Embed Size (px)
DESCRIPTION
Aeromonas hydrophila
Citation preview

6
BAB II
IKAN GURAME, BAKTERI Aeromonas hydrophila, PENANDA GENETIK,
PCR, MIKROSATELIT & APLIKASINYA, DAN GEL
POLIAKRILAMIDA
A. Ikan Gurame (Osphronemus goramy Lac.)
Ikan gurame (Osphronemus goramy Lac.) merupakan salah satu ikan asli
perairan Indonesia. Menurut Jangkaru (2004) gurame mempunyai bentuk badan
agak panjang, pipih dan tertutup sisik yang berukuran besar serta terlihat kasar
dan kuat. Punggungnya tinggi dan mempunyai sirip perut dengan jari pertama
sudah berubah menjadi alat peraba. Gurame jantan yang sudah tua terdapat
tonjolan seperti cula. Mulutnya kecil dengan bibir bawah menonjol sedikit
dibandingkan bibir atas. Pada jantan bibir bawah relatif tebal.
Gurame memiliki lima buah sirip, yaitu: sirip punggung, sirip dada, sirip
perut, sirip dubur dan sirip ekor. Sirip punggung tidak begitu panjang, atau
pendek dan berada hampir di bagian belakang tubuh. Sirip dada kecil berada di
belakang tutup insang. Sirip perut juga kecil berada di bawah sirip dada. Sirip
ekor berada di belakang tubuh dengan bentuk bulat, sedangkan sirip dubur
panjang, mulai dari belakang sirip perut hingga pangkal bawah sirip ekor (Arie,
2008)
Menurut Jangkaru (2004) ujung sirip punggung dan sirip dubur dapat
mencapai pangkal ekor, ujung pangkal ekor berbentuk busur. Pada dasar sirip
dada gurame betina terdapat tanda berupa bundaran hitam. Bagian kepala gurame
muda berbentuk lancip dan akan menjadi tumpul bila sudah besar. Pada badan

7
gurame muda terdapat garis tegak atau vertikal berwarna hitam berjumlah 7-10
buah dan garis-garis tegak ini akan hilang setelah dewasa (Robert, 1992).
Badan gurame bluesafir muda pada umumnya berwarna biru kehitaman
dan bagian perut berwarna putih atau kekuningan. Warna tersebut akan berubah
menjelang dewasa, yakni pada bagian punggung berwarna kecoklatan dan pada
bagian perut berwarna keperakan atau kekuningan (Jangkaru, 2004).
Gambar 2.1. Ikan gurame varietas bluesafir (Sumber : Kusumawaty et al., 2008)
Klasifikasi gurame menurut Lacapede (1801 dalam Sitanggang dan
Sarwono, 2003) adalah sebagai berikut:
Kingdom : Animalia
Phylum : Chordata
Class : Actinopterygii
Order : Perciformes
Family : Anabantidae
Genus : Osphronemus
Species : Osphronemus gouramy (Lacepede)
Penyebaran ikan gurame, pada awalnya banyak ditemukan di Pulau
Sumatra, Jawa dan Kalimantan, tetapi karena sangat digemari masyarakat, maka

8
ikan ini menyebar ke beberapa pelosok tanah air. Bahkan sejak abad 18, ikan
gurami sudah diproduksi ke negara lain, diantaranya: Madagaskar, Mauritius,
Sycheles, Australia, Srilangka, Suriname, Guyane, Martinique dan Haiti (Robert,
1992).
Sesuai dengan sejarah perikanan Indonesia yang cukup panjang, ikan
gurami juga telah lama dikembangkan secara komersial oleh para pembudidaya,
baik yang khusus memelihara gurami atau memelihara dengan jenis ikan lainnya.
Bahkan di beberapa daerah sudah terbentuk sentra-sentra kawasan pengembangan
budidaya, sehingga apabila memerlukan benih atau konsumsi dapat dengan
mudah mendatanginya. Berikut ini merupakan kawasan pembudidaya gurami
diantaranya: Jawa Barat (yaitu di Bogor, Tasikmalaya, Ciamis dan Garut), Jawa
Tengah (Cilacap, Banyumas, Banjarnegara dan Purbalingga), Daerah Istimewa
Yogyakarta (Kulonprogo, Bantul dan Sleman), Jawa Timur (Tulungagung, Blitar
dan Lumajang) dan propinsi lainnya, yaitu Sumatra Barat, Riau, dan Kalimantan
Selatan (Arie, 2008).
Gurame sangat menyenangi perairan yang tenang, seperti rawa, situ, danau
dan perairan tenang lainnya. Menurut Sitanggang & Sarwono (2003), di sungai
yang berarus deras, gurame jarang ditemui. Kehidupan yang menyukai perairan
yang bebas arus itu terbukti, ketika gurami sangat mudah dipelihara di kolam-
kolam tergenang, Max Weber dan De Beau Fort dalam The Fishes of The
Australian Archipelago mengungkapkan bahwa gurame dapat menyesuaikan diri
pada perairan yang agak payau dan agak asin. Kegiatan ini banyak dilakukan di
Cengkareng, Kamal, dan Tegal Alur di Wilayah Jakarta Barat.

