Embed Size (px)
Citation preview

1
1
2
Phage specificity of the freshwater fish pathogen Flavobacterium 3
columnare 4
5
Elina Laanto, Lotta-Riina Sundberg and Jaana K. H. Bamford* 6
7
Department of Biological and Environmental Science and Nanoscience Center, 8
University of Jyväskylä, Jyväskylä, POBox 35, 40014-University of Jyväskylä, Finland 9
10
11
Running title: Bacteriophages of Flavobacteria 12
Key words: F. columnare, fish pathogen, bacteriophages, phage ecology 13
14
Address for correspondence: 15
Jaana Bamford 16
Department of Biological and Environmental Science and Nanoscience Center 17
University of Jyväskylä, 18
P.O.Box 35, 40014 University of Jyväskylä 19
FINLAND 20
Tel: +358 14 260 2272 21
Fax: +358 14 260 2221 22
E-mail: [email protected] 23
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.05574-11 AEM Accepts, published online ahead of print on 2 September 2011
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

2
Flavobacteria and their phages were isolated from Finnish freshwaters and fish 24
farms. Emphasis was set in finding phages infecting the fish pathogen 25
Flavobacterium columnare for further purposes as phage therapy agents. Host 26
ranges of the flavobacterial phages varied, phages infecting F. columnare being 27
more host-specific than the other isolated phages. 28
29
Species of the genus Flavobacterium are widely distributed in nature and they have been 30
recorded from diverse habitats (3, 19, 30, 32, 36, 37, 38, 39). In general flavobacteria are 31
non-pathogenic, but some species are opportunistic pathogens (2). Columnaris, a disease 32
caused by a fish pathogen Flavobacterium columnare, can cause up to 100% mortality 33
among the salmonid fingerlings (28). Antibiotic treatment must be applied to prevent 34
mass mortalities at fish farms. Despite effective treatment columnaris disease occurs 35
repeatedly during summer (18). Therefore the number of antibiotic treatments and the 36
amount of antibiotics used can be extremely high. Increased resistance to antibiotics at 37
fish farms and their surroundings has been recorded in environmental bacteria (10, 24, 26, 38
31), and antibiotic resistant fish-pathogenic flavobacteria have also emerged (7). To 39
avoid risks related to antibiotic use, enrichment of bacteriophages could be an ecological 40
method to decrease the number of F. columnare infections. To our knowledge this is the 41
first study on European F. columnare phages. 42
43
In this study, a total of 53 flavobacterial isolates were received during the warm water 44
period (May to August) in years 2008-2009 from water samples that included both open 45
freshwater environments (rivers and lakes not connected to fish farming) and three inland 46
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

3
land based fish farms rearing mainly salmonid fingerlings in Finland (Fig. 1, Tables 1 and 47
2). One bacterium (B67) was isolated from diseased fish during 2007. Water samples 48
were cultured on Shieh agar (25) and 1/5 Luria Bertani agar (22), and yellow and orange 49
pigmented colonies selected for analyses. The flavobacterial isolates were subjected to 50
UP-PCR (for methods see references 5, 6, 13) and the 16S rDNA of different groups 51
were sequenced (Table 1). All F. columnare isolates fell into same UP-PCR group, and 52
thus they were further analysed with Ribosomal Intergenic Spacer Analysis (RISA) (8, 53
29). According to RISA, the isolates were assigned into 5 groups (Table 1). According to 54
our data, occurrence of F. columnare seems to be connected to fish farming environment; 55
it was not isolated in natural waters. However, it is likely, that the initial source of F. 56
columnare at the farms is nature, because there is evidence that it is present also outside 57
fish farms (20, 21, Kunttu et al., unpublished data). 58
59
Phages infecting the genus Flavobacterium and their interaction with the bacterial host 60
have been previously described mostly in marine environments (4, 9, 12, 14) but also the 61
phage-host relationship of a fish pathogen Flavobacterium psychrophilum has been 62
studied (27). A total of 49 bacteriophages were isolated from water samples (Table 2). 63
Phages were enriched using flavobacterial isolates from freshwaters, fish farms and 64
previously described F. columnare strains. Phage stocks were prepared and selected 65
phages were grown by infecting the host bacterium (multiplicity of infection 5-10) at 66
proper cell density and concentrated and purified (22). Many of the isolated phages 67
produced low titer lysates, and they were used only for infection tests (see section for 68
host range studies). Phage genomic DNA was extracted (for methods see references 1 and 69
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

