Embed Size (px)
Citation preview

Ecology and Evolution 2017; 7: 771–779 | 771www.ecolevol.org
Received:6July2016 | Revised:13October2016 | Accepted:29October2016DOI:10.1002/ece3.2640
O R I G I N A L R E S E A R C H
Plant invasion and speciation along elevational gradients on the oceanic island La Palma, Canary Islands
Manuel J. Steinbauer1,2† | Severin D. H. Irl1† | Juana María González-Mancebo3 | Frank T. Breiner4,5 | Raquel Hernández-Hernández3 | Sebastian Hopfenmüller6 | Yohannes Kidane1 | Anke Jentsch7 | Carl Beierkuhnlein1
ThisisanopenaccessarticleunderthetermsoftheCreativeCommonsAttributionLicense,whichpermitsuse,distributionandreproductioninanymedium,providedtheoriginalworkisproperlycited.©2016TheAuthors.Ecology and EvolutionpublishedbyJohnWiley&SonsLtd.
†Equalcontribution.
1DepartmentofBiogeography,BayCEER,UniversityofBayreuth,Bayreuth,Germany2SectionEcoinformatics&Biodiversity,DepartmentofBioscience,AarhusUniversity,Aarhus,Denmark3DepartamentodeBotánica,EcologíayFisiologíaVegetal,UniversidaddeLaLaguna,SanCristóbaldeLaLaguna,Tenerife,Spain4SwissFederalResearchInstituteWSL,Birmensdorf,Switzerland5DepartmentofEcologyandEvolution,UniversityofLausanne,Lausanne,Switzerland6DepartmentofAnimalEcologyandTropicalBiology,UniversityofWürzburg,Würzburg,Germany7DepartmentofDisturbanceEcology,BayCEER,UniversityofBayreuth,Bayreuth,Germany
CorrespondenceManuelJ.Steinbauer,SectionEcoinformatics&Biodiversity,DepartmentofBioscience,AarhusUniversity,Aarhus,Denmark.Email:[email protected]
Funding informationEuropeanUnion’sHorizon2020,Grant/AwardNumber:641762;Net-Biometransnational;GermanResearchFoundation(DFG);UniversityofBayreuth;DanishCarlsbergfondet,Grant/AwardNumber:CF14-0148.
AbstractEcosystems that provide environmental opportunities but are poor in species andfunctionalrichnessgenerallysupportspeciationaswellasinvasionprocesses.Theseprocessesareexpectednottobeequallyeffectivealongelevationalgradientsduetospecificecological,spatial,andanthropogenicfilters,thuscontrollingthedispersalandestablishment of species. Here, we investigate speciation and invasion processesalongelevationalgradients.Weassessthevascularplantspeciesrichnessaswellasthenumberandpercentageofendemicspeciesandnon-nativespeciessystematicallyalongthreeelevationalgradientscoveringlargepartsoftheclimaticrangeofLaPalma,CanaryIslands.Speciesrichnesswasnegativelycorrelatedwithelevation,whilethepercentageofCanaryendemicspeciesshowedapositiverelationship.However,thepercentageofCanary–Madeiraendemicsdidnotshowarelationshipwithelevation.Non-native species richness (indicating invasion) peaked at 500m elevation andshowedaconsistentdeclineuntilabout1,200melevation.Abovethatlimit,nonon-nativespecieswerepresentinthestudiedelevationalgradients.Ecological,anthropo-genic,andspatialfilterscontrolrichness,diversification,andinvasionwithelevation.With increase in elevation, richness decreases due to species–area relationships.Ecologicallimitationsofnativeruderalspeciesrelatedtoanthropogenicpressureareinlinewiththeabsenceofnon-nativespeciesfromhighelevationsindicatingdirec-tionalecologicalfiltering.Increaseinecologicalisolationwithelevationdrivesdiversi-fication and thus increased percentages of Canary endemics. The best preservedeasterntransect,includingmaturelaurelforests,isanexception.ThehighpercentageofCanary–Madeiraendemicsindicatesthecloudforest’senvironmentaluniqueness—andthusecologicalisolation—beyondtheMacaronesianislands.
K E Y W O R D S
alienspecies,altitudinalgradient,colonization,diversification,diversity,endemism,exotic,high-elevationecosystems,islandbiogeography

772 | STEINBAUER ET Al.
1 | INTRODUCTION
Elevational gradients on isolated mountains (especially on oceanichigh-elevationislands)poseauniqueopportunitytostudyimportantecological processes, including evolutionary dynamics (Steinbaueretal.,2016)andplantinvasion(Alexanderetal.,2011;Daehler,2005).For both processes, oceanic high-elevation islands offer a uniquesetting provided by the high rates of species evolved in situ (neo-endemism;Whittakeretal.,2007).
Islandstendtobemorepronetoinvasionthanareaswithhigherdegreesofconnectivityonthecontinent(island susceptibility hypothe-sis,Simberloff,1995)duetolowspeciesrichness(i.e.,ahigherrelativeecological opportunity), functional diversity, and reduced competi-tiveness of species compared to those of themainland (Sol, 2000).However,recentstudiesindicatethattheislandsusceptibilityhypoth-esisisnotgenerallyvalid.Presumably,theisolationofislands(smallerregional speciespool) also impedes theestablishmentofnon-nativespecies(Etherington,2015;Jeschke,2008;Vila,Pino,Montero,&Font,2010),whichnowadaysmainlyoccursviadirectanthropogenicvectors.
Introductionsofnon-nativespeciesarenotequallyprobablealongelevationalgradients.Accordingly,themajorityofnon-nativeplantspe-ciesisexpectedinlowerelevationsandlessextremehabitats,whereclimaticconditionsaremorefavorable,humandisturbancesaremostfrequent,and isolation is lowest (Becker,Dietz,Billeter,Buschmann,&Edwards,2005;Pauchardetal.,2009).Extremeclimaticconditionsof high-elevation ecosystems and low anthropogenic disturbancespreventtheestablishmentofnon-nativespecies(butseeexceptions,e.g.,Irl,Jentsch,&Walther,2013;Alexanderetal.,2016).Hence,itisprobablethatthemajorityofintroducednon-nativesarepre-adaptedtomid-orlowelevations(Pauchardetal.,2009).Accordingly,thelow-elevationalienfloramaybethemainspeciespoolforthealienfloraofhigh-elevationecosystems(McDougalletal.,2011).Incontrasttohigh-elevationspecialists,invasivenon-nativespeciestendtobegen-eraliststoleratingawiderangeofclimaticconditions(Alexanderetal.,2011;Higgins&Richardson,2014),althoughthedistributionpatternin the area of introduction depends on the interaction of traits oftheinvadingspeciesandtheenvironmentalconditions(Haideretal.,2010).
Onmountainousoceanic islands, low-elevationecosystemshavelarger spatial extent than high-elevation ecosystems—a typical fea-tureofmountainsystems(butseeElsen&Tingley,2015).Especiallytropicalandsubtropicalislandspossessabroadclimaticrangeduetowindward/leeward effects and thus a comparatively high absolutenumberofhabitats(Hortal,Triantis,Meiri,Thebault,&Sfenthourakis,2009;Irletal.,2015).Asaconsequence,mostspeciesonoceanicis-lands—non-natives, natives, or endemics—will be found in low- andmid-elevationecosystems(Zobeletal.,2011).Inordertounderstandinvasionprocesses,however,itisofhighinterestwhetherthepercent-ageofnon-nativespeciesincreaseswithelevation.Thepercentageofnon-nativespeciescanbeseenasanindicatorforinvasionprocessesindependent of overall richness (c.f.,Gillespie,Claridge,&Roderick,2008)althoughitisstillsusceptibletodifferencesindisturbances.
