Embed Size (px)
Citation preview

Reduced production of sulfated glycosaminoglycans occurs inZambian children with kwashiorkor but not marasmus1–4
Beatrice Amadi, Andrew O Fagbemi, Paul Kelly, Mwiya Mwiya, Franco Torrente, Camilla Salvestrini, Richard Day,Michael H Golden, Erik A Eklund, Hudson H Freeze, and Simon H Murch
ABSTRACTBackground: Kwashiorkor, a form of severe malnutrition with highmortality, is characterized by edema and systemic abnormalities.Although extremely common, its pathophysiology remains poorlyunderstood, and its characteristic physical signs are unexplained.Objective: Because kwashiorkor can develop in protein-losing en-teropathy, which is caused by a loss of enterocyte heparan sulfateproteoglycan (HSPG), and previous observations suggest abnormalsulfated glycosaminoglycan (GAG) metabolism, we examinedwhether intestinal GAG and HSPG are abnormal in children withkwashiorkor.Design: Duodenal biopsy samples collected from Zambian childrenwith marasmus (n ¼ 18), marasmic kwashiorkor (n ¼ 8), andkwashiorkor (n ¼ 15) were examined for expression of HSPG,GAGs, and immunologic markers and compared against referencesamples from healthy UK control children. GAG and HSPG expres-sion density and inflammatory cell populations were quantitated bycomputerized analysis.Results: The kwashiorkor group was less wasted and had a lowerHIV incidence than did the other groups. All duodenal biopsy sam-ples showed inflammation compared with the histologically unin-flamed control samples. Biopsy samples from marasmic childrenhad greater inflammation and greater CD31 and HLA-DR (humanleukocyte antigen DR)–positive cell densities than did samples fromchildren with kwashiorkor. Expression of both HSPG and GAGs wassimilar between marasmic and well-nourished UK children but wasmarkedly lower in children with kwashiorkor in both the epitheliumand lamina propria. Although underglycosylated and undersulfated,epithelial syndecan-1 protein was normally expressed in kwashiorkor,which confirmed that abnormalities arise after core protein synthesis.Conclusions: Intestinal HSPG loss occurs in kwashiorkor, whichmay precipitate protein-losing enteropathy to cause edema. If oc-curring systemically, impaired HSPG expression could cause sev-eral previously unexplained features of kwashiorkor. We speculatethat a genetic predisposition to reduced HSPG biosynthesis mayoffer a contrasting selective advantage, by both diminishing proteincatabolism during transient undernutrition and protecting againstspecific infectious diseases. Am J Clin Nutr 2009;89:592–600.
INTRODUCTION
Edema that occurs because of malnutrition has been recog-nized since biblical times (1). The name kwashiorkor is derivedfrom a Ghanaian term for a characteristic disease arising in young
children and is clinically distinct from marasmus (1). In additionto edema, features distinguishing kwashiorkor from marasmusinclude hypoalbuminemia, skin exfoliation, fatty liver, suscep-tibility to infection, heart failure, thrombosis, profound apathy,and suppressed appetite (1). In marasmic kwashiorkor, edemacoexists with severe wasting. In Zambia, children with marasmickwashiorkor have mild edema and are clinically closer to ma-rasmus than kwashiorkor (2). Kwashiorkor is both extremelycommon and the most lethal form of malnutrition—fatality ratesfor established kwashiorkor remain at ’25% or more in manyfacilities (3, 4).
Early theories that disease is caused by a low-protein, carbo-hydrate-based diet (1) have been rendered unlikely by findings ofsimilar protein intakes in children developing kwashiorkor andmarasmus (5, 6). Resolution of all signs, including edema, occurswith low-protein treatment regimens, and low-protein diets do notreproduce kwashiorkor in animals (1). Speculation that kwashior-kor might be caused by aflatoxin poisoning (7) has not been sup-ported by analyses of postmortem tissues (1). Although enhancedfree radical production occurs, causing lipid peroxidation (1),a large field trial of an antioxidant cocktail did not preventkwashiorkor (8). Golden et al (1, 9) speculated that sulfated gly-cosaminoglycans (GAGs) might be disrupted in kwashiorkor on the
1 From the Department of Paediatrics and Child Health, University Teach-
ing Hospital of Lusaka, Lusaka, Zambia (BA and MM); the Centre for
Paediatric Gastroenterology, Royal Free & University College School of
Medicine, London, United Kingdom (AOF, FT, and CS); the Tropical Gas-
troenterology and Nutrition Group, University of Zambia School of Medi-
cine, Lusaka, Zambia (PK); the Queen Mary, University of London, London,
United Kingdom (PK); the Centre for Gastroenterology and Nutrition, Uni-
versity College London, London, United Kingdom (RD); the University of
Aberdeen, Aberdeen, United Kingdom (MHG); the Burnham Institute for
Research, La Jolla, CA (EAE and HHF); and the Clinical Sciences Research
Institute, Warwick Medical School, University of Warwick, Coventry, United
Kingdom (SHM).2 BA and AOF are joint first authors.3 PK was supported by The Wellcome Trust, and HHF and SHM were
supported by the Children’s Heart Fund for research into protein-losing
enteropathy.4 Address correspondence and reprint requests to SH Murch, Warwick
Medical School, Clinical Sciences Research Institute, Clifford Bridge Road,
Coventry CV2 2DX, United Kingdom. E-mail: [email protected].
Received October 13, 2008. Accepted for publication November 22, 2008.
First published online December 30, 2008; doi: 10.3945/ajcn.2008.27092.
592 Am J Clin Nutr 2009;89:592–600. Printed in USA. � 2009 American Society for Nutrition
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018

basis of findings of diffusion of interstitial dye, renal podocyteeffacement, resistance of these children to cholera (1), and reportsof low urinary excretion of sulfate and sulfated GAGs (10, 11). Thegeneralized nature of this defect in complex carbohydrate metab-olism is suggested by findings of striking reductions in brain sul-fated GAGs in kwashiorkor, with relative sparing of undersulfatedchondroitin sulfate over heparan sulfate proteoglycan (HSPG) (12).These findings agreewith magnetic resonance imaging data showingmarked brain volume shrinkage that recovers with nutritional re-habilitation (13).