9
Djarijah dan Puspowardoyo (1992) mengungkapkan gurame umumnya
hidup dan banyak dipelihara di perairan tawar, terutama pada perairan yang
tenang dan dalam. Gurame dapat tumbuh dan berkembang pada perairan tropis
dan subtropis. Ikan ini mempunyai daya adaptasi tinggi terhadap lingkungan,
tetapi lebih cocok hidup pada ketinggian maksimal 800 m di atas permukaan laut.
Selanjutnya keduanya mengatakan, bahwa suhu ideal untuk pertumbuhan gurami
antara 24–29oC, derajat keasaman (pH) antara 6,5–8, kandungan oksigen terlarut
3–5 ppm, dan air yang tidak terlalu keruh dengan kecerahan pada pengukuran alat
secchi disk.
B. Penyakit yang Disebabkan oleh Aeromonas hydrophila
Walaupun pemeliharaan ikan gurame relatif mudah, namun masih ada
kendala yang terjadi, yaitu terserangnya ikan oleh penyakit yang disebabkan oleh
bakteri. Salah satu penyakit bakterial yang merupakan kendala dalam budidaya air
tawar adalah penyakit yang disebabkan oleh Aeromonas hydrophila. Bakteri
Aeromonas tersebut umumnya hidup di air tawar, tanaman air, dan tubuh ikan,
sehingga berpeluang besar untuk dapat menginfeksi pada saat sistem pertahanan
tubuh ikan menurun akibat stres. Aeromonas dapat menyerang semua jenis ikan
air tawar dan jenis penyakitnya disebut Motil Aeromonas Septicemia (MAS)
(Plumb, 1994).
Sinonim dari penyakit MAS berhubungan dengan gejala serangan penyakit
yang disebabkan bakteri atau racun yang ditimbulkan yaitu septicemia pada
permukaan tubuh ikan dan organ tubuh ikan lainnya. Bakteri A. hydrophila adalah
bakteri gram negatif berbentuk batang yang biasanya diisolasi dari kolam air

10
tawar. Bakteri ini adalah organisme yang banyak ditemui pada saluran pencernaan
ikan (Cipriano, 2001).
Aeromonas hydrophila dikategorikan sebagai patogen oportunis, yaitu
dapat menyebabkan penyakit bila kondisi memenuhi syarat. Pada situasi alami,
kehadiran bakteri ini dianggap normal, akan tetapi pada kondisi akuakultur yang
intensif keberadaan bakteri ini patut diperhitungkan. Penyakit akibat bakteri ini
biasanya muncul akibat dari kondisi stres pada ikan. Ahli akuakultur sepakat
bahwa ikan dapat mengalami stres apabila terkondisikan pada penanganan yang
kurang baik, kepadatan yang terlalu tinggi, transportasi dalam kondisi yang buruk,
nutrisi yang tidak memadai dan kualitas air yang buruk. Beberapa faktor kualitas
air yang dapat menyebabkan ikan rentan terserang A. hydrophila antara lain
tingginya kandungan nitrit, rendahnya kandungan oksigen terlarut dalam air atau
tingginya kandungan karbon dioksida terlarut (Robert, 1992).
Ikan yang terserang penyakit dapat menunjukkan beberapa gejala antara
lain kematian yang mendadak, kurangnya nafsu makan, gerakan berenang yang
tidak normal, insang yang pucat, pembengkakan tubuh atau luka-luka pada ikan.
Kurniadie (2005) dan Meita (2005) mengemukakan bahwa ikan yang terinfeksi
bakteri A. hydrophila mengalami gejala klinis seperti organ dalam ikan hancur,
terjadi pendarahan, sisik mengelupas, tubuh luar memerah sampai borok dan
terjadi pembengkakan perut dan mata sehingga terlihat menonjol. Simptom
penyakit bervariasi karena tergantung dari beberapa faktor antara lain virulensi
dari bakteri, resistensi ikan terhadap infeksi, hadirnya atau tiadanya septicemia
dan bacterimia dan faktor yang diasosiasikan dengan stres pada ikan (Cipriano,

11
2001). Berikut beberapa gambar yang menunjukkan gejala klinis dari ikan yang
mati atau sensitif terhadap bakteri A. hydrophila
Gambar 2.2. Ikan gurame yang terserang bakteri A. hydrophila, bagian mata memerah (Sumber : Kusumawaty et al., 2008).
a
b
Gambar 2.3. Ikan gurame yang terserang bakteri A. hydrophila, (a) sisiknya
mengelupas dan (b) terdapat bercak-bercak merah di tubuhnya (Sumber : Kusumawaty et al., 2008)
Terdapat beberapa data tentang kasus serangan bakteri A. hydrophila
terhadap ikan air tawar. Serangannya terjadi sepanjang bulan Mei sampai Agustus
2002. Pada bulan Mei ratusan ton ikan mati secara mendadak akibat serangan
bakteri ini di Kabupaten Subang. Pada bulan Juni, wabah ini menyerang petani
ikan di Kabupaten Bandung dan Kabupaten Garut. Serangan bakteri yang terjadi
di Kabupaten Bandung menyebabkan kematian ikan mas sebanyak 63,3 ton.
Kejadian ini menyebabkan kerugian sebanyak 506,7 juta rupiah (Holipah, 2006).

12
Wabah ini juga menyerang empat kecamatan di Kabupaten Garut, yaitu
Kecamatan Bayongbong, Kadungora, Sukawening dan Cikajang. Kejadian di
Kabupaten Garut ini menyebabkan petani ikan mas tidak berhasil menjual hasil
tambaknya ke pasaran, bahkan ikan yang tersisa dipanen secara terpaksa (Holipah,
2006).
Pada bulan Juli 2002, di Kabupaten Bandung kembali terjadi wabah
serangan bakteri A. hydrophila dengan jumlah keseluruhan ikan yang terserang
adalah 442,4 ton dan hanya 10% dari jumlah tersebut yang berhasil diselamatkan.
Pada bulan yang sama, terjadi juga di daerah Kuningan serangan bakteri ini
menyebabkan kerugian sebesar 3,2 milyar rupiah dan di daerah tersebut, bakteri
Aeromonas hydrophila menyerang ikan mas, ikan tawes dan ikan gurame
(Holipah, 2006).
Pada bulan Mei 2003 ribuan petani ikan mas di sepanjang Sungai Cimala
dan Waduk Cirata di Kecamatan Cipeundeuy, Cikalong Wetan, dan Cililin,
Kabupaten Bandung, Jawa Barat, juga mengalami gagal panen karena ikan mas-
nya terserang bakteri Aeromonas. Sekitar 1.430 petani ikan dengan jumlah kolam
dan karamba mencapai 2.000 buah di kedua kecamatan tersebut dan 400
pembudidaya ikan mas yang menggunakan jaring terapung di Waduk Cirata
terancam bakteri Aeromonas. Ikan mas yang mati terserang bakteri Aeromonas
tampak mengambang di kolam air deras yang memanfaatkan aliran Sungai
Cimala. Ikan yang mati itu terlihat menggembung dengan insang membengkak.
Akibat wabah ini, diperkirakan produksi ikan mas dari wilayah ini menurun 70-
80% (Tn. 2003a).

