Embed Size (px)
Citation preview

JGOFS REPORT No. 35
REPORT of the INDIAN OCEAN SYNTHESIS GROUP
on the ARABIAN SEA PROCESS STUDY

Published in Bergen, Norway, January 2002 by:
Scientific Committee on Oceanic Research and JGOFS International Project OfficeDepartment of Earth and Planetary Sciences Centre for Studies of Environment and ResourcesThe Johns Hopkins University University of BergenBaltimore, MD 21218 5020 BergenUSA NORWAY
The Joint Global Ocean Flux Study of the Scientific Committee on Oceanic Research (SCOR) is a Core Project of the International Geosphere-Biosphere Programme (IGBP). It is planned by aSCOR/IGBP Scientific Steering Committee. In addition to funds from the JGOFS sponsors,SCOR and IGBP, support is provided for international JGOFS planning and synthesis activitiesby several agencies and organizations. These are gratefully acknowledged and include the USNational Science Foundation, the International Council of Scientific Unions (by funds from theUnited Nations Education, Scientific and Cultural Organization), the IntergovernmentalOceanographic Commission, the Research Council of Norway and the University of Bergen,Norway.
Citation: Report of the Indian Ocean Synthesis Group on the Arabian Sea Process Study. January 2002
ISSN: 1016-7331
Cover: JGOFS and SCOR Logos
The JGOFS Reports are distributed free of charge to scientists involved in global change research.
Additional copies of the JGOFS reports are available from:
Ms. Judith R. Stokke, Administrative Assistant Tel: (+47) 55 58 42 46JGOFS International Project Office Fax: (+47) 55 58 96 87Centre for Studies of Environment and Resources E-mail: [email protected] of BergenHigh-Technology CentreN-5020 Bergen, NORWAY
or, from our website: http://ads.smr.uib.no/jgofs/jgofs.htm

JOINT GLOBAL OCEAN FLUX STUDY
– JGOFS –
REPORT No. 35
REPORT of the INDIAN OCEAN SYNTHESIS GROUP
on the ARABIAN SEA PROCESS STUDY

i
Editorial Group
Louisa Watts
Peer Review Committee Secretary - Terrestrial SciencesTerrestrial and Freshwater Sciences TeamNatural Environment Research CouncilPolaris HouseNorth Star AvenueSwindon, SN2 1EUUNITED KINGDOM
Peter Burkill
Plymouth Marine LaboratoryNatural Environment Research CouncilProspect PlaceWest HoePlymouth, PL1 3DHUNITED KINGDOM
Sharon Smith
Division of Marine Biology and FisheriesRosenstiel School of Marine and Atmospheric Science University of Miami4600 Rickenbacker CausewayMiami, FL 33149-1098United States of America

ii
ABSTRACT
The JGOFS Process Studies of the Arabian Sea have resulted in more than 120 publications thataddress different aspects of the biogeochemistry and physical forcing of this fascinating region.These publications arose from an integrated international study involving ca. 80 research cruises carried out by seven countries in the Arabian Sea.
Our understanding of the region has improved considerably from such disciplinary publicationsand suggest, for instance• Seasonal monsoonal forcing results in levels of primary production that are as high during
the weaker NE Monsoon as they are during the stronger SW Monsoon;• Bacterial production would seem to be less strongly coupled to monsoonal forcing than
primary production with an amplification of ca. 2, considerably lower than primaryproduction;
• While microzooplankton grazing continues at high levels throughout all seasons, they havelittle impact on diatoms that bloom during the SW Monsoon;
• The fate of these diatoms depends on the trophic coupling to herbivorous crustaceans, suchas Calanoides carinatus;
• Export flux to the deep ocean has been shown to be strongly seasonal and spatially variable.
These and other findings are presented in specialist disciplinary chapters in this report. Abibliography of research papers and other seminal publications including eight Special Issues of Deep-Sea Research has been compiled at the end of this report.
The next step is to focus on syntheses of the primary literature and data by scientific teamsworking across national boundaries and aiming for interdisciplinary publications in the peer-reviewed literature. A time-line for this, led by the JGOFS Indian Ocean Synthesis Group(IOSG), is laid out in this report.

iii
CONTENTS
1. INTRODUCTION 1
2. SPECIFIC TOPICS
2.1 The Surface Circulation of the Northern Arabian Sea 3
2.2 Hydrography 9
2.3 Air-Sea Interactions and Exchange 13
2.4 Primary Productivity in the Arabian Sea 26
2.5 Remote Sensing of Ocean Colour in the Arabian Sea 33
2.6 Microbial Dynamics 39
2.7 Zooplankton 54
2.8 The Role of the Oxygen Minimum Zone in Biogeochemical Cycles 60
2.9 Sedimentation 68
2.10 Arabian Sea Benthic Dynamics 77
2.11 Modelling Physical-Biogeochemical Interactions 82
3. MAJOR ACHIEVEMENTS, RECOMMENDATIONS AND FUTURE TIME-LINE
3.1 Major Achievements 88
3.2 Recommendations 89
3.3 Future Time-line and Research 90
4. APPENDICES
APPENDIX 4.1: JGOFS IOSG Membership 91
APPENDIX 4.2: Reference List from Section 2 92
APPENDIX 4.3 Seminal Literature: 1990-Present 115

