Embed Size (px)
Citation preview

11
http://journals.tubitak.gov.tr/botany/
Turkish Journal of Botany Turk J Bot(2017) 41: 11-24© TÜBİTAKdoi:10.3906/bot-1606-8
Relationships between Sorghum bicolor (Poaceae) and its closerelatives based on genomic in situ hybridization evidence
Qing LIU1,*, Huan LIU1, Jun WEN2
1Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou, P.R. China
2Department of Botany, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA
* Correspondence: [email protected]
1. IntroductionAmong global cereal crops, sorghum [Sorghum bicolor (L.) Moench] ranks fifth in terms of yield tonnage, exceeded only by wheat, maize, rice, and barley (http://faostat.fao.org/). Among these cereal crops, wheat (Triticum aestivum L.) is a hexaploid originated from diploid ancestors around the 0.4 million years ago (Marcussen et al., 2014). Maize (Zea mays L.) is a tetraploid with the last whole genome duplication dated to 11.9 million years ago (Paterson et al., 2004; Swigoňová et al., 2004). Major progress on the phylogenetic origin of rice (Oryza sativa L.) and barley (Hordeum vulgare L.) has been made through genome sequencing and transcriptome analyses (e.g., Dai et al., 2004; Huang et al., 2012). In contrast, the origin and diversity of S. bicolor have never been fully disentangled as the sorghum relatives are extremely complicated. Four main hypotheses have been proposed to explain the early phylogenetic history of S. bicolor: 1) it was an ancient allotetraploid derived from a hybrid between two diploid species (2n = 2x = 10) (Tang and Liang, 1988); 2) it was a natural autotetraploid originated via chromosome doubling from a single diploid ancestor (2n = 2x = 10)
(Swigoňová et al., 2004); 3) S. bicolor shared a common ancestor with maize through ancient polyploidization events (Paterson et al., 2004; Messing, 2009); and 4) some members of Sorghum subg. Sorghum were inferred to be immediate wild progenitors of S. bicolor based on cytological and molecular phylogenetic studies (Liu Q et al., 2014b). A better understanding of the genomic relationships between S. bicolor and its close relatives is critical for better utilization of Sorghum biodiversity.
Nuclear DNA data have suggested that S. × almum Parodi, S. arundinaceum (Desv.) Stapf, S. × drummondii (Nees ex Steud.) Millsp. & Chase, S. halepense (L.) Pers., and S. propinquum (Kunth) Hitchc. are closely related to S. bicolor in Sorghum subg. Sorghum (Liu Q et al., 2014b; Hawkins et al., 2015). Sorghum × almum is the fertile hybrid between S. bicolor and S. halepense, while S. × drummondii is the fertile hybrid between S. bicolor and S. arundinaceum (Doggett, 1970; Ng’uni et al., 2010). Analysis of meiotic chromosome behavior suggested that S. halepense (2n = 4x = 40) might be a segmental tetraploid involving two genomes, with one genome very similar to S. bicolor and another relatively divergent one (Tang and Liang, 1988).
Abstract: Sorghum bicolor (L.) Moench (sorghum) is a naturally well-established diploid species with 2n = 2x = 20. Here we provide evidence to identify the genomic relationships of S. bicolor and its extant relatives using genomic in situ hybridization. The genomic divergences between S. bicolor and its close relatives may be presented as: S. bicolor-S. × drummondii (Nees ex Steud.) Millsp. & Chase < S. bicolor-S. × almum Parodi < S. bicolor S. arundinaceum (Desv.) Stapf < S. bicolor-S. propinquum (Kunth) Hitchc. Sorghum arundinaceum probe signals showed spotted or painted patterns on the S. bicolor chromosomes, indicating that the degree of genomic divergence between S. bicolor and S. arundinaceum is distinct. It is reasonable to infer that S. bicolor might have diverged from S. arundinaceum in the early diversification history of the subgenus. The probe signal intensity of S. arundinaceum and S. propinquum is greater on the 17 chromosomes than on the remaining 23 chromosomes of S. halepense (L.) Pers., whereas S. bicolor and S. propinquum probes produced spotted or patchy signals on the 40 S. halepense chromosomes. Therefore, the degree of genomic divergence may be presented as: S. halepense-S. bicolor < S. halepense-S. arundinaceum < S. halepense-S. propinquum.
Key words: Cereal crops, cultivated sorghum, genomic in situ hybridization, genomic relationship, Sorghum halepense
Received: 07.06.2016 Accepted/Published Online: 30.08.2016 Final Version: 17.01.2017
Research Article

LIU et al. / Turk J Bot
12
However, the degree of genomic divergence between S. bicolor and its extant relatives remains ambiguous due to their unresolved phylogenetic positions among the short paraphyletic branches in molecular studies (Ng’uni et al., 2010; Liu Q et al., 2014b; Hawkins et al., 2015). Elucidating the genomic relationships of S. bicolor and its close relatives is not only of phylogenetic interest to reconstruct its origin, but is also of practical value, allowing more efficient utilization of wild germplasm resources in S. bicolor breeding.
Repetitive sequences occupy as much as 90% of the genomic DNA (gDNA) in cereal crops (Heslop-Harrison, 2000; Dodsworth et al., 2015). The renowned geneticist Hitoshi Kihara noted that the history of all organisms was inscribed in the chromosomes based on wheat genome studies (Crow, 1994). Hence, comparisons of repetitive sequences among chromosomes have considerable potential for understanding the genomic diversity of cereal crops (Katsiotis et al., 2000; Price et al., 2005; Dodsworth et al., 2015). Genomic in situ hybridization (GISH) studies offer a powerful system for detection of repetitive sequence (mostly transposable elements) distribution using labeled probes on denatured chromosome DNA spread onto microscope slides, owing to the repetitive sequences of chromosomal target regions comprising several kilobases of homologous sequences of probes (Biscotti et al., 2015). GISH experiments have shown genome-wide evidence of sequence homogenization among crops and their candidate progenitors, thus supporting hypotheses derived from molecular phylogenetic investigations
(Hunt et al., 2014; Liu Q et al., 2014a). In addition, the repetitive sequences were enormously variable over time even among very closely species; therefore, GISH experiments complementing molecular phylogenetic data are more effective in estimating the degree of genomic differentiation as previously proposed (Li et al., 2001a), e.g., the diploid Hordeum marinum Huds. (sea barley) (Carmona et al., 2013), the tetraploid Eleusine coracana (L.) Gaertn. (finger millet) (Liu et al., 2011), and the tetraploid Panicum miliaceum (proso millet) (Hunt et al., 2014). In this study, GISH was applied to determine the degree of genomic differentiation between S. bicolor and its extant close relatives in Sorghum subg. Sorghum, and to elucidate the degree of relatedness between S. halepense and its close relatives with a focus on identifying the degree of genomic differentiation between S. bicolor and S. halepense.
2. Materials and methods2.1. Plant materials and root collectionThe material sources and the ploidy levels of species investigated here are summarized in Table 1. Mature seeds were collected from plants grown in the South China Botanical Garden, Chinese Academy of Sciences (Guangzhou, China). All voucher specimens were deposited at the South China Botanical Garden Herbarium (IBSC). Excised root tips (5–10 mm) from vigorously growing plants were pretreated with 4 mmol/L of 8-hydroxyquinoline for 2 h at room temperature and then fixed in 3:1 (v/v) absolute ethanol and glacial acetic acid for 12 h at 4 °C, and stored in 70% ethanol at –20 °C.
Table 1. Sources and chromosome numbers of materials used in the study. -1 and -2 refer to different accessions of the same species, respectively.IL, Accession number in the International Livestock Research Institute (ILRI) at Addis Ababa, Ethiopia; PI, plant introduction number of United States Department of Agriculture (USDA) at Beltsville, MD, USA; Herbarium (IBSC, South China Botanical Garden Herbarium; CANB, Australian National Herbarium); RQHN, Ru Qin Hua Nan (the abbreviation of the Chinese phonetic spelling of “invasive species in South China”).
Species Accessiona Herbarium Life cycleChromosomenumber
Locality
Sorghum × almum Parodi-1 Liu 237 (ILRI 13515) IBSC/US Perennial 2n = 2x = 40 Ethiopia
S. × almum-2 Liu 432 (PI 190579) IBSC Argentina
S. × drummondii (Nees ex Steud.) Millsp. & Chase Liu 238 (ILRI 15974) IBSC/US Annual 2n = 2x = 20 China
S. bicolor (L.) Moench-1 Mitchell 8765 CANB Annual 2n = 2x = 20 Australia
S. bicolor-2 Liu 433 (PI 287556) IBSC Zimbabwe
S. arundinaceum (Desv.) Stapf-1 Liu 434 (ILRI 17109) IBSC Annual 2n = 2x = 20 Mali
S. arundinaceum-2 Liu 232 (ILRI 13283) IBSC/US Kenya
S. halepense (L.) Pers.-1 Liu 234 (PI 302268) IBSC/US Perennial 2n = 4x = 40 Tanzania
S. halepense-2 RQHN 00603 (IBSC) IBSC China
S. propinquum (Kunth) Hitchc. Liu 233 (PI 653737) IBSC/US Perennial 2n = 2x = 20 USA

LIU et al. / Turk J Bot
13
2.2. Chromosome preparationsSomatic chromosome spreads were prepared according to Li et al. (2001b). Root tips were digested in an enzyme mixture of 1% (w/v) cellulase OnozukaR-10 (Sangon Biotech, Shanghai, China) and 5% (w/v) pectolase Y-23 (Sangon Biotech) for 270 min at 37 °C, and then squashed in 45% acetic acid on poly-L-lysine coated slides (Citglas) by applying pressure to an 18 × 18 mm coverslip. Slides were examined under an Axioplan-2 imaging microscope (Carl Zeiss, Oberkochen, Germany) equipped with a phase-contrast device, and those with acceptable numbers of somatic metaphase cells (5–10) were kept at –80 °C prior to use.2.3. Genomic in situ hybridization and signal detectionTotal gDNA was extracted from green leaves of Sorghum bicolor, S. × almum, S. arundinaceum, S. × drummondii, S. propinquum, and S. halepense using the standard 2X CTAB method (Doyle and Doyle, 1987). DNA concentrations were determined by spectrophotometer (NanoDrop 2000, Wilmington, DE, USA) and gel electrophoresis. After preshearing by ultrasonic processor (Scientz IID, Ningbo, China) (Jiang et al., 2012), total gDNA from sorghum relatives used as probes was labeled with digoxigenin-11-dUTP or biotin-16-dUTP (Roche, Mannheim, Germany) by nick translation. The probe length was estimated by gel electrophoresis to be 200–500 bp after purification with the DNA Probe Purification Kit (Omega Bio-Tek Inc., Norcross, GA, USA). The blocking DNA was autoclaved for 10 min to obtain 100–300 bp fragments (Heslop-Harrison et al., 1990).
Chromosome preparations were pretreated with 100 µg/mL RNase A for 60 min and with 0.01% pepsin for 40 min, followed by denaturation in 4% paraformaldehyde in 1X phosphate-buffered saline (pH 7.4) for 10 min at 37 °C, then washed in 2X standard saline citrate (SSC) and dehydrated using a gradient ethanol series (70%, 80%, and 99.8%, 2 min at each stage). The hybridization mix contained DNA probes at a concentration of 2.5–3.5 ng/µL, 50% (v/v) deionized formamide, 10% (w/v) dextran sulfate, 0.1% (w/v) sodium dodecyl sulfate (SDS) in 2X SSC, and salmon sperm-blocking DNA at 80- to 100-fold more than labeled probes. The slides with chromosomes were denatured together with the hybridization mixture (90 °C for 10 min), cooled quickly on ice for 10 min, and allowed to hybridize for 12–16 h in a humid chamber at 37 °C (Schwarzacher and Heslop-Harrison, 2000; Liu Q et al., 2014a).
After overnight hybridization at 37 °C, slides were sequentially washed at room temperature in 0.1% (v/v) SDS in 2X SSC at 42 °C three times and 0.1% (v/v) SDS in 0.2X SSC three times (5 min each), followed by a blocking with 5% (w/v) bovine serum albumin (BSA) in 4X SSC/Tween-20 at 37 °C for 30 min. The washing
stringency for S. bicolor slides was 84%–87%, while that for other species slides was 77%–88%. The signals from the digoxigenin-labeled probe were detected with 30 µL of rhodamine-conjugate anti-DIG antibody (Roche) in 1X PBS and 5% (w/v) BSA, and those of the biotin-labeled probe were detected with 30 µL of avidin-FITC antibody (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA). Chromosome preparations were counterstained with 2 µg/mL 4’,6-diamidino-2-phenylindole (DAPI, Roche) and mounted with Vectashield medium (Vector Laboratories, Burlingame, VT, USA).2.4. Image capturing and GISH signal quantificationAll images were obtained using a monochrome AxioCam HRC CCD camera (Carl Zeiss) attached to an Axioplan-2 imaging fluorescence microscope and processed using ISIS v.5.5.2 (MetaSystems, Altlußheim, Germany). Digital images were combined using Zeiss Axiovision v.4.6, then imported into Photoshop CS6 v.13.0 (Adobe, San Jose, CA, USA) for further processing. The relative intensity of GISH signals was quantified using AxioVisionRel v.4.6 (Carl Zeiss) (Iourov et al., 2005) with three technical replicates per signal. Descriptive statistics of signal intensity were presented as values of mean ± standard deviation, and their color coding was processed by styles-conditional format of the default setting in Office Excel 2007 v.12.0 (Microsoft, Redmond, WA, USA) (Table 2).
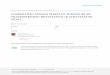
3. Results3.1. GISH analyses of Sorghum bicolor and its extant relativesFigures 1a–1e show results for S. bicolor (2n = 2x = 20) using biotin-labeled gDNA of S. × almum and digoxigenin-labeled gDNA of S. arundinaceum as probes. Under 84%–87% washing stringency, the 20 chromosomes showed green and red signals of variable intensity (Figure 1f) and painted dispersion pattern. The values of green/red fluorescent intensity for the 13 chromosomes (no. 1, 2, 3, 4, 5, 8, 9, 10, 11, 12, 13, 19, and 20) were 0.62–0.80. In contrast, the values of green/red fluorescent intensity for the remaining seven chromosomes (no. 6, 7, 14, 15, 16, 17, and 18) were 0.55–0.58, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the seven chromosomes were 0.76–1.01 (Supplementary Material: Table S1). In short, the intensity of the proportion of green signals (S. × almum probe) in red signals (S. arundinaceum probe) was greater for the 13 chromosomes than for the 7 chromosomes, and the intensity proportion of green signals in labeling signals was greater for the 7 chromosomes than for the 13 chromosomes. This result suggests that the degree of genomic divergence between S. bicolor and S. × almum is less than that between S. bicolor and S. arundinaceum (Table 2).