Recent studies suggest that diversification, as indicated by twofrequentlyusedindices(i.e.,thepercentageofsingle-islandendemicspeciesandthepercentageofarchipelagoendemicsspecies;Emerson&Kolm,2005;Whittakeretal.,2007),increaseswithelevation—apat-ternwhich has large-scale support for islands aswell as continen-tal mountains (Steinbauer etal., 2016). However, this trend can bemodifiedonan intra-insular levelbyotherenvironmental influencessuchasdifferentaspectsofclimate(esp.precipitation;Irletal.,2015)andshouldbestrongerthemorepronouncedenvironmentalchangesalongtheelevationalgradientare.Additionally,diversificationissup-portedbygeneticisolation(andhenceatendencytowardspeciation)andtheavailabilityofecologicalopportunitiesduetolowspeciesrich-ness(Hughes&Eastwood,2006;Steinbauer&Beierkuhnlein,2010).
Directional ecological filtering along elevational gradients(Alexander etal., 2011) causes isolation of high-elevation ecosys-tems(Gillespie,2004).Particularlyonoceanicislands,high-elevationecosystems tend to bemore isolated from the nearest ecosystemswithcomparableenvironment than their low-elevationcounterparts(Steinbaueretal.,2016).Thelatterarelessdistantfromsimilarhabitatsonotherislandsandonthemainland(Steinbauer,Irl,&Beierkuhnlein,2013).Inconsequence,high-elevationsystemsformislandswithinis-lands(Fernández-Palacios,Otto,Thebaud,&Price,2014).Elevationalgradientsthusprovideanexcellentnaturalsettingtoinvestigatetheeffectofisolationonspeciationandinvasionprocesses.
Here,wetestwithaspecificallydesignedfieldstudythefollowingpredictionsusingvegetationsurveysalongthreeelevationgradientsrangingmorethan2000monLaPalma(CanaryIslands).Basedoncur-rentknowledge,weexpect(1)thatthevascularplantspeciesrichnessaswellasthenumberofendemics(endemictoLaPalma,endemictotheCanaryIslandsorsharedbetweenMadeiraandtheCanaryIslands,respectively)decreases,whilethepercentageofendemicspeciesin-creaseswithelevation.(2)Theabsolutenumberofnon-nativespeciesdecreases,whilethepercentageofnon-nativespeciesincreaseswithelevation, if theavailabilityofunoccupiedhabitatsathigh-elevationecosystems significantly enhances the establishment of non-nativespecies.Thecontrary istobeexpected, iftheextremeenvironmen-talconditionsathighelevationsposeamoreeffectivefilterfornon-nativespeciesthanthealreadybypassedgeographicalisolation.
2 | MATERIALS AND METHODS
2.1 | Study area
LaPalmaisoneofthemostwesternandyoungestislandsoftheCanaryarchipelago(~1.7Ma;Carracedoetal.,2002)expressingcontinuousvol-canicactivity(Prietoetal.,2009).Theislandcovers706km²andreachesamaximumelevationof2426ma.s.l.attheRoquedelosMuchachos(Figure1).ThenorthernpartofLaPalmaiscomprisedbytheCalderadeTaburientecomplexandistheoldestandsteepestpartoftheisland,thusofferingalsosteepenvironmentalgradients(Irletal.,2015).
Owingtoitsnorthwesternposition,LaPalmareceivesfairamountsofprecipitationinwinter,yetextensivedryperiodsarecommononthewesternsideoftheislandandathighelevations(Irl&Beierkuhnlein,

| 773STEINBAUER ET Al.
2011). Besides elevation, the dominating climatic influence factorsarehumidtradewindsregularlyblowingfromthenortheast(Garzón-Machado,Otto&delArcoAguilar,2014)anddividingtheislandintoahumidwindwardsideandadroughtandfire-proneleewardside.Thus,themainprecipitationgradientoftheislandrunsalonganortheast–southwestaxisreachingfromthewesterncoast(approx.120mm/a)tothehigh-elevationnortheasternslopesoftheCalderadeTaburiente(approx.1,300mm/a;AEMET,2012).Abovethecloudbank,athermalinversion layer exists that impedes an orographic rise of moist airmasses eliminating the tradewind influence.Above the thermal in-version layer,highsolar radiationandhighdaily temperatureampli-tudesarecommon.There,wintersnowappearsregularlyandevenicestormscanoccur(Garzón-Machadoetal.,2014).
2.2 | Sampling design
Threetransectsforfloristicinventorywereinstalledacrossthecom-pleteelevationalgradient.Onereachedthecalderarimfromthedrywestcoast,onefromthemorehumidnorthcoast,andonefromthenortheastthroughthelaurelforestbelt,thus,systematicallydesignedtocoverallvegetationzonesandthecompleteprecipitationgradientofferedbyLaPalma(Figure1).Withineachtransect,weinstalledplots(10×10m) atpreselectedelevational levels.Criteria forplot selec-tionwere accessibility and a slope not higher that 35°, but alwaysuninfluenced by roads. Along the western and northern transects,weestablished21plotsdistributedevenlybetweensevenelevationlevels spanning100mand separatedby300-m intervals (note: theintervalbetweenthesecondhighestandthehighestelevationlevelonly measured 200m, because a maximum elevation of 2,400ma.s.l.couldnotbeexceeded).Threereplicatesper levelwereestab-lished,which is in accordancewith guidelines for optimal samplingalong environmental gradients (Schweiger, Irl, Steinbauer, Dengler,&Beierkuhnlein,2016).DatasamplingonthewesternandnortherntransectstookplaceinFebruary2009.Alongtheeasterntransect,24plotsdistributedacross12elevationlevelswith200-mintervalswereinstalled.Thus,tworeplicateswereestablishedper level.Datasam-plingfortheeasterntransecttookplacefromFebruarytoApril2013.AllrawdataareprovidedinonlineAppendix.
Werecordedallfloweringplantspeciesandclassifiedthemassingle-island endemics, archipelago endemics (i.e., endemic to the CanaryIslands), and non-native and nonendemic (including definitely native,possiblynativeandprobablynative)accordingtoAcebesGinovésetal.(2010).Inourstudy,thegroupofnon-nativesconsistsofallsub-classesgivenbyAcebesGinovésetal.(2010),thatis,differentiatedintointro-ducedspecies,probablyintroducedspeciesandintroducedinvasives.
As the Canary Islands share a high number of endemic specieswithMadeira,wealsoclassifiedCanary–Madeira endemics.ThisgroupincludesalltheCanaryendemicsplusthespecieswithagloballyre-stricted distribution range to the Canary Islands andMadeira. TheCanary Islands andMadeira currently are located at a distance lessthan400kmandwereevencloserduringtimeperiodswithlowersealevel(Fernández-Palaciosetal.,2011).Wefurtheridentifiedasubsetof Madeira–Canary endemics by excluding those species that onlyoccurredontheCanaries—thusfocusingonlyonendemicspeciesin-habitingbotharchipelagos(furtheronreferredtoasMadeira–Canary twins).
2.3 | Statistical analysis
Therelationbetween indices (forrichness,endemism,and invasion)with elevationwas assessed using generalized linearmixedmodels(R package lme4 version 1.1-7, Bates,Maechler, Bolker, &Walker,2015). Fullmodels includedelevation,elevation2, transect aswell astheinteractionsbetweentransectandelevationasexplanatoryvari-ables (y~elevation+elevation2+transect+transect:elevation). In afurther step, elevation, elevation2, and/or the interaction betweentransectandelevationwasremoved ifnotsignificantlycontributingto theexplanatorypowerof themodel. The laterwas testedusingaFtest(ANOVA)betweenamodelincludingandoneexcludingthefocalvariable (backwardselection;Table1providesanoverviewonfinalmodelspecifications).Asvegetationplotsarereplicatedperel-evationallevel,weincludedablockfactorintherandomstructureofthemodelstoaccountforthisspatialclustering.
Poisson error distribution (log-link function) was used for allrichness-based indices, while binomial error distribution (logit-link function) was used for percentage values. The binomial error
F IGURE 1 MapofLaPalma.LaPalmaisthenorthwesternmostislandoftheCanaryIslandssituatedoffthecoastofMorocco(Africa)intheAtlanticOcean.Themapillustratesthelocationofthewestern(circles,n = 21),northern(squares,n = 21),andeasterntransects(triangle,n = 24)coveringalargeelevationalgradientofmorethan2400mindetail.Duetothepredominanttradewinddirectionfromthenortheast,precipitationincreasesfromwesterntoeasterntransect,especiallyatmid-elevations
774 | STEINBAUER ET Al.
distribution forpercentagevalueshas theadvantage toaccount forthefactthataparticularpercentagevalueismorepreciseifitisbasedonalargernumberofobservations(herethenumberofspecies).Notethatpercentageofendemicspeciesiscalculatedexcludingnon-nativespecies to get a cleaner signal for diversification. Patterns remain,however,ifnon-nativeswereincluded.
Goodness-of-fitstatisticsfortheimplementedmodelswereper-formedcalculatingmarginalandconditionpseudo-R2asimplementedinRpackagepiecewiseSEM (version1.1.3;Lefcheck,2015)basedonNakagawa and Schielzeth (2013). All statistical analyses were per-formedinRversion3.2.0(RDevelopmentCoreTeam,2015).
3 | RESULTS
3.1 | Diversification along an elevation gradient
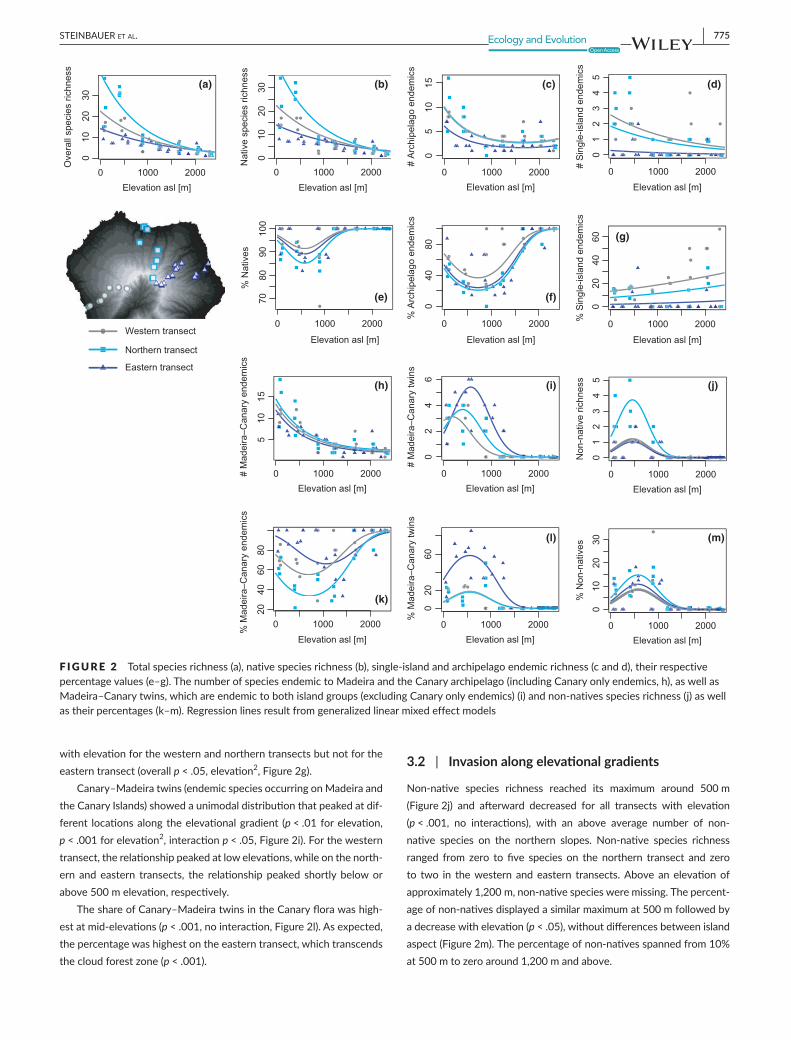
Plantspecies richnessshowedahighlysignificantdecreasewith in-crease in elevation (p<.001). The northern transect showed bothhigherspeciesrichness(p > .01)andasteeperslope(Figure2a),asin-dicatedbyasignificantinteractionwithelevation(p > .05).Plantspe-ciesrichnessdecreasedfromvaluesatthelowestelevation(approx.100ma.s.l.).Richnessvaluesreachedfrommorethan30speciesonthenorthern througharoundhalfof thaton thewesternandeast-erntransectstolessthanthreespeciesinthesummitregion(approx.2,300melevation).Trendsremainedsimilarifnativespeciesrichnesswasanalyzed(non-nativespeciesnotincluded), indicatingtheirhighshareintotalspeciesnumbers(Figure2b).
Canary archipelago endemic richnesswas low atmid- and highelevationswithfewerthanfourspeciesperplot.Canaryarchipelagoendemicrichnessgenerallydecreasedwithelevationforalltransects,with similar declines from about ten species at 100m elevation toabout four speciesat thesummit (p < .001 forelevation,p < .01 for
elevation2,nointeractionbetweenelevationandtransect,Figure2c).ThenumberofCanaryarchipelagoendemicswassignificantlyhigheron the northern and western transects, with the difference beingabouttwotothreespecies.ThisdifferencedisappearsifthedefinitionofendemismincludessharedCanary–Madeiraarchipelagoendemics(Figure2h).
ThepercentageofCanaryarchipelagoendemicsdisplayedamin-imum around 750m for all transects (p < .001)with overall highestpercentage values (100%) in high elevations. Thewestern transectexpressedonaveragehigherpercentagevaluesthanthenorthernandeasternones(Figure2f,p < .001).ThepatternswitchesifthedefinitionofendemismincludedMadeira.TheproportionofCanary–Madeiraar-chipelagoendemicsishighestintheeasterntransectfollowedbythewesterntransect(p < .01).ThepercentageofMadeira–Canaryarchi-pelagoendemicsshowsamid-elevationminimumat1,000–1,400minteractingwithanoverallincreasewithelevationforthewesternandnortherntransectsthatisnotpresentontheeasterntransect(interac-tionp < .05,Figure2k).Theeasterntransectalsodifferedbyshowingamuchhigherproportionofendemicsreachingupto80%–100%.
Theoverallnumberofsingle-islandendemicspeciesislow(n = 14),makingtheir interpretationtentative.Single-islandendemicrichnessperplotshowedsignificantdifferencesbetweentransects (no inter-action), slightly declined on thewestern transect and increased onthenortherntransect(p < .01)butdidnotchangealongtheelevationgradientontheeasterntransect.Thenumberofsingle-islandendem-icsperplot increased from theeastern transect through thenorth-erntransecttothewesterntransect(p < .001,Figure2d).Overall,thewesterntransecthadnineandthenortherntransecteightsingle-islandendemicspeciescomparedtofourspeciesontheeasterntransect.
Thepercentageofsingle-islandendemicsdifferedsignificantlybe-tweentransects,withthewesterntransecthavingthehighestvalues.Thepercentageofsingle-islandendemicspeciesshowedanincrease
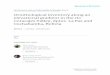
TABLE 1 Modelstructurefordifferentdependentvariables
Variable Model structure Family R2
MargR2
Cond
Overallrichness y~elevation***+transectn.sig+elevation:transect*+(1|block) Poisson .74 .79
Nativerichness y~elevation***+transectn.sig+elevation:transect*+(1|block) Poisson .72 .77
%Natives y~elevation*+elevation2*+transectn.sig+(1|block) Binomial .81 .81
Archipelagoendemics y~elevation***+elevation2**+transect*+(1|block) Poisson .40 .48
%Archipelagoendemics y~elevation***+elevation2***+transect*+(1|block) Binomial .71a –
Islandendemics y~elevation*+transect***+(1|block) Poisson .44 .56
%Islandendemics y~elevation2*+transect***+(1|block) Binomial .32a –
Canary–Madeiraendemics y~elevation***+elevation2*+transectn.sig+(1|block) Poisson .59 .59
%Canary–Madeiraendemics y ~elevation***+elev.2***+transect**+elev.:transect*+(1|block) Binomial .66a –
Madeira–Canarytwins y~elevation***+elev.2***+transectn.sig+elev.:transect**+(1|block) Poisson .83a –
%Madeira–Canarytwins y~elevation**+elevation2**+transect***+(1|block) Binomial .82 .83
Non-natives y~elevation*+elevation2**+transect**+(1|block) Poisson .77a –
%Non-natives y~elevation***+elevation2*+transectn.sig+(1|block) Binomial .50a –
Formodelsmarkedwithan“a”attheR2Marg,pseudo-R2wascalculatedafterremovingtherandomstructure(blockeffect)fromthemodelasthefunction
piecewiseSEMfailedtoprovidepseudo-R2valuesforthemixedeffectmodel.Significanceofexplanatoryvariables(basedonF-statistics)isindicatedwithstars(p < .05*;p < .01**;p < .001***).Marginal(R2
Marg)andconditional(R2
Cond)
pseudo-R2werecalculatedbasedonRpackagepiecewiseSEM(Lefcheck,2015).
| 775STEINBAUER ET Al.
withelevationforthewesternandnortherntransectsbutnotfortheeasterntransect(overallp < .05,elevation2,Figure2g).
Canary–Madeiratwins(endemicspeciesoccurringonMadeiraandtheCanaryIslands)showedaunimodaldistributionthatpeakedatdif-ferent locationsalongtheelevationalgradient (p < .01forelevation,p < .001forelevation2,interactionp < .05,Figure2i).Forthewesterntransect,therelationshippeakedatlowelevations,whileonthenorth-ern andeastern transects, the relationshippeaked shortlybeloworabove500melevation,respectively.
TheshareofCanary–MadeiratwinsintheCanaryflorawashigh-estatmid-elevations(p < .001,nointeraction,Figure2l).Asexpected,thepercentagewashighestontheeasterntransect,whichtranscendsthecloudforestzone(p < .001).
3.2 | Invasion along elevational gradients
Non-native species richness reached its maximum around 500m(Figure2j) and afterward decreased for all transects with elevation(p < .001, no interactions), with an above average number of non-native species on the northern slopes. Non-native species richnessranged from zero to five species on the northern transect and zeroto two in thewestern andeastern transects.Aboveanelevationofapproximately1,200m,non-nativespeciesweremissing.Thepercent-ageofnon-nativesdisplayedasimilarmaximumat500mfollowedbyadecreasewithelevation(p < .05),withoutdifferencesbetweenislandaspect(Figure2m).Thepercentageofnon-nativesspannedfrom10%at500mtozeroaround1,200mandabove.
F IGURE 2 Totalspeciesrichness(a),nativespeciesrichness(b),single-islandandarchipelagoendemicrichness(candd),theirrespectivepercentagevalues(e–g).ThenumberofspeciesendemictoMadeiraandtheCanaryarchipelago(includingCanaryonlyendemics,h),aswellasMadeira–Canarytwins,whichareendemictobothislandgroups(excludingCanaryonlyendemics)(i)andnon-nativesspeciesrichness(j)aswellastheirpercentages(k–m).Regressionlinesresultfromgeneralizedlinearmixedeffectmodels
Western transect
Northern transect
Eastern transect
(a) (b) (c) (d)
(e) (f)
(g)
(h) (i) (j)
(k)
(l) (m)
Elevation asl [m]
Nat
ive
spec
ies
richn
ess
Elevation asl [m]
# A
rchi
pela
go e
ndem
ics
Elevation asl [m]
# S
ingl
e-is
land
end
emic
s
Elevation asl [m]
Non
-nat
ive
richn
ess
Elevation asl [m]
# M
adei
ra–C
anar
y en
dem
ics
Elevation asl [m]
# M
adei
ra–C
anar
y tw
ins
Elevation asl [m]
% N
ativ
es
Elevation asl [m]
% A
rchi
pela
go e
ndem
ics
Elevation asl [m]
% S
ingl
e-is
land
end
emic
s
Elevation asl [m]
% N
on-n
ativ
es
Elevation asl [m]
% M
adei
ra–C
anar
y en
dem
ics
Elevation asl [m]
% M
adei
ra–C
anar
y tw
ins
Elevation asl [m]
Ove
rall
spec
ies
richn
ess
010
2030
0 1000 2000
010
2030
0 1000 2000
7080
9010
0
0 1000 2000
040
80
0 1000 2000
020
4060
0 1000 2000
010
2030
0 1000 2000
05
1015
0 1000 2000
2040
6080
0 1000 2000
020
60
0 1000 2000
01
23
45
0 1000 2000
02
46
0 1000 2000
01
23
45
0 1000 2000
510
15
0 1000 2000

776 | STEINBAUER ET Al.
4 | DISCUSSION
4.1 | Species diversity and diversification along elevational gradients
Inour study,plant species richnessaswell as thenumberofarchi-pelago (Canary and Canary–Madeira) endemic species decreasedwithelevation,acommonpatternwhichwaspreviouslyfoundonLaPalma(Irl,Steinbauer,Epperlein,etal.,2014)andotherislands(Trigas,Panitsa,&Tsiftsis,2013).Adecreaseinsurfaceareaofindividualel-evationalzoneswithincreaseinaltitudeismaderesponsibleforthiseffect(Lomolino,2000;butseeElsen&Tingley,2015).Furthermore,area is often seen as a surrogate for habitat diversity (Hortal etal.,2009). Larger areasmight support larger population sizes,which inturndecreasestheriskofextinction(MacArthur,1972).However,asweusedastandardizedsamplingdesignwithequallysizedplots,thereisnoeffectof(plot)areaonspeciesrichnessinourstudy.Rather,localspecies richnessmay be enhanced due to “sampling” from a largerspeciespoolgenerallyavailableatlowelevations(Kargeretal.,2011).Harshenvironmentalconditionswithclimaticextremeeventsathighelevationsmightadditionallydecreasetotalandnativespeciesrich-nessanddensity(habitatharshnesshypothesissensuDefeo,Gomez,&Lercari,2001).
Thegeographic isolation that reduces thenumberof individualspre-adaptedtohigh-elevationecosystemsarrivingonanislandmayen-hancediversificationbygeneticisolation(nonadaptiveallopatricspe-ciation).Speciescommunitiesfilteredbyisolationmayfurtherprovideecologicalopportunity for adaptive radiation tofill gaps in resourceusagethatareleftunoccupied(Steinbaueretal.,2016).Opportunity-drivendiversification is supportedbyphylogenetic studies (Bennett&O’Grady,2013).BothmechanismscanhelptoexplaintheincreaseinthepercentageofCanaryarchipelagoendemicsaswellastheper-centageofsingle-islandendemicswithelevation,expressingahigherdegreeofendemisminhigh-elevationecosystems(Steinbauer,Otto,Naranjo-Cigala,Beierkuhnlein,&Fernández-Palacios,2012;Verboom,Bergh,Haiden,Hoffmann,&Britton,2015). In contrast to richness-based indices (like the number of endemics), the percentage of en-demicspeciesindicatestowhichdegreelocalspeciespoolsonoceanicislandsoriginate from insituspeciation (thealternativeoriginbeingcolonizationfromotherareas,Irletal.,2015).
In summary, two explanations for the observed pattern in thethreeelevationalgradientsareinaccordancewiththesuggestedrel-ativeimmaturityofhigh-elevationecosystemscomparedtolowerel-evations (Fernández-Palaciosetal., 2014). First, in lowerelevations,species cannot realize their complete potential distribution due tobioticinteractions.Suchcompetitiveinteractions,however,decreasewithelevation,whichisparticularlytrueforhigh-elevationecosystemsowingtotheirlowspeciesrichness(butseeIrletal.,2012).Second,thespatial isolationofhigh-elevationspeciesorcladesprovidesthepossibilityfordiversification(speciationenhancedbyisolationaswellasbyecologicalopportunity;Steinbaueretal.,2016).Theimmigrationofpopulationsfromotherislandsdecreaseswithelevationduetolon-gerdistancestosourceregionsandsmallerareas(Merckxetal.,2015;
Steinbaueretal.,2013).Inaddition,locallydisturbancesareanaddi-tionalkeyfactorinfluencingthehigherpercentageofCanaryarchipel-agoandsingle-islandendemics.Particularly,plotsinthelowerpartsofthewesternandnortherntransects(0–800ma.s.l.)haveahighfrac-tionofnativespeciesthatareassociatedwithhabitatsmodifiedand/ordisturbedbyhumans (ruderalspecies),whichmight influencethepercentageofendemics.
On theeastern transect,which isbetterpreservedcompared tothenortherntransect(especiallythelaurelforest),theproportionofCanary–Madeiraendemicsdoesnot increasewithelevation. Incon-trast,single-islandendemicspeciesarevirtuallyabsentfromthistran-sect, not showingdifferencesbetween lowerandhigherelevations,andCanaryarchipelagoendemicsrichnessislow.Onereasonforthismaybethefactthatthelaurelforestsharesalargenumberoffloralel-ementswithMadeira(asisreflectedinthenumberofCanary–Madeiraarchipelagoendemics;Figure2i).Thishighlightsthatdispersalvectors(andthusisolation)differbetweenthemorehumidlaurophyllouseast-ernvegetationandthedrierwesternpartsofLaPalmaindicatingthatdispersalcharacteristicsvarybetweendifferentvegetationunitsevenwithinasingle island(Nogalesetal.,2015).However,futurestudieswillneedtoidentifywhetherdispersaltraitsdifferbetweenvegetationzonesonislandsandhowthisinfluencesdiversification.
Finally,ecologicalisolation,notonlygeographicalisolation,mightberesponsibleforthehighdiversificationinthelaurelforests.SincetheonsetoftheMediterraneanclimate, laurelforestsonlypersistedinMacaronesia,wheretheoceanicenvironmentbufferedtheclimaticoscillationsofthePleistocene(Fernández-Palaciosetal.,2011;Patiñoetal.,2016).Insummary,wesuggestthatenvironmentalfiltering,an-thropogenicdisturbance,andspatialandecologicalisolationofcom-munitiestobethemainfactorsresponsiblefortheobservedchangesinspatialtrendswithelevation.
4.2 | Elevational threshold for non- natives along the elevational gradients
Non-nativespeciesrichnessonLaPalmashowsaclearpeakat500m,probablyassociatedwiththemaincentersofpopulationandagricul-ture in this elevational zone (del Arco Aguilar, González-González,Garzón-Machado,& Pizarro-Hernández, 2010). Beyond that, eleva-tionexhibitsastrongnegativeinfluenceonnon-nativespeciesrich-nessandthepercentageofnon-nativespecies.Aboveabout1,200ma.s.l., non-native species weremissing in this study. Environmentalconditions above this threshold seem to be unfavorable for non-nativeplantspeciesonLaPalma,atleastatthisstateofthe(dynamic)invasionprocess.Inthefollowing,wemakeseveralassumptionsfortheexistenceofthisthreshold:
1. The1,200-m threshold corresponds verywellwith the transitionfrom cultivated lowland vegetation (i.e., plots located mainly inextensivelymanagedrangelandorrecentlyabandonedcultivation)to the pine forests (although cultivation can reach higher ele-vations in other areas of La Palma). Above 1,200m elevation,only extensive forestry is practiced in the sampled area due to

| 777STEINBAUER ET Al.
increasing steepness and deteriorating climatic conditions. As aconsequence,humandisturbancesassociatedwithagricultureandthe probability of an anthropogenic introduction of non-nativespeciesaregreatlyreducedabovethe1,200-mthreshold.However,historiclanduseinformofherding,fire,andhuntinghasaffectedandstillaffectsthewholeenvironmentalgradient(Garzón-Machadoetal., 2010; Irl, Steinbauer, Messinger, etal., 2014).
2. Pinus canariensisforestsarehighlydominantabovethisthreshold(1,200–1,500m). This vegetation type has the highest fire fre-quencyontheCanaryIslands.Onlynon-nativespecieswitharela-tivelyhighfiretolerancecanpotentiallyestablishinthevegetationzoneof thepine forest; thus,firemightactasa strongfilter forpotential invaders particularly asmany fire-prone ecosystems inotherpartsof theworldarenonforested.However,firecanalsocauseashort-termincreaseinnativeandnon-nativeannualplants(mostlyruderals;GonzálezGómezetal.,2011).Athighelevations,firetogetherwithharshconditions(Kueffer,Pyšek,&Richardson,2013),suchashighsolarradiation,strongdiurnaltemperatuream-plitude,extensivesummeraridity,andwintersnowandicestorms(Garzón-Machadoetal.,2014),probablyfilterpotentialinvadersinthesummitscrub.
3. The sampling design of this study avoided plant communities ofpurehumanoriginsuchasroadsidecommunities,whichareoftenappliedforquestionsregardingplantinvasioninmountainsystems(see,e.g.,Alexanderetal.,2011;Arévaloetal.,2010;Ottoetal.,2014).RoadshavebeenshowntobeveryeffectivevectorsofplantinvasionandhomogenizationontheneighboringislandsofTenerife(Arévaloetal.,2010;Haideretal.,2010)andinothermountainousareasoftheworld(Alexanderetal.,2011).RoadsidecommunitiesathigherelevationsexistonLaPalmaaswell,wherenon-nativespeciessuchasReseda luteolaL.reachup intothehighesteleva-tions (unpublisheddata). In general, roadside communitiesmighthelp to identifymaximumelevations reachedbynon-nativespe-cies,indicatingphysiologicallimitsofnon-natives(e.g.,temperaturethresholds;Poll,Naylor,Alexander,Edwards,&Dietz, 2009), yetfocusingonmorenaturalcommunitiesisabettertooltodeterminethestateof invasibilityandmighthelp to identifyhowbiologicalconstraintssuchasinterspecificcompetitionlimitplantinvasion.
Introducedplantstendtobepre-adaptedtolow-elevationsitesbe-causeintroductionusuallytakesplaceatlowelevations.Thelowamountof non-native high-elevation specialists and the filtering effect of theP. canariensisforest(Alexanderetal.,2011)leadtotheabsenceofnon-nativespeciesinhigh-elevationecosystems.However,thismaybeonlyatransitionalsituation,asglobalclimatechangehasthepotentialtoalterthehigh-elevationharshnessfilteron islands, creatingpossibilities forinvasionatincreasingelevationsinthenearfuture(Harteretal.,2015).
Besides increasing temperatures, increasing moisture availabilityhas been shown to facilitate the invasion process (Maron&Marler,2007; Stohlgren etal., 1999).We observed highest values for non-nativespeciesrichnessonthenortherntransect.Thissupportsenvi-ronmentalfiltering resulting in lowernon-nativespecies richnessonthewesternsideoftheisland.Ontheotherhand,thedenseevergreen
canopyofthelaurelforestontheeasterntransectpreventsmostlightfrompenetratingtotheground(Delgado,Arroyo,Arévalo,&Fernández-Palacios,2007),whichpotentiallymightpreventtheestablishmentofheliophyllousnon-nativespecies.Generally,thetotalnumberofnon-nativeplants and their contribution to thevegetation structure andcompositionarelowonLaPalmacomparedwithmanyoceanicislands(Daehler,2005;Macdonald,Thébaud,Strahm,&Strasberg,1991).Thisfact, togetherwith the identifiedspatialpatterns,maycontribute tocopingandmanagementstrategiesinordertomitigatepossiblefuturenegativeimpactsofinvasivespeciesatanearlystage.
4.3 | Theoretical concept of invasion and speciation on oceanic islands
Invasionofnon-nativeplantspeciesanddiversificationofnativeanden-demicplantsaregenerallyseenastwoindependentprocesses.However,theunderlyingenvironmentaldriversonoceanicislands’elevationalgra-dients,thatis,environmentalfiltering,degreeofhumaninfluence,andgeographicfilteringintheformofspatialandecologicalisolation,seemto influence both processes, but in contrasting directions. Aiming todissectthepresumedinterrelationofunderlyingenvironmentaldriverswiththeseprocesses,Levin(2003)suggestedusingislandsasresearchobjectsbecauseoftheirexcellentsettingforthispurpose.
Consistent with phylogenetic findings (Merckx etal., 2015),Steinbaueretal.(2016)foundelevation-drivenisolationtopositivelyinfluence speciation. This seems to be a general phenomenon onhigh-elevationislandsbutalsoappliestomountainsonthecontinent,highlighting its importanceforglobalbiodiversitypatterns.Althoughanthropogenicinfluencesaffectcurrentspeciesdistributionpatterns,forexample,via theoccurrenceof ruderal species, theevolutionarysignalisstillstronginthespatialgradientsdetectedinourstudy.Thelower immigration ratesathigherelevations results in small speciespools,whereas the degree of endemism, as a proxy for diversifica-tion,ishigh(Merckxetal.,2015).Acombinationofdriverseffectivelyimpedes colonization of high elevations by natural immigration andcolonization.Amongthesedriversareharshclimaticconditions,de-creasingareaandisolationcausedbytheeffectivegeographicalisola-tionfromcomparableecosystems(Steinbaueretal.,2013,2016).
Atthecurrentstageofinvasion,high-elevationecosystemsseemtobebetterprotectedfrominvasionprocessesthanlowelevationsonLaPalma.Recentsynthesesshowthatthisisageneralphenomenonbecauseharshnesstendstodecreasetheinvasibilityofasystem(inva-sion syndrome;Kuefferetal.,2013).However,changesinlanduseandclimateinthenearfuturemightalterinvasionfiltersathighelevations,posingathreattotheseenvironments(Alexanderetal.,2016).Whilemany lowlandsystemsarepresumablynonrestorable,high-elevationsystemsstilloffertheopportunitytopreventlarge-scalealterationsbynon-nativespecies,ifimmediateprotectionmeasuresaretaken.
ACKNOWLEDGMENTS
WewouldliketothankDr.FélixMedinaandFranciscoPrietooftheConsejería deMedioAmbiente delCabildo Insular de LaPalma for

778 | STEINBAUER ET Al.
theirsupportandJulioLealPérezforhishelp inthefield.TheEliteNetworkofBavariawithintheframeoftheGlobalChangeEcologystudyprogramsupportedthefieldsurvey.ThisworkcontributestotheprojectECOPOTENTIALfundedfromtheEuropeanUnion’sHorizon2020 research and innovation program under grant agreement no.641762. This work further contributes to the project MOVECLIMunder the frameworkof theNet-Biome transnational first callwithsupportoftheAgenceNationaledelaRecherche,theConseilrégionaldelaRéunion,theConseilrégionaldelaGuadeloupe,theGovernmentofAzores,andtheGovernmentofCanarias.Additionally,thispubli-cationwas fundedby theGermanResearchFoundation (DFG) andthe University of Bayreuth in the funding program Open AccessPublishing.MJSwaspartlysupportedbytheDanishCarlsbergfondet(CF14-0148).
CONFLICT OF INTEREST
Nonedeclared.
REFERENCES
AcebesGinovés,J.R.,LeónArencibia,M.C.,RodríguezNavarro,M.L.,delArcoAguilar,M.,GarcíaGallo,A.,PérezdePaz,P.L.,...WildpretdelaTorre,W. (2010). Spermatophyta. InArechavaleta,M., S. Rodríguez,N. Zurita & A. García (Eds.), Lista de especies silvestres de Canarias Hongos, plantas y animales terrestres 2009(pp.122–172).SantaCruzdeTenerife:GobiernodeCanarias.
AEMET (2012). Atlas climático de los archipiélagos de Canarias, Madeira y Azores—Temperatura del aire y precipitación (1971–2000). Madrid,España:MinisteriodeAgricultura,AlimentaciónyMedioAmbiente.
Alexander,J.M.,Kueffer,C.,Daehler,C.C.,Edwards,P.J.,Pauchard,A.,etal.(2011).Assemblyofnonnativeflorasalongelevationalgradientsexplainedbydirectionalecologicalfiltering.Proceedings of the National Academy of Sciences of the United States of America,108,656–661.
Alexander,J.M.,Lembrechts,J.J.,Cavieres,L.A.,Daehler,C.,Haider,S.,et al. (2016). Plant invasions intomountains and alpine ecosystems:Currentstatusandfuturechallenges.Alpine Botany,126,89–103.
Arévalo,J.R.,Otto,R.,Escudero,C.,Fernández-Lugo,S.,Arteaga,M.,etal.(2010).Doanthropogeniccorridorshomogenizeplantcommunitiesatalocalscale?AcasestudiedinTenerife(CanaryIslands).Plant Ecology,209,23–35.
Bates,D.,Maechler,M.,Bolker,B.,&Walker,S.(2015).Fittinglinearmixed-effectsmodelsusinglme4.Journal of Statistical Software,67,1–48.
Becker, T., Dietz, H., Billeter, R., Buschmann, H., & Edwards, P. (2005).Altitudinal distribution of alien plant species in the Swiss Alps.Perspectives in Plant Ecology Evolution and Systematics,7,173–183.
Bennett, G. M., & O’Grady, P. M. (2013). Historical biogeography andecological opportunity in the adaptive radiation of native Hawaiian leafhoppers (Cicadellidae:Nesophrosyne).Journal of Biogeography,40,1512–1523.
Carracedo, J. C., PérezTorrado, F. J.,Ancochea, E.,Meco, J.,Hernán, F.,etal.(2002).CenozoicvolcanismII:TheCanaryIslands.InW.Gibbons,&T.Moreno (Eds.),The geology of Spain (pp.439–472).Bath,UnitedKingdom:GeologicalSocietyofLondon.
Daehler, C. C. (2005). Upper-montane plant invasions in the HawaiianIslands: Patterns and opportunities. Perspectives in Plant Ecology, Evolution and Systematics,7,203–216.
Defeo,O.,Gomez,J.,&Lercari,D.(2001).Testingtheswashexclusionhy-pothesisinsandybeachpopulations:ThemolecrabEmerita brasiliensis inUruguay.Marine Ecology Progress Series,212,159–170.
del Arco Aguilar, M.-J., González-González, R., Garzón-Machado, V., &Pizarro-Hernández, B. (2010). Actual and potential natural vegeta-tionontheCanaryIslandsanditsconservationstatus.Biodiversity and Conservation,19,3089–3140.
Delgado, J. D.,Arroyo, N. L.,Arévalo, J. R., & Fernández-Palacios, J.M.(2007). Edge effects of roads on temperature, light, canopy cover,andcanopyheightinlaurelandpineforests(Tenerife,CanaryIslands).Landscape and Urban Planning,81,328–340.
Elsen,P.R.,&Tingley,M.W.(2015).Globalmountaintopographyandthefateofmontanespeciesunderclimatechange.Nature Climate Change,5,772–776.
Emerson,B.C.,&Kolm,N.(2005).Speciesdiversitycandrivespeciation.Nature,434,1015–1017.
Etherington, T. R. (2015). Geographical isolation and invasion ecology.Progress in Physical Geography,39,697–710.
Fernández-Palacios,J.M.,deNascimento,L.,Otto,R.,Delgado,J.D.,García-del-Rey,E.,etal.(2011).AreconstructionofPalaeo-Macaronesia,withparticularreferencetothelong-termbiogeographyoftheAtlanticis-landlaurelforests.Journal of Biogeography,38,226–246.
Fernández-Palacios,J.M.,Otto,R.,Thebaud,C.,&Price,J.(2014).Overviewof habitat history in subtropical oceanic island summit ecosystems.Arctic, Antarctic, and Alpine Research,46,801–809.
Garzón-Machado,V.,Otto,R.,&delArcoAguilar,M.J.(2014).Bioclimaticandvegetationmappingofa topographically complexoceanic islandapplying different interpolation techniques. International Journal of Biometeorology,58,887–899.
Garzón-Machado, V., González-Mancebo, J. M., Palomares-Martínez, A.,Acevedo-Rodríguez,A.,Fernández-Palacios,J.M.,etal.(2010).StrongnegativeeffectofalienherbivoresonendemiclegumesoftheCanarypineforest.Biological Conservation,143,2685–2694.
Gillespie, R. (2004). Community assembly through adaptive radiation inHawaiianspiders.Science,303,356–359.
Gillespie,R.G.,Claridge,E.M.,&Roderick,G.K.(2008).Biodiversitydy-namicsinisolatedislandcommunities:Interactionbetweennaturalandhuman-mediatedprocesses.Molecular Ecology,17,45–57.
González Gómez, S., Torres-Díaz, C., Valencia, G., Torres-Morales, P.,Cavieres,L.A.,&Pausas,J.G.(2011).AnthropogenicfireincreasealienandnativeannualspeciesintheChileancoastalmatorral.Diversity and Distributions,17,58–67.
Haider,S.,Alexander,J.,Dietz,H.,Trepl,L.,Edwards,P.J.,etal.(2010).Theroleofbioclimaticorigin,residencetimeandhabitatcontextinshapingnon-nativeplantdistributionsalonganaltitudinalgradient.Biological Invasions,12,4003–4018.
Harter,D.E.V.,Irl,S.D.H.,Seo,B.,Steinbauer,M.J.,Gillespie,R.,Triantis,K.A.,…Beierkuhnlein,C.(2015).Impactsofglobalclimatechangeonthe floras of oceanic islands—Projections, implications and currentknowledge.Perspectives in Plant Ecology, Evolution and Systematics,17,160–183.
Higgins, S. I., & Richardson, D.M. (2014). Invasive plants have broaderphysiologicalniches.Proceedings of the National Academy of Sciences of Sciences of the United States of America,111,10610–10614.
Hortal,J.,Triantis,K.A.,Meiri,S.,Thebault,E.,&Sfenthourakis,S.(2009).IslandSpeciesrichness increaseswithhabitatdiversity.The American Naturalist,174,E205–E217.
Hughes,C.,&Eastwood,R.(2006).Islandradiationonacontinentalscale:Exceptional rates of plant diversification after uplift of the Andes.Proceedings of the National Academy of Sciences of the United States of America,103,10334–10339.
Irl,S.D.H.,&Beierkuhnlein,C.(2011).Distributionofendemicplantspe-ciesonanoceanic island—Ageospatial analysisofLaPalma (CanaryIslands).Procedia Environmental Sciences,7,170–175.
Irl,S.D.H.,Harter,D.E.V.,Steinbauer,M.J.,GallegoPuyol,D.,Fernández-Palacios,J.M.,Jentsch,A.,&Beierkuhnlein,C.(2015).Climatevs.to-pography—Spatialpatternsofplantspeciesdiversityandendemismonahighelevationisland.Journal of Ecology,103,1621–1633.

| 779STEINBAUER ET Al.
Irl,S.D.H.,Jentsch,A.,&Walther,G.-R.(2013).PapavercroceumLEDEB:ArareexampleofanalienspeciesinalpineenvironmentsoftheUpperEngadine,Switzerland.Alpine Botany,123,21–30.
Irl, S.D.H., Steinbauer,M. J., Epperlein, L.,Harter,D. E.V., Jentsch,A.,Pätz, S.,…Beierkuhnlein, C. (2014).The hitchhiker’s guide to islandendemism—Biodiversityandendemicperennialplantspeciesinroad-side and surrounding vegetation. Biodiversity and Conservation, 23,2273–2287.
Irl,S.D.H.,Steinbauer,M.J.,Messinger,J.,Blume-Werry,G.,Palomares-Martínez, Á., Beierkuhnlein, C., & Jentsch, A. (2014). Burned anddevoured—Introduced herbivores, fire, and the endemic flora of thehigh-elevationecosystemonLaPalma,CanaryIslands.Arctic, Antarctic and Alpine Research,46,859–869.
Irl,S.D.H.,Steinbauer,M.J.,Strohmeier,S.,Babel,W.,Palomares-Martínez,A.,etal. (2012).Mono-dominanceofanendemic legumeshrubsug-gestsherbivoryasthemajorthreattohigh-elevationislandbiodiver-sity.Journal of Vegetation Science,23,1114–1125.
Jeschke,J.M. (2008).Across islandsandcontinents,mammals aremoresuccessfulinvadersthanbirds.Diversity and Distributions,14,913–916.
Karger,D.N.,Kluge,J.,Krömer,T.,Hemp,A., Lehnert,M.,&Kessler,M.(2011).Theeffectofareaonlocalandregionalelevationalpatternsofspeciesrichness.Journal of Biogeography,38,1177–1185.
Kueffer,C.,Pyšek,P.,&Richardson,D.M.(2013).Integrativeinvasionsci-ence: Model systems, multi-site studies, focused meta-analysis andinvasionsyndromes.New Phytologist,200,615–633.
Lefcheck,J.S.(2015).piecewiseSEM:Piecewisestructuralequationmodel-inginRforecology,evolution,andsystematics.Methods in Ecology and Evolution,7,573–579.
Levin,D.A. (2003).Ecological speciation:Lessons from invasive species.Systematic Botany,28,643–650.
Lomolino, M. (2000). Ecology’s most general, yet protean pattern: Thespecies-arearelationship.Journal of Biogeography,27,17–26.
MacArthur,R.H.(1972).Geographical ecology: Patterns in the distribution of species.NewYork,NY:Harper&Row.
Macdonald,I.A.,Thébaud,C.,Strahm,W.A.,&Strasberg,D.(1991).Effectsof alien plant invasions on native vegetation remnants on La Réunion(MascareneIslands,IndianOcean).Environmental Conservation,18,51–61.
Maron, J.,&Marler,M. (2007).Nativeplant diversity resists invasion atbothlowandhighresourcelevels.Ecology,88,2651–2661.
McDougall,K.L.,Alexander,J.M.,Haider,S.,Pauchard,A.,Walsh,N.G.,et al. (2011). Alien flora of mountains: Global comparisons for thedevelopment of local preventive measures against plant invasions.Diversity and Distributions,17,103–111.
Merckx,V.S.F.T.,Hendriks,K.P.,Beentjes,K.K.,Mennes,C.B.,Becking,L.E.,etal.(2015).Evolutionofendemismonayoungtropicalmountain.Nature,524,347–350.
Nakagawa,S.,&Schielzeth,H. (2013).Ageneral and simplemethod forobtainingR2fromgeneralizedlinearmixed-effectsmodels.Methods in Ecology and Evolution,4,133–142.
Nogales, M., Heleno, R., Rumeu, B., González-Castro, A., Traveset, A.,Vargas, P., & Olesen, J. M. (2015). Seed-dispersal networks on theCanariesandtheGalápagosarchipelagos:Interactionmodulesasbio-geographicalentities.Global Ecology and Biogeography,25,912–922.
Otto, R., Arteaga, M. A., Delgado, J. D., Arévalo, J. R., Blandino, C., &Fernández-Palacios,J.M.(2014).Roadedgeeffectandelevationpat-ternsofnativeandalienplantsonanoceanicisland(Tenerife,CanaryIslands).Folia Geobotanica,49,65–82.
Patiño,J.,Mateo,R.G.,Zanatta,F.,Marquet,A.,Aranda,S.C.,Borges,P.A.V.,…Vanderpoorten,A. (2016).Climate threaton theMacaronesianendemic bryophyte flora. Scientific Reports, 6, 29156. doi:10.1038/srep29156
Pauchard, A., Kueffer, C., Dietz, H., Daehler, C. C., Alexander, J., et al.(2009).Ain’tnomountainhighenough:Plantinvasionsreachingnewelevations.Frontiers in Ecology and the Environment,7,479–486.
Poll,M.,Naylor,B.J.,Alexander,J.M.,Edwards,P.J.,&Dietz,H. (2009).SeedlingestablishmentofAsteraceaeforbsalongaltitudinalgradients:Acomparisonoftransplantexperimentsinthenativeandintroducedranges.Diversity and Distributions,15,254–265.
Prieto,J.F.,Gonzalez,P.J.,Seco,A.,Rodriguez-Velasco,G.,Tunini,L.,etal.(2009). Geodetic and structural research in La Palma, Canary Islands,Spain:1992–2007results.Pure and Applied Geophysics,166,1461–1484.
RDevelopmentCoreTeam(2015).R: A language and environment for statisti-cal computing.Vienna,Austria:RFoundationforStatisticalComputing.
Schweiger,A.H.,Irl,S.D.H.,Steinbauer,M.J.,Dengler,J.,&Beierkuhnlein,C.(2016).Optimizingsamplingapproachesalongecologicalgradients.Methods in Ecology and Evolution,7,463–471.
Simberloff,D.(1995).Whydointroducedspeciesappeartodevastateis-landsmorethanmainlandareas?Pacific Science,49,87–97.
Sol,D.(2000).Areislandsmoresusceptibletobeinvadedthancontinents?Birdssayno.Ecography,23,687–692.
Steinbauer,M.J.,&Beierkuhnlein,C.(2010).Characteristicpatternofspe-ciesdiversityontheCanaryIslands.Erdkunde,64,57–71.
Steinbauer,M.J.,Field,R.,Grytnes,J.A.,Trigas,P.,Ah-Peng,C.,etal.(2016).Topography-drivenisolation,speciationandagloballyconsistentpat-ternofendemism.Global Ecology and Biogeography,25,1097–1107.
Steinbauer,M.J.,Irl,S.D.H.,&Beierkuhnlein,C.(2013).Elevation-drivenecologicalisolationpromotesdiversificationonMediterraneanislands.Acta Oecologica,47,52–56.
Steinbauer, M. J., Otto, R., Naranjo-Cigala, A., Beierkuhnlein, C., &Fernández-Palacios,J.M.(2012).Increaseofislandendemismwithal-titude—Speciationprocessesonoceanicislands.Ecography,35,23–32.
Stohlgren,T.,Binkley,D.,Chong,G.,Kalkhan,M.,Schell,L.,etal. (1999).Exoticplantspeciesinvadehotspotsofnativeplantdiversity.Ecological Monographs,69,25–46.
Trigas,P.,Panitsa,M.,&Tsiftsis,S.(2013).Elevationalgradientofvascularplantspecies richness and endemism in Crete—The effect of post-isolationmountainupliftonacontinentalislandsystem.PLoS One,8,e59425.
Verboom,G.A.,Bergh,N.G.,Haiden,S.A.,Hoffmann,V.,&Britton,M.N.(2015).TopographyasadriverofdiversificationintheCapeFloristicRegionofSouthAfrica.New Phytologist,207,368–376.
Vila,M., Pino, J.,Montero,A., & Font, X. (2010).Are island plant com-munitiesmore invaded than theirmainland counterparts? Journal of Vegetation Science,21,438–446.
Whittaker, R. J., Ladle, R. J., Araújo, M. B., Fernández-Palacios, J. M.,Delgado,D.J., et al. (2007).The island immaturity—Speciationpulsemodelofislandevolution:Analternativetothe“diversitybegetsdiver-sity”model.Ecography,30,321–327.
Zobel, M., Otto, R., Laanisto, L., Naranjo-Cigala, A., Paertel, M., et al.(2011).Theformationofspeciespools:Historicalhabitatabundanceaffects current local diversity. Global Ecology and Biogeography, 20,251–259.
SUPPORTING INFORMATION
AdditionalSupportingInformationmaybefoundonlineinthesupport-inginformationtabforthisarticle.
How to cite this article:Steinbauer,M.J.,Irl,S.D.H.,González-Mancebo,J.M.,Breiner,F.T.,Hernández-Hernández,R.,Hopfenmüller,S.,Kidane,Y.,Jentsch,A.andBeierkuhnlein,C.(2017),PlantinvasionandspeciationalongelevationalgradientsontheoceanicislandLaPalma,CanaryIslands.EcologyandEvolution,7:771–779.doi:10.1002/ece3.2640








![Thermophysicalpropertiesofdryandhumidair ... · range of temperature and pressure. Data on gW of compressed humid air [7], humid nitrogen, humid argon, and humid carbon dioxide [9]](https://img.pdfslide.net/doc/110x75/5e626081cfea87225a37645c/thermophysicalpropertiesofdryandhumidair-range-of-temperature-and-pressure.jpg)