1
1. INTRODUCTION
During the last decade, an intense study of the biogeochemistry of the Arabian Sea has takenplace. This has involved scientists from Canada, Germany, India, Netherlands, Oman, Pakistan,UK and USA, who participated in Process Studies run as part of the JGOFS Programme. Studies in the Arabian Sea were co-ordinated by the JGOFS Indian Ocean Planning Group (IOPG), who produced a Science Plan (SCOR 1995) that identified the nature of a co-ordinated,multidisciplinary study. The SCOR (1995) report also served as an Implementation Plan thatidentified the measurements to be taken by each nation on cruises, the PI’s responsible, and their contact details.
When the Process Studies ended in 1997, more than 80 research cruises had been carried out in the Indian Ocean, with most of these in the Arabian Sea over a 6-year period. During this time,the cruises mounted by Germany, India, Pakistan, UK and USA focused on the northern Arabian Sea during 1992/97 while Netherlands JGOFS cruises tackled the Somali system during 1992/93 (Figure 1).
Figure 1: JGOFS cruises mounted byGermany, India, Pakistan, UK and USAfocused upon coverage of the northern ArabianSea during 1992/97, while Netherlands JGOFScruises tackled the Somali system during1992/93.Details of each cruise are given onhttp://ads.smr.uib.no/jgofs/inventory/index.htm
National participation in the Arabian Sea Process Study and other regions of the Indian Oceanare given in Table 1. A detailed list of cruises is given on the Internet at:http://ads.smr.uib.no/jgofs/inventory/index.htm.
On completion of field studies in 1997, publication of the primary scientific papers began. Thishas been achieved largely through publication of Special Issues of Deep-Sea Research, Part II. A list of primary scientific papers, the Special Issues and other publications are given in Chapter 3.
The next step in developing our understanding of Arabian Sea biogeochemistry involvessynthesis of the primary literature to produce coherent integrated understanding of the keypathways, their interactions and controls. This was discussed by the JGOFS Indian OceanSynthesis Group (IOSG) at a meeting held in Bangalore in 19991; IOSG concluded that it should produce a report that “summarised advances in understanding of biogeochemical cycling in theArabian Sea derived from JGOFS Process Cruises and other associated activities”. Funds
1 A list of IOSG Committee Members is given in APPENDIX 4.1

2
released by JGOFS in 2000/01 allowed three scientists to meet and produce this report in June2001. This is based on statements of recent advances in scientific understanding viewed byrecognised experts in some dozen disciplinary fields (Chapter 2). However, we recognise thatthis report is an important, but not defining, step of Arabian Sea biogeochemical synthesis.Further international integration of syntheses will be required with peer-reviewed publications in the future. The time-line considered appropriate by IOSG is outlined in Chapter 3.
Table 1: A summary of cruises carried out in the Arabian Sea and other regions of the IndianOcean.
Country Region Cruises YearsAustralia SW Indian Ocean 4 1992/95France W Indian Ocean 1 1992Germany Arabian Sea 12 1995/97India Arabian Sea 7 1992/97Japan Indian Ocean 4 1994/96Netherlands Arabian Sea 11 1992/93Pakistan Arabian Sea 5 1992/94South Africa SW Indian Ocean 5 1990/93UK Arabian Sea 4 1994USA Arabian Sea 28 1994/96
Total 81

3
2. SPECIFIC TOPICS
2.1 The Surface Circulation of the Northern Arabian Sea
John C. Kindle
Oceanography Division, Naval Research LaboratoryStennis Space Center, MS 39529-5004
USA
Introduction
Prior to the Arabian Sea Expedition of 1994-1995, our understanding of the northern ArabianSea’s response to the seasonally reversing monsoon cycle was based on climatological ship drift reports (Cutler and Swallow, 1984). Although occasional expeditions to the region provided new insights (e.g., Elliot and Savidge, 1990; Bauer et al., 1991) an integrated understanding of themesoscale response to the monsoon cycle relative to the mean seasonal circulation was lacking.That the best estimates of the surface forcing resulted from monthly composites of ship windobservations complicated the efforts to understand this highly variable region. Hence, it is notsurprising that prominent among the recent discoveries are 1) the pronounced spatial andtemporal structure of the monsoon atmospheric forcing and 2) the importance of mesoscalevariability in the associated ocean response. Among the important mesoscale features are coastal jets and filaments along the Oman coast during the Southwest (SW) Monsoon that are capable of exporting nutrient rich upwelled water hundreds of kilometres offshore. This paper reviewsobservational and modelling studies of the seasonal response of the northern Arabian Seacirculation to the monsoon cycle with a focus on results from the Joint Global Ocean Flux Study(JGOFS) Arabian Sea Expedition of 1994-95. Emphasis is placed on the circulation featuresnorth of approximately 10°N. An excellent review of the basin-wide circulation features,including results from the World Ocean Circulation Experiment (WOCE) efforts of 1994-96 can be found the recent article by Schott and McCreary (2001).
Atmospheric Forcing
The basin-scale circulation of the Arabian Sea is governed by the response to a seasonallyreversing monsoon cycle, characterised by upwelling-favourable, south-westerly winds duringthe SW Monsoon, reversing to north-easterly (albeit weaker) winds during the Northeast (NE)Monsoon. Meteorological measurements from a moored array centred at approximately 15.5°N,61.5°E and deployed from October 1994 to October 1995, provide the first high frequency (i.e.,sub-monthly) direct observations of the monsoon cycle in the interior of the Arabian Sea(Rudnick et al., 1997; Weller et al., 1998). Based on these measurements, the four seasons forthe 1994-95 monsoon year were defined as: September 16 to October 31 for the FallIntermonsoon; November 1 to February 15 for the NE Monsoon; February 16 to May 31 for theSpring Intermonsoon and June 1 to September 15 for the SW Monsoon. The excellent agreement between these observations and results from operational atmospheric prediction models (Welleret al., 1998) (Figure 1) suggest that the atmospheric models may be used to describe theessential characteristics of the spatial and temporal variability of the of the atmospheric forcingthroughout the basin. Likewise, Halpern et al., (1998) discovered excellent agreement betweenthe observed winds from the mooring and the satellite-derived winds from the ERS-1scatterometer.