We identified a specific role for HSPG in protein-losing en-teropathy (PLE). Infants with congenitally absent enterocyteHSPG suffered fatal PLE (14), whereas mislocalization of en-terocyte HSPG in congenital disorder of glycosylation (CDG)caused episodes of life-threatening PLE (15). In vitro studiesconfirmed a critical role for HSPG in preventing PLE and iden-tified synergy between HSPG loss, cytokines (tumor necrosisfactor-a and interferon-c), and hydrostatic pressure in promotingintestinal protein leak (16, 17). We confirmed these findings invivo in transgenic mice, in which cytokines or pressure-inducedintestinal albumin leak was markedly increased when gut epi-thelial expression of proteoglycan syndecan-1 or HSPG is in-hibited or HSPG glycan chain synthesis is reduced (18, 19). Inview of the similarities between infants with kwashiorkor andinfants with abnormal intestinal HSPG expression, we evaluatedthe hypothesis that there may be abnormal intestinal expressionof HSPG in kwashiorkor.
SUBJECTS AND METHODS
All patients were treated, and the investigations were per-formed on the malnutrition ward of the University TeachingHospital, Lusaka, Zambia. We examined duodenal biopsysamples that had been taken previously for a study of enteralnutrition therapy in malnourished children, which was performedduring 1998–2000 (2, 20). The material came from 3 groups ofchildren: marasmus (n ¼ 18), marasmic kwashiorkor (n ¼ 8),and classic kwashiorkor (n ¼ 15). Their clinical details areshown in Table 1. Follow-up duodenal biopsy samples werestudied from 16 of these patients who successfully completednutritional rehabilitation (5 marasmus, 5 marasmic kwashiorkor,and 6 kwashiorkor).
Children were recruited if they had malnutrition and persistentdiarrhea lasting �14 d (2, 20). Children were excluded if they
had features of measles, chicken pox, or neurologic disorder orif they were exclusively breastfed. The study protocol was ap-proved by the Research Ethics Committee of the University ofLusaka, and formal consent obtained from parents or guardiansfor each procedure. A structured medical history was taken anda complete physical examination was performed, including an-thropometric measurements. Malnutrition was classified as un-derweight, marasmus, kwashiorkor, and marasmic kwashiorkor aspreviously reported (21), and analyses of infection, malnutrition,mortality, and response to nutrition have been published (2, 20).
Duodenal biopsy samples were collected by BA via endoscopyunder sedation with midazolam. No adverse events were iden-tified during or after these procedures, although 2 follow-upendoscopies were cancelled because of paradoxical agitationduring sedation.
Biopsy analysis
Avidin-biotin immunohistochemistry (Vectastain Elite; Vec-tor, Peterborough, United Kingdom) was performed after ap-propriate antigen retrieval and blocking of endogenousperoxidase. Antibodies used included anti-CD3 (1:50), humanleukocyte antigen DR-1 (HLA-DR; 1:50), Ki67 (1:50; allDako, Cambridge, United Kingdom), syndecan-1 (1:50; Sero-tec, Oxford, United Kingdom), and HSPG (10E4, 1:50; Seiga-kaku, AMS Biotech, Abingdon, United Kingdom), as previouslyreported (15, 22). The distribution of sulfated GAGs was studiedwith a poly-L-lysine probe at pH 1.2, as previously reported (14,15). Both the HSPG monoclonal and the poly-L-lysine probeshave been extensively characterized. Monoclonal 10E4 detectsN-acetylated and N-sulfated disaccharides within HSPG glycanchains, and its staining is abrogated by preincubation withheparitinase before application of the monoclonal probe (23).The poly-L-lysine probe is specific for sulfate residues in GAGchains of heparan, dermatan, and chondroitin sulfates (14, 15).Previous studies have established that the probe for sulfatedGAGs recognizes predominantly HSPG within the epitheliumand a combination of heparan sulfate, dermatan sulfate, andchondroitin sulfate within the lamina propria (14). Both methodsdemonstrate similar epithelial staining in normal small intestine,which is predominantly basolateral, although intracellularstaining may be variably seen toward the villus tip (14). Withinthe lamina propria, the poly-L-lysine probe for sulfatedGAGs may demonstrate more extensive interstitial staining
TABLE 1
Clinical characteristics of the patients at the time of admission1
Marasmus (n ¼ 18) Marasmic kwashiorkor (n ¼ 8) Kwashiorkor (n ¼ 15)
Age (mo) 16 (12.2, 17.4)2 17 (13.6, 19.1) 19 (16, 19.8)
Sex [M/F (n)] 11/7 4/4 11/4
Weight-adjusted z score 24.8 (23.2, 25.4) 24.73 (24.5, 25.3) 23.6 (23.1, 23.9)
Weight for height z score 23.53 (22.9, 25.2) 24.53 (23.4, 25.1) 22.5 (22.1, 23.2)
Height-adjusted z score 22.7 (21.3, 24) 23.1 (21.6, 23.8) 23 (22, – 3.5)
HIV positive (n) 14 6 7
Acid fast bacilli in gastric aspirate (n) 1 2 4
Fever at time of admission (n) 5 2 3
1 Comparisons between groups were made by using an unpaired t test if probable significant differences were identified
by ANOVA and a multiple range test.2 Median; 95% CI in parentheses (all such values).3 Significantly different from kwashiorkor, P , 0.05.
GLYCOSAMINOGLYCANS IN KWASHIORKOR 593
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018

than the HSPG monoclonal probe, although many cell types areHSPG immunoreactive, and extracellular HSPG immunoreac-tivity may also occur because of secretion by fibroblasts orcleavage of cell surface proteoglycans (15, 23).
Digital photomicrographs were prepared from all slides withuse of the same microscope (Leica DMLB) and image capturesystem (Nikon Digital Sight DL-S1). All quantitations were madeon unprocessed images with the use of ImageJ software (NationalInstitutes of Health, Bethesda, MD: http://rsb.info.nih.gov/ij/).For quantitation of staining densities, plug-ins for color decon-volution (AC Ruifrok, Netherlands) and for region-of-interestdensitometry (J Kuhn, A Padua, USA) were used to separatespecific di-amino-benzidine (DAB) or silver staining from hae-malum counterstain and to quantitate local density of DAB orsilver stain. Separate quantitation was done in the laminapropria, excluding crypts, and intraepithelial compartments. Forthe lamina propria, low-power images were used, and a singlefield was drawn around the entire compartment excluding crypts.If biopsy morphology prevented measurement of a single com-partment, a mean was obtained from separate compartments ofroughly equal areas. For the epithelium, high-power imageswere used, and a mean density was derived for each specimenfrom quantitation of multiple individual intercellular spaces inwell-orientated villi.
