Embed Size (px)
Citation preview

Yeast 15, 1511–1518 (1999)
Replicative Ageing in the Fission YeastSchizosaccharomyces pombe
M. G. BARKER1* AND R. M. WALMSLEY1
1Department of Biomolecular Sciences, UMIST, PO Box 88, Manchester M60 1QD, UK
Saccharomyces cerevisiae has been widely used as a model organism in studies of replicative ageing and senescence.The relevance of these studies to ageing in other organisms has, however, been questioned, since this yeast dividesby budding rather than fission, the more common pattern in higher organisms. Here we report that, contrary topopular belief, the fission yeast Schizosaccharomyces pombe also undergoes replicative senescence and in a mannersuperficially analogous to budding yeast. These experiments provide the first evidence of age asymmetry in cellfission and are consistent with the hypothesis of Jazwinski, that asymmetric division underlies culture immortality.Given their evolutionary divergence, comparison of the ageing determinants in fission and budding yeasts may helpidentify common mechanisms of the ageing process. Copyright ? 1999 John Wiley & Sons, Ltd.
*Correspondence to: M. G. Barker, Department of Bio-molecular Sciences, UMIST, PO Box 88, Manchester
INTRODUCTION
Mortimer and Johnston (1959) first demonstratedthat the unicellular budding yeast Saccharomycescerevisiae undergoes replicative senescence: afterabout 20 divisions individual cells die. This obser-vation was possible because, in the budding modeof cell division, the mother cell is readily dis-tinguished from the daughter. Generally each newdaughter cell has a full replicative lifespan anddaughters rarely inherit the age of their mothersunless they have very old mothers (Egilmez andJazwinski, 1989; Kennedy et al., 1994). Thus,whilst the culture is immortal, the individuals arenot. Subsequently, Hayflick (1965) demonstratedthat cultured mammalian fibroblasts also possessa limited proliferative capacity; however, in thiscase ageing is clonal—all cells in a culture die atapproximately the same time, suggesting thatfollowing cell division both cells inherit the sameage. Calleja et al. (1980) demonstrated that inSchizosaccharomyces pombe, fission scar segrega-tion between ‘sibs’ generated from a multi-scarredcell is equally likely to increase or decrease the scarnumber of the most scarred cell. This led theauthors to conclude that immortal fission yeastcultures, lacking such ‘mother–daughter’ distinc-tion in scar segregation, are comprised of immortal
M60 1QD, UK. E-mail: [email protected]
CCC 0749–503X/99/141511–08$17.50Copyright ? 1999 John Wiley & Sons, Ltd.
cells. To date this view has not been challenged,although work with S. cerevisiae has led Jazwinski(1993) to hypothesize that ‘asymmetric reproduc-tion (division) of yeast cells lies at the foundationof yeast ageing and yeast longevity’. He goes on tospeculate that mammalian cell lines follow thesame rules: asymmetric division maintaining thetotipotency of germ cell and stem cell lines, due tosome ‘unequal distribution of resources’. Thismode of division then switches to symmetricdivision in differentiated somatic cell lines (whichneed only possess a limited proliferative potential),to allow the ‘generation of populations ofequipotent, functional somatic cells’. Given theevolutionary conservation in eukaryotic cell cycleregulation (Wuarin and Nurse, 1996, and refer-ences therein), we speculated as to whether someform of asymmetric reproduction might alsounderlie culture immortality in S. pombe cells. Anumber of studies have demonstrated asymmetryin fission yeast division with regard to mating typeswitching (Egel and Eie, 1987; Grewal and Klar,1997). Furthermore, the molecular localization ofthe Cdc7 protein kinase, which regulates the initi-ation of septum formation, is localized asymmetri-cally to only one of the spindle poles duringanaphase B (Sohrmann et al., 1998). Interestingly,a slight asymmetry (4–10%) in septum locationwith respect to cell length has also been reported(Johnson et al., 1979; Miyata et al., 1986; May and
Mitchison, 1995). In this study, however, we haveReceived 20 February 1999Accepted 30 June 1999

1512 M. G. BARKER AND R. M. WALMSLEY
demonstrated progressive asymmetry, first in celldiameter and later in cell volume, followingrepeated divisions of individual S. pombe cells.Like budding yeast and animals cells (Barker andSmart, 1996; Hayflick, 1965; Jazwinski, 1993;Mortimer and Johnston, 1959), S. pombe increasein size with replicative age. From pedigree analysiswe have demonstrated that this distinction con-tinues to a replicative age at which cells die. Theseresults support the evolutionary ‘trade-off ’ theoriespromulgated by Kirkwood and Rose (1991).