4
Figure 1. Time-series of observed wind magnitude (dark line) versus atmospheric model winds(light line) at 15.5°N, 61.5°E for the period October 1994 to October 1995. Theatmospheric model winds are from the U.S. Navy’s Operational Global AtmosphericPrediction System (NOGAPS).
Both the atmospheric model winds (Weller et al., 1998) and the satellite-derived winds (Halpern et al., 1998) reveal significant regional differences in the timing of the onset and relaxation ofthe winds within a season. For example, Weller et al., (1998) used the operational wind product from the U.S. Navy to reveal a 7-10 day delayed onset of the 1995 SW Monsoon at the mooring site, whereas the onset near the Oman coast occurred normally. However, it should be noted that upwelling-favourable winds along the coast of Oman generally begin in April-May (Weller etal., 1998), even prior to the reversal of the winds in the interior of the Arabian Sea. This isconsistent with the analysis of ship reports by Fieux and Stommel (1977) indicating that theonset of upwelling-favourable winds along the Oman coast may begin in April. While the winds along the Oman coast in April-May are not as strong as during the SW Monsoon period, theyappear to be sufficient to generate a coastal upwelling response as detected in AVHRR imagery(Rixen et al., 1996; Arnone et al., 2001). The potential roles of this weak-to-moderate coastalupwelling during April-May in setting the stage for the physical and biogeochemical responsesto the primary SW Monsoon onset in June warrant further study.
Ocean Response to the Monsoon Cycle
The Southwest Monsoon
Historical ship drift records show the existence of the Oman Coastal Current (OCC) north ofapproximately 14°N by early May (Cutler and Swallow, 1984). In their analysis of the ship drift records, Elliot and Savidge (1990) showed a north-eastward flowing OCC of around 0.4 m sec-1
in magnitude and extending to 200 km offshore during the SW Monsoon. The current turnedabruptly to the east off Ras al Hadd (Figure 2). However, direct measurements of the coastalflow from ADCP instruments during this season in 1987 showed that the north-eastward coastal flow weakened toward the southwest from an estimated transport of ~10 Sv. near Ras al Hadd to weak and variable flow at ~ 17°N-18°N (Elliot and Savidge, 1990). Additionally, the ADCP data analysed by Flagg and Kim (1998) for the 1995 SW Monsoon displayed no mean coastal current

5
to the northeast, but rather the presence of variable flow characterised by current reversals overrelatively small distances. Flagg and Kim (1998) hypothesised that the differences between thedirect observations of the OCC and the historical ship drift data may be accounted for by asystematic bias in the ship observations due to the persistent nature of the high winds and sea-state during this period. Clearly, questions remain about the nature of the Oman Coastal Current:During the SW Monsoon, is the OCC comprised of several distinct flows of limited alongshoreextent or does it exist in the mean but disrupted by the pronounced mesoscale variability?Moreover, while we know the fate of the coastal flow after it leaves the coast at Ras al Hadd, we know little of its origins. The southeastern-most extent of the OCC has not been documented.
Figure 2. AVHRR image from 06 August 1994 showing major filaments extending offshorefrom Ras ash Sharbatat, Ras al Madrakah and Ras al Hadd (Image courtesy of R. Arnone).
Among the most prominent SW Monsoon features along the Oman coast are filaments and jets(Figure 2) which are capable of exporting cool, nutrient rich, upwelled coastal waters hundredsof kilometres offshore (Young and Kindle, 1994; Brink et al., 1998). The first directmeasurements of these features were described by Elliot and Savidge (1990) who discoveredplumes of cold water extending offshore from the coast in the region between Ras ash Sharbatat and Ras al Madrakah. This is in approximately the same region as a major filament that wasobserved during the 1995 SW Monsoon (Brink et al., 1998; Flagg and Kim, 1998; Arnone et al.,2000; Lee et al., 2000; and Manghnani et al., 1998). Some properties of the filaments have been described as similar to those off the U.S. Pacific west coast (Elliot and Savidge, 1990; Brink etal., 1998). However, Flagg and Kim (1998) view the offshore-directed plumes as part of a major anticyclonic feature that remains essentially in place for approximately six months, therebyextending through the SW Monsoon into the winter season. Manghnani et al. (1998)hypothesised that the plume was not part of the offshore deflection of the coastal current, butrather resulted from an interaction between the wind field and anticyclonic mesoscale featuresthat may have existed prior to the onset of the SW Monsoon.
When the Oman Coastal Current reaches Ras al Hadd, the flow turns offshore to the northeastand east to form the Ras al Hadd Jet (Figure 2). This feature is also referred to as the Ras alHadd Front because it forms the seasonal boundary between the northern Arabian Sea and theGulf of Oman. The measurements of Elliot and Savidge (1990) of the OCC just prior to reaching

