Embed Size (px)
Citation preview

Behavioural Processes 68 (2005) 145–163
Recognition of partially concealed leopards bywild bonnet macaques (Macaca radiata)
The role of the spotted coat
Richard G. Cossa,∗, Uma Ramakrishnanb, Jeffrey Schanka
a Department of Psychology, University of California, Davis, CA 95616, USAb The Connecticut Agricultural Experiment Station, New Haven, CT 06504, USA
Received 24 September 2004; accepted 16 December 2004
Abstract
Wild bonnet macaques (Macaca radiata) have been shown to recognize models of leopards (Panthera pardus), based on theirconfiguration and spotted yellow coat. This study examined whether bonnet macaques could recognize the spotted and darkmelanic morph when partially concealed by vegetation. Seven troops were studied at two sites in southern India, the MudumalaiWildlife Sanctuary and the Kalakad-Mundanthurai Tiger Reserve. The forequarters and hindquarters of the two leopard morphswere presented from behind thick vegetation to individuals at feeding stations 25 m away. Flight reaction times and frequency offlight were obtained from video for only those individuals who oriented towards the models prior to hearing alarm calls. Bonnetm arter thana s ignoredc le of spotsa ntributed tol al features.©
K
1
t
f
these ofr the. Inres-
t de-shes
0
acaques exhibited faster reaction times and greater frequency of flight after looking at the spotted morph’s forequfter looking at either its spotted hindquarter or the dark morph’s forequarter. The hindquarter of the dark morph waompletely. Artificial neural network modeling examined the perceptual aspects of leopard face recognition and the ros camouflage. When spots were integrated into the pattern recognition process via network training, these spots co
eopard face recognition. When networks were not trained with spots, spots did not act as camouflage by disrupting faci2004 Elsevier B.V. All rights reserved.
eywords:Antipredator behavior; Leopards; Bonnet macaques; Artificial neural network; Pattern recognition
. Introduction
Research on the recognition of solitary felid preda-ors by mammalian prey has been hampered by
∗ Corresponding author. Tel.: +1 530 752 1626;ax: +1 530 752 2087.
E-mail address:[email protected] (R.G. Coss).
the rarity of encounters, their reclusiveness, andpropensity of these predators to hunt at night. The uvegetation as cover during the stalk is essential fosuccessful daytime hunting of large cursorial preysome contexts in which alarm calls announce the pence of a predator, such as the tiger (Panthera tigris),the uncertainty of predator whereabouts does noter hunting of large ungulate prey if grass and bu
376-6357/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.beproc.2004.12.004

146 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
provide sufficient cover to continue the stalk undetected(Thapar, 1999). This tactic is less successful for hunt-ing primates that typically forage on the ground neartrees (Stacey, 1986) or can maintain visual surveillanceof the predator’s location from their arboreal refuge(Zuberbuhler et al., 1999). This necessity to continu-ously monitor a predator’s location has engendered an-tipredator tactics in which some ungulates and primatesapproach felid predators to keep them in view (Bailey,1993; Boesch, 1991; Gandini and Baldwin, 1978). ForAfrican ungulates, vision plays an essential role for de-tecting and monitoring the activity of stealthy predatorsat a distance, especially when they are partly concealedby vegetation or elevated terrain (Baenninger et al.,1977; Caro, 1994; Stanley and Aspey, 1984).
2. Leopard recognition
The present study of wild bonnet macaques(Macaca radiata) in southern India examined whichperceptual features of leopards (P. pardus) were im-portant for predator recognition in microhabitats withthick vegetation that afforded some concealment. Ob-servations of bonnet macaques responding to leopards(Ali, 1981; Ramakrishnan and Coss, 2000) character-ize the diversity of daytime and nighttime microhab-itats in which leopards are encountered; these are notunlike the circumstances in which other primates arehunted by leopards (e.g.,Boesch, 1991; Busse, 1980;C ctq rdsa l.,1 ssfulh atricN nl
a -n yel-l re-v . As leop-a dis-t ardm re-s r theu of a
leopard standing, freezing briefly while looking at themacaques, and then disappearing from view. Resultsof this study revealed that the spotted upright modelwas the most provocative to bonnet macaques livingin forests where leopards are common and in an ur-ban site where leopards are absent. Comparisons of theupright and inverted positions of each morph providedevidence that spots on a yellow coat were still provoca-tive to bonnet macaques despite their appearance on theinverted felid form.
Consistent with the finding that the spotted yel-low coat was provocative, follow-up exploratory studyat one forest and one urban site examined the ef-fects of crouching leopard models constructed of light-brown towels with a leopard print or bluish-yellowtowels with a flower print. Only the towel modelwith the leopard print elicited alarm calling by bon-net macaques, prompting troop members in the forestto abandon their sleeping tree and mobbing by mem-bers of the urban troop on the top of a building. Al-though shaped the same as the spotted towel model, thetowel leopard with the flower print was ignored by bothtroops. Coupled with the finding that the inverted leop-ard model with spots was still provocative (Coss andRamakrishnan, 2000), these observations prompted thetheoretical conjecture that natural selection might haveoperated on the ability of bonnet macaques to detectleopards via visible patches of spots unobstructed byvegetation or rocks.
The ability to recognize leopards via their coat colora nec-d om-mc ob-s2 t ofC rveta tedt ,1
toi ofl iallyw undv n,e res,s olor,m ary
owlishaw, 1994; Isbell, 1990). Bonnet macaques reauickly by fleeing up trees when they detect leopand evidence from leopard scat (Ramakrishnan et a999) suggests that leopards are much less succeunters of bonnet macaques than they are of sympilgiri langurs (Trachypithecus johnii) and Hanuma
angurs (Semnopithecus entellus).Our initial research on leopard recognition (Coss
nd Ramakrishnan, 2000) examined whether wild bonet macaques differentiated the common spotted
ow morph from the rare dark melanic morph asealed by their alarm calling and flight responsesecond facet of this research examined whetherrd body configuration and coat texture afforded
inct recognition cues by presenting the two leoporphs in upright and inverted positions. Model p
entation employed a pop-up procedure which, fopright presentations, simulated the appearance
nd texture is not restricted to bonnet macaques. Aotal observations provide some evidence that con chimpanzees (Pan troglodytes) will mob or alarm
all after detecting realistic leopard models partlycured by vegetation (Kortlandt, 1967; Zuberbuhler,000). Hunters of vervet monkeys in the thick foresameroon locate their prey by capitalizing on velarm calling to the hunters’ ovoid headgear pain
o resemble the leopard’s spotted yellow coat (McRae997).
Study of the visual features used by preydentify leopards is complex due to the interplayeopard configuration and habitat features, espechen the leopard is partly obscured by foregroegetation. For primates with trichromatic visioven partial exposure of specific predator featuuch as the coupling of face, coat texture, and cight have acquired significance in the evolution

