalcareous assemblages in the lower part of the boundaryclay and a strong increase in the abundance of Spiroplec-tammina israelskyi at the top of this clay layer. They doc-umented a drastic increase in the relative abundance ofopportunistic species just after the K/Pg boundary duringa period of reduced primary productivity, and a gradualreestablishment of a normal trophic structure that tookplace, at least several hundred thousand years after the K/Pg boundary. A similar pattern of recovery has been re-ported from the South Atlantic (Widmark and Malmgren,1992), Mexico (Alegret and Thomas, 2001; Alegret et al.,2001, 2002a, b), Austria (Peryt et al., 1997), Tunisia (Spei-jer and Van der Zwaan, 1996; Peryt et al., 2002), andsoutheastern Spain (Alegret et al., 2003). The benthic fo-raminiferal turnover is compatible with the collapse andrecovery of surface productivity as the result of the K/Pg-boundary mass extinction of calcareous primary produc-ers (Zachos and Arthur, 1986; Kuhnt and Kaminski, 1993;d’Hondt et al., 1998; Alegret et al., 2001, 2003). The lack ofsignificant extinction of benthic foraminifera at a time ofmass extinction of planktic fauna suggests a weak bentho-pelagic coupling at that time (Thomas et al., 1999; Alegretet al., 2001). Nevertheless, the drastic reorganization ofbenthic foraminiferal assemblages coincident with the K/Pg boundary at Bidart, and at many other sections world-wide is compatible with a geologically instantaneous,widespread, and catastrophic event, such as the impact ofa large asteroid at the K/Pg boundary.

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 581
CONCLUSIONS
Upper Maastrichtian and lower Danian benthic forami-niferal assemblages from the Bidart section (southwesternFrance) are dominated by calcareous taxa, and indicateupper–middle bathyal depths of deposition for the wholestudied interval.
Upper Maastrichtian benthic foraminiferal assemblag-es are diverse and heterogeneous, and contain a mixtureof infaunal and epifaunal morphogroups, thus indicatingmesotrophic conditions during the latest Cretaceous. Thestrong decrease in diversity and heterogeneity of the as-semblages, as well as the drastic drop in the percentage ofinfaunal morphogroups, indicate a dramatic decrease inthe food supply to the sea-bottom floor coincident with theK/Pg boundary, which is related to the collapse of the foodweb triggered by the mass extinction of primary produc-ers. The benthic foraminiferal turnover, together with thegeochemical and mineralogical evidence, is compatiblewith an asteroid impact at the K/Pg boundary.
Primary productivity during the early Danian was dom-inated by blooms of non-calcareous primary producers;such a change in phytoplankton composition induced astressful environment for the benthic fauna. The low di-versity of the assemblages, as well as the low percentage ofinfaunal morphogroups through the Pv. eugubina and Ps.pseudobulloides Biozones, suggests that primary produc-tivity had not completely recovered more than 200 kyr af-ter the K/Pg boundary.
ACKNOWLEDGEMENTS
This research was funded by the DGES BTE2001–1809project (Spanish Ministry of Science and Technology), andthe DGA 2002–4 project. L. Alegret thanks the SpanishMinisterio de Educacion, Cultura y Deporte for her post-doctoral grant EX2003–0007. We thank S. J. Culver andM. B. Hart for their constructive reviews.
REFERENCES
ABRAMOVICH, S., and KELLER, G., 2002, High stress late Maastrich-tian paleoenvironment: inference from planktic foraminifera inTunisia: Palaeogeography, Palaeoclimatology, Palaeoecology, v.178, p. 145–164.
ALEGRET, L., ARENILLAS, I., ARZ, J.A., LIESA, C., MELENDEZ, A., MO-LINA, E., SORIA, A.R., and THOMAS, E., 2002a, The Cretaceous/Ter-tiary boundary: sedimentology and micropaleontology at El Mu-lato section, NE Mexico: Terra Nova, v. 14, p. 330–336.
ALEGRET, L., ARENILLAS, I., ARZ, J.A., and MOLINA, E., 2002b, Envi-ronmental changes triggered by the K/T impact event at Coxqui-hui (Mexico) based on foraminifers: Neues Jahrbuch fur Geologieund Palaontologie, v. 5, p. 295–309.
ALEGRET, L., MOLINA, E., and THOMAS, E., 2001, Benthic foraminiferaat the Cretaceous/Tertiary boundary around the Gulf of Mexico:Geology, v. 29, p. 891–894.