4
23) and digested with BamHI, EcoRI, HindIII and PstI. For the genomes that were cut, 70
the genome size was calculated from the resulting restriction profile (Table 2). For the 71
phages that were sequenced (FKj-2, FL-1, FCL-2, FCV-1), approximately 3000 base 72
pairs of each genome (except from FCV-1 1600 base pairs) and BLASTed 73
((http://blast.ncbi.nlm.nih.gov/Blast.cgi, May 2011), no significant DNA-sequence 74
sequence similarity was found in the database. Putative protein-coding genes were 75
analyzed using Vector NTI 11.0.0 (Invitrogen). Best matches for FCL-2 were to a 76
hypothetical protein of the Vibrio phage VP16T (score 57.8) and to a hypothetical protein 77
of the Vibrio phage VP16C (score 55.1). For FCV-1 best match was to a hypothetical 78
protein B40-8030 of a Bacteroides phage B40-8 (score 53.9). Transmission electron 79
microscopy (TEM) was used to study phage morphology. All of the Flavobacterial 80
phages that were characterized were tailed phages of the families of Myoviridae, 81
Podoviridae or Siphoviridae (Fig. 1 and Table 2). Most of these phages had an average 82
head size from 50 to 70 nm but some of the Myovirus isolates had capsid sizes around 83
and larger than 100 nm (FJy-3, FKo-2, FKj-2, FKy-1 and FKy-3). 84
85
Phages infecting F. columnare were isolated only from fish farms during disease 86
outbreaks. These phages might survive inside the host cell during cold water period and 87
start a lytic cycle when nutrients come available for the host cell and enough amount of 88
energy is available. 89
90
Studies on aquatic phage-host interplay have been conducted extensively in the marine 91
environments (15) but is less known in freshwaters (15, 34, 35). In our study the host 92
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

5
ranges differed greatly between the phage isolates. In the initial screening for 93
susceptibility of the bacteria for phages, each bacterium was infected with each phage 94
isolate by spotting the phage lysate on top agar containing the host bacterium. Each 95
bacterium was then infected with each phage using plaque assay. Some of the isolated 96
phages infecting Flavobacterium sp. species were detected to have broad host range. 97
These phages infected bacteria isolated from both freshwater and fish farm samples 98
although it has been suggested that single host enrichments select for phages of narrow 99
host range from sewage and marine environments (11, 33). Some of the phages 100
(especially all F. columnare phages) were more host-specific according to our collection 101
of Flavobacterium strains. F. columnare strains isolated from the same location as the 102
phage were the only ones susceptible for that specific phage. 103
104
No infection was observed in F. columnare strains by Flavobacterium sp. phages and 105
vice versa. However, 11 Flavobacterium sp. phage lysates (FTs-1, FKo-2, FL-1, FV-1, 106
FKy-1, FKy-2, FKy-3, FV-3, FV-4, FV-5, FV-6 and FV-8) inhibited the growth or lysed 107
the underlying bacterial culture on all F. columnare strains tested but produced no 108
individual plaques (data not shown). The phages could have been able to bind to the 109
bacteria and cause death but were not able to produce progeny, or the clear spots could be 110
an indication of bacteriocin activity. The causative agent of this strong inhibition or lysis 111
should be studied further for the possibility of developing antimicrobial agents. One of 112
the phages (FCL-1) was isolated from a fish suffering from columnaris disease. The 113
presence of the F. columnare phages in the fish indicates the possibility to control a fish 114
disease by enrichment of these phages. A number of successful reports on phage therapy 115
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

6
against fish diseases have been published when the phages are applied directly to the 116
water (16, 17). Other possible benefits related to phages are that they could be developed 117
to be used as diagnostic tools. In the present study we show that bacteriophages and 118
Flavobacteria are widespread also in the northern fresh waters. We found phages of F. 119
columnare to be host specific making them good candidates for phage therapy. 120
121
All sequences have been submitted to EMBL Nucleotide Sequence Database, 122
http://www.ebi.ac.uk/embl/, under the accession numbers given in Table 1 and 123
#FR714876 (FCL-2), #FR714877 (FL-1), #FR714878 (FKj-2) and #FR865436 (FCV-1). 124
125
Mr Petri Papponen, Ms Katja Ryymin and Ms Irene Helkala are thanked for their 126
excellent technical assistance. Some bacterial strains used in this study were kindly 127
donated by Dr. Päivi Rintamäki and Finnish Food safety authority EVIRA. Station 128
manager Kari Saikkonen and field master Esa Karpoff from The Kevo Research Station 129
(University of Turku) are acknowledged for providing water samples. This work was 130
supported by the Finnish Centre of Excellence Program of the Academy of Finland 131
(2006-2011) grant 1129648 (J.K.H.B), Academy of Finland grant 127500 (L.-R.S.) and a 132
grant from Maj and Tor Nessling Foundation (J.K.H.B.) 133
134
References 135
1. Bamford, J. K., and D. H. Bamford. 1991. Large-scale purification of membrane-136
containing bacteriophage PRD1 and its subviral particles. Virology. 181:348-352. 137
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

