Embed Size (px)
Citation preview

Anim Cogn (2008) 11:255–266
DOI 10.1007/s10071-007-0107-2ORIGINAL PAPER
Can brown lemurs (Eulemur fulvus) learn to deceive a human competitor?
Emilie Genty · Jane Foltz · Jean-Jacques Roeder
Received: 25 August 2006 / Revised: 7 August 2007 / Accepted: 29 August 2007 / Published online: 18 September 2007© Springer-Verlag 2007
Abstract In the present study we asked whether lemurscould learn to manipulate information in order to deceive ahuman competitive trainer. Four brown lemurs were trainedto communicate about the location of a hidden reward to acooperative trainer, who rewarded the subject if he indi-cated the baited bowl. Next, a competitive trainer was intro-duced who kept the reward for himself if the subjectindicated the baited bowl. In a Wrst experiment, sessionswere randomly assigned to be with either the cooperative orcompetitive trainer. No subject was able to show aneYcient tactic with both trainers. In a second experiment,the participation of the two trainers was randomized acrossthe trials for each session. When trials were mixed, onesubject signiWcantly chose baited location when interactingwith the cooperative trainer, and reliably increased hischoices of the unbaited location when presented with thecompetitive trainer. As with most other primate speciestested under the same paradigm, associative learning mayexplain deceptive pointing by lemurs in this study.
Keywords Lemurs · Cognition · Deception · Cooperation · Competition
Introduction
Deception is widespread in the animal kingdom and doesnot appear to be a uniquely human ability. Recent studieshave shown that 2½-year-old children may learn to lie atthe same time they learn to speak (Newton et al. 2000), butit is generally admitted that intentional deception in humanchildren is infrequent until 4 years of age (La Frenière1988; Russell et al. 1991; Sodian 1991). Children underfour Wnd it diYcult not to indicate the container covering adesired item to a “competitor” even though they know thatthe competitor would keep the item. However, this diY-culty may correspond to an inability to inhibit their gesturetoward the baited container, rather than failure to under-stand false belief (Carlson et al. 1998).
Withholding or conveying incorrect information to othershas been recorded in a wide variety of species includinginsects, birds, carnivores, elephants and primates (Mitchelland Thompson 1986). Although cognitive ethologists andpsychologists generally consider deception to imply inten-tionality and awareness of another’s awareness on the partof the deceiver, deception is not always intentional and doesnot necessarily require advanced cognitive abilities (Coussi-Korbel 1994). Mitchell (1986) has distinguished four levelsof deception, each one implying diVerent cognitive abilities.In level one, deception is eVected by appearance (e.g. plantswhich mimic); in level two, deception is achieved by coor-dination of perception and action (e.g. birds which feigninjury); in level three, deception is achieved by learning;and in level four, deception is eVected by planning (e.g.chimps which mislead about food location). Nevertheless itremains diYcult to identify the intentions of animals. Toavoid such diYculties, Byrne and Whiten (1985) introducedthe term of “tactical deception”. Tactical deception wasdeWned as “acts from the normal repertoire of an individual,
E. Genty · J. Foltz · J.-J. RoederIPHC, DEPE, CNRS, 23 rue Becquerel, 67000 Strasbourg, Francee-mail: [email protected]
J. Foltze-mail: [email protected]
J.-J. Roeder (&)DEPE UMR 7178, 23 rue Becquerel, 67000 Strasbourg, Francee-mail: [email protected]
123

256 Anim Cogn (2008) 11:255–266
used at low frequency and in contexts diVerent from those inwhich it uses the high frequency (honest) version of the act,such that another familiar individual is likely to misinterpretwhat the acts signify, to the advantage of the actor” (p. 672).The Wrst evidence of tactical deception comes from Weldobservations of primates (Whiten and Byrne 1988; Byrneand Whiten 1992). However, such anecdotal observationswere not suYcient to assess deception, and experimentalsettings are also necessary to clearly reveal the presence ofintentional/tactical deception in primates (e.g. Andersonet al. 2001; Coussi-Korbel 1994; Ducoing and Thierry 2003;Menzel 1974; Mitchell and Anderson 1997; WoodruV andPremack 1979). One method that has been extensively usedin several primate species is one in which the subject mustdeceive an experimenter as to the location of a hiddenreward. WoodruV and Premack (1979) trained four chim-panzees to point to a box containing food to a “cooperative”trainer. After several sessions in which a “competitive”trainer ate the food when he found it in the box, two of thesubjects started to point to the empty box when the competi-tive trainer approached. DiVerential pointing has also beendemonstrated in tufted capuchins (Mitchell and Anderson1997) and in squirrel monkeys (Anderson et al. 2001).Important questions emerged from both naturalistic andexperimental observations: do the deceptive behavioursobserved in the wild imply speciWc cognitive abilities anddo the results of experimental studies really reXect inten-tionality as opposed to conditional discrimination? Further-more, these previous studies only concerned great apes ormonkeys, and it remains to be understood if the absence ofrecords of some taxa/species is due to a lack of interest insuch species or to real diVerences in cognitive abilities(Whiten and Byrne 1988).