ALEGRET, L., MOLINA, E., and THOMAS, E., 2003, Benthic foraminif-eral turnover across the Cretaceous/Tertiary boundary at Agost(southeastern Spain): paleoenvironmental inferences: Marine Mi-cropaleontology, v. 48, p. 251–279.
ALEGRET, L., and THOMAS, E., 2001, Upper Cretaceous and lower Pa-leogene benthic foraminifera from northeastern Mexico: Micropa-leontology, v. 47, p. 269–316.
ALVAREZ, L.W., ALVAREZ, W., ASARO, F., and MICHEL, H.V., 1980, Ex-traterrestrial cause for the Cretaceous–Tertiary extinction: Sci-ence, v. 208, p. 1195–1208.
APELLANIZ, E., BACETA, J.I., BERNAOLA-BILBAO, G., NUNEZ-BETELU,
K., ORUE-ETXEBARRıA, X., PAYROS, A., PUJALTE, V., ROBIN, E., andROCCHIA, R., 1997, Analysis of uppermost Cretaceous–lowermostTertiary hemipelagic successions in the Basque Country (westernPyrenees): evidence for a sudden extinction of more than halfplanktic foraminifer species at the K/T boundary: Bulletin de laSociete Geologique de France, v. 168, p. 783–793.
ARENILLAS, I., ARZ, J.A., and MOLINA, E., 2004, A new high-resolutionplanktic foraminiferal zonation and subzonation for the lowerDanian: Lethaia, v. 37, p. 79–95.
BERGGREN, W.A., and AUBERT, J., 1975, Paleocene benthonic forami-niferal biostratigraphy, paleobiogeography and paleoecology of At-lantic–Tethyan regions; Midway-type fauna: Palaeogeography,Palaeoclimatology, Palaeoecology, v. 18, p. 73–192.
BERGGREN W.A., KENT D.V., SWISHER, C.C., and AUBRY, M.P., 1995,A revised Cenozoic geochronology and chronostratigraphy: inBerggren, W.A., Kent, D.V., Aubry, M.P., and Hardenbol, J., eds.,Geochronology, Time Scales and Global StratigraphicCorrelation:Society of Economic Paleontologists and Mineralogists SpecialPublication 54, Tulsa, p. 129–212.
BERNHARD, J.M., 1986, Characteristic assemblages and morphologiesof benthic foraminifera from anoxic, organic-rich deposits; Juras-sic through Holocene: Journal of Foraminiferal Research, v. 16, p.207–215.
BONTE, P., DELACOTTE, O., RENARD, M., LAJ, C., BOCHET, D., JEHAN-NO, C., and ROCCHIA, R., 1984, An iridium-rich layer at the Cre-taceous/Tertiary boundary in the Bidart section (southernFrance): Geophysical Research Letters, v. 11, p. 473–476.
CORLISS, B.H., 1985, Microhabitats of benthic foraminifera withindeep-sea sediments: Nature, v. 314, p. 435–438.
CORLISS, B.H., 1991, Morphology and microhabitat preferences ofbenthic foraminifera from the northeast Atlantic Ocean: MarineMicropaleontology, v. 17, p. 195–236.
CORLISS, B.H., and CHEN, C., 1988, Morphotype patterns of Norwe-gian Sea deep-sea benthic foraminifera and ecological implica-tions: Geology, v. 16, p. 716–719.
CULVER, S.J., 2003, Benthic foraminifera across the Cretaceous–Ter-tiary (K–T) boundary: a review: Marine Micropaleontology, v. 47,p. 177–226.
DELACOTTE, O., 1982, Etude magnetostratigraphique et geochemiquede la limite Cretace–Tertiaire de la coupe de Bidart (Pyrenees At-lantiques): Unpublished Ph.D. Dissertation, lUniversite Pierre etMarie Curie, Paris 6, 162 p.
DELACOTTE, O., RENARD, M., LAJ, C., PERCH-NIELSEN, K., PREMOLI-SILVA, I., and CLAUSER, S., 1985, Magnetostratigraphie et biostra-tigraphie du pass Cretace–Tertiaire de la coupe de Bidart (Pyre-nees Atlantiques): Bulletin des Centres de Recherches Geolo-giques et Minieres 1, Geologie de la France, v. 3, p. 243–254.
D’HONDT, S., DONAGHAY, P., ZACHOS, J.C., LUTTENBERG, D., and LIN-DINGER, M., 1998, Organic carbon fluxes and ecological recoveryfrom the Cretaceous–Tertiary mass extinction: Science, v. 282, p.276–279.