6
Ras al Hadd, revealed a transport of ~ 10 Sv. within 100 km off the coast. Flagg and Kim (1998) discovered that the Ras al Hadd Jet intensified in August 1995 following the reversal of the flow along the northeastern Oman coast from northward to southward, thereby adding to the flowalong the Ras al Hadd Front. They speculated that the reversal of the flow along the northeastern Oman coast in August was related to the intensification and/or propagation of a cyclonic eddy in the Gulf of Oman during this period. Similarly, Baker et al., (1996) suggested that such an eddyplays a role in the dynamics of the Ras al Hadd Jet: They hypothesize that the OCC at Ras alHadd forms a double vortex as it extends offshore during the Fall Intermonsoon period. It isspeculated that to the south, an anticyclonic vortex (or eddy) forms while to the north, in theGulf of Oman, a cyclonic eddy forms, both of which are driven by the extension of the Ras alHadd Jet into open waters. Böhm et al. (1999) provided an extensive description of the Ras alHadd Jet and the associated cyclonic and anticyclonic eddies based on ADCP measurements and remotely sensed observations in 1994 and 1995. They estimated that the transport of the jetvaried from 2-8 Sv. in magnitude and exhibited a maximum flow in September. Böhm et al.(1999) suggested that, while the counter-rotating eddies may interact with the jet, they may notnecessarily be generated by this feature. The relationship(s) between the Ras al Hadd Jet and the respective anticyclonic and cyclonic eddies to the south and north deserves further attention.
Among the key results of the ADCP analysis by Flagg and Kim (1998) is that the eddy kineticenergy (EKE) in the northern Arabian Sea dominates that of the mean and is enhanced in thenear coastal region. In an extension of the ADCP-based study, Kim et al. (2001) utilisedsatellite-derived sea level anomalies from the TOPEX/Poseidon altimeter to provide additionalanalyses of the EKE properties in the northern Arabian Sea. The results were in agreement with the ADCP study in affirming the importance of eddy activity in this region. The seasonalvariation of EKE was such that it increased in magnitude and areal extent during the SWMonsoon. Horizontal scales of the eddies varied from ~200-500 km nearshore to ~100-200 kmin the interior. Interestingly, they also discovered that the primary area of reduced eddy presence coincided with the core of the Oxygen Minimum Zone (OMZ). Furthermore, the study revealedthat eddy activity within the Arabian Sea basin was confined primarily to the region north of15°N. The generating mechanisms for the eddy field remain to be determined.
The behaviour of the surface mixed-layer during this season was addressed in both observational and modelling studies. Lee et al., (2000) used high-resolution observations from a towed profiler during 1994 and 1995, as well as climatological records of surface forcing and mixed-layerdepths, to examine the seasonal behaviour of the surface mixed-layer in the northern ArabianSea. Their analysis suggests that during the SW Monsoon, the shallow mixed-layer inshore ofthe wind maximum is maintained by a balance between wind-driven entrainment and thecombined effects of horizontal advection and Ekman pumping. Offshore of the wind maximum,Lee et al., (2000) show that Ekman pumping and wind-driven entrainment act together to deepen the mixed-layer with minimal influence from horizontal advection. Fischer (2000) utilised datafrom the Woods Hole Oceanographic Institute (WHOI) mooring at ~15.5°N, 61.5°E, a locationthat was chosen to approximately coincide with the climatological axis of the wind stressmaximum during the SW Monsoon, and a three-dimensional model to examine the mixed-layerresponse during this season. His results indicated that, at this location, local forcing dominatedthe mixed-layer response except during the latter stages of the SW Monsoon, when horizontaladvection played an important role. Similarly, Rochford et al., (2000), in a modelling studyusing a three-dimensional numerical model forced by 12-hourly atmospheric model winds,suggested that local forcing dominated the mixed-layer response at the mooring site during thefirst half of the SW Monsoon, but horizontal advection was also an important component during the second half of the summer (SW) Monsoon period. Finally, the modelling studies of Fischer(2000) and McCreary et al. (2000) found that rectification to the diurnally forced signal needs to