No example of tactical deception in lemurs was men-tioned in Whiten and Byrne’s paper (1988). As it is gener-ally assumed, intelligence has emerged from social andenvironmental constraints linked to access to resources(Byrne and Whiten 1992). Most lemur species live in com-plex social groups (multi-male/multi-female) in unstableenvironments (see Kappeler and Ganzhorn 1993a, b) andtherefore face similar social and environmental constraintsto simians. It would not be surprising, then, to Wnd compa-rable cognitive skills in lemurs and monkeys. In brownlemurs (Eulemur fulvus), group size ranges from 2 to 29individuals including several adult males and adult females(Ganzhorn 1988; Harrington 1975; Tattersall 1982). Con-trary to other lemur species, like the ring-tailed lemur(Lemur catta: Jolly 1966; Pereira et al. 1990) and the blacklemur (Eulemur macaco: Fornasieri et al. 1993; Pereira andKappeler 1997), which are characterized by female domi-nance, in brown lemurs dominance is not related to genderand dominance relations are weak (Roeder and Fornasieri1995; Pereira and Kappeler 1997). Therefore, lemurs could
oVer a valuable model for testing how social constraintsmay inXuence cognitive abilities.
Only a few studies have focused on deception in prosi-mians. One unpublished experimental study recordeddeception in Lemur catta in semi-naturalistic conditions(Deaner 2000) using Menzel’s (1974) methodology. A sub-ordinate male was informed of the location of a hidden fooditem and was subsequently released into the enclosuresimultaneously with an uninformed dominant female. Malering-tailed lemurs did not reliably deceive the dominantfemale. These results did not provide evidence for decep-tion in lemurs and therefore supported the hypothesis ofcognitive diVerences between prosimians and simians.However, in a previous study with black lemurs (Eulemurmacaco), Genty and Roeder (2006) have shown that thisspecies, after having been trained to communicate about thelocation of a hidden reward to a cooperative trainer, showedvarious behavioural adjustments when faced with a com-petitive trainer. Such behavioural adjustments involved“participation refusal” and “information withholding”.Deceptive pointing occurred rarely and was only used bythe subject over a short period. Therefore, the possibility ofdeception in lemurs remains questionable and additionalstudies are necessary to increase our knowledge of lemurs’deceptive ability, and more generally, of their cognitivecapacities.
In this paper, we analyse whether brown lemurs (Eulemurfulvus) behave in the same way as black lemurs or simian spe-cies when presented with a competitive trainer, and whetherthey learn to mislead this trainer. In other words: wouldbrown lemurs trained to point to a baited box reverse thisresponse when conditions/trainers change? Would they usecooperative or deceptive signalling according to the context?
Experiment 1
Methods
Subjects
The subjects were four brown lemurs (Eulemur fulvus),three adult females (Hy, Ho and Hi), and one adult male(Hu). All were born at the Centre de Primatologie in Stras-bourg, France. They were housed in spacious cages com-prising two compartments: an inside compartment(4.9 £ 2 m and 2.6 m high) maintained at 20°C, and an out-door compartment (8 £ 2.9 m and 2.6 m high). A tunnellinking these two compartments could be closed to facili-tate isolation of subjects for testing. Outside of test ses-sions, the lemurs could move freely between the indoor andoutdoor areas. The cages were furnished with tree trunks,ropes and shelters. The outdoor area contained natural
123

Anim Cogn (2008) 11:255–266 257
grasses and several bushes. The lemurs received commer-cial food pellets each day and fresh fruits and vegetablesonce a week, outside of experimentation and observationperiods. The lemurs were not food deprived for testing.
Procedure
The subjects were used to separation from the group, as allhad been previously trained to enter the inside compartmentof the cage where testing took place. They were given oneweek to habituate to the test apparatus and the experi-menter. Preliminary trials established that raisins were ahighly attractive food for all subjects.
An initial training period was conducted to train thelemurs to reliably indicate (by pointing through the cagemesh) to a cooperative trainer the location of a raisin hid-den under one of two bowls (by an experimenter), withouttrying to grab the bowls. Competence in pointing was deW-ned as the lemur pointing toward the baited bowl in at leasteight of ten trials over at least Wve consecutive sessions.
Two diVerently coloured plastic bowls, each placed atone end of a tray (60 £ 17 £ 30 cm), were used to hide therewards. The tray was placed out of reach of the subject onthe exterior side of the mesh of the subjects’ cage. Theexperimenter used a stopwatch to record the time taken bythe lemur to indicate the location of the bait.
At the start of each trial, the experimenter stood 3 mfrom the cage and a trainer sat facing away on a chairplaced approximately 2 m away from the cage. The experi-menter approached the cage, talked softly to the lemur andshowed it the raisin on the tray top. The experimenter thencovered the raisin with one of the bowls, ensuring that thesubject paid attention (the subject had to stay in front of theapparatus and look at both the rewarded and the unbaitedlocation). The experimenter then lifted the unbaited bowl,then the baited bowl, and replaced both to their originalpositions. Both bowls were lifted and replaced once more.The baited bowl was then lifted again for 1–2 s so that thelemur could again see the raisin. The experimenter thenreturned to his starting place from where he was able tonote the subject’s behaviour, and started the stopwatch. Thelemur typically sat in front of the baited bowl, but its bodyposition was not used by the trainer as a cue to the locationof the reward: only pointing was taken into account by thetrainer to decide which bowl to lift up. The trainer, unawareof which bowl was baited, approached the tray, placed oneWnger equidistant between the two bowls, and used thepointing behaviour of the lemur to decide which bowl to liftup. If the lemur did not point within 60 s, the trial wasstopped and a new one was started. During the cooperativecondition, if the bowl indicated by the lemur was baited, thetrainer congratulated the lemur and gave it the raisin. If thebowl that the lemur pointed to was unbaited, the trainer
expressed regret, lifted the two bowls simultaneously, andtook the raisin away.