Definitions of terms used in this manuscriptThe term ‘replicative age’ corresponds to the
number of divisions an individual cell has com-pleted. It is related to the Hayflick limit of mam-malian cell lines, which refers to the maximumnumber of divisions a cell/cell line can accomplish.It is different from the chronological age of a cell,which is measured in hours/days, etc. Chronologi-cal and replicative age are largely independent,even in yeast (Muller et al., 1980).
The following nomenclature was used in thetracking of individual cells during pedigree analy-sis: upon identification of a visibly asymmetricdivision, the terms ‘M0’ and ‘Ds’ are employed,referring to the sib with greater and lesser diameterat septation, respectively. The M0 cell maintainsthis nomenclature through subsequent divisionsand is assumed to be older than the Ds cells itgenerates. By analogy with ageing in buddingyeast, the M0 sib corresponds to the original‘mother’ cell, dragged from the cell streak. Giventhis assumption, the M0 cell inherits a replicativeage equivalent to 1+the number of cell doublingsobserved. For mortality analysis, the Ds sibs (or‘daughter sibs’) are discarded, since it is assumedtheir replicative age is reset to zero (by analogywith budding yeast, they correspond to ‘virgindaughter’ cells).
MATERIALS AND METHODS
Strains and growth conditionsThe wild-type S. pombe strain NCYC132 was
used in this study since it has been used in a pre-vious study with implications for ageing (Callejaet al., 1980). The lifespan determination exper-iments were performed at 25)C during the day andthe cells visually inspected every 2 h to monitorgrowth. Overnight the cells were incubated at 8)Cto limit growth. All media components were
Copyright ? 1999 John Wiley & Sons, Ltd.
supplied by Oxoid. Nutrient-agar contained: 2%(w/v) peptone, 1% (w/v) yeast extract, 1·2% (w/v)agar and 2% (w/v) glucose. Cell manipulationswere performed using a manual Tetrad-dissector(Singer). Other details regarding cell manipulationsare within the body of the text. Cell volume calcu-lations were based on those of Miyata et al. (1986)and performed using Vernier hand callipers onenlarged photographs taken at 800# magnifi-cation using an Olympus camera mounted on aZeiss microscope.
RESULTS
Cells were selected at random from a 2 day-old cellstreak and dragged to a fresh sector of the nutrientagar by micromanipulation. After two doublings,the four progeny generated from the majority oforiginal cells were visually indistinguishable,although in about a quarter of the groups, one cellhad a greater diameter. Two different types ofmanipulations were then carried out on the identi-cal groups of cells. In the first series of exper-iments, the two outer cells were discarded(arrowed in Figure 1) by dragging them to oneedge of the agar slab, and the two inner cells wereallowed to double again. In most of these newgroups of four cells, one cell could be visuallydistinguished from the other three due to itsgreater diameter and shorter length (arrowed inFigure 2). Where the cells were indistinguishable,all four were separated and a further round ofdivision was permitted. In all cases, one of thefour pairs of cells showed an asymmetrical divisionwith respect to diameter. Once identified, thisasymmetry persisted through subsequent divisions:one of the sibs, termed ‘M0’ (see definitions,above), would maintain the larger diameter of theparent cell, whilst the other, termed ‘Ds’, woulddisplay a normal (narrower) average diameter. Inthe second series of experiments, the outer cellswere observed. Following two divisions, the cellswere indistinguishable and again the groups offour cells were separated and allowed to divide.When these separated cells had gone throughtwo divisions, each giving rise to a further fourprogeny, one cell was again distinguishable due toits greater diameter. These two strategies, whichwere pursued with more than 50 isolated cells ineach case, both led to the same conclusion: thatasymmetric division is a visible property of allS. pombe cells within approximately four divisions
Yeast 15, 1511–1518 (1999)

1513REPLICATIVE AGEING IN S. POMBE
Figure 1. Removal of the distal cell-pair from the groupof four progeny generated after two divisions. Cells to bediscarded are marked by arrows. The internal cell pair(unmarked) would subsequently be monitored through furtherrounds of division, until a cell with a greater diameter than itssibs could be unambiguously identified.
