Embed Size (px)
Citation preview

Regulation of protein function through expression of chimaeric proteins
Didier Picard
Un ive rs i t y of Geneva , Geneva , Swi tzer land
The hormone-binding domain of steroid receptors can be used to subject heterologous protein functions to hormonal control in cis. Recent studies have established that these regulatory domains constitute a set of up to five different reversible molecular switches for the post-translational regulation of a wide variety of cytoplasmic and nuclear proteins, including kinases. This approach
has been applied both in vertebrate cells and in budding yeast.
Current Opinion in Biotechnology 1994, 5:511-515
Introduction
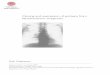
It is often necessary to regulate the expression and/or the activity of a protein of interest. As yet, very few approaches exist that allow a specific protein to be regu- lated post-translationally. The need for such techniques is obvious if one considers that transcriptionally inducible systems are hampered by drawbacks, such as the lag phase between induction and optimal protein activity and the difficulty of activating genes in isolation from other cel- lular genes. Table 1 compares the features of different approaches that allow the regulation of protein activities at the protein level. The emphasis of this article is on the use of the hormone-binding domain (HBD) of steroid receptors as a regulatory system (see Fig. 1). I first com- pare this system with the other approaches for regulating protein function (see Table 1), and then discuss progress achieved during the past eighteen months.
(a) [ ProteinX I Active
(b)
Protein X [ H~D Inactive
H O ~ ~ > hsp90 complex
[ ProteinX Active
©1994 Current Opinion in Biotechno[ogy
The hallmarks of steroid-binding domains as regulatory cassettes
It was discovered several years ago that the HBD of the glucocorticoid receptor (GI<) can be used as a molec- ular switch to regulate heterologous proteins in cis [1]. A constitutively expressed cytoplasmic or nuclear fusion protein is maintained in an inactive state in the absence of hormone and can be rapidly activated (derepressed) by the addition of cognate steroid hormone. It was subsequently shown that the HBDs of other steroid receptors can also function as autonomous regulatory cassettes [2,3 °°] and that a large variety of different pro- tein activities can be subjected to hormonal control. The
Fig. 1. The use of hormone-binding domains (HBDs) of steroid re- ceptors to regulate heterologous proteins. (a) Activity of a hetero- Iogous protein (Protein X) is under no regulatory control. (b) Once fused to a HBD, however, Protein X is inactivated. Inactivation me- diated by the hsp90 complex is relieved upon hormone (H) binding.
list o f regulable proteins now includes various oncopro- teins, transcription factors, an P,.NA-binding protein, as well as serine/threonine and tyrosine kinases (reviewed in [4°,5°]).
How does this system function? The unliganded HBDs of all vertebrate steroid receptors are known to assemble into a protein complex containing heat-shock protein 90 (hsp90). My colleagues and I have previously speculated that this complex, which is released upon hormone
Abbreviations AR~androgen receptor; CR---conserved region; EcR--ecdysone receptor; EGF--epidermal growth factor; ER--estrogen receptor;
GR--glucocorticoid receptor; HBD--hormone-binding domain; hsp90~heat-shock protein 90; MR--mineralocorticoid receptor; PR--progesterone receptor; TR--thyroid receptor.
© Current Biology Ltd ISSN 0958-1669 511

512 Expression systems
Table 1. Approaches for regulating protein functions at the protein level.
Features Approaches
Fusion to Chimaeric Temperature- Dimerization Microinjection steroid-binding transmembrane sensitive mediated by FK506
domain receptor mutants analogue
Side effects on cells No No Yes ? No (heat-shock)
Regulation of a wide Yes No No variety of heterologous (rarely available) ~roteins
No Yes
ntermediate levels of Yes Yes No Probable Yes induction
Reversibility Yes Yes No Probable No (only 'on' to 'off')
~s ~s NA No NA Choice of several regulatory domains
Species range Yeast to animals Animals Universal ? Tissue-culture cells
Applicable to whole Probable Yes Not in warm-blooded Probable organisms animals
NA
Abbreviations: NA, not applicable; ?, not known.
binding (see Fig. 1), may effect inactivation of protein function in the heterologous moiety by steric hindrance [1,4°,5°,6]. Although this remains unproven, it is clear that the HBD alone provides a sufficient regulatory ma- chinery to control the activity of a fusion protein [7]. Thus, the hallmarks of this system are the wide range of regulable functions, the choice of several regulatory domains, the possibility for fine-tuning using subsat- urating concentrations of hormone, the rapid kinetics, the reversibility of control, and the applicability to many different organisms.