13
Pada bulan Oktober 2005, warga Kanagarian Lubuk Pandan Kabupaten
Padang Pariaman dikejutkan oleh kematian ikan gurame lebih kurang 47 ton
gurame konsumsi dan 2,1 juta ekor benih yang siap untuk dipasarkan milik
kelompok tani ikan Mutiara Sukma di Kanagarian Lubuk Pandan. Lokasi yang
terserang luasnya sekitar 50 hektar dengan jumlah petani sekitar 80 KK, ditaksir
nilai kerugian lebih kurang Rp. 1,5 milyar. Masyarakat mengatakan penyakit aneh
dengan gejala ikan yang terserang nafsu makannya berkurang, juga menyerang
bagian mata dan kulit ikan. Kematian ikan sudah sudah berlangsung sejak awal
tahun 2005 tapi dalam jumlah yang sedikit (Diraja, 2007).
Beberapa kasus serangan bakteri A. hydrophila tersebut menimbulkan
banyak kematian pada ikan, tetapi masih ada ikan yang tetap hidup dalam jumlah
sedikit. Ikan yang tetap hidup tersebut merupakan ikan berkualitas baik karena
tahan terhadap serangan bakteri A. hydrophila. Selama ini, para petani ikan
gurame pada umumnya melihat kualitas ikan dari morfologi saja. Namun, petani
belum bisa menentukan secara pasti antara ikan yang resisten dengan ikan yang
sensitif terhadap bakteri A. hydrophila. Penanda DNA sangat potensial untuk
membantu memecahkan masalah ini, karena teknik ini sudah dapat menunjukkan
adanya perbedaan antara individu dalam satu species yang tinggi (Kusumawaty et
al., 2008). Dengan diperolehnya penanda DNA tersebut akan sangat membantu
para pemulia ikan gurame dengan program perbaikan mutu ikan untuk
mendapatkan ikan gurame yang unggul.

14
C. Penanda Genetik
Perkembangan ilmu pengetahuan yang sangat pesat dewasa ini,
memungkinkan saling menunjangnya perkembangan berbagai disiplin ilmu
pengetahuan. Beberapa kemajuan tersebut antara lain adalah perkembangan ilmu
biologi molekul yang memungkinkan diperolehnya suatu marka (penanda) gen
yang mengendalikan karakter suatu individu. Teknik-teknik yang digunakan
dalam genetika banyak menggunakan penanda genetik sebagai alat bantu
mengidentifikasi genotipe suatu individu. Penanda genetik merupakan ekspresi
pada individu yang terlihat oleh mata atau terdeteksi dengan alat tertentu, yang
menunjukkan dengan pasti genotif suatu individu (Widodo, 2003).
Aplikasi penanda genetik sangat luas. Bidang-bidang kedokteran,
pertanian, ilmu pangan, lingkungan, antropologi, sejarah, hukum
menggunakannya sebagai alat analisis atau alat pembuktian. Beberapa penanda
genetik sangat terpercaya karena tidak mudah berubah akibat pengaruh
lingkungan. Terdapat beberapa penanda genetik dalam menganalisis genom, yaitu
penanda morfologi, penanda biokmia dan penanda molekul.
Penanda morfologi merupakan penanda yang mudah dilihat oleh mata dan
telah banyak digunakan sejak masa awal genetika. Contohnya adalah warna,
ukuran, atau bentuk organ tertentu. Dalam jumlah besar penanda morfologi
dipelajari dan dipetakan untuk manusia, mencit, Drosophila, jagung, tomat serta
hewan dan tumbuhan lainnya (Liu, 1998). Penanda morfologi ini dapat
termodifikasi oleh pengaruh lingkungan sehingga dianggap tidak stabil. Selain itu,
penanda morfologi jumlahnya sangat terbatas dan untuk mengamatinya orang