7
be included to produce the most realistic mixed-layer response. This is particularly true forcoupled bio-physical models attempting to simulate the seasonal surface chlorophyll distribution in the interior of the northern Arabian Sea (McCreary et al., 2000).
The Fall Intermonsoon
Prominent features along the Oman coast generated during the SW Monsoon tend to linger inplace during the Fall Intermonsoon, as noted above for the cyclonic and anticyclonic eddiesassociated with the Ras al Had Jet (Arnone et al., 2001). Flagg and Kim (1998) noted thecontinued presence of the Ras al Hadd Jet, which appeared to maintain its intensity during theFall Intermonsoon period due to the intensification of the southward flow along the northeastcoast of Oman. Shi et al. (2000) reported that the cold surface water upwelled during the SWMonsoon lingered for nearly a month following the end of the monsoon in bays along the Oman coast.
The Northeast Monsoon
The transition from the Fall Intermonsoon to the NE (winter) Monsoon occurs in November. Inthe northern Arabian Sea, the primary circulation response to the onset of the northeasterlywinds is the reversal of the Oman Coastal Current to southeastward flow, thereby yielding acontinuous southward current that extends along the northeast coast of Oman, turns the corner at Ras al Hadd and continues southward along the coast until it is entrained into offshore directedsquirts and jets south of ~ 20°N (Flagg and Kim (1998). Anticyclonic features that were oncedirectly connected to the coastal circulation during the SW Monsoon and Fall Intermonsoonperiods evolve into separated eddies that exhibit a tendency to propagate southward along thecoast and may occasionally directly impact the coastal circulation. Along the northeast coast ofOman, the southward flow decays and reverses to weak northward flow that persists fromJanuary to July (Flagg and Kim, 1998).
The mixed-layer behaviour during the NE Monsoon is characterized by increased deepeningwith distance offshore and is dominated by convective overturning (Lee et al., 2000). This isconsistent with the earlier work of Weller et al., (1998) who utilised observations from theWHOI mooring to demonstrate the importance of convective overturning to the mixed-layerresponse in the central Arabian Sea during this season. Wiggert et al. (2000) used observationsfrom the mooring and a one-dimensional coupled bio-physical model to show the importance of diurnal forcing to the surface chlorophyll a distributions. They also hypothesised that theinterannual variation of this mechanism may be an important factor in the observed interannualvariability of chlorophyll a distributions during this monsoon season. Such interannualvariability was also examined by Kumar et al. (2001) who discovered deeper mixed-layerdepths, colder SST values and elevated chlorophyll concentrations during the NE Monsoon of1997 relative to that of 1995.
Recommendations
1. To perform retrospective synthesis studies that combine in situ and remotely sensed data together with data-assimilative model simulations capable of representing observedfeatures during the Arabian Sea Expedition of 1994-95. Such simulations would be ableto significantly aid the investigation of the relative importance of physical forcingmechanisms governing the biological response during the monsoon cycle. For example,such studies are needed to help interpret the statistical relationships between the sediment flux measurements and surface forcing.

8
2. To conduct additional field work just prior to and following the onset of the SWMonsoon to investigate such features as 1) the role of the upwelling along the Omancoast during April-May prior to the onset of the SW Monsoon, 2) the generatingmechanisms for the pronounced filaments observed during the SW Monsoon with a focus on examining the role(s) of pre-existing coastal features in the development of coastaljets and filaments, 3) the intriguing hypothesis of the role(s) diapausing copepods play in the timing of diatom blooms relative to the onset of the SW Monsoon.
3. To conduct additional fieldwork during the NE Monsoon to better understand and to testemerging hypotheses to explain the pronounced interannual variation of phytoplanktonblooms during this season.

9
2.2 Hydrography
John M. Morrison
Department of Marine, Earth and Atmospheric SciencesNorth Carolina State University, Raleigh, NC 27695
USA
Introduction
The semi-annual reversal in wind stress associated with the monsoon, water mass intrusionsfrom marginal seas and other oceans, geographical effects (the northern Indian Ocean beingblocked by the Eurasian landmass) and therefore no subtropical convergence or deep waterformation, give the upper waters of the Arabian Sea a unique thermohaline structure andcirculation (Wyrtki, 1971; Morrison and Olson, 1992; You and Tomczak, 1993; Morrison et al.,1998). The complex water mass structure (Figure 1) is due, in part, to advection and interleaving of water masses, and, in part, to formation of high-salinity waters in the Red Sea, Persian Gulfand northern portion of the basin (Arabian Sea Water) which sink to moderate depths in thecentral basin. Wyrtki (1971) provides an overall view of water mass structure for the IndianOcean. The yearly cycle in the Arabian Sea is normally divided into a Northeast (NE) Monsoon, characterised by moderate winds from the northeast during the late fall and winter, a Southwest(SW) Monsoon, characterised by strong winds from the southwest during the late spring andsummer, and Fall and Spring Intermonsoons. As Düing (1970) notes, the transition periods canvary from year to year and can occur at different times from region to region. Off Somalia, forexample, SW Monsoon conditions begin to “spin-up” in May, but in the central Arabian Sea,May is a period of relatively weak winds and lowest Coastal Zone Color Scanner (CZCS)chlorophyll levels (Smith et al., 1991). Winds during the peak of the SW Monsoon are amongst the strongest tropospheric winds on the planet (Smith et al., 1991) and are associated withextremely high inputs of dust (Prospero, 1996), coastal upwelling, and, in the canonicaldescriptions (e.g., Smith and Bottero, 1977), offshore upwelling.
The canonical description of the wind-driven upwelling regime during the SW Monsooninvolves coastal upwelling driven by the wind stress and an offshore upwelling regime driven by the curl of the wind stress (e.g., Smith and Bottero, 1977) that occurs inshore of the high velocity core (the Findlater Jet) of the SW Monsoon’s wind field. Observations made on sub-mesoscalesampling grids, in combination with satellite sea surface temperature (SST) data, suggest thathigh surface nutrients in the offshore regions of the Arabian Sea during the SW Monsoon areassociated with complex dynamics in the upwelling zone that include cool filaments, similar tothose formed in other upwelling regions (Abbott and Barksdale, 1995). These observations alsosuggest that it is difficult for the wind regime to produce organised upwelling circulationsoffshore (Brink et al., 1998). The hypothesis is that any coherent offshore upwelling northwestof the Findlater Jet quickly becomes unstable and collapses to an eddy field, as shown in theacoustic Doppler current profiler data by Flagg and Kim (1998). Offshore upwelling, however,has been suggested for other portions of the Arabian Sea (Muraleedharan and Kumar, 1996).
Recently, the northern Arabian Sea was extensively studied, as part of the Joint Global OceanFlux Study (JGOFS) during 1994-1996. The observations consisted of repeated ship surveys;special frontal surveys in the upwelling regions; and satellite sea surface temperature (SST) and altimetry data. For the SW (summer) Monsoon circulation off the Arabian peninsula, it wasdetermined that a northeastward coastal jet develops along the south coast of Oman in early May and persists throughout the SW Monsoon, called the Ras al Hadd Jet by Böhm et al. (1999). Itflows past Ras al Hadd at the southeast corner of the Gulf of Oman and meanders into theinterior, transporting cooler, coastally upwelled waters into the warmer environment of the Gulf