LIU et al. / Turk J Bot
14
Table 2. Degree of genome differentiation in GISH experiments.
No. Slides Probe combination*Dispersion pattern of hybridization signals
Summary of degree of genome differentiation
1 S. bicolor-2BIO-S. × almum-2 Painted
S. bicolor-S. × almum < S. bicolor-S. arundinaceumDIG-S. arundinaceum-2 Painted
2 S. bicolor-2BIO-S. × almum-2 Spotted
S. bicolor-S. × drummondii < S. bicolor-S. × almumDIG-S. × drummondii Painted
3 S. bicolor-2BIO-S. × drummondii Painted
S. bicolor- S. × drummondii < S. bicolor- S. arundinaceumDIG-S. arundinaceum-2 Painted
4 S. bicolor-2BIO-S. arundinaceum-2 Spotted
S. bicolor-S. arundinaceum < S. bicolor-S. propinquumDIG-S. propinquum Patchy
5 S. bicolor-2BIO-S. propinquum Patchy
S. bicolor-S. arundinaceum < S. bicolor-S. propinquumDIG-S. arundinaceum-2 Spotted
6 S. arundinaceum-1BIO-S. × drummondii Spotted
S. arundinaceum-S. × drummondii < S. arundinaceum-S. × almumDIG-S. × almum-2 Patchy
7 S. × drummondiiBIO-S. bicolor-1 Spotted
S. × drummondii-S. bicolor < S. × drummondii-S. arundinaceumDIG-S. arundinaceum-2 Patchy
8 S. halepense-1BIO-S. propinquum Patchy
S. halepense-S. arundinaceum < S. halepense-S. propinquumDIG-S. arundinaceum-2 Spotted
9 S. halepense-1BIO-S. propinquum Patchy
S. halepense-S. bicolor < S. halepense-S. propinquumDIG-S. bicolor-2 Spotted
*BIO probe labeled with biotin; DIG probe labeled with digoxigenin.
cba
d e f
Figure 1. Genomic in situ hybridization of Sorghum bicolor probed with gDNA of S. × almum and S. arundinaceum and GISH signal quantification. (a) GISH signals of S. × almum gDNA labeled with biotin (green); the enlargement illustrates green painted signals on chromosome no. 12 (inset). (b) GISH signals of S. arundinaceum gDNA labeled with digoxigenin (red); the enlargement illustrates red painted signals on chromosome no. 12 (inset). (c–d) Combined images of DAPI labeling S. bicolor metaphase chromosomes with Figures 1a and 1b, respectively. (e) A combined image of Figures 1a and 1b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. bicolor chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
15
Figures 2a–2e show results for S. bicolor using probes from biotin-labeled gDNA of S. × almum and digoxigenin-labeled gDNA of S. × drummondii. Under 84%–87% washing stringency, the 20 chromosomes of S. bicolor show green spotted and red painted dispersion signals of variable fluorescent intensity (Figure 2f). The values of green/red fluorescent intensity for 20 chromosomes were 0.60–0.71. In contrast, the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 20 chromosomes were 0.28–0.57 (Supplementary Material: Table S2). Thus, the intensity proportion of green signals (S. × almum probe) in red signals (S. × drummondii probe) was greater for the 20 chromosomes, whereas the intensity proportion of green signals was low for the 20 chromosomes. These results indicate that the degree of genomic divergence between S. bicolor and S. × drummondii is less than that between S. bicolor and S. × almum (Table 2).
Figures 3a–3e show the results for S. bicolor using probes from biotin-labeled gDNA of S. × drummondii and digoxigenin-labeled gDNA of S. arundinaceum. Under 84%–87% washing stringency, the 18 S. bicolor chromosomes showed green and red signals with similar fluorescent intensity and painted dispersion pattern, while red signal intensity was stronger than green signal intensity for chromosomes no. 5 and 6 (Figure 3f). The values of
green/red fluorescent intensity for the 18 chromosomes were 0.80–1.37, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 18 chromosomes were 0.81–2.97 (Supplementary Material: Table S3). The intensity proportion of green signals (S. × drummondii probe) in red signals (S. arundinaceum probe) was greater for the 18 chromosomes than for the 2 chromosomes, and the intensity proportion of green signals in labeling signals was greater for the 18 chromosomes. This result supports that the degree of genomic divergence between S. bicolor and S. × drummondii is less than that between S. bicolor and S. arundinaceum (Table 2)
Figures 4a–4e show the results for S. bicolor using probes from biotin-labeled gDNA of S. arundinaceum and digoxigenin-labeled gDNA of S. propinquum. Under 84%–87% washing stringency, the 19 S. bicolor chromosomes showed green spotted and red patchy signals of variable fluorescent intensity, while red signal intensity was similar to green signal intensity for chromosome no. 5 (Figure 4f). The values of green/red fluorescent intensity for the 19 chromosomes (except for chromosome no. 20) were 0.57–0.84, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 19 chromosomes were 0.32–1.16 (Supplementary Material: Table S4). Briefly, the intensity proportion of green signals (S. arundinaceum
d
cba
e f
Figure 2. Genomic in situ hybridization of Sorghum bicolor probed with gDNA of S. × almum and S. × drummondii and GISH signal quantification. (a) GISH signals of S. × almum gDNA labeled with biotin (green); the enlargement illustrates green spotted signals on chromosome no. 11 (inset). (b) GISH signals of S. × drummondii gDNA labeled with digoxigenin (red); the enlargement illustrates red painted dispersion signals on chromosome no. 11 (inset). (c–d) Combined images of DAPI labeling S. bicolor metaphase chromosomes with Figures 2a and 2b, respectively. (e) A combined image of Figures 2a and 2b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. bicolor chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
16
cba
d e f
Figure 3. Genomic in situ hybridization of Sorghum bicolor probed with gDNA of S. × drummondii and S. arundinaceum and GISH signal quantification. (a) GISH signals of S. × drummondii gDNA labeled with biotin (green); the enlargement illustrates green painted signals on chromosome no. 8 (inset). (b) GISH signals of S. arundinaceum gDNA labeled with digoxigenin (red); the enlargement illustrates red painted signals on chromosome no. 8 (inset). (c–d) Combined images of DAPI labeling S. bicolor metaphase chromosomes with Figures 3a and 3b, respectively. (e) A combined image of Figures 3a and 3b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. bicolor chromosomes. Scale = 5 µm.
cba
d e f
Figure 4. Genomic in situ hybridization of Sorghum bicolor probed with gDNA of S. arundinaceum and S. propinquum and GISH signal quantification. (a) GISH signals of S. arundinaceum gDNA labeled with biotin (green); the enlargement illustrates green spotted signals on chromosome no. 16 (inset). (b) GISH signals of S. propinquum gDNA labeled with digoxigenin (red); the enlargement illustrates red patchy signals on chromosome no. 16 (inset). (c–d) Combined images of DAPI labeling S. bicolor metaphase chromosomes with Figures 4a and 4b, respectively. (e) A combined image of Figures 4a and 4b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. bicolor chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
17
probe) in red signals (S. propinquum probe) was high for the 19 chromosomes, and the intensity proportion of green signals in labeling signals was high for the 19 chromosomes. The result suggests that the degree of genomic divergence between S. bicolor and S. arundinaceum is less than that between S. bicolor and S. propinquum (Table 2).
Figures 5a–5e show the results for S. bicolor using probes from biotin-labeled gDNA of S. propinquum and digoxigenin-labeled gDNA of S. arundinaceum. Under 84%–87% washing stringency, the 11 S. bicolor chromosomes (no. 2, 4, 6, 7, 8, 9, 10, 12, 14, 16, and 18) showed green patchy and red spotted signals of variable fluorescent intensity, while red signal intensity was similar to green signal intensity for the remaining 9 chromosomes (no. 1, 3, 5, 11, 13, 15, 17, 19, and 20) of S. bicolor (Figure 5f). The values of green/red fluorescent intensity for the 11 chromosomes were 0.45–0.81, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 11 chromosomes were 0.38–1.43 (Supplementary Material: Table S5). Therefore, the intensity proportion of green signals (S. propinquum probe) in red signals (S. arundinaceum probe) was lower for the 11 chromosomes than for the 9 chromosomes, and the intensity proportion of green signals in labeling signals was low for the 11 chromosomes. This demonstrates that the degree of genomic divergence between S. bicolor and
S. arundinaceum is less than that between S. bicolor and S. propinquum (Table 2).3.2. GISH analysis of Sorghum arundinaceum and its relativesFigures 6a–6e show the results for S. arundinaceum using probes from biotin-labeled gDNA of S. × drummondii and digoxigenin-labeled gDNA of S. × almum. Under 77%–88% washing stringency, the 20 S. arundinaceum chromosomes showed green spotted (S. × drummondii probe) and red patchy (S. × almum probe) signals with similar fluorescent intensity (Figure 6f). The values of green/red fluorescent intensity for the 20 chromosomes were 1.07–1.28, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 20 chromosomes were 0.50–0.82 (Supplementary Material: Table S6). In other words, the intensity proportion of green signals (S. × drummondii probe) in red signals (S. × almum probe) was high for the 20 chromosomes, and the intensity proportion of green signals in labeling signals was high for the 20 chromosomes, demonstrating that the degree of genomic divergence between S. arundinaceum and S. × drummondii is less than that between S. arundinaceum and S. × almum (Table 2).3.3. GISH analysis of S. × drummondii and its relativesFigures 7a–7e show the results for S. × drummondii using probes from biotin-labeled gDNA of S. bicolor and
cba
d e f
Figure 5. Genomic in situ hybridization of Sorghum bicolor probed with gDNA of S. propinquum and S. arundinaceum and GISH signal quantification. (a) GISH signals of S. propinquum gDNA labeled with biotin (green); the enlargement illustrates green patchy signals on chromosome no. 19 (inset). (b) GISH signals of S. arundinaceum gDNA labeled with digoxigenin (red); the enlargement illustrates red spotted signals on chromosome no. 19 (inset). (c–d) Combined images of DAPI labeling S. bicolor metaphase chromosomes with Figures 5a and 5b, respectively. (e) A combined image of Figures 5a and 5b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. bicolor chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
18
cba
d e f
Figure 6. Genomic in situ hybridization of Sorghum arundinaceum probed with gDNA of S. × drummondii and S. × almum and GISH signal quantification. (a) GISH signals of S. × drummondii gDNA labeled with biotin (green); the enlargement illustrates green spotted signals on chromosome no. 13 (inset). (b) GISH signals of S. × almum gDNA labeled with digoxigenin (red); the enlargement illustrates red patchy signals on chromosome no. 13 (inset). (c–d) Combined images of DAPI labeling S. arundinaceum metaphase chromosomes with Figures 6a and 6b, respectively. (e) A combined image of Figures 6a and 6b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. arundinaceum chromosomes. Scale = 5 µm.
d
cba
e f
Figure 7. Genomic in situ hybridization of Sorghum × drummondii probed with gDNA of S. bicolor and S. arundinaceum and GISH signal quantification. (a) GISH signals of S. bicolor gDNA labeled with biotin (green); the enlargement illustrates green spotted signals on chromosome no. 16 (inset). (b) GISH signals of S. arundinaceum gDNA labeled with digoxigenin (red); the enlargement illustrates red patchy signals on chromosome no. 16 (inset). (c–d) Combined images of DAPI labeling S. × drummondii metaphase chromosomes with Figures 7a and 7b, respectively. (e) A combined image of Figures 7a and 7b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 20 S. × drummondii chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
19
digoxigenin-labeled gDNA of S. arundinaceum. Under 77%–88% washing stringency, the 18 S. × drummondii chromosomes showed green spotted (S. bicolor probe) and red patchy (S. arundinaceum probe) signals of variable fluorescent intensity, while red signal intensity was similar to green signal intensity for 2 S. × drummondii chromosomes (no. 6 and 10) (Figure 7f). The values of green/red fluorescent intensity for the 10 chromosomes (no. 6, 9, 10, 11, 12, 13, 14, 16, 19, and 20) were 0.81–1.16, and the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 10 chromosomes were 0.52–1.04 (Supplementary Material: Table S7). Thus, the intensity proportion of green signals (S. bicolor probe) in red signals (S. arundinaceum probe) was greater for the former 10 chromosomes than for the remaining 10 chromosomes (no. 1, 2, 3, 4, 5, 7, 8, 15, 17, and 18), and the intensity proportion of green signals in labeling signals was high for the former 10 chromosomes. The result suggests that the degree of genomic divergence between S. × drummondii and S. bicolor is less than that between S. × drummondii and S. arundinaceum (Table 2).3.4. GISH analyses of S. halepense and its relativesFigures 8a–8e show the results for S. halepense using probes from biotin-labeled gDNA of S. propinquum and digoxigenin-labeled gDNA of S. arundinaceum. Under 77%–88% washing stringency, the 38 S. halepense
chromosomes showed green patchy and red spotted signals of variable fluorescent intensity, while red signal intensity was similar to green signal intensity for 2 S. halepense chromosomes (no. 1 and 4) (Figure 8f). The values of green/red fluorescent intensity for the 17 chromosomes (no. 1, 2, 3, 4, 5, 6, 10, 16, 18, 25, 26, 28, 29, 31, 32, 35, and 38) were 0.70–0.96, while the values of (green/red)/[(green+red)/blue] fluorescent intensity for the 40 chromosomes were 0.46–0.60 (Supplementary Material: Table S8). In short, the intensity proportion of green signals (S. propinquum probe) in red signals (S. arundinaceum probe) was greater for the 17 chromosomes than for the remaining 23 chromosomes, and the intensity proportion of green signals in labeling signals was low for the 40 chromosomes. The result indicates that the degree of genomic divergence between S. halepense and S. arundinaceum is less than that between S. bicolor and S. propinquum (Table 2).
Figures 9a–9e show the results for S. halepense using probes from biotin-labeled gDNA of S. propinquum and digoxigenin-labeled gDNA of S. bicolor. Under 77%–88% washing stringency, the 40 S. halepense chromosomes showed green patchy and red spotted signals of variable fluorescent intensity (Figure 9f). The values of green/red fluorescent intensity for the 40 chromosomes were 0.26–0.63, and the values of (green/red)/[(green+red)/blue]
d f
cba
e
Figure 8. Genomic in situ hybridization of Sorghum halepense probed with gDNA of S. propinquum and S. arundinaceum and GISH signal quantification. (a) GISH signals of S. propinquum gDNA labeled with biotin (green); the enlargement illustrates green patchy signals on chromosome no. 30 (inset). (b) GISH signals of S. arundinaceum gDNA labeled with digoxigenin (red); the enlargement illustrates red spotted signals on chromosome no. 30 (inset). (c–d) Combined images of DAPI labeling S. halepense metaphase chromosomes with Figures 8a and 8b, respectively. (e) A combined image of Figures 8a and 8b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 40 S. halepense chromosomes. Scale = 5 µm.

