Embed Size (px)
Citation preview

Rädler/WS 2009 BPM §1.3 1

Rädler/WS 2009 BPM §1.3 2
Protein Structure
special sequences function conformation
Peptide chain, 20 amino acids
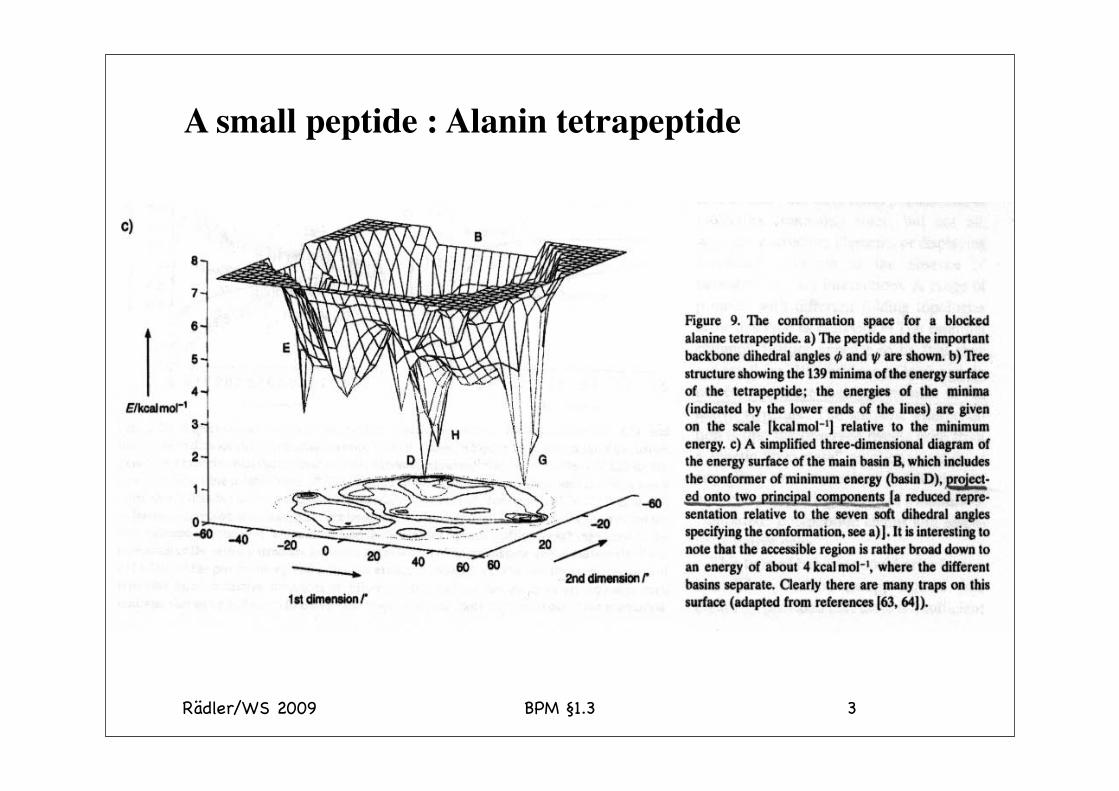
Rädler/WS 2009 BPM §1.3 3
A small peptide : Alanin tetrapeptide

Rädler/WS 2009 BPM §1.3 4
Folding of RNAse A in the test tube
denaturation renaturation
Incubate proteinin guanidine
hydrochloride(GuHCl)or urea
100-folddilution of proteininto physiological
buffer
Anfinsen, CB (1973) Principles that govern the folding of protein chains. Science 181, 223-230.
- the amino acid sequence of a polypeptide is sufficient to specify its three-dimensional conformation
Thus: “protein folding is a spontaneous process that does not require the assistance of extraneous factors”
(aggregation)

Rädler/WS 2009 BPM §1.3
Ribonuclease kann durch Oxidation (Spaltung der S-S Bindung)denaturiert werden
5
o) Nofür t icheRibonuclea se
Abb. 3.12: Die zwei Zuslände der Ribonuklease:links: KomDakt€ Funktionsfomrechts: Oenatudert€ Form
b )0eno tu r ie r teRibonu cleose
95H H65
Das Enzym hat 8 s-s Bindungen. Im Prinzip könnten 56 verschiedene Zustände (Isomere) gebildet werden. Es gibt aber offenbar nur einen Zustand niedrigster Energie.

