Embed Size (px)
Citation preview

The Journal of Neuroscience, November 1994, f4(11): 6392-6401
Regional Modulation of Neurofilament Organization by Myelination in Normal Axons
Sung-Tsang Hsieh,lv5 Grahame J. Kidd,2 Thomas 0. Crawford, 2-4 Zuoshang XU,~ Whei-Min Lin,2 Bruce D. Trapp,2 Don W. Cleveland,1~3 and John W. Griffin1v2
Departments of ‘Neuroscience, 2Neurology, 3Biological Chemistry, and 4Pediatrics, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21287 and 5Department of Neurology, National Taiwan University Hospital, Taipei, Taiwan, Republic of China
Previous studies in the hypomyelinating mouse mutant Trembler have suggested that demyelinating axons are smaller in caliber compared to normal axons, and that there are differences in the organization of axonal neurofilaments. In the normal PNS, however, the relationship between neu- rofilament organization and myelination has not been inves- tigated extensively. In normal axons, only the initial seg- ments, the nodes of Ranvier (- 1 pm), and the terminals are not covered by myelin. We took advantage of an unusual feature of the primary sensory neurons in the dorsal root ganglion, the relatively long nonmyelinated stem process (up to several hundred micrometers), to determine if the pres- ence of myelination correlates with differences in cytoskel- eta1 organization and neurofilament phosphorylation. Axonal caliber and neurofilament numbers were substantially great- er in the myelinated internodes than in the stem process or nodes of Ranvier. Neurofilament spacing, assessed by mea- suring the nearest-neighbor neurofilament distance, was 25- 50% less in the stem processes and nodes of Ranvier than in the myelinated internodes. In the myelinated internodes, neurofilaments had greater immunoreactivity for phosphor- ylated epitopes than those in the stem process. These find- ings indicate that interactions with Schwann cells modulate neurofilament phosphorylation within the ensheathed axonal segments, and that increased phosphorylation within my- elinated internodes leads to greater interfilament spacing. Lastly, the myelinated internodes had three fold more neu- rofilaments, but the same number of microtubules. Both the increased neurofilament spacing and the increase in neu- rofilament numbers in myelinated internodes contribute to a greater axonal caliber in the myelinated internodes.
[Key words: neurofilament phosphorylation, nearest- neighbor distance, myelination, Schwann cell, dorsal root ganglion, initial segment, stem process, node of Ranvier, axonal caliber, morphometty]
Received Aug. 27, 1993; revised Mar. 11, 1994; accepted Mar. 29, 1994. This work was supported by NIH Grants to J.W.G. (NINDSGrant ROl-14784);
to J.W.G., D.W.C., and B.D.T. (NS POl-22849), to S.-T.H. (NIMH Grant 5T32 MH 18030); and to T.O.C. (NINDS CIDA NS 0 14 12). We thank Eileen Mulrenin and Carol Haney for technical assistance; Drs. Paul Hoffman, Michael Shelanski, and Phil Wong for suggestions and discussion, and Rod Graham for editorial assistance. G.J.K. and Z.X. are postdoctoral fellows of the National Multiple Sclerosis Society, and the Muscular Dystrophy Association, respectively.
Correspondence should bc addressed to Dr. John W. Griffin, Johns Hopkins Hospital, Department of Neurology, Meyer 6-109, 600 North Wolfe Street, Bal- timore, MD 21287-7609.
Copyright 0 1994 Society for Neuroscience 0270-6474/94/146392-10$05.00/0
In large neurons neurofilaments are the most abundant cyto- skeletal organelles, and have been postulated to act as important intrinsic determinants of axonal caliber (Hoffman et al., 1987). Neurofilaments in vertebrates are composed of three related proteins with apparent molecular weights of 68, 150, 200 kDa (NF-L, NF-M, and NF-H, respectively) (Hoffman and Lasek, 1975; Liem et al., 1978; Geisler et al., 1983; Willard and Simon, 1983; Hirokawa et al., 1984; Schlaepfer, 1987). All three ofthese proteins share a central rod domain, but they differ in their C-terminal regions. In NF-H, this region contains over 40 ly- sine-serine-proline repeats (Julien et al., 1986, 1988; Lees et al., 1988), which provide potential phosphorylation sites (Julien et al., 1983; Geisler et al., 1983; Lee et al., 1988). The extent of phosphorylation of this region, which contributes to the side- arms of the neurofilament (Hirokawa et al., 1984; Carden et al., 1985; Hisanaga and Hirokawa, 1988) appears to differ within different regions of the neuron, with a higher phosphorylation state in the axon than in the cell body and dendrite (Stemberger and Sternberger, 1983; Lee et al., 1986, 1987). One attractive hypothesis is that phosphorylation on the side arms may in- crease spacings between nearest-neighbor neurofilaments (Car- den et al., 1987), and thereby achieve a larger axonal caliber.
Axonal caliber is a fundamental phenotypic feature of neu- rons. For each of the different functional classes of peripheral nerve fibers, there is a restricted range of normal axonal calibers. The conduction velocity of nerve action potential is a function offiber diameter (for review, see Waxman, 1980). Axonal caliber is considered to be largely determined developmentally. A spe- cial role of neurofilaments in influencing caliber has been de- duced from the observation that neurofilament density is rela- tively constant among axons of different calibers (Friede and Somarajski, 1970; Hoffman et al., 1984), although regional dif- ferences (Nixon and Logvinenko, 1986; Lasek, 1988; Price et al, 1990; Reles and Friede, I99 l), changes in axonal diseases (Parhad et al., 1987; Dyck et al., 1990) and alterations due to interrupted axon-target interactions (Hoffman et al., 1984) all indicate that neither neurofilament number nor density is fixed or immutable.
Few studies have examined the possible effects of ensheathing cells or myelination on the axon or its cytoskeleton (Aguayo et al., 1977; Windebank et al., 1985; de Waegh et al., 1992). In the PNS, all axons are ensheathed by Schwann cells. Schwann cells elaborate specialized lamellated plasmalemma sheaths, the myelin sheaths, around some of the PNS axons. These myelin sheaths are interrupted periodically at the nodes of Ranvier. Myelination is associated with increased axonal caliber in mye-

The Journal of Neuroscience, November 1994, 14(11) 6393
linating tissue culture (Windebank et al., 1985) and the nodes of Ranvier are smaller than the neighboring myelinated inter- nodes (Berthold, 1978). From these observations, it has been presumed that myelination has some effect on the axonal cy- toskeleton. In a series of studies by nerve-graft experiments, de Waegh and colleagues provided a possible basis for this effect in studies of the hypomyelinating/demyelinating mouse mutant, Trembler (de Waegh and Brady, 1990, 1991; de Waegh et al., 1992). Compared to normally myelinated internodes, the Trem- bler internodes were smaller, had closer neurofilament spacing, and had lower levels of phosphorylated NF-H. Based on these observations, de Waegh et al. (1992) proposed that the regional variations in neurofilament organization is modulated by mye- linating Schwann cells.
The effect of myelination on the regional variations of axonal cytoskeleton in the normal PNS has not been examined com- prehensively. In most large axons, only the short initial segments and the nodes of Ranvier (- 1 pm) are not covered by myelin. In this study, we have taken advantage of an unusual feature of the large primary sensory neurons in the dorsal root ganglion (DRG) and trigeminal ganglion, the nonmyelinated axonal stem process, which is usually of tens of micrometers in length (Fig. 1.4). We asked whether the stem process differed from the my- elinated internodes in terms of axonal size, cytoskeletal com- position, neurofilament spacing, and the level of neurofilament phosphorylation. The results indicate that those regions covered by myelin are different in all of these features of the axonal cytoskeleton, and are consistent with the hypothesis that mye- linating Schwann cells exert a major extrinsic influence on the organization of the axon.