15
harus menunggu hingga sifat penanda itu muncul sehingga penggunaan penanda
ini sangat terbatas.
Perkembangan ilmu pengetahuan yang begitu pesat, menyebabkan
ditemukanlah suatu penanda molekul yang mengandalkan sifat-sifat aplikatif
DNA. Penanda ini mulai dipakai semenjak ditemukannya secara berturut-turut
enzim endonuklease restriksi, teknik Southern blot, dan PCR sekitar tahun 1970.
Teknik elektroforesis gel, yang juga menjadi prasyarat penggunaan penanda ini,
telah dipakai bertahun-tahun sebelumnya. Dukungan dari bidang automasi,
robotika, dan bioinformatika terhadap teknik sekuensing pada tahun 1990 hingga
awal abad ke-21 menjadikan penanda molekul menjadi hal yang relatif ekonomis
untuk dikerjakan. Berikut ini adalah beberapa penanda molekul berdasarkan DNA
yang dikenal hingga saat ini, yaitu RFLP (Restriction Fragment Length
Polymorphisms), RAPD (Random Amplified Polymorphic DNA), ISSR (Inter
Simple Sequence Repeat) dan mikrosatelit (Widodo, 2003).
Restriction Fragment Length Polymorphisms atau RFLP adalah penanda
yang berasal dari susunan DNA yang terjadi karena proses evolusi, mutasi titik
pada situs enzim restriksi, insersi atau delesi dalam fragmen DNA. Restriction
Fragment Length Polymorphisms merupakan marker kodominan yang dapat
dipercaya dalam analisis keterpautan dan breeding. Penggunaan RFLP
menyebabkan proses analisis keterpautan dan breeding dapat ditentukan dengan
mudah jika karakter terdapat dalam bentuk homozigot atau heterozigot. Kelebihan
dari penanda RFLP adalah konsistensi yang tinggi, sifat pewarisan kodominan,
dapat diulang antarlaboratorium, tidak memerlukan informasi sekuen, dan relatif
mudah diinterpretasikan menjadi data matrik karena perbedaan yang besar antar

16
fragmen, tetapi penggunaan RFLP memerlukan DNA dalam jumlah yang besar
untuk pemotongan dengan enzim restriksi. Kekurangan lain dalam penggunaan
RFLP adalah penggunaan isotop radioaktif yang relatif mahal dan berbahaya serta
waktu yang diperlukan cukup lama (Semagn et al, 2006). Penanda lain adalah
RAPD merupakan penanda molekul yang menggunakan teknik PCR. Primer yang
digunakan dalam RAPD adalah Oligonukleotida tunggal pendek (10-12 basa)
dengan urutan acak untuk mengamplifikasi genomik DNA dengan temperatur
annealing yang rendah (Bardakci, 2001). Penanda lainnya adalah ISSR,
merupakan penanda molekul yang melibatkan amplifikasi segmen DNA yang
berada pada jarak yang dapat teramplifikasi antara dua daerah mikrosatelit
berulang yang identik tetapi dengan orientasi arah yang berbeda dan penanda yang
terakhir adalah mikrosatelit yang dikenal juga dengan simple sequence repeats
(SSRs) adalah kelas terkecil dari sekuen berulang yang terdiri dari dua, tiga atau
empat nukleotida. Penanda ini sering menunjukkan polimorfisme inter dan
intraspesifik dengan level tinggi dan terdapat melimpah dalam genom suatu
species. Salah satu contoh umum mikrosatelit adalah dinukleotida berulang
(CA)n, dimana n menunjukkan jumlah total nukeotida.
Karakteristik jenis-jenis penanda molekul tersebut dapat digunakan untuk
berbagai macam kepentingan yang biasanya bersifat diagnostik serta forensik.
Selain itu, penanda genetik bisa dipakai sebagai alat bantu seleksi dan pengukur
keanekaragaman genetik. Penanda genetik juga berguna dalam proses mengetahui
sidik jari DNA pada pembuktian forensik, uji serologi untuk mengetahui
kehadiran penyakit tertentu, pembuatan peta genetik, seleksi berbantuan marker
(marker-assisted selection, MAS), deskripsi keanekaragaman genetik, analisis

17
hubungan kekerabatan etnis manusia, analisis kekerabatan/taksonomi, analisis
kualitas lingkungan dan analisis kandungan bahan pangan/pakan (Widodo, 2003).
D. Teknik PCR (Polymerase Chain Reaction)
Untuk amplifikasi DNA digunakan teknik PCR yaitu suatu metode yang
digunakan untuk memperbanyak DNA secara in-vitro dengan dasar reaksi
enzimatik. Amplifikasi DNA secara eksponen terjadi melalui peningkatan dan
penurunan suhu di dalam mesin thermocycler sehingga terjadi tahapan sebagai
berikut, yaitu tahap denaturasi, penempelan primer (annealing) dan tahap
ekstensi. Tahap pertama denaturasi menggunakan suhu tinggi yaitu sekitar 90-
95oC. Selama denaturasi DNA rantai ganda akan tepisah menjadi DNA rantai
tunggal, semua reaksi enzimatik terhenti. Tahap selanjutnya adalah penempelan
primer, suhu yang digunakan berkisar antara 35-65oC (tergantung primer yang
digunakan) sehingga primer menempel pada DNA rantai tunggal. Setelah primer
menempel pada DNA cetakan, primer akan mengalami polimerisasi, yaitu suatu
proses penambahan deoksinukleosida trifosfat yang dikatalis oleh enzim DNA
polymerase, kemudian terjadi pemanjangan sehingga dihasilkan dua DNA rantai
ganda baru, pemanjangan ini terjadi pada suhu 72oC. Gambar 2.4.
Gambar 2.4 Gambaran tahapan-tahapan pada reaksi PCR yang dimulai dari
denaturasi, annealing sampai ekstensi (Vierstraete, 1999)

18
Pada Gambar 2.5 menunjukkan jumlah salinan DNA yang dihasilkan dari
satu siklus yang terdiri atas tiga tahapan. Jumlah salinan DNA target meningkat
secara eksponensial. Bila n jumlah siklus, maka jumlah salinan DNA target adalah
2n
Gambar 2.5 Ilustrasi sejumlah salinan DNA target yang meningkat secara
eksponensial. Bila n jumlah siklus, maka jumlah salinan DNA target adalah 2n ((Vierstraete, 1999)
Bahan bahan yang digunakan dalam PCR terdiri atas oligonukleotida
sebagai sumber energi dan donor basa untuk sintesis DNA. Primer, DNA
polymerase, DNA cetakan, buffer dan MgCl2. Siklus dalam PCR dapat berulang
beberapa kali sesuai dengan kebutuhan karena deoksinukleotida dan primer
terdapat dalam jumlah berlebih.
Enzim DNA polymerase hanya akan menambahkan deoksinukleotida
trifosfat pada nukleotida yang komplementer dengan nukleotida pada DNA
cetakan. Denaturasi rantai ganda memerlukan suhu tinggi (90-95oC), sehingga
mendenaturasi enzim yang digunakan. Oleh karena itu, digunakan enzim yang
tetap aktif dalam suhu tinggi. Enzim tersebut berasal dari bakteri temofilik,
Thermus aquaticus yang lebih dikenal dengan sebutan Taq polymerase (Saiki,
1988). Penggunaan enzim tahap panas ini mempermudah pengerjaan PCR karena
tidak membutuhkan panggantian enzim untuk setiap siklus dan setiap reaksi.

