Embed Size (px)
Citation preview

Peroxynitrite Production, DNA Breakage, and Poly(ADP-ribose) Polymerase Activation in a Mouse Model ofOxazolone-Induced Contact Hypersensitivity
EÂva SzaboÂ,*²1 LaÂszlo ViraÂg,*³1 Edina Bakondi,³ LaÂszlo GyuÈre,§ GyoÈrgy HaskoÂ,* PeÂter Bai,³JaÂnos Hunyadi,² PaÂl Gergely,³ and Csaba SzaboÂ**Inotek Corporation, Beverly, Massachusetts, U.S.A.; ²Department of Dermatology, University Medical School of Debrecen, Hungary; ³Department
of Medical Chemistry, University Medical School of Debrecen, Hungary; §Biogal Pharmaceutical Works Ltd, Debrecen, Hungary
Peroxynitrite-induced poly(ADP-ribose) polymeraseactivation has been implicated in the pathogenesis ofvarious in¯ammatory conditions. Here we haveinvestigated whether peroxynitrite and poly(ADP-ribose) polymerase may play a role in the patho-physiology of the elicitation phase of contact hyper-sensitivity. We have detected nitrotyrosine, DNAbreakage, and poly(ADP-ribose) polymerase acti-vation in the epidermis of mice in an oxazolone-induced contact hypersensitivity model. As tyrosinenitration is mostly mediated by peroxynitrite, anitric-oxide-derived cytotoxic oxidant capable ofcausing DNA breakage, we have applied peroxynitritedirectly on mouse skin and showed poly(ADP-ribose)polymerase activation in keratinocytes and in somescattered dermal cells. We have also investigated thecellular effects of peroxynitrite in HaCaT cells, ahuman keratinocyte cell line. We found that peroxy-nitrite inhibited cell proliferation and at higher con-centrations also caused cytotoxicity. Peroxynitrite
activates poly(ADP-ribose) polymerase in HaCaTcells and poly(ADP-ribose) polymerase activationcontributes to peroxynitrite-induced cytotoxicity, asindicated by the cytoprotective effect of thepoly(ADP-ribose) polymerase inhibitor 3-aminoben-zamide. The cytoprotective effect of 3-aminobenza-mide cannot be attributed to inhibition of apoptosis,as apoptotic parameters (caspase activation and DNAfragmentation) were not reduced in the presence of3-aminobenzamide in peroxynitrite-treated cells.Moreover, poly(ADP-ribose) polymerase inhibitionby 3-aminobenzamide dose-dependently reducedinterferon-induced intercellular adhesion molecule 1expression as well as interleukin-1b-induced interleu-kin-8 expression. Our results indicate that peroxyni-trite and poly(ADP-ribose) polymerase regulatekeratinocyte function and death in contacthypersensitivity. Key words: apoptosis/in¯ammation/interleukin-8/keratinocyte/necrosis. J Invest Dermatol117:74±80, 2001
Contact hypersensitivity (CHS) is a form of delayedtype hypersensitivity caused by various lipophiliccompounds (Grabbe and Schwarz, 1998). Thesensitization phase of CHS has been the focus ofinterest of dermato-immunologists for decades and it
is therefore well characterized. The elicitation phase of the responseis not well understood, however. Nitric oxide, a free radicalsynthesized from L-arginine by nitric oxide synthase enzymes(NOS), has been proposed to be a major mediator of in¯ammationin the elicitation phase of CHS (Morita et al, 1996; Ormerod et al,1997; Rowe et al, 1997; Ross et al, 1998). Enhanced expression ofthe mRNA of inducible NOS (iNOS) has been detected inLangerhans cells and keratinocytes in the elicitation phase of 2,4-dinitro¯uorobenzene-induced CHS (Ross et al, 1998). In the samemodel, as well as in picryl-chloride-induced CHS, pharmacologic
NOS inhibition suppressed the hypersensitivity reaction (Morita etal, 1996; Ross et al, 1998). The question arises, however, whetherNO directly or indirectly through the formation of a more reactiveintermediate such as peroxynitrite contributes to tissue injury inCHS.
Peroxynitrite, a cytotoxic oxidant formed in the near diffusion-limited reaction of nitric oxide and superoxide (Beckman andKoppenol, 1996; Groves, 1999), is an important mediator of tissueinjury in various forms of in¯ammation, shock, and ischemia-reperfusion injury (SzaboÂ, 1996a; 1996b; Ma et al, 1997). Thecytotoxic effect of peroxynitrite is attributed to inhibition of themitochondrial respiratory chain, inactivation of ion channels,initiation of lipid peroxidation, protein oxidation, DNA damage,and interference with protein tyrosine phosphorylation (Gow et al,1996; Kong et al, 1996; Groves, 1999). In addition, peroxynitritetriggers an indirect suicidal pathway mediated by poly(ADP-ribose)polymerase (PARP). PARP, a nuclear nick sensor enzyme, cleavesNAD+ to nicotinamide and ADP-ribose and polymerizes the latteron nuclear acceptor proteins such as histones, transcription factors,and PARP itself (de Murcia and Menissier de Murcia, 1994).Excessive PARP activation depletes cellular NAD+ and ATP poolsand results in necrotic cell death (Cochrane, 1991). Pharmacologicinhibition of PARP or the PARP±/± phenotype provided remark-
0022-202X/01/$15.00 ´ Copyright # 2001 by The Society for Investigative Dermatology, Inc.
74
Manuscript received October 18, 2000; revised March 8, 2001; acceptedfor publication March 15, 2001.
Reprint requests to: Dr. LaÂszlo ViraÂg, Inotek Corp., 100 CummingsCenter, Suite 419E, Beverly, MA 01915.Email: [email protected]
Abbreviations: 3-AB, 3-aminobenzamide; iNOS, inducible nitric oxidesynthase; PARP, poly(ADP-ribose) polymerase.
1The ®rst two authors contributed equally to the work.