In the second phase a competitive trainer was intro-duced. The procedure for competitive trials was the same asthat of the training period, except that when the lemur indi-cated the baited bowl the competitive trainer said loudly (inFrench), “Yes, it’s mine, now I am going to eat it!” and pre-tended to eat the raisin after overturning the selected bowlon the tray, and then left. If the lemur indicated the unbaitedbowl, the competitive trainer said “Oh, no!” returned thebowl to the tray and left. At this point the experimenterapproached, congratulated the lemur, lifted the second bowland gave the lemur the raisin.
The two trainers dressed distinctively. The cooperativetrainer wore a white gown; the competitive trainer wore adark jacket, a cap and sunglasses. Additionally, the cooper-ative and competitive trainers were consistently played bythe same person. Cooperative and competitive sessionswere randomly conducted with the constraint that neitheroccurred more than three consecutive times. Right and leftlocations of the bait were randomly distributed across trials.
A session consisted of ten trials, with one session perday. A session had to be completed in a maximum of30 min otherwise it was stopped and the remaining trialswere recorded as “uncompleted trials” (previous observa-tion have shown that after that time, the lemurs would notparticipate at all). Inter-trial intervals were approximately10 s. All lemurs received the same type of session (cooper-ative or competitive) on the same day. Each lemur receiveda total of 27 cooperative and 27 competitive sessions.Figure 1 gives an overview of the general set up of theexperiment.
The diVerent recorded behaviours were readily categor-ised (Table 1). Before pointing, the subject typically movedand sat at one end of the apparatus; then he extended onehand or forearm through the wire-mesh to select a bowl(without trying to grasp it). When withholding information,the lemur sat in front of the experimenter, looked at theapparatus and stayed in this location until the trainerarrived, but did not point in front of the trainer. When par-ticipation was refused, the subject did not approach theapparatus and generally moved toward the access tunnel atthe opposite side of the testing area. For a trial to be consid-ered as “achieved”, the experimenter and the trainer had toagree about the subject’s response. This occurred in morethan 99% of all cases (number of achieved trials = 2,761/total number of trials = 2,772). If the subject’s responsewas not clear, the trial was re-run.
Results
The subjects had to reach a criterion of 80% in choosing thebaited bowl during Wve consecutive sessions before enter-
123

258 Anim Cogn (2008) 11:255–266
ing the second phase of study. In other words, the subjecthad to point toward the baited bowl in the presence of thecooperative trainer in at least eight trials in a 10-trial ses-sion during Wve consecutive sessions. All the subjects hadpreviously been trained to point toward a bowl under whicha raisin was placed in full view by the experimenter.
When the cooperative trainer was introduced, Hu and Hyscored 80% correct in the Wrst session and maintained thislevel of performance during Wve consecutive sessions. Hireached the criterion after 7 sessions and Ho after 12 ses-sions. All the subjects received the same number of ses-sions (12). After these 12 cooperative sessions, thecompetitive trainer was introduced and 27 sessions wereconducted with both trainers.
Participation rate
All the subjects participated in all the 27 sessions with thecooperative trainer (Fig. 2). Hy and Ho participated in allthe competitive sessions (Fig. 2). Hi did not participate dur-ing the Wnal session and Hu completed fewer than Wve trialsin seven sessions. Although Hu continued to participatefully in cooperative sessions, his participation declined incompetitive sessions. The decrease in Hu’s participationappeared to be due to the presence of the competitivetrainer and the lack of reward associated with this trainer.
For further analysis, only those sessions in which at leastWve out of the ten scheduled trials were completed were
considered to avoid any biased result. Therefore, the analy-sis includes only 26 sessions (out of the 27 conducted) forHi and 20 sessions for Hu.
Performance and behavioural analysis
To analyse the performance and behaviour of each subject inthe two contexts (cooperative and competitive), we assessedthe percentage of choices of the baited bowl and the occur-rence of other possible behaviours (see terminology: Table 1).
During cooperative and competitive sessions, choices of thebaited bowl remained signiWcantly more frequent than theother behaviours (selection of the unbaited bowl and informa-tion withholding) (Binomial test: Cooperative session: Hy,z = ¡12.48, P < 0.001; Ho, z = ¡9.80, P < 0.001; Hi,z = ¡9.11, P < 0.001; Hu, z = ¡13.65, P < 0.001; Competitivesessions: Hy, z = ¡4.32, P < 0.001; Ho, z = ¡6.39, P < 0.001;Hi, z = ¡2.42, P < 0.05; Hu, z = ¡4.59, P < 0.001) (Table 2).
Nevertheless, the four subjects presented distinct behav-iours according to the experimental context. They showed asigniWcantly higher percentage of choices of the baitedbowl during cooperative sessions than during competitivesessions (Mann–Whitney: Hy, n1 = n2 = 27, U = 84.5,P < 0.0001; Ho, n1 = n2 = 27, U = 228, P = 0.018; Hi,n1 = 27, n2 = 26, U = 153.5, P = 0.0008; Hu, n1 = 27,n2 = 20, U = 13, P < 0.0001) (Table 2).