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 147
time frame as leopard-recognition cues if detection ofthese features afforded survival. On the other hand,yellowish vegetation during the dry season blends wellwith leopard coloration, a property which might forcereliance on the detection of the spotted coat texture. InSouth American primates in which only heterozygousfemales are trichromats (Smith et al., 2003), thespotted coat texture alone might provide a sufficientrecognition cue for identifying partially occluded fe-lids. For example,Herzog and Hopf (1986)found thatcinematic presentations of spotted textures with yellowbackgrounds moving briefly across a rectangular frameelicited alarm calling in captive squirrel monkeys(Saimiri sciureus). Such alarm calling might character-ize recognition of the spotted coats of felid predatorstypified byFelis tigrina,F. geoffroyi, andF. jaguarundi.
Similar to our research using a model of the darkleopard morph,Brown et al. (1992)presented a movingleopard silhouette to captive vervet monkeys (Chloro-cebus aethiops), eliciting alarm calling among severalmembers of the colony. However, the high rate of alarmcalling typical of individuals encountering leopards innatural settings (cf.Cheney and Seyfarth, 1990; Isbell,1990) was emitted by the only wild-caught and pre-sumably experienced individual in the colony. Lack ofalarm calling by inexperienced monkeys might simplyreflect the impoverish properties of the leopard silhou-ette possibly coupled with the effects of developmentaldeprivation due to captive rearing (seeStell and Riesen,1987; Struble and Riesen, 1978).
3
yel-l uest h ont rents peri-m rdsp tec-t ivedt of al thev f itsh
phw dark
morph (Coss and Ramakrishnan, 2000), we predictedthat partial body concealment by vegetation would notalter this difference in the two morphs. Our predictionsfor the effects of forequarter and hindquarter viewswere less confident. We knew from our field researchthat bonnet macaques are sensitive to the facing ori-entation of other troop members (Coss et al., 2002)and that the ability to recognize two facing eyes is anevolved trait in a variety of vertebrate taxa (Coss, 1978,1979a; Coss and Goldthwaite, 1995; Emery, 2000). Be-cause ambush predators typically use concealment af-forded by biotic and abiotic substrates while maintain-ing their visual fixation on prey, natural selection ismore likely to have shaped the recognition of two fac-ing eyes by prey under the stochastic circumstances inwhich the two eyes of the predator were still exposed.As such, two facing eyes might retain their provocativeproperties without surrounding facial features, an effectknown to occur in some mammals (Aiken, 1998; Coss,1970, 1978, 1979b; Hess, 1975; Topal and Csanyi,1994). From this perspective, we predicted that fore-quarter views showing the face would be more provoca-tive than hindquarters views. Yet for the spotted yellowmorph, the spots and flecks would likely reduce thecontrast of facial features and might disrupt face recog-nition, possibly yielding more equivalent provocativeeffects for forequarter and hindquarter views.
To explore this possibility further from a theoreti-cal perspective, we created an artificial neural network(ANN) as a simulation tool to investigate whether spotso andw itionb ofp risticv on-s atorr ysi-o
4c
4
4pril
a dia.
. Experimental questions and predictions
Our previous research showing that the spottedow morph was more provocative to bonnet macaqhan the dark morph prompted further field researche effects of partial leopard concealment. The curtudy of wild bonnet macaques addressed two exental questions involving the recognition of leopaartly exposed from behind vegetation: (1) Does de
ion of the spotted morph constitute a greater percehreat than the dark morph? (2) Does perceptioneopard’s forequarter with its face turned towardiewer engender greater alarm than perception oindquarter?
Since in full view, the upright spotted yellow moras shown to be more evocative than the upright
n the leopard’s face have camouflaging propertieshether dark pelage would hinder leopard recogny masking facial features. Thus, ANN modelingerceptual processes was expected to have heualue in pinpointing specific facial–feature relatihips that might prompt further field studies of predecognition and mechanistic studies of its neurophlogical underpinnings.
. Field research on recognition of partiallyoncealed leopards
.1. Methods
.1.1. Study sitesThe experiments were carried out between A
nd October 1997, at two study sites in southern In
148 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
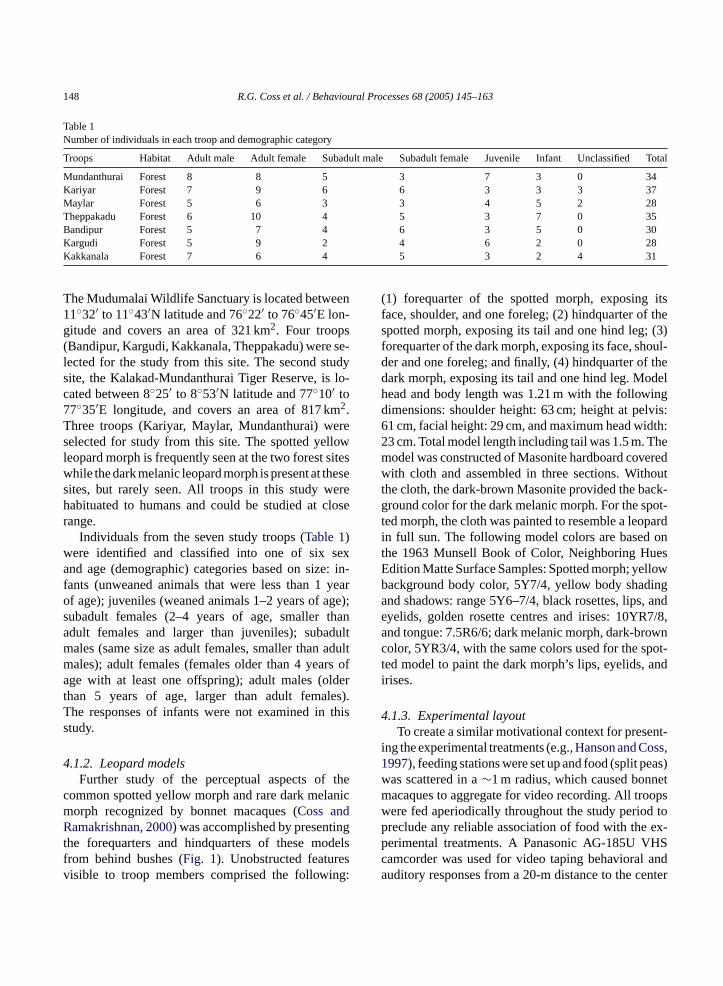
Table 1Number of individuals in each troop and demographic category
Troops Habitat Adult male Adult female Subadult male Subadult female Juvenile Infant Unclassified Total
Mundanthurai Forest 8 8 5 3 7 3 0 34Kariyar Forest 7 9 6 6 3 3 3 37Maylar Forest 5 6 3 3 4 5 2 28Theppakadu Forest 6 10 4 5 3 7 0 35Bandipur Forest 5 7 4 6 3 5 0 30Kargudi Forest 5 9 2 4 6 2 0 28Kakkanala Forest 7 6 4 5 3 2 4 31
The Mudumalai Wildlife Sanctuary is located between11◦32′ to 11◦43′N latitude and 76◦22′ to 76◦45′E lon-gitude and covers an area of 321 km2. Four troops(Bandipur, Kargudi, Kakkanala, Theppakadu) were se-lected for the study from this site. The second studysite, the Kalakad-Mundanthurai Tiger Reserve, is lo-cated between 8◦25′ to 8◦53′N latitude and 77◦10′ to77◦35′E longitude, and covers an area of 817 km2.Three troops (Kariyar, Maylar, Mundanthurai) wereselected for study from this site. The spotted yellowleopard morph is frequently seen at the two forest siteswhile the dark melanic leopard morph is present at thesesites, but rarely seen. All troops in this study werehabituated to humans and could be studied at closerange.
Individuals from the seven study troops (Table 1)were identified and classified into one of six sexand age (demographic) categories based on size: in-fants (unweaned animals that were less than 1 yearof age); juveniles (weaned animals 1–2 years of age);subadult females (2–4 years of age, smaller thanadult females and larger than juveniles); subadultmales (same size as adult females, smaller than adultmales); adult females (females older than 4 years ofage with at least one offspring); adult males (olderthan 5 years of age, larger than adult females).The responses of infants were not examined in thisstudy.
4the
c anicmR ingt delsf sv ng:
(1) forequarter of the spotted morph, exposing itsface, shoulder, and one foreleg; (2) hindquarter of thespotted morph, exposing its tail and one hind leg; (3)forequarter of the dark morph, exposing its face, shoul-der and one foreleg; and finally, (4) hindquarter of thedark morph, exposing its tail and one hind leg. Modelhead and body length was 1.21 m with the followingdimensions: shoulder height: 63 cm; height at pelvis:61 cm, facial height: 29 cm, and maximum head width:23 cm. Total model length including tail was 1.5 m. Themodel was constructed of Masonite hardboard coveredwith cloth and assembled in three sections. Withoutthe cloth, the dark-brown Masonite provided the back-ground color for the dark melanic morph. For the spot-ted morph, the cloth was painted to resemble a leopardin full sun. The following model colors are based onthe 1963 Munsell Book of Color, Neighboring HuesEdition Matte Surface Samples: Spotted morph; yellowbackground body color, 5Y7/4, yellow body shadingand shadows: range 5Y6–7/4, black rosettes, lips, andeyelids, golden rosette centres and irises: 10YR7/8,and tongue: 7.5R6/6; dark melanic morph, dark-browncolor, 5YR3/4, with the same colors used for the spot-ted model to paint the dark morph’s lips, eyelids, andirises.
4.1.3. Experimental layoutTo create a similar motivational context for present-
ing the experimental treatments (e.g.,Hanson and Coss,1 eas)w etm opsw d top ex-p HSc anda nter
.1.2. Leopard modelsFurther study of the perceptual aspects of
ommon spotted yellow morph and rare dark melorph recognized by bonnet macaques (Coss andamakrishnan, 2000) was accomplished by present
he forequarters and hindquarters of these morom behind bushes (Fig. 1). Unobstructed featureisible to troop members comprised the followi
997), feeding stations were set up and food (split pas scattered in a∼1 m radius, which caused bonnacaques to aggregate for video recording. All troere fed aperiodically throughout the study perioreclude any reliable association of food with theerimental treatments. A Panasonic AG-185U Vamcorder was used for video taping behavioraluditory responses from a 20-m distance to the ce