Densitometry was performed on an inverse scale (1–250units), based on light transmission. The staining density indexpresented is reciprocal [1/transmission intensity 3 104 2 40(background subtraction)], but all statistical analyses were per-formed on unprocessed data. With this derived index, values�150 represent intense staining, whereas values �60 representfaint staining. Reviewing previously reported specimens (15), anepithelial HSPG expression density of 60 was associated withacute severe PLE, whereas an expression density of 100 wassufficient to prevent intestinal protein leak. Cell densities werequantified by using the Nucleus Counter Plug-in (ImageJ soft-ware). Previously stained biopsy samples from UK children,reported in other studies (14, 15, 22), that were found not tohave significant gastrointestinal disease were similarly quanti-tated for expression of sulfated GAGs and HSPG.
Serum analysis
HIV serology was performed in all patients as previouslyreported (2). Isoelectric focusing for analysis of transferringlycosylation was performed in 9 HIV-negative patients (15).
Statistical analysis
Data were assessed for normal distribution by analysis ofstandardized skewness and standardized kurtosis by using Sta-graphic 5 Plus software (Manugistics Inc, Rockville, MD). Datawere compared initially by analysis of variance (ANOVA) orKruskal-Wallis ANOVA and multiple range testing. When po-tentially significant differences were identified, comparison wasmade between individual groups by using an unpaired t test(clinical groups) or a Mann-Whitney U test (all histochemicaland immunohistochemical measurements), with appropriate ad-justment for multiple comparisons. P values � 0.05 were con-sidered statistically significant.
RESULTS
There were some differences between the groups at pre-sentation (Table 1). Other than a lower incidence of tuberculosis(acid fast bacilli detected in gastric aspirate) in marasmus and ofHIV in kwashiorkor, infectious complications were similar be-tween groups. This lower prevalence of HIV in kwashiorkor wasrepresentative of the overall study population of 200 childrenpreviously reported (2).
No significant differences in biopsy samples by conventionalhistology were observed between the groups (Table 2). However,mucosal architecture was markedly abnormal compared withthat in healthy UK children, who had a median villus:crypt ratioof ’2:1 (22). Villous height was lower and crypt depth increasedto a similar extent as in UK specimens in all malnutrition groups,with minimal change after nutritional rehabilitation (Table 2).
Mucosal inflammation in marasmus and kwashiorkor
Mucosal inflammatory responses were different between theclinical groups (Figure 1, Table 3), although no significantdifference in overall lamina propria cell density was observedbetween the groups (Table 2). All groups had a significantlyhigher CD31 intraepithelial lymphocyte (IEL) density than didUK control children, as previously noted in Gambian children(22). This was significantly higher in the children with marasmickwashiorkor than in all other groups, with IEL density reachinglevels seen in celiac disease (Table 3, Figure 1). Within thelamina propria, all groups had a 4–5-fold higher total CD31 Tcell density (Table 3) than did UK children, with an overall celldensity similar to that in malnourished Gambian children (22).The children with kwashiorkor had significantly lower laminapropria T cell counts than did those with marasmus (Table 3).
TABLE 2
Mucosal characteristics by hematoxylin and eosin staining of biopsy samples, by group1
Marasmus
(n ¼ 18)
Marasmic
kwashiorkor (n ¼ 8)
Kwashiorkor
(n ¼ 15)
Nutritional
rehabilitation (n ¼ 15)
UK controls
(n ¼ 19)
Villous height (lm) 241.9 6 10.82 243.2 6 25.8 282.4 6 27.5 250.8 6 12.9 355 6 35
Crypt depth (lm) 263.4 6 25.4 263.0 6 23.4 274.4 6 18.2 245.8 6 15.9 170 6 20
Villus:crypt ratio 1.12 6 0.15 1.14 6 0.17 1.00 6 0.16 1.01 6 0.11 2.1 6 0.3
Lamina propria cell density 7290 (5780, 8801)3 7584 (6696, 8472) 6842 (5175, 8509) 8202 (6930, 9474) —
1 No significant differences were found between the study groups in villous height, crypt depth, villus:crypt ratio, or lamina propria cell density by
ANOVA and multiple range testing; thus, further Mann-Whitney testing was not performed. Normal data for comparison of villus and crypt architecture with
UK control children were obtained from previous studies (22). The nutritional rehabilitation group includes some patients from each group in the first
3 columns, for whom biopsy had been repeated after a period of nutritional therapy.2 Mean 6 SE (all such values).3 Mean; 95% CI in parentheses (all such values).
594 AMADI ET AL
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018
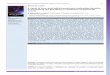
There was a strongly up-regulated expression of the interferon-c–induced class II major histocompatability complex moleculeHLA-DR on surface epithelium in all Zambian cases. This in-dicated a Th1-dominated mucosal response in these childreninitially enrolled into the nutritional intervention study (2, 20),which has also been observed in both malnourished and well-nourished Gambian children (13), although in UK children sucha finding is considered abnormal. On formal quantitation ofstaining intensity, no overall difference in HLA-DR expression
in the epithelium was detected between clinical groups (Table 3).In contrast, there was a significantly lower density of HLA-DR1
cells in the lamina propria in the children with kwashiorkor thanin the other groups (Table 3).
We found a strikingly higher expression of the crypt epithelialnuclear proliferation marker Ki67 in all groups compared withthe UK control children (21) (Table 3, Figure 1). Although therewere no differences identified between the malnourished groups,there was a reduction in crypt epithelial Ki671 cell density in
FIGURE 1. Mucosal inflammatory responses. Distribution of CD31 T cells, cells expressing human leukocyte antigen DR (HLA-DR), and the proliferativeantigen Ki67 in Zambian children. Immunoreactive cells are stained brown. The top row shows slides from children with marasmus (Maras), the second fromchildren with marasmic kwashiorkor (M-K), the third from children with kwashiorkor (Kwas), and the bottom from children participating in a nutritionalrehabilitation program (Nutr rehab). All were taken from specimens near to overall mean values (data shown in Table 2).