Copyright ? 1999 John Wiley & Sons, Ltd.
Figure 2. (A) and (B) Isolation of cells visually perceived topossess a greater diameter (marked with arrows) after 3–4divisions. The unmarked cells would be discarded at this stage.
of ‘birth’ and that this phenotype then persiststhrough subsequent rounds of division.
We hypothesized that these ‘M0’ cells (the sibswith the greater diameter at septation) representedolder cells. Applying the conventions of buddingyeast ageing to fission yeast, the M0 cells corre-sponded to ‘mother’ cells, whilst the narrower Dscells represented ‘virgin’ daughter cells. Giventhese assumptions, an explanation of why a minor-ity of the originally selected cells were able togive rise to asymmetrical divisions within twodoublings becomes readily available. In S.cerevisiae, randomly selected cells have a mean ageof one division: half the cells in a culture arevirgins, a quarter have divided once, an eighthhave divided twice, etc. (giving a mean age of onedivision for the culture as a whole). Thus, in theoriginal selection of S. pombe cells, the minoritywhich gave rise to non-identical groups of four areconcluded to have arisen from cells which hadalready divided one or more times. Havingdemonstrated asymmetric cell division and theability to follow pedigrees, we then had a methodto determine whether individual cells were indeedimmortal.
M0 cells with increased diameters (48 in total),identified by the method above, were moved tonew co-ordinates on the agar surface and againallowed to divide. At the same time a control cell(a ‘normal’ cell selected at random), was moved so
as to appear in the same field of view. This wasto ensure that the process of manipulation wasnot causing any of the observed phenomena. Theasymmetry in S. pombe division became moreand more apparent with subsequent divisions(Figure 3). As the M0 cell increased in replicativeage, the difference in size between M0 and Ds sibsbecame more obvious (Figure 4). Following eachdivision, the M0 cell was separated and moved tonew coordinates to prevent the depletion ofnutrients in the cell’s local environment and herprogeny discarded (by dragging to one edge of theagar). In parallel with this procedure, a cell fromthe control group was also selected and positionedadjacent to the M0 cell. Continual dissection of thecontrol cells (or the Ds ‘daughter sibs’ of M0 cells)led to the predicted appearance of asymmetrywithin four doublings. There was no evidence of abias in the appearance of this asymmetry—singlemanipulated cells, as well as those observed within
Yeast 15, 1511–1518 (1999)

1514 M. G. BARKER AND R. M. WALMSLEY
Figure 3. The emergence of obvious asymmetry in celldiameter during fission yeast division. The M0 cell on the rightgave rise to the cell in the centre (first generation progeny),which in turn gave rise to the cell on the left (second generationprogeny). The diameter of the M0 cell (five divisions old) isclearly greater than that of the cell to the left, which is about todivide for the first time. The central cell is the Ds sib from theM0 cell’s first division and has subsequently been through fourrounds of division itself. The asymmetry in cell diameter duringgrowth by terminal extension of the central cell is obvious andcontrasts with that of its previous sib to the left. The majority ofthe progeny generated by the central Ds cell and the M0 cell tothe right have been discarded.
Figure 4. Asymmetry in a five divisions-old S. pombe cellabout to divide. The M0 sib to the left of the septum (arrow) hasinherited the greater diameter of the parent cell, in markedcontrast to its Ds sib to the right of the septum. The cell pair tothe right of the photograph belong to another pedigree andshould be ignored.
unmanipulated microcolonies, followed the simplerules defined above.