able protection in disease models of hemorrhagic shock, stroke,myocardial ischemia, streptozotocin-induced diabetes, diabetes-associated endothelial dysfunction, arthritis, and traumatic braininjury, indicating that PARP activation may be involved in thepathomechanism of these diseases (Szabo and Dawson, 1998;Liaudet et al, 2000; Soriano et al, 2001). Very limited information isavailable in the literature regarding the possible role of peroxynitriteand PARP activation in skin pathology. Nitric oxide, a parentmolecule of peroxynitrite, has been shown to be produced byiNOS in keratinocytes, Langerhans cells, and dermal microvascularendothelial cells (Arany et al, 1996; Qureshi et al, 1996; Hoffmannet al, 1999). As superoxide is continously ``leaking'' from themitochondrial respiratory chain and is also produced by NADPHoxidase and xanthine oxidase enzymes under in¯ammatory condi-tions, the possibility exists that peroxynitrite is formed in thein¯amed skin and PARP activation may also occur. Indeed,peroxynitrite has been shown to be produced by ultraviolet-B-irradiated keratinocytes and endothelial cells and has been impli-cated in ultraviolet-B-induced skin in¯ammation (Deliconstantinoset al, 1996a; 1996b; 1996c). Furthermore, nitrotyrosine, as anindicator of in vivo peroxynitrite production, has been detected inskin homogenates from burns, chronic ultraviolet-B exposure,ischemia reperfusion, murine leishmaniasis, and systemic sclerosis(Giorgio et al, 1996; Hattori et al, 1996; Cotton et al, 1999; Um etal, 1999; Rawlingson et al, 2000).
The aim of this study was to investigate (i) whether peroxynitriteis produced in the skin during CHS, (ii) whether DNA breakageand PARP activation occur in CHS, (iii) whether peroxynitrite canactivate PARP in the skin, (iv) the cytostatic and cytotoxic effect ofperoxynitrite on the human keratinocyte cell line HaCaT, (v) therole of PARP in the peroxynitrite-induced cytostasis andcytotoxicity in HaCaT cells, and (vi) whether PARP is involvedin the cytokine-induced activation of HaCaT cells.
MATERIALS AND METHODS
Materials Unless speci®ed otherwise, all chemicals and materials listedwere purchased from Sigma-Aldrich (St. Louis, MO). 3-Aminophenanthridinone was from the rare chemical library of Sigma-Aldrich. Peroxynitrite was a kind gift from Dr. Harry Ischiropoulos(University of Pennsylvania Medical Center, Philadelphia, PA).
Animals Animal experiments conform with the Guide for the Care andUse of Laboratory Animals published by the US National Institute ofHealth and the treatment protocol was approved by the InstitutionalAnimal Care and Use Committee. Female CD1 mice were used in theexperiments and were allowed free access to food and water.
CHS model and in vivo peroxynitrite treatment A total of 12 micewere randomized into two groups (control group and CHS group).Sensitization was carried out on the shaved abdominal wall by theadministration of 100 ml 2% oxazolone solubilized in acetone:olive oil4:1 (CHS group, n = 6) or 100 ml vehicle (control group, n = 6). After7 d, all animals were challenged by applying 20 ml 0.5% oxazolone oneach ear. After 24 h, mice were sacri®ced by CO2 and ears wereimmediately removed and placed in formalin or frozen incryoembedding medium.
For in vivo peroxynitrite treatment, peroxynitrite was diluted inphosphate-buffered saline (PBS, pH 11.0) and 400 nmol was smeared onthe shaved abdominal wall in a volume of 200 ml. Control mice weretreated with the vehicle (PBS, pH 11.0). After 30 min, skin was excisedand frozen in cryoembedding medium.
Immunohistochemistry The immunohistochemical procedure wascarried out basically as described previously (ViraÂg et al, 1998b) withslight modi®cations, as follows. Paraf®n sections (5 mm) were treatedwith 0.3% hydrogen peroxide for 15 min to block endogenousperoxidase activity and then rinsed brie¯y in PBS. Non-speci®c bindingwas blocked by incubating the slides for 1 h in 2% goat serum (in PBS).To detect nitrotyrosine, rabbit polyclonal antinitrotyrosine antibody(Upstate Biotechnology, Lake Placid, NY) was applied for 2 h in adilution of 1:1000 at room temperature. (Control sections wereincubated with either normal rabbit serum or with the primary antibodyin the presence of 10 mM nitrotyrosine.) Following extensive washing(®ve 3 5 min) with PBS, immunoreactivity was detected with a
biotinylated goat antirabbit secondary antibody and the avidin±biotin-peroxidase complex (ABC), both supplied in the Vector Elite kit (VectorLaboratories, Burlingame, CA). Color was developed using nickel-DABsubstrate. Sections were counterstained with nuclear fast red for 2 min,dehydrated, and mounted in Permount medium.
Detection of DNA strand breaks DNA breakage was detected with acommercially available kit (Roche Molecular Biochemicals, Indianapolis,IN) following the manufacturer's instructions. Dewaxed and rehydratedsections were treated with 20 mg per ml proteinase K (in 10 mMTris/HCl pH 7.8) for 30 min at 37°C. DNA breaks were labeled withterminal deoxyribonucleotide transferase (TdT) and a deoxyribo-nucleotide mix containing ¯uorescein isothiocyanate (FITC) labeleddUTP for 60 min at 37°C. After washing, sections were incubated withanti-FITC peroxidase conjugate (30 min at room temperature), andperoxidase was detected as described above for immunohistochemistry.
In situ PARP activity assay PARP activity in tissues was detected byan in situ histochemical reaction utilizing biotinylated NAD+ as substrateof PARP (Zhang, 1997). Cryosections were ®xed in 95% ethanol at±20°C and then rinsed in PBS. Reaction mixture (10 mM MgCl2,1 mM dithiothreitol, 70 mM biotinylated NAD+, in 100 mM Tris,pH 8.0) was applied to the sections for 30 min at room temperature. Areaction mix containing 5 mM 3-aminobenzamide (3-AB) or biotinyl-NAD+-free reaction mix was used as control. After three washes in PBS,incorporated biotin was detected by peroxidase-conjugated streptavidine(Trevigen, Gaithersburg, MD). Color development, counterstaining,and mounting were carried out exactly as described for theimmunohistochemistry.