Other approaches to the post-translational regulation of proteins
In contrast to the flexibility of HBD systems, other tech- niques tend to be more limited. Microinjection and the use of temperature-sensitive mutants are very powerful approaches, but they are often either not applicable or not available (see Table 1). It should, however, be emphasized that temperature-sensitive mutants are un- matched as systems for regulating protein function in all cellular compartments and for almost any species.
A novel approach has recently been proposed by Schreiber, Crabtree and collaborators [8]. They have developed a divalent analogue of the immunosuppres- sant drug FK506, which can induce the dimeriza-
tion of fusion proteins containing FK506-binding im- munophilin domains. This ingenious compound can be used as an inducer for proteins that employ dimeriza- tion/oligomerization as a key step in activation. Even though this technique has been received with consid- erable enthusiasm--it promises to be applicable even in whole animals--the general applicabihty of the ap- proach remains to be established. In particular, one lim- itation may be that the dimerization of a fusion protein, and thus its activation, must be inhibited (e.g. by an al- ternative conformation or by a regulatory protein) before addition of the drug. Moreover, drug-mediated dimer- ization has to be dominant over the normal control mechanism.
My colleagues and I had already proposed that steroid binding domains might also be used to promote dimer- ization of a fusion protein because HBDs are themselves ligand-dependent dimerization domains. We found that the oncogenicaUy inactive normal c-Abl tyrosine kinase can be strongly activated in a hormone-dependent fash- ion by fusion to the HBD of the estrogen receptor (ELK) [9°°]. This implies that the EIK HBD-mediated dimer- ization can override normal negative control and that dimerization results in activation of wild-type Abl.
The construction of chimaeric transmembrane receptors has been applied very successfully to study signal trans- duction by candidate receptors with unknown ligands [10-15]. In these cases, a well characterized extracellu- lar domain, for example that of the epidermal growth

Regulation of protein function through expression of chimaeric proteins Picard 513
factor (EGF) or insulin receptors, is substituted for the extracellular domain of a novel receptor. Signalling by the intracellular domain(s) of the chimaeric receptor can then be induced by the cognate heterologous ligand. By definition, this approach is limited to a certain class of transmembrane receptor. Within these constraints, its versatility is further underlined by a recent report on an EGF receptor-CD45 chimaera by Desai et al. [16]. They have shown that the activity of the intracellu- lar CD45 domain, a protein tyrosine phosphatase, is switched off by the binding of EGF to the extracellu- lar EGF receptor domain.
Steroid-binding domains as molecular switches: recent progress
During the past year, notable progress has been made in the following areas: first, the HBDs of four out of five vertebrate steroid receptors are now known to function in fusion proteins; second, the-range of regulable pro- teins has been extended to kinases; and third, regulation by HBDs has been demonstrated to function in budding yeast.
A choice of several regulatory domains My colleagues and I have recently shown that the HBD of the mineralocorticoid receptor (MR) can also subject a heterologous protein to hormonal control [3°°]. A fu- sion protein consisting, for example, of the transcription factor GCN4 and the M R HBD can be turned on at very low concentrations of aldosterone in mammalian tissue culture cells, thereby avoiding cross-activation of endogenous GR. This feature is highly useful as the GR is ubiquitously expressed, whereas expression of ER and MR is restricted to certain tissues (and tissue-culture cells). Although the HBD of the progesterone receptor (PR) has not yet been tested to my knowledge, positive data has now been obtained for the HBD of the only other remaining vertebrate steroid receptor, the andro- gen receptor (AR). The AR HBD has been used to reg- ulate the yeast FLP recombinase both in mammalian cells and in yeast (M Nichols, F Stewart, personal communi- cation). Because the PR is also known to form ligand- reversible complexes with hsp90, it can be assumed that the PR HBD behaves like the other four HBDs. Thus, five different regulatory domains are available for regulat- ing heterologous protein activities. This implies not only that cross-activation of endogenous steroid receptors can be avoided by choosing the appropriate HBD, but also that up to five different fusion proteins can be regulated independently in the same cell.