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 149
Fig. 1. Partially exposed leopard models presented to bonnet macaques. Top left: forequarter of spotted yellow leopard morph. Top right:forequarter of dark melanic leopard morph. Bottom left: hindquarter of spotted yellow leopard morph. Bottom right: hindquarter of dark melanicleopard morph that was ignored completely by all subjects.
of the feeding station. Camera field of view encom-passed the entire feeding area.
Four of the seven troops were exposed to more thanone leopard view with Mundanthurai and Bandipurtroops exposed to three views. For these troops, thefour partly exposed leopard morphs were presented ina random order, with minimum and maximum intervalsbetween presentations of 4 and 12 days, respectively.Experiments were conducted between 06:00a.m. and10:00a.m. and between 03:00p.m. and 05:00p.m., cor-responding to the peak foraging periods of this species.Video recording was initiated after the animals arrivedat the feeding station. After 2 min of video recording,the forequarter or hindquarter of one of the two leopard
models was presented at a distance of approximately25 m to monkeys gathered at a feeding station. Forthese presentations, the assistant, hidden behind thickvegetation, removed the green cloth envelope used fortransporting the model. On cue, the assistant movedthe front or rear section of the model forward into viewand withdrew the model after the monkeys respondedby flight and alarm calling or after a 1-min intervalif there was no response. This procedure simulated aleopard emerging from behind a bush into partial viewof the monkeys, freezing and then retreating from viewafter being detected (Fig. 1). Video recording contin-ued for 3 min after the each model was no longer inview.

150 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
4.1.4. Behavioral measures and statisticalanalyses
A flight reaction time measure was calculated as theinterval between lifting or turning the head in the direc-tion of the model and initiation of flight. Reaction timescould not be obtained from individuals already lookingin the direction of the models when they were movedinto view. These individuals were not included in thisand the subsequent analyses. Rather than exhibiting anormal distribution, reaction times are typically skewedto the right due to physiological limitations on the as-sessment and recognition of visual information (Rogalet al., 1985). Therefore, nonparametric tests were ap-plied to the data. We used survival analyses (Gail et al.,1980; Gehan, 1975) with log-ranked tests on pairwisecomparisons to measure differences in flight reactiontimes after individuals detected the models. Individu-als were censored if they did not flee within the 1-minsampling period. We employed multinomial log–linearanalyses (Agresti, 1990) to examine the interaction ofleopard models and the proportion of individuals thateither fled or stayed within camera view.
4.1.5. Monte Carlo simulations of larger subjectpool
A basic problem faced by field research in animalbehavior concerns the exact identification of animals.In some cases, we have exact information with whichto identify individuals, but in many other cases we onlyhave partial information. In this study, we could not ex-a di-t /agec sim-u ands matet l as ap itht siblet theK testg(
4
rphf inh had
attended to the model. Although this model might havebeen looked at briefly via glances undetected on video,the absence of relevant antipredator behavior precludedstatistical comparisons of the hindquarter of the darkmorph with the other leopard views. As a consequence,the statistical null hypothesis (not the theoretical hy-pothesis) was that animals do not respond differentiallyto the three remaining leopard models.
4.2.1. Analysis of the synthetic data setTo determine both the range and probability of re-
sampling across test conditions and to determine theconsequences of resampling on the probability of aType I error, Monte Carlo simulation was used to con-struct synthetic data sets based on the original data set.This was accomplished as follows. For all 42 obser-vations, we had information on the troop and sex/ageclass. FromTable 1, we knew how many individuals be-longed to each troop and age/sex class within troops.Thus, within each of the three leopard model condi-tions, individuals were drawn randomly from the cor-responding troop and age/class without replacement.Between leopard model conditions, individuals weredrawn with replacement. Thus, resampling of individ-uals was possible between conditions.
For each individual, we assumed that it was a re-sponder (and have a measured latency to leave) or anonresponder (indicated by a censored data point). Theprobability of responding was 19/42 based on the fre-q auser datas t over
F ionsu odels(
ctly identify individuals across leopard-model conions. However, we did know the troop and the sexlasses and this allowed us to use Monte Carlolation of synthetic data sets based on the troopex/age class of each individual observed to estihe frequency range of possible resampling as welrobability distribution for this range. In addition, w
he synthetic data sets thus generated, it was poso do a robustness analysis of the application ofruskal–Wallis nonparametric analysis of varianceeneralized to survival analysis data using theMantel1967)method for ranking the data (Lee, 1980).
.2. Results
Presentation of the hindquarter of the dark moor the maximum of 1 min failed to elicit changesead or body posture to indicate that individuals
uency of responding in the actual data set. Becesponding was treated as a probability, any givenet could have more or less than 19 responders, bu
ig. 2. A comparison of the empirical and theoretical distributsed for animals that were assumed to respond to leopard mbased on 100,000 simulations).

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 151
many replications, the frequency of responders (or non-responders) was normally distributed about 19/42 (bythe central limit theorem).
The data collected were either uncensored (“re-sponded to a leopard model”) or censored (“did notrespond to a leopard model”). For those animals thatresponded, the distribution of latencies to respond wasmodeled by a truncated normal distribution of laten-cies because it was likely more similar to the actualdistribution than, for example, a Poisson distribution.If a randomly generated latency fell in the truncatedregion, a new random normal latency was generated.It was assumed that the fastest latency was 0.08 s. The
truncated normal distribution was generated with aninitial mean of 55 and standard deviation of 85, whichproduced a truncated normal distribution with a meanof 94.4, very close to the data mean of 94.1 as illustratedin Fig. 2.
The simulation program used was written in Code-warrior 6.0 for the Macintosh computer using ANSI C.Synthetic data sets were constructed as just describedfor each replication and the frequency of resamplingwas calculated for the data set. Ten million simulatedsynthetic data sets were generated yielding the fre-quency distribution of probable resampling frequenciesillustrated inFig. 3A. As illustrated, the most likely
Fr1I
ig. 3. (A) Frequency distribution of possible resampling frequencieange of possible resampling across conditions, with the most likely re0,000,000 Monte Carlo generated synthetic data sets and survival anerror rate due to the increasing homogeneity of individuals across te
s during multiple model presentations. The double arrow indicates thesampling frequency at about 14%. (B) Plots of 0.05 and 0.01�-levels foralyses. As can be seen clearly, the effect of resampling is to lower the Typest conditions.

152 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
frequency of resampling was about 14% with a rangeof 5–40%.
With these synthetic data sets, survival analysiscould be applied to determine the effects of resamplingon Type I errors. Using the Kruskal–Wallis ANOVAgeneralized to survival data by the Mantel scoring pro-cedure (Lee, 1980), we plotted theP-values for syn-thetic data sets for�-levels of 0.05 and 0.01. We foundthat resampling did have an effect on Type I errorrate, a property that decreased as resampling increased
(Fig. 3B). This implies that resampling in this study didnot increase the likelihood of a Type I error; rather, itlikely decreased it.
4.2.2. Analysis of the actual data setAnalysis of the interval between model detection
and the onset of flight, using the Kaplan–Meier esti-mate of the survivor function (Fig. 4A), showed thatthe three views of the spotted and dark morph dif-fered at a statistically significant level (χ2 = 18.146;
Fct
ig. 4. (A) Latency to flee after looking at the models as the proportensored if they did not flee within the 1-min sampling period, truncatehe models.
ion of individuals that have not fled at a specific time. Individuals wered graphically at 26 s. (B) Percentage of individuals fleeing after observing