Cell culture and peroxynitrite treatment HaCaT cells (kindlyprovided by Dr. Ulrich Rodeck, Thomas Jefferson University,Philadelphia, PA, with the permission of Professor Norbert E. Fusenig,German Cancer Research Center, Heidelberg, Germany) were culturedin RPMI 1640 medium supplemented with 10% fetal bovine serum,10 mM glutamine, 10 mM HEPES, 100 U per ml penicillin, and 100 mgper ml streptomycin. The concentration of authentic peroxynitrite wasdetermined spectrophotometrically by measuring absorbance at 302 nmusing an extinction coef®cient of 1670 M±1 cm±1. Peroxynitrite wasdiluted in PBS (pH 11.0) and was added to the cells in 1/10 of thevolume of the cell suspension. Under these conditions addition ofperoxynitrite does not shift the pH of the medium. Control sampleswere treated with PBS (pH 11.0) only. The effect of decomposedperoxynitrite (kept in PBS pH 7.2 at 37°C for 30 min) has also beentested in all of the assays and was found to have no effect on anyparameters measured.
Cell proliferation and cytotoxicity assays Cell proliferation wasdetermined with a commercially available enzyme-linked immunosorbentassay (ELISA) kit (Amersham Pharmacia Biotechnology, Piscataway, NJ)based on the measurement of BrDU incorporation. The assay was carriedout in 96-well tissue culture plates following the manufacturer'sinstructions. Cytotoxicity was quantitated with the colorimetric MTTtest, as described previously (ViraÂg et al, 1995).
Cellular PARP activity assay PARP activity was measured aspreviously described (ViraÂg et al, 1998a). Medium was removed from thecells 20 min after peroxynitrite treatment and cells were incubated at37°C in 0.5 ml assay buffer [56 mM Hepes pH 7.5, 28 mM KCl,28 mM NaCl, 2 mM MgCl2, 0.01% digitonin, and 0.125 mM 3H-NAD(0.5 mCi per ml)]. Cells were then scraped and transferred intoEppendorf tubes. Next, 200 ml ice-cold 50% trichloroacetic acid (TCA)was added to the samples and tubes were incubated for 4 h at 4°C.Samples were then spun (10,000g, 10 min) and the pellets were washedtwice in ice-cold 5% TCA and solubilized overnight in 250 ml 2%sodium dodecyl sulfate/0.1 N NaOH at 37°C. The contents of the tubeswere added to 7 ml ScintiSafe Plus scintillation liquid (Fisher Scienti®c)and radioactivity was determined in a liquid scintillation counter (Wallac,Gaithersburg, MD).
Measurement of caspase activation Caspase activity was measuredby the cleavage of the ¯uorogenic tetrapeptide-amino-4-methylcoumarine conjugate (DEVD-AMC), as described previously(ViraÂg and SzaboÂ, 1998). Cells were harvested 6 h after peroxynitritetreatment, washed in 1 3 PBS, and then lyzed in a lysis buffer (10 mMHEPES, 0.1% CHAPS, 5 mM dithiothreitol, 2 mM ethylenediaminetetraacetic acid, 10 mg per ml aprotinin, 20 mg per ml leupeptin, 10 mgper ml pepstatin A, and 1 mM phenylmethylsulfonyl ¯uoride, pH 7.25).Cell lysates and substrates (50 mM) were combined in triplicate in thecaspase reaction buffer (100 mM HEPES, 10% sucrose, 5 mM
VOL. 117, NO. 1 JULY 2001 PEROXYNITRITE AND PARP IN CHS 75
dithiothreitol, 0.1% CHAPS, pH 7.25). AMC liberation was monitoredover time with a Shimadzu ¯uorimeter using 380 nm excitation and460 nm emission wavelength.
Measurement of DNA fragmentation DNA fragmentation wasmeasured with a commercially available ELISA kit (Roche MolecularBiochemicals, Indianapolis, IN) following the manufacturer's instructions,as previously described (ViraÂg et al, 1998a). The assay is based on thedetection of histone-associated DNA fragments from the cytoplasmicfraction of cell lysates. HaCaT cells (2 3 105) were seeded into 12-wellplates in 1000 ml culture medium and treated with differentconcentrations of peroxynitrite. After 6 h incubation (37°C, 5% CO2)cells were washed with cold PBS, scraped into 400 ml incubation buffer(supplied with the kit), and incubated at 4°C for 60 min. Cell lysates
were then transferred to Eppendorf tubes and centrifuged (10,000g,10 min). Supernatants were diluted 103 and measured in an ELISA(antihistone capturing antibody, peroxidase-conjugated anti-DNAsecondary antibody, ABTS substrate). Absorbance was then measured(405 nm) versus substrate solution as a blank using a Spectramax 250microplate reader (Molecular Devices, Sunnyvale, CA).
Determination of intercellular adhesion molecule 1 (ICAM-1)expression HaCaT cells were stimulated by 30 ng per ml (300 U perml) recombinant human interferon-g (R&D Systems, Minneapolis, MN)for 24 h. Cells were then scraped and washed twice in ice-cold PBS.Staining was performed on ice with FITC-conjugated monoclonalantihuman ICAM-1 antibody (Pharmingen, San Diego, CA) andisotype-matched monoclonal antibody (Pharmingen) as a control for 1 h.