19
Selain itu enzim ini juga sangat efektif untuk proses polimerisasi pada suhu 72oC
karena enzim ini bekerja efektif pada suhu 70-75oC. Spesifitas reaksi amplifikasi
juga tergantung pada suhu penempelan primer yang ditentukan oleh urutan dan
panjang nukleotida primer (Lusiana, 1997).
E. Mikrosatelit
Mikrosatelit adalah sekuen sederhana yang berulang-ulang terdiri dari 2-4
motif sekuen nukleotida sebagai sekuen konservatif yang melimpah dalam genom
suatu species. Sebagai contoh adalah motif (GAG)10 =
(GAGGAGGAGGAGGAGGAGGAGGAGGAGGAG) atau (AT)20 =
(ATATATATATATATATATATATATATATATATATATATAT) yang aka n
berulang-ulang tanpa interupsi. Panjang pengulangan ini bervariasi tergantung
individu/varietas dan diwariskan secara baka kepada generasi berikutnya
(Prasetiyono et al., 2003).
Gambar 2.6. Penanda mikrosatelit dan aplikasinya. (Nagaraju et al.,2002)

20
Bentuk pengulangan sekuen DNA sederhana yang berulang-ulang
menjadikan penanda mikrosatelit sering disebut Simple Sequence Repeat (SSR),
Short Tandem Repeats (STRs) atau Simple Sequence Length Polymorphisms
(SSLPs) yang sekarang menjadi salah satu penanda paling banyak digunakan
secara luas untuk pemetaan genetik, analisis keragaman genetik, dan studi evolusi
(Temnykh et al., 2000). Mikrosatelit juga merupakan salah satu tipe polimorfisme
yang berulang-ulang, yang biasa dikelompokkan ke dalam Simple Tandem Repeat
Polymorphism (STRP), karena perbedaan genetik diantara molekul-molekul DNA
yang mengandung sejumlah kopi sekuen DNA pendek yang diulang beberapa
kali. STRP dengan pengulangan 10-60 pasang basa sering disebut minisatelit atau
variable number of tandem repeats (VNTR) sedangkan yang memiliki
pengulangan 2-4 pasang basa sering disebut mikrosatelit (Brown, 2002).
Gupta et al. (1996) menyebutkan bahwa sebagian besar minisatelit
berpusat dekat telomere, sedangkan mikrosatelit tersebar di seluruh genom. Salah
satunya adalah mikrosatelit kloroplas (cpSSRs) yang pada dasarnya mikrosatelit
ini sama dengan mikrosatelit di dalam inti sel, tetapi ulangan pada mikrosatelit
kloroplas hanya terdiri dari 1 pasang basa (misal (T)n). Setiap spesies biasanya
memiliki ciri khas dalam pengulangan sekuen sederhana ini. Misalnya pada padi,
sekuen mikrosatelit ini memiliki urutan dari yang terbanyak, yaitu: (GA)n, (GT)n,
(TTG)n, (ATT)n, (CGG)n, (TCT)n, (CAG)n, (TGG)n, (GATA)n, (ATC)n,
(CTTT)n, dan (CATG)n (McCouch et al., 1997). Secara umum sekuen AT paling
banyak terdapat pada tanaman, sedangkan AC/TG pada hewan (Morgante &
Olivieri, 1993 dalam prasetiyono et al., 2003). Motif lain yang juga banyak
didapatkan di dalam genom tanaman adalah (GA)n. Selain itu, motif (AAG)n dan

21
(AAT)n merupakan motif trinukleotida yang sering ditemukan dalam genom
tanaman. Pengulangan (CA)n terdapat pada genom manusia sekitar 50.000 buah,
dengan n sekitar 10 sampai 60. Pengulangan tri dan tetra nukleotida juga
umumnya terdapat pada genom manusia. Pengulangan motif CA juga banyak
didapatkan pada mammalia, namun motif ini sangat sedikit ditemukan di dalam
genom tanaman. Setiap 100 pb DNA akan didapatkan 2-3 lokus mikrosatelit pada
primata, 1,8 lokus pada ragi, dan 0,224 pada tanaman (Prasetiyono et al. 2003).
Menurut Weber (1990 dalam Kusumawaty et al.,2008) mengemukakan
bahwa panjang unit ulangan motif mikrosatelit, jumlah ulangan, dan
kesempurnaan atau kemurnian lokus mikrosatelit ini menentukan tingkat
polimorfisme suatu individu. Lokus mikrosatelit dinukleotida dengan jumlah
ulangan kurang dari 10 kali sering menghasilkan lokus yang monomorfik
sehingga hanya merupakan informasi untuk aplikasi mikrosatelit. Primer yang
digunakan untuk mengamplifikasi sampel sebaiknya berasal dari lokus
mikrosatelit dinukleotida yang mempunyai jumlah ulangan sedikitnya sepuluh
kali, sedangkan untuk trinukleotida sedikitnya tujuh kali dengan jumlah total
nukleotida dari ulangan tersebut sedikitnya adalah 20 nukleotida (Weber et al.,
1990 dalam Kusumawaty et al., 2008).
Ketika urutan yang berisi motif mikrosatelit sudah diperoleh, primer yang
spesifik dapat dirancang untuk mengamplifikasi lokus mikrosatelit. Hal ini dapat
dilakukan karena mikrosatelit memiliki daerah pengapit (Flankin region) yang
sifatnya lestari, sehingga dapat diaplikasikan pada individu atau varietas lain
(Mulyani 2003 dalam Kusumawaty et al., 2008). Primer merupakan