With subsequent cell divisions, the increase inM0 cell diameter was accompanied by a morpho-logical change to an ever more spheroidal shape(Figure 5a): rather like the ‘round bottomflask’ (RBF) morphology of cdc10-121 h" cells,described by Miyata et al. (1986), following treat-ment with the antifungal Aculeacin-A. This drug
Copyright ? 1999 John Wiley & Sons, Ltd.
inhibits â-glucan synthase (Yamaguchi et al.,1982) and at higher concentrations disrupts cellwall formation, causing cell lysis (Miyata et al.,1980). A preliminary study of cell volume atseptation, calculated for 10 individual cells only(with a mean age of approximately 8 divisions),confirmed the visual impression that the M0 cellvolume increased with age. The volume ratiobetween sibs, normally 1:1 (Johnson et al., 1979),was over 1·5:1 in those M0:Ds pairs measured(mean 1·63&0·5). A larger study was not per-formed because every photograph of an ageing S.pombe cell, from which the cell volume calculationswere made, represented a sacrifice of data from themortality analysis (it was necessary to place acoverslip onto the agar surface in order to obtain aclear image of the cells, precluding further micro-manipulation). Interestingly, the only otherreported deviation from the 1:1 volume ratiobetween sibs is the Aculeacin-A study cited above(Miyata et al., 1986). The older M0 cells werestill capable of producing normal-looking Ds‘daughter’ cells (Figure 5b), although the oldest ofthem would sometimes give rise to spheroidal firstgeneration sibs that in turn would divide, produc-ing normal, sausage-shaped, second generation‘daughters’ (Figure 5c). This apparent phenotypiccarry-over (division producing no normal sib)could persist to the second division before anormal cell was produced. As the pedigree contin-ued, a marked increase in division time wasobserved, apparent for example in Figure 5c,where the Ds (first generation) sib has undergone adivision before its M0 sib. A similar phenomenonis encountered during ageing in S. cerevisiae(Barker and Smart, 1996; Egilmez and Jazwinski,1989; Mortimer and Johnston, 1959).
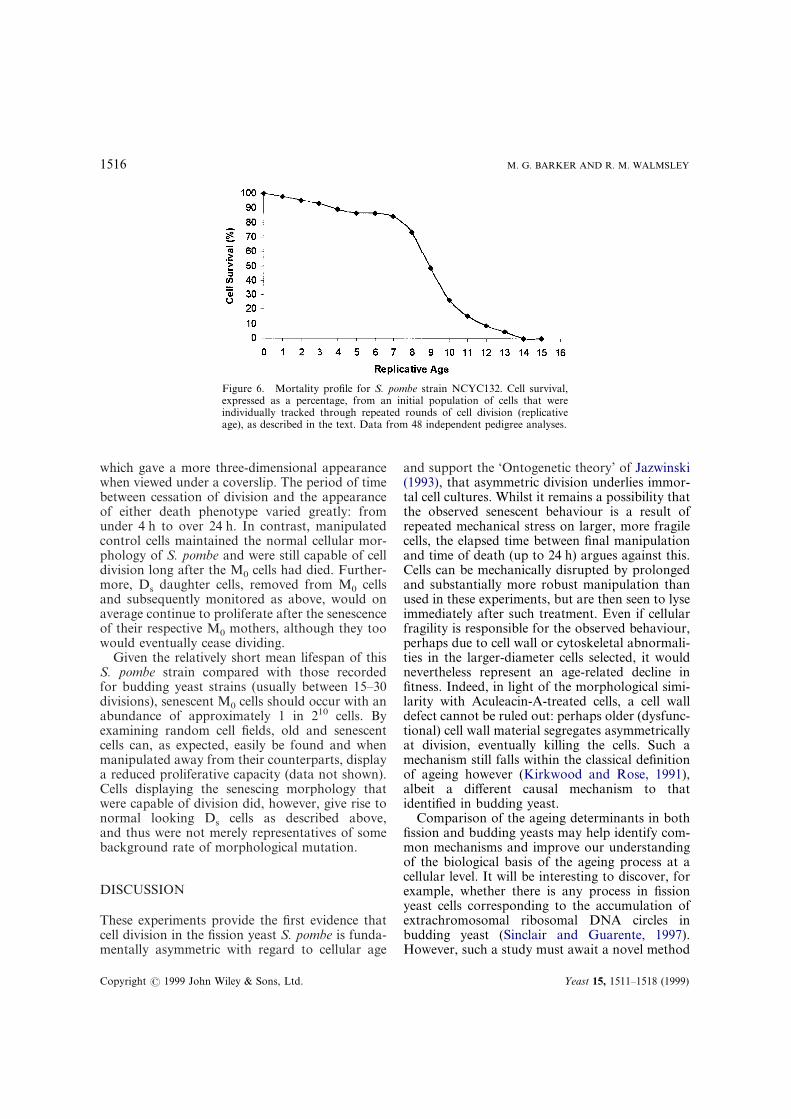
Eventually all the cells senesced and the mor-tality profile (calculated for 48 cells) followed theexpected Gompertz distribution (Figure 6), givinga mean lifespan of 9·2&2·5 divisions for thisstrain. The greatest number of divisions observedwas 14, and the least was 2. The most long-livedcells displayed the most abnormal (spheroidal)morphology, although over 50% of cells eventuallyassumed an RBF-like appearance. Two distinctdeath phenotypes were evident. The most drasticphenotype (Figure 7a and b) was cell lysis, occur-ring in approximately 30% of M0 cells, that wouldoften leave a collapsed cell ‘ghost’ from which thecell contents could clearly be seen to have spilledout onto the agar surface. The other phenotype(Figure 8a and b) was a solidified ‘cast-like’ cell,
