Embed Size (px)
Citation preview

JOURNAL OF ULTRASTRUCTURE RESEARCH 69, 307-315 (1979)
Patterns of Thylakoid System Formation 1
GIORGIO CASADORO AND NICOLETTA RASCIO
Istituto di Botanica e Fisiologia Vegetale dell'Universit~t, Via Orto Botanico n. 15, 1-35100 Padova, Italy
Received Apri l 18, 1979, and in revised form, July 9, 1979
Plastid differentiation has been studied in leaves of both tobacco and wheat. We could point out a correlation between the differentiation speed of the plastids and that of the cell containing them. The ontogenetic patterns of both tobacco and wheat plastids appear to be different from the standard ones. The significance of these particular ontogeneses is discussed with references to others in the literature.
It has been known for a long time that in many leaves their differentiation occurs in gradients, especially along their major leaf axis (N/igeli, 1846; Prantl, 1883; Avery, 1933; Foster, 1936). We have recently observed in belladonna and in sunflower that such gra- dients affect the differentiation of the chlo- roplasts which exhibit peculiar ontogenetic situations differing from the standard ones (Casadoro and Rascio, 1977; Casadoro and Rascio, in press). We have now extended our study to the plastids of other plants with foliar differentiation gradients, i.e., wheat and tobacco. We have chosen the monocotyledonous wheat because its leaves have a basal meristem which allows one to examine variously differentiated tissues along the same leaf.
Also the dicotyledonous tobacco is inter- esting to examine. According to Avery (1933) in tobacco the foliar tip already ma- tures in 3-mm long leaves. On the contrary, "marginal growth continues centrally and toward the basal end together with growth through the rest of the lamina, until the leaf is several centimeters long. The distal portion of the leaf is therefore relatively more mature, and the gradient from tip to base is a gradient from senescent to young tissues." Still in 6- to 7-cm-long leaves "the degree of maturity of all cells is dependent upon their distance from the tip and their distance from the midrib." Therefore to- bacco gives us the possibility of having not
1 Supported by a grant of CNR.
307
only successively older leaves but, above all, successively maturer tissues in the same leaf.
MATERIALS AND METHODS
Glasshouse-grown wheat (Triticum aestivum L.) plants and open air-grown plants of Nicotiana taba- cum L. from the Botanic Gardens of Padua University were used. Tobacco tissue samples were obtained, during spring, from the apical and the basal zones of successively older leaves. Wheat tissues were sampled serially from the leaf basal meristem to the leaf subap- ical zone; the samples were obtained from: the first leaf of a 7-day-old plant; the second leaf of a 14-day- old plant; the first leaf of a 21-day-old plant. All the tissue samples were fixed for 2 hr in 6% glutaraldehyde in 0.1 M cacodylate buffer (pH 6.9), washed in buffer, then postfixed 2 hr in 1% osmium tetroxide in 0.1 M cacodylate buffer (pH 6.9), and dehydrated in a graded series of ethyl alcohol and propylene oxide. Staining with uranyl acetate was carried out while dehydrating with 75% alcohol. Tissues were embedded in an Epon- Durcupan ACM mixture; thin sections, cut with an LKB Ultrotome III, were poststained with lead citrate and examined with a Hitachi HS 9 electron microscope operating at 75 kV.
RESULTS AND DISCUSSION
Tobacco
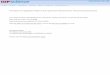
In very young leaves (about 5 mm) the mesophyll plastids of the basal zone gener- ally appeared like those in Fig. la. Rather voluminous prolamellar bodies are ob- served, which may be interconnected with thylakoid membranes. At a higher magni- fication (Fig. lb) these membranes ap- peared to be of the LS type (Casadoro and Rascio, 1978a) and to border compartments filled with very electron-dense material.
0022-5320/79/120307-09502.00/0 Copyright © 1979 by Academic Press, Inc. All rights of reproduction in any form reserved.
c~
Z

THYLAKOID SYSTEM FORMATION 309
Some plastids (Fig. la) showed an electron- dense inclusion-previously described by Stet ler and Laetsch (1969) and by other authors-whose content looked like the one of the intrathylakoid compartments .