22
oligonukleotida untai tunggal yang menempel pada fragmen DNA yang
diamplifikasi. Menurut Weber (1990 dalam Kusumawaty et al., 2008), motif-
motif mikrosatelit pada genom manusia dapat dikelompokkan ke dalam empat
kategori, yaitu :1. Sempurna (Perfect) : bilamana diantara motif mikrosatelit
tidak diselingi oleh nukleotida lain. Contoh (GC)10: 2. Tidak Sempurna
(imperfect) : bilamana diantara motif mikrosatelit terdapat nukleotida yang bukan
motif mikrosatelit contoh (CT)gg(CT)6t(CT)5; Campuran sempurna (Perfect
Compound) merupakan gabungan dua atau lebih motif mikrosatelit dan diantara
motif mikrosatelit tidak terdapat nukleotida yang bukan motif mikrosatelit.
Contoh (CA)12(GC)10; dan 4. Campuran tidak sempurna (Imperfect compound) :
merupakan gabungan dua atau lebih motif mikrosatelit dan diantara kedua motif
mikrosatelit terdapat nukleotida yang bukan motif mikrosatelit. Contoh
(CA)7gcc(GT)6(CCG)5. Pemakaian motif mikrosatelit dengan jumlah
pengulangan dibawah 10 kali serta jumlah total nukleotida dari ulangan tersebut
kurang dari 20 nukleotida untuk keempat kategori ataupun motif mikrosatelit
kurang sempurna dengan jumlah ulangan lebih dari 10 kali dapat menghasilkan
alel-alel yang polimorfik (Kusumawaty et al., 2008)
Pada penelitian ini digunakan penanda mikrosatelit dengan motif
sempurna dan campuran sempurna dengan jumlah ulangan 4-15 kali. Penggunaan
mikrosatelit motif tersebut diharapkan dapat menghasilkan alel-alel yang
polimorfik. Selain itu, penggunaan mikrosatelit sebagai penanda yang bersifat
polimorfisme dan daya pembeda yang sangat tinggi, diharapkan dapat mendeteksi
keragaman alel pada level yang tinggi terutama membedakan individu yang

23
memiliki hubungan kekerabatan sangat dekat (Kusumawaty et al., 2008).
Mikrosatelit memiliki kemampuan segregasi populasi seakurat RFLP, yaitu dapat
membedakan antara yang homozigot dan heterozigot, dan dengan kemampuan
multiple loading pada gel poliakrilamid pendeteksian populasi segregasi dengan
menggunakan penanda mikrosatelit akan dapat dilakukan secara cepat, tepat, dan
efisien (McCouch et al., 1997). Berdasarkan hal tersebut maka mikrosatelit
banyak digunakan untuk mengetahui genetika populasi, segregasi populasi,
pemetaan genetik dan tingkat kekerabatan suatu individu (Skirnisdottir et al.,
2008).
Selain keunggulan, penanda mikrosatelit mempunyai kelemahan. Dalam
pembuatan primernya, mikrosatelit memerlukan investasi yang besar, karena
harus melakukan sekuensing dan primer mikrosatelit ini bersifat spesifik species
(sukar dipertukarkan antarspecies) (Tn.2008c).
F. Aplikasi Mikrosatelit
Mikrosatelit sudah digunakan di beberapa kepentingan, diantaranya
adalah menganalisis keanekaragaman pada hewan dan tumbuhan, genetika
populasi suatu individu, tingkat kekerabatan suatu individu, sidik jari DNA pada
pembuktian forensik dan pembuatan peta genetik. Prasetiyono et al., (2003)
menggunakan teknik mikrosatelit untuk menganalisis keterpautan marka
mikrosatelit dengan sifat toleransi terhadap keracunan Al pada padi hasil
persilangan Dupa x ITA131. Pada penelitian tersebut digunakan 190 galur
tanaman padi generasi kedua (F2) hasil persilangan Dupa x ITA131 dan

24
menggunakan 243 pasang primer mikrosatelit. Hasil survei primer diperoleh hasil
bahwa dari 243 primer mikrosatelit, hanya 110 primer yang menghasilkan pita
yang polimorfik, tiga primer tidak menghasilkan pita, tujuh primer menghasilkan
pita yang banyak (lebih dari empat pita) dan 123 primer lainnya menghasilkan
pita yang monomorfik. 110 primer yang polimorfik tersebut digunakan 76 primer
untuk amplifikasi DNA dimana dari 76 primer tersebut hanya 67 yang primer
yang dapat dinterpretasikan menjadi data matrik, sembilan primer tidak dapat
dinterpretasikan menjadi data matrik karena pita polimorfiknya terlalu dekat, pita
terlalu banyak atau pita terlalu tipis. Berdasarkan hasil penelitian tersebut
didapatkan dua penanda mikrosatelit pada kromosom VII (RM248 dan RM445)
yang diduga terpaut dengan sifat toleransi terhadap keracunan Al.
Elfstrom et al. (2005) juga menggunakan teknik mikrosatelit untuk
mengetahui karakteristik lokus mikrosatelit pada Patinopecten caurinus utara
yaitu satu-satunya golongan bivalvia yang mempunyai kemampuan berenang.
Primer yang digunakan untuk analisisnya berjumlah 30 primer dan dihasilkan 16
primer yang menghasilkan pita yang cukup bersih untuk dianalisis dan bersifat
polimorfik.
Skirnisdottir et al. (2008) menggunakan teknik mikrosatelit untuk
pencarian segmen DNA genom Gadus morhua L. berisi motif mikrosatelit dengan
menggunakan 10 dinukleotida, lima trinukleotida, dua tetranukleotida dan
heptanukleotida. Pada penelitiannya, digunakan dua kondisi PCR dengan suhu
annealing berbeda. Delapan primer mikrosatelit diamplifikasi pada suhu
annealing 55oC dan sepuluh primer diamplifikasi pada suhu annealing 58oC yang