TABLE 3
Results of immunohistochemical analysis of mucosal biopsy samples, by group1
Marasmus Marasmic kwashiorkor Kwashiorkor Nutritional rehabilitation
CD31 IEL (per 100 enterocytes) 302 (26.4, 37.7) 45.5 (37.4, 58.3) 313 (26, 37.6) 303 (22.5, 36.9)
Epithelial HLA-DR staining intensity 118.7 (90.7, 202.6) 118.7 (80.5, 190.9) 120 (70.4, 194.2) 150.5 (113.2, 190.9)
Ki 671 cells within crypts (%) 76.44 (69.5, 89.5) 764 (68.2, 84.4) 75 (57.6, 85.8) 68.3 (66.6, 71.4)
Lamina propria CD31 cells (per mm2) 19535 (1708, 2054) 1842 (1358, 2457) 1318 (1187, 1862) 1893 (1527, 2060)
Lamina propria HLA-DR1 cells (per mm2) 25226 (2005, 2862) 23475 (1938, 3181) 1907 (1462, 2208) 25276 (1972, 2819)
Lamina propria area HLA-DR1 (%) 0.76 (0.41, 0.85) 0.61 (0.36, 0.95) 0.35 (0.18, 0.57) 0.52 (0.28, 0.89)
Lamina propria Ki671 cell density (per mm2) 716 (311, 991) 845 (386, 1253) 488 (394, 632) 810 (504, 1119)
1 All values are medians (95% CIs). Reference data for healthy UK control children were obtained from previous studies (22): median ,20 for CD31
intraepithelial lymphocytes (IEL; per 100 enterocytes), median ,30 for Ki 671 cells within crypts (%), and 290–390 for lamina propria CD31 cells (per
mm2). HLA-DR, human leukocyte antigen DR. Differences that were potentially significant by ANOVA or Kruskal-Wallis test were examined further by
a Mann-Whitney U test.2,3 Significantly different from marasmic kwashiorkor: 2P , 0.05, 3P , 0.01.4 Significantly different from nutritional rehabilitation, P , 0.01.5,6 Significantly different from kwashiorkor: 5P , 0.05, 6P , 0.01.
GLYCOSAMINOGLYCANS IN KWASHIORKOR 595
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018
children after rehabilitation. Large numbers of cells within thelamina propria expressed Ki67 in all groups; the density was notsignificantly lower in kwashiorkor. Thus, the mucosal inflam-matory responses were lower overall in the children with kwash-iorkor than in the children with marasmus, although both groupshad similar degrees of persistent diarrhea.
Reduced mucosal HSPG expression in classic kwashiorkor
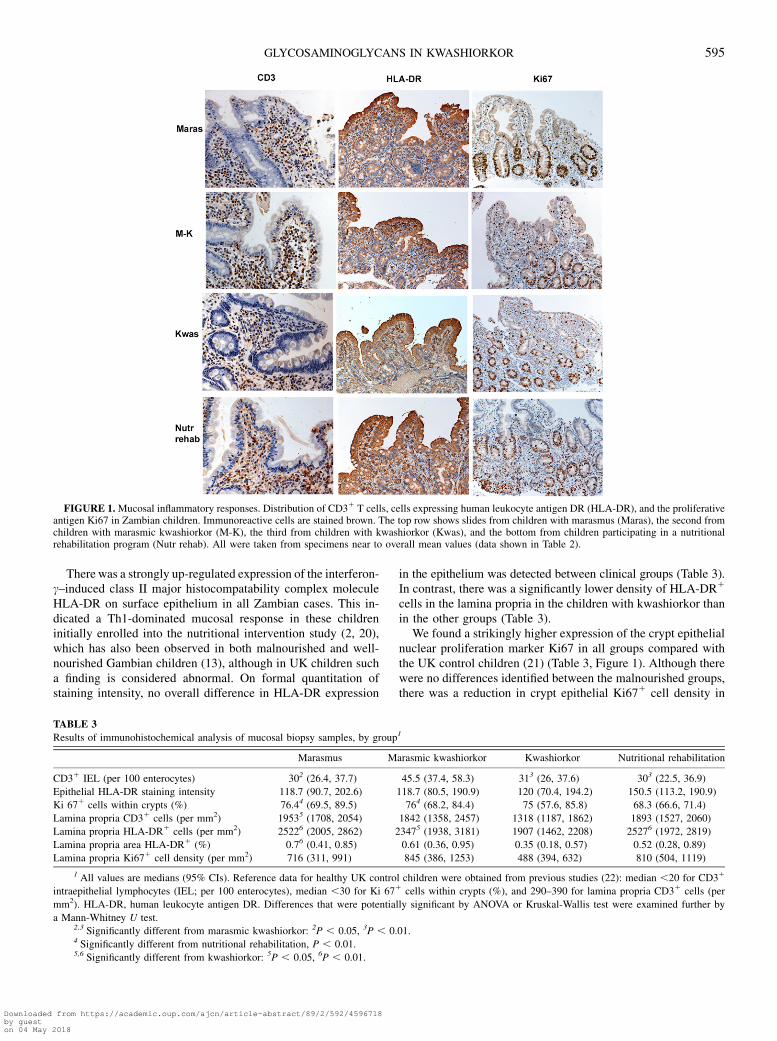
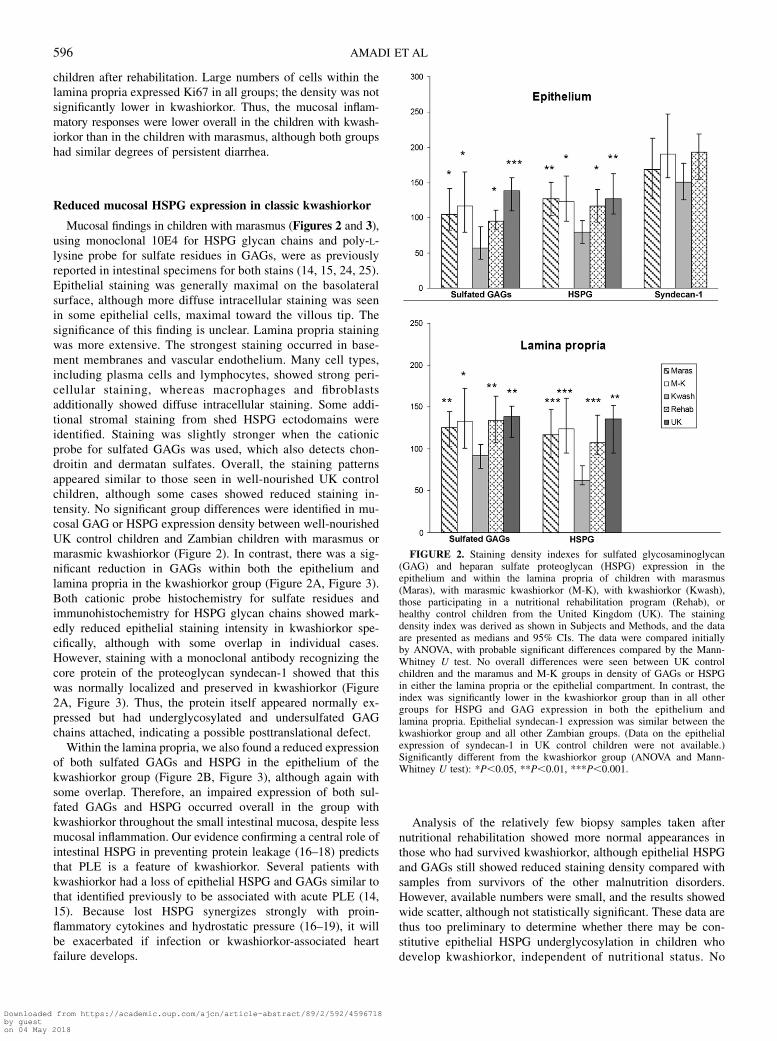
Mucosal findings in children with marasmus (Figures 2 and 3),using monoclonal 10E4 for HSPG glycan chains and poly-L-lysine probe for sulfate residues in GAGs, were as previouslyreported in intestinal specimens for both stains (14, 15, 24, 25).Epithelial staining was generally maximal on the basolateralsurface, although more diffuse intracellular staining was seenin some epithelial cells, maximal toward the villous tip. Thesignificance of this finding is unclear. Lamina propria stainingwas more extensive. The strongest staining occurred in base-ment membranes and vascular endothelium. Many cell types,including plasma cells and lymphocytes, showed strong peri-cellular staining, whereas macrophages and fibroblastsadditionally showed diffuse intracellular staining. Some addi-tional stromal staining from shed HSPG ectodomains wereidentified. Staining was slightly stronger when the cationicprobe for sulfated GAGs was used, which also detects chon-droitin and dermatan sulfates. Overall, the staining patternsappeared similar to those seen in well-nourished UK controlchildren, although some cases showed reduced staining in-tensity. No significant group differences were identified in mu-cosal GAG or HSPG expression density between well-nourishedUK control children and Zambian children with marasmus ormarasmic kwashiorkor (Figure 2). In contrast, there was a sig-nificant reduction in GAGs within both the epithelium andlamina propria in the kwashiorkor group (Figure 2A, Figure 3).Both cationic probe histochemistry for sulfate residues andimmunohistochemistry for HSPG glycan chains showed mark-edly reduced epithelial staining intensity in kwashiorkor spe-cifically, although with some overlap in individual cases.However, staining with a monoclonal antibody recognizing thecore protein of the proteoglycan syndecan-1 showed that thiswas normally localized and preserved in kwashiorkor (Figure2A, Figure 3). Thus, the protein itself appeared normally ex-pressed but had underglycosylated and undersulfated GAGchains attached, indicating a possible posttranslational defect.