10
of Oman and northern Arabian Sea, and therefore is easily identified in satellite SSTobservations. The jet is associated with an anticyclonic gyre just south of its offshore turningpoint and a cyclonic one to its north (Flagg and Kim, 1998; Böhm et al., 1999). Shi et al. (1999) determined that the cause of this alongshore, northeastward flow is an onshore pressure gradient (established by the upwelling) that reduces sea level near the coast by about 30 cm. During theNortheast Monsoon, warm water flows southward past Ras al Hadd into the open Arabian Sea,so that the coastal jet is fed from the north, rather than the west as during the SW Monsoon.Along the rim of the northern Arabian Sea, the mean flow during the NE Monsoon is westwardwith downwelling along the coast and an offshore pressure gradient (Shi et al., 2000).
Other circulation cells exist elsewhere along the Oman coast, their locations apparently linked to protruding capes of the Arabian Peninsula. Between these cells, upwelled waters can betransported offshore in filaments or squirts, which extend as far as 400 km offshore and can bedetected in satellite SST and colour imagery. In these filaments with widths of up to 100 kmoffshore, currents in excess of 50 cm s-1 have been observed (Brink et al., 1998).
Water Mass Formation in the Upper Ocean
Because of this intense variability in the circulation associated with the annual monsoon cycle,water mass structure in the upper kilometre of the Arabian Sea is extremely complex (Figure 1).Low salinity waters of southern origin are advected into the region by the Somali and NorthEquatorial Currents. In addition, during the SW Monsoon, the waters that upwell along theArabian Coast have relatively low salinities as a consequence of poleward advection associatedwith the Somali Current. Precipitation and runoff from rivers in the eastern Arabian Sea mustcontribute to the local formation of low salinity waters, although salinities in the upper severalhundred metres are always greater than 35.0 and generally greater than 35.5.
During the NE Monsoon, the cold and dry winter winds from Asia, combined with Ekmanpumping, cause subduction of high-salinity surface waters in the interior of the northern Arabian Sea (Morrison, 1997; Schott and Fischer, 2000). This generates the widespread Arabian SeaWater (ASW) salinity maximum (Figure 1) just underneath the mixed-layer with a potentialdensity (σθ) of ~25 (Morrison, 1997). It should be noted that Donguy and Meyers (1996) define a surface water mass with salinity between 35.5-36.5 and temperatures greater than 22°C asArabian Sea Water. The most dense ASW as defined by Donguy and Meyers (1996) would have a σθ of ~25.35 kg m-3 which would mean that it could overlap with the ASW. However, weprefer the definition that associates this water mass with a salinity maximum at a σθ of ~25.0 kg m-3. The formation of ASW is a complex process involving seasonal heating, cooling, andevaporation, and advection of low salinity waters. Morrison (1997) describes the formation ofASW and its associated salinity. Briefly, waters with ASW salinity and temperaturecharacteristics can be formed at the sea surface in the northern Arabian Sea by cooling andevaporation during the NE Monsoon. This relatively dense water can then be capped by lowerdensity surface waters advected into the region or arising from heating during the Intermonsoons and the SW Monsoon. A major cause of the ASW salinity maximum (when it is present) is theadvection of low salinity water from the Oman upwelling region in the Ras al Hadd Jet duringthe SW Monsoon. This upwelled water can sink beneath the warm surface waters and lie aboveASW, thereby contributing to the salinity maximum that is associated with ASW.
During the Indian JGOFS programme, observations during February-March 1995 suggest thepresence of a relatively cool, high salinity water mass (Arabian Sea High Salinity Water -ASHSW) that has been formed due to the action of the NE Monsoon winds and reducedinsulation (Kumar and Prasad, l996). The mode of formation of this water mass is similar to that for ASW, and the main distinction is that its density tends to be lower than the 25.0 kg m-3σθvalue that is often used to characterise the ASW salinity maximum. Kumar and Li (1996)