An open issue is whether the ligand-binding domains of other ligand-regulated transcription factors may also be exploited. Thus far, the HBDs of two other members of the nuclear receptor family have been tested. The HBDs
of the thyroid receptor (TR) and the ecdysone receptor (EcR) have been demonstrated to regulate MyoD [17 °] and certain chimaeric transactivators [18], respectively. Because the HBDs of receptors such as the T R and the EcR do not form stable complexes with hsp90, it re- mains unclear whether regulation by these two domains is more generally applicable. Several other laboratories have indeed failed to get ligand-reversible inactivation of heterologous activities with the EcR HBD (M Worek, D Picard, unpublished data; M Busslinger, personal com- munication; F Stewart, personal communication). The dioxin receptor is another candidate that needs its utility as a regulatory element confirmed. Although it belongs to another family of proteins, it forms ligand-reversible complexes with hsp90 and is able to subject a constitu- tively active truncated GR to dioxin control [19°]. Even so, the wild-type receptor is widely expressed, and re- lease ofhsp90 may require additional protein interactions [20].
The range of regulable proteins The versatility of the HBD technique has been con- firmed in the past year by its application to various kinases. Tyrosine kinases, such as c-Abl [9°°], Src and erbB1 (JM Bishop, personal communication), and the serine/threonine kinases Raft [21] and its yeast homo- logue, STE11 (JF Louvion, D Picard, unpublished data), become conditional upon fusion to a HBD. The list of regulable proteins (for comprehensive list, see [5°]), as extended [3"°,9°',17°,21,22°-24°,25,26"°,27 °] and ap- plied [28] during the past year, now includes proteins that bind DNA and RNA as well as proteins that un- dergo a variety of different protein-protein interactions.
Clearly, certain functions are not, or cannot be, regulated by HBDs. Those that remain constitutively active upon fusion to a HBD include several enzymatic functions that may not be affected sterically by the hsp90 complex (D Picard, unpublished data). A particularly well charac- terized example is a fusion protein of the dihydrofolate reductase and the GR HBD [29°]. The function of this fusion protein becomes even slightly worse in the pres- ence of hormone. Moreover, many examples of fusion proteins have been found where only a subset of func- tions becomes subjected to hormonal control (for further discussion, see [4°,5°]). E1A and Myc fusion proteins are particularly well studied in this respect (see below). The data are compatible with the notion that the spacial rela- tionship between the regulatory machinery and the reg- ulated function is important.
An E1A chimaera, constructed by insertion of a HBD into the carboxy-terminal domain of E1A, has a fully hormone-dependent transcriptional activation function [1], which requires the presence of conserved region (CR)3. In contrast, its nuclear localization [1] and its transformation function (D Picard, unpublished data), which depends on the more amino-terminal CR1 and CR2, remain constitutively active. Moving the HBD

514 Expression systems
closer to CIL1 and CIL2 on the linear protein map yields a fusion protein that conditionally immortalizes and transforms primary rat fibroblast cells in collabora- tion with an activated ras oncogene [22°]. Interestingly, when E1A activity is turned off by removal of hormone, cells become arrested in G1 phase.
The conditional c-Myc chimaera, MycEiL, has been ex- ploited extensively in fibroblasts to study a variety of Myc-mediated phenomena: oncogenic transformation [2]; entry into, and progression through, the cell cycle [30,31,32"]; apoptosis [33]; and transcriptional induc- tion of Myc target genes [30,31,34,35]. All of these Myc functions appear to require the interaction with Max, and this interaction is hormone-dependent in MycER [32"]. In contrast, transcriptional repression of Myc target genes, such as the gene for cyclin D1, es- capes hormonal control in MycEIL. Interestingly, inter- action with Max is dispensable for this function, which maps to a novel amino-terminal domain [32"].
A novel inducible system for Saccharomyces cerevisiae My colleagues and I have shown that the approach is also applicable to yeast, suggesting that the regulatory ma- chinery has been conserved in evolution. HBDs were used to control the activity o f two proteins: the chi- maeric transcriptional activator GAL4-VP16 [26"] and the serine/threonine kinase STE11 OF Louvion, D Pi- card, unpublished data). The former fusion protein turns out to be a very valuable regulatory molecule. It allows the induction of proteins expressed under the control of a GAL4-activated galactose-inducible promoter by a gra- tuitous steroid instead of a shift in carbon source, which is the conventional method.