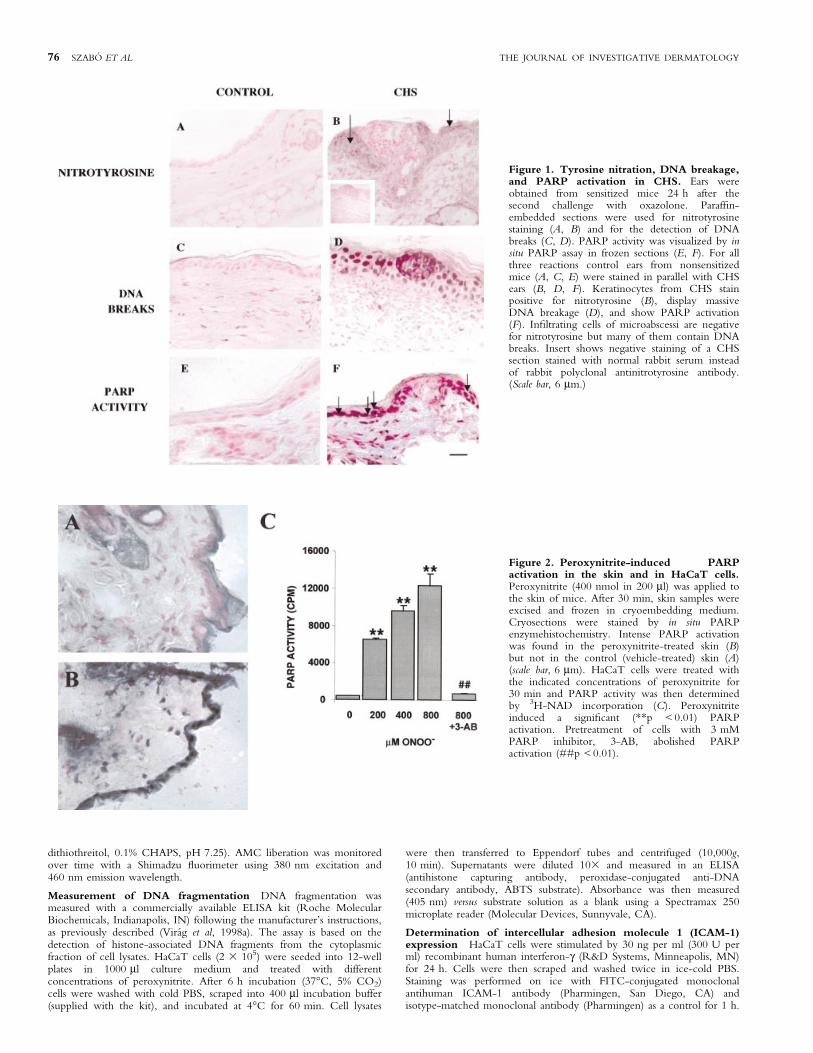
Figure 1. Tyrosine nitration, DNA breakage,and PARP activation in CHS. Ears wereobtained from sensitized mice 24 h after thesecond challenge with oxazolone. Paraf®n-embedded sections were used for nitrotyrosinestaining (A, B) and for the detection of DNAbreaks (C, D). PARP activity was visualized by insitu PARP assay in frozen sections (E, F). For allthree reactions control ears from nonsensitizedmice (A, C, E) were stained in parallel with CHSears (B, D, F). Keratinocytes from CHS stainpositive for nitrotyrosine (B), display massiveDNA breakage (D), and show PARP activation(F). In®ltrating cells of microabscessi are negativefor nitrotyrosine but many of them contain DNAbreaks. Insert shows negative staining of a CHSsection stained with normal rabbit serum insteadof rabbit polyclonal antinitrotyrosine antibody.(Scale bar, 6 mm.)
Figure 2. Peroxynitrite-induced PARPactivation in the skin and in HaCaT cells.Peroxynitrite (400 nmol in 200 ml) was applied tothe skin of mice. After 30 min, skin samples wereexcised and frozen in cryoembedding medium.Cryosections were stained by in situ PARPenzymehistochemistry. Intense PARP activationwas found in the peroxynitrite-treated skin (B)but not in the control (vehicle-treated) skin (A)(scale bar, 6 mm). HaCaT cells were treated withthe indicated concentrations of peroxynitrite for30 min and PARP activity was then determinedby 3H-NAD incorporation (C). Peroxynitriteinduced a signi®cant (**p < 0.01) PARPactivation. Pretreatment of cells with 3 mMPARP inhibitor, 3-AB, abolished PARPactivation (##p < 0.01).
76 SZABOÂ ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

Cells were then washed twice in ice-cold PBS and ®xed in 1%paraformaldehyde. Samples were analyzed on a FACSCalibur ¯owcytometer (Becton Dickinson, San Diego, CA) and ICAM-1 expressionwas determined as an increase in mean ¯uorescence intensity.
Measurement of interleukin-8 (IL-8) production HaCaT cells werestimulated for 24 h with 20 ng per ml human recombinant IL-1b (R&DSystems) in the presence or absence of PARP inhibitors. Supernatantswere then collected and assayed for IL-8 in a commercially availablesandwich ELISA system (R&D Systems).
Statistical analysis Results are reported as mean 6 SD ofquadruplicate samples. Experiments were repeated at least n = 3±4 times.Student's t test was used to compare mean values. Statistical differenceswere declared signi®cant for p < 0.05.
RESULTS
Tyrosine nitration in the in¯amed skin during CHS Asperoxynitrite nitrates the tyrosine residues of various proteins, in vivoperoxinitrite production is indicated by the presence ofnitrotyrosine-containing proteins in the tissues. Nitrotyrosinecould be detected by immunohistochemistry in the sections ofears from CHS mice but not from control (nonsensitized) ones(Fig 1). Tyrosine nitration of keratinocytes was most intense in thevicinity of in¯ammatory foci (microabscessi). Somewhatsurprisingly, only a small number of in®ltrating granulocytesdisplayed immunoreactivity for nitrotyrosine. Nitrotyrosine wasnot detectable in control nonsensitized skin (Fig 1).
Increased DNA breakage during CHS-induced skinin¯ammation As the formation of nitrotyrosine is mostprobably caused by peroxynitrite, which is a potent inducer ofDNA breakage, we performed TUNEL staining on skin samplesfrom CHS and control nonsensitized animals (Fig 1). Only faintpositivity could be detected in some scattered cells in control skintissues. In contrast, CHS-induced dermatitis resulted in a massiveincrease of TUNEL-positive cells both in the in®ltrating cells ofmicroabscessi and in keratinocytes (Fig 1). It is worthwhile notinghere that the TUNEL assay is widely used to detect apoptotic cells;however, what it detects are free 3¢-OH ends in DNA, which canbe caused by either single or double strand breaks. In fact, theTUNEL assay has been used to detect single strand breakage inseveral systems (Didier et al, 1996; Ahmadi and Ong, 1999; Jung etal, 2000; Mihm et al, 2000; Shen and Ong, 2000). Moreover, it hasalso been reported recently that proteinase K treatment (applied in
Figure 3. Antiproliferative and cytotoxic effect of peroxynitritein HaCaT cells. Proliferation (left panel) and viability (right panel) ofHaCaT cells have been determined after treatment of cells with theindicated concentrations of peroxynitrite. Treatment was carried out inthe presence or absence of 3-AB (3 mM). Asterisks indicate signi®cant(*p < 0.05, **p < 0.01) protection provided by the PARP inhibitor.
