Embed Size (px)
Citation preview

1
Basic Sciences
Principles and Practice Of OphthalmologyEds. D M ALBERT, F A JAKOBIEC
W B SAUNDERS Philadelphia, 1994
Note error in caption to Figure 116-13. The Noell damage should be solid squares; the Ham, damage solid circles;the rat damage, solid triangles and the bovine RPE potential, solid stars.
Chapter 116
Light Effects on the Retina
JOHN MELLERIO
The retina has evolved especially to be sensitive to light.The way it responds to light as part of the visual processis not the concern here-that is for the physiologist and forearlier chapters. The manner in which the retina interactswith light to cause deleterious changes in function is thesubstance of this chapter.
Two problems often stand out for those who work onlight-induced retinal damage. These are the properquantification of the light insult and the detection ofchanged function. Specifying and understanding the exactnature of the stimulus to damage should not be a problemif the physics is considered. The following discussionsattempt to ease the reader through the complexities of thetwo different systems of units that are used, sometimesincorrectly, to specify the stimulus. The way in whichfunctional deficits are detected are not considered here;this can be learned from texts concerned directly withvision. Often, techniques that search for morphologicchanges are substituted for functional tests, as the lesionsare small, and the resolution of behavior studies cannotpick up the small lesions that may be produced in theretinas of animals that are not renowned for their abilityto perform in laboratory studies. Histologic techniques,like those of testing vision, also are not discussed. Rather,consideration is given to stimuli and the effects theyproduce.
QUANTIFYING THE SOURCEThe electromagnetic (EM) energy that is our subject istermed radiation, and its importance lies in its capacity tobring about effects in the world. It has parameters of
intensity and wavelength (or frequency), which, togetherwith certain properties of the substrate. determine whateffects it can produce.
At intensities that might be described as everydayand at which nonlinear effects do not occur, some fractionof the incident radiation must be absorbed in the substratebefore any effect can occur. The extent of the effectdepends on the intensity of the radiation and on thesubstrate's absorption and sensitivity. Consequently, thetask of predicting the size of an effect might start byconsidering the quality and intensity of the radiationacross the part of the spectrum of interest. These dependon the source of the radiation.
The quality of the radiation includes the parametersof wavelength, coherence, geometric distribution, andtemporal characteristics (i.e., whether continuous orpulsed). Table 116-1 shows the way in which the EMspectrum is conventionally divided into regions thatdelimit the spread of wavelengths having similarities inthe way they behave with substrates. Strictly, only visibleradiation may be called light, but often the term is looselyapplied to include ultraviolet (UV) and infrared (IR)radiation. The energy output of a source varies withwavelength and is shown in a spectral power distribution(SPD) plot (i.e., power versus wavelength). Solidincandescent sources have SPDs that usually approximatethat of a black body or planckian radiator, which meansthat peak emission occurs at a wavelength that decreaseswith increasing temperature. Indeed, the radiationcharacteristics can be described by the three radiationlaws of Planck, Stefan-Boltzmann, and Wien. It is the last
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
2
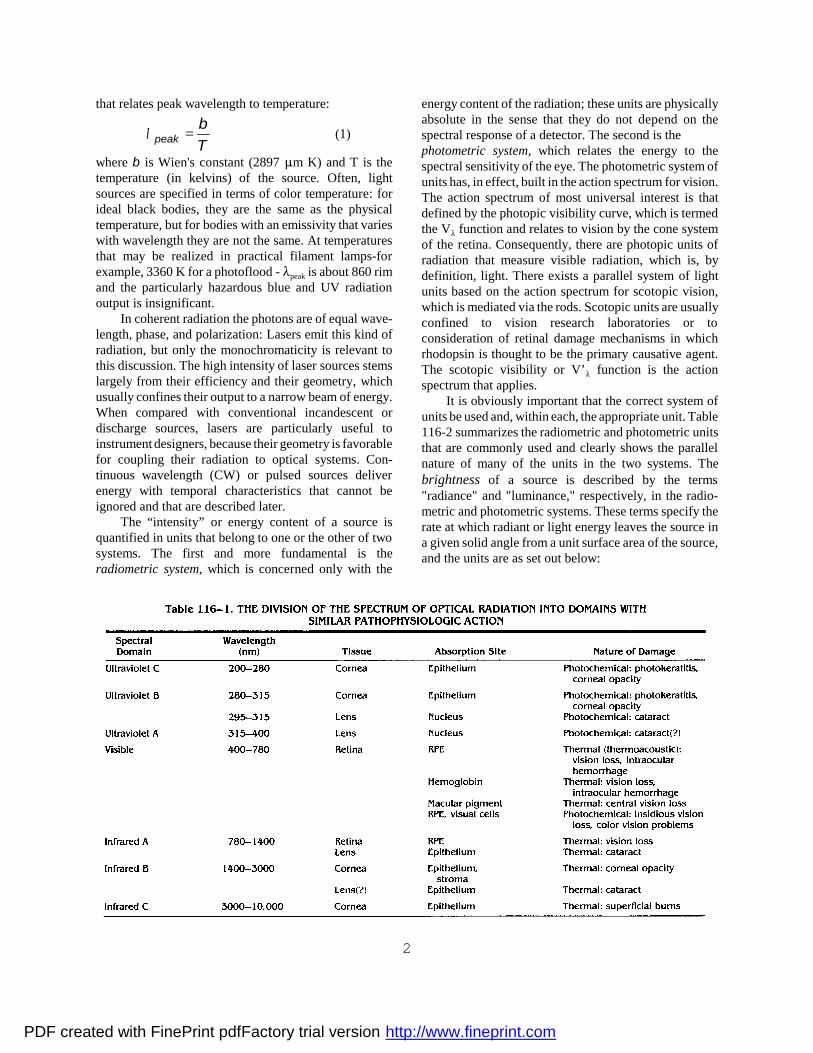
that relates peak wavelength to temperature:
(1)λ peakbT
=
where b is Wien's constant (2897 m K) and T is thetemperature (in kelvins) of the source. Often, lightsources are specified in terms of color temperature: forideal black bodies, they are the same as the physicaltemperature, but for bodies with an emissivity that varieswith wavelength they are not the same. At temperaturesthat may be realized in practical filament lamps-forexample, 3360 K for a photoflood - peak is about 860 rimand the particularly hazardous blue and UV radiationoutput is insignificant.
In coherent radiation the photons are of equal wave-length, phase, and polarization: Lasers emit this kind ofradiation, but only the monochromaticity is relevant tothis discussion. The high intensity of laser sources stemslargely from their efficiency and their geometry, whichusually confines their output to a narrow beam of energy.When compared with conventional incandescent ordischarge sources, lasers are particularly useful toinstrument designers, because their geometry is favorablefor coupling their radiation to optical systems. Con-tinuous wavelength (CW) or pulsed sources deliverenergy with temporal characteristics that cannot beignored and that are described later.
The “intensity” or energy content of a source isquantified in units that belong to one or the other of twosystems. The first and more fundamental is theradiometric system, which is concerned only with the
energy content of the radiation; these units are physicallyabsolute in the sense that they do not depend on thespectral response of a detector. The second is thephotometric system, which relates the energy to thespectral sensitivity of the eye. The photometric system ofunits has, in effect, built in the action spectrum for vision.The action spectrum of most universal interest is thatdefined by the photopic visibility curve, which is termedthe V function and relates to vision by the cone systemof the retina. Consequently, there are photopic units ofradiation that measure visible radiation, which is, bydefinition, light. There exists a parallel system of lightunits based on the action spectrum for scotopic vision,which is mediated via the rods. Scotopic units are usuallyconfined to vision research laboratories or toconsideration of retinal damage mechanisms in whichrhodopsin is thought to be the primary causative agent.The scotopic visibility or V’ function is the actionspectrum that applies.
It is obviously important that the correct system ofunits be used and, within each, the appropriate unit. Table116-2 summarizes the radiometric and photometric unitsthat are commonly used and clearly shows the parallelnature of many of the units in the two systems. Thebrightness of a source is described by the terms"radiance" and "luminance," respectively, in the radio-metric and photometric systems. These terms specify therate at which radiant or light energy leaves the source ina given solid angle from a unit surface area of the source,and the units are as set out below:
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

3
RADIANCEJoule per second per steradian per square meter, whichsimplifies to Watt.sr-1.m-1. The defining equation is
(2)Ld
d dAee=
⋅ ⋅
2ΦΩ cosθ
where e is the radiant flux, the solid angle, and A thesurface area of the source. It also includes the cosine law,which allows for measurement at anangle from the normal to the surface of the sourcebecause the projected unit area along the line ofmeasurement decreases with the cosine of .LUMINANCELumen per steradian per square meter, which becomescandela per square meter (cd.m-1), as a candela is a lumenper steradian. The defining equation is
(3)L dd dAv
v=⋅ ⋅
2ΦΩ cosθ
and is the same as for the radiance except that the fluxterm in the numerator is the luminous flux, whichcontains the all-important luminous efficiency ofradiation, Km in its definition:
(4)ΦΦ
v meK d
dV d= ∫
λλλ
The value of Km is usually taken as 680 lumens per wattand is defined as the maximum number of lumensproduced by 1 watt of broadband radiation extendingfrom the UV to the IR (i.e., over the visible range of thespectrum) in the relation:
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