Within the lamina propria, we also found a reduced expressionof both sulfated GAGs and HSPG in the epithelium of thekwashiorkor group (Figure 2B, Figure 3), although again withsome overlap. Therefore, an impaired expression of both sul-fated GAGs and HSPG occurred overall in the group withkwashiorkor throughout the small intestinal mucosa, despite lessmucosal inflammation. Our evidence confirming a central role ofintestinal HSPG in preventing protein leakage (16–18) predictsthat PLE is a feature of kwashiorkor. Several patients withkwashiorkor had a loss of epithelial HSPG and GAGs similar tothat identified previously to be associated with acute PLE (14,15). Because lost HSPG synergizes strongly with proin-flammatory cytokines and hydrostatic pressure (16–19), it willbe exacerbated if infection or kwashiorkor-associated heartfailure develops.
Analysis of the relatively few biopsy samples taken afternutritional rehabilitation showed more normal appearances inthose who had survived kwashiorkor, although epithelial HSPGand GAGs still showed reduced staining density compared withsamples from survivors of the other malnutrition disorders.However, available numbers were small, and the results showedwide scatter, although not statistically significant. These data arethus too preliminary to determine whether there may be con-stitutive epithelial HSPG underglycosylation in children whodevelop kwashiorkor, independent of nutritional status. No
FIGURE 2. Staining density indexes for sulfated glycosaminoglycan(GAG) and heparan sulfate proteoglycan (HSPG) expression in theepithelium and within the lamina propria of children with marasmus(Maras), with marasmic kwashiorkor (M-K), with kwashiorkor (Kwash),those participating in a nutritional rehabilitation program (Rehab), orhealthy control children from the United Kingdom (UK). The stainingdensity index was derived as shown in Subjects and Methods, and the dataare presented as medians and 95% CIs. The data were compared initiallyby ANOVA, with probable significant differences compared by the Mann-Whitney U test. No overall differences were seen between UK controlchildren and the maramus and M-K groups in density of GAGs or HSPGin either the lamina propria or the epithelial compartment. In contrast, theindex was significantly lower in the kwashiorkor group than in all othergroups for HSPG and GAG expression in both the epithelium andlamina propria. Epithelial syndecan-1 expression was similar between thekwashiorkor group and all other Zambian groups. (Data on the epithelialexpression of syndecan-1 in UK control children were not available.)Significantly different from the kwashiorkor group (ANOVA and Mann-Whitney U test): *P,0.05, **P,0.01, ***P,0.001.
596 AMADI ET AL
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018

differences between groups were apparent in recovery in thelamina propria, where cellular turnover is slower.
The normal location of epithelial syndecan-1 in kwashiorkorwas unlike findings for CDG-I during PLE (15). Additionalserologic analysis of transferrin glycosylation in 9 HIV negativepatients with kwashiorkor showed no evidence of defectiven-glycosylation (data not shown). Thus, we found no evidenceof a CDG-I–like defect in Zambian children with kwashiorkor.
DISCUSSION
Our findings support data from other organs, which indicatesa generalized loss of sulfated GAGs in kwashiorkor (1, 9–12). Weacknowledge limitations imposed by studies of biopsy samples,too small for in-depth glycobiological analysis with currenttechnology. Analysis of staining intensity, even with computer-ized densitometry, is a relatively crude approach to a complexbiosynthetic process. Potential sources of artifact may ariseduring tissue handling or staining. However, using 2 well-characterized stains specific for glycan carbohydrates (15) andsulfates (14, 15), we identified a similar pattern of reducedsulfated GAG expression throughout the small intestinal mu-cosa. Children with marasmus and persistent diarrhea generallyshowed HSPG expression similar to that of normal Europeanchildren. Thus, it is unlikely that nutritional deficiencies causingwasting and stunting per se induce this loss. Because sulfatedGAGs are now identified as critical in many aspects of physi-ology (24), their disruption may play a role in the signs andpathophysiology of kwashiorkor.