Figure 4. Peroxynitrite-induced caspaseactivation and DNA fragmentation inHaCaT cells. HaCaT cells were treated withincreasing concentrations of peroxynitrite in thepresence or absence of 3 mM 3-AB. After 6 h,caspase activity (left panel) and DNA fragmentationwere determined. Asterisks indicate signi®cantly(*p < 0.05, **p < 0.01) elevated caspase activityor DNA fragmentation. Hash marks indicatesigni®cant difference between 3-AB-treated anduntreated cells.
Figure 5. Effect of PARP inhibition onICAM-1 expression and IL-8 production ofimmune-stimulated HaCaT cells. (A)ICAM-1 expression; (B) IL-8 production.HaCaT cells were stimulated for 24 h with30 ng per ml human recombinant interferon-gto induce ICAM-1 expression or with humanrecombinant IL-1b to induce IL-8 secretion.Induction was carried out in the presence orabsence of 3-AB. 3-AB (3 mM) causedsigni®cant (*p < 0.05, **p < 0.01) inhibitionof both ICAM-1 expression and IL-8secretion.
VOL. 117, NO. 1 JULY 2001 PEROXYNITRITE AND PARP IN CHS 77

TUNEL kits) enhances the detection of single strand breaks in non-apoptotic cells (Gal et al, 2000). In our system, we believe thatDNA single strand breaks are responsible for the majority ofTUNEL positivity, as most TUNEL-positive nuclei do not showapoptotic morphology. Moreover, the vast majority of TUNEL-positive cells stain negative for p85 (a fragment of PARP generatedby caspase-mediated cleavage of the 116 kDa enzyme duringapoptosis) (not shown), indicating that no caspase activation occurs.Moreover, we found that almost 100% of keratinocytessurrounding the in¯ammatory foci are TUNEL positive, whichwould be an improbably high apoptotic percentage.
PARP activation in CHS We detected PARP activation inCHS by in situ PARP activity assay. PARP activity was notdetectable in control sections (Fig 1E). In CHS, we found strongstaining in keratinocytes and also in various dermal cells (Fig 1F).When the reaction was carried out in the presence of 5 mM 3-AB,no staining could be detected (not shown).
Peroxynitrite induces PARP activation in the skin and inHaCaT cells As peroxynitrite has been shown to cause DNAsingle strand breaks in human keratinocytes (Spencer et al,1996), we investigated whether poly(ADP-ribose) polymerase, anuclear nick sensor enzyme, becomes activated in peroxynitrite-treated keratinocytes. To prove that peroxynitrite can activatePARP in the skin, we applied 400 nmol peroxynitrite on theskin of mice for 30 min. Control animals were treated withvehicle (PBS, pH 11.0). We found intense PARP activation inthe peroxynitrite-treated skin but not in control skin, asdetected by in situ PARP activity assay (Fig 2A, B). PARPactivation was most intense in keratinocytes; however, somedermal cells also stained positive. The presence of the PARPinhibitor 3-AB (5 mM) in the reaction mixture abolished thestaining.
We also investigated the cellular effects of peroxynitrite in ahuman keratinocyte-derived cell line (HaCaT). Peroxynitriteinduced a concentration-dependent PARP activation (Fig 2C),which was completely blocked by the PARP inhibitor 3-AB.
Antiproliferative and cytotoxic effect of peroxynitrite inHaCaT cells Peroxynitrite caused a dose-dependent inhibitionof cell proliferation (Fig 3A) and also caused cytotoxicity (Fig 3B).PARP is not likely to be involved in proliferation inhibition causedby the lowest concentrations of peroxynitrite, as 3-AB had noeffect on this parameter. At higher concentrations, theantiproliferative effect of peroxynitrite was in part mediated byPARP activation, as indicated by the signi®cant cytoprotectiveeffect of 3-AB. 3-AB also provided signi®cant protection againstperoxynitrite-induced cytotoxicity (Fig 3B). Similar results wereobtained with another PARP inhibitor, 3-amino-phenanthridinone (5 mM) (not shown).
Peroxynitrite-induced caspase activation and DNAfragmentation in HaCaT cells Peroxynitrite has beenreported to cause apoptosis in various cell types includingthymocytes, HL-60 cells, and PC12 cells (Estevez et al, 1995; Linet al, 1995; ViraÂg and SzaboÂ, 1998; ViraÂg et al, 1998a). Here weshow that peroxynitrite also causes apoptosis in HaCaT cells, asindicated by increased caspase-3-like activity (Fig 4A) and DNAfragmentation (Fig 4B). With other cell types we have previouslyshown that pharmacologic PARP inhibition or the absence ofPARP (in PARP±/± cells) shifts the oxidant-induced necrotic celldeath toward apoptosis and toward the normal phenotype (ViraÂg etal, 1998a; 1998c). The increase in DNA fragmentation is usuallymore pronounced than the increase in caspase activation, indicatingthat DNA fragmentation is more sensitive to the consequences(e.g., ATP depletion) of PARP activation. In peroxynitrite-treatedHaCaT cells, in the presence of 3-AB we have detected amoderately increased DNA fragmentation but virtually unchangedcaspase activity (Fig 4). 3-AB (2±16 mM) alone, however, did notcause caspase activation or DNA fragmentation (data not shown).
Involvement of PARP in the regulation of ICAM-1expression and IL-8 production Human recombinantinterferon-g (30 ng per ml) induced a ninefold increase in thecell surface expression of ICAM-1. Interferon-g-induced ICAM-1expression was dose-dependently inhibited by 3-AB. The highestconcentration (4 mM) of the PARP inhibitor caused a 50%reduction in the expression of ICAM-1. Similar results wereobtained with another PARP inhibitor, 3-amino-phenanthridinone (5 mM) (not shown).
In serum-containing media, HaCaT cells are known to spon-taneously produce IL-8 (Stein et al, 1997). We found that IL-1bincreased the production of IL-8 by HaCaT cells. Both the basaland the IL-1b-induced IL-8 production were inhibited by 3-AB(4 mM), indicating a possible involvement of PARP in theregulation of IL-8 expression.