11
suggest a characteristic temperature, salinity, and σθ for this water mass of 26.345°C, 36.387,and 24.082 kg m-3, respectively. Nevertheless, the denser fractions of ASHSW have σθ’s > 25.0 kg m-3. It might be appropriate, therefore, to think of ASW as a relatively dense fraction ofASHSW.
Intermediate Water Masses
At intermediate depths, You and Tomczak (1993) suggest that Indian Central Water (ICW) andan “aged” version of this water mass that they call North Indian Central Water (NICW) are themain source for thermocline ventilation for the northern Indian Ocean with only minorcontributions from the Red Sea and the Persian Gulf outflows. The conversion of ICW to NICW is a slow process, accompanied by a significant fall in oxygen values due to consumption oforganic matter and a corresponding increase of nutrient concentrations (You, 1997). Theysuggest a transition path of ICW: subduction in the South Indian Ocean at ~ 40-45 °S, advection with the subtropical gyre, and exit to the northern Indian Ocean as part of the western boundarycurrents. The northward spreading of ICW into the equatorial and northern Indian Ocean is byway of the Somali Current centred at 300-400 m with σθ of ~26.7 during the SW Monsoon andby way of the Equatorial Countercurrent during the NE Monsoon (You, 1997). This is consistent with the indications of Warren et al. (1996) that northward-moving Subtropical SubsurfaceWater which corresponds to the ICW of You and Tomczak (1993) appears to penetrate wellnorth of the equator in the Somali Basin. Kumar and Li (1996) call ICW Central Indian Waterand suggest a core σθ of ~27.5 kg m-3. They agree that it is of major importance in ventilatingthe thermocline of the northern Indian Ocean, but they consider it less important for supplyingwaters to the thermocline of the western basin (Arabian Sea). They suggest that evaporation inthe Arabian Sea, Red Sea, and Persian Gulf contributes to the formation of North Indian

12
Intermediate Water (NIIW). This is a high salinity water mass with a characteristic σθ of ~27.1kg m-3 that is not very different from Wyrtki’s RSW (see below).
The northern marginal seas, the Red Sea and Persian Gulf (sometimes referred to as the Arabian Gulf), inject water masses into the Indian Ocean that are identifiable over long distances by their high salinities at core densities. The dynamics of both marginal seas are quite different, with the Red Sea being deep and separated from the Gulf of Aden by a shallow sill, similar to thesituation of the Mediterranean Sea, while the Persian Gulf shallows northward from its outflowchannel. Salinity maxima (Figure 1) arising from Persian Gulf (PGW) and Red Sea (RSW)waters are centred at 26.6 kg m-3 and 27.2 kg m-3 σθ, respectively (Wyrtki, 1971). Persian GulfWater has two spreading paths into the Arabian Sea. First, a narrow subsurface jet of PGW flows to the south as a western boundary undercurrent under the upwelling zone where it spreadseastward into the central basin (Morrison, 1997). Second, PGW spreads to the east in thenorthern extremes of the Arabian Sea and down the coast of India during NE Monsoon andadvects to the west into the central basin during SW Monsoon (Morrison, 1997). PGW is present throughout the entire northern Arabian Sea, identified by it core salinity maximum, but it rapidlylooses its identity further to the south.
A seasonal cycle of the outflow from the Red Sea through the Gulf of Aden and into the IndianOcean with a winter maximum has been documented (Murray and Johns, 1997). Its preferredspreading route into the Indian Ocean is through the passage between Socotra Island and theAfrican continent (Schott and Fischer, 2000) with a second branch of this undercurrent goingaround Socotra (Schott and Fischer, 2000). This water mass is observed as far south as theMozambique Channel (Beal et al., 2000a). Water mass properties show that the lower part ofthis undercurrent is the main supplier of RSW out of the Gulf of Aden into the Indian Ocean.Meanwhile, in the central basin, RSW is a rapidly mixed away with the spread of RSW out ofthe Gulf of Aden being very inhomogeneous in the Arabian Sea due to the intense eddy activity(Quadfasel and Schott, 1982). RSW is almost mixed away by 64° E in the central basin and has little effect on the water masses in the central Arabian Sea (Morrison, 1997). You and Tomczak (1993) found that the influence of RSW in the upper thermocline is mainly restricted to theArabian Sea, where it contributes up to 30% to the thermocline water. In contrast, Olson et al.(1973), using chlorofluorocarbon (CFC) data, found an even smaller contribution of RSW(<10%). Meanwhile, Antarctic Intermediate Water (AAIW) is spreading northward into theArabian Sea on essentially the same density horizon as RSW (core density of 27.1- 27.3 kg m-3
σθ; Figure 1). AAIW has entered the Indian Ocean basin from the Antarctic in the southeastern region (Fine, 1993). It is generated by subduction in the subpolar frontal zone, and is marked bylow salinities due to the excess of precipitation over evaporation in that area and oxygenmaximum.
Deep Ocean Water Masses
As noted above, the Indian Ocean has no northern sources of deep water. Therefore, deep water can only be supplied from the south. The deep water has the cold, fresh, oxygen-richcharacteristics of Lower Circumpolar Water (CDW) which flows northward in the Madagascarand West Australian basins (Toole and Warren, 1993), but it cannot directly enter the centralIndian Basin because of obstruction by the deep ridges. The western pathway of this water mass into the Northern Hemisphere is through the Somali Basin (Johnson et al., 1991b. Beal et al.,2000b) where it must disappear through gradual upwelling into the overlying Deep Water toform Indian Deep Water (IDW) which is specific to the northern Indian Ocean (Figure 1). This water eventually enters the Arabian Basin. The depth range of IDW in the Arabian Basin is from 3,800 m upward to about 1,500 m (Johnson et al., 1991a). Like the flow of Bottom Water, theflow of Indian Deep Water is northward and concentrated in western boundary currents (Warren and Johnson, 1992).