4
(5)Kmv
e=
ΦΦ
The maximum value is taken for Km as the ratio inequation 5 varies with the spectral distribution of energy(e.g., radiation with a relatively large amount of IRenergy is not visually very effective). Km is the linkbetween radiometric and photometric units.
The rate at which energy arrives at a substrate hascorresponding units in each system.IRRADIANCEEnergy per time per unit area, which is usually measuredin watts per square meter (W.m-2) , as a watt is a joule (J)per second.ILLUMINANCEAlso energy per time per unit area, but the units arelumens per square meter (lm.m-2) , as lumens are definedin terms of e which itself is a time flux of energy.
An important radiometric term is that of radiantenergy density (J.m-1), which defines the amount ofenergy within a certain volume of substrate that isavailable to influence the substrate. Often the term“energy density" is used incorrectly when the radiantexposure (J.m-2) is meant. This latter unit is easier tomeasure or calculate but is not always as useful as manyof those who employ it think. The corresponding pho-tometric unit of luminous energy density (1m.s.m-3) is notoften used.
For completeness the troland (td) must be added tothe units in the table. This is a unit of retinal illuminationdefined as the product of the source luminance (cd.m-1)and the pupillary diameter squared (mm2). The troland isused in studies of retinal photochemistry and variousaspects of visual performance.
Figure 116-1 summarizes some of the concepts thatunderlie the radiometric units described above.
QUANTIFYING THE DOSEThe dose, the quantity of energy deposited in a substrate,depends on three parameters: the "intensity" of theradiation, the substrate's absorbing properties, and theduration of exposure.
Intensity and absorption are dealt with by the processof "convolution." The absorption of the substrate isusually determined at each wavelength, in the region ofthe EM spectrum of interest, to give the
Figure 116-1. Summary diagrams of some concepts ofradiometry: The photometric system has similar concepts.The solid angle is shown bounded by a square; the crosssection may have any shape and is shown here as squarefor clarity. Solid angle: The area A bounded by JKLMsubtends a solid angle steradians at the source Sdefined as = A/R2 steradians. Radiant intensity: Thesource emits a power of in all directions and deliversinto the solid angle a reduced quantity known as theradiant intensity defined as l = d/d with units of wattsper steradian. Radiance: Each small area dA of thesurface of an emitting source emits power into a solidangle of : this is the radiance defined as L = d2/dA.dwith units of watts per steradian per m2.
spectral absorption curve. This curve is multiplied,wavelength by wavelength, by the spectral powerdistribution of the radiation source. This convolutionprocess yields the relative absorbed spectral dose curve,which specifies the energy deposited in the substrateacross the spectrum. If the intensity of the radiationsource is increased, the area under this curve increasesalso. Indeed, this area, is the spectral integral defined so:
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com
5
(6)( )Q a p dmax = ⋅∑ λ λλ
λλ
1
2
where a and p are the spectral absorption and spectralpower and represent the maximum possible energy that isavailable to influence the substrate. It is a useful, butnecessarily rough, guide in comparing the effects ofdifferent radiation sources on one substrate, or howdifferent substrates might behave if exposed to the sameradiation source.
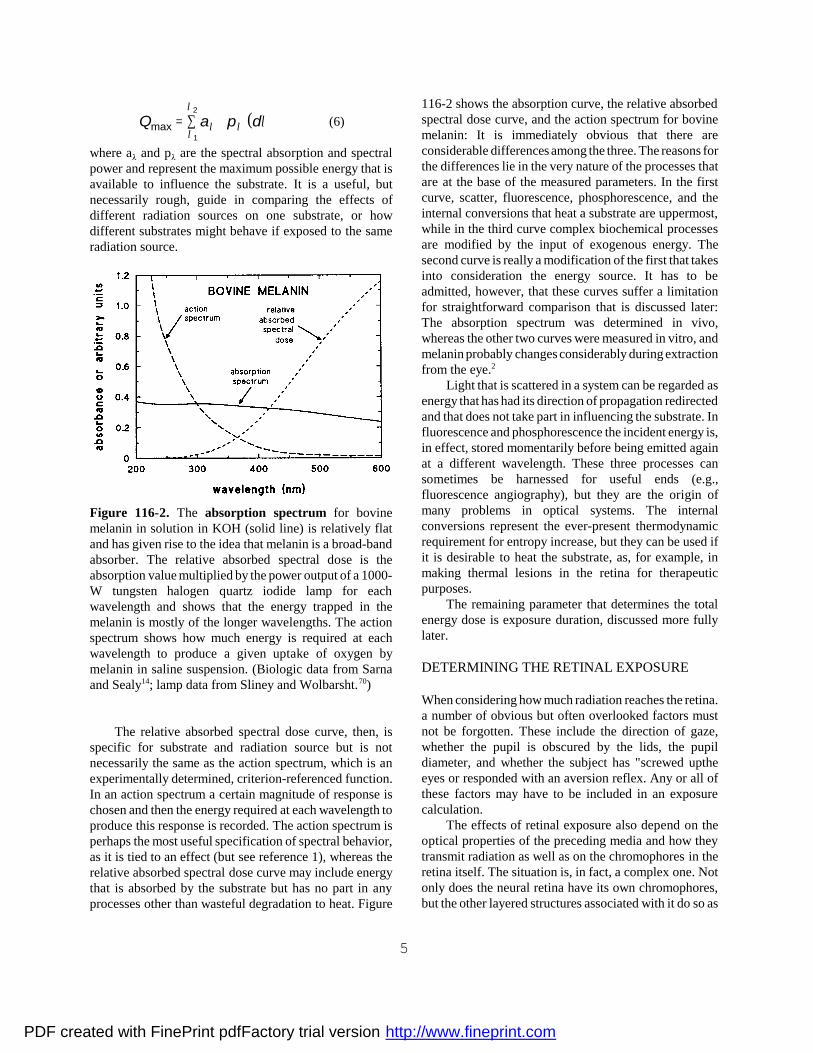
Figure 116-2. The absorption spectrum for bovinemelanin in solution in KOH (solid line) is relatively flatand has given rise to the idea that melanin is a broad-bandabsorber. The relative absorbed spectral dose is theabsorption value multiplied by the power output of a 1000-W tungsten halogen quartz iodide lamp for eachwavelength and shows that the energy trapped in themelanin is mostly of the longer wavelengths. The actionspectrum shows how much energy is required at eachwavelength to produce a given uptake of oxygen bymelanin in saline suspension. (Biologic data from Sarnaand Sealy14; lamp data from Sliney and Wolbarsht.70)
The relative absorbed spectral dose curve, then, isspecific for substrate and radiation source but is notnecessarily the same as the action spectrum, which is anexperimentally determined, criterion-referenced function.In an action spectrum a certain magnitude of response ischosen and then the energy required at each wavelength toproduce this response is recorded. The action spectrum isperhaps the most useful specification of spectral behavior,as it is tied to an effect (but see reference 1), whereas therelative absorbed spectral dose curve may include energythat is absorbed by the substrate but has no part in anyprocesses other than wasteful degradation to heat. Figure
116-2 shows the absorption curve, the relative absorbedspectral dose curve, and the action spectrum for bovinemelanin: It is immediately obvious that there areconsiderable differences among the three. The reasons forthe differences lie in the very nature of the processes thatare at the base of the measured parameters. In the firstcurve, scatter, fluorescence, phosphorescence, and theinternal conversions that heat a substrate are uppermost,while in the third curve complex biochemical processesare modified by the input of exogenous energy. Thesecond curve is really a modification of the first that takesinto consideration the energy source. It has to beadmitted, however, that these curves suffer a limitationfor straightforward comparison that is discussed later:The absorption spectrum was determined in vivo,whereas the other two curves were measured in vitro, andmelanin probably changes considerably during extractionfrom the eye.2
Light that is scattered in a system can be regarded asenergy that has had its direction of propagation redirectedand that does not take part in influencing the substrate. Influorescence and phosphorescence the incident energy is,in effect, stored momentarily before being emitted againat a different wavelength. These three processes cansometimes be harnessed for useful ends (e.g.,fluorescence angiography), but they are the origin ofmany problems in optical systems. The internalconversions represent the ever-present thermodynamicrequirement for entropy increase, but they can be used ifit is desirable to heat the substrate, as, for example, inmaking thermal lesions in the retina for therapeuticpurposes.
The remaining parameter that determines the totalenergy dose is exposure duration, discussed more fullylater.
DETERMINING THE RETINAL EXPOSURE
When considering how much radiation reaches the retina.a number of obvious but often overlooked factors mustnot be forgotten. These include the direction of gaze,whether the pupil is obscured by the lids, the pupildiameter, and whether the subject has "screwed uptheeyes or responded with an aversion reflex. Any or all ofthese factors may have to be included in an exposurecalculation.
The effects of retinal exposure also depend on theoptical properties of the preceding media and how theytransmit radiation as well as on the chromophores in theretina itself. The situation is, in fact, a complex one. Notonly does the neural retina have its own chromophores,but the other layered structures associated with it do so as
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

6
well. It is necessary, therefore, when considering the retinato take account of the ocular media and these other layersand the energy that they trap. This is because this energycan either spread to the retina proper as a thermal insult or,by compromising the function of closely related tissues,compromise the retina too by upsetting normal physiologicprocesses. The chromophores in the retina and the retinalpigment epithelium (RPE), together with the melanin in theRPE and choroid and the hemoglobin in the choriocap-illaris, must be included in any discussion of radiationinsult to the retina.
Figure 116-3. Spectral transmission curves of rhesusmonkey ocular media. C, Cornea (path length = 0.55 mm),.A aqueous (path length 2.86 mm); V, vitreous (path length= 11.55 mm); and, for comparison, the dotted curve, W, ofdistilled water (path length = 18.5 mm). (Data fromMaher.11)
Of the pre-retinal ocular media, only the lens hassignificant absorption, scatter, or fluorescence in theregion of the spectrum from UVA to IRC. Figure 1163shows absorption curves for the cornea, aqueous, andvitreous; the extent of absorption changes with age in thesemedia, but only significantly in the cornea at the shorterwavelengths. Also shown is the curve for water, whichobviously determines most of the absorption of thesemedia. the effect on the passage of radiation to the retinaof these media is slight, except for the lens, and occasionslittle disagreement among published authorities.Obviously, in aphakic eyes or those with intraocular lens(IOL) implants the situation is very different, and in theformer eves visibility of UV light does diminish with age.In those with IOLs the material of the lens controls thequality of radiation that reaches the retina. These lenses are
usually made of polymethyimethacrylate (PMMA) andmay have a UV blocker, and perhaps pigmentation, tomimic the transmission properties of an aging crystallinelens.3
Figure 116-4. Spectral curves for the optical density atthe thickest part of the lens (pole) for six typical lensestaken from the study by Mellerio.11 The age of each lensis shown in italic figures on the ordinate at the start ofeach curve.
The crystalline is well known for its absorption ofUV and blue light but there is in the literature aconsiderable range of transmission values.4 Figure 116-4shows transmission curves for six lenses of different ages,but authors have not always specified the age of theirmaterial or whether it was in any way cataractous. Figure116-5 shows results at one wavelength (400 nm) fromnine studies that do show the effects of aging. The verylarge discrepancies -well over 1 log unit at 400 nm - arethought to derive partly from a real variation of opticalproperties, scatter as well as absorption,' and partly fromthe various techniques of measurement. Weale6 criticizesWerner7 for the improper inclusion of some of the data inhis figures, which are, nevertheless, included in Figure116-5. The methods of determining lens absorption rangefrom subjective color matching or photography of thecrystalline in vivo to various ways of dealing with lensestaken out of the eye. Another difficulty arises because theloss of light transmission in the lens is due to scattering(or fluorescence) and not only to absorption.5, 6 It isnecessary to bear this in mind when using lenstransmission figures, as the method of measuring themmay have influenced their magnitude. At shortwavelengths and especially with older lenses, the spreadof lens transmission values is so great that individual
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