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 153
d.f. = 2; P< 0.0005). The forequarter of the spottedmorph (N= 11) elicited a flight reaction time thatwas significantly faster than those elicited by itsspotted hindquarter (N= 16, log-ranked test = 3.265;P< 0.0025), and the dark morph’ forequarter (N= 15,log-ranked test = 3.653;P< 0.0005). Unlike the spot-ted forequarter, the flight reaction time after seeing thespotted hindquarter was not significantly different thanthat after seeing the forequarter of the dark morph (log-ranked test = 0.429;P= 0.668).
Multinomial log–linear analyses (Fig. 4B) were em-ployed to examine the proportions of individuals thatfled after looking at the three views of the two morphs(Table 2). The interaction between models and the fre-quency of flight was statistically significant (likelihoodratio χ2 = 13.869; d.f. = 2;P< 0.001). Differences inthe proportion of individuals fleeing from each modelfollowed the same trend as differences in flight reactiontimes, with the largest proportion of bonnet macaquesfleeing after they looked at the forequarter of the spot-ted morph. As such, statistically significant interac-tions with very large standardized effect sizes appearedfor the proportion of individuals that fled after theylooked at the spotted forequarter and the proportionsof individuals that fled after they looked at the spot-ted hindquarter (likelihood ratioχ2 = 10.519; d.f. = 1;P= 0.001,d= 1.65) and dark morph’ forequarter (like-lihood ratioχ2 = 11.790; d.f. = 1;P< 0.001,d= 1.89).The proportion of individuals fleeing after looking atthe spotted hindquarter was not significantly differ-e thedd
4.2.3. Post hoc comparisons of partially and fullyexposed leopard models
The reaction times of individuals after they orientedtoward the partially exposed leopard models clearlyrevealed differences in the excitatory effects of modelassessment. However, the generality of these findingis limited to this particular experimental context. Aninteresting question then is whether bonnet macaquesreact to partially exposed leopards in a manner simi-lar to when they detect leopards in full view. Such acomparison can be made post hoc because, in order todetermine the onset of model exposure for measuringreaction time, individuals were only sampled if theyhad looked up or turned their heads in the direction ofthe modelsafter the models had been positioned intoview. Thus, the marked differences in the dynamics ofpresenting pop-up models in full view for 10 s (Cossand Ramakrishnan, 2000) and the method of presentingthe forequarters and hindquarters of the models frombehind a bush in the present study were irrelevant fordetermining individual reaction times (seeTable 2).Differences in the time interval between the two exper-iments and composition of troops further reduced thelikelihood of resampling.
Pairwise comparison of the full view and forequarterview of each morph revealed that the prepotent proper-ties of these views did not differ appreciably (Fig. 5).Sight of the spotted morph in full view (N= 11) elicitedflight reaction times that were significantly similar(e.g., reliably identical) to the flight reaction times ofi re-qt ight
TN rd mod
L Sub Total
S 0 00
S 1 50
D 00
S 00
D 10
T embe r analyses.makris
nt from the proportion fleeing after looking atark morph’s forequarter (likelihood ratioχ2 = 0.079;.f. = 1;P> 0.5).
able 2umber of individuals who looked at partially concealed leopa
eopard views Flight Adult male Adult female
potted forequarter Yes 3 3No 1 0
potted hindquarter Yes 0 3No 7 4
ark forequarter Yes 1 2No 5 4
potted in full viewa Yes 5 4No 0 0
ark in full viewa Yes 2 5No 5 1
he hindquarter of the dark morph was not looked at by troop ma Leopard models that popped up into full view inCoss and Ra
ndividuals (N= 11) who looked at the spotted fouarter (log-ranked test = 0.034;P= 0.973). Although
he dark morph was less provocative overall, the fl
els after the models were positioned into view
adult male Subadult female Juvenile Unclassified
2 2 0 10 0 0 1
1 0 00 0 0 11
0 1 0 41 0 1 111 1 0 110 0 0 00 0 0 80 0 0 6
rs in video view. Total proportions were examined by log–lineahnan (2000)for post hoc comparisons of reaction times.

154 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
Fig. 5. Latency to flee after looking at models of the forequarters ofthe spotted and dark morphs compared will fully exposed leopardmodels (fromCoss and Ramakrishnan, 2000). The forequarter andfully exposed spotted leopard models elicited significantly similarreaction times. Graph is truncated at 26 s.
reaction times after individuals saw its full view(N= 14) and forequarter (N= 15) were not signifi-cantly different (log-ranked test = 1.398;P= 0.162).On the whole, these findings suggest that the completebody and forequarter views of the same leopardmorphs are similarly provocative.
4.3. Discussion
Models of the spotted yellow and dark melanic leop-ard morphs were presented briefly to bonnet macaquesas partly exposed views of the leopard’s forequarterand hindquarter. Such comparisons provided insightinto which perceptual features of leopards were im-portant for predator recognition in microhabitats withthick vegetation that afforded some concealment. Thefrequency of flight of individuals in different troopsthat looked at the these models provided evidence thatthe forequarter view of the spotted yellow morph wasperceived as much more threatening than either itshindquarter or the forequarter view of the dark morph.Despite this difference in flight elicitation, it must benoted that at least one individual alarm called fromarboreal refuge in every troop exposed to these threeleopard views. With exception of the hindquarter ofthe dark morph that was ignored completely, detectionof motionless models by individuals in video viewwas hindered by the early emission of alarm callsfrom individuals already in trees or by the sight ofn hoh they
were first exposed, events which typically triggeredimmediate flight without pauses to scan for the sourceof the disturbance (cf.Coss and Ramakrishnan, 2000;Ramakrishnan and Coss, 2000). As such, reaction timedata were typically restricted to a small number ofindividuals in each troop who were the first to detectthe models (Table 2).
4.3.1. Role of the forequarter and face in leopardrecognition
It is reasonable to assert that the primary featuresfor leopard recognition are exhibited by the leopard’santerior portion and spotted yellow coat. The most pre-dominant finding supporting this supposition was themuch faster reaction time and higher frequency of flightelicited by the forequarter of the spotted morph com-pared with its hindquarter. Only one individual from alltroops (Table 2) failed to flee after looking at the fore-quarter of the spotted morph, a frequency of flight thatapproaches the 100% flight engendered by the spottedmorph in full view (Coss and Ramakrishnan, 2000).Also, the reaction times elicited by the spotted fore-quarter were nearly identical to those engendered bythe spotted morph in full view (Fig. 5). The lack of re-sponse to the hindquarter of the dark morph, comparedwith its forequarter, also supports the argument that theessential information for assessing the leopard’s preda-tory threat is exhibited by the perceptual features of theleopard’s forequarter, which includes the face. Whilebonnet macaques exhibited a complete lack of atten-t thed coatc nfig-u thef on-s evi-d to thec ndV heu cog-n rds,b rapidp sim-i on-s hent ex-p l.,1
eighboring monkeys running on the ground wad detected the models by their motion as
ion and vigilance directed at the hindquarter ofark morph, the presence of the spotted yellowhanged this apparently irrelevant hindquarter coration into a relevant one, equivalent to that of
orequarter of the dark morph. This parity in respiveness might also reflect the neurophysiologicalence showing that macaques are most sensitiveolor yellow (Yoshioka and Dow, 1996; Yoshioka aautin, 1996), a property which might contribute to tnderlying neurological processes of leopard reition. Thus for recognizing partly exposed leopaonnet macaques would not need to engage in theerceptual operations of surface completion using
lar surface fragments or edge interpolation to rectruct the entire image hypothesized for humans whey recognize partly occluded patterns in learnederimental tasks (Sekular and Palmer, 1992; Yin et a997).