Materials and Methods Tissue preparation: primary sensory ganglion neurons We studied the primary sensory neurons and their axons in the rat lumbar dorsal root ganglia, and trigeminal ganglion. The stem process, the nonmyelinated portion of the axon that extends from the cell body of the sensory neurons to the first myelin heminode (Spencer et al., 1973), varies in its length and course. Long stem processes wrap around their cell bodies in a glomerular fashion. On cross sections, the stem process was easily recognized as a nonmyelinated axon lying near the cell body and surrounded by satellite cells (Fig. 1B); in neurons with glomerular arrangement the stem process could be seen more than once in a single cross section.
Groups ofsprague-Dawley rats (8-12 weeks ofage, with body weights of 200-300 gm) and C57 mice (6-8 weeks of age) were anesthetized with 4% chloral hydrate and animals were fixed by intracardiac perfusion with 4% paraformaldehyde/3% glutaraldehyde (for ultrastructural mor- phometry), 4% paraformaldehyde (for light microscopic immunocyto- chemistry), or 4% paraformaldehvde/OS% dutaraldehvde (for electron microscopic immunocytochemistry) in 0.1 M Sorenson’s phosphate buffer. The lumbar DRGs and trigeminal ganglia with roots attached were dissected out and postfixed in the same fixatives for another 24 hr. and then transferred to buffers.
Ultrastructural morphometry Preparalions. The lumbar DRGs and dorsal roots were postfixed in 2% osmium tetroxide, embedded in Epon and sectioned in an ultramicro- tome. Relatively thick thin-sections (silver-gold to gold reflection) were picked up on Formvar-coated grids, stained with uranyl acetate and lead citrate, and observed under electron microscope. Individual stem processes were identified and photographed at 10,000 x and 40,000 x . For each stem process the adjacent myelinated axon and the nerve cell body were photographed at the same magnifications.
To compare the cytoskeletal organization between the node of Ran- vier and the internode of the same axon, serial thin sections intermixed
- Hgure 1. The diagram of a dorsal root ganglion (DRG) neuron. The stem process, the nonmyelinated axonal portion, extends from the cell body to the first internode. If a cross section was made through the dashed line of the schematic DRG neuron (A), the perikaryon (p), stem process (s), and the myelinated internodes (m) would appear on the same section, as illustrated in the electron micrograph in B. The stem process was covered by satellite cells, which did not make myelin sheaths.
with thick sections of the dorsal roots (total of 300 thin sections and 20 thick sections) were cut. The criteria of defining a node of Ranvier included (1) the absence of myelin, (2) the presence of Schwann cell microvilli, and (3) the presence of dense undercoatings on axonal mem- brane. When a node of Ranvier was identified, we traced forward and backward to find its corresponding internode and photographed both sites at 6000 x , 10,000 x , and 40,000 x
Cytoskeletal composition. To estimate the cytoskeletal composition, we modified the previous method of Parhad et al. (1987). Briefly, we divided the whole cross-section axonal area into tiles of 1 pm2, assigned consecutive numbers (beginning from 1) on each tile, chose one to five tiles based on the numbers of a random table, and counted the whole axonal neurofilament densities (expressed as neurofilaments/~m* axonal area) on those selected squares. The mean of the whole axonal neuro- filament densities for each sampled axon was multiplied by the axonal area to give the estimated total number of neurofilaments for that axon. The same procedure was repeated for the estimation of the total number of microtubules per axon. For stem process and node of Ranvier, one to five 1 pm* tiles constituted 50-90% of axonal area. For myelinated axons, the sampled regions would be responsible for 1640% of axonal area. In the myelinated axons, the region occupied by membrane-bound organelles (mitochondria, smooth endoplasmic reticulum, and vesicular organelles) was negligible (less than 5% of axonal area) (Berthold, 1978; Hoffman et al., 1984). In the stem process and nodes particulate or- ganelles occupied a greater area of the cross-sectional area, sothat the whole axonal density data cannot be used for comparing neurofilament spacing. Price et al. (1990) and de Waegh et al. (1992) calculated neu- rofilament packing density by excluding the area occupied by particulate organelles. In this study, we used nearest-neighbor distance to compare neurofilament spacing (see below); the whole axonal density data were used to calculate the total number of neurofilaments and microtubules.
Nearest-neighbor distance. We used the analysis of nearest-neighbor distance to measure neurofilament spacing. The nearest-neighbor dis- tance approach identifies obligate minimum spacing and the degree of variations in spacing of neurofilaments. Both nearest-neighbor distance and packing density are complementary and necessary to understand neurofilament organization and distribution (Hsieh et al., 1994). Briefly, the negatives of electron micrographs were illuminated on a view box, imaged with a video camera. Each neurofilament that was oriented in true transverse section was accepted and digitized with the BIOQUANT
software (R&M Biometrics, Inc., Nashville, TN). The data of x-y or- dinates for each neurofilament were exported to a database. A program was written to calculate the nearest-neighbor distance for each neuro- filament. In addition, axonal area, axonal circumference, and myelin sheath thickness were measured with the BIOQUANT software to compare their relationships with neurofilament spacing.

6394 Hsieh et al. l Myelination Modulates Neurofilament Organization and Phosphorylation
200.
116-
97-
66-
45
Figure 2. Immunoblotting to characterize the polyclonal antibody pAb- NFH. The antibody pAb-NFH was made against the C-terminus ofNF- H. Protein extract of mouse spinal cord was electrophoresed on 7% SDS-polyacrylamide gel and stained with Coomassie blue (lane I). A duplicate gel was transferred onto a nitrocellulose paper (lane 2). Im- munoblotting in lane 2 indicated that pAb-NFH recognized NF-H as a discrete band.
Antibodies against d@erent epitopes of neuroJilament proteins We used immunoreactivity against different epitopes of NF-H (phos- phorylated, nonphosphorylated and phosphorylation-independent ep- itopes) to assess the regional difference of neurofilament phosphoryla- tion. SMI-3 1 and SMI-32 are mouse monoclonal antibodies (Stemberger Monoclonals Inc., Baltimore, MD). They have been characterized in terms of their phosphorylation status. SMI-31 is directed against the extensively phosphorylated form of NF-H, the sequence motif contain- ing highly phosphorylated lysine-serine-proline. The SMI-32 recognize a nonphosphorylated or relatively hypophosphorylated form of this motif on neurofilaments (Stemberaer and Stemberger, 1983; Lee et al., 1988).
For the purpose of recognizing all the NF-H, independent of phos- phorylation state, a polyclonal antibody (termed pAb-NFH in this re- port) was made against NF-H. The peptide CYEKTTEDKATKGEK (one-letter code for amino acid), which corresponds to the carboxyl terminal 13 amino acids of mouse NF-H (Julien et al., 1986; Julien et al., 1988) plus C and Y at its N-terminus for coupling and UV detection, was chemically synthesized using a peptide synthesizer. The peptide was then coupled to keyhole limpet hemocyanin and used to immunize rabbits. Purification of the antibody was carried out as described pre- viously (Lopata and Cleveland, 1987).
To characterize the antibodv nAb-NFH, approximatelv 20 pg of total protein extract from mouse spinal cord wabklectrophoresed on a 7% SDS-polyacrylamide gel (Laemmli, 1970), and stained with Coomassie blue. A duplicate gel was transferred onto a nitrocellulose filter as de- scribed before (Lopata and Cleveland, 1987). The nitrocellulose filter was then incubated with the purified antibody pAb-NFH in 2% powder milk and PBS for 2 hr. After the filter was washed five times with PBS, it was incubated with ‘*+protein A (Amersham Corp., Arlington Heights, IL) for 1 hr. The filter was then repeatedly washed and exposed to x-ray film. On immunoblotting, pAb-NFH recognized neurofilaments as a band at the 200 kDa position on immunoblotting (Fig. 2). Because there are no potential se&k phosphorylation sites in this region ofthe carboxy terminus of NF-H (Julien and Mushynski, 1983; Julien et al., 1986, 1988) pAb-NFH should recognize the total NF-H independent of phos- phorylation status.