Yeast 15, 1511–1518 (1999)

1515REPLICATIVE AGEING IN S. POMBE
Figure 5. Rounded morphology of senescing S. pombe. (A) Nine divisionsold M0 cell undergoing septation. The M0 sib (arrow) displays the charac-teristic spheroidal morphology of an ageing cell, which has not beeninherited by its Ds sibs. (B) An old spheroidal M0 ‘mother’ cell, giving riseto a more normal looking Ds ‘daughter sib’. The cell above and to the rightis a Ds sib from a previous division. (C) First and second generation progenyfrom an ageing spheroidal M0 cell, displaying the gradual return to thenormal ‘sausage-shaped’ morphology of S. pombe. The Ds sib (first gener-ation) has already performed a new round of cell division to produce thesecond generation sib since the M0 cell last divided, illustrating the cell cycledelay that occurs in extremely old cells. The unlabelled cell to the right is arandomly selected control cell.
Copyright ? 1999 John Wiley & Sons, Ltd. Yeast 15, 1511–1518 (1999)
1516 M. G. BARKER AND R. M. WALMSLEY
which gave a more three-dimensional appearancewhen viewed under a coverslip. The period of timebetween cessation of division and the appearanceof either death phenotype varied greatly: fromunder 4 h to over 24 h. In contrast, manipulatedcontrol cells maintained the normal cellular mor-phology of S. pombe and were still capable of celldivision long after the M0 cells had died. Further-more, Ds daughter cells, removed from M0 cellsand subsequently monitored as above, would onaverage continue to proliferate after the senescenceof their respective M0 mothers, although they toowould eventually cease dividing.
Given the relatively short mean lifespan of thisS. pombe strain compared with those recordedfor budding yeast strains (usually between 15–30divisions), senescent M0 cells should occur with anabundance of approximately 1 in 210 cells. Byexamining random cell fields, old and senescentcells can, as expected, easily be found and whenmanipulated away from their counterparts, displaya reduced proliferative capacity (data not shown).Cells displaying the senescing morphology thatwere capable of division did, however, give rise tonormal looking Ds cells as described above,and thus were not merely representatives of somebackground rate of morphological mutation.
Figure 6. Mortality profile for S. pombe strain NCYC132. Cell survival,expressed as a percentage, from an initial population of cells that wereindividually tracked through repeated rounds of cell division (replicativeage), as described in the text. Data from 48 independent pedigree analyses.