Rädler/WS 2009 BPM §1.3 6

Rädler/WS 2009 BPM §1.3 7

Rädler/WS 2009 BPM §1.3 8
Probing Proteins in SolutionHigh-resolution techniques (local):• FTIR• Fluorescence• NMR• UV absorption
Low-resolution techniques:• SAXS• DLS siehe BPE
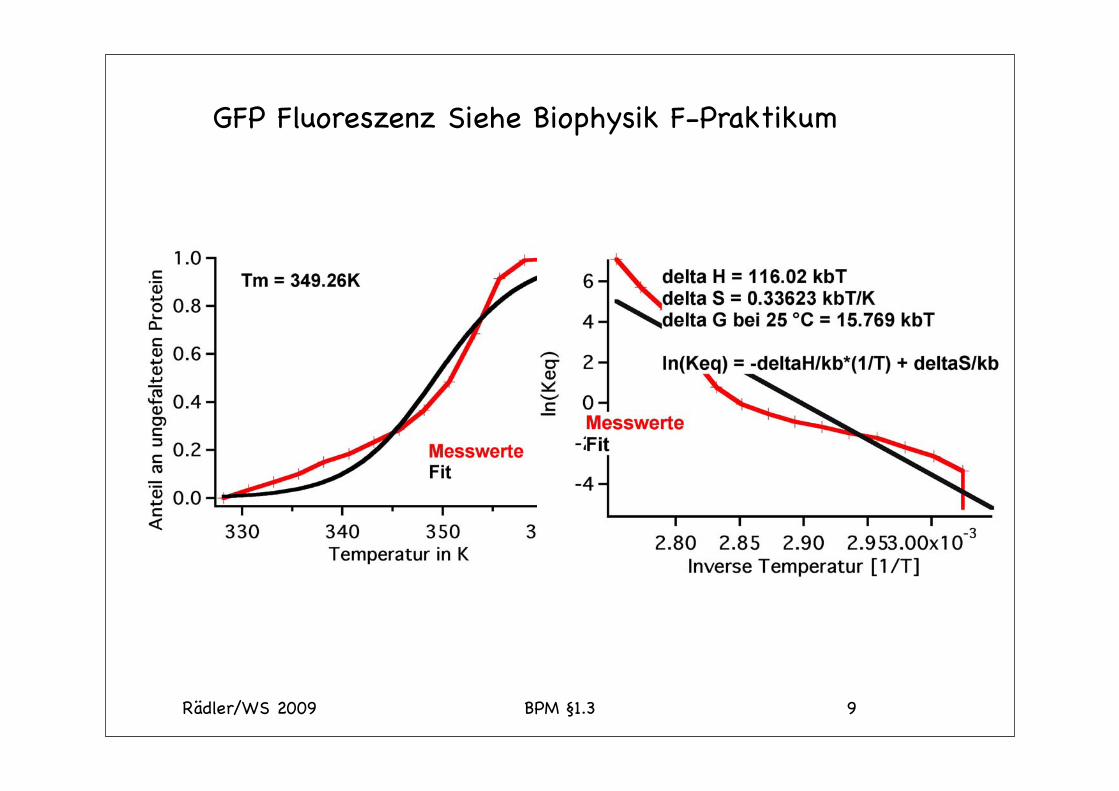
Rädler/WS 2009 BPM §1.3 9
GFP Fluoreszenz Siehe Biophysik F-Praktikum
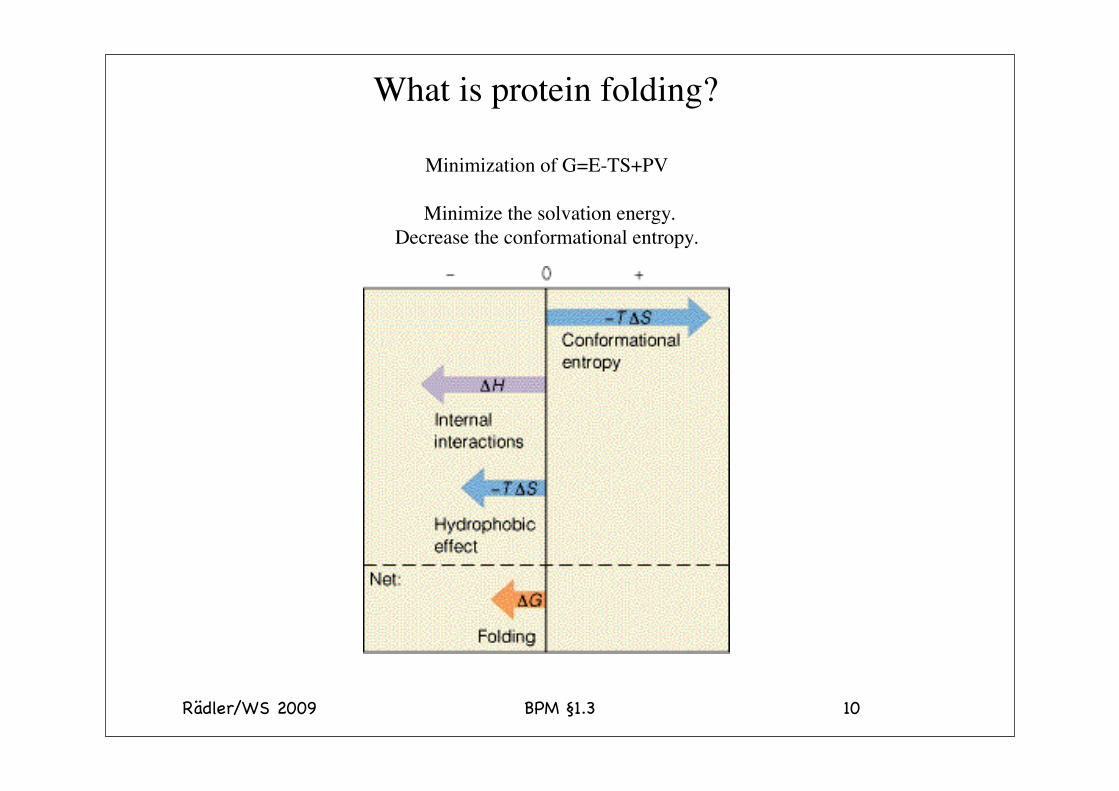
Rädler/WS 2009 BPM §1.3 10
What is protein folding?
Minimization of G=E-TS+PV
Minimize the solvation energy.Decrease the conformational entropy.