7
2. Bernardet J.F., and Bowman J. P. 2006. The genus Flavobacterium, p. 481-531. In 138
M. Dworkin, S. Falkow, E. Rosenberg, K. H. Schleifer, E. Stackebrandt. (ed.), The 139
Prokaryotes: a Handbook on the Biology of Bacteria, 3rd ed., vol. 7. Springer-Verlag, 140
New York. 141
3. Bernardet, J. F., P. Segers, M. Vancanneyt, F. Berthe, K. Kersters, and P. 142
Vandamme. 1996. Cutting a Gordian knot: emended classification and description of the 143
genus Flavobacterium, emended description of the family Flavobacteriaceae, and 144
proposal of Flavobacterium hydatis nom. nov.(basonym, Cytophaga aquatilis Strohl and 145
Tait 1978). Int. J. Syst. Evol. Microbiol. 46:128. 146
4. Borriss, M., E. Helmke, R. Hanschke, and T. Schweder. 2003. Isolation and 147
characterization of marine psychrophilic phage-host systems from Arctic sea ice. 148
Extremophiles. 7:377-384. 149
5. Brandt, K. K., A. Petersen, P. E. Holm, and O. Nybroe. 2006. Decreased abundance 150
and diversity of culturable Pseudomonas spp. populations with increasing copper 151
exposure in the sugar beet rhizosphere. FEMS Microbiol. Ecol. 56:281-291. doi: 152
10.1111/j.1574-6941.2006.00081.x. 153
6. Bulat, S., N. Mironenko, M. Lapteva, and P. Strelchenko. 1994. Polymerase chain 154
reaction with universal primers (UP-PCR) and its application to plant genome analysis. 155
Conservation of Plant Genes II: Utilization of Ancient and Modern DNA. 48:113-129. 156
7. Ekman, E. 2003. Natural and experimental infections with Flavobacterium 157
psychrophilum in salmonid fish. Swedish University, Uppsala. 158
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

8
8. Hartmann, M., B. Frey, R. Kolliker, and F. Widmer. 2005. Semi-automated genetic 159
analyses of soil microbial communities: comparison of T-RFLP and RISA based on 160
descriptive and discriminative statistical approaches. J. Microbiol. Methods. 61:349-360. 161
doi: 10.1016/j.mimet.2004.12.011. 162
9. Holmfeldt, K., M. Middelboe, O. Nybroe, and L. Riemann. 2007. Large 163
variabilities in host strain susceptibility and phage host range govern interactions between 164
lytic marine phages and their Flavobacterium hosts. Appl. Environ. Microbiol. 73:6730-165
6739. doi: 10.1128/AEM.01399-07. 166
10. Huys, G., G. Rhodes, P. McGann, R. Denys, R. Pickup, M. Hiney, P. Smith, and 167
J. Swings. 2000. Characterization of oxytetracycline-resistant heterotrophic bacteria 168
originating from hospital and freshwater fishfarm environments in England and Ireland. 169
Syst. Appl. Microbiol. 23:599-606. 170
11. Jensen, E. C., H. S. Schrader, B. Rieland, T. L. Thompson, K. W. Lee, K. W. 171
Nickerson, and T. A. Kokjohn. 1998. Prevalence of broad-host-range lytic 172
bacteriophages of Sphaerotilus natans, Escherichia coli, and Pseudomonas aeruginosa. 173
Appl. Environ. Microbiol. 64:575. 174
12. Jiang, S. C., C. A. Kellogg, and J. H. Paul. 1998. Characterization of marine 175
temperate phage-host systems isolated from Mamala Bay, Oahu, Hawaii. Appl. Environ. 176
Microbiol. 64:535. 177
13. Lübeck, M., I. A. Alekhina, P. S. Lübeck, D. F. Jensen, and S. A. Bulat. 1999. 178
Delineation of Trichoderma harzianum into two different genotypic groups by a highly 179
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

9
robust fingerprinting method, UP-PCR, and UP-PCR product cross-hybridization. Mycol. 180
Res. 103:289-298. 181
14. Middelboe, M., K. Holmfeldt, L. Riemann, O. Nybroe, and J. Haaber. 2009. 182
Bacteriophages drive strain diversification in a marine Flavobacterium: implications for 183
phage resistance and physiological properties. Environ. Microbiol. 11:1971-1982. 184
15. Middelboe, M., S. Jacquet, and M. Weinbauer. 2008. Viruses in freshwater 185
ecosystems: an introduction to the exploration of viruses in new aquatic habitats. 186
Freshwat. Biol. 53:1069-1075. 187
16. Nakai, T., and S. C. Park. 2002. Bacteriophage therapy of infectious diseases in 188
aquaculture. Res. Microbiol. 153:13-18. 189
17. Park, S. C., and T. Nakai. 2003. Bacteriophage control of Pseudomonas 190
plecoglossicida infection in ayu Plecoglossus altivelis. Dis. Aquat. Organ. 53:33-39. 191
18. Pulkkinen, K., L. R. Suomalainen, A. F. Read, D. Ebert, P. Rintamaki, and E. T. 192
Valtonen. 2010. Intensive fish farming and the evolution of pathogen virulence: the case 193
of columnaris disease in Finland. Proc. Biol. Sci. 277:593-600. doi: 194
10.1098/rspb.2009.1659. 195
19. Qu, J. H., H. L. Yuan, H. F. Li, and C. P. Deng. 2009. Flavobacterium cauense sp. 196
nov., isolated from sediment of a eutrophic lake. Int. J. Syst. Evol. Microbiol. 59:2666-197
2669. doi: 10.1099/ijs.0.009688-0. 198
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