LIU et al. / Turk J Bot
20
fluorescent intensity for 37 chromosomes (except for no. 6, 7, and 39) were 0.32–0.69 (Supplementary Material: Table S9). Consequently, the intensity proportion of green signals (S. propinquum probe) in red signals (S. bicolor probe) was low for the 40 chromosomes, and the intensity proportion of green signals in labeling signals was lower for the 37 chromosomes than for the 3 chromosomes. This indicates that the degree of genomic divergence between S. halepense and S. bicolor is less than that between S. halepense and S. propinquum (Table 2).
4. DiscussionGISH results show the degree of genomic divergence between S. bicolor and its extant relatives. The degree of genomic divergence between S. bicolor and S. × drummondii is less than that between S. bicolor and S. ×
almum, which, in turn, is less than that between S. bicolor and S. arundinaceum, which again is less than that between S. bicolor and S. propinquum. The degree of genomic divergence may presented as the ranked species pairs as follows: S. bicolor-S. × drummondii < S. bicolor-S. × almum < S. bicolor-S. arundinaceum < S. bicolor-S. propinquum (Figure 10).
Inferring the degree of genomic divergence between S. bicolor and S. propinquum and that between S. arundinaceum and S. × drummondii is challenging. The separation of S. propinquum from S. bicolor is supported by geographic distribution and morphological characters. The natural distribution of S. bicolor is African, whereas S. propinquum occurs in Sri Lanka, southern India, and Myanmar, extending as far as the Southeast Asian Islands. Annual nonrhizomatous S. bicolor can easily
ba c
d e f
Figure 9. Genomic in situ hybridization of Sorghum halepense probed with gDNA of S. propinquum and S. bicolor and GISH signal quantification. (a) GISH signals of S. propinquum gDNA labeled with biotin (green); the enlargement illustrates green patchy signals on chromosome no. 34 (inset). (b) GISH signals of S. bicolor gDNA labeled with digoxigenin (red); the enlargement illustrates red spotted signals on chromosome no. 34 (inset). (c–d) Combined images of DAPI labeling S. halepense metaphase chromosomes with Figures 9a and 9b, respectively. (e) A combined image of Figures 9a and 9b. (f) Scatter diagram of blue (circle), green (square), and red (triangle) fluorescence signal intensity. Numbers indicate 40 S. halepense chromosomes. Scale = 5 µm.
Sorghum bicolorSorghum arundinaceum
Sorghum × drummondiiSorghum × almum
Sorghum halepense
Sorghum propinquum
Figure 10. Schematic diagram of genomic relationships of Sorghum bicolor and its extant close relatives. Numbers in circles 1-5 indicate degrees of genomic divergence of S. bicolor and its extant relatives from low to high. Numbers in squares 1-3 indicate degrees of genomic divergence in S. halepense and its relatives from low to high.

LIU et al. / Turk J Bot
21
be distinguished from the perennial rhizomatous S. propinquum by its broadly obovate sessile spikelets (Clayton and Renvoize, 1982; Chen and Phillips, 2006). Since the S. propinquum probe painted S. bicolor chromosomes with patchy signals (Figures 4 and 5), and a similar genome size was observed for S. bicolor (1.67 pg) and S. propinquum (1.52 pg) (Price et al., 2005), the degree of genomic divergence between them is low. By contrast, S. × drummondii is an interspecific hybrid between S. bicolor and S. arundinaceum, and therefore an ancestor-descendant relationship exists between S. arundinaceum and S. × drummondii. Being an ancient forest-savanna species native to tropical Africa (House, 1985), S. arundinaceum extends eastwards to India and Australia and has been introduced to the tropical Americas (Liu H et al., 2014). Sorghum × drummondii is native to an area extending from southern Egypt to Sudan (Hacker, 1992). Geographic distribution heterogeneity may be a predictor of genetic differentiation in Sorghum subg. Sorghum. This assumption is substantiated by allozyme studies, S. arundinaceum having a low level of genetic similarity (Nei’s genetic identity = 0.876) with S. × drummondii (Morden et al., 1990). The Sorghum arundinaceum probe painted the 20 S. × drummondii chromosomes with patchy signals (Figure 7), suggesting that the degree of genomic divergence between them is high. Therefore, it is reasonable to suppose that the degree of genomic divergence between S. bicolor and S. propinquum is less than that between S. arundinaceum and S. × drummondii (Figure 10).
Sorghum arundinaceum was assumed to be a wild progenitor of S. bicolor (De Wet, 1978). This hypothesis is supported by the allozyme and GISH data. Allozyme data show that there are genetic structural differences between S. arundinaceum and S. bicolor. The average number of alleles per locus for S. arundinaceum (3.2) is much higher than that observed for S. bicolor (1.8). This finding suggests that the genetic structure of S. bicolor has been modified since its origin (Morden et al., 1990). The apportionment of genetic variation between S. arundinaceum and S. bicolor is consistent with the GISH evidence. The labeled gDNA of S. arundinaceum chromosomes and that of other taxa (i.e. S. × almum or S. × drummondii or S. propinquum) hybridized with S. bicolor reveals that signals of the S. arundinaceum probe cover S. bicolor chromosomes with spotted or painted patterns, (Figures 1, 5, 7, and 9), indicating that an obvious degree of genomic divergence between S. arundinaceum and S. bicolor is evident. The crown age of Sorghum subg. Sorghum is estimated to be 8.6 million years ago, in the middle Miocene (Liu Q et al., 2014b). Both GISH and phylogenetic studies suggest that sorghum diverged from S. arundinaceum early in the diversification history of the subgenus Sorghum. This hypothesis is consistent with their
evident genetic variation (Morden et al., 1990; Liu Q et al., 2014b).
Several sorghum descendants, including S. × almum and S. × drummondii, may have had a relatively short phylogenetic history. Sorghum × almum is a recent fertile hybrid between S. bicolor and S. halepense (Doggett, 1976; Lazarides et al., 1991), and the species was hypothesized to have originated in Argentina (Celarier, 1958). Sorghum × drummondii was inferred to be an interspecific hybrid based on intermediate spikelet morphology between S. bicolor and S. halepense (De Wet, 1978), and it occurs in southern Egypt to Sudan (Morden, 1990; Mutegi et al., 2010). In allozyme studies, S. × almum had a relatively high level of genetic similarity (Nei’s genetic identity = 0.927) to S. × drummondii (Morden et al., 1990). Sorghum × almum and S. × drummondii probes hybridized with S. bicolor chromosomes with painted signals (Figure 2), supporting that the genome composition is homoeologous between S. bicolor and the probe species. The crown age of S. × almum and S. × drummondii was dated to the Pliocene to Pleistocene interval, which was considered to be a recent time frame in the evolution of Poaceae (Hoffmann et al., 2013). Sorghum bicolor exhibited a lower level of genomic divergence with the probe combinations containing the sorghum descendants (i.e. S. × almum or S. × drummondii) than with the probe combinations without sorghum descendant species (Figures 4 and 5), providing further evidence for the relatively short phylogenetic history of the sorghum descendants. Although the hybrids (S. × almum and S. × drummondii) may include the S. bicolor genome as one presumed paternal (or maternal) progenitor (Liu Q et al., 2014b), the distinct hybridization signals between different descendants and the common progenitor (Figure 2) revealed the repetitive sequences with genome-wide divergence across different descendant genomes by combinatorial interactions during speciation, as in previous findings (Swanson-Wagner et al., 2006; He et al., 2010).
A novel insight has been detected using the S. bicolor probe on the hybrids and the allopolyploid (Figures 1, 7, and 9), while results of hybrid probes’ spread on S. bicolor chromosomes provided unexpected results. For example, S. × almum has a mixture of genomes due to the presumption of a recent fertile hybrid between S. bicolor and S. halepense, and the hybridization signal intensity values from descendant species S. × almum probe were weaker than those from progenitor species A. arundinaceum probe, but the dispersion pattern was similarly painted, supporting that the genomic divergence between S. × almum and S. bicolor is less than the divergence between S. bicolor and S. arundinaceum at the level of repetitive sequences (Figure 1). Hypothesized differentiation among progenitor-descendant species has