In the basal zone of a leaf about 3 cm long the mesophyll chloroplasts showed a fairly developed thylakoid system, already arranged into grana and intergrana thyla- koids (Fig. 2). The membranes no longer appeared to be of the LS type but were strongly stained, and electron-dense mate- rial was still visible in the in t ramembranal compartments . Prolamel lar bodies were scat tered among the thylakoids.
In older leaves (about 5-6 cm) the mes- ophyll plastids of the basal zone (Fig. 3) showed a well-developed thylakoid system. The in t ramembranal compar tments ap- peared to be electron t ransparent and lim- ited by strongly stained membranes; pro- lamellar bodies were still visible.
In the basal zone of leaves about 12 cm long (Fig. 4) the mesophyll plastids looked like mature chloroplasts with abundant thylakoid systems formed by numerous grana and intergran~ thylakoids. Prolamel- lar bodies were no longer visible.
T he above ontogenetic steps were ob- served also in the apical zone of the leaves. However, concomitant with the faster mat- urat ion of the apical cells (Avery, 1933), plastid differentiation was also faster.
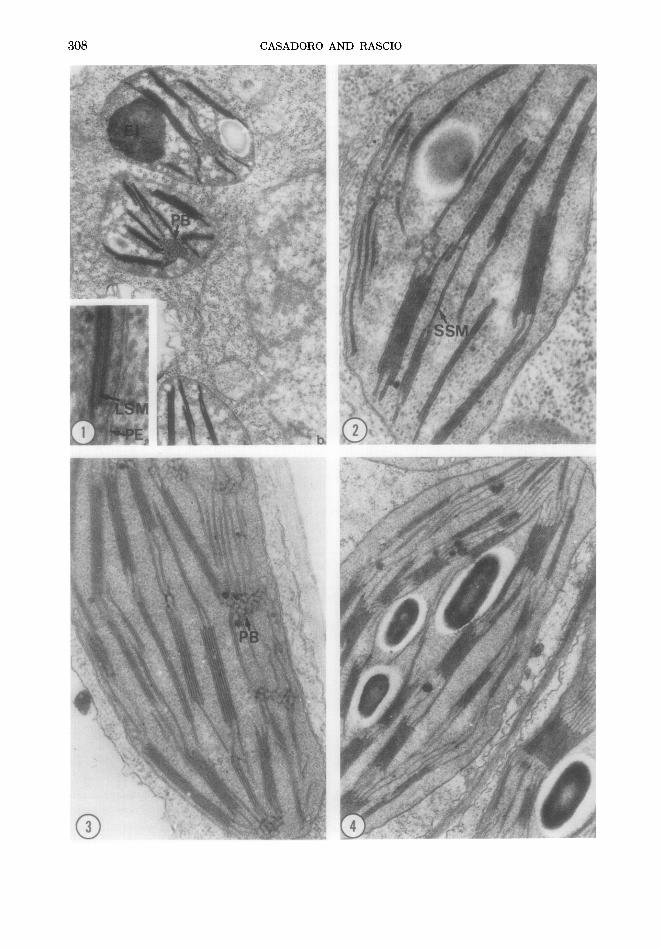
In a leaf about 3 cm long (Fig. 2 illustrates the plastids of its basal region), the chloro- plasts of its apical region (Fig. 5) showed a
well-arranged thylakoid system with strongly stained membranes and electron- t ransparent in t ramembranal compart- ments. Prolamel lar bodies were visible just as small residues.
As regards the plastids of bo th the epi- dermises, they showed a more delayed on- togenesis than the mesophyll plastids. For instance, plastids with reduced thylakoid system formed by LS membranes (Fig. 6) were still visible in the apical epidermises of a leaf about 12 cm long. However, in the longest leaves examined by us (20 cm), the epidermal plastids showed strongly stained thylakoid membranes and electron-trans- parent compartments , a l though their thy- lakoid system appeared to be less developed than the one of the mesophyll chloroplast.