Immunocytochemistry and confocal laser scanning microscopy The dissected rat lumbar DRGs, rat, and mouse trigeminal ganglia were cryoprotected with 20% glycerol in the Sorenson’s buffer for 24 hr and
then cut on a freezing microtome. Sections of 20-30 pm thickness were rinsed in PBS solution, incubated with 25% Triton X-100 in PBS for 24 hr to increase antibody penetration, and then blocked with 5% normal horse serum for 30 min. Tissue sections were labelled by incubating them in a combination of mouse and rabbit antibodies in 1% Triton X-loo/PBS at 4°C for 24 hr. The primary antibodies used in this study included the monoclonal antibodies against phosphorylated neurofila- ments (SMI-3 1) and nonphosphorylated neurofilaments (SMI-32), and a polyclonal antibody against myelin protein P, (Trapp et al., 1981). The sections were washed and then incubated in a mixture of fluorescein isothiocyanate (FITC)-conjugated sheep anti-mouse and Texas red-con- jugated donkey anti-rabbit antibodies (Amersham Corp., Arlington Heights, IL) for 2 hr. The sections were washed in PBS and mounted in a fade-resisting mowiol-based mounting medium containing para- phenylene diamine.
The sections were examined with standard epifluorescence and im- aged with a Leica confocal laser scanning microscope. Optical sections of approximately 0.5-l pm thick were collected using a line averaging technique. The Texas red and FITC images were collected simulta- neously. In some cases, serial sequences of optical sections were obtained at l-2 pm intervals through the profiles. The continuity and staining patterns of myelinated and non-myelinated portions of the same axon were investigated using these “z-series” either viewed as a sequence of single images or in three-dimensional reconstructions and rotations.
Electron microscopic immunocytochemistry The procedure of electron microscopic immunocytochemistry followed that previously reported (Trapp et al., 1989). Briefly, the rat lumbar DRGs were infiltrated with 1 M, 2 M, and 2.3 M sucrose solution con- taining 30% polyvinylpyrrolidone. Frozen sections were cut on a Reich- ert ultramicrotome in a liquid nitrogen<ooled chamber (around - 100°C). Thin sections around 100 nm in thickness were picked up on Formvar- coated grids, rinsed with PBS, blocked with ovalbumin/normal goat serum, and incubated with primary antibodies (SMI-3 1 and pAb-NFH) for 16-24 hr. After washing in PBS, grids were incubated with 10 nm colloidal gold-labeled goat anti-mouse IgG (for SMI-3 1) and 5 nm col- loidal gold-labeled goat anti-rabbit IgG (for pAb-NFH) for 2 hr. The sections were fixed with glutaraldehyde, stained with uranyl acetate, embedded in uranyl acetate-containing methyl cellulose, and observed under a Hitachi 600 electron microscope.
To assess the relative intensity of labeling, stem processes and my- elinated axons on the grids were photographed at 60,000 x . The densities of immunogold particles in the axons were measured using BI~QUANT
software.
Statistical analysis T test, Wilcoxon ranked sums test, and regression statistics were gen- erated by using the STATISTICAL ANALYSIS SYSTEM program (SAS Institute Inc., Cary, NC). Differences with p < 0.05 were considered statistically significant.
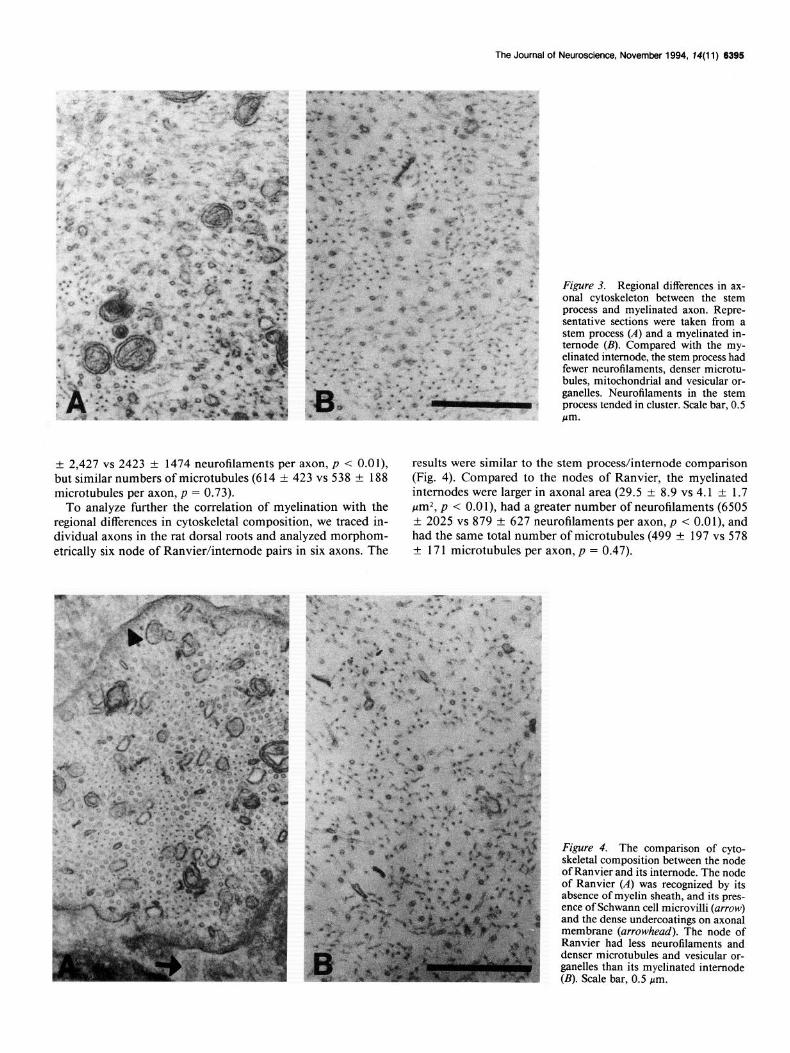
Results Axonal caliber and cytoskeletal composition in myelinated and nonmyelinated segments To test whether variations of axonal caliber and cytoskeletal composition were related to the myelination status of the axonal segment, we sampled eight stem processes and nine neighboring myelinated axons and compared the following parameters: ax- onal caliber, and the total number of neurofilaments, and mi- crotubules per axon. Myelinated axons were larger than stem processes [axonal area: 26.3 f 9.4 vs 11.0 f 4.8 pm2 (mean f SD), p < 0.011. Ultrastructurally, neurofilaments appeared as 10 nm electron-dense dots in axonal cross section and had side arms extending as thin threads from the core of neurofilaments. The most impressive feature of the cytoskeleton in the stem process was the greater proportion of microtubules, and other membrane-bound organelles and the smaller proportion of neu- rofilaments compared to the pattern in the myelinated segments (Fig. 3; Berthold, 1978). Compared to the stem process, the myelinated internodes had threefold more neurofilaments (6760
The Journal of Neuroscience, November 1994, 14(11) 6395
Figure 3. Regional differences in ax- onal cytoskeleton between the stem process and myelinated axon. Repre- sentative sections were taken from a stem process (A) and a myelinated in- ternode (II). Compared with the my- elinated internode, the stem process had fewer neurotilaments, denser microtu- bules, mitochondrial and vesicular or- ganelles. Neurofilaments in the stem process tended in cluster. Scale bar, 0.5 m
+ 2,427 vs 2423 f 1474 neurofilaments per axon, p < O.Ol), results were similar to the stem process/internode comparison but similar numbers of microtubules (6 14 f 423 vs 538 k 188 (Fig. 4). Compared to the nodes of Ranvier, the myelinated microtubules per axon, p = 0.73). internodes were larger in axonal area (29.5 f 8.9 vs 4.1 rt 1.7
To analyze further the correlation of myelination with the pmZ, p < O.Ol), had a greater number of neurofilaments (6505 regional differences in cytoskeletal composition, we traced in- + 2025 vs 879 + 627 neurofilaments per axon, p < O.Ol), and dividual axons in the rat dorsal roots and analyzed morphom- had the same total number of microtubules (499 k 197 vs 578 etrically six node of Ranvier/internode pairs in six axons. The f 171 microtubules per axon, p = 0.47).