Conclusions
The HBDs of steroid receptors are now established as versatile and powerful regulatory elements. Given the many examples of proteins and cell types where HBD fusions have been successful, these regulatory switches should be adopted as a first choice whenever post-trans- lational control of cytoplasmic or nuclear protein ac- tivity is required. Nevertheless, further improvements are anticipated in the future. In the short term, the transcriptional activation function o f the HBD, which can modify the properties of weak transcription fac- tors [23"], can be, avoided through the use of specific HBD mutants [36-38]. In the long term, an artificial HBD-ligand couple should be developed, providing a completely heterologous regulatory system both for ver- tebrates and even for gene therapy in humans. The syn- thetic ligand would be capable o f exclusively turning on a particular mutant HBD without any effect on endoge- nous wild-type steroid receptors.
Acknowledgements
1 am grateful to Jean-Francois Louvion for critical comments on the manuscript and to many colleagues for communicating unpublished results. Our work is supported by the Canton de Gen~ve and the Swiss National Science Foundation.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as: • of special interest • , of outstanding interest
1. Picard D, Salser SJ, Yamamoto KR: A Movable and Regulable Inactivation Function within the Steroid Binding Domain of the Glucocorlicoid Receptor. Cell 1988, 54:1073-1080.
2. Eilers M, Picard D, Yamamoto KR, Bishop JM: Chimeras of the MYC Oncoprotein and Steroid Receptors Cause Hormone- Dependent Transformation of Cells. Nature 1989, 340:66-68.
3. Fankhauser CP, Briand P-A, Picard D: The Hormone Binding • , Domain of the Mineralocorticoid Receptor Can Regulate Het-
erologous Activities in Cis. Biochem Biophys Res Commun 1994, 200:] 95-201.
Using the transcription factor GCN4 as a marker protein, the utility of MR HBD as a regulator of protein activity is demonstrated. The MR HBD can be activated by very low concentrations of aldosterone, suggesting that it may be employed in many cell types.
4. Picard D: Steroid-Binding Domains for Regulating the Func- • tions of Heterologous Proteins in Cis. Trends Cell Biol 1993,
3:278-280. A recent review with a discussion of the model for the mechanism of regulation by HBDs.
S. Mattioni T, Louvion J-F, Picard D: Regulation of Protein Activ- • ities by Fusion to Steroid Binding Domains. Methods Cell Biol
1994, 43:335-352. A recent comprehensive review that discusses the advantages and limi- tations of the HBD fusion protein approach. Contains explicit protocols.
6. Yamamoto KR, Godowski PJ, Picard D: Ligand Regulated Non- specific Inactivation of Receptor Function: A Versatile Mecha- nism for Signal Transduction. Cold Spring Harbor Symp Quant Biol 1988, 53:803-811.
7. Scherrer LC, Picard D, Massa E, Harmon JM, Simons SS Jr, Yamamoto KR, Pratt WB: Evidence that the Hormone Bind- ing Domain of Steroid Receptors Confers Hormonal Control on Chimeric Proteins by Determining their Hormone-Regu- lated Binding to Heat-Shock Protein 90. Biochemistry 1993, 32:5381-$386.
8. Spencer DM, Wandless TJ, Schreiber SL, Crabtree GR: Control- ling Signal Transduction with Synthetic Ligands. Science 1993, 262:1019-1024.
9. Jackson P, Baltimore D, Picard D: Hormone-Conditional Trans- • , formation by Fusion Proteins of c-Abl and its Transforming
Variants. EMBO J 1993, 12:2809-2819. First paper showing that fusion of a kinase to a HBD can be used as a means of controlling its activity. Surprisingly, the HBD not only strongly inhibits an active Abl tyrosine kinase in a hormone-reversible fashion, but also strongly activates the normally silent wild-type Abl (see text for further comments).
10. Ellis L, Morgan DO, Jong SM, Wang LH, Roth RA, Rutter WJ: Heterologous Transmembrane Signaling by a Human Insulin Receptor-v-ros Hybrid in Chinese Hamster Ovary Cells. Proc Natl Acad Sci USA 1987, 84:5101-5105.
11. Lee J, Dull TJ, Lax I, Schlessinger J, UIIrich A: HER2 Cytoplas- mic Domain Generates Normal Mitogenic and Transforming Signals in a Chimeric Receptor. EMBO J 1989, 8:167-173.
12. Lev S, Yarden Y, Givol D: Receptor Functions and tigand- Dependent Transforming Potential of a Chimeric kit Proto- Oncogene. Mol Cell Biol 1990, 10:6064--6068.