Even if not marked as in the epidermal plastids, a similar delay in ontogenesis was also observed in the plastids of the small vein regions. In fact, in young leaves such plastids always appeared to be less differ- ent ia ted than those of the surrounding mes- ophyll. However, their ontogenetic stages were the same and, in adult leaves, they also showed an abundant , well-arranged thylakoid system.
The differentiation of tobacco plastids shows a characterist ic pat tern. The very young plastids have an internal membrane system formed by LS membranes which enclose electron-dense material in the in- t r amembrane compartments . During their subsequent ontogenetic stages, the LS membranes increase in stainability and, at
FIGS. 1-12. EI, electron-dense inclusion; LSM, lightly stained membranes; PB, prolamellar body; PE, plastid envelope; S, starch; SSM, strongly stained membranes.
Fins. 1-4. Plastids of the basal zone of successively older tobacco leaves. Fig. 1. Leaf about 5 mm long. Its very young plastids have a reduced membrane system and show some starch granules (a). Voluminous prolamellar bodies appear to be interconnected with thylakoid membranes. Very electron-dense material appears to fill both the intramembrane compartments and a large roundish body. A higher magnification (b) reveals that the internal membrane system is formed by LS membranes. Fig. 2. Leaf about 3 cm long. The plastid internal membranes appear to be strongly stained. Electron-dense material is still visible in the intramembrane compartments. Fig. 3. Leaf 5-6 cm long. The thylakoid membranes appear to be well arranged into grana and intergrana thylakoids, and to enclose electron-transparent compartments. Numerous prolamellar bodies are scattered among the thylakoids. Fig. 4. Leaf about 12 cm long. The chloroplast thylakoid system is very abundant and well arranged; prolamellar bodies are no longer visible. Starch granules are scattered among the thylakoids. Fig. la, × 20 000; Fig. lb, × 90 000; Fig. 2, × 48 000; Fig. 3, × 32 000; Fig. 4, × 23 000.
310 CASADORO AND RASCIO
FIGS. 5-6. Plastids of the apical zone of successively older tobacco leaves. Fig. 5. Leaf about 3 cm long. In the leaf tip the mesophyll plastids have their thylakoid systems formed by membranes which appear to be strongly stained and to enclose electron-transparent compartments. A few residual prolamellar bodies can be seen among the thylakoids. Fig. 6. Epidermal plastid of a leaf about 12 cm long. Its thylakoid system is rather reduced and formed by LS membranes. Very electron-dense material is present both in the intrathylakoid compartments and in a large roundish body. Fig. 5, × 25 000; Fig. 6, × 33 000.
the same time, the electron-dense mater ia l is reduced in s tainabil i ty until the la t ter disappears, all of which suggests t ha t the stainable mater ia l is inserted into the mem- branes themselves. This is in accordance with the observat ions recent ly made in leaves of sesame (Platt-Aloia and T h o m - son, 1977), sunflower (Casadoro and Rascio, 1978b; Mares et al., 1979) and Lippia ci- triodora L. (Casadoro and Rascio, 1978c). Moreover, it suppor ts the in terpre ta t ion by Stet ler and Laetsch (1969)--reaff irmed later by other authors (Ames and Pivorun, 1974; Henry, 1975a,b; H u r k m a n and Ken- nedy, 1977)--according to which, the ma- terial tha t they had seen and described in the m e m b r a n e - b o u n d bodies of the tobacco plastids would serve to build up thylakoid
membranes . Finally, in well-differentiated tobacco chloroplasts all the internal m e m - branes are normal ly s tained and the intra- ' m e m b r a n a l c o m p a r t m e n t s are electron t ransparent .
Bo th the presence and the long persist- ence of prolamel lar bodies in chloroplasts had a l ready been noted by Ste t ler and Lae tsch (1969) who considered the phe- nomenon to be independent of light; also H u r k m a n and Kennedy (1977) observed prolamel lar bodies, in still 6- to 8-cm-long leaves. The above-descr ibed sequences of thylakoid sys tem ma tu ra t ion is the same for all tobacco chloroplasts. On the con- trary, the speeds a t which the various de- ve lopmenta l stages occur are not the same, but differ in different leaf zones. A ma tu -

THYLAKOID SYSTEM FORMATION 311
ration gradient of the thylakoid system par- allels the already described cell maturation gradient along the major leaf axis (Avery, 1933).