7
cases can readily be found that would support the choiceof almost any figure for transmission-you pay your moneyand you take your choice.
Figure 116-5. Optical density of human lenses at 400 rimplotted against age. The large filled circles and the fittedsecond-order polynomial curve are from the paper byMellerio72 and the small solid triangles are from Werner,7
who took the data from eight other authors for comparison.Werner's data are fitted by a straight line regression (D =0.021 age + 0.94). The two sets of data clearly show theincrease of density with age and the increase in range ofvalues that accompanies this.
Reflection at, and light scatter within, the layers of theneural retina, RPE, and choroid are not usually consideredimportant in assessing the possibility of retinal lightdamage, because the fraction distributed away from theretina in this way is small compared with the directirradiation within the retinal image (typically 10 to 15percent but see reference 8). The various chromophores inthese layers absorb light, which may initiate damagingprocesses, and thus modify it before it is passed on todeeper chromophores. A complete picture of what happensin these layers may be complex, and it is necessary toconsider the known chromophores in turn to gain anunderstanding of the processes involved. Again, it must beremembered that absorption spectra may not adequatelydescribe the possibility of damage, because they are notaction spectra. Indeed, those action spectra that arecurrently available for light-induced damage in the retina
do not all closely match the absorption curves of the mostobvious chromophores. Nevertheless, it is usual toconsider those discussed below as the prime movers ofdamage.
The yellow macular pigment xanthophyll is confinedto the outer plexiform layer; its absorption spectrum isshown in Figure 116-6. If this pigment is a damageoriginator, it might be involved in the photochemicaldamage that is described below. Its contribution tothermal damage might be small because its total absorp-tion is much less than that of melanin, but in the foveaeven small lesions can have enormous visual effects.
Figure 116-6. Spectral absorption curves for melanin,xanthophyll, cone pigments, and oxygenated hemoglobin.The absorption is the fraction of incident radiation at thecornea that is absorbed in the pigment-bearing structurein the eye. Consequently, the curves include absorption inthe lenticular pigment: the age of the lens chosen for thecalculations was 52 years.72 The cone pigment absorptionis calculated for a cone of average outer segment lengthand pigment loading. The hernoglobin is assumed to bein a retinal vessel 10 m in diameter and the macularpigment uses the values found in whole retinal prepa-rations by Marshall (personal communication). Themelanin figures are for the combined absorption in theRPE and choroid and are derived from unpublished datafrom Marshall and the paper by Boulton and coworkers.13
The visual pigments are also obvious chromophoresthat could originate damage. Their total optical density issmall, and their primary involvement in thermal damageis not likely to be extensive or, in clinical situations,important, except in the fovea. Undoubtedly, they makea contribution to photochemical damage and theirabsorption spectra (Fig. 116-6; see also Fig. 11614) are
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

8
not incompatible with the presumptive action spectra forone type of this damage. For example, Harwerth andSperling9, 10 were able to produce damage in blue cones orgreen cones by exposing primates to appropriately coloredlights. However, the fact that the blue cones had a lowerthreshold than that of the green (and presumably the red)does not correlate with the total quantity of pigmentpresent in the outer segments of each cone type. Also, itturns out that cones are more sensitive than rods, althoughmore rhodopsin is present in the longer outer segments ofthe latter. This situation has little to do with pigmentabsorption and arises because the repair processes withinthe cones act more slowly to restore normality than thosein the rods.
Figure 116-7. Spectral absorption curves of melanin invarious situations. BG, a suspension of human melaningranules pooled from the RPE of eyes over 50 years old;suspension in buffered saline. BS, a solution of humanmelanin granules as in BG, solvent was 1M KOH. PG,mean results from 16 flat mounts of human RPE measuredunder a microscope; age not specified. GB, mean results of28 human retinas, RPE, and choroid, calculated to includethe preretinal ocular media (which explains the short-wavelength cutoff); age of donors ranged from 23 to 78 years.(Data from: BG and BS13; PG17; GB8.)
Hemoglobin is present in the internal retinal circula-tion and is often a target for clinicians trying to close off ordestroy blood vessels and circulatory anomalies. Usually,an immediate effect is required, so the intention is toproduce a thermal lesion by focusing radiation on theoffending vasculature. There is also a "lake" ofhemoglobin in the choriocapillaris. The absorption of thisblood pigment is, therefore, held to be important; Figure116~6 shows this for the oxygenated form. The
expectation that yellow light from a dye laser should beused to destroy blood-bearing structures has been onlypartially established because in total there is not muchabsorbing power in the thin layers of blood that retinal orchoroidal capillaries contain. In addition, the flow ofblood in retinal vessels carries away heat, so that it isdifficult to close a vessel without spreading energy toneighboring structures.11 Blood flow is not an importantfactor for the choriocapillaris, because, as Birngruber andcoworkers12 showed, threshold doses for thermal lesionsof the retina are not affected by the presence or absenceof choroidal blood flow.
Undoubtedly, the most avidly absorbingchromophore present is the melanin of the RPE andchoroid. Figure 116-7 depicts the spectral absorptioncurve for a solution of melanin; it is not possible tomeasure absorption for an individual granule, as it is sogreat. The absorption of the solution is believed bysome.13, 14 to describe adequately melanin absorption inthe eye, but the conditions used to obtain solution (hot,concentrated potassium hydroxide [KOH]) are so harshand unphysiologic that it is difficult to believe thatmelanin solution can tell us much that is of use forunderstanding the situation in vivo. Spectral absorptioncan be measured for suspensions of granules (Fig. 116-7),but the sieve effect15 gives a curve that is too flat. Inaddition, scattering of light among the granules leads toartifactual wavelength-dependent effects. The curves ofBoulton and coworkers13 show minor peaks and troughsin the plots for melanin granule suspensions; these maybe due to selective scatter or to contaminants that do notfind their way into alkali solution.
The relative absorbed spectral dose cannot be calcu-lated without knowing the spectral absorption of melanin.So, if solutions yield values that are doubtful and one issuspicious of suspensions, might it be possible to measureabsorption of melanin in vivo or in vitro? Several reportshave raised such questions, and not only in ocularcircles,16 and though some have attempted measurementsin vitro, to date none has convincingly measured melaninabsorption in the living eye. Geeraets and Berry8 were thefirst to measure absorption of the RPE and choroid inhuman tissue (Fig. 116-7), but this measurement isexpressed in terms of light incident at the cornea and soincludes the transmission properties of the ocular media.Also shown is the curve Gabel and colleagues17 madeusing a microscope to examine pieces of human RPEsheet mounted on a slide. The solution and the tissuecurves have approximately the same slope, but in linearplots this signifies little: Curves such as these speak ofconsiderable variation between studies that arise eitherfrom the different techniques used to extract and measure
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

9
melanin absorption or, more interestingly, from differencesin the melanin itself, e.g., those due to genetic factors.Tissue curves such as those in Figure 116-7 give the meanpicture, but as the melanin is distributed in granules theyare not useful when considering retinal heating withmicroscale models.
EXPERIMENTAL LIGHT DAMAGE TO THERETINA
The damage that radiation produces is usually classifiedunder one of three heads (Fig. 116-8). In reality, there is acontinuum of observable effects, and the three divisionsare more for convenience of discussion than they aretheoretically significant. The intermediate classes ofthermally assisted chemical effects and thermoacousticeffects underline the seamless nature of the continuum.
Photochemical Damage
By definition, photochemical damage is brought about byphotons of such high intrinsic energy (i.e., shortwavelength) that chemical bonds of the molecules thatcompose the tissues are directly split. Once a bond isbroken in a biomolecule the molecule may change itsconformation and thus lose its function. For example, across-linking bond in a protein may be split, allowing themolecule to uncoil and change shape dramatically. Ofcourse, with such complex molecules as proteins there maybe no conformational change if one or even a few bondsare broken, but whether this happens depends very muchon which protein is under consideration. The evolutionarysignificance of neutral mutations, some of which resultfrom photochemical damage to DNA, has a bearing on theineffectiveness of photochemical damage, as may beargued from the views set forth by Jukes.18
The portion of the molecule, or the functional group,that absorbs the incident photon is termed a chromophore.The electronic transition to the excited state absorbs theenergy of the photon, but the decay and release of thisenergy may proceed by a number of routes. One may bethe transfer of the energy to split a bond either in thechromophore or elsewhere in the molecule, evensometimes at sites quite remote from the absorption locus.If a covalent bond is split, each fraction of the moleculemay retain one of the shared electrons to become a freeradical. These radicals are highly reactive and exist onlybriefly before they react with neighboring molecules.19
If the energy in the incident photon can be coupled tothe molecule more efficiently, the likelihood of bondsplitting is enhanced. This may be done with a vital dye,which is an efficient chromophore that can pass on its
trapped energy to the bound molecule, thus leading tobond breaking. Such vital stains as toluidine blue, rosebengal, hematoporphyrin, and many others can so in-crease the energy trapped that irradiated cells containingthem may die.20 This process is used therapeutically totreat certain tumors in the eye and elsewhere.21 Vitalstains are exogenous sensitizers. There is a range ofendogenous sensitizers too: chlorophyll, riboflavin, andbilirubin are three that have been suggested to havebiologic importance.22 Lipcifuscin, which is found inincreasing amounts in the aging retina, might also act asa chromophorell in elderly eyes, in which age-relatedmacular degeneration is often a problem. Interestingly,the visual pigment retinol can act as a sensitizer.
Figure 116-8. The mechanisms for light-induced retinaldamage related to exposure duration.
The formation of free radicals is undoubtedly moreimportant in producing tissue damage than the splitting ofbonds. Free radicals, such as singlet oxygen, can attackmany molecule types and render them ineffective. It isthis hyperactivity that makes the free radicals so toxic.24
There is an additional risk in tissues where there is a largeconcentration of cell membranes, which are, of course,composed largely of lipids. Once one of the -CH2- groupsin the chain of a polyunsaturated fatty acid in a lipidreacts with a free radical, a chain reaction of lipidperoxidation starts. and it ends eventually in thedecomposition of the molecule. As lipids form thebackbone of membranes, the membranes are brokendown. This process is seen in Figure 116-9, which showsthe early stages of light damage in the outer segment of apigeon cone.25 The retina is thus well placed to sustainthis kind of damage, as it has an endogenous sensitizerand an abundant supply of oxygen for singlet oxygenformation and it is nicely placed under a transparentfocusing system designed especially to collect light.26, 27
Its most important structures. the membrane discs ofexposure to a bank of standard fluorescent lamps thatwere as bright as an overcast London sky. Note thepunctate damage to the disc membranes resulting from
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