FONDECAVE-WALLEZ, M.J., PEYBERNES, B., and EICHENE, P., 1995,Mise en evidence micropaleontologique dun flysch ypreso-lute-tien, excretace, a materiel triasique resedimente, dans la coupe deBidart Nord (Pyrenees atlantiques, Sud–Ouest de la France): Geo-logie de la France, v. 3, p. 47–56.
FONTANIER, C., JORISSEN, F.J., LICARI, L., ALEXANDRE, A., ANSCHUTZ,P., and CARBONEL, P., 2002, Live benthic foraminiferal faunasfrom the Bay of Biscay: faunal density, composition and microhab-itats: Deep-Sea Research I, v. 49, p. 751–785.
HASLETT, S.K., 1994, Planktonic foraminiferal biostratigraphy andpaleoceanography of the Cretaceous–Tertiary boundary section atBidart, South-west France: Cretaceous Research, v. 15, p. 179–192.
HILDEBRAND, A.R., PENFIELD, G.T., KRING, D.A., PILKINGTON, M.,CAMARGO, Z.A., JACOBSEN, S.B., and BOYNTON, W.V., 1991, Chi-cxulub crater: a possible Cretaceous–Tertiary boundary impactcrater on the Yucatan Peninsula, Mexico: Geology, v. 19, p. 867–871.
HUBER, B.T., MACLEOD, K.G., and NORRIS, R.D., 2002, Abrupt extinc-tion and subsequent reworking of Cretaceous planktonic forami-nifera across the Cretaceous–Tertiary boundary: Evidence fromthe subtropical North Atlantic: in Koeberl, C., and MacLeod, K.G.,

582 ALEGRET ET AL.
eds., Catastrophic Events and Mass Extinctions: Impacts and Be-yond: Geological Society of America Special Paper No. 356, Geo-logical Society of America, Boulder, p. 227–289.
ITTEKOT, V., 1993, The abiotically driven biological pump in the oceanand short-term fluctuations in atmospheric CO2 contents: Globaland Planetary Change, v. 8, p. 17–25.
JONES, R.W., and CHARNOCK, M.A., 1985, ‘‘Morphogroups’’ of agglu-tinated foraminifera; their life positions and feeding habits andpotential applicability in (paleo)ecological studies: Revue de Paleo-biologie, v. 4, p. 311–320.
JORISSEN, F.J., STIGTER, H.C., and WIDMARK, J.G.V., 1995, A concep-tual model explaining benthic foraminiferal microhabitats: Ma-rine Micropaleontology, v. 26, p. 3–15.
KAIHO, K., KAJIWARA, Y., TAZAKI, K., UESHIMA, M., TAKEDA, N., KA-WAHATA, H., ARINOBU, T., ISHIWATARI, R., HIRAI, A., and LAMOL-DA, M.A., 1999, Oceanic primary productivity and dissolved oxy-gen levels at the Cretaceous/Tertiary boundary: their decrease,subsequent warming, and recovery: Paleoceanography, v. 14, p.511–524.
KAMINSKI, M.A., KUHNT, W., and RADLEY, J.D., 1996, Paleocene–Eo-cene deep water agglutinated foraminifera from the NumidianFlysch (Rif, Northern Morocco): their significance for the paleo-ceanography of the Gibraltar gateway: Journal of Micropaleontol-ogy, v. 15, p. 1–19.
KAMINSKI, M.A., and MALMGREN, B.A., 1989, Stable isotope and traceelement stratigraphy across the Cretaceous/Tertiary boundary inDenmark: Geologiska Foreningens i Stockholm Forhandlingar, v.111, p. 305–312.
KELLER, G., 1989a, Extended Cretaceous/Tertiary boundary extinc-tions and delayed population changes in planktonic foraminiferafrom Brazos River, Texas: Paleoceanography, v. 4, p. 287–332.
KELLER, G., 1989b, Extended period of extinctions across the Creta-ceous/Tertiary boundary in planktonic foraminifera of continentalshelf sections: implications for impact and volcanism theories:Geological Society of America Bulletin, v. 101, p. 1408–1419.
KELLER, G., LE, L., and MACLEOD, N., 1995, The Cretaceous–Tertiaryboundary stratotype section at El Kef, Tunisia: how catastrophicwas the mass extinction?: Palaeogeography, Palaeoclimatology,Palaeoecology, v. 119, p. 221–254.