DISCUSSION
These experiments provide the first evidence thatcell division in the fission yeast S. pombe is funda-mentally asymmetric with regard to cellular age
Copyright ? 1999 John Wiley & Sons, Ltd.
and support the ‘Ontogenetic theory’ of Jazwinski(1993), that asymmetric division underlies immor-tal cell cultures. Whilst it remains a possibility thatthe observed senescent behaviour is a result ofrepeated mechanical stress on larger, more fragilecells, the elapsed time between final manipulationand time of death (up to 24 h) argues against this.Cells can be mechanically disrupted by prolongedand substantially more robust manipulation thanused in these experiments, but are then seen to lyseimmediately after such treatment. Even if cellularfragility is responsible for the observed behaviour,perhaps due to cell wall or cytoskeletal abnormali-ties in the larger-diameter cells selected, it wouldnevertheless represent an age-related decline infitness. Indeed, in light of the morphological simi-larity with Aculeacin-A-treated cells, a cell walldefect cannot be ruled out: perhaps older (dysfunc-tional) cell wall material segregates asymmetricallyat division, eventually killing the cells. Such amechanism still falls within the classical definitionof ageing however (Kirkwood and Rose, 1991),albeit a different causal mechanism to thatidentified in budding yeast.
Comparison of the ageing determinants in bothfission and budding yeasts may help identify com-mon mechanisms and improve our understandingof the biological basis of the ageing process at acellular level. It will be interesting to discover, forexample, whether there is any process in fissionyeast cells corresponding to the accumulation ofextrachromosomal ribosomal DNA circles inbudding yeast (Sinclair and Guarente, 1997).However, such a study must await a novel method
Yeast 15, 1511–1518 (1999)

1517REPLICATIVE AGEING IN S. POMBE
of isolating age-synchronous cell populations,since some cell wall material is distributed betweensibs during S. pombe division—precluding thebiotin-labelling techniques employed in Guarente’slaboratory (Sinclair and Guarente, 1997).
It is interesting to note that these experimentsprovide data that accommodate the behaviour ofS. pombe into the evolutionary ‘trade-off’ theoriespromulgated by Kirkwood and Rose (1991). Suchtheories posit that from a finite energy budget,maximum fitness is achieved if the balance ofenergy invested in cellular maintenance and repro-duction is tipped in favour of more reproductionat the expense of maintenance. There is thus
Copyright ? 1999 John Wiley & Sons, Ltd.
insufficient maintenance for individual cellularimmortality, but an immortal and evolutionarily‘fit’ population is produced.
ACKNOWLEDGEMENTS
Thanks to Steve Oliver for the loan of his ageingSinger micromanipulator, to Katherine Smart forproviding space in Oxford Brookes for some ofthis work and to Judy Barrow for advising a virginS. pombe worker.
Figure 7. Cell death phenotype 1. Following a prolongedsenescent phase, approximately 30% of cells would abruptlylyse. (A) A recently lysed spheroidal M0 cell (arrow), with a‘halo’ of spilled cellular material visible around its lower half.Its Ds sib displays an abnormal long-celled phenotype. (B) Alysed M0 cell in an advanced stage of decomposition (arrow)and its surviving Ds sib (adjacent), displaying a normal mor-phology. The other cells (below and to the right) belong tounrelated pedigrees and should be ignored.
Figure 8. Cell death phenotype 2. The second death pheno-type was a gradual assumption of a solidified ‘cast-like’ appear-ance. (A) An eight-divisions-old senescent S. pombe cell(arrow). The rougher, more three-dimensional morphology ofthe senescent cell contrasts with the group of cells from anunrelated pedigree to the right. (B) A senescent M0 cell (arrow)and spheroidal Ds sib, above. The unmarked spheroidal cell tothe right of the photograph belongs to an unrelated pedigree.
REFERENCES
Barker, M. G. and Smart, K. A. (1996). Morphologicalchanges associated with the cellular aging of a brew-ing yeast strain. J. Am. Soc. Brew. Chem. 54(2),121–126.