10
the early stages of light damage induced by lipidperoxidation the outer segments of the photoreceptors. aremade almost entirely of lipid just waiting to beperoxidized.
Figure 116-9. Electron micrograph of part of the outersegment of a pigeon cone 7 days after several hours' .
Whether a bond is broken depends on the energy ofthe incident photons: If the energy is not great enough, thebond will never be parted. The probability that the bondwill be broken depends on the number of photons incidenton a chromophore, provided that they have sufficientintrinsic energy. The changes that a particular wavelengthof radiation brings about are always the same, but theyproceed at a rate that varies only with the incident flux.Thus, photochemical reactions are dose dependent: Acertain number of photons are needed to complete thereaction by breaking a certain number of bonds, and thereaction proceeds at a rate determined by the rate at whichphotons arrive, that is, by the irradiance. Consequently, areciprocal relationship exists between irradiance andexposure duration (Fig. 116-10). Indeed, the
demonstration of a direct reciprocal relation betweenthese two quantities is usually taken as confirmation thata process is photochemical.
Figure 116-10 shows that two distinct processes cancause damage to the retina. One has the expected slope of- 1, establishing this as photochemical; the other has avery different and much smaller slope, representingthermal damaging processes (see further on). There is awave le ngth-depende nt effect; the argon laser, with itsenergetic photons, readily causes photochemical damage,whereas the infrared Nd:YAG laser, with photons onlyhalf as energetic as those from the argon device, causesonly thermal damage.
Figure 116-10. Log retinal irradiance (as measured at thecornea) plotted against log exposure duration forproduction of just visible threshold retinal damage fromsingle exposures to argon laser radiation (514 nm, #) andto Nd:YAG laser radiation (1064 nm, !). The dashed linehas a slope of - 1 and is a by-eye fit: it clearlydemonstrates the reciprocal nature of the damage atlonger exposures for argon radiation. The dotted lines arebyeye fits of very low slope and represent damageinduced by thermal processes. These data are selectedfrom Sliney.21
Photochemical damage, of which sunburn is a goodexample, usually demonstrates delayed onset. In theretina the delay may be several minutes or several hours.This is because overall cellular processes have turnovercycles of minutes or hours and photochemical damageworks by destroying perhaps just one molecular com-ponent of a cell's biochemistry. In addition, photochem-ical damage to the retina produced in laboratory studiesis often widely distributed and diffuse. as would beexpected if the chromophore were some endogenouscomponent of a basic biochemical process, such as
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

11
glycolysis or cell respiration. There is a second reason forthe widespread nature of this damage in the retina. As theirradiances are low and the exposures are long, the lightsources used are often extended (e.g., fluorescent lamps).In extralaboratory situations, damage may result from, forexample, reflection from snow, and this, too, represents anextended light source.
It has become customary in recent years to classifyretinal photochemical damage as either type 1 or type 2.This is an unfortunate choice of name, however, for itleads to confusion with the terms generally used inphotochemistry for reactions in which the photosensitizedmolecule reacts either directly with the substrate (type 1reaction) or with molecular oxygen to form singlet oxygenor superoxide free radicals, which in turn react with thesubstrate (type 2 reaction). To avoid this confusion, weprefer the eponymous Noell- and Ham- type damage, toacknowledge the respective discoverers of thesephenomena. The classification is based on damagedistribution within the retinal layers and on the exposureparameters, but the division is not wholly satisfactory.Some authorities have found the division artificial, andrecent evidence points to several damage mechanisms thatoverlap to produce a continuum of effects. from Noell toHam damage. Nevertheless, consideration ofphotochemical retinal damage can well begin with adiscussion of these two classes of damage; they aredescribed here together with some criticisms. It must alsobe remembered that once a damage process is under way,the end results are often very similar. Therefore, workershave had to pay particular attention to damage caused byjust suprathreshold doses of radiation. This makes thework difficult and very dependent on subjective evaluationof histologic material.
NOELL DAMAGE
Noell damage is caused by long exposures (typicallyhours or even days) to low irradiances; at threshold, thefirst manifestations of damage are seen in the photore-ceptors. The area of retina damaged is usually large, forthe reasons mentioned earlier. The first report of this typeof damage was by Noell and coworkers,28 who showeddamage in the retinas of rats in cages surrounded byfluorescent lamps. It was soon established that cones weremore sensitive than rods to light damage, which initially isintriguing but is now explicable in terms of photoreceptorrepair processes (see later). By exposing animals tocolored lights it is possible to damage the group of conessensitive to these colors,25 and even to produce animals thatare effectively colorblind.10 It is also possible that Noelldamage is responsible for the reduction in blue lightsensitivity that develops in ophthalmic surgeons who have
been cumulatively exposed to argon laser radiation formany hours when treating patients in the laser clinic.29 Ifthe damage appears first in the photoreceptors, and conepopulations can be selectively destroyed, thechromophore responsible might be in the receptor outersegments. The finding that the action spectrum of Noelldamage corresponds relatively well with the absorptionspectrum of the visual pigments supports this claim.28, 30-32
HAM DAMAGEHam damage is caused by irradiances higher than thosethat produce Noell damage, and the exposures are shorter(seconds to minutes). The area of retinal damage issmaller because the irradiances required are usuallypossible only with light sources that are presented to theeye in collimated beams. The damage. first described byHam and colleagues,33 is held to originate in the RPE.Ham measured the action spectrum of the damage34 andshowed that its sensitivity increased with reducingwavelength of light (Fig. 116-11). Consequently, thisHam-type damage is sometimes known as Blue LightDamage. In earlier papers, Ham's group34 thought that thesite of origin of the damage was “located in the outersegment of the photoreceptor and perhaps in the pigmentepithelium.” Later, they, were able to show that thegreatest damage appeared in the RPE but that thephotoreceptor was also affected.26 Because of theinvolvement of the RPE, melanin has been suggested asthe chromophore responsible, but as Figure 116-11shows, the blue light damage action spectrum does notremotely match the melanin action spectrum for uptake ofoxygen (see Fig. 116-2) or the absorption spectrum formelanin solution.14 Since the Ham action spectrum wasmade for a whole eye and the absorption of the ocularmedia is included in the curve, the disparity would beeven greater for direct retinal exposure.
Sarna2 reviewed the role of melanin as a protector ofcell integrity. It is established that melanin protects thecells containing it from oxidative damage-the RPE andchoroidal cytoplasm have very high oxygen concentra-tions-probably by binding oxidative catalytic metal ions,which are thus removed from the biochemical arena ofthe cell. Sarna also reviewed the evidence for a toxic rolefor melanin, which has frequently been suggested, butconcluded that in vivo, as opposed to in vitro, melaninwould not normally release free radicals or promoteoxidative processes. With aging the constant exposure tohigh oxygen levels and years of daily light doses mightcause the antioxidant properties of melanin to diminishand allow its pro-oxidative proclivities to gain the upperhand, so inducing the damage we know as age-relatedmacular degeneration. Sarna's conclusions would seem tosupport the doubts that melanin is the chromophore that
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

12
initiates Ham-type damage in the retina.
Figure 116-11. The blue light action spectrum from Hamand coworkers34 for 100 sec exposures of monkey eyes; theordinate (note the logarithmic scale) is reciprocal retinalirradiance (cm2/W) for just fundoscopically visible lesions24 hours after exposure; the data points are shown fitted bya second-order polynomial curve. Shown for comparisonare the melanin action spectrum for oxygen uptake andmelanin solution absorption curve (see Fig. 116-2 fordetails)14; both curves have been shifted vertically to becoincident with the Ham curve at wavelengths above 600nm; for both these curves the ordinate units must beconsidered arbitrary.
Though the existence of blue light damage has beenconfirmed by several reports and the data have beenincorporated into the codes of practice for the safe use ofintense light sources, accounts of where the damageoriginates do not completely agree. The view of Ham andcoworkers26 that it is not easy to distinguish betweenprimary damage to the photoreceptors and secondarydamage resulting from damage to the RPE must be givendue weight. Ham has suggested34 that blue light damageprobably is not the result of a single process, and theliterature supports this view. Rapp and associates31 havesuggested that mitochondrial enzymes may be thechromophore for the blue light damage and cite the initialswelling of these organelles to substantiate their claim.Pautler and coworkers35 also implicate mitochondria, andin particular the cytochrome enzymes resident therein.Using electron microscopy they showed that the initialdamage resulting from light exposure was to themitochondria. Kremers and van Norren36 suggest that theproducts of photobleaching may constitute thechromophore. Whatever the chromophore, Ham and
coworkers26 have shown that oxygen seems to be nec-essary for Ham damage to occur and that if oxygentensions are increased in the retinas of irradiated animals,the damage increases and the threshold drops. They alsodemonstrated that certain free radical quenchers protectedthe retina from the effects of increased oxygen tension.Thus Ham damage would appear to involve free radicals,especially oxygen.
Kremers and van Norren36 pointed out that mostlaboratory studies that show Noell damage have usedrodents exposed for a long time, whereas studies thathave reported Ham damage have been made with pri-mates exposed for up to 4 hours. Reasoning that thisdivision of species and exposure parameters might ex-plain some or all of the differences between Noell andHam damage, they exposed primate retinas for as long as12 hours. They then compared their data with those fromthe literature by plotting exposure duration against thedamage threshold irradiance for both types of damage.37
Their graph is replotted as irradiance against exposure inFigure 116-12, which convincingly shows a bipartitecurve that separates the two damage types. The Hamregion shows a reciprocal relation between irradiance andduration, with the expected photochemical slope of - 1.There is an abrupt change at 12 hours, when the thresholddrops by a factor of 100 before Noell damage continuesfor longer periods, also with a slope of - 1.
Kremers and van Norren's examination of the datashows no obvious reason for a correlation between animalspecies and type of damage except that it is difficult andmore expensive to expose primate retinas continuously tolight. Their own extended exposures of monkeyssuggested that the correlation of damage type withspecies was fortuitous. The report of Rapp andcoworkers31 confirms this view: They claim to haveproduced in one animal type, the Long-Evans rat, lightdamage due to two different mechanisms initiated byUVA and green radiation. One was initiated by UVAradiation peaking around 355 nim and 500 W.cm-2 at thecornea and revealed by minimal disorganization of thephotoreceptor outer segments after 1 day, pyknosis oftheir nuclei, and severe swelling of the rod innersegments and RPE mitochondria. Exposure to 5000W.cm-2 green light, on the other hand, caused severederangement of the inner and outer segments but left theRPE appearing normal. The intensities of the radiationfor threshold damage are markedly different at thecornea, and this difference increases greatly for retinalintensities if the high absorption of the ocular media toUVA radiation is taken into account. As might beexpected, the UVA radiation was also markedly inferiorin bleaching rhodopsin compared with the green light.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