Table 2 shows that, in competitive sessions, the foursubjects sometimes withheld information about the location
Fig. 1 Representation of the general set up of the experiment. B baited bowl, U unbaited bowl
REWARDED
B BU U
NON REWARDED REWARDEDNON REWARDED
COOPERATIVE SESSION COOPERATIVE SESSION
Table 1 Recorded behaviours: behavioural responses are deWned and rewarded as indicated above
Subject behaviour Response coding Reward: cooperative sessions
Reward: competitive sessions
Participates: extends arm toward the baited bowl Choice of the baited bowl Rewarded Unrewarded
Participates: extends arm toward the unbaited bowl Choice of the unbaited bowl Unrewarded Rewarded
Participates: orients head toward experiment during baiting, but does not extend arm toward either bowl within 60 s
Information withholding Unrewarded Unrewarded
No participation: fails to approach the apparatus Participation refused Unrewarded Unrewarded
123

Anim Cogn (2008) 11:255–266 259
Fig. 2 Percentage of completed trials during cooperative (black square) and competitive (open circle) sessions (Experiment 1)
0
20
40
60
80
100
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 2324 25 26 27
Sessions
% c
om
ple
ted
tri
als
% c
om
ple
ted
tri
als
0
20
40
60
80
100
% c
om
ple
ted
tri
als
(Hy) and(Ho)
(Hi)
(Hu)
Table 2 Percentage of the diVerent recorded behaviours for each subject in cooperative and competitive sessions in Experiment 1
Cooperative sessions Competitive sessions
Hy (%) Ho (%) Hi (%) Hu (%) Hy (%) Ho (%) Hi (%) Hu (%)
Choice of the baited bowl 88 80 78.5 98.5 63 69.6 57.7 66.5
Choice of the unbaited bowl 12 20 21.5 1.5 33 29.6 41.5 19
Information withholding 4 0.8 0.8 14.5
123

260 Anim Cogn (2008) 11:255–266
of the food, but the infrequent occurrence of this behavioursuggests that it was not a consistent tactic used to misleadthe competitive trainer.
The question arises whether the decrease in selecting thebaited bowl in competitive sessions is due to discriminationof the two contexts, or to an increase in “errors” caused bythe disturbing presence of the competitive trainer. Toanswer this question the behaviour of each subject was ana-lysed individually.
Hy During cooperative sessions Hy’s performance (% ofchoice of the baited bowl) remained stable across sessions:she selected the baited 80% or more in 23 sessions over 27sessions (Fig. 3). The presence of the competitive trainerdid not appear to modify the behaviour she had acquiredduring the training phase.
During competitive sessions, Hy’s performance wasmuch less stable; her percentage of choice of the baitedbowl did not decrease across sessions (Fig. 3). Nonetheless,she chose the unbaited bowl more frequently during com-petitive sessions compared to cooperative sessions (Mann–Whitney: n1 = 32, n2 = 89, U = 127, P < 0.0001). Therefore,the results indicate that Hy discriminated between the twocontexts, but she did not develop an eYcient and consistenttactic in the presence of the competitive trainer, whichwould be to select the baited bowl less frequently, and selectthe unbaited bowl more frequently across sessions.
The increased percentage of choices of the unbaitedbowl in competitive sessions compared with cooperativesessions does not appear to be the result of an intentionaltactic, but may be more parsimoniously interpreted as theresult of a higher “error rate” in a novel context.
Ho During cooperative sessions Ho’s performance wasvery variable: she selected the baited bowl 80% or more dur-ing the Wrst Wve sessions, then her performance declined untilthe 13th session, and increased again without stabilizing(Fig. 3). During competitive sessions her performance wasalso very variable, with no obvious pattern across sessions.
Ho’s performance is comparable to that of Hy, in thatcompetitive sessions did not give rise to the emergence of aparticular tactic. As with Hy, the increased selection of theunbaited bowl during competitive sessions is more attribut-able to the increased number of “errors” (Ho did not alwaysselect the baited bowl during cooperative sessions) than tointentional choices. But during cooperative sessions, Ho’sperformance, unlike Hy’s, was not consistent. These resultsindicate that the introduction of the competitive trainer dis-turbed Ho to the extent that she could not performeYciently, whichever trainer she was presented with.
Hi During cooperative sessions, Hi’s performance wasstable only for the Wrst ten sessions, then it decreased. The
decreased percentage of choices of the baited bowl was cor-related with the number of sessions (Spearman correlation:n = 17, r = ¡0.77, P < 0.001) (Fig. 3) and was in fact dueto the development of a position bias. From session 11, Hichose the bowl placed on the right side of the table signiW-cantly more often, and she chose it in at least eight of theten trials in 11 of the last 16 sessions. The development of aposition bias could represent a tactic for receiving thereward on 50% of the trials. This kind of tactic (alreadyfound in another study with black lemurs, see Genty et al.2004) often reXects the complicated character of a task.
During competitive sessions, Hi’s selection of the baitedbowl signiWcantly decreased across sessions (Spearmancorrelation: n = 17, r = ¡0.67, P < 0.001) and developed inthe same way as in cooperative sessions (Fig. 3).
Hi’s performance is quite diVerent from that of Hy andHo. Although she chose the baited bowl signiWcantly moreoften with the cooperative trainer than with the competitivetrainer, her global performance with both the trainersdecreased across sessions, as her frequency of choosing thebaited bowl was negatively correlated with the number ofsessions.
Hu During cooperative sessions, Hu’s performanceremained stable: he chose the baited bowl in at least 90% ofthe trials over all sessions. This result indicates that theintroduction of the competitive trainer did not aVect theperformance level Hu acquired during the training phase.