HSPG loss promotes albumin leakage and reduced tissueturgor (9, 14–18, 24, 25). Although urinary protein loss is rare inkwashiorkor (9), PLE has been reported and may precipitatedisease (26, 27). Although we found normal HSPG in marasmickwashiorkor, IEL density was high, which indicated a likelyrecent infection. We previously showed that reglycosylationcapacity limits PLE after HSPG loss (16–18) and suggests thatthese previously marasmic children may have suffered an acuteepisode of PLE on infection but were then able to reglycosylateto restore the epithelial barrier.
HSPG deficiency is consistent with known associations ofkwashiorkor. Its frequent onset after displacement from the breastmay relate to both infections and substrate deficiency, becausebreast milk is rich in N-acetyl glucosamine (GlcNAc) and sulfur-containing amino acids (28). It is intriguing that consumption ofinsects, high in chitin GlcNAc, appears to protect infants againstkwashiorkor, which suggests the ironic possibility that Westernmissionaries may have increased the incidence of kwashiorkorin the early 20th century because of their campaign againstentomophagy (29). Aflatoxin liver damage is associated withendothelial HSPG loss, and heparin is strongly protective (30).The known state of oxidative stress (1) may also relate to HSPGdeficiency, because the constitutive antioxidant function of su-peroxide dismutase is dependent on HSPG binding (31) andcomplex carbohydrates are a target for oxidative damage.
Even under conditions of extreme privation, it is unusual formore than ’10% of children to develop kwashiorkor. Why thendo 90% of children living in the same household and eating thesame diet fail to develop kwashiorkor? A potential geneticpredisposition was postulated by Gopalan (5). We note in thiscontext that mice with congenitally reduced HSPG biosynthesis
appear normal but show exaggerated enteric protein leakage oncytokine challenge (11, 32) or under conditions of under-nutrition (L Bode, H Freeze, unpublished observations, 2008).Undernutrition induces competition between metabolic path-ways for limited nutritional substrates. Low glycosylators maythen better maintain body mass, because reduced consumptionof substrates in glycosylation reduces the need for protein ca-tabolism. There is indeed evidence that protein breakdown isreduced (33) and body mass is preserved (1, 2) in children withkwashiorkor. However, an infectious enteric challenge thatcauses inflammatory HSPG breakdown (25), mandating rapidepithelial reglycosylation for repair (15–18), may cause HSPGdeficiency sufficient to induce PLE. Chronic enteric proteinlosses would then exacerbate nutritional inadequacy, causinga vicious circle.
Because of the multiple roles of HSPG (24), deficient productionduring starvation may explain several features of kwashiorkor inaddition to oxidative stress (31). Fatty liver, caused by low he-patic lipoprotein lipase expression and high serum free fattyacids (34), is explicable on the basis that hepatic lipoproteinlipase localization and fatty acid uptake are HSPG dependent(35, 36). Hepatic knockdown of GlcNAc N-deacetylase/N-sulfotransferase 1 (Ndst1) in mice markedly inhibits lipoproteinclearance (36). Exfoliative dermatitis may develop becauseHSPG glycosylation of CD44 is important for keratinocyteadhesion (37). Vascular thrombosis is a known consequenceof lost HSPG-dependent antithrombotic mechanisms (24, 27),whereas myocardial contractility is decreased by protamine,which blocks GAGs (38) and is increased by heparin (39). Reducedglycosylation of syndecans (as for syndecan-1) may inducehypophagia, because hypothalamic syndecan-3 GAG chainsmediate the normal hunger response (40) and may contribute toincreased infectious mortality, as observed in lipopolysaccharide–challenged, syndecan-4–deficient mice (41). We thus suggestthat inadequate HSPG synthesis under malnourished con-ditions may underpin several major features of kwashiorkor.Such loss of GAGs may also have therapeutic implications,particularly because blood-brain barrier integrity is HSPGdependent (24). In view of the major loss of brain HSPG inkwashiorkor (12), drugs that are highly toxic if they cross theblood-brain barrier, notably ivermectin (used widely in Africa totreat onchocerciasis), may provide particular risks for those withnutritional edema.
Polymorphisms reducing HSPG glycosylation or sulfationmay have benefits other than limiting protein catabolism. Afurther predisposition to evolutionary advantage would be pro-tection against infective diseases, because many pathogens bindspecifically to domains within sulfated GAGs. Plasmodium fal-ciparum erythrocyte membrane protein-1 induces intravascularsequestration in infected erythrocytes by binding to endothelialHSPG (42). HSPG binding avidity determines pathogenicity(43), and exogenous GAGs reverse microvascular complica-tions (42). It is thus notable that kwashiorkor protects againstsevere P. falciparum malaria and that cerebral malaria is almostnever seen in this disorder. Vibrio cholerae survives in water byspecifically binding to GlcNAc residues in crustacean chitinsand then infects host cells by binding to glycocalyx GlcNAc(44). Patients with kwashiorkor showed a remarkable 25-foldreduction in cholera infection compared with those with ma-rasmus during an outbreak in a therapeutic feeding center (1).
GLYCOSAMINOGLYCANS IN KWASHIORKOR 597
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018
HIV entry across endothelial barriers is mediated by interactionbetween gp120 and HSPG (45), and transplacental infection isthus likely to be HSPG dependent. The finding of 50% less HIVseropositivity in kwashiorkor in the overall cohort of 200children from this unit (2) is in line with previous findings of 3-fold lower HIV seropositivity in Malawian children (46), 4-foldlower HIV seropositivity in Indian children with kwashiorkor(47), and 2–4-fold lower HIV seropositivity in numerous otherAfrican countries. Thus, vertical transmission of HIV appears tobe reduced in infants who later develop kwashiorkor. Wespeculate that genetic polymorphism or polymorphisms in GAGexpression by the mother or infant may protect the infant fromboth intrauterine and breast-milk transmission of the virus.
Delayed clinical progression of HIV also occurs in children whodevelop kwashiorkor (48).