KUHNT, W., and KAMINSKI, M.A., 1993, Changes in the communitystructure of deep water agglutinated foraminifers across the K/Tboundary in the Basque Basin (Northern Spain): Revista Espano-la de Micropaleontologıa, v. 25, p. 57–92.
LOEBLICH, A.R., and TAPPAN, H., 1988, Foraminiferal Genera andTheir Classification: Van Nostrand Reinhold Company, NewYork, 1182 p.
LUTERBACHER, H.P., and PREMOLI-SILVA, I., 1964, Biostratigrafia dellimite Cretaceo–Terziario nell’Appennino centrale: Rivista Itali-ana di Paleontologia, v. 70, p. 67–117.
LY, A., and KUHNT, W., 1994, Late Cretaceous benthic foraminiferalassemblages of the Casamanca Shelf (Senegal, NW Africa): indi-cation of a late Cretaceous oxygen minimum zone: Revue de Mi-cropaleontologie, v. 37, p. 49–74.
MACNEIL, D.H., and CALDWELL, W.G.E., 1981, Cretaceous rocks andtheir foraminifera in the Manitoba Escarpment: Geological Asso-ciation of Canada Special Publication 21, 435 p.
MOLINA, E., ALEGRET, L., ARENILLAS, I., and ARZ, J.A., 2002, Taphon-omy in analysis of the pattern of extinction at the Cretaceous/Ter-tiary boundary: in De Renzi, M., Pardo, M.V., Belinchon, M., Pen-alver, E., Montoya, P., and Marquez-Aliaga, A., eds., Current Top-ics on Taphonomy and Fossilization: Encontres Collection, Barce-lona, v. 5, p. 89–95.
MOLINA, E., ARENILLAS, I., and ARZ, J.A., 1998, Mass extinction inplanktic foraminifera at the Cretaceous/Tertiary boundary in sub-tropical and temperate latitudes: Bulletin de la Societe Geologiquede France, v. 169, p. 351–363.
MURRAY, J.W., 1991, Ecology and Palaeoecology of Benthic Forami-nifera: Elsevier, Amsterdam, 397 p.
NELSON, B.K., MACLEOD, G.K., and WARD, P.D., 1991, Rapid changein strontium isotopic composition of sea water before the Creta-ceous/Tertiary boundary: Nature, v. 351, p. 644–647.
NYONG, E.E., and OLSSON, R.K., 1984, A paleoslope model of Cam-panian to Lower Maestrichtian foraminifera in the North Ameri-
can basin and adjacent continental margin: Marine Micropaleon-tology, v. 8, p. 437–477.
PERCH-NIELSEN, K., 1979, Calcareous nannofossils at the Cretaceous/Tertiary boundary near Biarritz, France: in Christensen, W.K.,ed., Cretaceous–Tertiary Boundary Events: University of Copen-hagen, Copenhagen, v. 2, p. 151–155.
PERYT, D., ALEGRET, L., and MOLINA, E., 2002, The Cretaceous/Paleo-gene (K/P) boundary at Aın Settara, Tunisia: restructuring of ben-thic foraminiferal assemblages: Terra Nova, v. 14, p. 101–107.
PERYT, D., LAHODYNSKY, R., and DURAKIEWICZ, T., 1997, Deep-wateragglutinated foraminiferal changes and stable isotope profilesacross the Cretaceous/Paleogene boundary in the Rotwandgrabensection, Eastern Alps (Austria): Palaeogeography,Palaeoclimatol-ogy, Palaeoecology, v. 132, p. 287–307.
PEYBERNES, B., FONDECAVE-WALLEZ, M.J., EICHENE, P., BOST, J.,SIBE, B., MARAIS, M., and QUILES, G., 1996, La Limite Cretace–Paleocene: Phenomenes Biologiques, Evenements GeologiquesdApres les Sites de la Cote Basque: Centre Regional de Documen-tation Pedagogique dAquitaine, Bordeaux, 91 p.
PUJALTE, V., BACETA, J.I., PAYROS, A., ORUE-ETXEBARRIA, X., andSERRA-KIEL, J., 1994, Late Cretaceous–Middle Eocene sequencestratigraphy and biostratigraphy of the SW and W Pyrenees(Pamplona and Basque Basins, Spain): Field-Seminar of the Grou-pe d’etude du Paleogene and IGCP Project 286, Barcelona, 118 p.
RENARD, M., DELACOTTE, O., and LETOLLE, R., 1982, Le strontium etles isotopes stables dans les carbonates totaux de quelques sites delAtlantique et de la Tethys: Bulletin de la Societe Geologique deFrance, v. 14, p. 519–534.