LIU et al. / Turk J Bot
22
indeed been confirmed by GISH evidence; perhaps an intensive study with more population material of this case based on advanced tools of molecular cytogenetics would help quantify the differentiation degree between S. bicolor and its close relatives.
GISH results suggest that genomes of S. halepense and those of the closely related species have been well differentiated. The degrees of genomic differentiation for S. halepense-S. arundinaceum and S. halepense-S. bicolor are both less than the degree of genomic differentiation of S. halepense-S. propinquum. The red spotted signals of the S. arundinaceum probe are divided into two groups comprising the 17 strong signal chromosomes (red fluorescent intensity = 50.36–79.08) and the 23 weak signal chromosomes (red fluorescent intensity = 25.28–49.86) for S. halepense (Table S8; Figure 8f), whereas the red spotted signals of S. bicolor probe located on the 40 S. halepense chromosomes have relatively uniform signal intensity (18.74–53.59; Table S9; Figure 9f). Sorghum arundinaceum is hybridized to the wild progenitor of S. bicolor (Lazarides et al., 1991); this hypothesis is consistent with the results of nuclear DNA data, which placed S. arundinaceum as being closely related to S. bicolor with strong support (Liu Q et al., 2014b). It is appropriate to suggest that the degree of genomic divergence between S. halepense and S. bicolor is less than that between S. halepense and S. arundinaceum (a sorghum progenitor). Therefore, the degree of genomic divergence may be presented as the following ranked species pairs: S. halepense-S. bicolor < S. halepense-S. arundinaceum < S. halepense-S. propinquum (Figure 10).
The genomic origin of S. halepense has been debated for years. The GISH results reveal that the S. bicolor, S. arundinaceum, and S. propinquum probes hybridize to S. halepense chromosomes with different signal patterns (Figures 8f and 9f). The Sorghum bicolor probe showed spotted signals for the 40 S. halepense chromosomes, indicating less genetic distance between S. halepense and S. bicolor than between S. halepense and S. arundinaceum or S. propinquum (Tang and Liang, 1988). Based on the allozyme alleles, S. halepense is not well differentiated from S. bicolor (Morden et al., 1990), suggesting that S. halepense is of relatively recent origin from the parental species S.
bicolor. Although the placements of S. halepense differed in the nuclear gene trees, the species formed a clade with S. bicolor, S. arundinaceum, and S. propinquum (Liu Q et al., 2014b; Hawkins et al., 2015). The data available so far are consistent with the hypothesis that S. halepense may have arisen via a complex phylogenetic history involving several progenitor species (Paterson et al., 1995).
It has been suggested that repetitive sequences played an important role in genomic divergence leading to plant speciation (Paterson et al., 2009; Biscotti et al., 2015). Some repetitive sequences such as ribosomal DNA sequences are relatively conservative, while others are variable, especially in more distantly related species (Piednoël et al., 2013; Renney-Byfield et al., 2013). The evolutionary changes in repetitive sequences often result in diverged or novel variants, which can serve as species-specific DNA probes, providing important insights into phylogenetic relationships (Schmidt et al., 1998; Spangler et al., 1999; Nouzova et al., 2001). GISH data provide a novel and fine-scale understanding of the genomic relationships of S. bicolor and its extant close relatives in the context of repetitive sequences; therefore, the present study represents not only important progress to disentangle the interspecific relationships in Sorghum subg. Sorghum, but also practical assistance to meet geneticists’ and breeders’ demands in sorghum breeding.
AcknowledgmentsThis work was supported by the Chinese Academy of
Sciences President’s International Fellowship Initiative (no. 2016VBA010), the China Scholarship Council Awards (no. 201604910096), the National Natural Science Foundation of China (no. 31270275), the Special Basic Research Foundation of the Ministry of Science and Technology of the People’s Republic of China (no. 2013FY112100), and College Students’ Innovative Practice of the Chinese Academy of Sciences Training Plan (No.27). We thank ILRI-Addis Ababa and USDA-Beltsville Germplasm System for providing caryopses, Dr Kai Wang (Fujian Agriculture and Forestry University, China) for technical support, and the editor and reviewers for their constructive comments that improved the manuscript.
References
Biscotti MA, Olmo E, Heslop-Harrison JS (2015). Repetitive DNA in eukaryotic genomes. Chromosome Res 23: 415-420.
Carmona A, Friero E, de Bustos A, Jouve N, Cuadrado A (2013). The evolutionary history of sea barley (Hordeum marinum) revealed by comparative physical mapping of repetitive DNA. Ann Bot-London 112: 1845-1855.
Celarier RP (1958). Cytotaxonomic notes on the subsection Halepensia of the genus Sorghum. B Torrey Bot Club 85: 49-62.
Chen SL, Phillips SM (2006). Sorghum Moench. In: Wu ZY, Raven PH, editors. Flora of China, Vol. 22. Beijing, China: Science Press, pp. 602-604.
Clayton WD, Renvoize SA (1982). Gramineae (Part 3). In: Polhill RM, editor. Flora of Tropical East Africa. Rotterdam, the Netherlands: August Aimé Balkema, pp. 320-734.
Crow JF (1994). Anecdotal, historical and critical commentaries on genetics. Genetics 137: 891-894.

LIU et al. / Turk J Bot
23
Dai F, Chen ZH, Wang XL, Li ZF, Jin GL, Wu DZ, Cai SG, Wang N, Wu FB, Nevo E et al. (2004). Transcriptome profiling reveals mosaic genomic origins of modern cultivated barley. P Natl Acad Sci USA 111: 13403-13408.
De Wet JMJ (1978). Systematics and evolution of Sorghum sect. Sorghum (Gramineae). Am J Bot 65: 477-484.
Dodsworth S, Chase MW, Kelly LJ, Leitch IJ, Macas J, Novák P, Piednoël M, Weiss-Schneeweiss H, Leitch AR (2015). Genomic repeat abundances contain phylogenetic signal. Syst Biol 64: 112-126.
Doggett J (1970). Sorghum. London, UK: Green and Company, Longmans.
Doggett H (1976). Sorghum. In: Simmonds NW, editor. Evolution of Crop Plants. London, UK: Longman Scientific and Technical, pp. 112-117.
Doyle JJ, Doyle JL (1987). A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19: 11-15.
Hacker JB (1992). Sorghum × drummondii (Steud) Millsp. & Chase. In: Mannetje LT, Jones RM, editors. Plant Resources of South-East Asia. No. 4. Forages. Wageningen, Germany: Pudoc Scientific Publishers, pp. 206-208.
Hawkins JS, Ramachandran D, Henderson A, Freeman J, Carlise M, Harris A, Willison-Headley Z (2015). Phylogenetic reconstruction using four low-copy nuclear loci strongly supports a polyphyletic origin of the genus Sorghum. Ann Bot-London 116: 291-299.
He GM, Zhu XP, Elling AA, Chen LB, Wang XF, Guo L, Liang MZ, He H, Zhang HY, Chen FF et al. (2010). Global epigenetic and transcriptional trends among two rice subspecies and their reciprocal hybrids. Plant Cell 22: 17-33.
Heslop-Harrison JS (2000). Comparative genome organization in plants: from sequence and markers to chromatin and chromosomes. Plant Cell 12: 617-636.
Heslop-Harrison JS, Leitch AR, Schwarzacher T, Anamthawat-Jónsson K (1990). Detection and characterization of IB/IR translocations in hexaploid wheat. Heredity 65: 385-392.
Hoffmann MH, Schneider J, Hase P, Röser M (2013). Rapid and recent world-wide diversification of bluegrasses (Poa, Poaceae) and related genera. PLoS One 8: e60061.
House LR (1985). A Guide to Sorghum Breeding. Andhra, India: Pradesh Institute for the Semi-Arid Tropics.
Huang XH, Kurata N, Wei XH, Wang ZX, Wang AH, Zhao Q, Zhao Y, Liu KY, Lu HY, Li WJ et al. (2012). A map of rice genome variation reveals the origin of cultivated rice. Nature 490: 497-503.
Hunt HV, Badakshi F, Romanova O, Howe CJ, Jones MK, Heslop-Harrison JSP (2014). Reticulate evolution in Panicum (Poaceae): the origin of tetraploid broomcorn millet, P. miliaceum. J Exp Bot 65: 3165-3175.
Iourov IY, Soloviev IV, Vorsanova SG, Monakhov VV, Yurov YB (2005). An approach for quantitative assessment of fluorescence in situ hybridization (FISH) signals for applied human molecular cytogenetics. J Histochem Cytochem 53: 401-408.
Jiang B, Liu LQ, Hu XY, Liu Q (2012). Distribution of Eleusine indica AA genome on finger millet (Poaceae) chromosomes identified by genomic in situ hybridization and optimization method of probe length. J Trop Subtrop Bot 20: 44-50.
Katsiotis A, Loukas M, Heslop-Harrison JS (2000). Repetitive DNA, genome and species relationships in Avena and Arrhenatherum (Poaceae). Ann Bot-London 86: 1135-1142.
Lazarides M, Hacker JB, Andrew MH (1991). Taxonomy, cytology and ecology of indigenous Australian sorghums (Sorghum Moench, Andropogoneae, Poaceae). Aust Syst Bot 4: 591-635.
Li CB, Zhang DM, Ge S, Lu BR, Hong DY (2001a). Differentiation and inter-genomic relationships among C, E and D genomes in the Oryza officinalis complex (Poaceae) as revealed by multicolor genomic in situ hybridization. Theor Appl Genet 103: 197-203.
Li CB, Zhang DM, Ge S, Lu BR, Hong DY (2001b). Identification of genome constitution of Oryza malampuzhaensis, O. minuta, and O. punctata by multicolor genomic in situ hybridization. Theor Appl Genet 103: 204-211.
Liu H, Zeng FY, Liu Q (2014). Geographical distribution of Sorghum Moench (Poaceae). J Trop Subtrop Bot 22: 1-11.
Liu Q (2015). Research progress on structure and evolution of plant centromeres. J Trop Subtrop Bot 23: 576-586.
Liu Q, Jiang B, Wen J, Peterson PM (2014a). Low-copy nuclear gene and McGISH resolves polyploidy history of Eleusine coracana and morphological character evolution in Eleusine. Turk J Bot 38: 1-12.
Liu Q, Liu H, Wen J, Peterson PM (2014b). Infrageneric phylogeny and temporal divergence of Sorghum (Andropogoneae, Poaceae) based on low-copy nuclear and plastid sequences. PLoS One 9: e104933.
Liu Q, Triplett JM, Wen J, Peterson PM (2011). Allotetraploid origin and divergence in Eleusine (Chloridoideae, Poaceae): evidence from low-copy nuclear gene phylogenies and a plastid gene chronogram. Ann Bot-London 108: 1287-1298.
Marcussen T, Sandve SR, Heier L, Spannagl M, Pfeifer M, The International Wheat Genome Sequencing Consortium, Jakobsen KS, Wulff BBH, Steuernagel B, Mayer KF et al. (2014). Ancient hybridizations among the ancestral genomes of bread wheat. Science 345: 1250092.
Messing J (2009). The polyploidy origin of maize. In: Bennetzen JL, Hake S, editors. Maize Handbook, Vol. II: Genetics and Genomics. New York, NY, USA: Springer, pp. 221-238.
Morden CW, Doebley J, Schertz KF (1990). Allozyme variation among the spontaneous species of Sorghum section Sorghum (Poaceae). Theor Appl Genet 80: 296-304.
Mutegi E, Sagnard F, Muraya M, Kanyenji B, Rono B, Mwongera C, Marangu C, Kamau J, Parzies H, De Villiers S et al. (2010). Ecogeographical distribution of wild, weedy and cultivated Sorghum bicolor (L.) Moench in Kenya: implications for conservation and crop-to-wild gene flow. Genet Resour Crop Ev 57: 243-253.