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 155
5. Artificial neural network modeling ofleopard face recognition
Reliance on specific morphological features forleopard recognition, rather than the necessity of seeingthe whole image before recognition occurs, is consis-tent with the properties of the innate perceptual systemsof other species (Coss, 1991, 1999; Curio, 1975, 1993).This reliance on specific cues is best demonstrated bythe perceptual aspect of two facing eyes, which havebeen available historically as a visual cue that the per-ceiver is being watched (Coss and Goldthwaite, 1995).The ability to recognize two facing eyes operates in-dependent of other facial features (Coss, 1970, 1978;Perrett et al., 1982), in part, because the primary sourceof selection shaping this ability centers on the successof assessing the direction of gaze of both conspecificsand predators. As mentioned above, stealthy predatorsusing visual obstruction to approach prey have to ex-pose their eyes to monitor prey activity from cover,thereby revealing this reliable visual cue that prey arebeing watched. Two facing eyes are also provocativeunder low-contrast conditions if there is sufficient lightto detect the eyes and surrounding eyerings (Coss,1978) or under nighttime conditions, as suggested byexperimental simulation (Topal and Csanyi, 1994), ifthe eyes shine via moonlight reflectance on the tapetum.
Consistent with our experimental manipulations inthe field, our ANN simulations addressed the issueof whether the dark face of the rare melanic leop-a gni-t rph.T issueo mou-fl re-gan aleA
5
iche redi rtex( ls eciest icala ects
of distributed inputs and connectivity from adjacentand nearby neural columns are amenable to computersimulations (seeMiikkulainen et al., 1998). Thus in atheoretical domain, ANN simulations can complementthe aforementioned empirical research on bonnetmacaque responses to the spotted and dark leopardmorphs by examining whether facial spots and flecksdisrupt leopard face recognition.
The ANN simulator (tlearn) developed byPlunkettand Elman (1997)was used to construct a neuralnetwork and train it using back-error propagation forpattern classification tasks. In the backpropagationprocedure, network learning is accomplished bynumerous iterations of forwards and backwards prop-agation steps (Rumelhart et al., 1986). During eachforwards step, the input pattern generates an outputpattern, which is compared with a desired target pat-tern. In the backwards step, error computations fromthis output-target comparison are propagated downthrough the network to adjust connection weights;the global discrepancy between these forwards andbackwards activations is presented as a single rootmean square (RMS) error (Plunkett and Elman, 1997).In the current set of experiments, the number ofactivation sweeps selected to train the network wasdetermined by the number of patterns in the trainingset and the desire to obtain RMS errors that approachedzero in the output-target pattern comparison.
The network’s ability to distinguish the target pat-tern from novel patterns outside the training set wasd singa n tog aredw reeo r,2 st-i of ab ut,a ition( un-d extf emai -l c-t cog-n sin-g ningr rks
rd morph truly compromises leopard-face recoion by macaques experienced with the spotted mohese simulations also addressed the theoreticalf whether spots adjacent to the leopard’s eyes caage the leopard’s face by either masking the eyeion or disrupting eye-schema recognition (cf.Gavishnd Gavish, 1981; Ortolani, 1999) for prey historicallyaıve to leopards, but not to large felids, such as femsiatic lions (P. leo).
.1. Methods
Extensive networks of clustered neurons, whxhibit partially redundant processing of sha
nformation, are ubiquitous in mammalian neocoFujita et al., 1992; Lund et al., 1993). In the visuatream of domestic cats and macaques, the sphat are the primary subjects of electrophysiolognd neuroanatomical studies, the functional eff
etermined by testing each pattern separately usingle forwards step of output-target compariso
enerate its RMS error. This RMS error was compith the target pattern’s RMS error to infer the degf pattern generalization (seeBasheer and Hajmee000). It is important to note that this nonlearning te
ng procedure simulated the recognition processiological neural network attuned to a specific inpproperty that occurs in innate predator recogn
Coss, 1999). Thus for this state of adaptation, theerlying biological neural network awaits the cont
or its specific activation when the appropriate schs encountered (seeCoss, 1993). However, unlike bioogical neural networks, which exhibit multiple funional states subserving a wide range of pattern reition tasks, the ANN described herein exhibited ale awaiting state that characterized its specific traiegime. Also analogous to biological neural netwo

156 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
from this state perspective, the global RMS error canbe viewed as the emergent property of the entire neu-ral network’s image processing, reflecting the arrange-ment of network connectivity and connection weights(Gochin, 1996).
5.1.1. Artificial neural network architectureThe ANN architecture consisted of 1210 nodes, or-
ganized as three hidden layers sandwiched betweeninput and output layers (Fig. 6). These hidden layers
consisted of a 13× 13 array of nodes, each with a non-linear response property characterized by a “squash-ing” sigmoidal slope in activation function. Computa-tional descriptions of the network activation functionand weight adjustment with backpropagation appear inPlunkett and Elman (1997). The first two hidden layersreceived pattern input from a 14× 14 lattice of pixelswith luminance values ranging for 0 to 0.9. Each nodein these hidden layers received luminance inputs fromtwo adjacent pixels in the input lattice, as dominoes ar-
FFl
ig. 6. Neural network architecture of a portion of the tlearn simulatoror the whole network, feedforward connectivity with adjustable weigh
ines) within each hidden layer is accomplished by 506 nonlearning c
is shown for three overlapping input vectors centered in the input array.ts (solid lines) is provided by 703 nodes. Internodal connectivity (dashed
opy-back nodes (not shown).

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 157
ranged in either the vertical plane (input to hidden layer1) or horizontal plane (input to hidden layer 2). Thispairing of luminance inputs to each node yielded smallvertical or horizontal receptive fields. To enhance edgecontrast, each node in the first and second hidden layersreceived input from adjacent nodes via a single “copy-back” node with nonadjustable connections (Plunkettand Elman, 1997). In this arrangement, each copy-backnode acted as a linear filter, restricting luminance in-formation from surrounding nodes to 33% of the totalluminance input to each node, a property approximat-ing the percentage of suppression of texture surroundson receptive field activity in area V1 of the macaque vi-sual cortex (Knierim and van Essen, 1992; Nothdurft etal., 1999). Nodes in the first and second hidden layersprojected to their topographic counterparts in hiddenlayer 3 and their adjacent nodes. The dispersion of 4–9feedforward connections to hidden layer 3 from eachnode in hidden layers 1 and 2, yielded a tessellationof vertical and horizontal receptive fields, providing abiomimetic analog to the visual stream described forcat striate cortex (Gilbert and Wiesel, 1989; Ts’o et al.,1986) and macaque visual cortex (Lund et al., 1995).Within hidden layer 3, each node exhibited, via dis-tinct copy-back nodes, feedforward connections withadjacent nodes in the horizontal plane and with nonad-jacent nodes in a vertical radial pattern (Fig. 6), roughlyemulating the topography of intercolumnar connectiv-ity within macaque inferotemporal and prefrontal cor-tex (Fujita and Fujita, 1996; Levitt et al., 1993). Forn erali ldedl in-p ioni par-t mnsi ;T ro-jo andr di-t tputl f theo
5gni-
t cale
Fig. 7. Tests of neural network learning for two vector sets used innetwork training. Note the low root mean square (RMS) error for thetarget pattern compared with the other training patterns within eachrespective vector set.
target pattern was developed depicting a generic spot-ted leopard face (with pixel luminance values rangingin 0.1 increments from 0.0 for black spots, black nos-trils, and black eyerings, to 0.7 for whitish eye andmuzzle patches). The background framing the muz-zle was 0.9 luminance value. A second target patternexhibited the same pixel configuration and luminancevalues except that all facial spots were replaced by pix-els with 0.5 luminance values. This schematic imageresembled that of a female lion. Two vector sets wereconstructed (Fig. 7) in which the outputs of these spot-ted and unspotted target patterns were compared duringeight 1000 sweep epochs with the outputs of their re-spective spotted and unspotted patterns exhibiting twoschematic eyes in vertical and diagonal planes. Thechoice of these contrasting patterns for training eye-schema recognition as a component of leopard facerecognition was based on empirical research on humansand other mammals showing that patterns with twovertically and diagonally positioned eyes were muchless provocative than patterns with two facing eyes inthe horizontal plane (Aiken, 1998; Coss, 1970, 1978,
odes in hidden layer 3, the combination of collatnputs and those from the hidden layers 1 and 2 yiearge receptive fields, spanning up to 9 pixels in theut lattice. This dispersion of luminance informat
n hidden layer 3 was analogous to the spread ofial pattern information among adjacent neural colun macaque inferotemporal cortex (Fujita et al., 1992anaka, 1996). Finally, these hidden layer 3 nodes p
ected to their topographic counterparts in a 14× 14utput layer, with the exception of nodes in the topight side of hidden layer 3 which projected to an adional output node in the top and right side of the ouayer. This output layer thus afforded comparison output and target patterns.
.1.2. Artificial neural network trainingTo address the questions of leopard-face reco
ion and the effects of facial camouflage, a grays