ROHL, U., OGG, J.G., GEIB, T.L., and WEFER, G., 2001, Astronomicalcalibration of the Danian time scale: in Kroon, D., Norris, R.D.,and Klaus, A., eds., Western North Atlantic Palaeogene and Cre-taceous Palaeoceanography: Geological Society Special Publica-tion No. 183, p. 163–183.
ROMEIN, A.J.T., and SMIT, J., 1981, The Cretaceous/Tertiary bound-ary: calcareous nannofossil and stable isotopes: Proceedings Kon-inklijke Nederlandse Akademie van Wetenschappen, v. 84, p.295–314.
SEUNES, J., 1891, Recherches geologiques sur les terrains secondaireset lEocene inferieur de la region sous-pyreneenne du Sud-Ouestde la France (Basses-Pyrenees et Landes): Annuales Minerales,Paris, v. 18, 250 p.
SEYVE, C., 1984, Le passage Cretace–Tertiaire a Pont Labau:Bulletindes Centres de Recherches Exploration-Production Elf-Aquitaine,v. 8, p. 385–423.
SEYVE, C., 1990, Nannofossil biostratigraphy of the Cretaceous–Ter-tiary boundary in the French Basque Country: Bulletin des Cen-tres de Recherches Exploration-Production Elf-Aquitaine, v. 14, p.553–572.
SHARPTON, V.L., DALRYMPLE, G.B., MARıN, L.E., RYDER, G., SCHU-RAYTZ, B.C., and URRUTIA-FUCUGAUCHI, J., 1992, New links be-tween the Chicxulub impact structure and the Cretaceous/Tertia-ry boundary: Nature, v. 359, p. 819–821.
SMIT, J., 1982, Extinction and evolution of planktonic foraminiferaaf-ter a major impact at the Cretaceous/Tertiary boundary: in Silver,L. T., and Schultz, P.H., eds., Geological Implications of Impacts ofLarge Asteroids and Comets on the Earth: Geological Society ofAmerica Special Paper, v. 190, p. 329–352.
SMIT, J., 1990, Meteorite impact, extinctions and the Cretaceous–Ter-tiary Boundary: Geologie en Mijnbouw, v. 69, p. 187–204.
SMIT, J., and HERTOGEN, J., 1980, An extraterrestrial event at theCretaceous–Tertiary boundary: Nature, v. 285, p. 198–200.
SMIT, J., and TEN KATE, W.G.H.Z., 1982, Trace-element patterns atthe Cretaceous–Tertiary boundary: consequences of a large im-pact: Cretaceous Research, v. 3, p. 307–332.
SORIA, A.R., LIESA, C., MATA, M.P., ARZ, J.A., ALEGRET, L., ARENIL-LAS, I., and MELENDEZ, A., 2001, Slumping and a sandbar depositat the K/T boundary in the El Tecolote section (northeastern Mex-ico): an impact-induced sediment gravity flow: Geology, v. 29, p.231–234.
SPEIJER, R.P., and VAN DER ZWAAN, G.J., 1996, Extinction and survi-vorship of Southern Tethyan benthic foraminifera across the Cre-taceous/Paleogene boundary: in Hart, M. B., ed., Biotic Recovery

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 583
from Mass Extinction Events: Geological Society Special Publica-tion 102, p. 343–371.
THOMAS, E., 1990a, Late Cretaceous through Neogene deep-sea ben-thic foraminifers (Maud Rise, Weddell Sea, Antarctica): in Barker,P.F., Kennett, J.P., O’Connell, S.B., and Pisias, N.G., eds., Pro-ceedings ODP Scientific Results 113: College Station, Texas,Ocean Drilling Program, p. 571–594.
THOMAS, E., 1990b, Late Cretaceous–early Eocene mass extinctionsin the deep sea: in Sharpton, V.L. and Ward, P.W., eds., Global Ca-tastrophes in Earth History: The Proceedings of an Interdisciplin-ary Conference on Impacts, Volcanism and Mass Mortality: Geo-logical Society of America Special Publication 247, Geological So-ciety of America, Boulder, p. 481–495.
VAN MORKHOVEN, F.P.C.M., BERGGREN, W.A., and EDWARDS, A.S.,1986, Cenozoic cosmopolitan deep-water benthic foraminifera:Bulletin des Centres de Recherches Exploration-Production Elf-Aquitaine, Pau, v. 11, 421 p.