25
menghasilkan frekuensi alel antara 6-45 alel dan menunjukkan sifat heterozigot
yang berbeda signifikan sesuai dengan teori Hardy-Weinberg. Dari hasil
penelitian tersebut diperoleh kesimpulan bahwa penggunaan teknik mikrosatelit
dengan dua kondisi PCR (suhu annealing) berbeda diperoleh keuntungan yang
besar yaitu selain waktu dan biaya yang murah juga resiko kesalahan pada saat
penelitian dapat diperkecil.
Keberhasilan teknik mikrosatelit juga diperkuat oleh Alpermann et al.
(2006) dalam pencarian segmen DNA Alexandrium tamarense yang berisi lokus
mikrosatelit. Pada penelitiannya diperoleh hasil bahwa DNA Alexandrium
tamarense dapat diamplifikasi. Jumlah alel dan diversitas gen pada Alexandrium
tamarense menunjukkan bahwa lokus mikrosatelit yang dihasilkan cukup
polimorfik untuk digunakan dalam studi populasi dan menunjukkan
keanekaragaman yang tinggi dalam Alexandrium tamarens.
Untuk menganalisis lokus mikrosatelit yang dihasilkan melalui proses
amplifikasi PCR, para peneliti di atas ada yang menggunakan gel agarosa atau gel
poliakrilamida untuk visualisasi lokus mikrosatelit tersebut. Hal tersebut
berpengaruh terhadap jumlah larik yang dihasilkan. Jumlah larik yang dihasilkan
dengan visualisasi menggunakan gel poliakrilamida lebih banyak dibanding gel
agarosa. Hal ini terjadi karena kemampuan separasi gel poliakrilamida lebih tinggi
dibanding gel agarose. Menurut Reddy et al. (1999) gel poliakrilamida secara
efektif dapat digunakan untuk pemisahan fragmen DNA dari ukuran 200-1000 pb.

26
G. Gel Poliakrilamida
Gel poliakrilamida adalah matriks separasi yang digunakan pada
elektroforesis biomolekul, seperti protein atau fragmen DNA. Teknik sekuensing
DNA tradisional seperti Maxam-Gilbert menggunakan gel poliakrilamida untuk
memisahkan fragmen DNA yang berbeda berdasarkan ukuran panjang base pair
(bp) sehingga urutannya bisa dibaca. Sekarang ini metode paling modern untuk
separasi DNA banyak menggunakan gel agarosa, tetapi untuk pemisahan fragmen
DNA yang kecil, gel poliakrilamida paling sering digunakan terutama dalam
bidang imunologi dan analisis protein.
Pada penelitian analisis variasi genetik menggunakan penanda
mikrosatelit, gel poliakrilamid lebih banyak digunakan dibandingkan dengan gel
agarosa. Clark et al., (2004) menggunakan gel poliakrilamida 6% untuk proses
separasi DNA dalam mempelajari genetik populasi ikan tuna atlantik dengan
menggunakan primer mikrosatelit. Dari 25 primer mikrosatelit yang digunakan,
dihasilkan alel sebanyak 3-23 alel dengan nilai heterozigositas tiap lokus antara
0,32-0,93. Elfstrom et al., (2005) juga menggunakan gel poliakrilamida 5% untuk
memvisualisasi hasil amplifikasi lokus mikrosatelit pada Patinopecten caurinus.
Penelitian tersebut menggunakan 30 primer mikrosatelit dan hanya 16 primer
yang menghasilkan larik yang bersih dan dapat dinterpretasikan menjadi data
matrik. Jumlah alel yang dihasilkan sebanyak 4-26 alel dengan nilai
heterozigositas tiap lokus antara 0,437-1. Chaix et al., (2002) menggunakan gel
agarosa sebagai media untuk memvisualisasi hasil amplifikasi Eucalyptus grandis
menggunakan penanda mikrosatelit. Penelitian tersebut menggunakan tujuh
primer mikrosatelit dan berhasil mengamplifikasi semua sampel Eucalyptus