Prolamellar bodies--which are just small remnants in the apical plastids and more prominent in the basal ones--also show different aspects in the same leaf. This con- firms the interpretation of Stetler and Laetsch (1969) that prolamellar bodies are independent of light and represent a normal ontogenetic feature in tobacco plastids.
A further correlation between cell matu- ration and plastid development can also be found through the leaf lamina thickness. In fact, in the small-vein regions--which, ac- cording to Avery (1933), remain meriste- matic for a longer time than the rest of the mesophyll--the plastids show a slightly slower differentiation than those of the sur- rounding mesophyll.
Therefore plastid differentiation seems to depend on cell maturation and, conse- quently, on the cellular environment in which the plastids develop. The control from the cellular environment over plastid differentiation appears somewhat stronger in those cells where the plastids, besides showing a delayed ontogenesis, succeed in building a less abundant membrane system than that of the mesophyll plastids (viz. in the epidermis).
Wheat
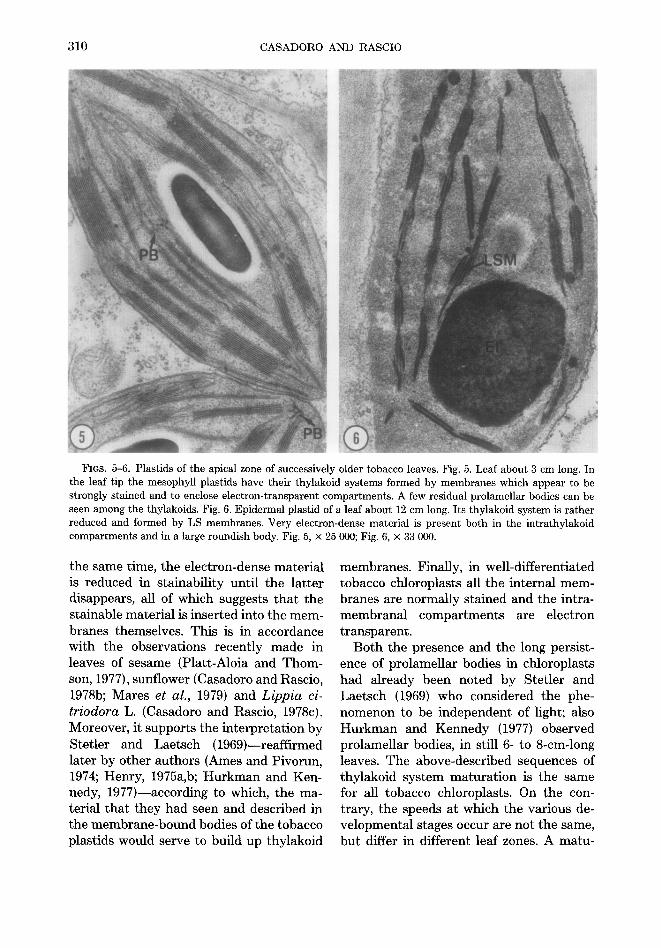
First leaf of a 7-day-old plant. In the basal meristem the proplastids showed a few, strongly stained thylakoid membranes which enclosed electron-transparent spaces (Fig. 7). The proplastids often contained large starch granules. In a region 1.5-2 cm distal to the basal meristem the plastids showed small grana together with fret membranes. In the same leaf section almost all the plastids had paracrystalline prola- mellar bodies interconnected with the thy- lakoid membranes (Fig. 8).
The paracrystalline prolamellar bodies were still ubiquitous in the plastids of ma- turer leaf-regions (3-3.5 cm from the basal
meristem). However, the thylakoid system of such plastids appeared to be well devel- oped and arranged in numerous grana and intergrana thylakoids (Fig. 9). In some chlo- roplasts of the subapical region (6-6.5 cm from the basal meristem) it was still possi- ble to see small and residual prolamellar bodies scattered among the large and nu- merous grana (Fig. 10).