We speculate that HSPG loss in kwashiorkor may representa genetically determined reduction in biosynthesis, promotingdisease if dietary substrates are inadequate and GI infectionsnecessitate reglycosylation, but limiting body protein catabolismin undernutrition and protecting against some infectious diseases.Protection against HIV clearly predates the development ofkwashiorkor. If HSPG biosynthesis is indeed genetically alteredin children at risk of kwashiorkor, those who survive it shouldshow a subclinical reduction in HSPG glycan chain length orsulfation at sites of rapid cellular turnover. This is testable, and, ifconfirmed, would suggest the need for studies of functional
FIGURE 3. Distribution of sulfated glycosaminoglycans (GAGs) and heparan sulfate proteoglycan (HSPG) within the mucosa. A: High-power (originalmagnification 340) images of histochemical staining for sulfated GAGs (left column) and immunohistochemical staining for HSPG and the proteoglycansyndecan-1 in the villous of children with marasmus (Maras) or kwashiorkor (Kwash). The arrows indicate basolateral epithelial localization of sulfated GAGsand HSPG in marasmus. Similar basolateral localization occurs in well-nourished UK children, as reported previously (14, 15) and is functionally important inpreventing intestinal protein leakage (16–18). Basolateral epithelial localization of sulfated GAGs and HSPG was reduced in kwashiorkor, and thesubepithelial basement membrane was less well defined. In contrast, the core protein of epithelial syndecan-1 was normally expressed on the basolateralepithelial border in kwashiorkor as in marasmus. The findings differ from those observed in congenital disorders of N-glycosylation, where the core protein ismislocalized intracellularly (15). Syndecan-1 is also expressed in plasma cells within the lamina propria. B: Low-power (original magnification 320) imagesshowing overall mucosal expression of sulfated GAGs and HSPG in serial sections from children with marasmus, marasmic kwashiorkor (M-K), orkwashiorkor. Expression of GAGs and HSPG was also reduced within the lamina propria in kwashiorkor. Similar staining patterns can be seen in theserial sections stained for GAGs and HSPG (lower row). Staining density index data for the images shown in A and B were similar to the mean values for theoverall groups (data shown in Figure 2).
598 AMADI ET AL
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018

polymorphisms in genes involved in GAG glycan chain synthesisand sulfation (49). Our findings also have potential therapeuticimplications. It was shown that mucosal GAG synthesis in-creased in inflammatory bowel disease after enteral adminis-tration of GlcNAc (50) and PLE was inhibited in animals bya heparin analog (18, 19).
We thank all the scientists and clinicians who contributed to the initial part
of the nutritional intervention study, particularly John Walker-Smith and Mike
Thomson for their contributions to the study design and to SHS International
Ltd for a grant that facilitated the study. We particularly wish to express grat-
itude to members of the team of nurses who worked so hard to achieve a high
standard of clinical care and record keeping, including Elwyn Chomba,
Chifumbe Chintu, Stella Sinyangwe, Justina Hachungula, Bernice Simpito,
Evelyn Mukuka, Angela Watuka, Nayoto Sitwala, Harriet Ngulube, Sandra
Chipepo, Asled Banda, Euphrasia Bwalya, Nalishebo Muyunda, Joyce Nganga,
and Eugenia Kachenjela. We are grateful to Saqib Ansari and Paul Ashwood
for help with the initial biopsy assessment and to Anatoly Shmygol for gen-
erous guidance with the ImageJ analysis.
The authors’ responsibilities were as follows—BA, MM, and PK: per-
formed the clinical studies, including collection of the tissue samples and
the clinical data; AOF, FT, CS, and RD: performed the laboratory analysis
of tissue specimens and contributed to the quantitation; MHG: contributed
to the planning, hypothesis development, and manuscript writing; EAE
and HHF: performed the n-glycosylation studies; SHM: designed and led
the laboratory analysis of tissue specimens, performed quantitative and data
analysis, and drafted the paper; and HHF and SHM: developed the program
of protein-losing enteropathy analysis. None of the authors are aware of any
potential conflicts of interest.
REFERENCES1. Golden MH. Edematous malnutrition. Br Med Bull 1998;54:433–44.2. Amadi B, Kelly P, Mwiya M, et al. Intestinal and systemic infection,
HIV, and mortality in Zambian children with persistent diarrhea andmalnutrition. J Pediatr Gastroenterol Nutr 2001;32:550–4.
3. Courtright P, Canner J. The distribution of kwashiorkor in the southernregion of Malawi. Ann Trop Paediatr 1995;15:221–6.
4. Manary MJ, Brewster DR. Intensive nursing of kwashiorkor in Malawi.Acta Paediatr 2000;89:203–7.
5. Gopalan C. Kwashiorkor and marasmus: evolution and distinguishingfeatures. In: McCance RA, Widdowson EM, eds. Calorie deficienciesand protein deficiencies. London, United Kingdom: Churchill, 1968:48–58.
6. Lin CA, Boslaugh S, Ciliberto HM, et al. A prospective assessment offood and nutrient intake in a population of Malawian children at risk forkwashiorkor. J Pediatr Gastroenterol Nutr 2007;44:487–93.
7. Hendrickse RG. Of sick turkeys, kwashiorkor, malaria, perinatal mor-tality, heroin addicts and food poisoning: research on the influence ofaflatoxins on child health in the tropics. Ann Trop Med Parasitol 1997;91:787–93.
8. Ciliberto H, Ciliberto M, Briend A, Ashorn P, Bier D, Manary M.Antioxidant supplementation for the prevention of kwashiorkor inMalawian children: randomised, double blind, placebo controlled trial.BMJ 2005;330:1109–14.
9. Golden MH, Brooks SE, Ramdath DD, Taylor E. Effacement of glo-merular foot processes in kwashiorkor. Lancet 1990;336:1472–4.
10. Mohanram M, Reddy V. Urinary excretion of acid mucopolysaccharidesin kwashiorkor and vitamin A-deficient children. Clin Chim Acta 1971;34:93–6.
11. Ittyerah TR. Urinary excretion of sulfate in kwashiorkor. Clin ChimActa 1969;25:365–9.
12. Chandrasekaran EV, Mukherjee KL, Bachhawat BK. Isolation andcharacterization of glycosaminoglycans from brain of children withprotein-calorie malnutrition. J Neurochem 1971;18:1913–20.
13. Gunston GD, Burkimsher D, Malan H, Sive AA. Reversible cerebralshrinkage in kwashiorkor: an MRI study. Arch Dis Child 1992;67:1030–2.
14. Murch SH, Winyard PJD, Koletzko S, et al. Congenital enterocyteheparan sulfate deficiency is associated with massive albumin loss, se-cretory diarrhea and malnutrition. Lancet 1996;347:1299–301.
15. Westphal V, Murch S, Kim S, et al. A congenital disorder of glycosyl-ation (CDG-Ic) impairs heparan sulfate proteoglycan accumulation insmall intestine epithelial cells and contributes to protein-losing enter-opathy. Am J Pathol 2000;157:1917–25.
16. Bode L, Eklund EA, Murch SH, Freeze HH. Heparan sulfate loss causesprotein leakage and amplifies TNFa-response in an in vitro model ofprotein-losing enteropathy. Am J Physiol Gastrointest Liver Physiol2005;288:G1015–23.
17. Bode L, Murch S, Freeze HH. Heparan sulfate plays a central role ina dynamic in vitro model of protein-losing enteropathy. J Biol Chem2006;281:7809–15.
18. Bode L, Salvestrini C, Park PW, et al. Heparan sulfate and syndecan-1are essential in maintaining intestinal epithelial barrier function. J ClinInvest 2008;118:229–38.
19. Lencer WI. Patching a leaky intestine. N Engl J Med 2008;359:526–8.20. Amadi B, Mwiya M, Chomba E, et al. Improved nutritional recovery on
an elemental diet in Zambian children with persistent diarrhea andmalnutrition. J Trop Pediatr 2005;51:5–10.
21. Classification of infantile malnutrition. Lancet 1970;296:302–3.22. Campbell DI, Murch SH, Lunn PG, et al. Chronic T cell-mediated en-
teropathy in rural West African children: Relationship with nutritionalstatus and small bowel function. Pediatr Res 2003;54:306–11.
23. David G, Bai XM, Van der Schueren B, Cassiman JJ, Van den Berghe H.Developmental changes in heparan sulfate expression: in situ detectionwith mAbs. J Cell Biol 1992;119:961–75.
24. Bishop JR, Schuksz M, Esko JD. Heparan sulfate proteoglycansfine-tune mammalian physiology. Nature 2007;446:1030–7.
25. Murch SH, MacDonald TT, Walker-Smith JA, Levin M, Lionetti P,Klein NJ. Disruption of sulfated glycosaminoglycans in intestinal in-flammation. Lancet 1993;341:711–4.
26. Dossetor J. Persistent intestinal protein loss after measles. BMJ 1976;2:1231.
27. Shukry AS, Gabr M, Sallam F, Saadani M. Protein-losing gastro-enteropathy in kwashiorkor. J Trop Med Hyg 1965;68:269–71.
28. Bode L. Recent advances on structure, metabolism, and function ofhuman milk oligosaccharides. J Nutr 2006;136:2127–30.
29. De Foliart GR. Insects as food: why the Western attitude is important.Annu Rev Entomol 1999;44:21–50.
30. Luyendyk JP, Copple BL, Barton CC, Ganey PE, Roth RA. Augmen-tation of aflatoxin B1 hepatotoxicity by endotoxin: involvement of en-dothelium and the coagulation system. Toxicol Sci 2003;72:171–81.
31. Karlsson K, Lindahl U, Marklund SL. Binding of human extracellularsuperoxide dismutase C to sulfated glycosaminoglycans. Biochem J1988;256:29–33.
32. Bode L, Freeze HH. Applied glycoproteomics—approaches to studygenetic-environmental collisions causing protein-losing enteropathy.Biochim Biophys Acta 2006;1760:547–59.
33. Jahoor F, Badaloo A, Reid M, Forrester T. Protein metabolism in severechildhood malnutrition. Ann Trop Paediatr 2008;28:87–101.
34. Agbedana EO, Johnson AO, Taylor GO. Studies on hepatic and extra-hepatic lipoprotein lipases in protein-calorie malnutrition. Am J ClinNutr 1979;32:292–8.
35. Kolset SO, Salmivirta M. Cell surface heparan sulfate proteoglycans andlipoprotein metabolism. Cell Mol Life Sci 1999;56:857–70.
36. MacArthur JM, Bishop JR, Stanford KI, et al. Liver heparan sulfateproteoglycans mediate clearance of triglyceride-rich lipoproteins in-dependently of LDL receptor family members. J Clin Invest 2007;117:153–64.
37. Milstone LM, Hough-Monroe L, Kugelman LC, Bender JR, Haggerty JG.Epican, a heparan/chondroitin sulfate proteoglycan form of CD44,mediates cell-cell adhesion. J Cell Sci 1994;107:3183–90.
38. Hird RB, Crawford FA, Spinale FG. Differential effects of protaminesulfate on myocyte contractile function with left ventricular failure.J Am Coll Cardiol 1995;25:773–80.
39. Karli J, Stamatelopoulos S, Karikas G, Levis G, Moulopoulos S. Effectof heparin on myocardial contractility of the dog and on Na, K-ATPase.Res Commun Chem Pathol Pharmacol 1984;43:79–96.
40. Reizes O, Clegg DJ, Strader AD, Benoit SC. A role for syndecan-3 inthe melanocortin regulation of energy balance. Peptides 2006;27:274–80.
41. Ishiguro K, Kadomatsu K, Kojima T, et al. Syndecan-4 deficiency leadsto high mortality of lipopolysaccharide-injected mice. J Biol Chem2001;276:47483–8.
GLYCOSAMINOGLYCANS IN KWASHIORKOR 599
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018

42. Vogt AM, Pettersson F, Moll K, et al. Release of sequestered malaria par-asites upon injection of a glycosaminoglycan. PLoS Pathog 2006;2:e100.
43. Heddini A, Pettersson F, Kai O, et al. Fresh isolates from children withsevere Plasmodium falciparum malaria bind to multiple receptors. InfectImmun 2001;69:5849–56.
44. Kirn TJ, Jude BA, Taylor RK. A colonization factor links Vibrio chol-erae environmental survival and human infection. Nature 2005;438:863–6.
45. Bobardt MD, Salmon P, Wang L, et al. Contribution of proteoglycans tohuman immunodeficiency virus type 1 brain invasion. J Virol 2004;78:6567–84.
46. Kessler L, Daley H, Malenga G, Graham S. The impact of the humanimmunodeficiency virus type 1 on the management of severe malnu-trition in Malawi. Ann Trop Paediatr 2000;20:50–6.
47. Angami K, Reddy SV. Singh KhI, Singh NB, Singh PI. Prevalence of
HIV infection and AIDS symptomatology in malnourished children:
a hospital based study. J Commun Dis 2004;36:45–52.48. Bachou H, Tylleskar T, Downing R, Tumwine JK. Severe malnutrition
with and without HIV-1 infection in hospitalised children in Kampala,
Uganda: differences in clinical features, haematological findings and
CD41 cell counts. Nutr J 2006;5:27.49. Esko JD, Selleck SB. Order out of chaos: assembly of ligand binding
sites in heparan sulfate. Annu Rev Biochem 2002;71:435–71.50. Salvatore S, Heuschkel RB, Tomlin S, et al. Therapeutic efficacy of
N-acetyl glucosamine, a nutritional substrate for glycosaminoglycan
synthesis, in chronic inflammatory bowel disease. Aliment Pharmacol
Ther 2000;14:1567–79.
600 AMADI ET AL
Downloaded from https://academic.oup.com/ajcn/article-abstract/89/2/592/4596718by gueston 04 May 2018