Yeast 15, 1511–1518 (1999)

1518 M. G. BARKER AND R. M. WALMSLEY
Calleja, G. B., Zuker, M., Johnson, B. F. and Yoo, B. Y.(1980). Analyses of fission scars as permanent recordsof cell division in Schizosaccharomyces pombe.J. Theor. Biol. 84, 523–544.
Egel, R. and Eie, B. (1987). Cell lineage asymmetry inSchizosaccharomyces pombe—unilateral transmissionof a high-frequency state for mating-type switching indiploid pedigrees. Curr. Genet 12(6), 429–433.
Egilmez, N. K. and Jazwinski, S. M. (1989). Evidencefor the involvement of a cytoplasmic factor in theaging of the yeast Saccharomyces cerevisiae. J.Bacteriol. 171, 37–42.
Grewal, S. I. S. and Klar, A. J. S. (1997). A recombina-tionally repressed region between mat2 and mat3 locishares homology to centromeric repeats and regulatesdirectionality of mating-type switching in fissionyeast. Genetics 146(4), 1221–1238.
Hayflick, L. (1965). The limited in vitro lifetime ofhuman diploid cell strains. Exp. Cell Res. 37, 614–636.
Jazwinski, S. M. (1993). The genetics of aging in theyeast Saccharomyces cerevisiae. Genetica 91, 35–51.
Johnson, B. F., Calleja, G. B., Boisclair, I. and Yoo,B. Y. (1979). Cell division in yeasts. III. The biased,asymmetric location of the septum in the fission yeastcell, Schizosaccharomyces pombe. Exp. Cell Res. 123,253–259.
Kennedy, B. K., Austriaco, N. R. and Guarente, L.(1994). Daughter cells of Saccharomyces cerevisiaefrom old mothers display a reduced life span. J. CellBiol. 127, 1985–1993.
Kirkwood, T. B. L. and Rose, M. R. (1991). Evol-ution of senescence: late survival sacrificed for
Copyright ? 1999 John Wiley & Sons, Ltd.
reproduction. Phil. Trans. R. Soc. Lond. Ser. B 332,15–24.
May, J. W. and Mitchison, J. M. (1995). Pattern ofpolar extension of the cell wall in the fission yeastSchizosaccharomyces pombe. Can. J. Microbiol. 41,273–277.
Miyata, M., Kitamura, J. and Miyata, H. (1980). Lysisof growing fission-yeast cells induced by aculeacin A,a new antifungal antibiotic. Arch. Microbiol. 127,11–16.
Miyata, M., Miyata, H. and Johnson, B. F. (1986).Asymmetric location of the septum in morphologi-cally altered cells of the fission yeast Schizosaccharo-myces pombe. J. Gen. Microbiol. 132, 883–891.
Mortimer, R. K. and Johnston, J. R. (1959). Life span ofindividual yeast cells. Nature 183, 1751–1752.
Muller, I., Zimmermann, M., Becker, D. and Flomer,M. (1980). Calendar life span versus budding life spanof Saccharomyces cerevisiae. Mech. Ageing Dev. 12,47–52.
Sinclair, D. A. and Guarente, L. (1997). Extrachromo-somal rDNA circles—a cause of aging in yeast. Cell91(7), 1033–1042.
Sohrmann, M., Schmidt, S., Hagan, I. and Simanis, V.(1998). Asymmetric segregation on spindle poles ofthe Schizosaccharomyces pombe septum-inducingprotein kinase Cdc7p. Genes Dev. 12(1), 84–94.
Wuarin, J. and Nurse, P. (1996). Regulating S phase:CDKs, licensing and proteolysis. Cell 85, 785–787.
Yamaguchi, H., Hiratani, T., Iwata, K. and Yamamoto,Y. (1982). Studies on the mechanism of antifungalaction of aculeacin A. J. Antibiotics 35, 210–219.
Yeast 15, 1511–1518 (1999)





