11
Wechselwirkungen in Proteinen
- Elektrostatik (Ionenbrücken, Debeye-Hückel)
- H-Brücken (in Konkurrenz zu Wassser)
- Van der Waals (Lennard Jones Potential)
- Hydrophobe WW (Allg. Entropische LM-WW)
- Peptid-Bindungspotentiale

Rädler/WS 2009 BPM §1.3 12

Rädler/WS 2009 BPM §1.3 13
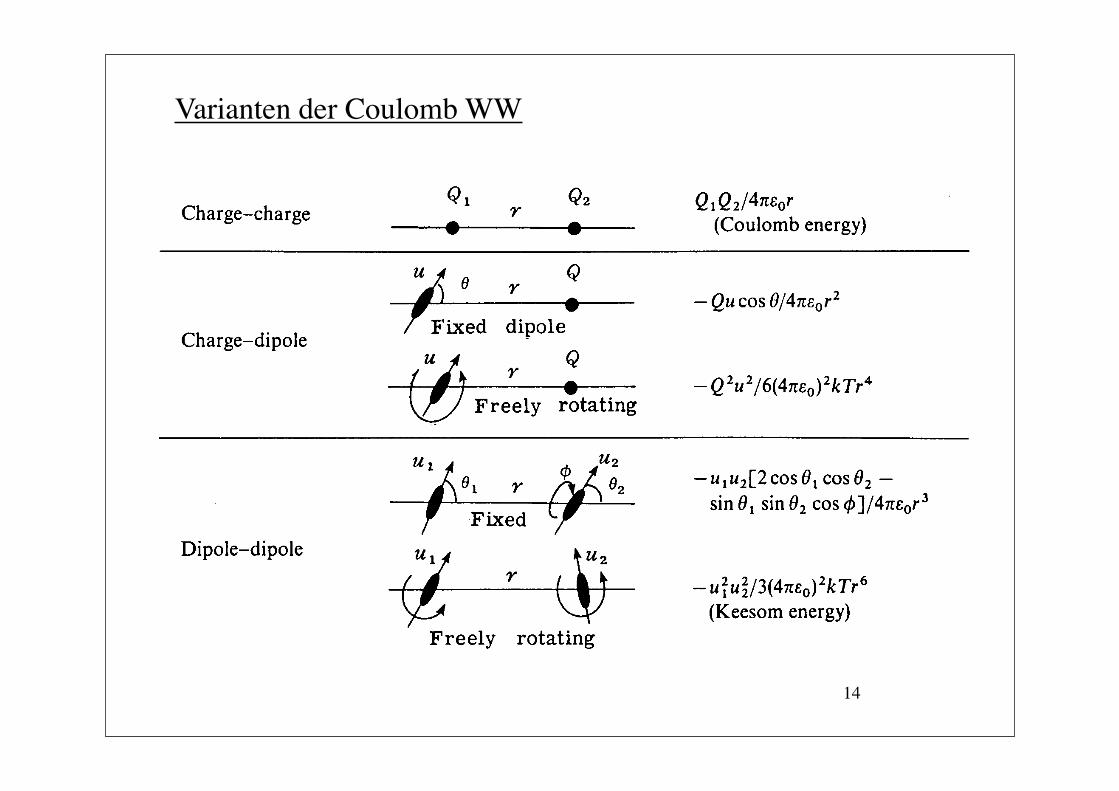
14
Varianten der Coulomb WW