First leaf of a 21-day-old plant. Just distal to the basal meristem the plastids appeared to be well differentiated with nu- merous grana and no prolamellar bodies (Fig. 11).
Second leaf of a 14-day-old plant. In the region just distal to the basal meristem the plastids appeared to be very young with a reduced thylakoid system. Well-arranged prolamellar bodies were visible among the thylakoids (Fig. 12).
Wheat chloroplasts, just like those of to- bacco, gain their full maturity by passing through an intermediate stage which is characterized by the contemporary pres- ence of both prolamellar bodies and thyla- koids. Such a stage--described and named etiochloroplast stage in maize plastids (Ras- cio et al., 1976)--persists for a long time in the young leaves (first leaf of a 7-day-old plant). On the contrary it becomes less and less evident with the increasing of leaf age, until it is no longer detectable (first leaf of a 21-day-old plant).
Such a phenomenon reappears in all the forming leaves. Therefore, owing to the characteristic developmental pattern of the wheat leaves, the presence of etiochloro- plasts appears to be related to the cell dif- ferentiation speed in a given leaf and to the age of that leaf in a given plant.
The above suggests that in wheat plas- rids, as observed in maize (Rascio et al., 1976), the etiochloroplast is a normal on- togenetic stage, since its persistence is re- lated to the maturation of the cells in which the plastids develop.
Neither LS membranes nor electron- dense intracompartment material have ever been observed in wheat plastids. When
>.
> ©
©
> > A
0
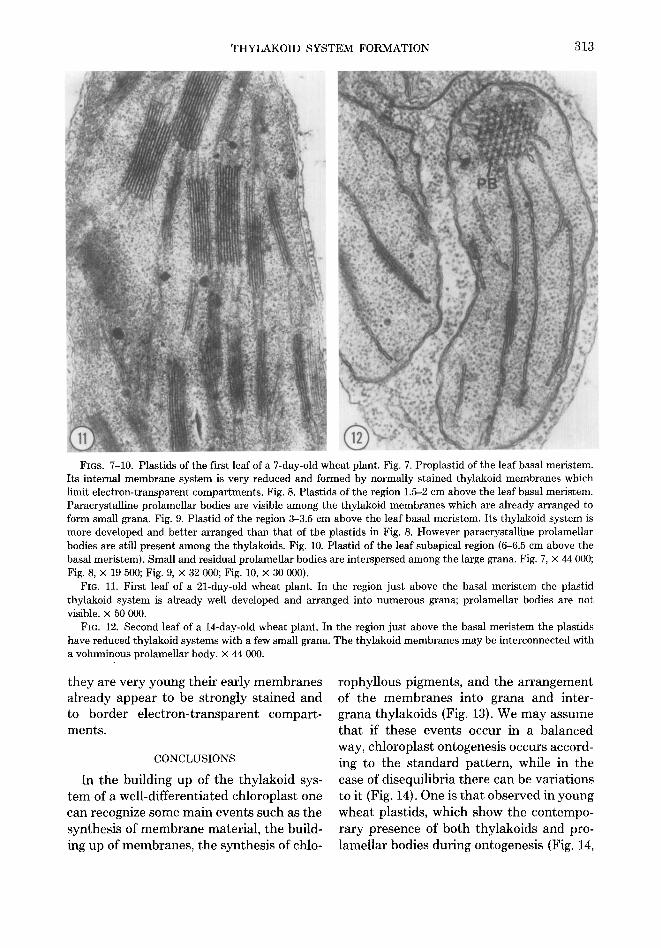
THYLAKOID SYSTEM FORMATION 313
FIGS. 7-10. Plastids of the first leaf of a 7-day-old wheat plant. Fig. 7. Froplastid of the leaf basal meristem. Its internal membrane system is very reduced and formed by normally stained thylakoid membranes which limit electron-transparent compartments. Fig. 8. Plastids of the region 1.5-2 cm above the leaf basal meristem. Paracrystalline prolamellar bodies are visible among the thylakoid membranes which are already arranged to form small grana. Fig. 9. Plastid of the region 3-3.5 cm above the leaf basal meristem. Its thylakoid system is more developed and better arranged than that of the plastids in Fig. 8. However paracrystalline prolamellar bodies are still present among the thylakoids. Fig. 10. Plastid of the leaf subapical region (6-6.5 cm above the basal meristem). Small and residual prolamellar bodies are interspersed among the large grana. Fig. 7, × 44 000; Fig. 8, × 19 500; Fig. 9, × 32 000; Fig. 10, × 30 000).