158 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
1979b; Topal and Csanyi, 1994). Also, at the level ofsingle units in neural columns in macaque inferotempo-ral cortex, facial configurations engendering maximumneural responsiveness were determined by empiricalsimplification of their provocative properties (Tanaka,1996). This process of simplification yielded an opti-mal schematic face pattern with two facing eyes in thehorizontal plane remarkably similar to those used inbehavioral research (cf.Altbacker and Csanyi, 1990;Coss, 1978, 1979b; Topal and Csanyi, 1994).
For network training using the tlearn simulator,teacher forcing was employed, with learning rate andmomentum = 0.3 and 0.9, respectively, and initial seed-ing = 1.0, with random training without replacement.After this procedure, network learning was revealed bypresenting each pattern in the vector set separately tothe network input lattice for one nontraining test sweepand examining its RMS error. Testing of the patternsin their respective training vector sets (Fig. 7) showedthat the outputs of spotted and unspotted faces with twohorizontally positioned eyes yielded the lowest RMSerrors. These low RMS errors for the target patternsreflect the specificity of successful ANN learning.
While testing patterns in the vectors sets character-ized target-pattern learning, the examination of target-pattern generalization to related novel patterns couldbe accomplished by developing a series of spottedfaces, unspotted faces, and dark faces depicting lin-ear changes in the number of eyes. Again, similar pat-terns varying in the number of eyes have been usedin behavioral studies of eye-schema recognition (cf.Altbacker and Csanyi, 1990; Coss, 1978, 1979b; Topaland Csanyi, 1994).
The perceptual differences of the spotted and darkmelanic morphs could be evaluated in simulations us-ing the network trained on the spotted target patternwith two horizontally positioned eyes. Comparison ofthese spotted and dark faces is roughly analogous to theaforementioned experimental presentations of the spot-ted and dark leopard forequarters to bonnet macaquesin the field. The role of dark spots in leopard facerecognition could be evaluated by comparing the ef-fects of spot removal using the network trained onthe spotted target face. Conversely, the question ofwhether dark spots camouflage the face could be eval-uated using the network trained on the unspotted targetface and presenting the series of novel faces with darkspots.
6. Results
6.1. Comparison of faces of spotted and darkleopard morphs
This simulation experiment compared the effective-ness of the network in recognizing its target pattern, thespotted face with two facing eyes, with that engenderedby the novel spotted and dark faces with different num-bers of eyes. The lowest and next to the lowest RMS er-rors for the two V-shaped generalization gradients arecentered, respectively, over the spotted target pattern(spotted leopard face) and the novel face of the darkmorph sharing the schema of two facing eyes (Fig. 8).It is important to note that the RMS error for the faceof the dark morph was nearly double that of the spot-ted target pattern. Nevertheless, the RMS error of thedark morph’s face was still substantially less than theRMS errors generated by the spotted and dark patterns
F orkw acinge otted( com-b char-a earlyd .
ig. 8. Generalization gradient of RMS errors when the netwas trained to recognized the spotted target pattern with two fyes and tested on this pattern and a continuum of novel spcrosshatched bars) and dark patterns (dark bars) with differentinations of eyes. The novel dark pattern with two facing eyes,cterizing the dark leopard morph, produced an RMS error nouble of that of the spotted target pattern with two facing eyes

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 159
with other eye arrangements. That is, darkening of theface by 0.4 luminance to reduce spot conspicuousnessreduced, but did not disrupt face recognition. On thewhole, the generalization gradient is most symmetricalfor the spotted series and appears remarkably similarto that generated from empirical study of adaptive eye-schema recognition (cf.Coss, 1978, 1979a).
6.2. Comparison of spotted and unspotted faces
The second simulation experiment tested the effectsof removal of the dark spots on leopard-face recogni-tion using the network trained on the spotted face. Be-cause of the physiognomic similarity of faces withinthe genusPanthera, this simulation could be viewedas analogous to leopard-experienced macaques detect-ing the novel face of a female lion. Removal of thedark spots on the novel face with two facing eyes pro-duced an RMS error substantially larger than that of thetrained spotted target face and nearly equivalent to thatgenerated by the spotted face with one eye (Fig. 9). De-spite this difference, there were similarities in the RMSerrors of the spotted and unspotted series of patterns,notably the relatively symmetrical V-shaped general-ization gradients with the lowest RMS error centeredfor each gradient on the face with two horizontally posi-tioned eyes. The reduction of network recognition withthe removal of darks spots demonstrates the integrationof the dark spots and two facing eyes when these spotsare part of the network training regime.
6
fac-i novelp theu ula-t ruptt ori-c twof oure ts off do t oft vir-tc spot-t d V-
Fig. 9. Generalization gradient of RMS errors when the networkwas trained to recognized the spotted target pattern with two facingeyes and tested on this pattern and a continuum of novel spotted andunspotted patterns. Removal of spots on the novel pattern with twofacing eyes produced a substantial increase in the RMS error, indi-cating a marked diminution of network recognition of this pattern.
shaped generalization gradients that were remarkablysimilar, also indicating that spots had little effect onpattern recognition. For the ANN architecture devel-oped herein, contrasting spots became important onlyif the network was trained to “expect” them as an inte-gral facial feature, whereas the addition of spots addedlittle additional information if the network was trainedto recognize two facing eyes on a backdrop of mod-erately contrasting facial features. A similar processmight occur in biological systems if natural selectionhas operated consistently on the prey’s ability to dis-tinguish the two facing eyes of ambush predators frompartially obscuring foreground vegetation.
7. Discussion
On the whole, our ANN modeling of leopard facerecognition yielded simulations that afforded some in-sights for interpreting experimental presentations of the
.3. Spots as facial camouflage
The pattern of dark spots on the face with twong eyes (spotted leopard face) was presented as aattern to the input lattice of the ANN trained onnspotted target pattern (lion-like face). This sim
ion is analogous to testing whether dark spots dishe face-recognition abilities of prey species histally attuned to the provocative qualities of theacing eyes of unspotted carnivores. Contrary toxpectations based on the aforementioned effecacial darkening, theadditionof dark spots producenly a slight elevation in the RMS error above tha
he target pattern, indicating that these spots hadually no camouflaging properties (Fig. 10). Furtheromparisons of the entire series of spotted and uned patterns with different numbers of eyes reveale

160 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
Fig. 10. Generalization gradient of RMS errors when the networkwas trained to recognize the unspotted target pattern with two facingeyes and tested on this pattern and a continuum of novel patterns withand without dark spots. The addition of spots to the novel test patternwith two facing eyes produced only a slight increase in the RMS error,indicating strong network generalization in pattern recognition.
spotted and dark morph’s forequarters to wild bonnetmacaques. For example, when the networks was trainedon the spotted target pattern with two facing eyes, dark-ening of the face produced a moderate mismatch whichnearly doubled the RMS error, but not to the extent ofthat of novel patterns with fewer or larger numbers ofeyes (Fig. 8). This mismatch can be best explained bythe loss of conspicuousness of the black spots and eye-rings on the dark schematic face. Further evidence thatcontrasting spots and eyerings were cohesively linkedas critical features for network recognition was appar-ent when the black spots were replaced by pixels of amedium luminance value (Fig. 9), thus characterizingthe novel schematic face of a female lion. The absenceof black spots increased the RMS error substantially, in-dicating a reduction of overall pattern coherence; albeit,this absence of spots did not compromise eye-schemarecognition completely as evinced by the lowest RMSerror in the V-shaped generalization gradient centeredon the unspotted face with two facing eyes.
Training the ANN on the unspotted target imageyielded unexpected results when, during testing, novelspots as black pixels replaced those of medium lu-minance value. Under this training regime, the ANNmaintained the ability to distinguish spotted patternsthat differed in number of eyes nearly as well as thatelicited by the unspotted patterns (Fig. 10). This find-ing contrasts with the effects of facial darkening whichdid impact face recognition, but likely characterizes themost prevalent adaptive context in which two facingeyes connote the exhibitor’s interest in the perceiveressential for risk assessment (Coss, 1978; Coss andGoldthwaite, 1995; Emery, 2000). From this simula-tion, it is reasonable to consider that dark spots smallerthan the eyes and dark eyerings would still permit theemergence of face recognition even though spots re-duce the contrast of facial features markedly. In hu-mans, for example, face-recognition performance is notdegraded until the contrast of facial images drops be-low 90% (Avidan et al., 2002). However, disruption offace recognition is likely to occur when larger patchesare paired bilaterally as in the dark muzzles patches ofsome carnivores. These patches have perceptual qual-ities like the additional pair of eyes used in the afore-mentioned test patterns with four eyes and may indeedhave camouflaging properties (Ortolani, 1999) similarto the effects of beards that can disguise familiar humanfaces (Patterson and Baddeley, 1977; Terry, 1994).
8
be-c ex-p pectf ardf tiont ema.S icals ed byn suals pledl eg-e calf
re-q s-h the
. General discussion
Our ANN modeling showed that facial spotsame relevant only when the ANN was trained toect them. When the network was not trained to ex
acial spots, the addition of spots did not disrupt leopace recognition because it added no new informahat altered the relationships of the perceptual schuch expectation might operate similarly in biologystems in which the visual system has been shapatural selection to await exposure to specific vichemata. For example, the face of a lion in dapight with leaf shadows or behind a spotty veil of vtation might still be recognized by prey if the criti
eatures of the lion’s face are not obstructed.Since previous exposure to leopards is not
uired for leopard recognition (Coss and Ramakrinan, 2000), the findings of this study suggests that