LIU et al. / Turk J Bot
24
Ng’uni D, Geleta M, Fatih M, Bryngelsson T (2010). Phylogenetic analysis of the genus Sorghum based on combined sequence data from cpDNA regions and ITS generate well-supported trees with two major lineages. Ann Bot-London 105: 471-480.
Nouzova M, Neumann P, Navratilova A, Galbraith DW, Macas J (2001). Microarray-based survey of repetitive genomic sequences in Vicia spp. Plant Mol Biol 45: 229-244.
Paterson AH, Bowers JE, Bruggmann R, Dubchak I, Grimwood J, Gundlach H, Haberer G, Hellsten U, Mitros T, Poliakov A et al. (2009). The Sorghum bicolor genome and the diversification of grasses. Nature 457: 551-556.
Paterson AH, Bowers JE, Chapman BA (2004). Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. P Natl Acad Sci USA 101: 9903-9908.
Paterson AH, Schertz KF, Lin YR, Liu C, Chang YL (1995). The weediness of wild plants: molecular analysis of genes influencing dispersal and persistence of Johnsongrass, Sorghum halepense (L.) Pers. P Natl Acad Sci USA 92: 6127-6131.
Piednoël M, Carrete-Vega G, Renner SS (2013). Characterization of the LTR retrotransposon repertoire of a plant clade of six diploid and one tetraploid species. Plant J 75: 699-709.
Price HJ, Dillon SL, Hodnett G, Rooney WL, Ross L, Johnston JS (2005). Genome evolution in the genus Sorghum (Poaceae). Ann Bot-London 95: 219-227.
Schmidt T, Heslop-Harrison JS (1998). Genomes, genes and junk: the large-scale organization of plant chromosomes. Trends Plant Sci 3: 195-199.
Schwarzacher T, Heslop-Harrison P (2000). Practical in situ Hybridization. Oxford, UK: BIOS Scientific Publishers.
Spangler RE, Zaitchik B, Russo E, Kellogg EA (1999). Andropogoneae evolution and generic limits in Sorghum (Poaceae) using ndhF sequences. Syst Bot 24: 267-281.
Swanson-Wagner RA, Jia Y, DeCook R, Borsuk LA, Nettleton D, Schnable PS (2006). All possible modes of gene action are observed in a global comparison of gene expression in a maize F1 hybrid and its inbred parents. P Natl Acad Sci USA 103: 6805-6810.
Swigoňová Z, Lai JS, Ma JX, Ramakrishna W, Llaca V, Bennetzen JL, Messing J (2004). Close split of sorghum and maize genome progenitors. Genome Res 14: 1916-1923.
Tang H, Liang GH (1988). The genomic relationship between cultivated sorghum [Sorghum bicolor (L.) Moench] and Johnsongrass [S. halepense (L.) Pers.]: a re-evaluation. Theor Appl Genet 76: 277-284.

LIU et al. / Turk J Bot
1
Table S1. GISH signal intensity from mitotic metaphase chromosomes of Sorghum bicolor (blue) hybridized with biotin-labeled gDNA of S. × almum (green) and digoxigenin-labeled gDNA of S. arundinaceum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
19 89.41 ± 5.73 40.45 ±9.91 61.31 ± 11.12 89.41 19 45.14 1 64.82 1 0.80 11 1.36 1 1.01 17
4 88.99 ± 5.38 29.36 ± 8.99 47.61 ± 9.86 88.99 4 44.80 8 63.07 8 0.80 9 1.27 8 1.00 16
20 88.58 ± 6.68 35.98 ± 11.31 56.87 ± 12.5 88.58 20 42.40 11 61.31 19 0.71 8 1.23 2 0.97 6
5 88.24 ± 6.52 32.82 ± 9.70 50.02 ± 9.02 88.24 5 41.40 9 59.03 10 0.70 1 1.16 9 0.88 14
11 85.80 ± 8.24 42.40 ± 10.43 53.03 ± 8.9 85.8 11 40.45 19 58.63 2 0.66 19 1.16 10 0.86 18
8 85.14 ± 8.13 44.80 ± 12.92 63.07 ± 11.44 85.14 8 38.70 10 56.87 20 0.66 5 1.14 19 0.77 15
3 84.88 ± 6.72 33.08 ± 9.86 52.87 ± 10.85 84.88 3 37.74 2 53.03 11 0.66 10 1.11 11 0.76 7
10 84.04 ± 8.51 38.70 ± 9.29 59.03 ± 9.58 84.04 10 35.98 20 52.87 3 0.64 2 1.05 20 0.72 11
6 83.99 ± 7.78 17.66 ± 3.82 31.31 ± 4.54 83.99 6 33.08 3 51.93 9 0.64 18 1.01 3 0.71 4
17 83.61 ± 9.03 17.87 ± 4.74 30.62 ± 6.18 83.61 17 32.82 5 50.02 5 0.63 20 0.98 12 0.70 5
15 83.43 ± 10.08 25.79 ± 6.58 41.50 ± 9.04 83.43 15 29.36 4 48.46 12 0.63 3 0.94 5 0.68 9
7 83.00 ± 6.94 22.06 ± 3.92 39.20 ± 5.41 83 7 28.18 12 47.61 4 0.62 15 0.88 13 0.66 13
18 81.94 ± 8.63 23.73 ± 6.17 37.22 ± 6.49 81.94 18 25.79 15 43.47 13 0.62 4 0.86 4 0.62 3
1 80.61 ± 8.85 45.14 ± 14.33 64.82 ± 13.97 80.61 1 25.02 13 41.50 15 0.58 17 0.81 15 0.60 20
9 80.19 ± 9.09 41.40 ± 11.49 51.93 ± 10.85 80.19 9 23.73 18 39.20 7 0.58 12 0.74 18 0.59 12
16 79.45 ± 10.7 15.29 ± 3.47 27.99 ± 5.63 79.45 16 22.06 7 37.22 18 0.58 13 0.74 7 0.58 19
2 78.29 ± 8.73 37.74 ± 10.02 58.63 ± 10.28 78.29 2 17.87 17 31.31 6 0.56 6 0.63 14 0.56 10
13 78.13 ± 10.31 25.02 ± 6.44 43.47 ± 8.56 78.13 13 17.66 6 30.62 17 0.56 7 0.58 6 0.56 8
12 77.92 ± 8.25 28.18 ± 6.83 48.46 ± 10.49 77.92 12 16.47 14 29.67 14 0.56 14 0.58 17 0.52 2
14 73.54 ± 10.15 16.47 ± 3.22 29.67 ± 5.09 73.54 14 15.29 16 27.99 16 0.55 16 0.54 16 0.51 1

LIU et al. / Turk J Bot
2
Table S2. GISH signal intensity from mitotic metaphase chromosomes of Sorghum bicolor (blue) hybridized with biotin-labeled gDNA of S. × almum (green) and digoxigenin-labeled gDNA of S. × drummondii (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
7 56.41 ± 20.31 28.61 ± 10.71 47.92 ± 12.9 56.41 7 31.31 2 47.92 7 0.71 19 2.24 20 0.57 9
10 52.72 ± 20.15 25.00 ± 11.51 40.04 ± 13.10 52.72 10 29.37 1 45.20 1 0.69 2 1.97 19 0.51 15
3 52.17 ± 21.21 27.80 ± 11.06 41.75 ± 12.97 52.17 3 28.88 4 45.17 4 0.67 14 1.84 18 0.51 10
2 51.43 ± 17.35 31.31 ± 10.45 45.11 ± 10.86 51.43 2 28.61 7 45.11 2 0.67 16 1.81 5 0.50 3
8 50.32 ± 19.42 26.29 ± 11.64 41.73 ± 14.35 50.32 8 27.80 3 42.42 11 0.67 3 1.75 16 0.50 14
9 49.21 ± 22.22 20.51 ± 10.36 33.11 ± 11.22 49.21 9 27.19 19 41.75 3 0.65 1 1.66 1 0.47 13
14 48.79 ± 22.88 26.32 ± 11.46 39.26 ± 12.02 48.79 14 26.32 14 41.73 8 0.64 4 1.53 4 0.47 2
4 48.45 ± 18.73 28.88 ± 12.16 45.17 ± 13.83 48.45 4 26.29 8 40.74 5 0.64 6 1.53 6 0.47 8
11 47.61 ± 22.91 25.33 ± 11.46 42.42 ± 11.99 47.61 11 26.29 16 40.61 6 0.64 5 1.49 2 0.45 12
1 44.96 ± 16.42 29.37 ± 12.7 45.20 ± 14.26 44.96 1 25.95 6 40.04 10 0.64 20 1.42 11 0.45 17
12 43.98 ± 21.13 23.41 ± 12.28 37.55 ± 14.53 43.98 12 25.95 5 39.26 14 0.64 17 1.42 17 0.44 7
6 43.58 ± 18.76 25.95 ± 10.33 40.61 ± 11.21 43.58 6 25.33 11 39.26 16 0.63 8 1.39 12 0.42 11
15 40.35 ± 19.10 18.97 ± 10.47 30.38 ± 11.70 40.35 15 25.00 10 38.38 20 0.62 15 1.36 7 0.42 6
13 40.00 ± 17.61 20.50 ± 9.76 32.88 ± 9.64 40.00 13 24.39 20 38.34 19 0.62 10 1.35 8 0.42 4
16 37.54 ± 15.21 26.29 ± 12.30 39.26 ± 12.65 37.54 16 23.41 12 37.55 12 0.62 18 1.34 14 0.39 1
17 37.09 ± 20.84 20.41 ± 12.60 32.13 ± 15.76 37.09 17 20.51 9 33.11 9 0.62 13 1.33 13 0.38 16
5 36.84 ± 14.37 25.95 ± 10.87 40.74 ± 13.11 36.84 5 20.50 13 32.88 13 0.62 12 1.33 3 0.36 19
19 33.18 ± 14.96 27.19 ± 13.23 38.34 ± 14.65 33.18 19 20.41 17 32.13 17 0.62 9 1.23 10 0.35 5
20 28.02 ± 11.14 24.39 ± 13.76 38.38 ± 14.76 28.02 20 18.97 15 30.38 15 0.60 11 1.22 15 0.34 18
18 21.50 ± 8.46 15.17 ± 8.14 24.33 ± 10.70 21.50 18 15.17 18 24.33 18 0.60 7 1.09 9 0.28 20