15
Varianten der Coulomb WW
Siehe J.N. Israelachvili, Intermolecular and Surface Forces with Applications to Colloidal and Biological Systems, Academic Press 1985

16
Partner µ in Debye β in 1077 Jm6 βKeesom βDebye βLondon
H2O 1.82 2.1 85 5 10Benzol 0 4.3 0 0 100Phenol 1.55 6.5 15 9 76Diphenylanilin 1.1 14.3 2 4 94
€
WVanderWaals= −βr6
= −βKeesom + βDebye + βLondon
r6
1 Debeye = 3.3 10-30 Cm
Ausser bei Wasser dominiert London (Dispersions) WW
Bsp: 2 Benzolringe; für r = 5 Å. ist Wdip ≈ kBT (bei 300 K).
Typische Werte

17
d
r
A
Bε = 1
dξ
ξ y
dy
ε BA A
B
ε ε BA εM
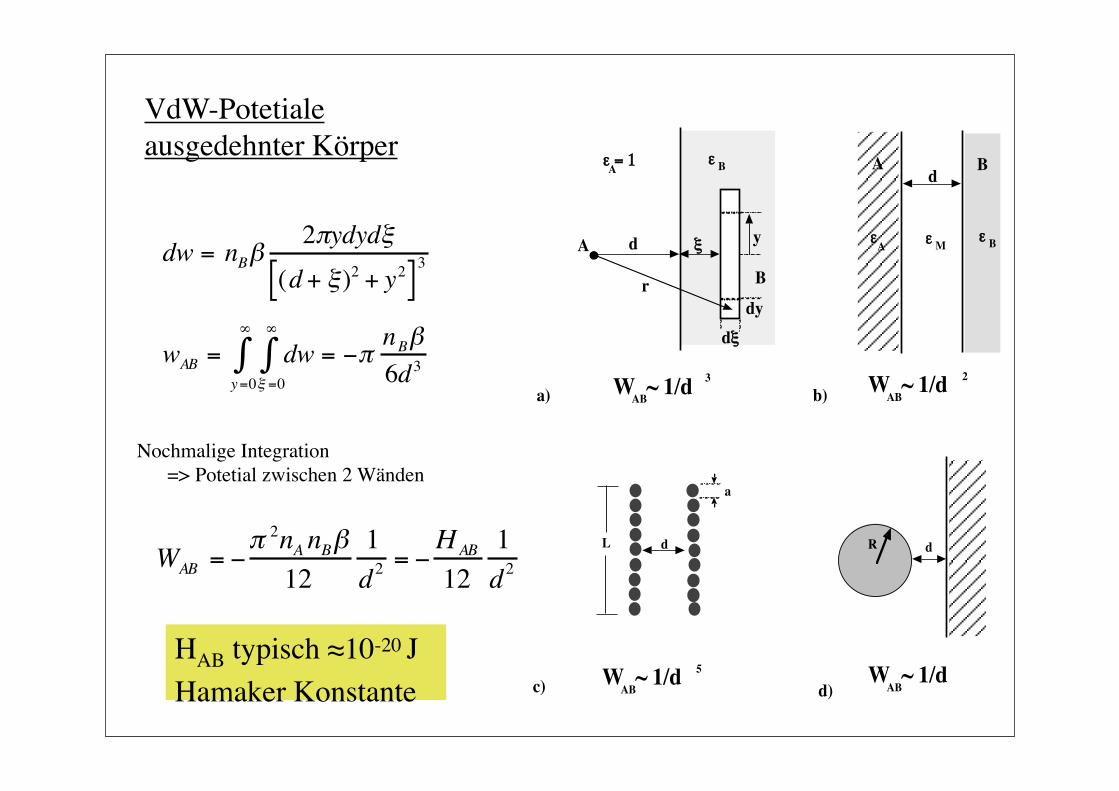
ABW ~ 1/d
ABW ~ 1/d 3
ABW ~ 1/d 2
ABW ~ 1/d 5
a) b)
c) d)
a
L R d
d
d
€
dw = nBβ2πydydξ
(d + ξ)2 + y2[ ]3
€
wAB = dw = −πnBβ6d3ξ =0
∞
∫y=0
∞
∫
VdW-Potetiale ausgedehnter Körper