10
20. Revetta, R. P., M. R. Rodgers, and B. K. Kinkle. 2005. Isolation and identification 199
of freshwater bacteria antagonistic to Giardia intestinalis cysts. Journal of Water and 200
Health. 3:83-85. 201
21. Rickard, A., A. McBain, R. Ledder, P. Handley, and P. Gilbert. 2003. 202
Coaggregation between freshwater bacteria within biofilm and planktonic communities. 203
FEMS Microbiol. Lett. 220:133-140. 204
22. Sambrook J., and D. W. Russell. 2001. Molecular cloning: a laboratory manual. 205
Cold Spring Harbor Laboratory Press, New York. 206
23. Santos, M. A. 1991. An improved method for the small scale preparation of 207
bacteriophage DNA based on phage precipitation by zinc chloride. Nucleic Acids Res. 208
19:5442. 209
24. Schmidt, A. S., M. S. Bruun, I. Dalsgaard, and J. L. Larsen. 2001. Incidence, 210
distribution, and spread of tetracycline resistance determinants and integron-associated 211
antibiotic resistance genes among motile aeromonads from a fish farming environment. 212
Appl. Environ. Microbiol. 67:5675-5682. doi: 10.1128/AEM.67.12.5675-5682.2001. 213
25. Shieh, H. Studies on the nutrition of a fish pathogen, Flexibacter columnaris. 214
Microbios letters. 13:129-133. 215
26. Sorum, H. 2006. Antimicrobial drug resistance in fish pathogens, p. 213-238. In F. M. 216
Aarestrup (ed.), Antimicrobial Resistance in Bacteria of Animal Origin. American 217
Society for Microbiology Press, Washington, DC, USA. 218
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

11
27. Stenholm, A. R., I. Dalsgaard, and M. Middelboe. 2008. Isolation and 219
characterization of bacteriophages infecting the fish pathogen Flavobacterium 220
psychrophilum. Appl. Environ. Microbiol. 74:4070-4078. doi: 10.1128/AEM.00428-08. 221
28. Suomalainen, L., M. Tiirola, and E. Valtonen. 2005. Effect of Pseudomonas sp. 222
MT 5 baths on Flavobacterium columnare infection of rainbow trout and on microbial 223
diversity on fish skin and gills. Dis. Aquat. Org. 63:61-68. 224
29. Suomalainen, L. R., H. Kunttu, E. T. Valtonen, V. Hirvela-Koski, and M. Tiirola. 225
2006. Molecular diversity and growth features of Flavobacterium columnare strains 226
isolated in Finland. Dis. Aquat. Organ. 70:55-61. 227
30. Tamaki, H., S. Hanada, Y. Kamagata, K. Nakamura, N. Nomura, K. Nakano, 228
and M. Matsumura. 2003. Flavobacterium limicola sp. nov., a psychrophilic, organic-229
polymer-degrading bacterium isolated from freshwater sediments. Int. J. Syst. Evol. 230
Microbiol. 53:519-526. 231
31. Tamminen, M., A. Karkman, A. Lõhmus, W. I. Muziasari, H. Takasu, S. Wada, 232
S. Suzuki, and M. Virta. 2011. Tetracycline Resistance Genes Persist at Aquaculture 233
Farms in the Absence of Selection Pressure. Environ. Sci. Technol. 45:386-391. 234
32. Van Trappen, S., J. Mergaert, and J. Swings. 2003. Flavobacterium gelidilacus sp. 235
nov., isolated from microbial mats in Antarctic lakes. Int. J. Syst. Evol. Microbiol. 236
53:1241-1245. 237
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