VERITY, P.G., STOECKER, D.K., SIERACK, M.E., and NELSON, J.R.,1996, Microzooplankton grazing of primary production at 1408Win the equatorial Pacific: Deep Sea Research, v. 43, p. 1227–1255.
WIDMARK, J.G.V., 2000, Biogeography of terminal Cretaceous benthicforaminifera: Deep-water circulation and trophic gradients in thedeep South Atlantic: Cretaceous Research, v. 21, p. 367–379.
WIDMARK, J., and MALMGREN, B., 1992, Biogeography of terminalCretaceous deep-sea benthic foraminifera from the Atlantic andPacific oceans: Palaeogeography, Palaeoclimatology, Palaeoecolo-gy, v. 92, p. 375–405.
ZACHOS, J.C., and ARTHUR, M.A., 1986, Paleoceanography of the Cre-taceous/Tertiary event: Inferences from stable isotope and otherdata: Paleoceanography, v. 1, p. 5–26.
ACCEPTED APRIL 14, 2004
584 ALEGRET ET AL.A
PP
EN
DIX
Ben
thic
fora
min
ifer
alsp
ecie
sco
un
tsin
the
stu
died
sam
ples
from
Bid
art.
Th
en
um
ber
ofea
chsa
mpl
e(a
bove
)co
rres
pon
dsto
the
cen
tim
eter
sbe
low
(2)
orab
ove
(1)
the
K/P
gbo
un
dary
.
K2
400
K2
300
K2
200
K2
100
K2
50K
225
K2
6210
K2
022
T1
012
T1
316
T1
131
17T
125
129
T1
50T
111
7T
115
5T
116
5T
121
0T
130
0T
136
0
Ala
bam
ina
ct.
mud
way
ensi
sA
llom
orph
ina
ct.
nav
arro
ana
All
omor
phin
asp
.A
llom
orph
ina
vela
scoe
nsi
sA
mm
odis
cus
cret
aceu
s
5 6 14
11
2 3 1
2 1
51 1
31
Am
mod
iscu
sgl
abra
tus
Am
mod
iscu
ssp
.A
ngu
loga
veli
nell
aav
nim
elec
hi
Ano
mal
inoi
des
acut
us
Ano
mal
inoi
ds
amm
onoi
des
1 2
1 4
1 6
1 10
1 5
1 1 5
72 5 5 2 4
12 3
6 27 36
2 1 25 214 3
92
93
1A
nom
alin
oide
ssp
p.A
rago
nia
sp.
Ara
gon
iave
lasc
oen
sis
Are
nobu
lim
ina
sp.
Ast
acol
us
cf.
jarv
isi
4 4
8 2
15
2 2
5
2
2 3
3
2 1
2 7 120 3
28 8
5 10 1
3 12 2
9 2 2
7 8 2
22 6
Ast
acol
us
spp.
Bat
hysi
phon
sp.
Bat
hysi
phon
sp.
CB
oliv
inoi
des
deli
catu
lus
Bol
ivin
oide
sd
raco
2 11 1
1 2 11
2 2 1 6
1 1 47
1 7
1 4
1 23
7 2 4
1 1 5
4 12 13
4
1
11
Bol
ivin
oide
ssp
.B
uli
min
am
idw
ayen
sis
Bu
lim
ina
pale
ocen
ica
Bu
lim
inia
spp.
Bu
lim
ina
trin
itat
ensi
s1
11
7 51 3 1 1
1
22
4 811
3 84
4 64 9
15 8B
uli
min
ave
lasc
oen
sis
Bu
lim
inel
lacf
.be
aum
onti
Bu
lim
inel
lasp
.C
auda
mm
ina
sp.
Cib
icid
oide
scf
.ab
udu
rben
sis
1
3 71 1
2 1
44 1
12
Cib
icid
oide
scf
.ek
blom
iC
ibic
idoi
des
cf.
nar
anjo
ensi
sC
ibic
idoi
des
dayi
Cib
icid
oide
sho
wel
liC
ibac
idoi
des
hyph
alu
s
1 51 7
3 2 15
8 1 253
4124
2 35
10 90
6
221
4151
2027
13 15
2 36
5 31
1 45
1 19C
ibic
idoi
des
prop
riu
sC
ibic
idoi
des
sp.