18
d
r
A
Bε = 1
dξ
ξ y
dy
ε BA A
B
ε ε BA εM
ABW ~ 1/d
ABW ~ 1/d 3
ABW ~ 1/d 2
ABW ~ 1/d 5
a) b)
c) d)
a
L R d
d
d
€
dw = nBβ2πydydξ
(d + ξ)2 + y2[ ]3
€
wAB = dw = −πnBβ6d3ξ =0
∞
∫y=0
∞
∫
VdW-Potetiale ausgedehnter Körper
€
WAB = −π 2nA nBβ12
1d2
= −HAB
121d2
Nochmalige Integration => Potetial zwischen 2 Wänden
HAB typisch ≈10-20 J Hamaker Konstante
19
d
r
A
Bε = 1
dξ
ξ y
dy
ε BA A
B
ε ε BA εM
ABW ~ 1/d
ABW ~ 1/d 3
ABW ~ 1/d 2
ABW ~ 1/d 5
a) b)
c) d)
a
L R d
d
d
€
dw = nBβ2πydydξ
(d + ξ)2 + y2[ ]3
€
wAB = dw = −πnBβ6d3ξ =0
∞
∫y=0
∞
∫
VdW-Potetiale ausgedehnter Körper
€
WAB = −π 2nA nBβ12
1d2
= −HAB
121d2
Nochmalige Integration => Potetial zwischen 2 Wänden
HAB typisch ≈10-20 J Hamaker Konstante

20
€
HAMB =34kBT
εA −εMεA +εM
εB −εMεB + εM
+
3h4π
εA (iv) −εM (iv )εA (iv) + εM (iv)
εB (iv) −εM (iv)εB (iv) +εM (iv)
dv
v1
∞
∫
Lifshitz Theorie der VdW-WW
0-Frequenz-Beitrag(Keesom und Debye)
Dispersions WW
Quantenfeldtheoretische Herleitung(lesbare Approximation siehe Israelachvili et al)
Vorsicht: - Durch Medium kann VdW-WW auch repulsiv werden!Aufgrund der endlichen Lichtgeschwindigkeit und damit der endlichen Übertragungszeit der ε-Fluktuationen (Retardierung ) fällt das Potential für große Abstände (d > 10 nm) schneller ab (1/d7)
-

21
Debye-Hückel Theorie der Makro-Ionen
+
+ +
++
+
+
Makro-Ionz.B. Protein
++ -
-
-
-
-
-Gelöste Gegen-Ionen
-
ρ(r) -
++
++
+
-
-
--
-
Elektrolyt-Lösung
€
niZii∑ + Z1 = 0Elektro-
Neutralität:Elektrolyt&Gegenionen
Makroion
PoissonGleichung
€
Δψ(r) = −4πεε0
ρ(r)=>
Bolzmann-Statistik
€
ni(r) = ni(0)exp −Zieψ(r)kBT
€
Δψ(r) = −4πεε0
Zieni(0)exp −Zieψ(r)kBT
i=1
N
∑Eingesetzt =>
Exakt, aber nur numerisch lösbar!

22
€
Δψ(r) = −4πεε0
Zieni(0)exp −Zieψ(r)kBT
i=1
N
∑
€
(exp(x) ≈1+x1!
+x 2
2!+ ...)
€
Δψ r( ) = −1εε0
zieni(0) 1−zieψ r( )kbT
i∑ = −
1εε0
zieni(0) −zi2e2ni(0)ψ r( )
kbTi∑
i∑
€
Δψ r( ) =1εε0
e2
kbTψ r( ) zi
2nii∑ (0)
=0 wegen Elektroneutralität
{
€
Δψ(r )− κ 2ψ(r) = 0
€
κ 2 =1
εε0kBTni
i=1
N
∑ (0)Zi2e2mit
κ = Debye-Länge
Näherung: Hohe Temperatur(problematisch)
€
<<1
{

23
€
Δψ(r )− κ 2ψ(r) = 0
Allg. Lösung:
€
ψ r( ) = Ae−κr
r+ B
e+κr
r
weil
€
limψ r( ) = 0r->∞
€
limψ r( ) =eZI4πε0ε
1rr-> 0
€
ψ r( ) =1εε0
Q4π
e−κr
r
€
κ 2 =1
εε0kBTni
i=1
N
∑ (0)Zi2e2