DISCUSSION
Here we provide evidence that intense tyrosine nitration occurs inthe epidermis during CHS. Detection of nitrotyrosine in tissues isregarded as an indicator of in vivo peroxynitrite production(Beckman and Koppenol, 1996). Although microabscessi in theskin contained a large number of neutrophil granulocytes, the vastmajority of these cells did not stain for nitrotyrosine. In contrast toin®ltrating cells, keratinocytes showed the most intense staining fornitrotyrosine, thus representing the prime targets of peroxynitrite inthe skin during CHS. The most likely scenario for increasedtyrosine nitration is that in¯ammatory cytokines induce theupregulation of iNOS synthase and overproduction of superoxide.iNOS-derived NO and superoxide, in turn, react to formperoxynitrite, which nitrates tyrosine. Other mechanisms ofin vitro tyrosine nitrations, most notably the one mediated bymyeloperoxidase, have also been described (Sampson et al, 1998);however, the in vivo signi®cance of these alternative tyrosinenitration pathways remains to be proven. Our ®nding thatin¯ammatory cells in microabscessi were negative for nitrotyrosinemakes it unlikely that myeloperoxidase-mediated tyrosine nitrationoccurs in our model.
We have also shown that, in CHS, intense DNA breakage occursin keratinocytes as well as in in®ltrating cells. Based on the lack ofapoptotic morphology, lack of caspase activation, and the unani-mous staining of keratinocytes surrounding the in¯ammatory foci,we propose that single stranded DNA breakage occurs in thekeratinocytes during CHS. In fact, peroxynitrite has been shown tocause DNA damage in various cell types including humankeratinocytes (Spencer et al, 1996; SzaboÂ, 1996b). In light of theproapoptotic effect of peroxynitrite, however, it cannot beexcluded that a fraction of TUNEL-positive cells are apoptotic.
We have investigated the cellular effects of peroxynitrite inHaCaT cells with special regard to the role played by PARP. Nitricoxide, one of the parent molecules of peroxynitrite, has beenshown to enhance the proliferation of HaCaT cells (Stallmeyer et al,1999). Peroxynitrite at lower concentrations was antiproliferative,further emphasizing the differences in the cellular effects of NO andperoxynitrite. The differences in the chemistry, reactivity, andtarget molecule spectrum may account for the often divergentcellular effects of NO and peroxynitrite. At higher concentrationsperoxynitrite induced cytotoxicity. Similarly to our previousobservations (Szabo et al, 1998; ViraÂg et al, 1998a; 1998c) wefound cytoprotection by PARP inhibitors against peroxynitrite-induced cytotoxicity in HaCaT cells. The cytoprotection providedby 3-AB is not related to inhibition of apoptosis, as caspaseactivation and DNA fragmentation were not reduced. On thecontrary, 3-AB caused a moderate increase in peroxynitrite-induced apoptotic DNA fragmentation. This is not due to thecytotoxic effect of 3-AB, as 3-AB alone (2, 4, 8, 16 mM) causedneither cytotoxicity nor caspase activation or DNA fragmentation(data not shown). The increased output of apoptotic parameters in3-AB-treated and oxidatively stressed cells is in line with ourprevious ®ndings as well as reports from other laboratories showing
78 SZABOÂ ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

a diversion by PARP inhibitors of oxidant-induced necrosis towardapoptosis (ViraÂg et al, 1998a; Filipovic et al, 1999; Ha and Snyder,1999). Switching the mode of cell death from necrosis to apoptosismay be anti-in¯ammatory in vivo, as it prevents the leakage of theproin¯amatory cell content into the tissues. In oxidatively injuredcells, the mechanism of the necrosis to apoptosis switch in thepresence of PARP inhibitors is probably the preservation of NAD+
and ATP pools that would be depleted by PARP activation. InHaCaT cells, however, the increase in DNA fragmentation in thepresence of 3-AB is less than in other cell types. This is likely to bedue to differences in the metabolism of different cell types.
The question arises, however, whether PARP inhibitors mayhave therapeutic potential in CHS. We believe that PARPinhibition may prevent oxidant-induced cell dysfunction andcytotoxicity, by preserving cellular energy stores. PARP inhibitorsmay also have bene®cial effects in other human skin diseases such asStevens±Johnson syndome or toxic epidermal necrolysis wherekeratinocyte necrosis is accompanied by iNOS expression (Lerneret al, 2000); therefore the NO±peroxynitrite±PARP activation±necrosis pathway may be involved in the pathogenesis of thesediseases.
Another important mechanism by which PARP may beinvolved in CHS and other in¯ammatory dermatoses is theregulation of keratinocyte activation. Upon cytokine stimulation,keratinocytes are known to produce numerous humoral factors,among them chemokines such as IL-8. IL-8 is responsible for therecruitment of neutrophil granulocytes into the sites of in¯amma-tion, so that inhibition of IL-8 production by PARP inhibitors mayreduce in¯ammation. Upregulation of adhesion molecules is alsoimportant in the regulation of cellular traf®cking during in¯am-mation; thus the inhibitory effect of PARP inhibition on ICAM-1expression might be anti-in¯ammatory (Zingarelli et al, 1998). Thetranscription factor NFkB is a key regulator of the expression ofin¯ammatory proteins (Baeuerle and Baltimore, 1996), includingchemokines and adhesion molecules. PARP has been shown toenhance the effect of NFkB (Oliver et al, 1999), indicating thatinhibition of NFkB activation by PARP inhibitors may beresponsible for the reduced IL-8 production and ICAM-1 expres-sion in 3-AB-treated HaCaT cells.
CONCLUSIONS
(i) CHS leads to peroxynitrite production, DNA breakage, andPARP activation in keratinocytes. (ii) Peroxynitrite activates PARPin the skin. (iii) Peroxynitrite causes apoptosis characterized bycaspase activation and DNA fragmentation in HaCaT cells. (iv) Athigher concentrations, peroxynitrite causes necrosis in HaCaT cells.(v) Peroxynitrite activates PARP in HaCaT cells. (vi) PARP plays arole in the cytokine-induced activation of keratinocytes.
Our ®ndings provide rationale for experimental therapies ofCHS and possibly other in¯ammatory dermatoses by targetingperoxynitrite or PARP.
This work was supported by a grant from the National Institutes of Health to C.S.