LIU et al. / Turk J Bot
3
Table S3. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum bicolor (blue) hybridized with biotin-labeled gDNA of S. × drummondii (green) and digoxigenin-labeled gDNA of S. arundinaceum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
14 74.90 ± 8.99 21.57 ± 6.53 23.58 ± 10.88 74.90 14 32.07 5 47.99 6 1.37 1 1.18 5 2.97 3
13 74.41 ± 9.15 18.30 ± 6.37 14.25 ± 6.25 74.41 13 31.75 6 47.06 5 1.28 13 1.13 6 2.94 13
12 73.82 ± 10.07 23.07 ± 7.83 28.71 ± 10.81 73.82 12 25.94 20 30.14 20 1.21 3 1.06 20 2.80 1
6 70.67 ± 8.61 31.75 ± 7.90 47.99 ± 15.16 70.67 6 24.26 18 29.79 18 1.16 2 1.01 18 2.16 15
15 69.76 ± 9.72 18.59 ± 8.32 16.93 ± 10.19 69.76 15 23.66 8 28.71 12 1.11 10 0.99 8 1.63 4
3 67.80 ± 13.34 15.09 ± 7.74 12.48 ± 8.38 67.80 3 23.64 2 25.12 8 1.10 15 0.96 9 1.63 10
17 67.45 ± 8.91 21.38 ± 7.48 21.66 ± 11.40 67.45 17 23.07 12 25.07 9 1.03 4 0.79 7 1.56 2
5 67.26 ± 9.07 32.07 ± 6.55 47.06 ± 16.16 67.26 5 22.04 19 23.58 14 1.02 11 0.77 19 1.55 17
1 62.47 ± 9.01 17.63 ± 6.82 12.89 ± 7.37 62.47 1 22.03 9 22.12 19 1.01 7 0.74 2 1.52 11
10 59.91 ± 9.09 21.39 ± 8.21 19.34 ± 9.37 59.91 10 21.57 14 21.66 17 1.00 19 0.70 12 1.52 14
4 59.31 ± 11.03 19.08 ± 7.13 18.48 ± 9.71 59.31 4 21.39 10 20.30 2 0.99 17 0.68 10 1.51 16
2 59.03 ± 6.10 23.64 ± 8.12 20.30 ± 9.40 59.03 2 21.38 17 19.39 7 0.96 16 0.67 11 1.30 19
19 57.46 ± 5.54 22.04 ± 5.22 22.12 ± 9.47 57.46 19 19.64 7 19.34 10 0.94 8 0.64 17 1.28 7
11 57.29 ± 7.51 19.41 ± 6.48 19.01 ± 8.60 57.29 11 19.41 11 19.01 11 0.91 14 0.63 4 1.15 12
16 55.95 ± 12.09 17.33 ± 7.24 18.1 ± 10.05 55.95 16 19.08 4 18.48 4 0.88 9 0.63 16 0.95 8
18 53.49 ± 6.22 24.26 ± 7.12 29.79 ± 9.60 53.49 18 18.59 15 18.10 16 0.86 20 0.60 14 0.91 9
20 52.72 ± 6.19 25.94 ± 7.91 30.14 ± 14.42 52.72 20 18.30 13 16.93 15 0.81 18 0.51 15 0.81 20
8 49.28 ± 4.69 23.66 ± 7.57 25.12 ± 11.67 49.28 8 17.63 1 14.25 13 0.80 12 0.49 1 0.81 18
7 49.19 ± 3.82 19.64 ± 7.51 19.39 ± 7.68 49.19 7 17.33 16 12.89 1 0.68 5 0.44 13 0.59 6
9 48.89 ± 5.24 22.03 ± 8.18 25.07 ± 10.40 48.89 9 15.09 3 12.48 3 0.66 6 0.41 3 0.58 5

LIU et al. / Turk J Bot
4
Table S4. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum bicolor (blue) hybridized with biotin-labeled gDNA of S. arundinaceum (green) and digoxigenin-labeled gDNA of S. propinquum. (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
17 64.54 ± 12.07 22.92 ± 8.24 38.38 ± 8.99 64.54 17 38.75 2 53.82 2 0.84 15 3.03 20 1.16 5
15 55.61 ± 12.44 31.81 ± 11.39 37.88 ± 14.26 55.61 15 37.66 1 51.99 3 0.84 18 2.36 1 0.67 15
2 55.57 ± 6.28 38.75 ± 8.78 53.82 ± 10.04 55.57 2 36.16 3 49.36 1 0.83 13 2.33 10 0.63 18
5 52.34 ± 13.08 14.47 ± 7.42 19.30 ± 9.53 52.34 5 31.81 15 41.41 12 0.82 10 1.87 12 0.63 17
16 51.73 ± 13.87 25.76 ± 13.36 35.56 ± 13.26 51.73 16 31.52 12 39.86 4 0.79 6 1.72 7 0.61 16
3 51.51 ± 11.40 36.16 ± 12.16 51.99 ± 11.93 51.51 3 31.09 4 38.94 6 0.78 4 1.71 3 0.60 14
4 47.46 ± 13.78 31.09 ± 12.80 39.86 ± 14.09 47.46 4 30.73 6 38.38 17 0.76 1 1.67 2 0.54 8
14 43.97 ± 10.01 23.57 ± 8.95 31.27 ± 10.29 43.97 14 30.15 13 37.88 15 0.76 12 1.60 6 0.52 4
6 43.48 ± 11.78 30.73 ± 13.55 38.94 ± 16.26 43.48 6 29.23 10 37.27 19 0.76 19 1.60 13 0.52 13
13 41.74 ± 11.10 30.15 ± 10.38 36.48 ± 12.56 41.74 13 28.16 19 36.48 13 0.75 14 1.57 19 0.50 11
19 41.66 ± 10.33 28.16 ± 8.57 37.27 ± 11.55 41.66 19 25.76 16 36.16 7 0.75 5 1.50 11 0.49 6
11 39.40 ± 10.00 25.31 ± 10.74 33.81 ± 11.25 39.40 11 25.31 11 35.74 10 0.75 11 1.49 4 0.48 19
12 38.94 ± 9.15 31.52 ± 12.42 41.41 ± 15.06 38.94 12 23.97 7 35.56 16 0.72 16 1.45 9 0.43 2
18 38.11 ± 7.17 23.17 ± 7.45 27.63 ± 7.21 38.11 18 23.57 14 33.81 11 0.72 2 1.33 18 0.41 3
1 36.86 ± 10.14 37.66 ± 11.09 49.36 ± 14.15 36.86 1 23.17 18 32.41 9 0.70 3 1.25 15 0.41 12
8 36.5 ± 8.81 16.78 ± 7.50 26.34 ± 8.70 36.50 8 22.92 17 31.27 14 0.66 7 1.25 14 0.39 9
9 35.02 ± 6.36 18.41 ± 7.77 32.41 ± 9.38 35.02 9 18.41 9 27.63 18 0.64 8 1.19 16 0.38 7
7 34.86 ± 8.66 23.97 ± 8.12 36.16 ± 11.9 34.86 7 16.78 8 26.96 20 0.60 17 1.18 8 0.35 10
10 27.86 ± 9.13 29.23 ± 11.19 35.74 ± 13.80 27.86 10 14.47 5 26.34 8 0.57 9 0.95 17 0.32 1
20 13.61 ± 4.71 14.30 ± 7.40 26.96 ± 5.12 13.61 20 14.30 20 19.30 5 0.53 20 0.65 5 0.17 20

LIU et al. / Turk J Bot
5
Table S5. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum bicolor (blue) hybridized with biotin-labeled gDNA of S. propinquum (green) and digoxigenin-labeled gDNA of S. arundinaceum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
17 53.12 ± 6.42 24.10 ± 13.52 19.78 ± 6.49 53.12 17 39.37 20 54.51 10 1.23 19 1.94 12 1.86 19
18 51.75 ± 8.00 7.81 ± 7.22 17.42 ± 9.56 51.75 18 39.24 10 48.82 1 1.22 17 1.93 7 1.47 17
2 51.56 ± 4.69 22.01 ± 6.89 32.11 ± 9.79 51.56 2 34.40 6 46.46 20 0.97 5 1.87 11 1.43 6
8 51.06 ± 6.55 39.24 ± 15.45 54.51 ± 14.30 51.06 8 33.96 14 43.24 17 0.89 20 1.84 8 1.34 5
1 49.98 ± 6.91 30.53 ± 13.37 36.25 ± 15.23 49.98 1 31.22 16 39.14 18 0.88 11 1.76 9 0.92 18
3 47.94 ± 6.15 30.77 ± 12.72 35.06 ± 11.83 47.94 3 30.88 8 38.46 6 0.88 3 1.74 10 0.84 13
19 46.8 ± 7.26 17.13 ± 9.09 13.91 ± 6.30 46.80 19 30.77 18 36.31 3 0.85 15 1.51 20 0.78 15
7 45.77 ± 4.67 39.37 ± 13.51 48.82 ± 15.26 45.77 7 30.53 15 36.25 11 0.84 1 1.39 4 0.74 14
9 45.73 ± 5.36 33.96 ± 13.84 46.46 ± 16.10 45.73 9 28.75 2 35.23 15 0.82 13 1.37 3 0.65 2
16 45.64 ± 7.33 24.29 ± 12.60 36.31 ± 13.93 45.64 16 27.30 7 35.06 12 0.81 7 1.34 1 0.64 3
15 45.52 ± 7.90 22.9 ± 10.56 26.86 ± 10.71 45.52 15 24.29 17 34.53 2 0.80 12 1.33 16 0.63 1
4 44.53 ± 5.16 27.3 ± 11.60 34.53 ± 12.81 44.53 4 24.10 1 32.11 9 0.79 6 1.09 15 0.59 20
20 43.95 ± 6.88 31.22 ± 13.34 35.23 ± 12.42 43.95 20 22.90 4 26.86 4 0.79 4 1.05 2 0.57 4
14 41.46 ± 5.23 18.79 ± 9.28 24.4 ± 7.35 41.46 14 22.01 13 24.40 14 0.77 14 1.04 14 0.50 16
10 41.42 ± 5.61 28.75 ± 9.91 43.24 ± 15.44 41.42 10 18.79 11 19.78 8 0.73 9 0.98 13 0.47 11
6 39.97 ± 5.49 9.76 ± 5.36 12.33 ± 6.35 39.97 6 17.13 12 19.76 5 0.72 8 0.83 17 0.42 7
11 39.27 ± 4.51 34.40 ± 13.91 39.14 ± 14.09 39.27 11 16.25 19 17.42 7 0.69 2 0.72 5 0.42 9
5 36.95 ± 3.75 13.13 ± 6.55 13.58 ± 6.53 36.95 5 13.13 5 13.91 16 0.67 16 0.66 19 0.41 12
13 36.78 ± 5.54 16.25 ± 7.66 19.76 ± 7.15 36.78 13 9.76 3 13.58 19 0.66 10 0.55 6 0.39 8
12 35.79 ± 5.24 30.88 ± 17.76 38.46 ± 16.42 35.79 12 7.81 9 12.33 13 0.45 18 0.49 18 0.38 10