FIG. 11. First leaf of a 21-day-old wheat plant. In the region just above the basal meristem the plastid thylakoid system is already well developed and arranged into numerous grana; prolamellar bodies are not visible. × 50 000.
FIG. 12. Second leaf of a 14-day-old wheat plant. In the region just above the basal meristem the plastids have reduced thylakoid systems with a few small grana. The thylakoid membranes may be interconnected with a voluminous prolameUar body. x 44 000.
t hey are very y o u n g the i r ear ly m e m b r a n e s a l ready appea r to be s t rongly s t a ined a n d to bo rde r e l e c t r o n - t r a n s p a r e n t compar t -
men t s .
CONCLUSIONS
In the bu i ld ing up of the t hy l ako id sys-
t e m of a wel l -d i f fe ren t ia ted ch loroplas t one
can recognize some m a i n even t s such as the
syn thes i s of m e m b r a n e mater ia l , the bui ld-
ing up of m e m b r a n e s , t he syn thes i s of chlo-
rophy l lous p igments , a nd the a r r a n g e m e n t of the m e m b r a n e s in to g rana a nd in ter - g rana thy lako ids (Fig. 13). We m a y a s sume
t h a t if these even t s occur in a b a l a n c e d
way, ch lorop las t on togenes i s occurs accord-
ing to the s t a n d a r d pa t t e rn , whi le in the
case of d i sequ i l ib r ia the re can be va r i a t ions
to it (Fig. 14). One is t h a t observed in young
whe a t plast ids, which show the c on t e mpo-
ra ry p resence of b o t h thy lako ids a n d pro-
l ame l l a r bodies du r ing on togenes i s (Fig. 14,
314
PROPLASTID
CASADORO AND RASCIO
I) Membrs~ne material synthesis
Lightly stained 2) Membrane /
building up "~gtrongly stained WELL-DIFFERENTIATED
CHLOROPLAST 3) Chlorophylls
synthesis
~ 4) Membrane/Prilamellar body arrangement
"~Gran a
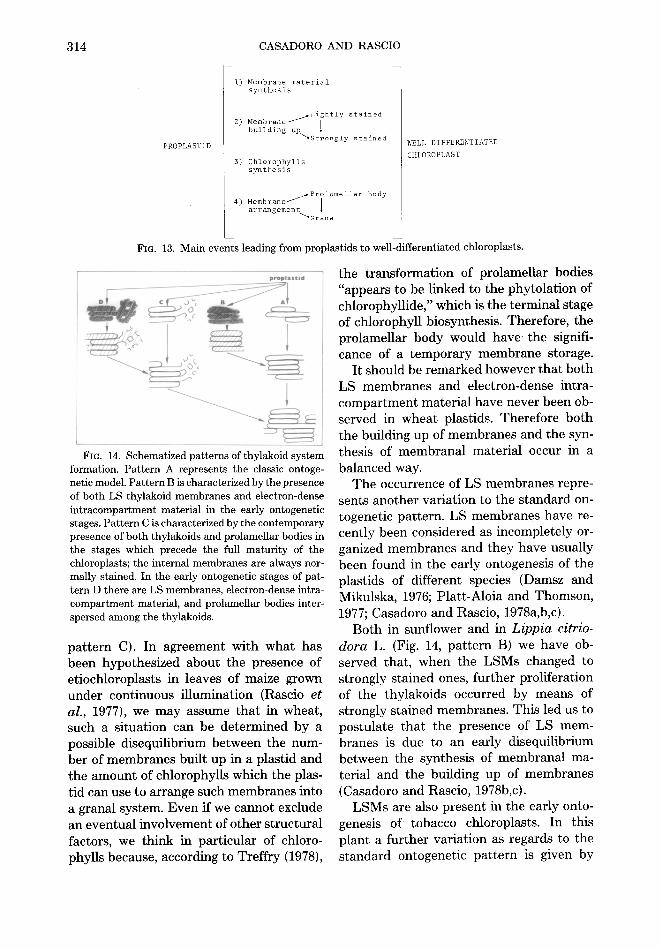
Fro. 13. Main events leading from proplastids to well-differentiated chloroplasts.