During competitive sessions, Hu’s choices of the baitedbowl decreased across the Wrst 11 sessions and were nega-tively correlated with the number of sessions (Spearmancorrelation: n = 11, r = 0.83, P < 0.001). From session 12,his selection of the baited bowl increased again and stabi-lized with a signiWcant level of choice of the baited bowl(Fig. 3).
These results reveal that Hu discriminated clearlybetween the two trainers. However, he was not able to dealadaptively with the situation, as he did not develop an eVec-tive tactic with the competitive trainer (i.e. to select thebaited bowl less frequently across sessions). Hu seemedable to adapt his behaviour according to the context butafter session 11, became too disturbed, probably by thetransfer from one context to another, and abandoned theeVective tactic developed with the competitive trainer with-out modifying his acquired tactic with the cooperativetrainer.
Discussion
In spite of the marked individual diVerences, the four sub-jects seemed to discriminate between the two trainers accord-ing to their performances in the two contexts. Nevertheless,only two subjects (Hy and Hu) were able to maintain their
123

Anim Cogn (2008) 11:255–266 261
Fig. 3 Percentage of choice of the baited bowl during coopera-tive (black square) and competi-tive (open circle) sessions (Experiment 1)
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
(Hy)
(Ho)
(Hi)
(Hu)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Sessions
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
123

262 Anim Cogn (2008) 11:255–266
signiWcant levels of choice of the baited bowl with the coop-erative trainer. With the competitive trainer, only two sub-jects (Hi and Hu) progressively decreased their percentage ofchoice of the baited bowl across sessions (only during theWrst 11 sessions for Hu). Hi was the only subject to develop astable tactic in that context, as she progressively increasedher percentage of choices of the unbaited bowl. However, shealso increased her percentage of choices of the unbaited bowlwith the cooperative trainer. Therefore, the subjects’ perfor-mances with the competitive trainer seem to be more attribut-able to an increased number of “errors” than to thedevelopment of an eYcient tactic. Only one subject (Hu)appeared to develop diVerent tactics according to the context(during the Wrst 11 sessions of the experiment) but did notstabilise these eYcient tactics subsequently.
None of the four subjects developed eVective tacticswith both the trainers, that is to say that none of them wereable to maintain a signiWcant choice of the baited bowl withthe cooperative trainer and, at the same time, to decreasethe number of choices of this bowl and simultaneouslyincrease the number of choices of the unbaited bowl withthe competitive trainer.
Experiment 2
A second experiment was conducted in which the subjectsexperienced trials with both trainers during a given session.Previous studies using sessions with mixed trainers havebeen conducted in squirrel monkeys to assess their Xexibil-ity in switching from honest to deceptive responses acrosstrials (Anderson et al. 2001). In the present study, it washypothesised that:
• mixed trials would help the subjects to keep a high levelof motivation (gain rewards from cooperative trainer in agiven session),
• the presence of the two trainers in a given session couldhelp the subjects to discriminate between them more eas-ily (would lemurs apply two diVerent rules for two diVer-ent trainers?), and would thus facilitate the emergence ofthe tactic most adapted to the context (would both “coop-erative” and “deceptive” signalling strategies be used?).
Methods
The same four individuals served as subjects. The experi-mental procedure was the same as in Experiment 1 exceptthat both trainers (cooperative and competitive) partici-pated in the same sessions. The same persons as in Experi-ment 1 acted as cooperator and competitor. Each sessionconsisted of ten trials comprising of Wve cooperative trialsand Wve competitive trials, randomly scheduled with theconstraint that neither occurred in more than three consecu-tive trials. The two trainers sat facing away on two chairsplaced side by side approximately 2 m away from the cagemesh. At the beginning of each trial, the experimenterannounced which kind of trial it was going to be before pro-ceeding to hide the bait. After the experimenter returned tohis place, there followed a 5-s delay, after which one of thetrainers stood up and approached the tray. Each subjectreceived a total of 17 sessions (85 cooperative trials and 85competitive trials).
Results
Participation rate
The four subjects participated fully in all 17 sessions exceptHu who completed only 7 trials during the Wrst 10-trial ses-sion. The analysis of the results includes the 17 sessions forall subjects.