13
Figure 116-12. Log retinal irradiance plotted against logexposure duration forjust detectable retinal photochemicaldamage. The data are taken from Kremers and vanNorren,37 who derived them from the literature ofphotochemical damage to different species of animal anddifferent exposure regimens. The arrow indicates exposureof 12 hours, and the dashed lines have a slope of - 1.
Van Norren and Schellekens32 have alsoproducedNoell and Ham damage in rats. They confirm thatthe action spectrum of Ham damage in the rat is like Ham'smonkey blue light spectrum, but they do not know whatpigment or pigments initiate the process (Fig. 116-13).Pautler and associates35 measured an action spectrum forreduction of the transepithelial potential in bovine RPE inan Ussing chamber. They argue that this spectrum fits verywell the absorption spectrum of cytochrome C oxidase,which is found in mitochondria, the organelles they believeto be the initial site of Ham damage. Their action spectrumis also shown in Figure 116-13: In their paper they claimthat it fits well with the action spectrum from Ham'sgroup34-between 400 and 500 nm-but the fit is not tooconvincing. They are on surer ground when they point outthe considerable disparity at shorter wavelengths and, as aresult, suggest that two mechanisms must be responsiblefor the damage, one dependent on blue light and one onUVA radiation. Some chromophore with higher absorptionin the UVA region is therefore required, and in Figure 4 of
their paper they compare the absorption spectrum of all-trans retinol (which peaks at 360 to 370 nm) with theiraction spectrum and reject it as a candidate for damageinitiation in favor of cytochrome C oxidase. This isreasonable, because their preparation contains only theRPE separated from the neuroretina an hour or morebefore; for an intact retina, the basis of Ham's group14
experiments, all-trans retinol and other products ofbleaching are formed continuously during exposure. Thisline of argument could be held to support the Kremersand van Norren model described below, but a fullerexamination of the absorption properties of the variousbleaching products should be undertaken, together witha consideration of the contribution of each that will ensuefrom the kind of dynamic equilibrium between bleachingand regeneration that this model requires for the longishexposures that produce Ham damage.
Because the action spectrum of Noell damagefits the absorption curve of the visual pigmentsreasonably well,28, 38 van Norren and colleagues39
proposed an explanation for their unifying curve. Withlow irradiances the most sensitive damage-inducingchromophores, the visual pigments, couple energy to aphotochemical damage mechanism (probably lipidperoxidation of disc membrane25, 27), and Noell damageprocesses are set in train. With higher irradiances thevisual pigments are rapidly bleached and effectivelyremoved, so that a less sensitive damage-inducingchromophore, nature unknown, couples energy to RPEsystems, which results in Ham damage. This schememight have two versions (Fig. 116-14). In version A,more intense radiation bleaches the visual pigment tosuch an extent that radiation can reach an unspecifiedsensitizer, presumably in the RPE. In version B. it is theproducts of bleaching themselves that are the sensitizer:The bleach-regeneration cycle of visual pigments usesseveral different products in dynamic equilibrium withinthe photoreceptor and a retinol store in the RPE. TheUVA absorption of retinol in the RPE35 (discussedearlier), coupled to that of mitochondrial enzymes, mightexplain the origin of Ham damage.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

14
Figure 116-13. Action spectra for Noell and Hamphotochemical damage to retina. !, Spectrum for Noelldamage, data points from Noell and coworker28 andWilliams and Howell,38 and the line is the scotopicvisibility curve. #, Spectrum for 100 s exposure of monkeyfrom fiam and coworkers. , Spectrum for rat damagefrom van Norren and Schellekens. 32 , Spectrum forbovine RPE transepithelial potential from Pautler andcoworkers.35 The spectra have been moved vertically byarbitrary amounts for comparison.
As photochemical reactions depend on the energy ofthe photons and the rate of reaction depends on the numberof photons incident per unit time, there is no "threshold"for a photochemical reaction. It simply happens faster orslower depending on the irradiance. That threshold criteriawere used by the various investigators whose data wereplotted by Ki-emers and van Norren might, therefore, seemconfusing, as the effects are photochemical. The fact is thatthresholds arise here because the effects have to be of acertain magnitude before they can be detected. Mostly.ophthalmoscopic visibility has been taken as the thresholdcriterion for retinal lesions, so the concept of threshold hassignificance only at a macroscopic level.
Figure 116-14. A summary diagram of two possibleversions of the Kremers and van Norren explanation ofphotochemical damage to the retina. In Noell damage theirradiance is low and a dynamic equilibrium is establishedbetween the bleaching of visual pigments (hatched box)and their regeneration: energy is coupled from the visualpigments acting as sensitizers to the damaging processes.For flam damage the irradiance is high and theequilibrium of bleached products and regenerated visualpigment is pushed so that hardly any pigment is present:in & the radiation can pass to the as yet unknownsensitizer to couple energy to the damaging processes,whilst in B, the products of bleaching are suggested to bethe sensitizer.
Photochemical damage is not immediately obviousbecause the damaged molecules may take part inbiochemical pathways that have finite rates of reaction, solack of products or excess of intermediates takes time tobe noticed. Also, all cells have repair and renewalprocesses that attempt to nullify any damage. Of course,if the incident radiation destroys molecules faster thanthey can be repaired, there will be a net increase indamage; the effect will he seen subsequently and athreshold established.
At normal body temperatures, thermal processes con-stantly denature biomolecules. but an equilibrium existsbetween thermal degradation and repair processes in ahealthy cell. There is always spare capacity in the repairprocesses to provide for any emergency situation, butelevation of body temperature increases the rate ofdegradation and reduces the spare repair capacity. Thisexplains the familiar observation that at high bodytemperatures photochemical damage is enhanced and theirradiance -thresholds- are lowered.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

15
The presence of repair processes leads to a departureof photochemical damage thresholds from the strictreciprocity relation that is expected.40 This will be es-pecially obvious at long-duration exposures. in which theirradiances are low and the repair processes very nearlybalance the damage mechanisms. This concept isillustrated in Figure 116-15, in which the ordinate rep-resents the irradiance, which is directly related to the rateof degradation, and the abscissa is the duration ofexposure. The area in which the rate of repair can exceedthe rate of degradation is shaded; the solid line shows theexpected relation between irradiance and durationdeparting from strict reciprocity at shorter exposures to anasymptote parallel to the time axis at longer times.
Thermal Damage
The term -thermal damage- implies a rise in temperaturewithin the tissues that are damaged: To understand theprocess it is necessary to know what is meant bytemperature rise and how incident energy can bring thisabout. Temperature is a measure of the kinetic energystored throughout a large population of molecules in theirvibrational, rotational. and intermolecular motions-it hasno meaning for individual or very small assemblages ofmolecules . An analogy to temperature would he theamount of jostling in a crowd at a football match, aconcept that becomes meaningless for a very fewindividuals in an otherwise empty stadium. If thetemperature or stored kinetic energy increases, there comesa point when molecules shake themselves free of theinfluence of other molecules (breaking intermolecularbonds) or even shake themselves to pieces (breakingintramolecular bonds). This energy comes initially (1)from internal conversions, where excited electronic statesdecay, releasing energy to vibrational or rotational states;or (2) by direct absorption of incident energy into vibrationor rotation. It is also possible to have transfer of energyfrom a variety of processes through the members of apopulation of molecules.
Figure 116-15. Damage rate (or irradiance) plottedagainst duration of exposure for photochemical retinaldamage (both axes in arbitrary units). The shaded arearepresents the area where repair processes exceed the rateof degradation, the dashed line shows the reciprocityrelation of slope - 1, and the solid line the practicaldivision between damage and no damage.
The kinetic energy that is temperature comes, as faras this discussions concerned, from the input of radiantenergy into the retina. Unlike photochemical reactions, inwhich there is no irradiance threshold and irradiance andexposure duration are linked by reciprocity, thermalreactions do have irradiance thresholds and do not showreciprocity. This is because there is no photon energyrequirement for bond breaking. Sufficient energy toincrease the temperature can come by any route. while atthe same time this energy can leak harmlessly away,especially if the exposure duration is long. Consequently,the energy required to produce a thermal effect increaseswith exposure time and the irradiance-duration relationfor just detectable lesions is not linear but is typically ofthe form:
(7)E K te = ⋅ −0 25.
where K is a constant and t is between 20 sec and 10sec.22, 40 In Figure 116-10 the “thermal” portions of theplots have slopes of about - 0.12, so for the particularexperimental circumstances that apply to this figure theexponent is about half the usually accepted value.
The major structural molecules of biologic tissuesare proteins, which are held in highly specificconfigurations by hydrogen bonds and Van der Waals'forces. These important cross-links are relatively weakand readily destroyed by heating. When the bonds arebroken, the protein may uncoil and recoil into a new
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

16
configuration, which could completely remove any usefulfunction that the protein had, This is the process ofdenaturation or coagulation and is easily seen whenpoaching or boiling an egg. The unfolding and refoldingtakes a finite time that may be about the same as theduration of some of the shorter laser pulses that can inducethermal damage. Proteins are particularly sensitive to heat,and they turn out to be the cell's Achilles' heel for thermaldamage. If even a few protein molecules are destroyed byheat, the repair processes may not be able to effect a repairand cell death could follow. Marshall41 has argued thatonly a small portion of a cell's total protein complementneed be damaged for cell death to occur. In such a highlyspecialized tissue as the retina, damage that leads to celldeath in any layer is readily spread to other cells whoseexistence depends on the functional integrity of the firstcells. A typical situation occurs when photoreceptors aredamaged by any type of insult and complete retinalatrophy ensues. For radiation damage in the retina thisprocess can be termed damage amplification, for it is onlynecessary to destroy a few pigment epithelial cells orphotoreceptors to induce full-thickness retinal atrophy.
Thermal effects can occur only if incident energy istrapped or absorbed in the substrate molecules, Thisabsorption is easily measured, and it is generally expectedthat an absorption value of a substrate will provide anindication of what increase in temperature might result.There is thus considerable interest in the absorbingproperties of ocular tissues on the part of investigatorstrying to evaluate what effects are produced or trying toproduce desired treatment schemes. Spectral absorptioncurves may yield a full specification for a substrate, but ifthe radiation comes from a wavelength-limited source,such as a laser with its monochromatic output, it is oftenmore convenient to specify the absorption at the chosenwavelength. This absorption is related to the incidentradiation by the relation
(8)T A S R+ + + =1where T is the fraction of radiation transmitted through thesubstrate, R is the fraction reflected, S is the amountscattered out of the substrate, and A is the fractionabsorbed. The figure of A or T by itself is not alwaysparticularly informative, and the concept of penetrationdepth is sometimes used. This is defined as the distance, d,through a substrate where the intensity of the radiation hasfallen to l/e of its value at the surface; so
(9)II et
o=
1
where Io is the incident radiation intensity and It is theintensity of the transmitted radiation. The penetrationlength d is particularly useful when it is necessary to
consider how deep into a substrate the radiation isdeposited.
There are a number of specific chromophores thatoccur in the retina and that readily absorb light. Figure116-6 shows the spectral absorption curves of the fourthought to be most important for inducing thermal effects.
The way in which hemoglobin absorbs light withinthe eye is obvious to anyone who has used an ophthalmo-scope to examine an albino or a lightly pigmented person.The red pupillary reflex is another example of the opticalimportance of blood. The two forms of hemoglobin,oxygenated and reduced, have slightly differentabsorption curves, but both peak in the yellow portion ofthe spectrum. This property has been exploited bysurgeons who seek to make burns in the retina, and someequipment manufacturers have taken up this argument tosell lasers that produce yellow light (e.g., certain dyesystems). But it is not always realized that the curves thatare produced in sales brochures showing how wellhemoglobin absorbs in the yellow were made with pathlengths many times longer than those found in retinalvessels or choroidal capillaries (Fig. 116-16). Path lengthis an important and, so it seems, often overlookedparameter, for A is related to it by Lambert's law, which,if expressed mathematically, may be rearranged thus:
(10)A e l= − −1 β
provided there is no loss due to scatter or reflection and is the extinction coefficient for unit path length (con-stant at fixed wavelength) and l is the path length. If thepath length is short, the value of A, and thus the amountof energy absorbed and the temperatures reached, will bereduced.
The value of the concept of penetration depth cannow be put to good use. It requires no calculation torealize that if d for hemoglobin is 0.1 min in the yellow,to coagulate a blood vessel thermally it should not bemuch greater than about d in diameter or sufficient energywill not penetrate to its center. Unfortunately, if the vesseldiameter is too small the bolus nature of blood flow maymean that the hemoglobin will not be equally distributedalong the vessel and it is again equally difficult tocoagulate the blood. It is possible to achieve bloodcoagulation by increasing the irradiance, but this isusually associated with destruction of tissues surroundingthe target vessel, a situation that is not clinicallydesirable. It is, however, possible to produce hernostasis,but only if the target vessel is associated with somestructure that is rich in melanin. The heat spreads fromthe melanin into the vessel to produce coagulation-ofcourse, the melanin-containing tissues are alsodamaged.11 Larger vessels are also difficult to closebecause the blood flows fast enough to carry away any
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