Figure 4. The comparison of cyto- skeletal composition between the node of Ranvier and its internode. The node of Ranvier (A) was recognized by its absence of myelin sheath, and its pres- ence of Schwann cell microvilli (arrow) and the dense undercoatings on axonal membrane (arrowhead). The node of Ranvier had less neurofilaments and denser microtubules and vesicular or- ganelles than its myelinated internode (B). Scale bar, 0.5 pm.
6396 Hsieh et al. - Myelination Modulates Neurofilament Organization and Phosphorylation
10 22 34 46 58 70 Neamd-neighbor nsurofilament distance (nm)
Nearest-neighbor neurofilament distance (nm)
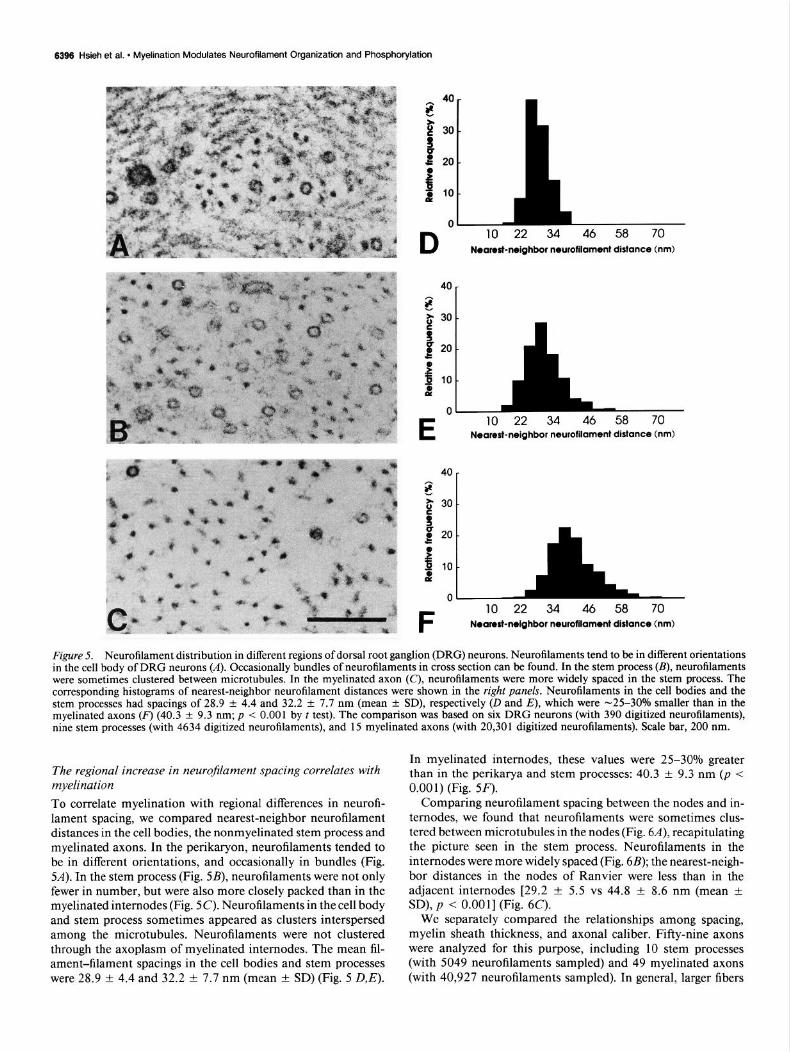
Figure 5. Neurofilament distribution in different regions of dorsal root ganglion (DRG) neurons. Neurofilaments tend to be in different orientations in the cell body of DRG neurons (A). Occasionally bundles of neurofilaments in cross section can he found. In the stem process (B), neurofilaments were sometimes clustered between microtubules. In the myelinated axon (C), neurofilaments were more widely spaced in the stem process. The corresponding histograms of nearest-neighbor neurofilament distances were shown in the right panels. Neurofilaments in the cell bodies and the stem processes had spacings of 28.9 & 4.4 and 32.2 f 7.7 nm (mean +- SD), respectively (D and E), which were -25-30% smaller than in the myelinated axons (F) (40.3 + 9.3 nm; p < 0.001 by t test). The comparison was based on six DRG neurons (with 390 digitized neurofilaments), nine stem processes (with 4634 digitized neurofilaments), and 15 myelinated axons (with 20,301 digitized neurofilaments). Scale bar, 200 nm.
The regional increase in neurofilament spacing correlates with myelination To correlate myelination with regional differences in neurofi- lament spacing, we compared nearest-neighbor neurofilament distances in the cell bodies, the nonmyelinated stem process and myelinated axons. In the perikaryon, neurofilaments tended to be in different orientations, and occasionally in bundles (Fig. 54). In the stem process (Fig. 5B), neurofilaments were not only fewer in number, but were also more closely packed than in the myelinated internodes (Fig. X). Neurofilaments in the cell body and stem process sometimes appeared as clusters interspersed among the microtubules. Neurofilaments were not clustered through the axoplasm of myelinated internodes. The mean fil- ament-filament spacings in the cell bodies and stem processes were 28.9 + 4.4 and 32.2 f 7.7 nm (mean f SD) (Fig. 5 D,E).
In myelinated internodes, these values were 25-30% greater than in the perikarya and stem processes: 40.3 + 9.3 nm (p < 0.001) (Fig. 5F).
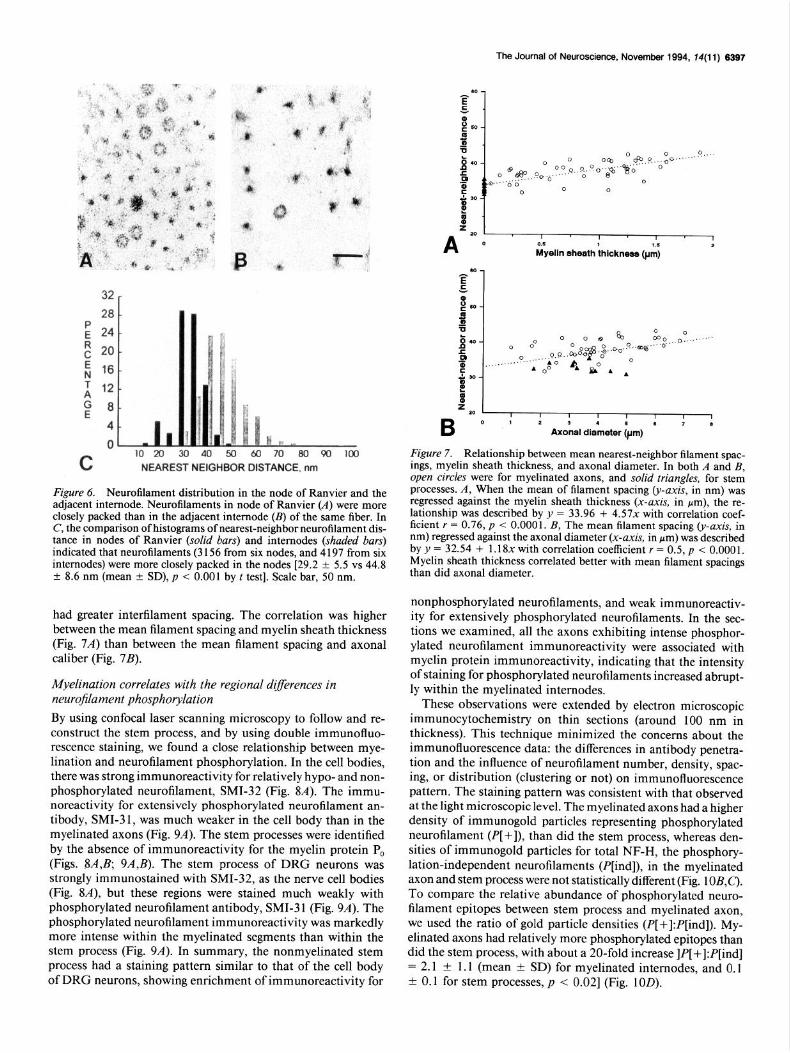
Comparing neurofilament spacing between the nodes and in- ternodes, we found that neurofilaments were sometimes clus- tered between microtubules in the nodes (Fig. 6A), recapitulating the picture seen in the stem process. Neurofilaments in the internodes were more widely spaced (Fig. 6B); the nearest-neigh- bor distances in the nodes of Ranvier were less than in the adjacent internodes [29.2 f 5.5 vs 44.8 f 8.6 nm (mean -t SD), p < O.OOl] (Fig. 6C).