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 161
visual system of bonnet macaques is attuned to the in-variant perceptual features of the leopard’s head regionand its spotted yellow coat, both of which might facil-itate the detection and recognition of leopards partlyconcealed by vegetation. For prey species like bonnetmacaques, any initial historical benefits to leopards ofcamouflage provided by rosettes, spots, and flecks havenow been circumvented by the evolution of perceptualsystems attuned to these patterns. Conversely, the re-cessive allele for the dark melanic coat (Sleeper, 1995)might be sustained in low frequency in leopard popula-tions simply because the absence of the spotted yellowcoat fosters faster prey habituation to leopards that re-main still in ambush mode for long periods (seeRice,1986).
Acknowledgements
This research was supported by Faculty Researchgrant D-922 to R.G. Coss and by the Foundationfor Ecological Research, Advocacy and Learning,Pondicherry, India, to U. Ramakrishnan. We thankthe Forest Department of Tamil Nadu for permis-sion to conduct research in the Kalakad-MundanthuraiTiger Reserve and Mudumalai Wildlife Sanctuary andtheir staff for facilitating our research. We also thankour field assistants, Anil Kumar, M. Siddhan, and V.Yashoda for their contribution in data collection andA. Dharawat and M. Park for their assistance in quan-t
R
AA spot
A nnetof
A da-fish
A E.,reas87,
B r be-ildl.
Bailey, T.N., 1993. The African Leopard. Columbia University Press,New York.
Basheer, I.A., Hajmeer, M., 2000. Artificial neural networks: funda-mentals, computing, design, and application. J. Microbiol. Meth.43, 3–31.
Boesch, C., 1991. The effects of leopard predation on grouping pat-terns in forest chimpanzees. Behaviour 117, 220–242.
Brown, M.M., Kreiter, N.A., Maple, J.T., Sinnott, J.M., 1992. Sil-houettes elicit alarm calls from captive vervet monkeys (Cerco-pithecus aethiops). J. Comp. Psychol. 4, 350–359.
Busse, C., 1980. Leopard and lion predation upon chacma baboonsliving in the Moremi Wildlife Reserve. Botswana Notes Rec. 12,15–21.
Caro, T.M., 1994. Ungulate antipredator behaviour: preliminaryand comparative data from African bovids. Behaviour 28, 189–228.
Cheney, D.L., Seyfarth, R.M., 1990. How Monkeys See the World.University of Chicago Press, Chicago, 377 pp.
Coss, R.G., 1970. The perceptual aspects of eye-spot patterns andtheir relevance to gaze behaviour. In: Hutt, C., Hutt, S.J. (Eds.),Behaviour Studies in Psychiatry. Pergamon Press, London, pp.121–147.
Coss, R.G., 1978. Perceptual determinants of gaze aversion by thelesser mouse lemur (Microcebus murinus), the role of two facingeyes. Behaviour 64, 248–270.
Coss, R.G., 1979a. Delayed plasticity of an instinct: recognition andavoidance of 2 facing eyes by the jewel fish. Dev. Psychobiol.12, 335–345.
Coss, R.G., 1979b. Perceptual determinants of gaze aversion by nor-mal and psychotic children: the role of two facing eyes. Behaviour69, 228–254.
Coss, R.G., 1991. Context and animal behavior. III. The relation-ship of early development and evolutionary persistence of groundsquirrel antisnake behavior. Ecol. Psychol. 3, 277–315.
Coss, R.G., 1993. Evolutionary persistence of ground squirrel anti-Psy-
C lutionVari-sms.
C signsthol-148.
C iron-onnet
C leop-
C ula-
C be-tion.
C dator
ifying video recordings.
eferences
gresti, A., 1990. Categorical Data Analysis. Wiley, New York.iken, N.E., 1998. Human cardiovascular response to the eye
threat stimulus. Evol. Cognit. 4, 1–12.li, R., 1981. The ecology and behaviour of the Agastyamalai bo
monkeys (Macaca radiatadiluta). Ph.D. Thesis, UniversityBristol.
ltbacker, V., Csanyi, V., 1990. The role of eyespots in pretor recognition and antipredator behaviour of the paradiseMacropodus opercularisL. Ethology 85, 51–57.
vidan, G., Harel, M., Hendler, T., Ben-Bashat, D., Zohary,Malach, R., 2002. Contrast sensitivity in human visual aand its relationship to object recognition. J. Neurophysiol.3102–3116.
aenninger, R., Estes, R.D., Baldwin, S., 1977. Anti-predatohaviour of baboons and impalas toward a cheetah. E. Afr. WJ. 15, 327–329.
snake behavior: reflections on Burton’s commentary. Ecol.chol. 5, 171–194.
oss, R.G., 1999. Effects of relaxed natural selection on the evoof behavior. In: Foster, S.A., Endler, J.A. (Eds.), Geographication in Behavior: Perspectives on Evolutionary MechaniOxford University Press, Oxford, pp. 180–208.
oss, R.G., Goldthwaite, R.O., 1995. The persistence of old defor perception. In: Thompson, N.S. (Ed.), Perspectives in Eogy 11: Behavioral Design. Plenum Press, New York, pp. 83–
oss, R.G., Marks, S., Ramakrishnan, U., 2002. Early envment shapes the development of gaze aversion by wild bmacaques (Macaca radiata). Primates 43, 217–222.
oss, R.G., Ramakrishnan, U., 2000. Perceptual aspects ofard recognition by wild bonnet macaques (Macaca radiata). Be-haviour 137, 315–335.
owlishaw, G., 1994. Vulnerability to predation in baboon poptions. Behaviour 131, 293–304.
urio, E., 1975. The functional organization of anti-predatorhaviour in the pied flycatcher: a study of avian visual percepAnim. Behav. 23, 1–115.
urio, E., 1993. Proximate and developmental aspects of antiprebehavior. Adv. Stud. Behav. 22, 135–238.