12
33. Wichels, A., G. Gerdts, and C. Schütt. 2002. Pseudoalteromonas spp. phages, a 238
significant group of marine bacteriophages in the North Sea. Aquat. Microb. Ecol. 239
27:233-239. 240
34. Wilhelm, S. W., and A. R. Matteson. 2008. Freshwater and marine virioplankton: a 241
brief overview of commonalities and differences. Freshwat. Biol. 53:1076-1089. 242
35. Wommack, K. E., and R. R. Colwell. 2000. Virioplankton: viruses in aquatic 243
ecosystems. Microbiol. Mol. Biol. Rev. 64:69-114. 244
36. Yi, H., H. M. Oh, J. H. Lee, S. J. Kim, and J. Chun. 2005. Flavobacterium 245
antarcticum sp. nov., a novel psychrotolerant bacterium isolated from the Antarctic. Int. J. 246
Syst. Evol. Microbiol. 55:637-641. doi: 10.1099/ijs.0.63423-0. 247
37. Yoon, J. H., S. J. Kang, J. S. Lee, and T. K. Oh. 2007. Flavobacterium terrigena sp. 248
nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 57:947-950. doi: 249
10.1099/ijs.0.64776-0. 250
38. Zhang, D. C., H. X. Wang, H. C. Liu, X. Z. Dong, and P. J. Zhou. 2006. 251
Flavobacterium glaciei sp. nov., a novel psychrophilic bacterium isolated from the China 252
No.1 glacier. Int. J. Syst. Evol. Microbiol. 56:2921-2925. doi: 10.1099/ijs.0.64564-0. 253
39. Zhu, F., S. Wang, and P. Zhou. 2003. Flavobacterium xinjiangense sp. nov. and 254
Flavobacterium omnivorum sp. nov., novel psychrophiles from the China No. 1 glacier. 255
Int. J. Syst. Evol. Microbiol. 53:853-857. 256
257
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

13
Tables and Figures 258
Table 1. Bacterial strains isolated and used in the study 259
aThe previously studied F. columnare strains (genomic groups from A to H and colony 260
morphology from 1 to 4) are referred in this study by colony morphology rhizoid, Rz 261
(previously 1), rough, R (previously 2 and 3) and smooth, S (previously 4), following the 262
genomic group A to H, for example Rz-C (previously C1). 263
bUP-PCR group in numbers for Flavobacterium sp. , RISA group in letters for 264
Flavobacterium columnare, ND = not determined 265
cfor the partial 16S rRNA sequence of the bacteria 266
267
Table 2. Bacteriophages isolated and characterized in this study 268
a ND = not determined 269
b First letters in the phage name indicate the isolation host, F for Flavobacterium sp. and 270
FC for F. columnare, following letters refer to the sampling site 271
c L; much larger than the typical tailed phage genome (50 kb) 272
d Letter refers to the F. columnare RISA group that the phage is specific to 273
274
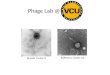
Figure 1. Panel A shows the isolation places of Flavobacteria and their phages from 275
Finland. Sampling sites are marked on the free map obtained from the National Land 276
Survey of Finland © Maanmittauslaitos 2005. The sites, their abbreviations and 277
coordinates are listed on the right. In the panel B are electron micrographs of purified and 278
negatively stained tailed Flavobacterium phages. (a) FCV-1; (b) FCL-2; (c) FJy-3; (d) 279
FKo-2; (e) FKy-1. Scale bar 50 nm. 280
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

14
281
Figure 2. Host ranges of the phages infecting different Flavobacterium sp. strains. Grey 282
squares indicate infection; white indicates no infection and light grey squares mark the 283
strain which was originally used for isolation of the phage. The numbers at right and 284
bottom are the total number of different host ranges of the phage and susceptibility of 285
bacteria to phages, respectively. The approximate titer of each phage on the bacteria is 286
marked with plusses (+ ≤ 105, ++ 106-108 and +++ ≥ 109, all pfu/ml). The titer (PFU/ml) 287
of the phage on the isolation strain is marked on the light grey squares. In parenthesis 288
after the name of the bacterial strain is the abbreviation of the isolation place, which is 289
also included in the phage nomenclature (see Figure 1). F.columnare phages infected 290
only and specifically to one F. columanre RISA group and they are listed in Table 2. 291
292
293
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