Performance and behaviour analysis
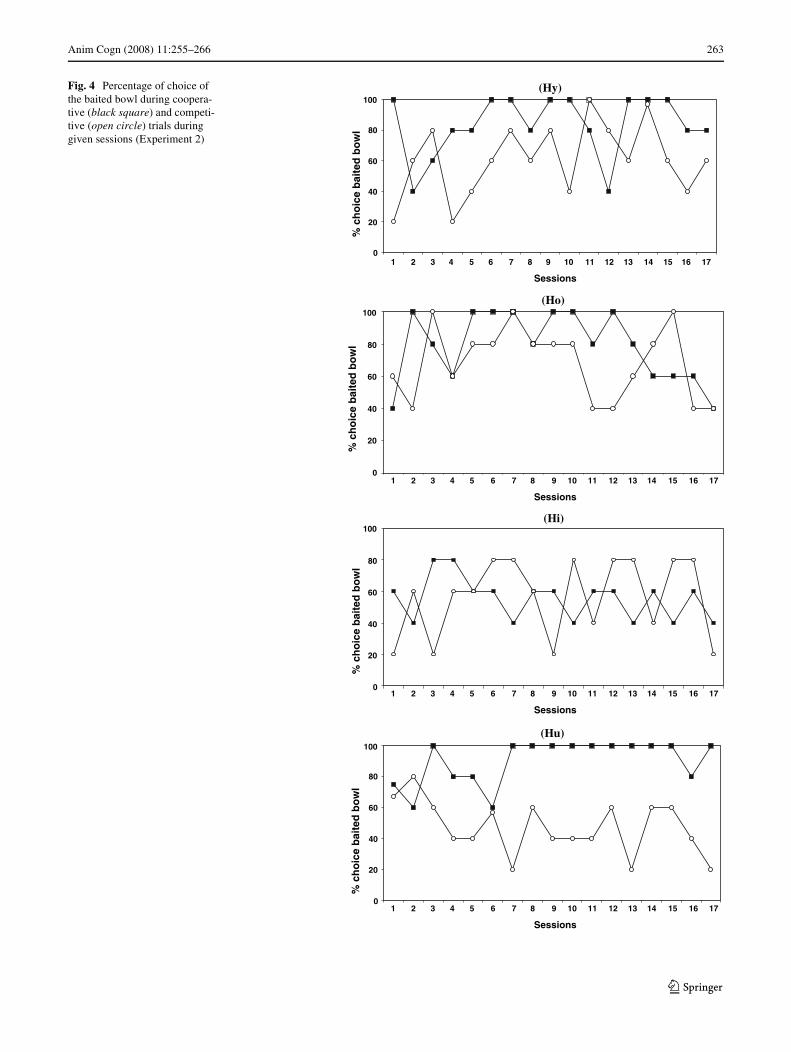
Ho and Hi did not behave diVerently with the two trainers,as these subjects’ percentage of choices of the baited bowlwas not signiWcantly diVerent between the cooperativetrainer and the competitive trainer (Mann–Whitney:n1 = n2 = 17; Ho: U = 105.5, P = 0.18; Hi: U = 129,P = 0.60) (Table 3). Moreover, Hi was the only subject notto show a signiWcant preference for the baited bowl duringcooperative trials (Binomial test: z = ¡0.9, P = 0.18). Hoand Hi’s performances did not evolve in any way sugges-tive of eYcient tactics (Fig. 4).
The other two subjects, Hy and Hu, selected the baitedbowl signiWcantly more often with the cooperative trainer
Table 3 Percentage of the diVerent recorded behaviours for each subject in cooperative and competitive trials in Experiment 2
Cooperative sessions Competitive sessions
Hy (%) Ho (%) Hi (%) Hu (%) Hy (%) Ho (%) Hi (%) Hu (%)
Choice of the baited bowl 83.5 79 55 89 61 68 56.5 45
Choice of the unbaited bowl 16.5 21 45 9.4 35 32 43.5 50
Information withholding 1.8
Participation deny 1.1 3.2
123
Anim Cogn (2008) 11:255–266 263
Fig. 4 Percentage of choice of the baited bowl during coopera-tive (black square) and competi-tive (open circle) trials during given sessions (Experiment 2)
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
0
20
40
60
80
100
(Hy)
(Ho)
(Hi)
(Hu)
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
% c
ho
ice
bai
ted
bo
wl
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Sessions
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Sessions
123

264 Anim Cogn (2008) 11:255–266
than the competitive trainer (Mann–Whitney: n1 = n2 = 17;Hy: U = 68, P = 0.0086; Hu: U = 12, P < 0.0001)(Table 3). It is interesting to note that in the Wrst session,Hu withheld information on the 6th trial and refused to par-ticipate in the last three trials. These behaviours might reX-ect the disturbance produced by the new experimentalparadigm, or simply a lack of motivation on that particularday. Hy also failed to develop eYcient tactics (Fig. 4). TheWnal subject, Hu, clearly diVerentiated the two trainers.Considering the overall competitive sessions, his total per-centage of choices of the unbaited bowl in competitive tri-als, although not signiWcant, was higher than thecorresponding total for the baited bowl (50% versus 45%)(Binomial test: z = ¡0.86, P = 0.19). From session 7, Huscored 80% or more choices of the baited bowl duringcooperative trials, compared to a score ranging from 60 to20% choice of the baited bowl (with a mean of 38%) incompetitive trials (Fig. 4). Thus, Hu’s performanceshowed, from session 7, the emergence of eYcient tacticswith both trainers. During cooperative trials, he maintainedthe tactic acquired during the training phase, and, whenfaced with competitive trials he selected the unbaited bowlmore often, sometimes reaching four choices of the unba-ited bowl over the Wve competitive trials in a session.
Discussion
The experimental procedure used in Experiment 2 was use-ful for one subject (Hu) out of the four tested, and permittedhim to optimise his behaviour according to the context.DiVerential pointing clearly occurred from the 7th sessionand was maintained through the end of the experiment.Other behaviours, such as “participation refusal” or “infor-mation withholding”, were seldom recorded. The behaviourof this subject is similar to the deception-like behavioursreported for chimpanzees (WoodruV and Premack 1979)capuchin monkeys (Mitchell and Anderson 1997) andsquirrel monkeys (Anderson et al. 2001) tested under simi-lar experimental paradigms. However, it is diYcult to con-clude that our lemur subjects’ deceptive behaviour wasdone to mislead the competitive trainer, in view of theintensive training required for the emergence of such tactics.