24
I in Mol/l 10-5 10-4 10-3 10-2 10-1 1κ-1 in nm 170 30 9.6 3 1 0.3d in nm 76 35 16 7.6 3.5 1.6
Typische Werte für Abstand zwischen den Ionen und Abschirmlängen für 1-wertige Elektrolyten
Bjerrum-Länge: Abstand zwischen zwei Ladungen bei dem Coulomb-Energie gleich thermischer Anregung ist
lBjerrum = e2/(εε0kBT) = 0,72 nm
150 mMPhysiologische Ionenstärke

25
Kolloidale Stabilisierung
d
potentielleEnergie W
d
potentielleEnergie W
b)a)
W el
VdWW VdWW
W el
€
W = −Ad6
+Bd12
+Cde−κd
WVdW≈ Wel
d ≈ 10 nm∆G ≈ kBT∆G* >> kBT
+++
++
++
+
++ +
++
--
---
--
--
---
-
-
-
-
d
WVdW> Wel

Rädler/WS 2009 BPM §1.3 26
Electrostatic Contributions• Φi=(zie/4πεo)(1/r2) coulomb potential• Sensitive to pH and ion concentrations• pH determines total charge (pI)• Ionic strength determines effective range of interactions• Ion pairs contribute 1-3 kcal/mol (on surface)• Ion pairs generally destabilizing if buried (cost up to 19 kcal/mol/ion
to completely bury• Ion pairs contribute ~5-15 kcal/mol per 150 aa’s

Rädler/WS 2009 BPM §1.3 27
Hydrogen Bonds
• ~90% of CO and NH groups H-bonded• Hydrogen bonds contribute 2-10 kcal/mol• Destabilizing by themselves (transfer of polar groups from
high to low dielectric medium• If driven by other forces (e.g. hydrophobic collapse) favour
“internal organization”

Rädler/WS 2009 BPM §1.3 28
Measuring Unfolding

Rädler/WS 2009 BPM §1.3 29
PNAS 45: 1601 (1959)

Rädler/WS 2009 BPM §1.3 30
Hydrophobic Effect• At normal temp’s the “hydrophobic effect” is entropic
water molecules form ordered structures around nonpolar compounds• Hydrophobic residues collapse in to exclude water• Additional forces can then stabilize (vdw, h-bond,intrinsic properties)• Hydrophobic effect is dependent on temperature
(unstable at high AND low temp).

Rädler/WS 2009 BPM §1.3 31
Hydrophobicity is the dominant driving force of protein folding

Rädler/WS 2009 BPM §1.3 32
How many conformations are there in the Native state?
There are only a small number of ways to configure a chain to maximize the number of nonpolar contacts.

Rädler/WS 2009 BPM §1.3 33
Thermodynamic considerations• Protein stability is composed of two components. ΔG = ΔH-TΔS• There is a complex temperature dependence for ΔH and TΔS which means that the contribution of the enthalpic and entropic terms changes with temperature.
• This temperature dependence arises from the anomalously high change in heat capacity on transferring hydrophobic compounds into water. This is the hall-mark of the hydrophobic effect and

Rädler/WS 2009 BPM §1.3 34
Heat Capacity• The heat capacity influences both the temperature
dependence of the enthalpy and entropy
• It is proportional to the buried non-polar surface area as are all of the thermodynamic parameters.
• The large heat capacity is indicative of a well ordered water structure around non-polar molecules in water as is evident from their partial specific volumes when dissolved in water
€
Cp =δHδT
=TδSδT

Rädler/WS 2009 BPM §1.3 35
Temperature dependence of ΔG
• Thermodynamics of transfer of a hydrocarbon from liquid to aqueous solution.
• The temperature dependence is the result of different heat capacities of the two phases.
• The large changes in ΔH and TΔS compensate so that ΔG is fairly constant with temperature

Rädler/WS 2009 BPM §1.3 36
Temperature dependence of ΔH and TΔS continued
ΔH becomes more favorable at lower temperatures, whereas the entropic term becomes less favorable. This is consistent with an increase in the order in the water surrounding the non-polar molecule.The water-ordering increases the interaction between solvent and solute and thus "enhances" the solubility that would occur in its absence. Even so, the interactions between solute and water eliminate hydrogen bonds within the water that cannot be compensated for by the ordering of the water.Significantly the van der Waals interactions are greater in the pure water and solute than in the dissolved solute.It is the loss of hydrogen bonds and van der Waals interactions that is the cause
ΔH is ~0 at room temperature
Terms counterbalance