LIU et al. / Turk J Bot
6
Table S6. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum arundinaceum (blue) hybridized with biotin-labeled gDNA of S. × drummondii (green) and digoxigenin-labeled gDNA of S. × almum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
16 72.29 ± 13.24 54.58 ± 16.47 49.09 ± 12.96 72.29 16 61.22 6 55.55 18 1.28 9 2.36 13 0.82 10
19 69.24 ± 16.77 53.59 ± 16.02 49.92 ± 14.78 69.24 19 60.85 18 54.04 6 1.26 5 2.30 9 0.80 20
1 67.60 ± 16.40 58.60 ± 17.64 49.22 ± 14.94 67.6 1 58.88 17 53.68 17 1.26 7 2.04 11 0.78 16
2 64.97 ± 18.17 56.33 ± 19.17 46.31 ± 13.13 64.97 2 58.6 1 52.43 4 1.26 8 1.99 7 0.77 2
18 64.74 ± 16.65 60.85 ± 17.80 55.55 ± 15.31 64.74 18 57.16 4 50.61 11 1.25 10 1.93 6 0.75 5
20 64.36 ± 13.65 48.00 ± 12.78 42.68 ± 9.60 64.36 20 57.06 7 49.92 19 1.22 2 1.88 14 0.75 1
3 63.84 ± 17.03 52.92 ± 15.87 47.16 ± 14.07 63.84 3 56.8 11 49.22 1 1.19 1 1.83 8 0.72 12
17 62.79 ± 18.20 58.88 ± 16.84 53.68 ± 16.77 62.79 17 56.33 2 49.09 16 1.19 13 1.80 18 0.72 19
4 61.07 ± 18.41 57.16 ± 19.62 52.43 ± 16.71 61.07 4 55.14 9 47.16 3 1.14 12 1.79 4 0.72 15
6 59.79 ± 19.76 61.22 ± 21.55 54.04 ± 17.04 59.79 6 54.58 16 46.31 2 1.13 6 1.79 17 0.72 3
5 58.25 ± 18.25 54.47 ± 17.39 43.16 ± 13.62 58.25 5 54.47 5 45.41 7 1.12 20 1.68 5 0.69 8
15 57.59 ± 17.85 47.79 ± 18.25 42.50 ± 10.61 57.59 15 53.59 19 43.37 13 1.12 15 1.59 1 0.63 7
10 54.82 ± 13.29 46.35 ± 20.60 37.00 ± 12.60 54.82 10 52.92 3 43.23 9 1.12 11 1.58 2 0.61 17
11 52.6 ± 15.08 56.80 ± 18.92 50.61 ± 15.61 52.6 11 51.62 13 43.16 5 1.12 3 1.58 12 0.61 18
7 51.49 ± 14.00 57.06 ± 19.57 45.41 ± 14.79 51.49 7 49.94 8 42.68 20 1.12 14 1.57 15 0.61 4
8 49.01 ± 15.16 49.94 ± 17.95 39.77 ± 13.30 49.01 8 48 20 42.5 15 1.11 16 1.57 3 0.60 14
14 47.74 ± 16.02 47.40 ± 20.53 42.25 ± 16.04 47.74 14 47.79 15 42.25 14 1.10 17 1.52 10 0.59 6
12 45.25 ± 15.95 38.01 ± 14.94 33.35 ± 9.96 45.25 12 47.4 14 39.77 8 1.10 18 1.49 19 0.56 9
9 42.84 ± 13.47 55.14 ± 17.55 43.23 ± 14.06 42.84 9 46.35 10 37 10 1.09 4 1.43 16 0.55 11
13 40.22 ± 15.09 51.62 ± 20.65 43.37 ± 16.85 40.22 13 38.01 12 33.35 12 1.07 19 1.41 20 0.50 13

LIU et al. / Turk J Bot
7
Table S7. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum × drummondii (blue) hybridized with biotin-labeled gDNA of S. bicolor (green) and digoxigenin-labeled gDNA of S. arundinaceum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
19 77.83 ± 9.25 35.49 ± 23.21 43.01 ± 9.16 77.83 19 51.15 9 58.4 8 1.16 12 1.70 9 1.04 14
17 76.45 ± 10.61 36.92 ± 21.57 48.09 ± 13.07 76.45 17 46.4 10 57.99 9 1.05 10 1.67 7 0.90 12
20 74.11 ± 11.38 31.38 ± 16.93 37.81 ± 11.05 74.11 20 44.75 12 56.91 7 1.04 6 1.46 8 0.89 20
13 73.65 ± 10.43 34.57 ± 20.24 42.12 ± 11.41 73.65 13 44.75 6 55.11 5 1.03 14 1.46 6 0.83 16
18 72.77 ± 13.06 39.79 ± 21.54 50.75 ± 10.82 72.77 18 43.35 7 52.71 4 0.88 9 1.44 11 0.82 19
2 72.34 ± 11.04 35.48 ± 21.07 48.88 ± 14.10 72.34 2 41.94 11 50.75 18 0.83 11 1.43 10 0.79 13
14 72.02 ± 13.74 36.42 ± 23.05 35.32 ± 13.38 72.02 14 39.79 18 50.25 11 0.83 20 1.38 4 0.73 10
5 69.79 ± 14.28 30.27 ± 11.57 55.11 ± 7.55 69.79 5 39 8 49.76 15 0.83 19 1.29 1 0.71 6
15 69.54 ± 12.82 29.69 ± 16.46 49.76 ± 10.83 69.54 15 37.5 4 48.88 2 0.82 13 1.28 12 0.69 17
3 68.87 ± 15.36 32.05 ± 19.95 46.29 ± 11.98 68.87 3 36.92 17 48.09 17 0.81 16 1.24 18 0.63 18
16 67.60 ± 15.78 29.24 ± 13.68 36.29 ± 15.75 67.6 16 36.42 14 46.29 3 0.78 18 1.22 5 0.62 2
8 66.69 ± 14.49 39.00 ± 19.10 58.4 ± 10.35 66.69 8 35.49 19 44.62 1 0.78 1 1.17 2 0.61 3
4 65.23 ± 14.54 37.50 ± 26.25 52.71 ± 13.34 65.23 4 35.48 2 44.13 10 0.77 17 1.14 15 0.60 1
12 64.96 ± 16.31 44.75 ± 25.11 38.62 ± 16.83 64.96 12 34.74 1 43.04 6 0.76 7 1.14 3 0.58 11
9 64.18 ± 12.47 51.15 ± 22.81 57.99 ± 15.90 64.18 9 34.57 13 43.01 19 0.73 2 1.11 17 0.52 15
11 63.94 ± 14.60 41.94 ± 21.88 50.25 ± 11.90 63.94 11 32.05 3 42.12 13 0.71 4 1.04 13 0.52 9
10 63.20 ± 12.42 46.4 ± 25.62 44.13 ± 18.03 63.2 10 31.38 20 38.62 12 0.69 3 1.01 19 0.51 4
1 61.62 ± 14.45 34.74 ± 19.03 44.62 ± 15.55 61.62 1 30.27 5 37.81 20 0.67 8 1.00 14 0.46 8
7 60.17 ± 13.82 43.35 ± 23.47 56.91 ± 12.72 60.17 7 29.69 15 36.29 16 0.60 15 0.97 16 0.46 7
6 60.15 ± 14.00 44.75 ± 25.98 43.04 ± 18.29 60.15 6 29.24 16 35.32 14 0.55 5 0.93 20 0.45 5

LIU et al. / Turk J Bot
8
Table S8. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum halepense (blue) hybridized with biotin-labeled gDNA of S. propinquum (green) and digoxigenin-labeled gDNA of S. arundinaceum (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
18 76.83 ± 11.43 51.30 ± 23.48 73.6 ± 11.03 76.83 18 72.88 4 79.08 4 0.96 2 2.05 23 0.60 3
4 75.32 ± 11.07 72.88 ± 17.31 79.08 ± 7.75 75.32 4 57.38 5 73.6 18 0.93 31 2.02 4 0.60 14
7 73.64 ± 11.85 22.55 ± 8.66 53.07 ± 9.23 73.64 7 51.3 18 70.12 5 0.92 4 1.93 25 0.60 15
5 73.14 ± 14.68 57.38 ± 23.71 70.12 ± 15.18 73.14 5 49.8 25 67 23 0.89 10 1.75 19 0.59 36
12 71.68 ± 14.77 22.86 ± 13.23 40.73 ± 7.48 71.68 12 46.94 35 62.42 17 0.89 32 1.74 5 0.58 32
34 71.41 ± 18.62 23.83 ± 12.23 40.02 ± 10.03 71.41 34 45.49 10 61.84 25 0.88 1 1.64 35 0.58 9
33 70.27 ± 18.30 29.96 ± 14.06 45.36 ± 14.14 70.27 33 45.28 16 60.84 19 0.87 35 1.63 18 0.58 30
15 70.21 ± 14.34 19.73 ± 9.68 43.42 ± 9.43 70.21 15 44.74 23 58.35 16 0.84 28 1.60 16 0.57 27
14 69.81 ± 15.55 19.92 ± 12.32 45.56 ± 7.64 69.81 14 39.35 38 54.76 20 0.82 5 1.54 17 0.57 8
38 66.61 ± 18.36 39.35 ± 24.90 50.36 ± 17.76 66.61 38 38.97 31 53.79 35 0.81 25 1.54 10 0.57 2
9 66.15 ± 17.99 17.82 ± 10.91 30.85 ± 10.31 66.15 9 38.94 6 53.07 7 0.78 38 1.52 20 0.56 7
39 65.63 ± 16.95 16.63 ± 13.41 37.64 ± 9.74 65.63 39 35.45 17 52.84 13 0.78 6 1.50 28 0.56 29
8 65.16 ± 18.06 27.45 ± 15.14 46.68 ± 15.47 65.16 8 35.05 19 51.12 10 0.78 16 1.49 26 0.56 24
16 64.61 ± 18.30 45.28 ± 27.62 58.35 ± 16.78 64.61 16 34.97 20 50.36 38 0.76 26 1.47 31 0.56 21
37 64.32 ± 17.16 12.04 ± 9.99 29.70 ± 9.97 64.32 37 34.43 40 49.86 6 0.71 29 1.39 6 0.56 33
40 63.96 ± 18.14 34.43 ± 22.13 49.80 ± 18.66 63.96 40 34.02 28 49.8 40 0.70 3 1.35 38 0.54 13
6 63.92 ± 17.48 38.94 ± 14.64 49.86 ± 16.57 63.92 6 30.4 2 47.55 24 0.70 18 1.32 40 0.54 34
1 63.42 ± 18.68 26.31 ± 16.33 29.76 ± 15.84 63.42 1 29.96 33 46.68 8 0.69 40 1.24 21 0.54 35
13 63.41 ± 17.68 25.03 ± 15.09 52.84 ± 11.88 63.41 13 29.83 26 45.56 14 0.67 23 1.23 13 0.54 18
17 63.40 ± 16.91 35.45 ± 16.11 62.42 ± 13.59 63.4 17 28.8 32 45.36 33 0.66 33 1.18 24 0.53 39
10 62.81 ± 19.91 45.49 ± 29.27 51.12 ± 22.25 62.81 10 27.45 8 45.14 21 0.66 11 1.18 29 0.53 12
3 62.02 ± 11.94 23.97 ± 10.06 34.28 ± 11.10 62.02 3 26.31 1 43.42 15 0.64 27 1.14 8 0.53 22
35 61.33 ± 21.50 46.94 ± 30.15 53.79 ± 20.05 61.33 35 25.9 29 41.98 31 0.64 20 1.11 32 0.53 11
36 61.16 ± 20.25 11.02 ± 9.16 30.74 ± 9.10 61.16 36 25.03 13 40.73 12 0.60 34 1.07 33 0.53 17
30 59.78 ± 19.39 14.14 ± 9.72 31.19 ± 8.63 59.78 30 24.25 21 40.34 28 0.59 8 1.04 2 0.53 40
24 59.64 ± 18.66 23.03 ± 13.73 47.55 ± 14.67 59.64 24 23.97 3 40.02 34 0.58 9 1.03 7 0.53 16
2 59.64 ± 17.17 30.40 ± 18.37 31.62 ± 14.58 59.64 2 23.83 34 39.07 26 0.58 19 1.01 27 0.52 10
20 59.06 ± 18.97 34.97 ± 20.74 54.76 ± 15.52 59.06 20 23.03 24 37.64 39 0.57 17 0.94 3 0.52 28
25 57.99 ± 17.44 49.80 ± 23.62 61.84 ± 16.50 57.99 25 22.86 12 36.52 29 0.56 12 0.94 14 0.52 26
21 55.84 ± 15.54 24.25 ± 12.22 45.14 ± 12.99 55.84 21 22.55 7 34.28 3 0.54 21 0.90 15 0.52 38
11 55.50 ± 16.85 18.79 ± 13.56 28.56 ± 9.92 55.5 11 19.92 14 32.39 32 0.48 24 0.89 34 0.52 31
32 55.37 ± 20.83 28.80 ± 27.55 32.39 ± 20.10 55.37 32 19.73 15 31.62 2 0.47 13 0.89 12 0.51 20
31 54.91 ± 20.26 38.97 ± 27.99 41.98 ± 22.49 54.91 31 18.79 11 31.19 30 0.45 15 0.88 1 0.51 1
19 54.89 ± 14.91 35.05 ± 13.19 60.84 ± 10.51 54.89 19 18.06 27 30.85 9 0.45 30 0.85 11 0.51 6
23 54.57 ± 11.76 44.74 ± 19.63 67.00 ± 12.09 54.57 23 17.82 9 30.74 36 0.44 39 0.83 39 0.51 5
29 52.98 ± 17.24 25.90 ± 15.77 36.52 ± 15.90 52.98 29 16.63 39 29.76 1 0.44 14 0.76 30 0.51 19
28 49.72 ± 17.87 34.02 ± 28.01 40.34 ± 20.04 49.72 28 14.14 30 29.7 37 0.42 7 0.74 9 0.49 37
22 49.65 ± 15.48 8.12 ± 5.82 25.28 ± 10.64 49.65 22 12.04 37 28.56 11 0.41 37 0.68 36 0.48 25
26 46.29 ± 15.15 29.83 ± 16.79 39.07 ± 15.63 46.29 26 11.02 36 28.03 27 0.36 36 0.67 22 0.47 23
27 45.82 ± 20.63 18.06 ± 12.39 28.03 ± 11.83 45.82 27 8.12 22 25.28 22 0.32 22 0.65 37 0.46 4