Conclusion
The results of this study reveal that brown lemurs are ableto discriminate between two distinct contexts (cooperativeand competitive) and can adapt their behaviours to thesecontexts. The four subjects showed distinct responses to thetwo trainers, with only one subject (Hu) developing eVec-tive tactics in the two contexts. Hu maintained his signiW-cant preference for the baited bowl with the cooperativetrainer and decreased his choice of the baited bowl while
choosing the unbaited bowl more frequently with the com-petitive trainer. Hu only developed these eVective behav-iours during Experiment 2, when the cooperative andcompetitive trainers were both present in the same session.Immediate comparison between the two trainers led to bet-ter overall performance in this subject. It is interesting tonote that in Experiment 1 Hu sometimes refused to partici-pate or withheld information about the location of the baitduring competitive sessions, but he did not do this duringthe competitive trials of Experiment 2 (except for the Wrstsession). It is possible that, because his choices did not leadto reward during competitive sessions of Experiment 1, hismotivation decreased and he consequently presented thosealternative behaviours. In Experiment 2, however, the pres-ence of the two trainers within the same session sustainedhis motivation level enough for Hu to develop an eVectivetactic with the competitive trainer while maintaining theeVective tactic with the cooperative trainer.
The results also show important individual diVerences,as only one of the four subjects (Hu) clearly adapted hisbehaviour to the context. However, our small sample sizedoes not allow any conclusions concerning the inXuence ofage or sex on performance and further experiments areneeded to assess these issues.
A procedure similar to that of Experiment 1 was previ-ously conducted in black lemurs (Eulemur macaco, Gentyand Roeder 2006). The main diVerence between the twospecies is that “participation refusal” and “informationwithholding” were much more frequent in black lemurs.The two species of lemurs (brown lemurs, Eulemur fulvusand black lemurs, Eulemur macaco) diVer in their socialorganization. In black lemurs, females are dominant overmales (Fornasieri et al. 1993) whereas in brown lemurs,males are the central individuals of the group (Roeder andFornasieri 1995). How diVerences in social organisationcould be related to deceptive tactics should require atten-tion in further studies.
From a comparative perspective we have assessed ourlemur subjects’ performance against that of the other pri-mate species similarly tested, considering the qualitative(behaviour) and quantitative (number of trials to reach asame performance criterion) aspects of their performance.Mitchell and Anderson (1997) considered that deceptionoccurred if their capuchin monkeys chose the unbaitedbowl on 80% or more of competitive trials. We used thesame criterion to compare the performances in the competi-tive context of the diVerent primate species studied. Onlyone capuchin reached this criterion in seven competitivesessions out of 36 (Anderson et al. 2001). Squirrel monkeysreached this criterion only after the introduction of colour-cued containers. No black lemurs reached this criterion.However one subject presented a maximum of 60% ofchoices of the unbaited bowl in seven of 19 sessions, and
123

Anim Cogn (2008) 11:255–266 265
her selections of the unbaited bowl with the competitivetrainer increased signiWcantly across the Wrst nine sessions,before she developed a position bias (Genty and Roeder2006). In the present study, three subjects reached the crite-rion, in four sessions for Hi, two sessions for Hy and threesessions for Hu out of 17 (Experiment 2). Hi and Hyreached this criterion in the Wrst session of Experiment 2and Hu in the seventh session, whereas the capuchin mon-key reached it in the 22nd session (220 trials). Thus, from aquantitative point of view, the performance of our brownlemurs in the competitive context is comparable to that ofthe simian species studied. If we analyse their behaviour incompetitive contexts, it is clear that the introduction of acompetitive trainer in Experiment 1 caused a decrease inperformance with the cooperative trainer in three of oursubjects, an eVect also noted in squirrel and capuchin mon-keys (Anderson et al. 2001; Mitchell and Anderson 1997).But during competitive sessions, the lemurs showed pro-gressive behavioural adjustments to the new context as theysometimes withheld information about food location,declined to participate, or increasingly selected the unbaitedbowl, before performance deteriorated or a position biasdeveloped. Similar tactics have been described in capuchinmonkeys, except that they were able to stabilize their tac-tics whereas brown lemurs did not in Experiment 1. Onlyone brown lemur subject stabilized an eYcient tactic withboth trainers in Experiment 2, within which the discrimina-tion between the 2 contexts was facilitated in mixed trainerssessions. Therefore, from a qualitative point of view,lemurs behave similarly to simian species as they expressedsimilar behaviours in competitive contexts but with the dis-tinction that they were not able to stabilize eYcient tacticsadapted to both the contexts, one subject excepted. In ourexperiment, all the animals were trained to be responsive tothe cooperative trainer Wrst. It remains uncertain how theymight have behaved if they had not been trained that way,and were just given random presentations of both trainers.
Our results lead us to conclude that lemurs may learn toreliably discriminate between a cooperative and a competi-tive trainer. However, although their behaviour correspondsto the deWnition for tactical deception (Byrne and Whiten1985), a crucial question is whether their behaviour in com-petitive context can be described as intentional or planneddeception. Is the pointing gesture toward the unbaited bowlin the competitive context really intentional, done to mis-lead the competitive trainer, or the result of simple condi-tional learning? Perhaps, it is the experimental protocoldescribed by WoodruV and Premack (1979) which requiresscrutiny. Mitchell and Anderson (1997) suggested that thebehaviour of their capuchin monkeys (Cebus apella) in asimilar protocol was explainable in terms of response inhi-bition and conditional discrimination learning. Indeed, thesubjects could simply have learned to indicate the baited
bowl in the presence of the cooperative trainer and to indi-cate the unbaited bowl in the presence of the competitivetrainer. These discriminative behaviours could result fromcomplex associative learning requiring the inhibition or thereduction of the behaviour learned in the cooperative con-text when placed in the competitive context. Constraints onlearning might therefore explain why lemurs had diYcul-ties in stabilizing the most eYcient tactic in the two con-texts. It is clear that nonhuman primates can learn to behavedeceptively as a result of intensive training, but it is diYcultto conclude that this deceptive behaviour reXects intention-ality as opposed to simple conditional discrimination.