17
heat formed in the hemoglobin. Stopping the flow in targetvessels by raising intraocular pressure improves theprospects for hemostasis, but it is easier to increase theirradiance, although this produces widespread damagebeyond the vessel. The larger vessels usually run in theinner retina and are overlaid by the nerve fiber layer.When the vessels are exposed to intense radiation, thenerve fibers may be severed and their ends swell and forma diffuse, fluffy white mass called a cottonwoof spot thatdevelops 12 hours or more after exposure.
Retinal thermal damage occurs only if the temperatureof the target site rises, and it is easy to see that this cannothappen unless energy is delivered to the retinais actuallyabsorbed in the tissue volume-faster than it leaks away. Atthe start of an exposure,
(11)E EABS LEAK=where E represents the energy being absorbed in, orleaking from, the tissue volume. Eventually an equilibriummay be reached where EABS = ELEAK, and a steadytemperature is established. At the end of the exposure, EABSis zero and the temperature decreases at a rate determinedby ELEAK. The time and temperature history of the tissueand its constituent molecules are generally held to be theparameters that determine how much damage is done, andthis is expressed by the Arrhenius equation
(12)Ω = ∫−C e dtC T r z
tt ci
f1
/ ( , )
where is the fraction of molecules functionally destroyedfor the temperature T between the initial time ti to the finaltime tf as a function of the radial distance r from the centerof the retinal image along the z axis of the beam of incidentradiation. Equations 11 and 12 could be the starting pointof a model that could predict what exposure intensities anddurations could lead to retinal damage.Indeed, manymodels have been based on this kind of argument, workingback through the Arrhenius equation and considerations ofEABS and ELEAK to specify ocular hazards or the likelyresults of exposing eyes to a specified light source. Mostauthors have set at 0.5 rather arbitrarily, it seems, if thearguments of Marshall41 apply. The models encountercomplexity, at least for n on-mathematicians, at the stageof calculating EABS and ELEAK which values depend not onlyon the kinds of optical transmission problems discussedearlier but also on the distribution and physical dimensionsof the chromophores that absorb the radiation. The modelsconsider only melanin, and most treat it as though it werespread in a thin, uniform layer under the retina: infinitelythin, Vos42; in a 10 m layer43-47; or 4 or 5 m layers,depending on species modeled.12 Most of these modelsincluded absorption in choroidal melanin. They have
enjoyed considerable success with longer-durationexposures (greater than 5 sec) but were perhaps wantingat very short exposure times.
Figure 116-16. A Spectral absorption curve of blood(oxygenated hemoglobin?), melanin (solution?), andxanthophyll as they appeared in advertising material forophthalmic laser instruments. Note, there is no scale onthe ordinate axis, the source or nature of the curves is notspecified, and no path lengths are given. B, A version ofA that seeks to rectify its errors. The melanin curve istaken from Gabel and coworkers17(see curve PG in Figure116-7) and represents data derived directly from RPEpreparations. The blood curve is for a path length of 10m of oxygenated blood. Neither A nor B makes anyallowance for transmission loss of preretinal media. Also,the five shaded bars are at the wavelengths of severalcommon or new ophthalmic lasers: 488-nm argon, 514-nm argon, 580-nm dye, 600-nm dye, 647-nm krypton red.
Besides considering the likely damaging effects oftemperature increases of several degrees producing tissuedamage, Vos42 proposed that steam production would beimportant for brief exposures. Eventually, it appearednecessary to redefine the models with the chromophore ingranules rather than in a homogeneous slab, because of
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

18
difficulties with fitting the models to the data from short-exposure experiments. Hansen and Fine48 proposed thatheating of a granule of melanin would produce a zone ofdenaturation around it and that this would spread withconduction of heat away from the granule source; this istheir single-mechanism model. They also proposed a two-mechanism model, in which steam would be generated atthe granule-cytoplasm interface if the energy influx weregreat enough. The heating would lead first to thermaldamage and then to steam and physical, disruptive damage.Hayes and Wolbarsht49 also developed a granule modelwith which to explain the effects of steam generation inshort exposures when power densities are usually high.They pointed out that there would be considerable amplifi-cation of lesion size (more than 1000 times) once steamwas formed and that further amplification would comefrom the ensuing biologic effects of inflammatory proc-esses.
Figure 116-17. Log retinal radiant exposure for thresholdlesions plotted against log of the diameter of the retinalimage for various exposure times. The dashed lines areleast squares fits through each set of data, and the dottedline has a slope of - 1. (Data from Sliney.22)
Examination of the models is informative, and,indeed, they are still used today in certain circumstances,but a few simple considerations might help our under-standing of thermal lesions in the retina without ourbecoming involved in calculus or numerical modeling. Theway EABS may be defined is derived directly from Beer'sand Lambert's laws and the considerations discussed
earlier, but ELEAK is more problematic.Energy may leave the volume of the absorbing site
by convection, not likely in a compartmentalized cellularstructure, or by bulk flow of the chromophore, plainly thesituation for hemoglobin in flowing blood in retinalvessels. Reradiation is not effective, as the temperaturesare too low for Stefan's law to suggest other thaninfinitesimal flow. Conduction is the process by whichmost heat flows from the absorption site. The retinalimage is usually circular and the chromophore is assumedto be in a homogeneously distributed layer, so the volumeof the absorber increases with increasing retinal imagediameter faster than the surface area of the absorber. Asheat is lost through the surface, the relative rate of heatloss drops and larger images are associated with greatertemperature increases at their centers or, put another way,the radiant exposure for threshold damage will fall. Thisis convincingly shown in Figure 116-17 for a range ofexposure durations that have slopes approximating - 1.This is what simple surface area:volume ratios wouldsuggest, as the ratio is inversely proportional to the radiusof the image.
Exposure duration is a more important determinantof the type and extent of damage produced than is retinalimage size. The concept of relaxation time50 can help,because it defines the time that the temperature falls to1/e of the initial value. It is related to the size of theabsorbing chromophore by the relation
(13)t C lr = ⋅ 2
where C is a constant (106s.m-2) and l is the smallestdimension of the heated object. For a typical melaningranule 1 m in diameter, tr is about 1 sec and for anarteriole 100 m, across, tr is 10 msec. The smallestretinal images used in the laser clinic are typically 50 min diameter, which suggests values for tr of about 2.5msec. But the layer of melanin is only about 5 m thick,which means tr will be approximately 25 sec.Consequently, if exposure times are greater than aboutthis value, considerable heat will flow from the heatedtarget chromophore to thermally damage the surroundingtissues. If laser surgical intervention is necessary in ornear the fovea, prudence suggests that retinal image sizeshould be kept small, but unless exposure time is alsokept short there will be considerable heat flow, andperhaps greater vision loss than is intended.
The drive to shorter exposures should not be pushedtoo far, because, as the granule models of thermal damageexplain and experiment shows, steam generation canbecome a problem with exposures of a few microsecondsor less. Explosive generation of steam producesmechanical transient51 that can tear tissues and evenrupture blood vessels. If the exposure duration is very
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

19
short (e.g., a few nanoseconds from a Q-switched laser),the risk of hemorrhage is great. The ratio of the energy thatjust produces a hemorrhage to the energy for a justophthalmoscopically visible retinal lesion falls fromseveral hundred, with exposure durations of milliseconds,to near unity for exposures of a few nanoseconds (Fig.116-18). This change can be interpreted to mean that, asthe exposures get shorter the energy absorbed istransformed not only into heat but also into othermodalities, such as mechanical forces.
The anomalies with the models of retinal light damageat short exposure times were referred to earlier. Themodels predict that there will be a relation between energyentering the eye, Ei, and exposure duration, te for thresholdretinal lesions of the form
(14)E m ti en= ⋅
where m and n are constants and n is approximately 0.75. Fig. 116-19 shows this relation and some experi-mental data points. It is apparent that the relation holds forvalues of te> 20 sec up to 10 sec, but that for exposuretimes less than that which corresponds to the thermalrelaxation time of the RPE layer, the relation no longerholds. Below this thermal relaxation time, energy wouldnot be expected to be lost from the absorbing chromophoreand the energy input for threshold damage should beconstant (i.e., the line in the figure should be horizontal).Obviously, the data points show a rise in threshold thatpeaks at durations of about 1 nsec. Sliney22 suggests thatthis effect may be due to the conversion of the incidentenergy into mechanical energy (compression waves) orthat the melanin acts as a saturable absorber, bleachingwhen the incident irradiance is too high and so lettingsome of the energy that would otherwise be expected to beabsorbed pass through the target.
From the foregoing, it is clear that short exposures ofaround 100 nsec to 10 sec are shorter than the thermalrelaxation times of the RPE (but not of its individualgranules) and thus on a gross scale will not cause heat flowinto surrounding structures. Neither are these times shortenough to lead to the mechanical problems that threatenexposures of a few nanoseconds or less. Consequently,they should be useful for limiting damage spread. If vitaldyes or stains tied to immunohistochemically targeted cellsor cell organelles could be administered to retinas, short-duration exposure to radiation matched to the absorptionproperties of the dye would allow selective destruction ofthese components.
Figure 116-18. Ratio of the energy threshold for retinalhemorrhage to that required for a just visible thermallesion (HIT) for two different pulse duration domains-nanoseconds (Q-switched laser) and milliseconds ("free-running" lasers). (Based on data from Wolbarsht andAllen73 and other sources.)
Figure 116-19. Log of the energy entering the eye thatcauses threshold thermal damage plotted against logexposure duration. The line has a slope of 0.75 and is aby-eye fit. (Data from Sliney.22)
If the models describe long exposures well and veryshort exposures less well, there remains the problem ofrepeated exposure. Many light sources, lasers in partic-ular, can be driven very effectively at high repetitionrates; if the interpulse interval is less than the thermalrelaxation time of the target chromophore there will be
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