We separately compared the relationships among spacing, myelin sheath thickness, and axonal caliber. Fifty-nine axons were analyzed for this purpose, including 10 stem processes (with 5049 neurofilaments sampled) and 49 myelinated axons (with 40,927 neurofilaments sampled). In general, larger fibers
The Journal of Neuroscience, November 1994, 14(11) 6397
28
(; 24
E 20
; 16
T
2 12 8
4
0 A 10 20 30 40 50 60 70 80 90 loo
L NEAREST NEIGHBOR DISTANCE, nm
Figure 6. Neurofilament distribution in the node of Ranvier and the adjacent internode. Neurofilaments in node of Ranvier (A) were more closely packed than in the adjacent internode (B) of the same fiber. In C, the comparison of histograms of nearest-neighbor neurofilament dis- tance in nodes of Ranvier (solid bars) and internodes (shaded bars) indicated that neurofilaments (3 156 from six nodes, and 4 197 from six internodes) were more closely packed in the nodes [29.2 + 5.5 vs 44.8 ? 8.6 nm (mean ? SD), p < 0.001 by t test]. Scale bar, 50 nm.
had greater interfilament spacing. The correlation was higher between the mean filament spacing and myelin sheath thickness (Fig. 74 than between the mean filament spacing and axonal caliber (Fig. 7B).
Myelination correlates with the regional d@erences in neurojlament phosphorylation
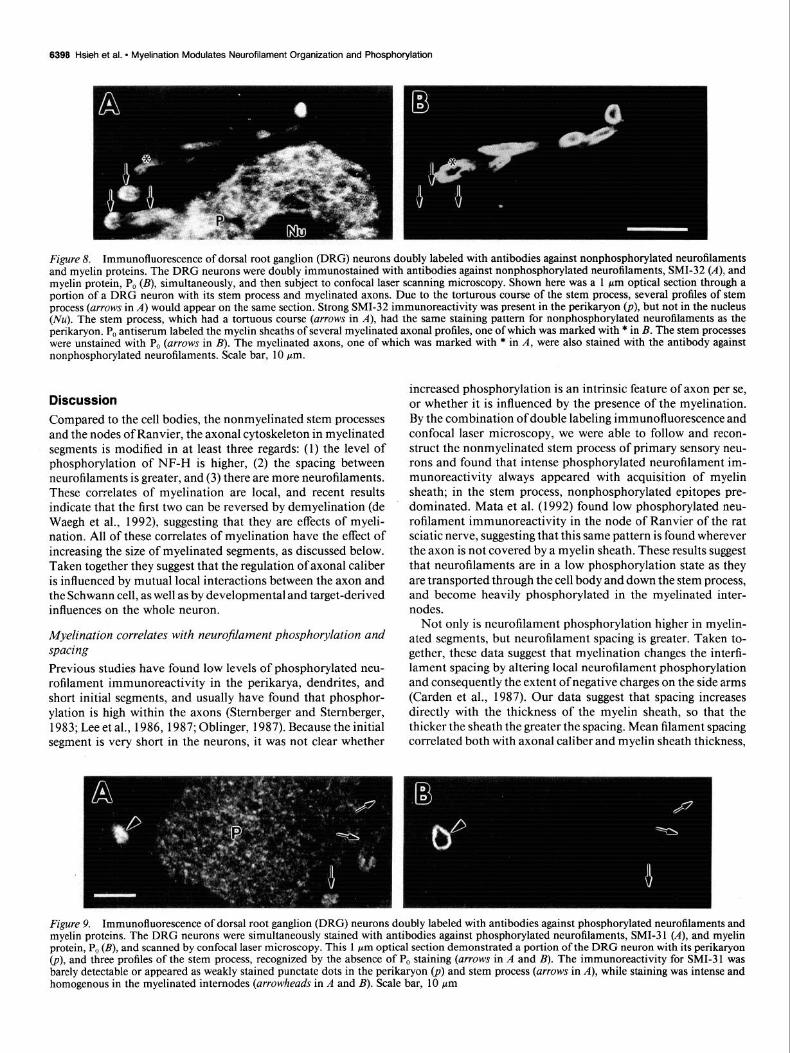
By using confocal laser scanning microscopy to follow and re- construct the stem process, and by using double immunofluo- rescence staining, we found a close relationship between mye- lination and neurofilament phosphorylation. In the cell bodies, there was strong immunoreactivity for relatively hypo- and non- phosphorylated neurofilament, SMI-32 (Fig. 84). The immu- noreactivity for extensively phosphorylated neurofilament an- tibody, SMI-3 1, was much weaker in the cell body than in the myelinated axons (Fig. 94. The stem processes were identified by the absence of immunoreactivity for the myelin protein P, (Figs. &4,B, 9A,B). The stem process of DRG neurons was strongly immunostained with SMI-32, as the nerve cell bodies (Fig. 84, but these regions were stained much weakly with phosphorylated neurofilament antibody, SMI-3 1 (Fig. 9.4). The phosphorylated neurofilament immunoreactivity was markedly more intense within the myelinated segments than within the stem process (Fig. 94. In summary, the nonmyelinated stem process had a staining pattern similar to that of the cell body of DRG neurons, showing enrichment of immunoreactivity for
Figure 7. Relationship between mean nearest-neighbor filament spac- ings, myelin sheath thickness, and axonal diameter. In both A and B, open circles were for myelinated axons, and solid triangles, for stem processes. A, When the mean of filament spacing (y-axis, in nm) was regressed against the myelin sheath thickness (x-axis, in pm), the re- lationship was described by y = 33.96 + 4.57x with correlation coef- ficient r = 0.76, p < 0.000 1. B, The mean filament spacing (y-axis, in nm) regressed against the axonal diameter (x-axis, in pm) was described by y = 32.54 + 1.18x with correlation coefficient r = 0.5, p < 0.0001, Myelin sheath thickness correlated better with mean filament spacings than did axonal diameter.
nonphosphorylated neurofilaments, and weak immunoreactiv- ity for extensively phosphorylated neurofilaments. In the sec- tions we examined, all the axons exhibiting intense phosphor- ylated neurofilament immunoreactivity were associated with myelin protein immunoreactivity, indicating that the intensity of staining for phosphorylated neurofilaments increased abrupt- ly within the myelinated internodes.
These observations were extended by electron microscopic immunocytochemistry on thin sections (around 100 nm in thickness). This technique minimized the concerns about the immunofluorescence data: the differences in antibody penetra- tion and the influence of neurofilament number, density, spac- ing, or distribution (clustering or not) on immunofluorescence pattern. The staining pattern was consistent with that observed at the light microscopic level. The myelinated axons had a higher density of immunogold particles representing phosphorylated neurofilament (P[+]), than did the stem process, whereas den- sities of immunogold particles for total NF-H, the phosphory- lation-independent neurofilaments (P[ind]), in the myelinated axon and stem process were not statistically different (Fig. 1 O&C). To compare the relative abundance of phosphorylated neuro- filament epitopes between stem process and myelinated axon, we used the ratio of gold particle densities (P[+]:P[ind]). My- elinated axons had relatively more phosphorylated epitopes than did the stem process, with about a 20-fold increase ]P[+]:P[ind] = 2.1 -t 1.1 (mean f SD) for myelinated internodes, and 0.1 f 0.1 for stem processes, p -C 0.021 (Fig. 1OD).