162 R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163
Emery, N.J., 2000. The eyes have it: the neuroethology, function andevolution of social gaze. Neurosci. Biobehav. Rev. 24, 581–604.
Fujita, I., Fujita, T., 1996. Intrinsic connections in the macaque in-ferior temporal cortex. J. Comp. Neuro. 368, 467–486.
Fujita, I., Tanaka, K., Ito, M., Cheng, K., 1992. Columns for visualfeatures of objects in monkey inferotemporal cortex. Nature 360,843–846.
Gail, M.H., Santner, T.J., Brown, C.C., 1980. An analysis of com-parative carcinogenesis experiments based on multiple times totumor. Biometrics 36, 255–266.
Gandini, G., Baldwin, P.J., 1978. An encounter between chimpanzeesand a leopard in Senegal. Carnivore 1, 107–109.
Gavish, L., Gavish, B., 1981. Patterns that conceal and bird’s eye. Z.Tierpsychol. 56, 193–204.
Gehan, E.A., 1975. Statistical methods for survival time studies. In:Staquet, M.J. (Ed.), Cancer Therapy: Prognostic Factors and Cri-teria. Raven Press, New York, pp. 7–35.
Gilbert, C.D., Wiesel, T.N., 1989. Columnar specificity of intrinsichorizontal and corticocortical connections in cat visual cortex. J.Neurosci. 9, 2432–2442.
Gochin, P.M., 1996. The representation of shape in the temporal lobe.Behav. Brain Res. 76, 99–116.
Hanson, M.T., Coss, R.G., 1997. Age differences in the response ofCalifornia ground squirrels (Spermophilus beecheyi) to avian andmammalian predators. J. Comp. Psychol. 111, 174–184.
Herzog, M., Hopf, S., 1986. Recognition of visual pattern compo-nents in squirrel monkeys. Eur. Arch. Psychiatr. Neurol. Sci. 236,10–16.
Hess, E.H., 1975. The Tell-Tale Eye, How your Eyes Reveal HiddenThoughts and Emotions. Van Nostrand Reinhold Company, NewYork.
Isbell, L.A., 1990. Sudden short-term increase in mortality of vervetmonkeys (Cercopithecus aethiops) due to leopard predation inAmboseli National Park, Kenya. Am. J. Primatol. 21, 41–52.
Knierim, J.J., van Essen, D.C., 1992. Neuronal responses to staticNeu-
K ild.nisseVer-
L ife-
L ra-on-
338,
L trin-bral
L andhe-pp.
M ob-
M 4.
Miikkulainen, R., Bednar, J.A., Choe, Y., Sirosh, J., 1998. Proceed-ings of the Fifth International Conference on Neur. Informat.Process, Kitakyushu, Japan. A self-organizing neural networkmodel of the primary visual cortex.
Nothdurft, H.C., Gallant, J.L., Van Essen, D.C., 1999. Responsemodulation by texture surround in primate area V1: corre-lates of “popout” under anesthesia. Visual Neurosci. 16, 15–34.
Ortolani, A., 1999. Adaptive Coloration in the Carnivora. Ph.D. dis-sertation, UMI Order Number: AAT 9958637. University of Cal-ifornia, Davis.
Patterson, K.E., Baddeley, A.D., 1977. When face recognition fails.J. Exp. Psychol.: Hum. Learn. Mem. 3, 406–417.
Perrett, D.I., Rolls, E.T., Caan, W., 1982. Visual neurones respon-sive to faces in the monkey temporal cortex. Exp. Brain Res. 47,329–342.
Plunkett, K., Elman, J.L., 1997. Exercises in Rethinking Innateness:A Handbook for Connectionists Simulations. The MIT Press,Cambridge, MA, 313 pp.
Ramakrishnan, U., Coss, R.G., 2000. Age differences in the re-sponses to adult and juvenile alarm calls by bonnet macaques(Macaca radiata). Ethology 105, 131–144.
Ramakrishnan, U., Coss, R.G., Pelkey, N.W., 1999. Tiger declinecaused by the reduction of large ungulate prey: supporting evi-dence from a comparative study of leopard diets in two reservesin southern India. Biol. Conserv. 89, 113–120.
Rice, C.G., 1986. Observations on predators and prey at Eraviku-lam National Park, Kerala. J. Bombay Nat. Hist. Soc. 83, 283–305.
Rogal, L., Reible, G., Fischer, B., 1985. Reaction times of the eyeand the hand of the monkey in a visual reach task. Neurosci. Lett.58, 127–132.
Rumelhart, D.E., Hinton, G., Williams, R., 1986. Learning inter-nal representations by error propagation. In: Rumelhart, D.E.,McClelland, J.L. (Eds.), Parallel Distributed Processing: Explo-
l. 1.
S d ob-–111.
S Inc.,
S D.,de-.
S ba-
S zoo-ual
S nkeyperi-hav.
S driticailed
texture patterns in area V1 of the alert macaque monkey. J.rophysiol. 67, 961–980.
ortlandt, A., 1967. Experimentation with chimpanzees in the wIn: Starck, D., Schneider, R., Kuhn, H.-J. (Eds.), Neue Ergebder Primatologie – Progress in Primatology. Gustav Fischerlag, Stuttgart, pp. 208–224.
ee, E.T., 1980. Statistical Methods for Survival Data Analysis. Ltime Learning Publications, Belmont, CA.
evitt, J.B., Lewis, D.A., Yoshioka, T., Lund, J.S., 1993. Topogphy of pyramidal neuron intrinsic connections in macaque mkey prefrontal cortex (areas 9 and 46). J. Comp. Neurol.360–376.
und, J.S., Yoshioka, T., Levitt, J.B., 1993. Comparison of insic connectivity in different areas of macaque monkey cerecortex. Cerebral Cortex 3, 148–162.
und, J.S., Wu, Q., Levitt, J.B., 1995. Visual cortex cell typesconnections. In: Arbib, M.A. (Ed.), The Handbook of Brain Tory and Neural Networks. The MIT Press, Cambridge, MA,1016–1021.
antel, N., 1967. Ranking procedures for arbitrarily restrictedservations. Biometrics 23, 65–78.
cRae, M., 1997. Road kill in Cameroon. Nat. Hist. 106, 36–7
rations in the Microstructure of Cognition: Foundations, voThe MIT Press, Cambridge, MA, pp. 318–362.
ekular, A.B., Palmer, S.E., 1992. Perception of partly occludejects: a microgenetic analysis. J. Exp. Psych: Gen. 121, 95
leeper, B., 1995. Wild Cats of the World. Crown Publishers,New York, 216 pp.
mith, A.C., Buchanan-Smith, H.M., Surridge, A.K., Osorio,Mundy, N.I., 2003. The effect of colour vision status on thetection and selection of fruits by tamarins (Saguinusspp.). J. ExpBiol. 206, 3159–3165.
tacey, P.B., 1986. Group size and foraging efficiency in yellowboons. Behav. Ecol. Sociobiol. 18, 175–187.
tanley, M.E., Aspey, W.P., 1984. An ethometric analysis in alogical garden: Modification of ungulate behavior by the vispresence of a predator. Zoo Biol. 3, 89–109.
tell, M., Riesen, A., 1987. Effects of early environments on mocortex neuroanatomical changes following somatomotor exence: effects on layer III pyramidal cells in monkey cortex. BeNeurosci. 101, 341–346.
truble, R.G., Riesen, A.H., 1978. Changes in cortical denbranching subsequent to partial social isolation in stumptmonkeys. Dev. Psychobiol. 11, 479–486.

R.G. Coss et al. / Behavioural Processes 68 (2005) 145–163 163
Tanaka, K., 1996. Representation of visual features of objects in theinferotemporal cortex. Neur. Net. 9, 1459–1475.
Terry, R.L., 1994. Effects of facial transformations on accuracy ofrecognition. J. Soc. Psychol. 134, 483–492.
Thapar, V., 1999. The Secret Life of Tigers. Oxford University Press,Oxford, 99 pp.
Topal, J., Csanyi, V., 1994. The effect of eye-like schema on shut-tling activity of wild house mice (Mus musculus domesticus):context-dependent threatening aspects of the eyespot patterns.Anim. Learn. Behav. 22, 96–102.
Ts’o, D.Y., Gilbert, C.D., Wiesel, T.N., 1986. Relationships betweenhorizontal interactions and functional architecture in cat striatecortex as revealed by cross-correlation analysis. J. Neurosci. 6,1160–1170.
Yin, C., Kellman, P.J., Shipley, T.F., 1997. Surface completion com-plements boundary interpolation in the visual integration of partlyoccluded objects. Perception 26, 1459–1479.
Yoshioka, T., Dow, B.M., 1996. Color, orientation and cytochromeoxidase reactivity in areas V1, V2 and V4 of macaque monkeyvisual cortex. Behav. Brain Res. 76, 71–88.
Yoshioka, T., Vautin, R.G., 1996. Neuronal mechanisms of colorcategorization in areas V1, V2 and V4 of macaque monkey visualcortex. Behav. Brain Res. 76, 51–70.
Zuberbuhler, K., 2000. Causal knowledge of predators’ behaviour inwild Diana monkeys. Anim. Behav. 59, 209–220.
Zuberbuhler, K., Jenny, D., Bshary, R., 1999. The predator de-terrence function of primate alarm calls. Ethology 105, 477–490.










![Snow leopards[1]](https://img.pdfslide.net/doc/110x75/548e7950b47959b3488b479a/snow-leopards1.jpg)