Cib
icid
oide
sve
lasc
oen
sis
Cla
vuli
noid
esam
orph
aC
.am
orph
aju
ven
ile
form
2312 1
10 36
9 5
8 4
14 1 1 15
15 6 1217
418 5
12 9
4 21 247
12 12
31 56 15
23 52 8
Cla
vuli
noid
essp
.C
lavu
lino
ides
tril
ater
aC
.tr
ilat
era
juve
nil
efo
rmC
anor
bin
asp
.C
oryp
host
oma
decu
rren
s
3 12
11
1 2
6 45
25
3 5
4 2
1
10
24 3
7
Cor
ypho
stom
ain
cras
sata
C.
incr
assa
tafo
rma
giga
nte
aC
oryp
host
oma
sp.
Den
tali
na
sp.
Dor
oth
iabu
llet
a
16 8 2
3 1 1 1
3 1 3
18 13 2
4 1
3 1
19 1
6910
0 1
657 1
5 14
1

RECOVERY POST CRETACEOUS/PALEOGENE BOUNDARY, BIDART, FRANCE 585
AP
PE
ND
IXC
onti
nu
ed. K
240
0K
230
0K
220
0K
210
0K
250
K2
25K
262
10K
202
2T
101
2T
131
6T
113
117
T1
251
29T
150
T1
117
T1
155
T1
165
T1
210
T1
300
T1
360
Dor
oth
iabu
llet
aju
ven
ile
form
Dor
oth
iapu
paD
orot
hia
pupa
juve
nil
efo
rmD
orot
hia
spp.
Ell
ipso
dim
orph
ina
sp.
4 2 53
3
7 3
2 1 11
8
1 3 11
12
Fis
suri
na
spp.
Fu
rsen
koin
asp
.G
aud
ryin
apy
ram
idat
aG
aud
ryin
asp
.ju
ven
ile
form
Gav
elin
ella
dan
ica
3 2
1 2
2 1 6
2 2 21
1 4 24 4
22
1 11 1
4
Gla
ndu
lin
asp
.G
lobo
rota
lite
ssp
.G
lobu
lin
apr
isca
Glo
buli
na
spp.
Glo
mos
pira
gord
iali
s
18 1 6
8 5
3 65 1 3
4 5
10 3
5 1 1 12
1 26
55 1
100
6610
11 2
244
1 8 2
12
Glo
mos
pira
serp
ens
calc
areo
us
isom
orph
Glo
mos
pira
spp.
Glo
mos
pire
lla
grzy
bow
zki
Gut
tuli
na
sp.
Gyr
oid
inoi
des
beis
seli
143 2
4 62 4
1 15
3 2 24
1 3 2232
39
3 16
2 2 28
1 1 22
9 12
15 20
14 2 17
1 1853
1 153 16
Gyr
oid
inoi
des
cf.
goud
koffi
Gyr
oid
inoi
des
depr
essu
sG
yroi
din
oide
sgi
rard
anu
sG
yroi
din
oide
sgl
obos
us
Gyr
oid
inoi
des
sp.
5 4 2
3 1 2
2 5 4 3
3 8
2 1 64 3 5
4 3 1 1
1 2 1 1
5 5 21
2 2 1
6 4 2 1
38 4 2 6
1 2 3 35 7
5 2 1
2 2 2 4 6
5 7 3H
aplo
phra
gmoi
des
prot
eus
Hap
loph
ragm
oide
ssp
.H
eron
alle
nia
lin
gula
taH
orm
osin
ave
lasc
oen
sis
Hyp
eram
min
asp
.1
21
1
11
15 1
5 3
1
31
2
Insc
ulp
tare
nula
sp.
Lae
vide
nta
lin
asp
p.L
agen
agl
obos
aL
agen
asp
p.L
enti
culi
na
spp.
26 1 21
19 2 6
25 4 10
16 2 4 7
14 10 11
9 7 4
2 24 5 20
5 4
3 6
5 7
8 13
7 17
5 8
3 2
4 2 5
4 2 42
4 7
8 7 12L
ingu
lin
asp
p.L
oxos
tom
um
eley
iM
argi
nuli
na
sp.
Mar
sson
ella
oxyc
ona
M.
oxyc
ona
juve
nil
efo
rm
1 18 5 2
39 1 2
34 3 1
24 3
18 3
14 11 6
1 242
147 2
1
1 54 2
Mar
sson
ella
sp.
Neo
flab
elli
na
sp.
Non
ione
lla
cret
acea
Non
ione
lla
sp.
Nut
tall
ides
spp.
1 1 1 5
3 1 1 1213
1227
2 1 29
1 19
2 810
1618
4138
50
2 3819
618
14N
utta
llid
estr
uem
pyi
Nut
alli
nell
aco
ronu
laN
utta
llin
ella
flore
alis
Nut
alli
nell
asp
p.O
olin
asp
p.