Regulation of protein function through expression of chimaeric proteins Picard 515
13. Lehvaslaiho H, Lehtola L, Sistonen L, Alitalo K: A Chimeric EGF-R-neu Proto-Oncogene Allows EGF to Regulate neu Tyro- sine Kinase and Cell Transformation. EMBO ] 1989, 8:159-166.
14. Riedel H, Dull TJ, Honegger AM, Schlessinger J, UIIrich A: Cytoplasmic Domains Determine Signal Specificity, Cellular Routing Characteristics and Influence Ligand Binding of Epi- dermal Growth Factor and Insulin Receptors. EMBO J 1989, 8:2943-2954.
15. Seedorf K, Felder S, Millauer 8, Schlessinger J, UIIrich A: Anal- ysis of Platelet-Derived Growth Factor Receptor Domain Func- tion Using a Novel Chimeric Receptor Approach. J Biol Chem 1991, 266:12424-12431.
16. Desai DM, Sap J, Schlessinger I, Weiss A: Ligand-Mediated Negative Regulation of a Chimeric Transmembrane Receptor Tyrosine Phosphatase. Cell 1993, 73:541-554.
17. Hollenberg SM, Cheng PF, Weintraub H: Use of a Conditional • MyoD Transcription Factor in Studies of MyoD Trans-Activa-
tion and Muscle Determination. Proc Nat/Acad 5ci USA 1993, 90:8028-8032.
Exploits the unique advantages of the HBD fusion protein approach. The reversible short-term direct effects (in the absence of protein synthesis) and the long-term effects of MyoD induction are distinguished using a conditional HBD-MyoD fusion.
18. Christopherson KS, Mark MR, Bajaj V, Godowski P J: Ecdysteroid-Dependent Regulation of Genes in Mammalian Cells by a Drosophila Ecdysone Receptor and Chimeric Trans- activators. Proc Nat/ Acad Sci USA 1992, 89:6314-6318.
19. Whitelaw ML, GOttlicher M, Gustafsson J-,~, Poellinger L: Def- • inition of a Novel Ligand Binding Domain of a Nuclear bHLH
Receptor: Co-Locallzatlon of Ligand and HSP90 Binding Activ- ities within the Regulable Inactivation Domain of the Dioxin Receptor. EMBO J 1993, 12:4169-4179.
This study suggests that regulatory domains from factors belonging to other protein families, which form ligand-reversible hspg0 complexes, may also be exploited to regulate heterologous proteins.
20. McGuire J, Whitelaw ML, Pongratz I, Gustafsson J-,~, Poellinger L: A Cellular Factor Stimulates Ligand-Dependent Release of hspg0 from the Basic Helix-Loop-Helix Dioxin Receptor. Mol Cell Biol 1994, 14:2438-2446.
21. Samuels ML, Weber MJ, Bishop JM, McMahon M: Condi- tional Transformation of Cells and Rapid Activation of the Mitogen-Activated Protein Kinase Cascade by an Estradiol- Dependent Human Raf-1 Protein Kinase. Mol Cell Biol 1993, 13:6241-6252.
22. Spitkovsky D, Steiner P, Lukas J, Lees E, Pagano M, Schulze • A, Joswig S, Picard D, Tommasino M, Eilers M, Jansen-D~irr
P: Modulation of Cyclin Gene Expression by Adenovirus EIA in a Cell Line with EIA-Dependent Conditional Proliferation. J Virol 1994, 68:2206-2214.
This paper illustrates the advantages of a regulable post-translational system over other ways to introduce a protein at a given moment. It describes a primary cell line that is conditionally 'immortalized' by an E1A fusion protein.
23. Schuermann M, Hennig G, MOiler R: Transcriptional Activa- • tion and Transformation by Chimaeric Fos-Estrogen Receptor
Proteins: Altered Properties as a Consequence of Gene Fusion. Oncogene 1993, 8:2781-2790.
A systematic analysis confirming that wild-type HBDs may qualitatively alter the properties of fusion proteins. This paper makes the important point that additional functions, such as the transactivation activity, of the HBD must be borne in mind. See text for solutions to this problem.
24. Briegel K, Lim K-C, Planck C, Beug H, Engel JD, Zenke M: • Ectopic Expression of a Conditional GATA-2/Estrogen Recep-
tor Chimera Arrests Erythroid Differentiation in a Hormone- Dependent Manner. Genes Dev 1993, 7:1097-1109.