(R01GM60915), by a ``Mecenatura'' grant from the Health Science Center of the
University of Debrecen to E.S. (Mec 10/99), and by a grant from the Hungarian
Ministry of Health (ETT 104/99) to L.V. L.V. was supported by a Bolyai
fellowship from the Hungarian Academy of Sciences. The technical assistance of Ms
Livia Szilagyi is greatly appreciated. The authors are grateful to Dr. U. Rodeck
(Thomas Jefferson University, Philadelphia, PA) for kindly providing HaCaT cells,
to Professor N.E. Fusenig (German Cancer Research Center, Heidelberg,
Germany) for permission to use this cell line, and to Dr. Harry Ischiropoulos
(University of Pennsylvania Medical Center, Philadelphia, PA) for donating
peroxynitrite.
REFERENCES
Ahmadi A, Ong SC: Fertilizing ability of DNA-damaged spermatozoa. Exp Zool284:696±704, 1999
Arany I, Brysk MM, Brysk H, Tyring SK: Regulation of inducible nitric oxidesynthase mRNA levels by differentiation and cytokines in humankeratinocytes. Biochem Biophys Res Commun 220:618±622, 1996
Baeuerle PA, Baltimore D: NF-kappa B: ten years after. Cell 87:13±20, 1996Beckman JS, Koppenol WH: Nitric oxide, superoxide, and peroxynitrite: the good,
the bad, and ugly. Am J Physiol 271:C1424±C1437, 1996Cochrane CG: Mechanisms of oxidant injury of cells. Mol Aspects Med 12:137±147,
1991Cotton SA, Herrick AL, Jayson MI, Freemont AJ: Endothelial expression of nitric
oxide synthases and nitrotyrosine in systemic sclerosis skin. J Pathol189:273±278, 1999
Deliconstantinos G, Villiotou V, Stavrides JC: Alterations of nitric oxide synthase andxanthine oxidase activities of human keratinocytes by ultraviolet B radiation.Potential role for peroxynitrite in skin in¯ammation. Biochem Pharmacol51:1727±38, 1996a
Deliconstantinos G, Villiotou V, Stavrides JC: Increase of particulate nitric oxidesynthase activity and peroxynitrite synthesis in UVB-irradiated keratinocytemembranes. Biochem J 320:997±1003, 1996b
Deliconstantinos G, Villiotou V, Stavrides JC: Nitric oxide and peroxynitrite releasedby ultraviolet B-irradiated human endothelial cells are possibly involved in skinerythema and in¯ammation. Exp Physiol 81:1021±1033, 1996c
Didier M, Bursztajn S, Adamec E, et al: DNA strand breaks induced by sustainedglutamate excitotoxicity in primary neuronal cultures. J Neurosci 16:2238±2250,1996
Estevez AG, Radi R, Barbeito L, Shin JT, Thompson JA, Beckman JS:Peroxynitrite-induced cytotoxicity in PC12 cells: evidence for an apoptoticmechanism differentially modulated by neurotrophic factors. J Neurochem65:1543±1550, 1995
Filipovic DM, Meng X, Reeves WB: Inhibition of PARP prevents oxidant-inducednecrosis but not apoptosis in LLC-PK1 cells. Am J Physiol 277:F428±F436,1999
Gal I, Varga T, Szilagyi I, Balazs M, Schlammadinger J, Szabo G Jr: Protease-elicitedTUNEL positivity of non-apoptotic ®xed cells. J Histochem Cytochem48:963±970, 2000
Giorgio S, Linares E, de Capurro ML, de Bianchi AG, Augusto O: Formation ofnitrosyl hemoglobin and nitrotyrosine during murine leishmaniasis. PhotochemPhotobiol 63:750±754, 1996
Gow AJ, Duran D, Malcolm S, Ischiropoulos H: Effects of peroxynitrite-inducedprotein modi®cations on tyrosine phosphorylation and degradation. FEBS Lett385:63±66, 1996
Grabbe S, Schwarz T: Immunoregulatory mechanisms involved in elicitation ofallergic contact hypersensitivity. Immunol Today 19:37±44, 1998
Groves JT: Peroxynitrite reactive, invasive and enigmatic. Curr Opin Chem Biol3:226±235, 1999
Ha HC, Snyder SH: Poly(ADP-ribose) polymerase is a mediator of necrotic celldeath by ATP depletion. Proc Natl Acad Sci USA 96:13978±13982, 1999
Hattori Y, Nishigori C, Tanaka T, et al: 8-hydroxy-2¢-deoxyguanosine is increasedin epidermal cells of hairless mice after chronic ultraviolet B exposure. J InvestDermatol 107:733±737, 1996
Hoffmann G, Schobersberger W, Rieder J, et al: Human dermal microvascularendothelial cells express inducible nitric oxide synthase in vitro. J Invest Dermatol112:387±390, 1999
Jung M, Davis WP, Taatjes DJ, Churg A, Mossman BT: Asbestos and cigarettesmoke cause increased DNA strand breaks and necrosis in bronchiolar epithelialcells in vivo. Free Radic Biol Med 28:1295±1299, 2000
Kong SK, Yim MB, Stadtman ER, Chock PB: Peroxynitrite disables the tyrosinephosphorylation regulatory mechanism: lymphocyte-speci®c tyrosine kinasefails to phosphorylate nitrated cdc2(6±20)NH2 peptide. Proc Natl Acad Sci USA93:3377±3382, 1996
Lerner LH, Qureshi AA, Reddy BV, Lerner EA: Nitric oxide synthase in toxicepidermal necrolysis and Stevens±Johnson syndrome. J Invest Dermatol114:196±199, 2000
Liaudet L, Soriano FG, Szabo E, Virag L, Mabley JG, Salzman AL, Szabo C:Protection against hemorrhagic shock in mice genetically de®cient inpoly(ADP-ribose) polymerase. Proc Natl Acad Sci USA 97:10203±10208, 2000
Lin KT, Xue JY, Nomen M, Spur B, Wong PY: Peroxynitrite-induced apoptosis inHL-60 cells. J Biol Chem 270:16487±16490, 1995
Ma XL, Lopez BL, Liu GL, Christopher TA, Ischiropoulos H: Peroxynitriteaggravates myocardial reperfusion injury in the isolated perfused rat heart.Cardiovasc Res 36:195±204, 1997