6396 Hsieh et al. * Myelination Modulates Neurofilament Organization and Phosphorylation
Figure 8. Immunofluorescence of dorsal root ganglion (DRG) neurons doubly labeled with antibodies against nonphosphorylated neurofilaments and myelin proteins. The DRG neurons were doubly immunostained with antibodies against nonphosphorylated neurofilaments, SMI-32 (A), and myelin protein, P, (B), simultaneously, and then subject to confocal laser scanning microscopy. Shown here was a 1 pm optical section through a portion of a DRG neuron with its stem process and myelinated axons. Due to the torturous course of the stem process, several profiles of stem process (arrows in A) would appear on the same section. Strong SMI-32 immunoreactivity was present in the perikaryon (p), but not in the nucleus (Nu). The stem process, which had a tortuous course (arrows in A), had the same staining pattern for nonphosphorylated neurofilaments as the perikaryon. P, antiserum labeled the myelin sheaths of several myelinated axonal profiles, one of which was marked with * in B. The stem processes were unstained with P, (arrows in B). The myelinated axons, one of which was marked with * in A, were also stained with the antibody against nonphosphorylated neurofilaments. Scale bar, 10 pm.
Discussion Compared to the cell bodies, the nonmyelinated stem processes and the nodes of Ranvier, the axonal cytoskeleton in myelinated segments is modified in at least three regards: (1) the level of phosphorylation of NF-H is higher, (2) the spacing between neurofilaments is greater, and (3) there are more neurofilaments. These correlates of myelination are local, and recent results indicate that the first two can be reversed by demyelination (de Waegh et al., 1992), suggesting that they are effects of myeli- nation. All of these correlates of myelination have the effect of increasing the size of myelinated segments, as discussed below. Taken together they suggest that the regulation of axonal caliber is influenced by mutual local interactions between the axon and the Schwann cell, as well as by developmental and target-derived influences on the whole neuron.
Myelination correlates with neurojilament phosphorylation and spacing
Previous studies have found low levels of phosphorylated neu- rofilament immunoreactivity in the perikarya, dendrites, and short initial segments, and usually have found that phosphor- ylation is high within the axons (Stemberger and Stemberger, 1983; Lee et al., 1986, 1987; Oblinger, 1987). Because the initial segment is very short in the neurons, it was not clear whether
increased phosphorylation is an intrinsic feature of axon per se, or whether it is influenced by the presence of the myelination. By the combination of double labeling immunofluorescence and confocal laser microscopy, we were able to follow and recon- struct the nonmyelinated stem process of primary sensory neu- rons and found that intense phosphorylated neurofilament im- munoreactivity always appeared with acquisition of myelin sheath; in the stem process, nonphosphorylated epitopes pre- dominated. Mata et al. (1992) found low phosphorylated neu- rofilament immunoreactivity in the node of Ranvier of the rat sciatic nerve, suggesting that this same pattern is found wherever the axon is not covered by a myelin sheath. These results suggest that neurofilaments are in a low phosphorylation state as they are transported through the cell body and down the stem process, and become heavily phosphorylated in the myelinated inter- nodes.
Not only is neurofilament phosphorylation higher in myelin- ated segments, but neurofilament spacing is greater. Taken to- gether, these data suggest that myelination changes the interfi- lament spacing by altering local neurofilament phosphorylation and consequently the extent of negative charges on the side arms (Carden et al., 1987). Our data suggest that spacing increases directly with the thickness of the myelin sheath, so that the thicker the sheath the greater the spacing. Mean filament spacing correlated both with axonal caliber and myelin sheath thickness,
Figure 9. Immunofluorescence of dorsal root ganglion (DRG) neurons doubly labeled with antibodies against phosphorylated neurofilaments and myelin proteins. The DRG neurons were simultaneously stained with antibodies against phosphorylated neurofilaments, SMI-3 1 (A), and myelin protein, P, (B), and scanned by confocal laser microscopy. This 1 pm optical section demonstrated a portion of the DRG neuron with its perikaryon @), and three profiles of the stem process, recognized by the absence of PO staining (arrows in A and B). The immunoreactivity for SMI-31 was barely detectable or appeared as weakly stained punctate dots in the perikaryon @) and stem process (arrows in A), while staining was intense and homogenous in the myelinated internodes (arrowheads in A and B). Scale bar, 10 pm
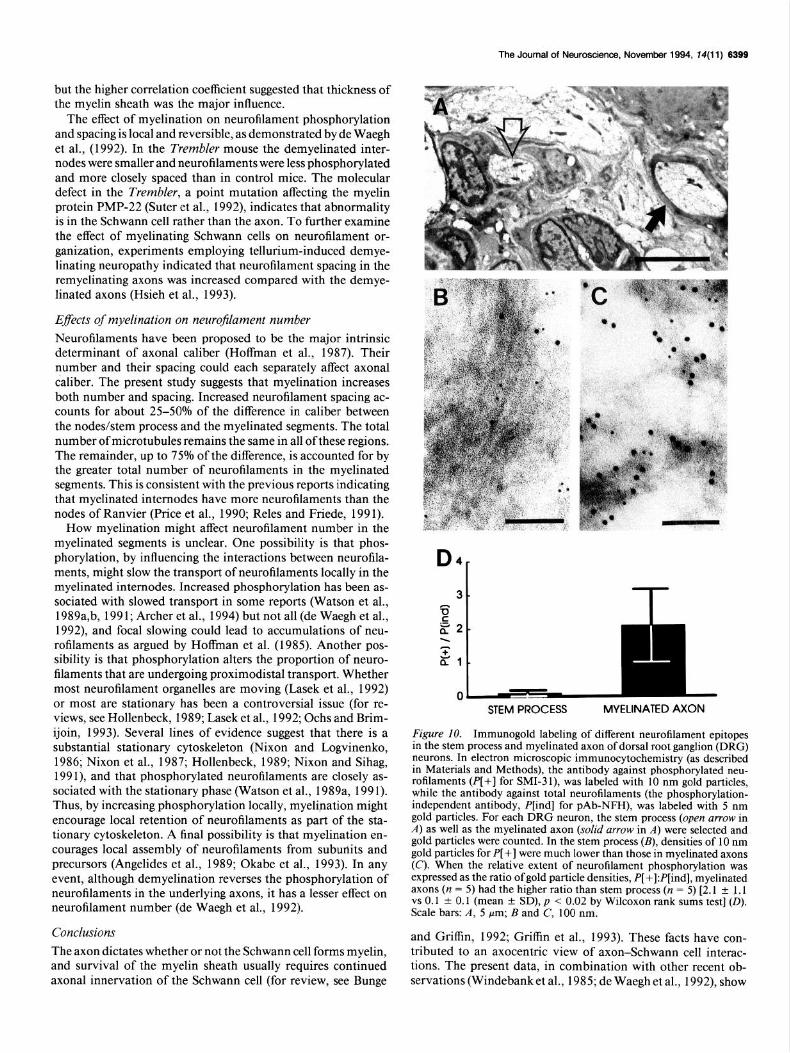
The Journal of Neuroscience, November 1994, f4(11) 8399
but the higher correlation coefficient suggested that thickness of the myelin sheath was the major influence.
The effect of myelination on neurofilament phosphorylation and spacing is local and reversible, as demonstrated by de Waegh et al., (1992). In the Trembler mouse the demyelinated inter- nodes were smaller and neurofilaments were less phosphorylated and more closely spaced than in control mice. The molecular defect in the Trembler, a point mutation affecting the myelin protein PMP-22 (Suter et al., 1992), indicates that abnormality is in the Schwann cell rather than the axon. To further examine the effect of myelinating Schwann cells on neurofilament or- ganization, experiments employing tellurium-induced demye- linating neuropathy indicated that neurofilament spacing in the remyelinating axons was increased compared with the demye- linated axons (Hsieh et al., 1993).