12 3
5 5 1
16 10
11 313 9 4
9 7
8 5
3 61 1
35
158
9
2437 4
33 36
45 7 18

586 ALEGRET ET AL.A
PP
EN
DIX
Con
tin
ued
. K2
400
K2
300
K2
200
K2
100
K2
50K
225
K2
6210
K2
022
T1
012
T1
316
T1
131
17T
125
129
T1
50T
111
7T
115
5T
116
5T
121
0T
130
0T
136
0
Ori
dors
alis
plu
mm
erae
Ori
dors
alis
sp.
Ori
dors
alis
um
bon
atu
sO
san
gula
ria
cf.
vela
scoe
nsi
sO
san
gula
ria
cf.
cord
ieri
ana
5 3
4 31
2 2
1 2 320
12 1
2 61
22 3 2
3
Osa
ngu
lari
acf
.pl
um
mer
aeO
san
gula
ria
spp.
Pal
iola
tell
aor
bign
yan
aP
alio
late
lla
sp.
Par
alab
amin
ah
ille
bran
dti
3 2
4 1 2
3 2
1 32 3
2 2 2
5 6 3
822
1939
16 35 2
18 4
8 21 47
11 3558
22 17
6 5
8 2 17P
lanu
lin
asp
.P
leu
rost
omel
lasp
.P
raeb
uli
min
acf
.re
uss
iP
raeb
uli
min
asp
.P
raeg
lobo
bulm
ina
spp.
28 1
2 142 8
2 3 12
1 5
21
14
Pse
udou
vige
rin
apl
um
mer
aeP
seud
ouvi
geri
na
sp.
Pu
llen
iacr
etac
eaP
ull
enia
jarv
asi
Pyr
amid
ina
rud
ita
2 3
2 2
1 1 7
1 93
13 6
3
3 12
21
1 11
Pyr
amid
ina
sp.
Pyr
uli
noid
essp
.Q
uad
rim
orph
ina
allo
mor
phi-
noid
esQ
uad
rim
orph
ina
sp.
Qu
inqu
eloc
uli
na
sp.
7 2
7 2
4 1
6 2
72
16
1
6 2
Ram
uli
na
spp.
Rec
urv
oide
ssp
.R
epm
ania
char
oide
sR
euse
lla
sp.
Rh
izam
min
asp
.
1 3 1
23
11
2 4 11
31 1
1
1S
acca
mm
ina
plac
enta
Sac
cam
min
asp
p.S
arac
enar
iasp
.S
chei
bner
ova
sp.
Sit
ella
sp.
1 13
1 11
3 41
11
2
Sli
teri
ava
rsov
ien
sis
Spi
ropl
ecta
mm
ina
cf.
den
tata
Spi
ropl
ecta
mm
ina
cf.i
srae
lsky
iS
piro
plec
tam
min
asp
ecta
bili
sS
piro
plec
tam
min
asp
p.2
1 4 1
1 33
13
65 1 1
11 3 8 5
34 1 3
299
7 2
525
15 1 2
3 2S
piro
plec
tine
lla
sp.
Spi
rill
ina
sp.
Ste
nsi
oein
abe
ccar
iifo
rmis
Ste
nsi
oein
aex
cola
taS
till
osto
mel
lasp
.
1 61 2 4
1 2
6 3 1
12 1 1
1 8 2 1
4 1 1
19 211
264 2
4 1 3
3 2
5 7 2
2 21 15 1
51 7 3
1 20 1
4 57 1 1
4 58 1
63
Tapp
anin
ase
lmen
sis
Th
alm
ann
amm
ina
sp.
Tro
cham
min
asp
.Va
ginu
lin
atr
ilob
ata
Valv
alab
amin
ale
nti
cula
3 11
11
3 1
1 1
1 6 46 4
73
1 13
2
1
TO
TA
L32
119
725
427
325
828
332
523
841
665
140
146
237
334
332
831
039
334
537
0





















![arXiv:1605.03844v2 [physics.ins-det] 27 Oct 2017 · 2017. 10. 30. · oDepartment of Physics and Astronomy, University College London, Gower Street, London WC1E 6BT, United Kingdom](https://img.pdfslide.net/doc/110x75/604329c01a656e088f29426f/arxiv160503844v2-27-oct-2017-2017-10-30-odepartment-of-physics-and-astronomy.jpg)