20
accumulation of energy in the target, and more damagethan expected will be produced. Put another way, theenergy in each pulse of a train that just produces damagewill be less than the threshold energy for a single pulse.Empirically it has been found that the energy content, Qs,of a pulse in a train is reduced for threshold so
(15)Q Ns ∝ −0 25.
where N is the number of pulses in the train. Sliney22
describes how this relation applies over a wide range ofpulse durations and repetition frequencies and to all but thelargest image sizes. He argues that this is due to theinterruption of a rapid repair process by the second andsubsequent pulses. Examination of the times over whichthe relation holds suggests that the repair process muststart within microseconds of the first exposure and becompleted by about 1 second.
Figure 116-8 shows the spectrum of damage types,and for completeness thermoacoustic effects must bedescribed. With sufficiently high retinal irradiance theexplosive formation of steam and the generation ofmechanical transients have already been mentioned. Theimportance of the mechanical transients lies in the waythey trap some of the incident energy and carry it awayfrom the site of the retinal image, perhaps over severalmillimeters. Similar but less intense mechanical energy canbe generated without steam formation, owing to the rapidexpansion of heated tissues that occurs with short(nanosecond-length) pulses.
By whichever mechanism the forces are generated(see under Photodisruption Damage), they promise damageof a different form to thermal denaturation and possibly atsome distance for the retinal image. Marshall andMellerio52 showed that Q-switched laser pulses induceddamage with mechanical displacement of cell constituentsoutside the image area, exactly as expected from theforegoing analysis. From their histologic examinationsthey were not able to say how much of the damage was dueto the passage of the mechanical compression waves, theactual steam bubble surface, or the blood flow inhemorrhages that were often induced.53
Photodisruption Damage
As Figure 116-8 shows, when light exposure times arevery short (nanoseconds and less), if enough energy ispresent to produce damage, this will be due to ionizationand is usually called photodisruption. This ionizationcomes about because the strength of the electromagneticfield is so great that electrons are stripped from the outerorbitals of the atoms. Inside the eye this happens onlywhen the radiation originates from Q-switched or mode-
locked lasers and the beam is focused into a small volumeeither by accessory optical power of an ophthalmicinstrument or contact lens or by the natural refractingpower of the eye.
Photodisruption has found considerable use in recentyears in ophthalmology, principally in the procedures ofiridotomy to improve drainage of the posterior chamberand in posterior capsulotomy after implantation of anintraocular lens. In these and other procedures, theionized tissue forms a plasma as the molecules areliterally torn apart, thus enabling very small lesions to be
made-an attribute that inspired the name "lasermicrosurgery”. Further, it can be done inside the eyewithout opening the globe. Laser iridotomy and capsu-lotomy are both processes inflicted on anterior structures,but laser photodisruption has been used farther back inthe eye. The problem of causing secondary damage to theretina when, for example, attempting to part a vitrealmembrane, is very real and cannot be ignored.54
Photodisruption might be considered a very goodtechnique for removing epiretinal membranes because ofthe precise nature of the lesions formed, but there hasbeen some concern that damage might be extendedbeyond the target site by mechanical processes (asdescribed below55) or because the plasma is not generatedexactly where expected but is displaced.` Because theincident beam has to be focused at the target site toensure power density great enough to cause a plasma, thegeometry of the beam in the focal area is an importantdeterminant of whether and where a plasma is formed.56
The cone angle of the focused beam specifies beamexpansion beyond the focal region and thus determinesthe irradiance at the retina, which could be hazardous ifthe focus is too close to the retina.57
Taking values for retinal irradiance for thresholddamage from the literature, Capon and coworkers58
calculated how near the retina typical ophthalmic pho-todisruptor lasers can be focused before they becomeunsafe with a retinal irradiance that might cause a lesion
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

21
(Table 116-3). The figures in Table 116-3 are probablyoptimistic, because they were derived for a minimalophthalmoscopically visible retinal lesion, but fluores-cence angiographic and electron microscopic techniquesare more sensitive at tracing retinal damage.59 If thresholdsdetermined by these techniques are used, the minimumdistances in Table 116-3 would probably need to bedoubled. Consequently, a surgeon should use as high acone angle as possible and, even with the minimumenergy, must still avoid working within several millimetersof the retina unless he is prepared to accept the possibilityof damage there.
The mechanisms of plasma formation are describedelsewhere,60 but they bear on the situation with the retinabecause the formation of a plasma in a liquid tends tooccur at the site of an impurity or an interface.56, 61
Consequently, if a Q-switched laser beam is brought to afocus in the vicinity of the retina, the inner surface of theretina may well be the site of plasma formation. Becausethe tissues at the plasma are vaporized the damage isconsiderable and the risk of hemorrhage very great. Evenif the plasma is formed deeper in the retina or even in thechoroid, because the focal volume of tissue is completelyremoved hemorrhage is very likely, and this could involvethe vitreous (Fig. 116-20).62 Visual function would becompromised, either by direct damage from the plasma tothe neural elements or by the obscuring and delayedimmunoreaction of the hemorrhage itself.
A plasma formed in a liquid environment, as in theeye, once initiated, loses heat (energy) very rapidly and isextinguished unless there is a sustaining inflow of energy.This is normally provided by the incident laser beam itself,so the plasma lasts only slightly longer than the laser pulse.Plasma maintenance absorbs energy from the laserradiation; in the early days of the development ofphotodisruption it was held that this absorption by theplasma would remove any hazard of radiation exposure tostructures downstream from the plasma. This plasmashielding was investigated by several authors63, 64 inconditions that approximate those in the eye, though overa range of energies that would be much higher than thosenormally used in the clinic.65 The shielding was found tobe variable and not to offer a very great safety factor atlow-energy inputs of the level that are used in the clinic foriridotomy and capsulotomy.58 It was estimates of theprobable amount of plasma shielding that would arise inthe attempt to section vitreal membranes that led to thefigures shown in Table 116-3.
Figure 116-20. Diagram of different types of retinalhemorrhages that are produced by Q-switched or mode-locked laser pulses (blood if shown in solid black). 1,subretinal; 2, intraretinal and subhyaloid; 3, subretinaland intraretinal; 4, vitreous hemorrhage with a blood stalktrapped in the vitreous; 5, vitreous hemorrhage where theblood has mixed with the vitreous.
The generation of a laser-induced plasma by a Q--switched or a mode-locked laser leads to such extremelyrapid expansion of the heated plasma60 that a compressionor shock wave is created that propagates away from theplasma throughout the eye. This wave is of concernbecause it carries energy to ocular locations remote fromthe initial site of exposure. The exact relationship be-tween the magnitude of the shock wave and damage totissues is one that still awaits elucidation, although it iswell established in tissues other than the eye that there isa link between the two.66-68 The bubble of heated gas leftby the extinguished plasma expands for severalmilliseconds, and must damage surrounding tissues,55
before collapsing destructively69 or oscillating to releasemore energy in bubble pulses before the final destructivecollapse. Should these cavitation bubbles be formed at ornear the retina there is little doubt that considerabledamage would be done by pressure waves and variousshearing forces generated by jets and similar phenomena,in addition to the removal of tissue by the plasma itself.Indeed, it is hard to think of any therapeutic process thatcould be executed safely in or near the retina byphotodisruptive methods because of the great risk ofcausing considerable tissue damage and massive
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