27
grandis yang digunakan. Jumlah alel yang dihasilkan dalam penelitian tersebut
antara 2-6 alel . Hal ini membuktikan bahwa dalam proses separasi gel agarosa
memiliki kapasitas pemisahan yang lebih rendah dibandingkan dengan gel
poliakrilamida, tetapi memilik spektrum pemisahan yang lebih besar. Teknik
elektroforesis gel agarosa ini biasanya digunakan untuk memisahkan fragmen
DNA dengan ukuran relatif kecil yaitu sekitar 200 pb sampai kira-kira 50 kb.
Fragmen dengan ukuran di atas 50 kb tidak dapat dipisahkan lagi dengan baik
(Suryanto, 2003). Gel poliakrilamida mempunyai keunggulan dibandingkan
dengan gel agarosa yaitu memiliki kapasitas lebih tinggi dalam proses separasi
fragmen DNA sampai ukuran satu pasang basa dan dapat memuat kuantitas DNA
dalam jumlah besar (Sambrook et al., 1989).
Terdapat 2 jenis Gel poliakrilamida, yaitu: denaturing poliakrilamida dan
non- denaturing poliakrilamida. Perbedaan keduanya terletak pada ada atau
tidaknya urea pada komposisi gel poliakrilamida. Berikut ini tabel komposisi gel
poliakrilamida
Tabel 2.1 Komposisi Gel Non Denaturing Poliakrilamida
(Sumber : Sambrook et al., 1989)
Komposisi Gel 6% 8% 10% 12% Deion water (ml) 6,4 5,9 5,4 4,9 5x TBE (ml) 2,0 2,0 2,0 2,0 40% akrilamid (ml) 1,5 2,0 2,5 3,0 10% APS (µl) 100 100 100 100 TEMED (µl) 10 10 10 10 Total Volume 10 10 10 10
28
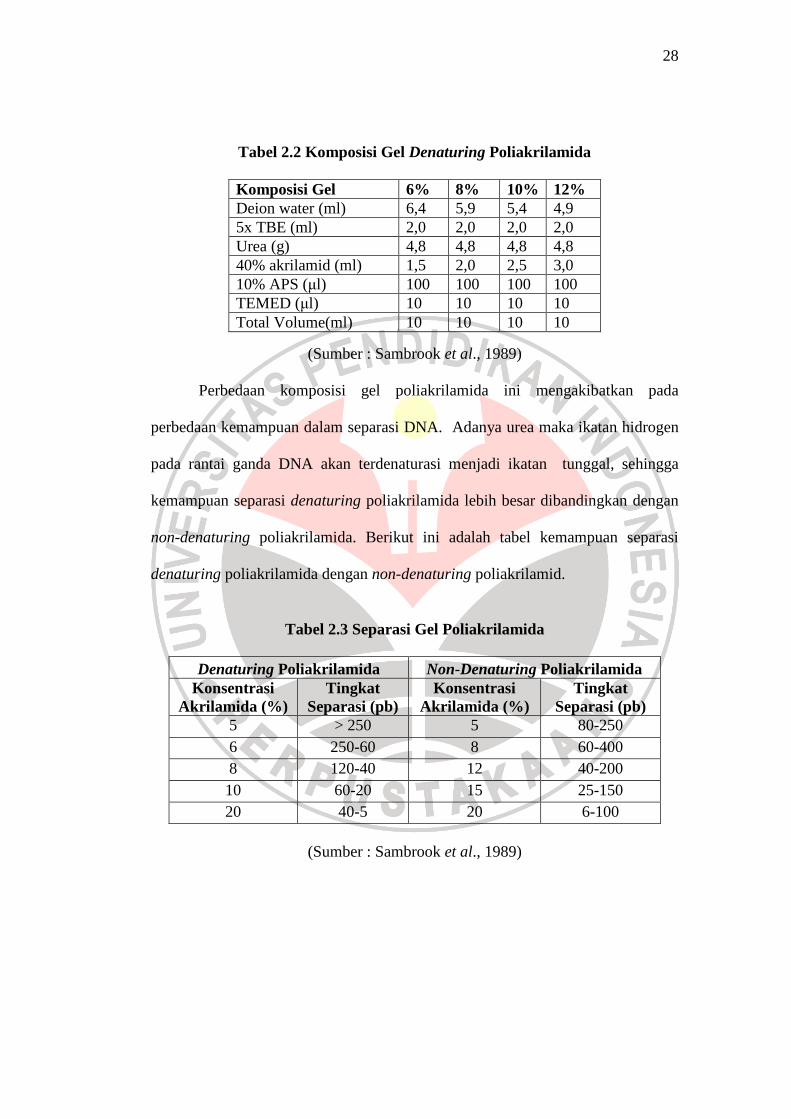
Tabel 2.2 Komposisi Gel Denaturing Poliakrilamida
(Sumber : Sambrook et al., 1989)
Perbedaan komposisi gel poliakrilamida ini mengakibatkan pada
perbedaan kemampuan dalam separasi DNA. Adanya urea maka ikatan hidrogen
pada rantai ganda DNA akan terdenaturasi menjadi ikatan tunggal, sehingga
kemampuan separasi denaturing poliakrilamida lebih besar dibandingkan dengan
non-denaturing poliakrilamida. Berikut ini adalah tabel kemampuan separasi
denaturing poliakrilamida dengan non-denaturing poliakrilamid.
Tabel 2.3 Separasi Gel Poliakrilamida
Denaturing Poliakrilamida Non-Denaturing Poliakrilamida
Konsentrasi Akrilamida (%)
Tingkat Separasi (pb)
Konsentrasi Akrilamida (%)
Tingkat Separasi (pb)
5 > 250 5 80-250 6 250-60 8 60-400 8 120-40 12 40-200 10 60-20 15 25-150 20 40-5 20 6-100
(Sumber : Sambrook et al., 1989)
Komposisi Gel 6% 8% 10% 12% Deion water (ml) 6,4 5,9 5,4 4,9 5x TBE (ml) 2,0 2,0 2,0 2,0 Urea (g) 4,8 4,8 4,8 4,8 40% akrilamid (ml) 1,5 2,0 2,5 3,0 10% APS (µl) 100 100 100 100 TEMED (µl) 10 10 10 10 Total Volume(ml) 10 10 10 10


![Clinical and Therapeutic Implications of Aeromonas Bacteremia: … · 2016-12-28 · Aeromonas bacteremia are malignancy and hepatobiliary dis-eases [5]. Aeromonas spp. tend to produce](https://img.pdfslide.net/doc/110x75/5ec79db8c2bd727c0b32cc58/clinical-and-therapeutic-implications-of-aeromonas-bacteremia-2016-12-28-aeromonas.jpg)


