KeKjTs
Äk
Vj
Jy
Ky
Va
KoLe
Äy
AB
C
KeKjTs
Äk
Vj
Jy
Ky
Va
KoLe
Äy
AB
C
KeKjTs
Äk
Vj
Jy
KyVa
KoLe
ÄyL
V
I
In
Location Abbrevation Longitude Latitude
Lake Konnevesi Ko E 26° 35.644' N 62° 31.446'
Lake Jyväsjärvi Jy E 25° 44.339' N 62° 13.779'
River Vantaanjoki Va E 24° 58.072' N 60° 15.948'
River Kevojoki Ke E 26° 59.429' N 69° 45.337'
River Tsarsjoki Ts E 26° 59.447' N 69° 45.430'
Lake Kevojärvi Kj E 26° 59.608' N 69° 45.430'
Lake Valtimojärvi Vj E 28° 51.000' N 63° 40.688'
River Kymijoki Ky E 26° 49.926' N 60° 41.391'
Lake Leppävesi Le E 25° 54.633' N 62° 16.064'
Lake Äkässaivo Äk E 24° 8.640' N 67° 41.840'
River Äyskoski Äy E 26° 40.756' N 63° 0.665'
Lake Inari In E 27° 53.097' N 68° 47.783'
Fishery L 1) L
Fishery V 1) V
Fishery I 2) I1) Central Finland2) Northern Finland
A
B a b
c d e
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Bacteria
Phages B80
(Jy)
B18
7(L
)
B20
7(I
n)
B28
(Ko)
B17
6(L
e)B
223
(Ky)
B22
2(K
y)
B18
3(L
)
B16
7(J
y)
B22
4(K
y)
B24
1(V
)
B25
7(V
)
B26
0(V
)
B26
2(V
)
B16
9(J
y)
B17
8(Ä
k)
B11
4(K
e)
B21
8(V
)B
110
(Va)
B17
1(V
j)
B22
5(V
)
B24
3(V
)B
105
(Va)
B13
0(K
j)
B17
4(K
y)
B18
0(Ä
y)
B20
9(I
n)
B21
4(V
)
B12
7K
j)
B12
1(T
s)
B22
6(V
)
FTs-1 ++ ++ ++ +++ ++ ++ ++ ++ ++ ++ ++ +++ ++ ++ + ++ +++ +++ +++ ++ ++ ++ + 10824
FJy-3 + + ++ ++ ++ ++ + ++ 1010+ + ++ + ++ + + + + + + ++ ++ + + 24
FV-10 ++ ++ ++ + ++ + + ++ ++ ++ ++ ++ ++ 103
++ ++ + ++ 18
FV-5 ++ ++ ++ + ++ + + ++ ++ ++ 109
++ ++ ++ ++ ++ +++ 17
FV-6 ++ ++ ++ + ++ + ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ 108
17
FV-8 ++ ++ ++ + ++ + ++ ++ ++ ++ ++ 108
++ ++ ++ ++ +++ 17
FJy-2 ++ ++ +++ 104++ ++ + ++ ++ ++ + ++ ++ + 14
FKj-2 ++ ++ ++ ++ ++ + + ++ ++ + ++ 1010++ 13
FKy-2 ++ ++ +++ ++ 1011
+++ ++ ++ + ++ ++ + 12
FKo-1 ++ ++ ++ 106++ + ++ ++ + ++ 10
FKy-1 + ++ 1010
++ ++ ++ ++ 7
FLe-1 1010+++ +++ +++ + + ++ 7
FV-9 ++ + + + 109
+++ + 7
FKo-2 ++ 106++ + ++ ++ 6
FKj-1 ++ ++ ++ + + 1036
FV-11 +++ ++ + + +++ ++ 6
FV-1 + ++ + ++ +++ 1010
6
FKy-3 ++ + 108
++ ++ 5
FL-1 ++ + 109+ + 5
FV-12 +++ + + +++ 4
FV-2 + + 106
3
FV-3 + 109
+++ 3
FKe-1 ++ 1082
FV-4 + 109
2
FJy-1 1091
18 12 12 12 11 11 10 9 9 9 9 9 9 8 8 7 7 7 6 6 6 6 6 6 4 4 4 4 3 3 1
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Bact
eria
l st
rain
aU
P-PC
R/
RIS
A g
roup
bEM
BL A
cces
sion
nu
mbe
rcSa
mpl
ing
site
Sour
ce/
Ref
eren
ceBa
cter
ial
stra
ina
UP-
PCR
/
R
ISA
gro
upb
EMBL
Acc
essi
on
num
berc
Sam
plin
g si
teSo
urce
/ R
efer
ence
B67
AFi
sher
y L
This
stud
yB2
4327
FR69
6354
Fish
ery
VTh
is st
udy
B28
1FR
6963
28La
ke K
onne
vesi
This
stud
yB2
44N
DFi
sher
y V
This
stud
yB8
02
FR69
6329
Lake
Jyvä
sjär
viTh
is st
udy
B245
CFi
sher
y V
This
stud
yB1
053
FR69
6330
Riv
er V
anta
anjo
kiTh
is st
udy
B247
CFi
sher
y V
This
stud
yB1
104
FR69
6331
Riv
er V
anta
anjo
kiTh
is st
udy
B257
26FR
6963
55Fi
sher
y V
This
stud
yB1
145
FR69
6332
Riv
er K
evoj
oki
This
stud
yB2
59C
Fish
ery
VTh
is st
udy
B121
6FR
6963
33R
iver
Tsa
rsjo
kiTh
is st
udy
B260
ND
FR69
6356
Fish
ery
VTh
is st
udy
B127
7FR
6963
34La
ke K
evoj
ärvi
This
stud
yB2
61C
Fish
ery
VTh
is st
udy
B130
8FR
6963
35La
ke K
evoj
ärvi
This
stud
yB2
62N
DFR
6963
57Fi
sher
y V
This
stud
yB1
679
FR69
6336
Lake
Jyvä
sjär
viTh
is st
udy
B263
ND
FR69
6358
Fish
ery
VTh
is st
udy
B169
10FR
6963
37La
ke Jy
väsj
ärvi
This
stud
yB2
67C
Fish
ery
VTh
is st
udy
B171
11FR
6963
38La
ke V
altim
ojär
viTh
is st
udy
B268
CFi
sher
y V
This
stud
yB1
7412
FR69
6339
Riv
er K
ymijo
kiTh
is st
udy
B269
ND
Fish
ery
VTh
is st
udy
B176
13FR
6963
40La
ke L
eppä
vesi
This
stud
yB2
70C
Fish
ery
VTh
is st
udy
B178
14FR
6963
41La
ke Ä
käss
aivo
This
stud
yB2
71C
Fish
ery
ITh
is st
udy
B180
15FR
6963
42R
iver
Äys
kosk
iTh
is st
udy
B272
JFi
sher
y I
This
stud
yB1
8316
FR69
6343
Fish
ery
LTh
is st
udy
B273
CFi
sher
y I
This
stud
yB1
85G
FR69
6344
Fish
ery
LTh
is st
udy
B274
CFi
sher
y I
This
stud
yB1
8717
FR69
6345
Fish
ery
LTh
is st
udy
B275
CFi
sher
y I
This
stud
yB2
0718
FR69
6346
Lake
Inar
iTh
is st
udy
Rz-
AA
49B2
0919
FR69
6347
Lake
Inar
iTh
is st
udy
R-B
B49
B214
20FR
6963
48Fi
sher
y V
This
stud
yR
z-C
C49
B218
21Fi
sher
y V
This
stud
yS-
CC
49B2
2222
FR69
6349
Riv
er K
ymijo
kiTh
is st
udy
R-D
D49
B223
23FR
6963
50R
iver
Kym
ijoki
This
stud
yS-
DD
49B2
2424
Riv
er K
ymijo
kiTh
is st
udy
Rz-
EE
49B2
2524
FR69
6351
Fish
ery
VTh
is st
udy
R-E
E49
B226
25FR
6963
52Fi
sher
y V
This
stud
yS-
EE
49B2
30I
Fish
ery
VTh
is st
udy
S-F
FF.
col
umna
re ty
pe st
rain
NC
IMB
2248
B234
CFi
sher
y V
This
stud
yR
z-G
G49
B235
CFi
sher
y V
This
stud
yR
-GG
49B2
36N
DFi
sher
y V
This
stud
yS-
GG
49B2
37C
Fish
ery
VTh
is st
udy
R-H
H49
B241
26FR
6963
53Fi
sher
y V
This
stud
yS-
HH
49
bU
P-PC
R g
roup
in n
umbe
rs fo
r Fla
voba
cter
ium
sp.
, R
ISA
gro
up in
lette
rs fo
r Fla
voba
cter
ium
col
umna
re, N
D =
not
det
erm
ined
cfo
r the
par
tial 1
6S rR
NA
sequ
ence
of t
he b
acte
ria
aTh
e pr
evio
usly
stud
ied
F. c
olum
nare
stra
ins (
geno
mic
gro
ups f
rom
A to
H a
nd c
olon
y m
orph
olog
y fro
m 1
to 4
) are
refe
rred
in th
is st
udy
by c
olon
y m
orph
olog
y rh
izoi
d,
Rz
(pre
viou
sly
1), r
ough
, R (p
revi
ousl
y 2
and
3) a
nd sm
ooth
, S (p
revi
ousl
y 4)
, fol
low
ing
the
geno
mic
gro
up A
to H
, for
exa
mpl
e R
z-C
(pre
viou
sly
C1)
.
TABL
E 1.
Bac
teria
l stra
ins i
sola
ted
and
used
in th
e st
udy
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from

Phag
ebSa
mpl
ing
site
Isol
atio
n str
ain
Phag
e fa
mily
App
roxi
mat
e ge
nom
e siz
e (k
bp)c
RIS
A
grou
pd
FJy-
1La
ke J
yväs
järv
iB
80M
yovi
rida
e30
FJy-
2La
ke J
yväs
järv
iB
167
ND
>48
FJy-
3La
ke J
yväs
järv
iB
169
Myo
viri
dae
LFK
o-1
Lake
Kon
neve
siB
28M
yovi
rida
e25
-48
FKo-
2La
ke K
onne
vesi
B28
Myo
viri
dae
20-3
0FK
e-1
Riv
er K
evoj
oki
B11
4M
yovi
rida
eN
DFT
s-1
Riv
er T
sars
joki
B12
1N
D25
-48
FKj-1
Lake
Kev
ojär
viB
127
ND
>48
FKj-2
Lake
Kev
ojär
viB
130
Myo
viri
dae
25-4
8FK
y-1
Riv
er K
ymijo
kiB
222
Myo
viri
dae
20-4
8FK
y-2
Riv
er K
ymijo
kiB
223
Podo
viri
dae?
25-4
8FK
y-3
Riv
er K
ymijo
kiB
224
Myo
viri
dae?
LFL
e-1
Lake
Lep
päve
siB
176
Siph
ovir
idae
?20
-30
FL-1
Fish
ery
LB
183
Myo
viri
dae
55FV
-1Fi
sher
y V
B21
4M
yovi
rida
e28
FV-2
Fish
ery
VB
218
ND
25-4
8FV
-3Fi
sher
y V
B22
5M
yovi
rida
eN
DFV
-4Fi
sher
y V
B22
6Po
dovi
rida
e25
-48
FV-5
Fish
ery
VB
241
ND
LFV
-6Fi
sher
y V
B24
3N
D30
-40
FV-8
Fish
ery
VB
257
ND
LFV
-9Fi
sher
y V
B26
0Po
dovi
rida
e?L
FV-1
0Fi
sher
y V
B26
2N
DL
FV-1
1Fi
sher
y V
B26
3N
D>4
8FV
-12
Fish
ery
VB
278
Podo
viri
dae?
60FC
L-1
Fish
ery
LB
67M
yovi
rida
e/Po
dovi
rida
e50
AFC
L-2
Fish
ery
LB
185
Myo
viri
dae
30G
FCL-
3Fi
sher
y L
R-G
ND
30G
FCL-
4Fi
sher
y L
R-G
ND
ND
GFC
V-1
Fish
ery
VR
z-C
Myo
viri
dae
50C
FCV
-2Fi
sher
y V
B23
5N
DN
DC
FCV
-3Fi
sher
y V
B23
6N
DN
DC
FCV
-4Fi
sher
y V
Rz-
CN
DN
DC
FCV
-5Fi
sher
y V
Rz-
CN
DN
DC
FCV
-6Fi
sher
y V
Rz-
CN
DN
DC
FCV
-7Fi
sher
y V
Rz-
CN
DN
DC
FCV
-8Fi
sher
y V
Rz-
CN
DN
DC
FCV
-9Fi
sher
y V
B24
5N
DN
DC
FCV
-10
Fish
ery
VB
247
ND
ND
CFC
V-1
1Fi
sher
y V
Rz-
CN
DN
DC
FCV
-12
Fish
ery
VR
z-C
ND
ND
CFC
V-1
3Fi
sher
y V
B26
1N
DN
DC
FCV
-14
Fish
ery
VR
z-C
ND
ND
CFC
V-1
5Fi
sher
y V
Rz-
CN
DN
DC
FCV
-16
Fish
ery
VR
z-C
ND
ND
CFC
V-1
7Fi
sher
y V
Rz-
CN
DN
DC
FCV
-18
Fish
ery
VR
z-C
ND
ND
CFC
V-1
9Fi
sher
y V
Rz-
CN
DN
DC
FCV
-20
Fish
ery
VR
z-C
ND
ND
Ca N
D =
not
det
erm
ined
c L;
muc
h la
rger
than
the
typi
cal t
aile
d ph
age
geno
me
(50
kb)
d L
ette
r ref
ers t
o th
e F.
col
umna
re R
ISA
gro
up th
at th
e ph
age
is sp
ecifi
c to
TAB
LE 2
. Bac
terio
phag
es is
olat
ed a
nd c
hara
cter
ized
in th
is stu
dya
b F
irst l
ette
rs in
the
phag
e na
me
indi
cate
the
isola
tion
host,
F fo
r Fl
avob
acte
rium
sp. a
nd F
C fo
r F. c
olum
nare
, fol
low
ing
lette
rs re
fer t
o th
e
on April 11, 2020 by guest
http://aem.asm
.org/D
ownloaded from



![F0AV [B50-35] F0AW [B40-30] Lenovo B40/B50 Series · 2014-12-15 · Version 1.0 Machine type: F0AU [B50-30] F0AV [B50-35] F0AW [B40-30] 2014.05 Lenovo B40/B50 Series Bedienungsanleitung](https://img.pdfslide.net/doc/110x75/5ec9daab23c48b12f13c35a5/f0av-b50-35-f0aw-b40-30-lenovo-b40b50-series-2014-12-15-version-10-machine.jpg)




















![B40 - annexe[ocr]](https://img.pdfslide.net/doc/110x75/58f247171a28ab113e8b456d/b40-annexeocr.jpg)