22
hemorrhage. Such exposure will probably be experiencedonly as a result of accident in industry or research, or ofintent from battlefield laser weapons,62 when the prognosisfor full recovery of vision may not be good.
CONCLUSION
The study of the effects of light on the retina coversmechanisms that correspond to situations only slightlymore intense than considered normal, up to franklytraumatic insults. Besides being fascinating, the study haspractical implications, just as investigation of repetitivestrain injury or of bone union in Colles' fracture has for atypist. Investigation of the processes that cause damage asa result of a lifetime's exposure to daylight, or to anindustrial laser, have already borne fruit. Codes of practicefor safe exposure to light sources now incorporate theknowledge derived from these investigations, andophthalmic surgeons purposely harness damagingprocesses to manipulate tissue relationships. As ourunderstanding of the way light interacts with tissuesincreases, we may look forward to better protection andmore effective surgical intervention.
REFERENCES
1. Coohill TP: Action spectra revisited. J Photochem Photobiol [B]13:95, 1992.
2. Sarna T: Properties and function of ocular melanin-a photobio-logical review. J Photochem Photobiol 12:215, 1992.
3. Mainster MA: Spectra, classification and rationale of ultravioletprotective intraocular lenses. Am J Ophthalmol 102:727. 1986.
4. Weale RA: Effects of the Ageing Lens on Vision. CIBA Symposium No. 19. London, Elsevier, 1973.
5. Ijspeert J, Dewaard P, van der Berg T, et al: Intritocular stray lightfunction in 129 healthy volunteers: Dependence on angle. age andpigmentation. Vision Res 30:699, 1990.
6. Weale RA: Human lenticular fluorescence and transmissivity andtheir effects on vision. Exp Eye Res 41:457, 1985.
7. Werner JS: Development of scotopic sensitivity and the absorptionspectrum of human ocular media. J Opt Soc Am 72:247, 1982
8. Geeraets WJ, Berry ER: Ocular spectral characteristics as relatedto hazards from lasers and other light sources. Am J Ophthalmol66:15, 1968.
9. Harwerth RG, Sperling HG: Prolonged color blindness induced byintense spectral lights in rhesus monkeys. Science 174:520, 1971.
10. Harwerth RG, Sperling HG: Effects of intense visible radiation onthe increment threshold spectral sensitivity of the rhesus monkey.Vision Res 15:1193, 1975.
11. Fankhauser F, Kwasniewska S, van der Zypen E: Basic mecha-nisms underlying laser thrombogenesis in vascular structures of theeye. Lasers Light Ophthalmol 2:223. 1989.
12. Birngruber R, Hillenkamp F. Gabel VP: Theoretical investigationsof laser thermal retinal injury. Health Phys 48:781. 1985.
13. Boulton M, Docchio F, Dayhaw-Barker P, et al: Age-relatedchanges in the morphology, absorption and fluorescence of mel-anosomes and lipofuscin granules of the retina and pigmentepithelium. Vision Res 30:1291, 1990.
14. Sarna T, SeAy RC: Photoinduced oxygen consumption in melanin
systems: Action spectra and quantum yields for eumelanin andsynthetic melanins. Photochem Photobiol 39:69, 1984.
15. Latimer P: Deconvolution of absorption spectra of green plantmaterials-improved corrections for the sieve effect. PhotochemPhotobiol 38:731, 1983.
16. Kollias N, Sayre RM, Zeise L, et al: Photoprotection by melanin.J Photochem Photobiol [B] 9:135, 1991.
17. Gabel VP, Birngruber R, Hillenkamp F: Visible and near-infraredlight absorption in pigment epithelium and choroid. In Interna-tional Congress Series No. 450, XXIII, ConciliumOphthalmologicum, Kyoto. Amsterdam, Elsevier, 1978.
18. Jukes T: Early development of the neutral theory. Perspect BiolMed 34:473, 1991.
19. Halliwell B, Gutteridge JM: Free Radicals in Biology and Medi-cine. Oxford, Oxford University Press, 1986.
20. Pooler JP, Valenzeno DP: Dye-sensitized photodynamicinactivation of cells. Med Phys 8:614, 1981.
21. Lingua RW, Parel JM: Photodynamic therapy for ocular tumors.J Photochem Photobiol [B) 9:119, 1991.
22. Sliney DH: Interaction mechanisms of laser radiation with oculartissues. In Bennett HE, Guenther AE, Milan D, et al (eds): Laser-Induced Damage in Optical Materials. Washington, DC, NBSPublications, 1984.
23. Dillon J: Photophysics and photobiology of the eye. J PhotochemPhotobiol 1131 10:23, 1991.
24. Dayhaw-Barker P, Barker FM II: Phototoxicity: The problem ofdrug and chemical enhancement. In Cronly-Dillon JR, MarshallJ (eds): Vision and Visual Dysfunction, Vol 16. London. Mac-millan, 1991.
25. Marshall J, Mellerio J, Palmer DA: Damage to pigeon retinae bymoderate illumination from fluorescent lamps. Exp Eye Res14:164, 1972.
26. Ham WT, Mueller HA, Ruffolo JJ, et al: Basic mechanismunderlying production of photochemical lesion in the mammalianretina. Curr Eye Res 3:165, 1984.
27. Mellerio J: Interaction of light with biological tissue and thepotential for damage. In Cronly-Dillon JR, Marshall J (eds):Vision and Visual Dysfunction, Vol 16. London, Macmillan,1991.
28. Noell WK, Walker VS, Kang BS, et al: Retinal damage by lightin rats. Invest Ophthalmol Vis Sci 5:450, l966
29. Arden GB, Berninger T, Hogg CR, et al: A survey of colourdiscrimination in German ophthalmologists: Changes associatedwith the use of lasers and operating microscope. Ophthalmology98:567, 1991.
30. Lawill T: Effects of prolonged exposure of rabbit retina to lowintensity light. Invest Ophthalmol Vis Sci 12:45, 1973.
31. Rapp LM, Tolman BL, Dhindsa HS: Separate mechanisms forretinal damage by ultraviolet A and mid-visible light. InvestOphthalmol Vis Sci 31:1186, 1990.
32. van Norren D, Schellekens P: Blue light hazard in rat. Vision Res30:1517, 1990.33. Ham WT, Ruffolo JJ, Mueller HA, et al: Histologic analysis of
photochemical lesion produced in rhesus retina by short wave-length light. Invest Ophthalmol Vis Sci 17:1029, 1978.
34. Ham WT, Mueller HA, Sliney DH: Retinal sensitivity to damagefrom short wavelength light. Nature (Lond) 260:153, 1976.35. Pautler EL, Morita M, Beezley D: Hemoproteins mediate blue
light damage in the retinal pigment epithelium. Photochem Pho-tobiol 51:599, 1990.
36. Kremers J.1M, van Norren D: Two classes of photochemicaldamage of the retina. Lasers Light Ophthalmol 2:41, 1988.
37. Kremers JJM, van Norren D: Retinal damage in macaque afterwhite light exposure lasting 10 minutes to 12 hours. InvestOphthalmol Vis Sci 30:1032, 1989.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com

23
38. Williams TP, Howell WL: Action spectrum of retinal light damagein albino rats. Invest Ophthalmol Vis Sci 24:285, 1983.
39. van Norren D: Photochemical damage to the eye. News Physiol Sci6:232, 1991.
40. Sliney DH: New chromophores for ophthalmic laser surgery.Lasers Light Ophthalmol 2:53, 1988.
41. Marshall J: Lasers in ophthalmology: the basic principles. Eye 2(Suppl):S98. 1988.
42. Vos JJ: A theory of retinal burns. Bull Math Biophys 24:115,1962.
43. Clarke AM, Geeraets WJ, Ham WT: Equilibrium thermal modelfor retinal injury from optical sources. Appl Opt 8:1051, 1969.
44. Mainster MA, White TJ, Tips JH, et al: Transient thermalbehaviour in biological systems. Bull Math Biophys 32:303, 1970.
45. Mainster MA, White U, Tips JH, et al: Retinal temperatureincreases produced by intense light sources. J Opt Soc Am 60:264,1970.
46. White TJ, Mainster MA, Tipps JH, et al: Choroidal thermalbehaviour. Bull Math Biophys 32:315, 1970.
47. Roulier A: Calculations of temperature increase in the eyeproduced by intense light. Bull Math Biophys 32:403, 1970.
48. Hansen WP, Fine S: Melanin granule models for pulsed laserinduced retinal injury. Appi Opt 7:155, 1968.
49. Hayes JR, Wolbarsht ML: Thermal model for retinal damageinduced by pulsed lasers. Aerospace Med 39:474, 1968.
50. Hillenkamp F: Laser radiation tissue interaction. Health Phys56:613, 1989.
51. Mellerio J: Laser light and its effect on the retina. TransOphthalmol Soc UK 87:335, 1967.
52. Marshall J, Mellerio J: Pathological development of retinal laserphotocoagulation. Exp Eye Res 6:303, 1967.
53. Marshall J, Mellerio J: Histology of retinal lesions produced withQ-switched lasers. Exp Eye Res 7:225, 1968.
54. Tassignon MJ, Stempels N, Brihaye M, et al: Vitreo-retinalmembranes treated with the Q-switched Nd:YAG laser: A clinicalstudy. Lasers Light Ophthalmol 1:205, 1987.
55. Mellerio J, Capon MRC, Docchio F: Nd:YAG lasers-a potentialhazard for cavitation bubble behaviour. in anterior chamber pro-cedures? Lasers Light Ophthalmol 1:185, 1987.
56. Capon MRC, Docchio F, Mellerio J: Investigation of multipleplasmas from ophthalmic Nd:YAG lasers. Lasers Light Ophtbal-mol 1:147, 1987.
57. Bonner RF, Meyers SM, Gaasteriand DE: Thresholds for retinaldamage associated with the use of high power Nd:YAG lasers inthe vitreous. Am J Ophthalmol 96:153, 1983.
58. Capon MRC, Docchio F, Mellerio J: Nd:YAG laser photodisrup-tion: An experimental investigation on shielding and multipleplasma formation. Graefes Arch Clin Exp Ophthalmol 226:362,1988.
59. Boriand RG, Brennen DH, Marshall J. et al: Role of fluoresceinangiography in the detection of laser-induced damage to the retina:A threshold study for Q-switched neodymium and ruby lasers. ExpEye Res 27:471, 1978.
60. Docchio F, Regondi P, Capon MRC, Mellerio J: Study of thetemporal and spatial dynamics of plasmas induced in liquids bynanosecond Nd:YAG laser pulses: 1. Analysis of the plasmastarting times. Appl Opt 27:3661, 1988.
61. Docchio F, Dossi L, Sacchi CA: 0-switched Nd:YAG irradiationof the eye and related phenomena: 1. Optical breakdown deter-mination for liquids and membranes. Lasers Life Sci 1:87, 1986.
62. Mellerio J. Marshall J, Tengroth B, et al: Battlefield laser weapons:an assessment of systems, hazards, injuries and ophthalmicresources required for treatment. Lasers Light Ophthalmol 4:41,1991.
63. Loertscher H: Laser induced breakdown for ophthalmic applica-
tions. In Trokel SL (ed): YAG Laser Ophthalmic Microsurgery.Norwalk, CT, Appleton-Century-Crofts, 1983.
64. Steinert RF, Puliafito CA, Kittrel C: Plasma formation andshielding by three ophthalmic Nd:YAG lasers. Am J Ophthalmot96:427, 1983.
65. Steinert RF, Puliafito CA, Kittrel C: Plasma shielding by Q-switched and mode locked Nd:YAG lasers. Ophthalmology90:1003, 1983.
66. Ara G. Anderson RR, Mandel KG, et al: Irradiation of pigmentedmelanoma cells with high intensity pulsed radiation generatesacoustic waves and kills cells. Lasers Surg Med 10:52, 1990.
67. Suneson A, Hansson HA, Lycke E, et al: Pressure wave injuriesto rat dorsal root ganglion cells in cultures by high energymissiles. J Trauma 29:10, 1989.
68. Yashima Y, McAuliffe DJ, Flotte TJ: Cell sensitivity to laserinduced photoacoustic injury of the skin. Lasers Surg Med10:280, 1990.
69. Vogel A, Lauterborn W, Timms R: Optical and acoustical inves-tigation of the dynamics of laser-produced cavitations near asolid boundary. J Fluid Mech 206: 299, 1989.
70. Sliney DH, Wolbarsht M: Safety with lasers and other opticalsources. New York, Plenum Press, 1980.
71. Maher EF: Transmission and Absorption Coefficients for OcularMedia of the rhesus Monkey. Report SAM-TR-78-32, USAFSchool of Aerospace Medicine. Brooks Air Force Base, Texas,1978.
72. Mellerio J: Yellowing of the lens-nuclear and cortical contribu-tions. Vision Res 27:1581, 1987.
73. Wolbarsht M, Alien R: New laser device for ophthalmology: Acomment. Appl Opt 25:1533, 1986.
PDF created with FinePrint pdfFactory trial version http://www.fineprint.com





