A good example of how fusion proteins can be used in a complex bio- logical system. The differential effects of members of the GATA family of transcription factors on the differentiation of chicken erythroid cells are investigated.
25. Braselmann S, Graninger P, Busslinger M: A Selective Transcrip- tional Induction System for Mammalian Cells Based on Gal4- Estrogen Receptor Fusion Proteins. Proc Nat/ Acad Sci USA 1993, 90:1657-1661.
26. Louvion J-F, Havaux-Copf B, Picard D: Fusion of GAL4-VP16 • . to a Steroid Binding Domain Provides a Tool for Gratuitous
Induction of Galactose-Responsive Genes in Yeast. Gene 1993, 131:129-134.
First demonstration that a HBD can regulate a protein in yeast. The paper presents a hormone-regulable GAL4 derivative that allows the induction of any pre-existing galactose-responsive construct without the standard need for a shift in carbon source.
27. Roemer K, Friedmann T: Modulation of Cell Proliferation and • Gene Expression by a p53-Estrogen Receptor Hybrid Protein.
Proc Nat/ Acad Sci USA 1993, 90:9252-9256. Interesting example of the application of the HBD fusion protein ap- proach to a multifunctional protein, in this case p53.
28. Wrighton C, Busslinger M: Direct Transcriptional Stimulation of the Ornithine Decarboxylase Gene by Fos in PC12 Cells but Not in Fibroblasts. Mol Cell Biol 1993, 13:4657-4669.
29. Israel DI, Kaufman Rl: Dexamethasone Negatively Regu- • lates the Activity of a Chimeric Dihydrofolate Reduc-
tase/Glucocorticoid Receptor Protein. Proc Natl Acad Sci USA 1993, 90:4290-4294.
The only published report, thus far, detailing a heterologous protein that fails to be inactivated upon fusion with an HBD. A detailed and careful enzymological study.
30. Eilers M, Schirm S, Bishop JM: The MYC Protein Activates Transcription of the Alpha-Prothymosin Gene. EMBO J 1991, 10:133-141.
31. Jansen-D(Jrr P, Meichle A, Steiner P, Pagano M, Finke K, Botz J, Wessbecher J, Draetta G, Eilers M: Differential Modulation of Cyclin Gene Expression by MYC. Proc Natl Acad Sci USA 1993, 90:3685-3689.
32. Philipp A, Schneider A, V~srik I, Finke K, Xiong Y, Beach D, • Alitalo K, Eilers M: Repression of Cyclin DI: A Novel Function
of MYC. Mol Cell Biol 1994, 14:4032-4043. A rare example of a study in which the actual function directly inacti- vated by the HBD is determined. In this case, the interaction of Myc with Max is shown to be affected.
33. Evan GI, Wyllie AH, Gilbert CS, Littlewood TD, Land H, Brooks M, Waters CM, Penn LZ, Hancock DC: Induction of Apoptosis in Fibroblasts by c-Myc Protein. Cell 1992, 69:119-128.
34. Gaubatz S, Meichle A, Eilers M: An E-Box Element Localized in the First Intron Mediates Regulation of the Protbymosin a Gene by c-myc. Mol Cell Biol 1994, 14:3853-3862.
35. Wagner AJ, Meyers C, Laimins LA, Hay N: c-Myc Induces the Expression and Activity of Ornithine Decarboxylase. Cell Growth Differ 1993, 4:879-883.
36. Danielian PS, White R, Lees JA, Parker MG: Identification of a Conserved Region Required for Hormone Dependent Transcriptional Activation by Steroid Hormone Receptors. EMBO J 1992, 11:1025-1033.
37. Ince BA, Zhuang Y, Wrenn CK, Shapiro DJ, Katzenellenbogen BS: Powerful Dominant Negative Mutants of the Human Es- trogen Receptor. J Biol Chem 1993, 268:14026-14032.
38. Wrenn CK, Katzenellenbogen BS: Structure-Function Analysis of the Hormone Binding Domain of the Human Estrogen Re- ceptor by Region-Specific Mutagenesis and Phenotypic Screen- ing in Yeast. J Biol Chem 1993, 268:24089-24098.
D Picard, D6partement de Biologie Cellulaire, Universit~ de Gen~ve, Sciences I!1, 30 Quai Ernest-Anserrnet, CH-1211 Gen~ve 4, Switzerland.