Mihm MJ, Jing L, Bauer JA: Nitrotyrosine causes selective vascular endothelialdysfunction and DNA damage. J Cardiovasc Pharmacol 36:182±187, 2000
Morita H, Hori M, Kitano Y: Modulation of picryl chloride-induced contacthypersensitivity reaction in mice by nitric oxide. J Invest Dermatol 107:549±552,1996
de Murcia G, Menissier de Murcia J: Poly(ADP-ribose) polymerase: a molecularnick-sensor. Trends Biochem Sci 19:172±176, 1994
Oliver FJ, Menissier-de Murcia J, Nacci C, et al: Resistance to endotoxic shock as aconsequence of defective NF-kappaB activation in poly(ADP-ribose)polymerase-1 de®cient mice. EMBO J 18:4446±4454, 1999
Ormerod AD, Dwyer CM, Reid A, Copeland P, Thompson WD: Inducible nitricoxide synthase demonstrated in allergic and irritant contact dermatitis. ActaDerm Venereol 77:436±440, 1997
Qureshi AA, Hosoi J, Xu S, Takashima A, Granstein RD, Lerner EA: Langerhanscells express inducible nitric oxide synthase and produce nitric oxide. J InvestDermatol 107: 815±821, 1996
VOL. 117, NO. 1 JULY 2001 PEROXYNITRITE AND PARP IN CHS 79

Rawlingson A, Greenacre SA, Brain SD: Generation of peroxynitrite in localised,moderate temperature burns. Burns 26:223±227, 2000
Ross R, Gillitzer C, Kleinz R, Schwing J, Kleinert H, Forstermann U, Reske-KunzAB: Involvement of NO in contact hypersensitivity. Int Immunol 10:61±69,1998
Rowe A, Farrell AM, Bunker CB: Constitutive endothelial and inducible nitricoxide synthase in in¯ammatory dermatoses. Br J Dermatol 136:18±23, 1997
Sampson JB, Ye Y, Rosen H, Beckman JS: Myeloperoxidase and horseradishperoxidase catalyze tyrosine nitration in proteins from nitrite and hydrogenperoxide. Arch Biochem Biophys 356:207±213, 1998
Shen H, Ong C: Detection of oxidative DNA damage in human sperm and itsassociation with sperm function and male infertility. Free Radic Biol Med28:529±536, 2000
Soriano FG, ViraÂg L, Jagtap P, et al: Diabetic endothelial dysfunction: the role ofpoly(ADP-ribose) polymerase activation. Nature Med 7:108±113, 2001
Spencer JP, Wong J, Jenner A, Aruoma OI, Cross CE, Halliwell B: Basemodi®cation and strand breakage in isolated calf thymus DNA and in DNAfrom human skin epidermal keratinocytes exposed to peroxynitrite or 3-morpholinosydnonimine. Chem Res Toxicol 9:1152±1158, 1996
Stallmeyer B, Kampfer H, Kolb N, Pfeilschifter J, Frank S: The function of nitricoxide in wound repair: inhibition of inducible nitric oxide-synthase severelyimpairs wound reepithelialization. J Invest Dermatol 113:1090±1098, 1999
Stein M, Bernd A, Ramirez-Bosca A, Kippenberger S, Holzmann H: Measurementof anti-in¯ammatory effects of glucocorticoids on human keratinocytes in vitro.Comparison of normal human keratinocytes with the keratinocyte cell lineHaCaT. Arzneimittelforschung 47:1266±1270, 1997
Szabo C: The pathophysiological role of peroxynitrite in shock, in¯ammation, andischemia-reperfusion injury. Shock 6:79±88, 1996a
Szabo C: DNA strand breakage and activation of poly-ADP ribosyltransferase: a
cytotoxic pathway triggered by peroxynitrite. Free Radic Biol Med 21:855±869,1996b
Szabo C, Dawson V: Role of poly(ADP-ribose) synthetase in in¯ammation andischaemia-reperfusion. Trends Pharmacol Sci 19:287±298, 1998
Szabo C, ViraÂg L, Cuzzocrea S, et al: Protection against peroxynitrite-induced®broblast injury and arthritis development by inhibition of poly(ADP-ribose)synthase. Proc Natl Acad Sci USA 95:3867±3872, 1998
Um SC, Suzuki S, Toyokuni S, Uchida K, Hiai H, Nishimura Y: Formation of 4-hydroxy-2-nonenal-modi®ed proteins and 3-nitro-L-tyrosine in rat island skin¯aps during and after ischemia. Ann Plast Surg 42:293±298, 1999
ViraÂg L, Szabo C: The crucial role of apopain in the peroxynitrite-induced apoptoticDNA fragmentation. Free Rad Biol Med 25:1075±1082, 1998
ViraÂg L, KereÂkgyarto C, Fachet J: A simple, rapid and sensitive ¯uorimetric assay forthe measurement of cell-mediated cytotoxicity. J Immunol Meth 185:199±208,1995
ViraÂg L, Scott GS, Cuzzocrea S, Marmer D, Salzman AL, Szabo C: Peroxynitrite-induced thymocyte apoptosis: the role of caspases and poly-(ADP-ribose)synthetase (PARS) activation. Immunology 94:345±355, 1998a
ViraÂg L, Hasko G, Salzman A, Szabo C: NADPH-diaphorase histochemistry detectsinducible nitric oxide synthetase (iNOS) activity in the thymus of naive andstaphylococcal enterotoxin B stimulated mice. J Histochem Cytochem46:787±792, 1998b
ViraÂg L, Salzman A, Szabo C: Poly(ADP-ribose) synthetase activation mediatesmitochondrial injury during oxidant-induced cell death. J Immunol161:3753±3759, 1998c
Zhang J: Use of biotinylated NAD to label and purify ADP-ribosylated proteins.Meth Enzymol 280:255±265, 1997
Zingarelli B, Salzman AL, Szabo C: Genetic disruption of poly(ADP-ribose)synthetase inhibits the expression of P-selectin and intercellular adhesionmolecule-1 in myocardial ischemia/reperfusion injury. Circ Res 83:85±94, 1998
80 SZABOÂ ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY