Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 93, pp. 8139-8144, July 1996Plant Biology
Preferential expression of an ammonium transporter and of twoputative nitrate transporters in root hairs oftomato
(nitrogen nutrition/permeases/gene regulation)
FRANK-ROMAN LAUTER*, OLAF NINNEMANNt, MARCEL BUCHER, JORG W. RIESMEIER, AND WOLF B. FROMMER4§Institut fur Genbiologische Forschung, Ihnestrasse 63, D-14195 Berlin, Germany
Communicated by Emanuel Epstein, University of California, Davis, CA, March 21, 1996 (received for review December 21, 1995)
ABSTRACT Root hairs as specialized epidermal cellsrepresent part of the outermost interface between a plant andits soil environment. They make up to 70%o of the root surfaceand, therefore, are likely to contribute significantly to nutri-ent uptake. To study uptake systems for mineral nitrogen,three genes homologous toArabidopsis nitrate and ammoniumtransporters (AtNrtl and AtAmtl) were isolated from a roothair-specific tomato cDNA library. Accumulation ofLeNrtl-l,LeNrtl-2, and LeAmtl transcripts was root-specific, with nodetectable transcripts in stems or leaves. Expression was rootcell type-specific and regulated by nitrogen availability.LeNrtl-2 mRNA accumulation was restricted to root hairsthat had been exposed to nitrate. In contrast, LeNrtl-1transcripts were detected in root hairs as well as other roottissues under all nitrogen treatments applied. Analogous toLeNrtl-1, the gene LeAmtl was expressed under all nitrogenconditions tested, and root hair-specific mRNA accumulationwas highest following exposure to ammonium. Expression ofLeAMTI in an ammonium uptake-deficient yeast strain re-stored growth on low ammonium medium, confirming itsinvolvement in ammonium transport. Root hair specificityand characteristics of substrate regulation suggest an impor-tant role of the three genes in uptake of mineral nitrogen.
Nitrogen availability is a major limiting factor in plant growthand, therefore, crop production. In the soil solution, nitrate(NO-) and ammonium (NH'), represent the most readilyavailable forms of nitrogen for root absorption. NO- is highlymobile in soils, whereas the concentration of NH' tends to bebuffered by interactions with negatively charged soil particles.NO is considered to be the preferred nitrogen source in manysoil types. However, assimilation of NOR, conversion of NOjto NO- and then to NH+, are among the most energy-intensiveprocesses in plants (1, 2). In addition, concurrent absorption ofNH+ and NO- maintains the cation-anion balance withinboth the rhizosphere and the plant (3). Therefore, mixed NH4and NOj nutrition is likely to be preferred by most plants tosatisfy their nitrogen demands.The principal evidence for the existence of specific transport
systems for NO- and NH+ in plant roots derived from uptakestudies. Multiphasic complex kinetics led to the proposal ofseveral transport systems for NO- and NH+ net uptake fromthe soil (3-5). At least three different transport systems forNO- could be distinguished. Two of these display saturablekinetics, one being a constitutive low capacity system and theother a NO--inducible system with higher transport capacity(6-8). In addition, a third, nonsaturable high-capacity systembecomes apparent at higher NO3 concentrations (9). For NH+uptake, saturable and nonsaturable systems operating at lowand high external NH+ concentrations, respectively, have beencharacterized (10-12). It should be noted that results fromkinetic studies do not allow us to conclude whether each
kinetic component is mediated by single transporters or resultsfrom activities of multiple transport proteins. In addition,interpretation of data on uptake kinetics is complicated byseveral other aspects. In many studies, only net uptake wasmeasured. Net uptake of mineral nitrogen reflects the sum ofinflux, efflux, and assimilation. The relative rates of NH' andNO- absorption vary with changes in environmental condi-tions such as the relative availabilities of these ions, pH (13,14), and temperature (15, 16). In addition, uptake is influencedby changes in the nitrogen content or composition (17) andcarbohydrate or energy availability (18) within the root as wellas by the nitrogen demand of the shoot (19). Finally, investi-gations of nutrient uptake have relied primarily on techniquesthat integrate uptake over the entire root system. Standardapproaches fail to reveal spatial differences in transport ac-tivities along the root axis (20, 21), as well as differencesbetween different root cell types. Thus, a dissection of trans-port systems for mineral nitrogen at the molecular level isneeded to understand better the available physiological data.
Recently, molecular techniques have made possible theisolation of a number of plant transporter genes (reviewed inrefs. 22 and 23). The first permeases for mineral nitrogen werecloned from Arabidopsis thaliana. The Arabidopsis mutantchlJ-1, defective in low-affinity nitrate transport, served as abasis for isolating a putative low-affinity NO- transporterAtNrtl (originally designatedAtCHLI; refs. 24 and 25). Actualnitrate transport activity was demonstrated by functionalexpression in Xenopus oocytes (25). The high-affinity ammo-nium transporter AtAmtl was identified and characterized byfunctional expression in a yeast mutant defective in ammoniumuptake (26, 27).The physiology of nitrogen uptake has been studied in
greater detail in tomato than inArabidopsis (3). The possibilityto isolate root hairs in sufficient quantities from tomato madeit possible to construct a root hair-specific cDNA library(M.B., B. Schroeer, L. Willmitzer, and J.W.R., unpublishedwork). Furthermore, tomato, a larger plant that can also beefficiently transformed, seems to be more appropriate for ananalysis of nitrogen transport in transgenic plants using phys-iological methods.AtNrtl and AtAmtl were used in the present study as
heterologous probes to isolate homologues from tomato (Ly-copersicon esculentum). The expression pattern of three nitro-gen transporter homologues was analyzed in different organsof tomato plants that had been exposed to different nitrogenconditions.
Data deposition: The sequences reported in this paper have beendeposited in the GenBank database [accession nos. X92853 (LeNrtl-1), X92852 (LeNrtl-2), and X92854 (LeAmtl)].*Present address: BioInside GmbH, Moosdorf Strasse 7, 12435, Berlin,Germany.
tPresent address: Anatomy Department, Medical Faculty, Charite,Humboldt University Berlin, D-10098 Berlin, Germany.4Present address: Institut fir Botanik, Eberhard-Karls-UniversitatTiubingen, Auf der Morgenstelle 1, D-72076 Tubingen, Germany.§To whom reprint requests should be addressed.
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
8139

Proc. Natl. Acad. Sci. USA 93 (1996)
MATERIALS AND METHODSStrains and Plasmids. The plant material was Lycopersicon
esculentum cv. Moneymaker. Yeast strain 26972c and expres-sion vector pFL61 were used for complementation of ammo-nium transport deficiency (26). Root hair-specific tomatocDNA library was in AUni-ZAP XR (M.B., B. Schroeer, L.Willmitzer, and J.W.R., unpublished work). For isolation ofLeNrtl-l, LeNrtl-2, and LeAmtl clones from the AZAPIIcDNA library, instructions of the manufacturer were followed(Stratagene) using AtNrtl (25) and AtAmtl (27) from Arabi-dopsis as probes. Le25s encoding 25S rRNA was used as acontrol.Hydroponic System. Nutrient solutions have been described
(29). The nitrogen source was varied and the pH of unbufferednutrient solutions was adjusted and maintained at 6.0 ± 0.3.KNO3 and NH4Cl were used as the sole nitrogen sources.Changes in mineral nitrogen source did not affect the pH of thesolution. All plant growth experiments described here weredone in a temperature-, light-, and humidity-controlled green-house. Supplementary light of 200 ,umol quanta PAR m-2.s-'was set at a 14-hr light (25 ± 2°C, 70% relative humidity)/10-hrdark (20 ± 0.5°C, 55% relative humidity) cycle. Seedlings weregerminated and grown on perlite for 6 weeks under conditionsof high nitrogen (2 mM NH4Cl and 5 mM KN03) applying thenutrient solution described (29). Plants were then transferredto hydroponic culture with a high nitrogen supply (2 mMNH4Cl and 5 mM KN03) nutrient solution (29). Nine plantswere grown in tanks holding 15 liters of an aerated nutrientsolution (29) containing 2 mM NH4Cl and 5 mM KN03.Nutrient solutions were replaced after 1 week and every 3 daysthereafter. After 2 weeks, plants were transferred to differentnitrogen treatments (see Fig. 5).Aeroponic System. Nutrient solutions (29) and growth con-
ditions were as described above. Details are given in Fig. 7.Isolation of Root Hairs. Isolation procedures were described
(ref. 30 and M.B., B. Schroeer, L. Willmitzer, and J.W.R.,unpublished work). In short, roots that had developed in anaeroponic system were frozen in liquid nitrogen. Frozen rootswere stirred carefully with a glass rod. Through the shearingforces applied, root hairs broke off and could be separated byfiltration through stainless steel filters with a cut-off size of 250,mm (Fig. 3).
Nucleic Acid Analysis. Total RNA was extracted fromvarious plant organs and Northern blot analyses were per-formed by- standard procedures (27, 31). RNA from eachsample was separated by electrophoresis through a 1.5%agarose/formaldehyde gel and transferred to Hybond-N (Am-ersham). RNA was probed with radiolabeled LeNrtl-l,LeNrtl-2, and LeAmtl probes. Radiolabeled DNA probeswere specific for the 3' untranslated region of the three genesand probes were prepared by hexamer-primed labeling. Allexperiments were repeated two to four times independentlywith comparable results. For DNA sequence analyses, cDNAswere excised with EcoRI, subcloned into pBluescript SK-(Stratagene) and both strands were sequenced using syntheticoligonucleotides.
RESULTSIsolation and Characterization of Transporters for Mineral
Nitrogen. Using the Arabidopsis clones AtNrtl and AtAmtl asheterologous probes, LeNrtl-l, LeNrtl-2, and LeAmtl couldbe isolated from a root hair-specific cDNA library preparedfrom 5-day-old tomato seedlings (27). The amino acid se-quence ofLeNrtl-l was 88% identical to LeNrtl-2, 71% to theArabidopsisAtNrtl (Fig. 1A), and 65% to BnNrtl from Brassicanapus (I. Muldin and B. Ingemarsson, unpublished data;GenBank accession no. U17987). In addition, LeNrts displayedsimilarities to oligopeptide transporters from Arabidopsis.
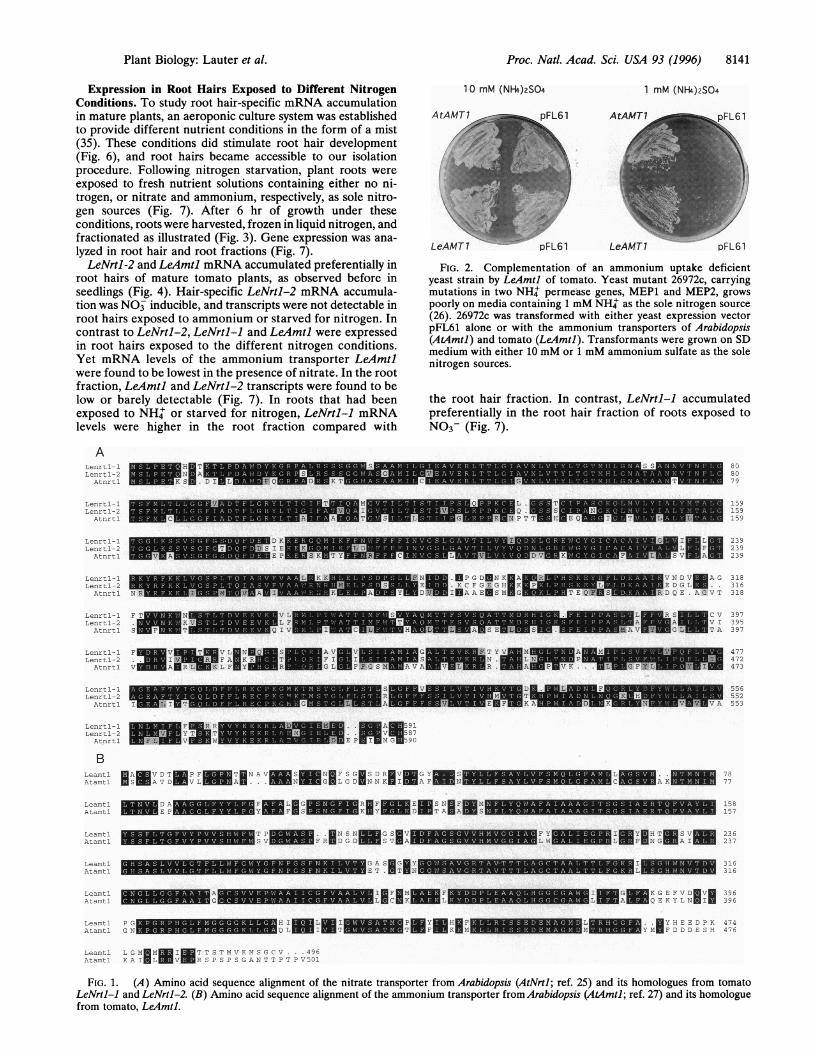
Amino acid sequence identity of LeNrtl-l was 22% withAtPTR2 (32) and 36% with AtNTRJ (33) (data not shown).LeAmtl was 78% identical to the Arabidopsis ammoniumtransporter AtAmtl (Fig. IB). Analysis of the distribution ofhydrophobic amino acid residues indicated that all 3 tomatogenes encode integral membrane proteins with 10-12 mem-brane spanning domains (data not shown, see also refs. 25 and27). Due to the high degree of homology to the Arabidopsistransporters that have been functionally characterized in het-erologous systems, tomato LeNrts and LeArmtl are likely toencode nitrate and ammonium transporters, respectively (25,27). To verify its function, LeAmtl was expressed in a yeastmutant deficient for ammonium uptake. LeAmtl was able tofunctionally complement the ammonium uptake deficiency ofthe yeast mutant (Fig. 2), indicating that it represents anortholog of AtAmtl.
Expression in Seedling Root Hairs. To analyze the expres-sion pattern of LeNrts and LeAmtl in root hairs, 5-day-oldseedlings were frozen in liquid nitrogen, and roots wereisolated. Subsequently, the roots were carefully stirred in liquidnitrogen and root hairs were separated by filtration. Micro-scopic analysis of the resulting two root fractions showed thatthe root hair fraction consisted of pure root hairs (Fig. 3A andB), whereas the stripped root fraction contained a mixture ofroot hair-free fragments (Fig. 3C), fragments carrying thebasal part of broken- off root hairs (Fig. 3D), and fragmentsstill carrying intact root hairs (Fig. 3 E and F). Therefore, pureroot hair fractions were compared with non-root hair fractionsstill contaminated with root hairs. Henceforth, these twofractions will be called root hair and root fraction. RNA wasisolated from both fractions and subjected to Northern blotanalysis (Fig. 4).LeNrtl-2 and LeAmtl mRNAs accumulated preferentially
in root hairs of seedlings. In addition, low levels of LeNrtl-2and very low levels of LeArmtl transcripts were also detectedin the root fraction. LeNrtl-l transcripts accumulated prefer-entially in the root fraction, although significant levels werefound in root hairs as well. These results indicate that expres-sion of LeNrtl-2 and LeAmtl was predominant in root hairsand that different members of the LeNrt gene family werepreferentially expressed in different cell types of the tomatoroot (Fig. 4).
Expression in Plants Exposed to Different Nitrogen Con-ditions. To study organ-specificity and substrate-inducibility oftransporter genes in mature plants, RNA was isolated fromdifferent organs of 2-months-old tomato plants exposed todifferent nitrogen sources. Following nitrogen starvation, hy-droponically grown plants were transferred to fresh nutrientsolutions containing either no nitrogen, 200 ,uM NO-, or 100,uM NH' as sole nitrogen sources (Fig. 5). After 1, 3, 6, or 18hr of growth under different nitrogen conditions, roots, stems,and leaves were isolated and mRNA accumulation of LeNrtsand LeAmtl was analyzed (Fig. 5).
Expression of all three genes was root-specific, with nodetectable transcripts in stems or leaves (Fig. 5). In roots,LeNrtl-l and LeAmtl mRNAs accumulated to high levelsunder all nitrogen treatments. In contrast, expression ofLeNrtl-2 was substrate-inducible. Following 1 hr of exposureto NO-, LeNrtl-2 mRNA accumulated to high levels, re-mained high during the following 5 hr, but decreased toundetectable levels after 18 hr of exposure to NO- (Fig. 5).LeNrtl-l transcript accumulation was only slightly stimulatedin the presence of NO-. The high levels of LeNrtl-l andLeNrtl-2 mRNA observed in roots exposed to NHW for 6 hr(Fig. 5) were not reproducible in three independent experi-ments (data not shown). It should be noted that even in fairlyclean hydroponic systems some nitrification of NH' to NO-can occur that might be sufficient to induce NO- absorptionand reduction (34).
8140 Plant Biology: Lauter et al.
Proc. Natl. Acad. Sci. USA 93 (1996) 8141
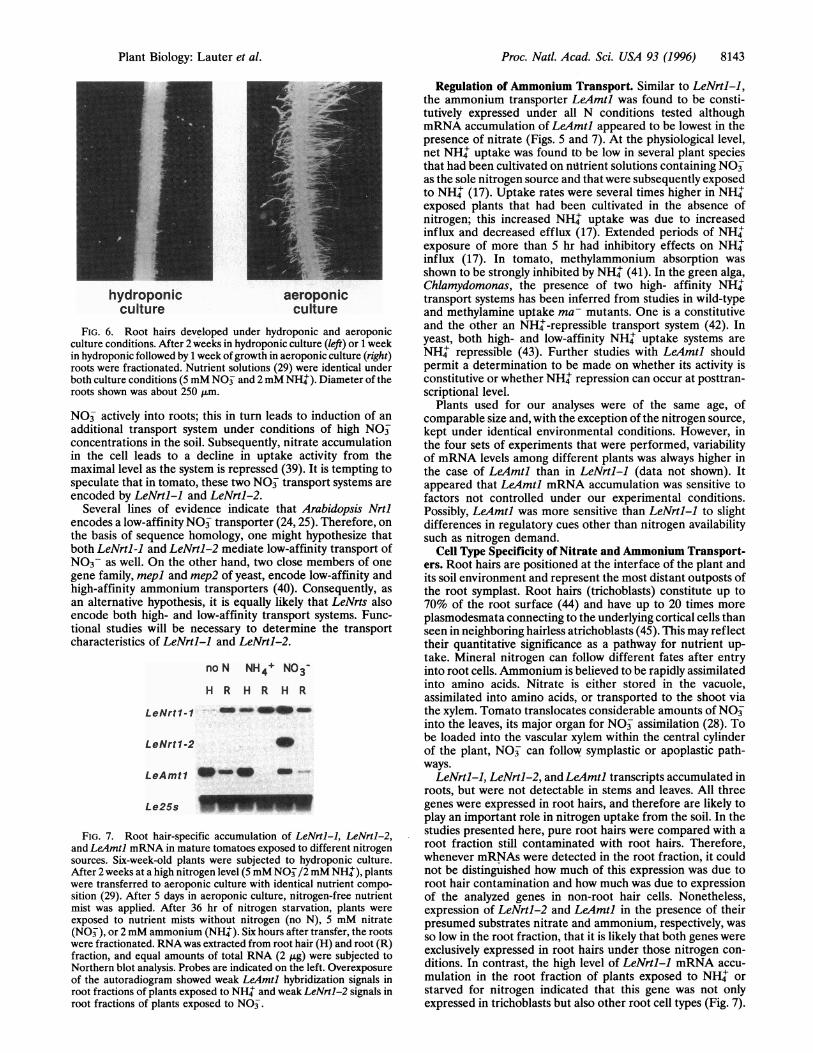
Expression in Root Hairs Exposed to Different NitrogenConditions. To study root hair-specific mRNA accumulationin mature plants, an aeroponic culture system was establishedto provide different nutrient conditions in the form of a mist(35). These conditions did stimulate root hair development(Fig. 6), and root hairs became accessible to our isolationprocedure. Following nitrogen starvation, plant roots wereexposed to fresh nutrient solutions containing either no ni-trogen, or nitrate and ammonium, respectively, as sole nitro-gen sources (Fig. 7). After 6 hr of growth under theseconditions, roots were harvested, frozen in liquid nitrogen, andfractionated as illustrated (Fig. 3). Gene expression was ana-lyzed in root hair and root fractions (Fig. 7).LeNrtl-2 and LeAmtl mRNA accumulated preferentially in
root hairs of mature tomato plants, as observed before inseedlings (Fig. 4). Hair-specific LeNrtl-2 mRNA accumula-tion was NO3 inducible, and transcripts were not detectable inroot hairs exposed to ammonium or starved for nitrogen. Incontrast to LeNrtl-2, LeNrtl-l and LeAmtl were expressedin root hairs exposed to the different nitrogen conditions.Yet mRNA levels of the ammonium transporter LeAmtlwere found to be lowest in the presence of nitrate. In the rootfraction, LeAmtl and LeNrtl-2 transcripts were found to below or barely detectable (Fig. 7). In roots that had beenexposed to NH' or starved for nitrogen, LeNrtl-l mRNAlevels were higher in the root fraction compared with
10 mM (NH4)2S04
AtAMTI _ pFL61
.40~#
LeAMT1 pFL61
1 mM (Nb4)2SO4
LeAMTI PFL61
FIG. 2. Complementation of an ammonium uptake deficientyeast strain by LeAmtl of tomato. Yeast mutant 26972c, carryingmutations in two NH+ permease genes, MEP1 and MEP2, growspoorly on media containing 1 mM NH+ as the sole nitrogen source(26). 26972c was transformed with either yeast expression vectorpFL61 alone or with the ammonium transporters of Arabidopsis(AtAmtl) and tomato (LeAmtl]). Transformants were grown on SDmedium with either 10 mM or 1 mM ammonium sulfate as the solenitrogen sources.
the root hair fraction. In contrast, LeNrtl-l accumulatedpreferentially in the root hair fraction of roots exposed toNO3- (Fig. 7).
8A
79
K_H D
.1A .I V
asW
_139159159
Ler.rtl-l D K G 1 2_ 239Lerrti-2 I E3 L_F 9
A-Kr T C L V PV K F L SVIA 239
Lenr-1 _j_N .EPGDENKMA K VNDVA C 318enrtL-2 , KCS__E__a H K E G L .. 3A:nrt NRIMVEAE A EASM*G TFQ7 DQE . AGST 318
_enr.r-1 F N -N S9 A
A:rr-:1> SK F T m
.An : ;
_ iGA:nr. l V T I VL C F G AA V LAG A *a.
Ler-r,.- F^ - T VL N S AV V V G _G T Y V _ M_ A477Lenrt:- 2 I VI R F AKT F I I S _ N L LP_ 472z
A--nrti V_ A-R L_KLF Y__R G L_F E! G S M_A V A _ I R .|A_ - K... _G F YL _ I 47 3
WAA.L F 556i n h r V
>5521 MICENH ~~~~~~~~5532
AA_591_ 587
:PI* M 9 C
B_eC-- 3| JA_ VD 3T1- dl9>.'It _ S_ - I.
z C S D*< 1 ' G C.-a.-~~~~~~ N_-G-'- _ G L 5 D|9 N, wrK I ft
- |-mJ _~~~~~~C AKLe=:__- CA F LpIG * * *
IYI7flR T_ 7;_ 2anL LFEG A:
316316
IjF_GEK G E F V DIDV 396|A__ E. YI LN )
L e~a: t L G M-I M'M_ I T 7 S !*V K Xf c G C ~ 4 9 F.
'--amt: K :. -1 ?- G ZD T p %'; :
FIG. 1. (A) Amino acid sequence alignment of the nitrate transporter from Arabidopsis (AtNrtl; ref. 25) and its homologues from tomatoLeNrtl-l and LeNrtl-2. (B) Amino acid sequence alignment of the ammonium transporter from Arabidopsis (AtAmtl; ref. 27) and its homologuefrom tomato, LeAmtl.
AL,enrE- i
Lenrr-1-1Lenrt1-2
A-:nrtl
Len.rtl-1Lenrtl-2
Atnrt 1
Lenrti- -:-enrtl-2
Atnrt'L
T P N S IN, G SIT."S _,,~~ F I D 'G D _~S -- A
Lear.t 1A.aritI
- eamtlI P G-art: CG N
IE F M FLU C K _ L
iv m LI QT p
i
R TD.L I I .. A K
... F RyTIEMF%Mvw
G k SIOIYE T T N I
Plant Biology: Lauter et al.
IIi
l_l1mM quL~I _. '1'un wX~ -I
_-ea-7~l't.L,- a-nt 1
A- Y 'z w- E D '--D K 4 "I 4.1 .-- D D, D ---:' S H 4' 7 E

Proc. Natl. Acad. Sci. USA 93 (1996)
. -4
0~0
LeNrtl-l _
LeNrtl-2
LeAmtl
Le25s ~FIG. 4. Root hair expression of LeNrtl-1, LeNrtl-2, and LeAmtl
mRNA in tomato seedlings. Seedlings were germinated and grown onfilter paper saturated with 0.5 x Hoagland medium with high nitrogenlevels (3 mM NO-/1 mM NHW) for 5 days. Root hair and root
F fractions were prepared (as shown in Fig. 3) and total RNA wasextracted. RNA (8 Zg per lane) was subjected to Northern blotanalysis. Membranes were probed with 32P-labeled LeNrt1-1,LeNrtl-2, and LeArmtl probes. The membrane used for LeArmtl wasstripped and rehybridized with Le25s as control. Probes are indicatedon the left. Overexposure of the autoradiogram showed a weakLeAmtl-specific hydridization signal in the root fraction.
FIG. 3. Light microscopy of root hair (A and B) and root fractions(C-F) of a 5-day-old tomato seedling (shown on the top lefthand side).Seedling roots were isolated and fractionated and analyzed with aZeiss Axiophot microscope. (Bars = 100 ,im.)
DISCUSSIONUptake of mineral nitrogen by higher plants has been analyzedin detail at the physiological level (3-5). The available dataindicate that entry of NH' and NO- into roots apparently ismediated by more than one transporter specific for each ion(11, 36). To dissect nitrogen transport systems at the molecularlevel, genes encoding putative transporters for mineral nitro-gen were cloned. Isolation ofLeNrtl-l, LeNrtl-2, and LeAmtlwill make it possible to analyze regulation and function ofindividual transport components. The focus of this analysis wasto study the regulation of transporter expression with respectto substrate-inducibility and cell type specificity.
Regulation of Nitrate Transport. The fact that two membersof the Nrtl gene family were isolated from tomato may supportevidence from earlier physiological studies that more than onetransporter is involved in NO- uptake. The response of twoclose members of the Nrtl family, LeNrtl-l and LeNrtl-2, tosubstrate exposure was found to be different. LeNrtl-l wasexpressed constitutively, in the presence or absence of NH4and NO-, whereas LeNrtl-2 expression was transiently NOR-inducible (Figs. 5 and 7). Nitrate inducibility of LeNrtl-2 isshared by its Arabidopsis homologue (25). LeNrtl-2 mRNAlevels were high during the first 6 hr of NO1 exposure but notdetectable after 18 hr (Fig. 5). Thus, the expression pattern ofLeNrtl-2 had features in common with the regulation of NO3uptake activity in plants. Nitrate uptake is subject to bothpositive and negative feedback regulation (37, 38). Physiolog-ical studies have identified a low capacity constitutive and ahigh capacity inducible NO- uptake system in barley (7-9).The constitutive system appears to be necessary to transport
Z I* + z.z +I no N NO3- NH4+ z +
0 0 o 0 X r-iF -~I- --1 l0e° °z2 z z r-- O'''1l0 0 0 - Io 0 mZ ZZ Z 0~C C CoC C Z z
0 000 000 U0 - U 0 W U U 0 00 .0 0 . a- a. a- Co 0 0 0 o0w -~~A0 o
m m
*[email protected] SeeSSeaSe esab
1 hour 3 hours 6 hours 18 hours
FIG. 5. Substrate-inducibility and organ-specificity of LeNrtl-J, LeNrtl-2, and LeAmtl mRNA accumulation. Six-week-old tomato plants weresubjected to hydroponic culture. After 2 weeks in nutrient solution (29) with a high nitrogen level (5 mM NO- and 2 mM NH'), plants weretransferred to fresh, nitrogen-free nutrient solution. After 36 hr of nitrogen starvation, plants were transferred to fresh nutrient solutions containingeither no nitrogen (no N), or 200 ,um NO-, or 100 ,um NHWt. Plants were harvested and roots were fractionated 1, 3, 6, and 18 hr after transfer.From plants exposed to various nitrogen conditions for 6 hr, stems as well as source [leaf (so)] and sink [leaf (si)] leaves were harvested in additionto roots. Equal amounts of total RNA (8 ,ug) were subjected to Northern blot analysis. Le25s was used as control. Probes are indicated on the left.
C
A
..
x
i.girb
LeNrtl-1
LeNrtl-2
Le 25s
LeAmtl
8142 Plant Biology: Lauter et al.
sam,owmaI 11
Proc. Natl. Acad. Sci. USA 93 (1996) 8143
hydroponicculture
aeroponicculture
FIG. 6. Root hairs developed under hydroponic and aeroponicculture conditions. After 2 weeks in hydroponic culture (left) or 1 weekin hydroponic followed by 1 week ofgrowth in aeroponic culture (right)roots were fractionated. Nutrient solutions (29) were identical underboth culture conditions (5 mM NO and 2 mM NH'). Diameter of theroots shown was about 250 ,um.
NO- actively into roots; this in turn leads to induction of anadditional transport system under conditions of high NO3concentrations in the soil. Subsequently, nitrate accumulationin the cell leads to a decline in uptake activity from themaximal level as the system is repressed (39). It is tempting tospeculate that in tomato, these two NO3 transport systems areencoded by LeNrtl-l and LeNrtl-2.
Several lines of evidence indicate that Arabidopsis Nrtlencodes a low-affinity NO- transporter (24, 25). Therefore, onthe basis of sequence homology, one might hypothesize thatboth LeNrtl-l and LeNrtl-2 mediate low-affinity transport ofNO3- as well. On the other hand, two close members of onegene family, mepl and mep2 of yeast, encode low-affinity andhigh-affinity ammonium transporters (40). Consequently, asan alternative hypothesis, it is equally likely that LeNrts alsoencode both high- and low-affinity transport systems. Func-tional studies will be necessary to determine the transportcharacteristics of LeNrtl-l and LeNrtl-2.
noN NH4+ N03
H R H R H R
LeNrtl-l - - 0
LeNrtl-2
LeAmtl
Le25s
FIG. 7. Root hair-specific accumulation of LeNrtl-l, LeNrtl-2,and LeAmtl mRNA in mature tomatoes exposed to different nitrogensources. Six-week-old plants were subjected to hydroponic culture.After 2 weeks at a high nitrogen level (5 mM NOy/2mM NHW), plantswere transferred to aeroponic culture with identical nutrient compo-sition (29). After 5 days in aeroponic culture, nitrogen-free nutrientmist was applied. After 36 hr of nitrogen starvation, plants wereexposed to nutrient mists without nitrogen (no N), 5 mM nitrate(NO-), or 2mM ammonium (NWt). Six hours after transfer, the rootswere fractionated. RNA was extracted from root hair (H) and root (R)fraction, and equal amounts of total RNA (2 ,ug) were subjected toNorthern blot analysis. Probes are indicated on the left. Overexposureof the autoradiogram showed weak LeAmtl hybridization signals inroot fractions of plants exposed to NWt and weak LeNrtl-2 signals inroot fractions of plants exposed to NO-.
Regulation of Ammonium Transport. Similar to LeNrtl-l,the ammonium transporter LeAmtl was found to be consti-tutively expressed under all N conditions tested althoughmRNA accumulation of LeAmtl appeared to be lowest in thepresence of nitrate (Figs. 5 and 7). At the physiological level,net NH' uptake was found to be low in several plant speciesthat had been cultivated on nutrient solutions containing NO3as the sole nitrogen source and that were subsequently exposedto NH' (17). Uptake rates were several times higher in NH'exposed plants that had been cultivated in the absence ofnitrogen; this increased NH' uptake was due to increasedinflux and decreased efflux (17). Extended periods of NH4exposure of more than 5 hr had inhibitory effects on NH4influx (17). In tomato, methylammonium absorption wasshown to be strongly inhibited by NH' (41). In the green alga,Chlamydomonas, the presence of two high- affinity NH4transport systems has been inferred from studies in wild-typeand methylamine uptake ma- mutants. One is a constitutiveand the other an NHt-repressible transport system (42). Inyeast, both high- and low-affinity NH' uptake systems areNH' repressible (43). Further studies with LeAmtl shouldpermit a determination to be made on whether its activity isconstitutive or whether NHW repression can occur at posttran-scriptional level.
Plants used for our analyses were of the same age, ofcomparable size and, with the exception of the nitrogen source,kept under identical environmental conditions. However, inthe four sets of experiments that were performed, variabilityof mRNA levels among different plants was always higher inthe case of LeAmtl than in LeNrtl-l (data not shown). Itappeared that LeAmtl mRNA accumulation was sensitive tofactors not controlled under our experimental conditions.Possibly, LeAmtl was more sensitive than LeNrtl-l to slightdifferences in regulatory cues other than nitrogen availabilitysuch as nitrogen demand.
Cell Type Specificity of Nitrate and Ammonium Transport-ers. Root hairs are positioned at the interface of the plant andits soil environment and represent the most distant outposts ofthe root symplast. Root hairs (trichoblasts) constitute up to70% of the root surface (44) and have up to 20 times moreplasmodesmata connecting to the underlying cortical cells thanseen in neighboring hairless atrichoblasts (45). This may reflecttheir quantitative significance as a pathway for nutrient up-take. Mineral nitrogen can follow different fates after entryinto root cells. Ammonium is believed to be rapidly assimilatedinto amino acids. Nitrate is either stored in the vacuole,assimilated into amino acids, or transported to the shoot viathe xylem. Tomato translocates considerable amounts of NO3into the leaves, its major organ for NO3 assimilation (28). Tobe loaded into the vascular xylem within the central cylinderof the plant, NO3 can follow symplastic or apoplastic path-ways.
LeNrtl-l, LeNrtl-2, and LeAmtl transcripts accumulated inroots, but were not detectable in stems and leaves. All threegenes were expressed in root hairs, and therefore are likely toplay an important role in nitrogen uptake from the soil. In thestudies presented here, pure root hairs were compared with aroot fraction still contaminated with root hairs. Therefore,whenever mRNAs were detected in the root fraction, it couldnot be distinguished how much of this expression was due toroot hair contamination and how much was due to expressionof the analyzed genes in non-root hair cells. Nonetheless,expression of LeNrtl-2 and LeAmtl in the presence of theirpresumed substrates nitrate and ammonium, respectively, wasso low in the root fraction, that it is likely that both genes wereexclusively expressed in root hairs under those nitrogen con-ditions. In contrast, the high level of LeNrtl-l mRNA accu-mulation in the root fraction of plants exposed to NWt orstarved for nitrogen indicated that this gene was not onlyexpressed in trichoblasts but also other root cell types (Fig. 7).
Plant Biology: Lauter et al.

Proc. Natl. Acad. Sci. USA 93 (1996)
Those cell types could be atrichoblasts on the surface of theroot. Alternatively, non-root hair cells could be cells inside theroot, for example cortex cells. Finally it should be noted, thatwe do not know whether all sections of the root were equallysusceptible to the root hair stripping technique employed here.Thus, the root hairs analyzed may represent only a subpopu-lation. To identify the cellular localization ofLeNrtl-2 mRNA,in situ hybridization technique has as of yet been appliedunsuccessfully. However, efforts are continuing. In addition,immunolocalization experiments are planned to localize thepolypeptides encoded by LeAmt], LeNrtl-1, and LeNrtl-2.
In summary, three genes that appear to be involved in rootmineral nitrogen absorption were isolated and characterizedwith respect to their expression in roots exposed to differentnitrogen regimes. Results from this study do not exclude thepossibility that additional transporters involved in the uptakeof mineral nitrogen are present in root hairs and other root celltypes. A more detailed molecular analysis will make possiblea better understanding of nitrogen uptake at the cellular andsubcellular level.
We are grateful to R. Breitfeld for competent assistance withgreenhouse work and B. Schroeer for valuable suggestions with respectto root hair manipulations. We thank K. Bienert and H. Spontjes forconstructing the aeroponic growth chambers, K. Breitkreuz for hiscomments, and N. Provart for providing the 25s rRNA clone. The yeastmutant 26972c from B. Andre (Brussels) and the AtNRT cDNA fromN. Crawford (University of California, San Diego) were essential forthe project. The project was supported by the European CommunitiesBiotech Programme as part of Technological Priority 1993-1996 andby KWS, Einbeck, Germany.
1. Chapin, F. S., III, Bloom, A. J., Field, C. B. & Waring, R. H.(1987) BioScience 37, 49-57.
2. Bloom, A. J., Sukurapanna, S. & Warner, R. L. (1992) PlantPhysiol. 99, 1294-1301.
3. Bloom, A. J. (1994) in Physiology and Determination ofCrop Yield,eds. Boote, K. J. & Sinclair, T. (ASA/CSA, Madison, WI), pp.303-310.
4. Glass, A. D. M. & Siddiqi, M. Y. (1995) in Nitrogen Nutrition inHigher Plants, eds. Srivastava, H. S. & Singh, R. P. (AssociatedPublishing, New Delhi, India), pp. 21-56.
5. Lauter, F.-R., Ninnemann, 0. & Frommer, W. B. (1995) inCarbon Partitioning and Source-Sink Interaction in Plants, eds.Madore, M. & Lucas, W. (Am. Soc. Plant Physiol., Rockville,MD), pp. 102-116.
6. Lee, R. B. & Drew, M. C. (1986) J. Exp. Bot. 37, 1753-1779.7. Behl, R., Tischner, R. & Raschke, K. (1988) Planta 176, 235-240.8. Aslam, M., Travis, R. L. & Huffaker, R. C. (1992) Plant Physiol.
99, 1124-1133.9. Siddiqi, M. Y., Glass, A. D. M., Ruth, T. J. & Rufty, T. W. (1990)
Plant Physiol. 93, 1426-1432.10. Ullrich, W. R., Larsson, M., Larsson, C.-M., Lesch, S. & Novacky,
A. (1984) Physiol. Plant. 61, 369-376.11. Smart, D. R. & Bloom, A. J. (1988) Oecologia 76, 336-340.12. Wang, M. Y., Siddiqi, M. Y., Ruth, T. J. & Glass, A. D. M. (1993)
Plant Physiol. 103, 1259-1267.
13. Raper, C. D., Jr., Vessey, J. K., Henry, L. T. & Chaillou, S. (1991)Plant Physiol. Biochem. 29, 205-212.
14. Vessey, J. K., York, E. K., Henry, L. T. & Raper, C. D., Jr. (1988)Biotronics 17, 79-94.
15. Bloom, A. J. & Chapin, F. S., III (1981) Plant Physiol. 68,1064-1067.
16. Macduff, J. H. & Wild, A. (1989) J. Exp. Bot. 40, 195-206.17. Morgan, M. A. & Jackson, W. A. (1988) J. Exp. Bot. 39, 179-191.18. Champigny, M. L. & Tolouizte, A. (1986) in Fundamental, Eco-
logical and Agricultural Aspects of Nitrogen Metabolism in HigherPlants, eds. Lambers, H., Neeteson, J. J. & Stulen, I. (Nijhoff,Dordrecht, The Netherlands), pp. 3-27.
19. Imsande, J. & Touraine, B. (1994) Plant Physiol. 105, 3-7.20. Henriksen, G. H., Raman, D. R., Walker, L. P. & Spanswick,
R. M. (1992) Plant Physiol. 99, 734-747.21. Lazof, D. B., Rufty, T. W. & Redingbaugh, M. G. (1992) Plant
Physiol. 100, 1251-1258.22. Crawford, N. M. (1995) Plant Cell 7, 859-868.23. Ninnemann, 0. & Frommer, W. B. (1995) Annu. Rev. Plant
Physiol. Plant Mol. Biol. 46, 419-444.24. Doddema, H. & Telkamp, G. P. (1979) Physiol. Plant. 45, 332-
338.25. Tsay, Y.-F., Schroeder, J. I., Feldmann, K. A. & Crawford, N. M.
(1993) Cell 72, 705-713.26. Marini, A. M., Vissers, S., Urrestarazu, A. & Andre, B. (1994)
EMBO J. 13, 3456-3463.27. Ninnemann, O., Jauniaux, J.-C. & Frommer, W. B. (1994)EMBO
J. 13, 3464-3471.28. Andrews, M. (1986) Plant Cell Environ. 9, 511-519.29. Smart, D. R. & Bloom, A. J. (1993) Plant Cell Environ. 16,
259-267.30. Rohm, M. & Werner, D. (1987) Physiol. Plant. 69, 129-136.31. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular
Cloning: A Laboratory Manual (Cold Spring Harbor Lab. Press,Plainview, NY), 2nd Ed.
32. Steiner, H.-Y., Song, W., Zhang, L., Naider, F., Becker, J. M. &Stacey. G. (1994) Plant Cell 6, 1289-1299.
33. Rentsch, D., Laloi, M., Rouhara, I., Schmelzer, E., Delrot, S. &Frommer, W. B. (1995) FEBS Lett. 370, 264-268.
34. Padgett, P. E. & Leonard, R. T. (1993) Plant Physiol. 101, 141-146.
35. Zobel, R. W., Tredici, P. D. & Torrey, J. G. (1976) Plant Physiol.57, 344-346.
36. Goyal, S. & Huffaker, R. C. (1986) Plant Physiol. 82, 1051-1056.37. Larsson, C.-M. & Ingemarsson, B. (1989) in MolecularAspects of
Nitrate Uptake in Higher Plants, eds. Wray, J. & Kinghorn, J.(Oxford Science, Oxford), pp. 3-14.
38. Redingbaugh, M. G. & Campbell, W. H. (1991) Physiol. Plant. 82,640-650.
39. Siddiqi, M. Y., Glass, A. D. M., Ruth, T. J. & Fernando, M.(1989) Plant Physiol. 90, 806-813.
40. Marini, A.-M., Vissers, S. & Andre, B. (1994) Arch. Int. Physiol.Biochim. 102, 86.
41. Kosola, K. R. & Bloom, A. J. (1994) Plant Physiol. 105, 435-442.42. Franco, A. R., Cardenas, J. & Fernandez, E. (1988)J. Biol. Chem.
263, 14039-14043.43. Dubois, E. & Grenson, M. (1979) Mol. Gen. Genet. 175, 67-76.44. Itoh, S. & Barber, S. A. (1983) Agron. J. 75, 457-461.45. Vakhmistrov, D. B. & Kurkova, E. B. (1979) Sov. Plant Physiol.
26, 763-771.
8144 Plant Biology: Lauter et al.





