proplsstld
J 1
Fro. 14. Schematized patterns of thylakoid system formation. Pattern A represents the classic ontoge- netic model. Pattern B is characterized by the presence of both LS thylakoid membranes and electron-dense intracompartment material in the early ontogenetic stages. Pattern C is characterized by the contemporary presence of both thylakoids and prolamellar bodies in the stages which precede the full maturity of the chloroplasts; the internal membranes are always nor- mally stained. In the early ontogenetic stages of pat- tern D there are LS membranes, electron-dense intra- compartment material, and prolamellar bodies inter- spersed among the thylakoids.
pa t te rn C). In agreement with what has been hypothesized about the presence of etiochloroplasts in leaves of maize grown under continuous illumination (Rascio et al., 1977), we may assume tha t in wheat, such a si tuation can be determined by a possible disequilibrium between the num- ber of membranes built up in a plastid and the amount of chlorophylls which the plas- tid can use to arrange such membranes into a granal system. Even if we cannot exclude an eventual involvement of other structural factors, we think in part icular of chloro- phylls because, according to Treff ry (1978),
the t ransformat ion of prolamellar bodies "appears to be linked to the phytola t ion of chlorophyllide," which is the terminal stage of chlorophyll biosynthesis. Therefore, the prolamellar body would h a v e the signifi- cance of a t emporary membrane storage.
It should be remarked however tha t both LS membranes and electron-dense intra- compar tment material have never been ob- served in wheat plastids. Therefore both the building up of membranes and the syn- thesis of membranal material occur in a balanced way.
The occurrence of LS membranes repre- sents another variat ion to the s tandard on- togenetic pat tern. LS membranes have re- cently been considered as incompletely or- ganized membranes and they have usually been found in the early ontogenesis of the plastids of different species (Damsz and Mikulska, 1976; Platt-Aloia and Thomson, 1977; Casadoro and Rascio, 1978a,b,c).
Both in sunflower and in Lippia citrio- dora L. (Fig. 14, pa t te rn B) we have ob- served that, when the LSMs changed to strongly stained ones, fur ther proliferation of the thylakoids occurred by means of strongly stained membranes. This led us to postulate tha t the presence of LS mem- branes is due to an early disequilibrium between the synthesis of membranal ma- terial and the building up of membranes (Casadoro and Rascio, 1978b,c).
LSMs are also present in the early onto- genesis of tobacco chloroplasts. In this plant a fur ther variation as regards to the s tandard ontogenetic pa t te rn is given by

THYLAKOID SYSTEM FORMATION 315
the presence of prolamellar bodies (Fig. 14, pattern D).
Such a complex ontogenesis--also ob- served in C i n n a m o m u m sp. plastids (Casa- doro and Rascio, unpublished data)-ap- pears to be very different from the classic one. We are induced to postulate that there may be present both a disequilibrium caus- ing the appearance of the LS membranes and a disequilibrium between the number of membranes and the amount of chloro- phylls. This would give rise to the occur- rence of the prolamellar bodies.
The following observations seem to sup- port the hypothesis that the variations from the standard ontogenetic pattern can be ascribed to particular disequilibria among the events which lead to the formation of a well-differentiated chloroplast, as already suggested by Whatley (1977). (1) Usually the above "particular" ontogeneses are found in leaves whose cells and tissues show various maturation speeds along the leaf major axis or throughout the lamina thick- ness. (2) Deviations from the standard pat- tern are progressively less evident as the differentiation speed of the cells in which the organelles develop increases.
In wheat plants, the older the leaves are the faster plastid differentiation is so that in wheat leaves of a given age the young plastids of the basal region do not show any prolamellar body. On the other hand, in maize leaves Rascio et al. (1976) already observed that the speed of cell ageing and, consequently, plastid differentiation speed are influenced by the age of the leaf. In tobacco leaves, during summer, the tip-to- base gradient is exhausted more rapidly than during spring (Casadoro and Rascio, unpublished observation) with a conse- quent acceleration of the chloroplast onto- genesis; we could in fact observe a rather short persistence of the LS membranes and an almost complete absence of prolamellar bodies. On the other hand Platt-Aloia and Thomson (1977) observed an influence of plant age on chloroplast ontogenesis in sesame.
It would thus appear that the plastids have an intrinsic differentiation speed. When the latter matches with the cell mat- uration speed, plastid ontogenesisproceeds in conformity with the usual ontogenetic model. On the contrary, if the cell differ- entiation speed is slowed down, there can be established anomalous plastid situations as a consequence of disequilibria between the speeds of the ontogenetic events occur- ring in the plastid and those occurring in the cell.
The display of a given ontogenetic model seems thus to depend on the more or less coordinate interactions between the plastid and the cell in the management of the events leading to the formation of a well- differentiated chloroplast.
REFERENCES
AMES, I. H., AND PIVORUN, J. P. (1974) Amer. J. Bot. 61, 794.
AVERY, G. S., JR. (1933) Amer. J. Bot. 20, 565. CASADORO, G., AND RASCIO, N. {1977) Caryologia 30,
189. CASADORO, G., AND RASClO, N. (1978a) J. Ultrastruct.
Res. 65, 30. CASADORO, G., AND RASCIO, N. (1978b) Protoplasma
97, 165. CASADORO, G., AND RASCIO, N. (1978c) Proceedings of
the 9th International Congress on Electron Micros- copy, Vol. 2, 416.
CASADORO, G., AND RASCIO, N. (1979) Cytobios, in press.
DAMSZ, B., AND MIKULSKA, E. (1976) Biochem. Phys- iol. Pf lanzen 169, 257.
FOSTER, A. S. (1936) Bot. Rev. 2, 349. HENRY, E. W. (1975a) J. Ultrastruct. Res. 52, 289. HENRY, E. M. (1975b) J. Microsc. 22, 109. HURKMAN, W. J., AND KENNEDY, G. S. (1977) Amer.
J. Bot. 64, 86. MARES, D. J., COOTE, M. A., AND POSSINGHAM, J. V.
(1979) Ann. Bot. 43, 191. N/4CELI, K. (1846) Z. Wiss. Bot. 3-4, 153. PLATT-ALOIA, K. A., AND THOMSON, W. W. (1977)
New Phytol. 78, 599. PRANTL, K. (1883) Ber. Deutsch. Ges. 1, 280. RASCIO, N., ORSENIGO, M., AND ARBOIT, D. (1976)
Protoplasma 90, 253. RASCIO, N., MARIANI COLOMBO, P., AND ORSENIGO,
M. {1977) Caryologia 30, 497. STETLER, D. A., AND LAETSCH, W. M. (1969) Amer. J.
Bot. 56, 260. TREFFRY, T. (1978) Int. Rev. Cytol. 52, 159. WHATLEY, J. M. (1977) New Phytol. 78, 407.





![Formation of morphogenetic patterns in cellular automata · Formation of morphogenetic patterns in cellular automata Manan'Iarivo Rasolonjanahary and Bakhtier Vasiev [0000-0002-3452-7885]](https://img.pdfslide.net/doc/110x75/5f0d0f6e7e708231d4387b62/formation-of-morphogenetic-patterns-in-cellular-automata-formation-of-morphogenetic.jpg)