LIU et al. / Turk J Bot
9
Table S9. GISH fluorescent signal intensity from mitotic metaphase chromosomes of Sorghum halepense (blue) hybridized with biotin-labeled gDNA of S. propinquum (green) and digoxigenin-labeled gDNA of S. bicolor (red).
Chr
omos
ome
no.
Blue
flu
ores
cent
inte
nsity
Gre
enflu
ores
cent
inte
nsity
Red
fluor
esce
ntin
tens
ity
Blue
fluo
resc
ent i
nten
sity
of S
. bico
lor
chro
mos
omes
in F
igur
e 1
Chr
omos
ome
no.
Gre
en fl
uore
scen
t int
ensit
y of
S. ×
alm
um p
robe
sign
als i
n Fi
gure
1
Chr
omos
ome
no.
Red
fluor
esce
nt in
tens
ity o
f S. a
rund
inac
eum
pr
obe
signa
ls in
Fig
ure
1
Chr
omos
ome
no.
Gre
en/R
ed fl
uore
scen
t int
ensit
yin
Fig
ure
1
Chr
omos
ome
no.
(Gre
en+r
ed)/
blu
e flu
ores
cent
Inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
(Gre
en/r
ed)/
[(gr
een+
red)
/blu
e]flu
ores
cent
inte
nsity
in F
igur
e 1
Chr
omos
ome
no.
26 78.48 ± 9.16 22.64 ± 8.11 53.59 ± 6.66 78.48 26 26.44 27 53.59 26 0.63 1 1.79 31 0.97 7
10 77.73 ± 10.01 24.02 ± 11.60 44.23 ± 11.90 77.73 10 25.69 25 52.39 27 0.61 34 1.53 20 0.83 39
32 73.64 ± 15.87 17.86 ± 10.06 52.38 ± 15.13 73.64 32 24.27 21 52.38 32 0.59 20 1.42 18 0.74 6
13 71.68 ± 13.39 18.33 ± 10.63 42.96 ± 11.20 71.68 13 24.02 10 50.21 2 0.59 31 1.37 33 0.69 35
8 70.80 ± 13.69 20.76 ± 9.10 44.54 ± 12.34 70.8 8 23.85 2 48.35 24 0.59 39 1.37 30 0.65 38
24 69.82 ± 12.19 23.58 ± 11.70 48.35 ± 10.79 69.82 24 23.58 24 46.69 9 0.58 35 1.36 21 0.62 10
11 69.37 ± 14.87 9.23 ± 8.57 35.22 ± 9.01 69.37 11 22.64 26 46.22 21 0.56 25 1.30 22 0.60 5
12 67.76 ± 14.24 17.61 ± 14.10 43.79 ± 11.54 67.76 12 21.58 18 45.83 25 0.56 5 1.28 17 0.59 4
25 65.93 ± 21.54 25.69 ± 7.99 45.83 ± 9.19 65.93 25 21.49 22 45.58 18 0.56 23 1.25 27 0.58 34
2 65.66 ± 17.33 23.85 ± 8.63 50.21 ± 12.97 65.66 2 21.33 9 44.54 8 0.54 10 1.22 29 0.54 1
6 65.16 ± 14.78 13.26 ± 8.48 28.11 ± 10.31 65.16 6 21.22 1 44.47 33 0.54 38 1.22 19 0.52 37
40 64.82 ± 16.83 19.75 ± 11.62 42.9 ± 10.29 64.82 40 20.76 8 44.23 10 0.53 7 1.17 1 0.52 25
9 64.38 ± 12.75 21.33 ± 7.61 46.69 ± 10.23 64.38 9 20.75 17 43.79 12 0.53 21 1.16 3 0.51 23
27 63.08 ± 16.18 26.44 ± 11.96 52.39 ± 12.14 63.08 27 20.31 33 42.96 13 0.52 16 1.15 16 0.51 8
39 59.18 ± 12.68 15.61 ± 9.01 26.51 ± 8.57 59.18 39 20.24 23 42.9 40 0.52 37 1.13 2 0.50 36
14 57.72 ± 20.04 14.61 ± 11.32 35.98 ± 14.66 57.72 14 20.1 3 42.54 3 0.51 28 1.09 23 0.50 13
36 57.16 ± 19.66 18.96 ± 12.64 38.04 ± 15.11 57.16 36 19.75 40 42.03 22 0.51 17 1.08 25 0.49 28
15 55.74 ± 19.25 18.96 ± 10.50 38.67 ± 13.27 55.74 15 19.15 16 40.52 17 0.51 22 1.07 34 0.48 40
5 54.27 ± 16.18 18.06 ± 9.99 32.27 ± 12.96 54.27 5 18.96 36 38.67 15 0.50 27 1.06 9 0.47 15
3 54.04 ± 18.40 20.10 ± 6.64 42.54 ± 9.03 54.04 3 18.96 15 38.04 36 0.50 29 1.05 28 0.47 24
4 53.77 ± 20.01 15.06 ± 7.66 30.42 ± 9.53 53.77 4 18.7 20 36.74 16 0.50 30 1.03 15 0.46 14
7 52.56 ± 18.32 9.88 ± 6.20 18.74 ± 8.82 52.56 7 18.33 13 36.22 23 0.50 36 1.03 24 0.45 16
23 51.87 ± 16.93 20.24 ± 8.53 36.22 ± 11.43 51.87 23 18.06 5 36.05 19 0.50 4 1.00 37 0.44 12
21 51.70 ± 19.38 24.27 ± 10.26 46.22 ± 12.74 51.7 21 17.86 32 35.98 14 0.49 15 1.00 36 0.43 26
35 49.73 ± 15.56 15.50 ± 9.73 26.56 ± 11.87 49.73 35 17.61 12 35.22 11 0.49 24 0.97 26 0.43 9
22 49.01 ± 19.80 21.49 ± 13.35 42.03 ± 14.81 49.01 22 17.42 31 34.22 29 0.48 2 0.97 40 0.42 2
16 48.75 ± 19.65 19.15 ± 9.55 36.74 ± 14.12 48.75 16 17.16 29 33.76 1 0.47 18 0.95 32 0.41 29
37 48.18 ± 19.35 16.52 ± 9.49 31.82 ± 12.79 48.18 37 16.52 37 32.27 5 0.47 3 0.93 5 0.41 11
17 48.01 ± 14.80 20.75 ± 11.39 40.52 ± 14.39 48.01 17 15.97 28 31.82 37 0.47 6 0.92 8 0.41 3
38 47.65 ± 14.11 13.98 ± 6.96 25.78 ± 10.89 47.65 38 15.61 39 31.56 20 0.47 8 0.91 12 0.40 27
33 47.23 ± 21.08 20.31 ± 9.13 44.47 ± 13.28 47.23 33 15.56 30 31.17 28 0.46 40 0.88 10 0.40 17
18 47.18 ± 17.82 21.58 ± 11.35 45.58 ± 13.60 47.18 18 15.5 35 31.15 30 0.46 9 0.88 14 0.39 22
1 47.10 ± 17.87 21.22 ± 8.17 33.76 ± 13.90 47.1 1 15.06 4 30.42 4 0.46 33 0.86 13 0.39 20
28 44.89 ± 15.35 15.97 ± 8.51 31.17 ± 11.17 44.89 28 14.61 14 29.51 31 0.43 13 0.85 4 0.39 21
29 41.97 ± 15.49 17.16 ± 8.98 34.22 ± 12.70 41.97 29 14.28 19 28.11 6 0.42 26 0.85 35 0.37 30
19 41.28 ± 18.76 14.28 ± 11.33 36.05 ± 13.96 41.28 19 13.99 34 26.56 35 0.41 14 0.83 38 0.36 32
34 34.36 ± 16.77 13.99 ± 11.89 22.75 ± 13.42 34.36 34 13.98 38 26.51 39 0.40 12 0.71 39 0.33 33
30 34.19 ± 15.18 15.56 ± 10.44 31.15 ± 13.92 34.19 30 13.26 6 25.78 38 0.40 19 0.64 11 0.33 18
20 32.78 ± 13.86 18.70 ± 10.61 31.56 ± 10.18 32.78 20 9.88 7 22.75 34 0.34 32 0.63 6 0.33 31
31 26.29 ± 11.95 17.42 ± 10.87 29.51 ± 12.05 26.29 31 9.23 11 18.74 7 0.26 11 0.54 7 0.32 19




![Sorghum bicolor (L.) Moench n Tohum ve Tohum Unu...[Sorghum bicolor (L.) Moench’in tohum, tohum ununun güvenilirliği] GKGM - Risk Değerlendirme Daire Başkanlığı, 2016 2 değeri](https://img.pdfslide.net/doc/110x75/60b9ad92d792884b1f35c195/sorghum-bicolor-l-moench-n-tohum-ve-tohum-unu-sorghum-bicolor-l-moenchain.jpg)

![THE POTENTIAL OF SWEET SORGHUM [Sorghum bicolor (L · The Potential of Sweet Sorghum [Sorghum Bicolor (L.) Moench] As A Bio- Resource for Syrup and Ethanol Production in Kenya EVANS](https://img.pdfslide.net/doc/110x75/5f0f52697e708231d44395b9/the-potential-of-sweet-sorghum-sorghum-bicolor-l-the-potential-of-sweet-sorghum.jpg)


![RESEARCH Open Access Genotypic variation in sorghum [Sorghum … · 2018-06-03 · RESEARCH Open Access Genotypic variation in sorghum [Sorghum bicolor (L.) Moench] exotic germplasm](https://img.pdfslide.net/doc/110x75/5f03a3af7e708231d40a0b15/research-open-access-genotypic-variation-in-sorghum-sorghum-2018-06-03-research.jpg)
![BAB 2 TINJAUAN PUSTAKA 2.1 Sorgum (Sorghum bicolor L. …eprints.umg.ac.id/283/3/[3] BAB 2.pdf · 4 BAB 2 TINJAUAN PUSTAKA 2.1 Sorgum (Sorghum bicolor L. Moench) Sorgum (Sorghum bicolor](https://img.pdfslide.net/doc/110x75/5d33713688c993bb3c8ce077/bab-2-tinjauan-pustaka-21-sorgum-sorghum-bicolor-l-3-bab-2pdf-4-bab-2-tinjauan.jpg)

![Quantificação de amido e proteínas totais em grãos de sorgo€¦ · Quantificação de amido e proteínas totais em grãos de sorgo [Sorghum bicolor (L.) Moench - Família: Poaceae]](https://img.pdfslide.net/doc/110x75/5fd1df498ca9665adf6eea3f/quantificao-de-amido-e-protenas-totais-em-gros-de-sorgo-quantificao.jpg)