Effects of myelination on neurojilament number Neurofilaments have been proposed to be the major intrinsic determinant of axonal caliber (Hoffman et al., 1987). Their number and their spacing could each separately affect axonal caliber. The present study suggests that myelination increases both number and spacing. Increased neurofilament spacing ac- counts for about 25-50% of the difference in caliber between the nodes/stem process and the myelinated segments. The total number of microtubules remains the same in all of these regions. The remainder, up to 75% of the difference, is accounted for by the greater total number of neurofilaments in the myelinated segments. This is consistent with the previous reports indicating that myelinated internodes have more neurofilaments than the nodes of Ranvier (Price et al., 1990; Reles and Friede, 199 1).
How myelination might affect neurofilament number in the myelinated segments is unclear. One possibility is that phos- phorylation, by influencing the interactions between neurofila- ments, might slow the transport of neurofilaments locally in the myelinated internodes. Increased phosphorylation has been as- sociated with slowed transport in some reports (Watson et al., 1989a,b, 1991; Archer et al., 1994) but not all (de Waegh et al., 1992), and focal slowing could lead to accumulations of neu- rofilaments as argued by Hoffman et al. (1985). Another pos- sibility is that phosphorylation alters the proportion of neuro- filaments that are undergoing proximodistal transport. Whether most neurofilament organelles are moving (Lasek et al., 1992) or most are stationary has been a controversial issue (for re- views, see Hollenbeck, 1989; Lasek et al., 1992; Ochs and Brim- ijoin, 1993). Several lines of evidence suggest that there is a substantial stationary cytoskeleton (Nixon and Logvinenko, 1986; Nixon et al., 1987; Hollenbeck, 1989; Nixon and Sihag, 1991), and that phosphorylated neurofilaments are closely as- sociated with the stationary phase (Watson et al., 1989a, 1991). Thus, by increasing phosphorylation locally, myelination might encourage local retention of neurofilaments as part of the sta- tionary cytoskeleton. A final possibility is that myelination en- courages local assembly of neurofilaments from subunits and precursors (Angelides et al., 1989; Okabe et al., 1993). In any event, although demyelination reverses the phosphorylation of neurofilaments in the underlying axons, it has a lesser effect on neurofilament number (de Waegh et al., 1992).
Conclusions The axon dictates whether or not the Schwann cell forms myelin, and survival of the myelin sheath usually requires continued axonal innervation of the Schwann cell (for review, see Bunge
D4-
STEM PROCESS MYELINATED AXON
Figure IO. Immunogold labeling of different neurofilament epitopes in the stem process and myelinated axon of dorsal root ganglion (DRG) neurons. In electron microscopic immunocytochemistrv (as described in Materials and Methods), the antibody against phosphorylated neu- rofilaments (PI+1 for SMI-3 1). was labeled with 10 nm eold oarticles. while the antibody against total neurofilaments (the phosphorylation- independent antibody, P[ind] for pAb-NFII), was labeled with 5 nm gold particles. For each DRG neuron, the stem process (open arrow in A) as well as the myelinated axon (solid arrow in A) were selected and gold particles were counted. In the stem process (B), densities of 10 nm gold particles for P[ +] were much lower than those in myelinated axons (C). When the relative extent of neurofilament nhosuhorvlation was expressed as the ratio of gold particle densities, P[ ;]:P[ind],myelinated axons (n = 5) had the higher ratio than stem process (n = 5) [2.1 + 1.1 vs 0.1 * 0.1 (mean t SD), p < 0.02 by Wilcoxon rank sums test] (0). Scale bars: A, 5 Frn; B and C, 100 nm.
and Griffin, 1992; Griffin et al., 1993). These facts have con- tributed to an axocentric view of axon-Schwann cell interac- tions. The present data, in combination with other recent ob- servations (Windebank et al., 1985; de Waegh et al., 1992), show

6400 Hsieh et al. - Myelination Modulates Neurofilament Organization and Phosphorylation
that the local axonal phenotype is profoundly modified by the ensheathing Schwann cell, necessitating a reappraisal of the rel- ative roles of the axon and the Schwann cell in their interactions. Understanding the mechanism of this Schwann cell-axon in- teraction has implications for developmental neurobiology and for demyelinating diseases.
References Aguayo AJ, Attiwell M, Trecarten J, Perkins S, Bray GM (1977) Ab-
normal myelination in transplanted Trembler mouse Schwann cells. Nature 265:73-75.
Angelides KJ, Smith KE, Takeda M (1989) Assembly and exchange of intermediate filament proteins of neurons: neurofilaments are dy- namic structures. J Cell Biol 108:1495-1506.
Archer DA, Watson DF, Griffin JW (1994) Phosphorylation-depen- dent immunoreactivity of neurofilaments and the rate of slow axonal transport in the central and peripheral axons of the rat dorsal root ganglion, J Neurochem, in press.
Berthold C-H (1978) Morphology of normal peripheral axons. In: Physiology and pathobiology of axons (Waxman SG, ed), pp 3-63. New York: Raven.
Bunge RP, Griffin JW (1992) The cell of Schwann. In: Diseases of the nervous system, clinical neurobiology (Asbury AK, McKhann GM, McDonald WI, eds), pp 87-100. Philadelphia: Saunders.
Carden MJ, Schlaepfer WW, Lee VM-Y (1985) The structure, bio- chemical properties, and immunogenicity of neurofilament peripheral regions are determined by phosphorylation state. J Biol Chem 260: 9805-98 17.
Carden MJ, Trojanowski JQ, Schlaepfer WW, Lee VM (1987) Two- stage expression of neurofilament polypeptides during rat neuroge- nesis with early establishment of adult phosphorylation patterns. J Neurosci 7:3489-3504.
de Waegh S, Brady ST (1990) Altered slow transport and regeneration in a myelin-deficient mutant mouse: the Trembler as an in vivo model for Schwann-axon interactions. J Neurosci 10: 1855-1865.
de Waegh SM, Brady ST (199 1) Local control of axonal properties by Schwann cells: neurofilaments and axonal transport in homologous and heterologous nerve grafts. J Neurosci Res 30:201-212.
de Waegh SM, Lee VM, Brady ST (1992) Local modulation of neu- rofilament phosphorylation, axonal caliber, and slow axonal transport by myelinating Schwann cells. Cell 68:45 l-463.
Dyck PJ, Lais AC, Giannini C, Engelstad JK (1990) Structural alter- ations of nerve during cuff compression. Proc Nat1 Acad Sci USA 87: 9828-9832.
Friede RL, Samorajski T (1970) Axon caliber related to neurofilaments and microtubules in sciatic nerve fibers of rats and mice. Anat Ret 167:379-388.
Geisler N, Kaufmann E, Fischer S, Plessmann U, Weber K (1983) Neurofilament architecture combines structural principles of inter- mediate filaments with carboxy-terminal extensions increasing in size between triulet oroteins. EMBO J 2: 1295-1302.
Griffin JW, Gdd G;, Trapp BD (1993) Interactions between axons and Schwann cells. In: Peripheral neuropathy (Dyck PJ, Thomas PK, Griffin JW, Low PA, Poduslo JF, eds), pp 317-330. Philadelphia: Saunders.
Hirokawa N, Glicksman MA, Willard MB (1984) Organization of mammalian neurofilament polypeptides within the neuronal cyto- skeleton. J Cell Biol 98: 1523-1536.
Hisanaga S, Hirokawa N (1988) Structure of peripheral domains of neurofilaments revealed by low angle rotary shadowing. J Mol Biol 202:297-305.
Hoffman PN, Lasek RJ (1975) The slow component of axonal trans- port: identification of major structural polypeptides of axon and their eeneralitv amone mammalian neurons. J Cell Biol 66:351-366.
H&man’ PN, Grit%n JW, Price DL (1984) Control of axonal caliber by neurofilament transport. J Cell Biol 99:705-714.
Hoffman PN, Griffin JW, Gold BG, Price DL (1985) Slowing of neu- rofilament transoort and the radial growth of developing nerve fibers. J Neurosci 5:2920-2929.