Rädler/WS 2009 BPM §1.3 37

Rädler/WS 2009 BPM §1.3 38

Rädler/WS 2009 BPM §1.3 39
Determination of protein stability.
• This can be measured with a variety of tools including, microcalorimetry, spectroscopy, and enzyme function.
• The transition can be accomplished with heat or denaturants.
• The area under the curve gives ΔH which agrees with measurements based on the van't Hoff equation
Rädler/WS 2009 BPM §1.3 40
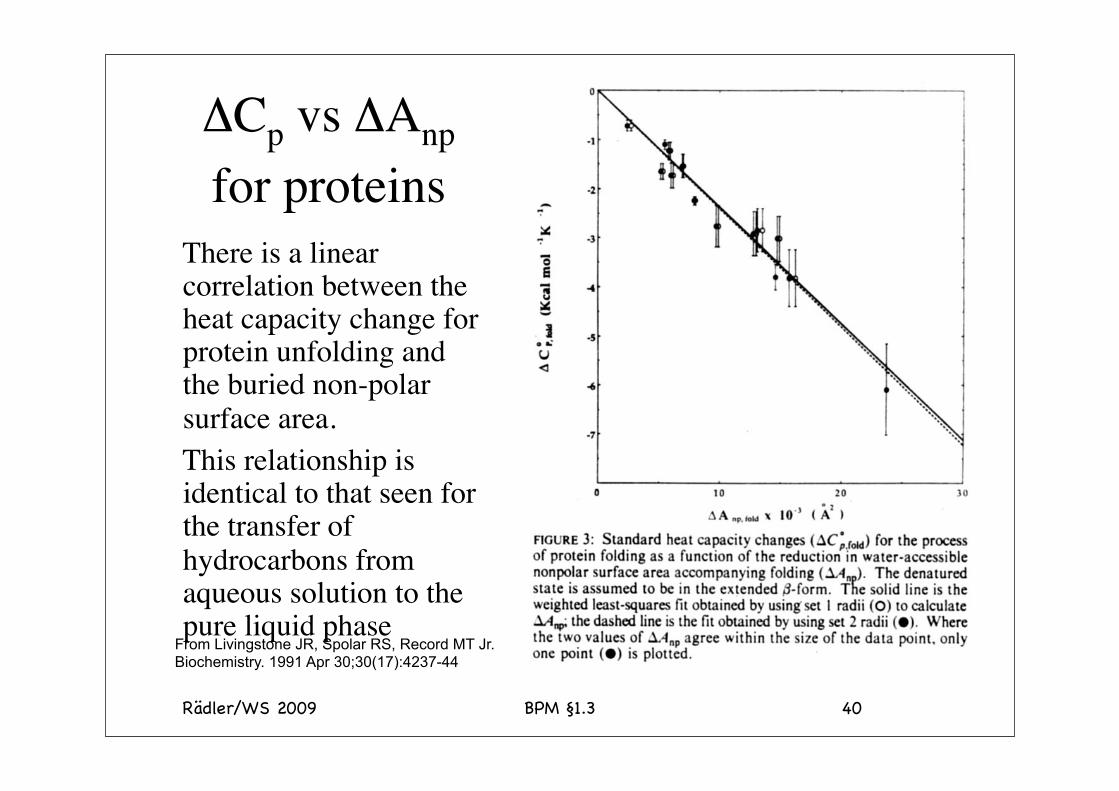
ΔCp vs ΔAnp for proteins
There is a linear correlation between the heat capacity change for protein unfolding and the buried non-polar surface area.This relationship is identical to that seen for the transfer of hydrocarbons from aqueous solution to the pure liquid phase
From Livingstone JR, Spolar RS, Record MT Jr. Biochemistry. 1991 Apr 30;30(17):4237-44

Rädler/WS 2009 BPM §1.3 41
Buried hydrophobic surface area• The buried hydrophobic surface area for a protein
correlates with the protein stability.
• Although it is difficult to predict the overall stability of a protein, it is possible to predict the worst case scenario that a mutation might produce based on changes in buried surface area.
• Occlusion of 1Å2 of hydrophobic surface area provides ~25 cal mol-1 of stability.
1kBT300K = 25 meV = 4 pNnm = 0.6 kcal/Mol = 2.5 kJ/Mol

Rädler/WS 2009 BPM §1.3 42
Denaturants• Temperature• pH
– Change the ionization state of critical residues• Detergents
– Bind strongly to the unfolded protein• High concentrations of water soluble organic
substances– Aliphatic alcohols. These disrupt the water structure
• Ionic or polar denaturants including urea and guanidinium

Rädler/WS 2009 BPM §1.3 43
Denaturants: The Hofmeister Series
• The ability of an ion to stabilize a protein follows the Hofmeister series
• Anions SO4
2->H2PO4->CH3COO->Cl->Br->I'->ClO4
->SCN-
• CationsNH4
+,Cs+,K+,Na+>Li+>Mg2+>Ca2+>Ba2+ >guanidinium>urea

Rädler/WS 2009 BPM §1.3 44
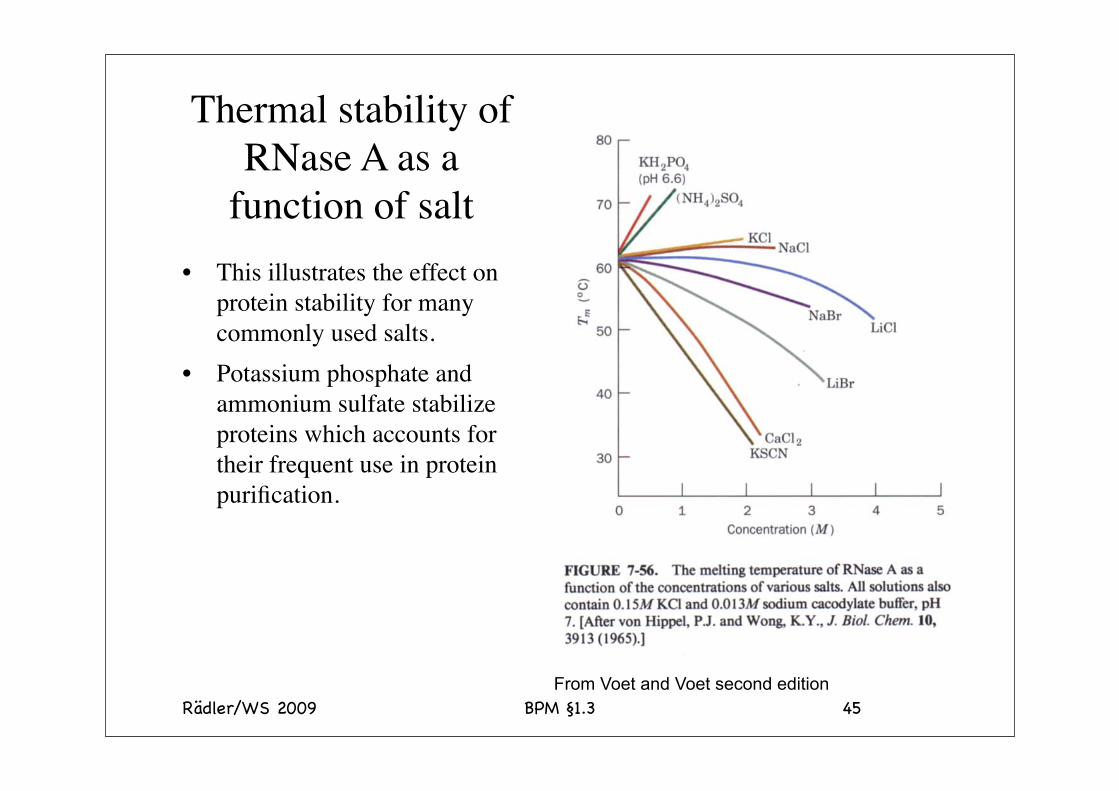
Thermal stability of RNase A
The thermal stability varies considerably depending on the nature of the precipitant. Note the brutal combination of guanidium and thiocyanate. In contrast when guanidinium is combined with sulfate the salt is stabilizing. This illustrates the effects of the salts in the Hofmeister series. Note: the effects are accumulative.
The temperature is the midpoint of thermal unfolding
Rädler/WS 2009 BPM §1.3 45
Thermal stability of RNase A as a
function of salt• This illustrates the effect on
protein stability for many commonly used salts.
• Potassium phosphate and ammonium sulfate stabilize proteins which accounts for their frequent use in protein purification.
From Voet and Voet second edition











![80 100 125 150 170 [BPM] STYLES & TEMPO IN ELECTRONIC … · 2019. 3. 6. · Dubstep [130-145 BPM] Trap [120-160 BPM] [140 BPM] Hardstyle [150 BPM] Breakbeat [140-170 BPM] Jungle](https://img.pdfslide.net/doc/110x75/6018bad90f937c130a7c6c52/80-100-125-150-170-bpm-styles-tempo-in-electronic-2019-3-6-dubstep.jpg)

