Acknowledgments We are grateful to Dr. J.R. Anderson and A.Ruiz for helpful comments on the manuscript and for correcting theEnglish. We thank four anonymous reviewers for their comments toimprove the manuscript. The authors attest that the experiments com-ply with the current French laws.
References
Anderson JR, Kuroshima H, Kuwahata H, Fujita K, Vick SJ (2001)Training squirrel monkeys (Saimiri sciureus) to deceive: acquisi-tion and analysis of behavior toward cooperative and competitivetrainers. J Comp Psychol 115:282–295
Byrne RW, Whiten A (1985) Tactical deception of familiar individualsin baboons (Papio ursinus). Anim Behav 33:669–673
Byrne RW, Whiten A (1992) Cognitive evolution in primates: evi-dence from tactical deception. Man 27:609–627
Carlson SM, Moses LJ, Hix HR (1998) The role of inhibitory processesin young children’s diYculties with deception and false belief.Child Dev 69:672–691
Coussi-Korbel S (1994) Learning to outwit a competitor in mangabeys(Cercocebus torquatus torquatus). J Comp Psychol 108:164–171
Deaner RO (2000) An experimental study of deception in ringtailedlemurs. AAPA Abstracts, p 135
Ducoing AM, Thierry B (2003) Withholding information in semifree-ranging Tonkean macaques (Macaca tonkeana). J Comp Psychol117:67–75
Fornasieri I, Caubère M, Roeder JJ (1993) Social dominance and pri-ority to drinking in Lemur macaco. Aggress Behav 19:455–464
Ganzhorn JU (1988) Food partitioning among Malagasy primates.Oecologia 75:436–450
Genty E, Palmier C, Roeder JJ (2004) Learning to suppress responsesto the larger of two rewards in two species of lemurs, Eulemur ful-vus and Eulemur macaco. Anim Behav 67:925–932
Genty E, Roeder JJ (2006) Learning to deceive in black lemurs(Eulemur macaco). J Exp Psychol: Anim Behav Process32:196–200
Harrington JE (1975) Field observations of social behaviour of Lemurfulvus fulvus E. GeoVroy 1812. In: Tattersall I, Sussman RW(eds) Lemur biology. Plenum, New York, pp 259–279
Jolly A (1966) Lemur social behaviour and primate intelligence.Science 153:501–506
Kappeler PM, Ganzhorn JU (1993a) The evolution of primate commu-nities and societies in Madagascar. Evol Anthropol 159–171
Kappeler PM, Ganzhorn JU (1993b) Lemur social systems and theirecological basis. Plenum, New York
La Frenière PJ (1988) The ontogeny of tactical deception in humans.In: Byrne RW, Whiten A (eds) Machiavellian intelligence.Oxford University Press, Oxford, pp 238–252
123

266 Anim Cogn (2008) 11:255–266
Menzel EW (1974) A group of young chimpanzees in a one-acre Weld.In: Schrier AM, Stollnitz F (eds) Behavior of nonhuman primates,vol 5. Academic, New York, pp 83–153
Mitchell RW (1986) A framework for discussing deception. In: Mitch-ell RW, Thompson NS (eds) Deception: perspectives on humanand nonhuman deceit. State University of New York Press, Alba-ny, pp 3–40
Mitchell RW, Thompson NS (1986) Deception: perspectives on hu-man and nonhuman deceit. State University of New York Press,Albany
Mitchell RW, Anderson JR (1997) Pointing, withholding information,and deception in capuchin monkeys (Cebus apella). J Comp Psy-chol 4:351–361
Newton P, Reddy V, Bull R (2000) Children’s everyday deception andperformance on false-belief tasks. Br J Dev Psychol 18:297–317
Pereira ME, Kappeler PM (1997) Divergent systems of agonisticbehaviour in Lemurid primates. Behaviour 134:225–274
Pereira ME, Kaufman R, Kappeler PM, OverdorV DJ (1990) Femaledominance does not characterize all of the Lemuridae. Folio Pri-matol 55:96–103
Roeder JJ, Fornasieri I (1995) Does agonistic dominance imply feed-ing priority in lemurs? A study in Eulemur fulvus mayottentis. IntJ Primatol 16:629–642
Russel J, Mauthner N, Sharpe S, Tidswell T (1991) The “window task”as a measure of strategic deception in preschoolers and autisticsubjects. Br J Dev Psychol 9:331–349
Sodian B (1991) The development of deception in young children. BrJ Dev Psychol 9:173–188
Tattersall I (1982) The primates of Madagascar. Columbia UniversityPress, New York
Whiten A, Byrne RW (1988) Tactical deception in primates. BehavBrain Sci 11:233–273
WoodruV G, Premack D (1979) Intentional communication in thechimpanzee: the development of deception. Cognition 7:333–362
123





