Hoffman PN. Cleveland DW. Griffin JW. Landes PW, Cowan NJ, Price DL (1987) Neurofilament gene expression: a major determinant of axonal caliber. Proc Nat1 Acad Sci USA 84:3472-3476.
Hollenbeck PJ (1989) The transport and assembly of axonal cyto- skeleton. J Cell Biol 108:223-227.
Hsieh S-T, Crawford TO, Bouldin TW, Griffin JW (1993) Influence of demyelination and remyelination on axonal organization. Brain Path01 3:307.
Hsieh S-T, Crawford TO, Griffin JW (1994) Neurofilament distri- bution and organization in the myelinated axons of the peripheral nervous system. Brain Res, in press.
Julien J-P, Mushynski WE (1983) The distribution of phosphorylation sites among identified proteolytic fragments of mammalian neuroti- laments. J Biol Chem 258:40194025.
Julien J-P, Meyer D, Flavell D, Hurst J, Grosveld F (1986) Cloning and developmental expression of the murine neurofilament gene fam- ily. Mol Brain Res 1:243-250.
Julien J-P, Cote F, Beaudet L, Sidky M, Flavell D, Grosveld F, Mu- shynski W (1988) Sequence and structure of the mouse gene coding for the largest neurofilament subunit. Gene 68:307-3 14.
Laemmli UK (1970) Cleavage of structural proteins during the assem- bly of the head of bacteriophage T4. Nature 227:680-685.
Lasek RJ (1988) Studying the intrinsic determinants of neuronal form and function. In: Intrinsic determinants of neuronal form and function (Lasek RJ, Black MM, eds), pp l-60. New York: Liss.
Lasek RJ, Paggi P, Katz MJ (1992) Slow axonal transport mechanisms move neurofilaments relentlessly in mouse optic axons. .I Cell Biol 117:607-616.
Lee VM-Y, Carden MJ, Trojanowski JQ (1986) Novel monoclonal antibodies provide evidence for the in situ existence of a nonphos- phorylated form of the largest neurofilament subunit. J Neurosci 6:850- 858.-
Lee VM-Y, Carden MJ, Schlaepfer WW, Trojanowski JQ (1987) Monoclonal antibodies distinguish several differentiallv phosphorv- lated states of the two largest rat neurofilament subuniis (NFIH and NF-M) and demonstrate their existence in the normal nervous system of adult rats. J Neurosci 7:3474-3488.
Lee VM-Y, Otvos L Jr, Carden MJ, Hollosi M, Dietzschold B, Lazzarini RA (1988) Identification of the major multiphosphorylation site in mammalian nemofilaments. Proc Nat1 Acad Sci USA 85: 1998-2002.
Lees JF, Shneidman PS, Skuntz SF, Carden MJ, Lazzarini RA (1988) The structure and organization of the human heavy neurofilament subunit (NF-H) and the eene encoding it. EMBO J 7: 1947-l 955.
Liem RKH, Yen’S-H, Salimon GD, Shelanski ML (1978) Interme- diate filaments in nervous tissues. J Cell Biol 79:637-645.
Lopata MA, Cleveland DW (1987) In vivo microtubules are copol- ymers of available p tubulin isotypes: localization of each of six ver- tebrate fl tubulin isotypes using polyclonal antibodies elicited by syn- thetic peptide antigens. J Cell Biol-105: 1707-1720.
Mata M. Kuoina N. Fink DJ (1992) Phosohorvlation-dependent neu- rofilament-epitopes are reduced at the node of Ranvier. J Neurocytol 21:199-210.
Nixon RA, Logvinenko KB (1986) Multiple fates of newly synthesized neurofilament proteins: evidence for a stationary neurofilament net- work distributed nonuniformly along axons of retinal ganglion cell neurons. J Cell Biol 102:647-659.
Nixon RA, Sihag RK (1991) Neurofilament phosphorylation: a new look at regulation and function. Trends Neurosci 14:501-506.
Nixon RA, Lewis SE, Marotta CA (1987) Posttranslational modifi- cation of neurofilament proteins by phosphate during axoplasmic transport in retinal ganglion cell neurons. J Neurosci 7: 1145-l 158.
Oblinger MM (1987) Characterization of posttranslational processing of the mammalian high-molecular-weight neurofilament protein in vivo. J Neurosci 7:2510-2521.
Ochs S, Brimijoin WB (1993) Axonal transport. In: Peripheral neu- ropathy (Dyck PJ, Thomas PK, Griffin JW, Low PA, Poduslo JF, eds), pp 33 l-360. Philadelphia: Saunders.
Okabe S, Miyasaka H, Hirokawa N (1993) Dynamics of the neuronal intermediate filaments. J Cell Biol 121:375-386.
Parhad IM, Clark AW, Griffin JW (1987) Effect of changes in neu- rofilament content on caliber of small axons: the @,p-iminodipro- pionitrile model. J Neurosci 7:2256-2263.
Price RL, Lasek RJ, Katz MJ (1990) Internal axonal cytoarchitecture is shaped locally by external compressive forces. Brain Res 530:205- 214.
Reles A, Friede RL (199 1) Axonal cytoskeleton at the nodes of Ran- vier. J Neurocytol 20:450-458.
Schlaepfer WW (1987) Neurofilaments: structure, metabolism and implications in disease. J Neuropathol Exp Neurol 46: 117-129.
Spencer PS, Raine CS, Wisniewski H (1973) Axonal diameter and

The Journal of Neuroscience, November 1994, 14(11) 6401
myelin thickness-unusual relationships in dorsal root ganglia. Anat Ret 176:225-244.
Stemberger LA, Stemberger NH (1983) Monoclonal antibodies dis- tinguish phosphorylated and nonphosphorylated forms of neurofila- ments in situ. Proc Nat1 Acad Sci USA 80:6 126-6 130.
Suter U, Welcher AA, Ozcelik T, Snipes GJ, Kosaras B, Francke U, Billings-Gagliardi S, Sidman RL, Shooter EM ( 1992) Trembler mouse carries a point mutation in a myelin gene. Nature 356:241-244.
Thomas PK, Ochoa J (1984) Microscopic anatomy of peripheral nerve fibers. In: Peripheral neuropathy (Dyck PJ, Thomas PK, Lambert EH, Bunge R, eds), pp 39-96. Philadelphia: Saunders.
Trapp BD, Itoyama Y, Stemberger NH, Quarles RH, Webster HdeF ( 198 1) Immunocytochemical localization of P, protein in Golgi com- plex membranes and myelin of developing rat Schwann cells. J Cell Biol 90: l-6.
Trapp BD, Andrews SB, Wong A, O’Connell M, Griffin JW (1989) Co-localization of myelin-associated glycoprotein and the microfi- lament components, F-actin and spectrin, in Schwann cells of my- elinated nerve fibers. J Neurocytol 18:47-60.
Watson DF, Griffin JW, Fittro KP, Hoffman PN (1989a) Phosphor- ylation-dependent immunoreactivity of neurofilaments increases dur- ing axonal maturation and @,/3’-iminodipropionitrile intoxication. J Neurochem 53:1818-1829.
Watson DF, Hoffman PN, Fittro KP, Griffin JW (1989b) Neurofila- ment and tubulin transport slows along the course of mature motor axons. Brain Res 4771225-232.
Watson DF, Fittro KP, Hoffman PN, Griffin JW (1991) Phosphory- lation-related immunoreactivity and the rate of transport of neuro- filaments in chronic 2,5-hexanedione intoxication. Brain Res 539: 103-109.
Waxman SG (1980) Determinants of conduction velocity in myelin- ated nerve fibers. Muscle Nerve 3: 141-150.
Willard M, Simon C (1983) Modulations of neurofilament axonal transport during the development of rabbit retinal ganglion cells. Cell 35:551-559.
Windebank AJ, Wood P, Bunge RP, Dyck PJ (1985) Myelination determines the caliber of dorsal root ganglion neurons in culture. J Neurosci 5: 1563-l 569.





