13
2.3 Air-Sea Interactions and Exchanges
S.W.A. Naqvi
National Institute of OceanographyDona Paula, Goa 403 004
India
Introduction
At the time of planning JGOFS Process Studies, it was recognised that the Arabian Sea makes a significant contribution to biogeochemical ocean-atmosphere transfer processes. However, themagnitudes of these exchanges were not well quantified even for constituents such as carbondioxide (CO2), nitrous oxide (N2O) and methane (CH4), for which some data were available. For others like ammonia (NH3) and dimethylsulphide (DMS), which had not been measuredpreviously, new measurements were planned. The work carried out during the JGOFS andrelated cruises on these climatically important atmospheric constituents, has greatly expandedthe database, allowing for improved estimates of their air-sea exchange rates.
Carbon Dioxide (CO2)
At the time of JGOFS planning, the Indian Ocean north of the Hydrochemical Front at ~10°Swas believed to serve as a source of CO2 to the atmosphere. In this respect there were severalcrucial questions concerning carbon cycling in the region that the JGOFS programme aimed toaddress. It was thought that since the upwelled water at the surface had very high pCO2,upwelling in conjunction with a wind-induced enhancement of the air-sea gas exchangecoefficient would result in a large efflux of CO2 to the atmosphere from the northwestern Indian Ocean. This effect would be aided by the secretion of CaCO3 by organisms (which raises pCO2),but opposed by the high rate of photosynthetic production (which lowers pCO2), sustained bycopious nutrients in the mixed-layer. The relative importance of these processes was not known. Also, as the physical forcings associated with the NE and SW Monsoons are highly variable inspace and time, large changes in the air-sea exchange of CO2 were expected.
Extensive surveys made under the JGOFS, WOCE (World Ocean Circulation Experiment) and,to a smaller extent, OACES (Ocean-Atmosphere Carbon Exchange Study) programmes havegreatly increased the existing carbon database for the Indian Ocean. This has enabled a muchmore reliable quantification of the air-sea CO2 exchange and an evaluation of its spatial andtemporal variations in the Indian Ocean, particularly in the Arabian Sea. These data weregenerated through both continuous underway pCO2 measurements and discrete water sampling.Results of these studies have conclusively established that the Arabian Sea is indeed a source ofCO2 to the atmosphere at almost all places and during all seasons, but with very large seasonaland geographical variability. The surface-water pCO2 levels are the highest in coastal upwelling zones: values reaching up to 700 µatm being observed off Oman (Figure 1) during theSouthwest (SW) Monsoon season (Körtzinger et al., 1997; Goyet et al., 1998; Mintrop et al.,1999). In the northern Arabian Sea, convective mixing during the NE Monsoon can elevate thepCO2 level to 500 µatm even in offshore areas (Sarma et al., submitted). pCO2 approaching 700 µatm was also recorded in the SW Monsoon upwelling zone off western India; however, as theupwelled water in this region is often overlain by a low-salinity layer formed as a result ofintense rainfall in the coastal zone, the surface water pCO2 (minimum 266 µatm) may sometimes be below saturation (Sarma et al., 2000). Ignoring these rare instances of pCO2 undersaturationcaused by a combination of physical and biological processes (a similar phenomenon has also

14
been reported from the Bay of Bengal; see Kumar et al., 1996), it would appear that theincreases in pCO2, through upwelling and vertical mixing of subsurface waters greatlyoverwhelm its biological removal in the northwestern Indian Ocean. Interestingly, theatmospheric pCO2, which also exhibits considerable seasonal variability, is the lowest (~340µatm) during or just after the SW Monsoon (Goyet et al., 1998) accentuating the sea-air CO2
gradient (∆pCO2 = pCO2sea - pCO2
air), while the exchange coefficient is the highest due to strong monsoon winds. The combination of these effects leads to some of the most intense emissions of CO2 (up to 119 mmol m-2 d-1, Körtzinger et al., 1997) from the sea surface.
Figure 1. Partial pressure of CO2 in surface water (bold curves) and in the atmosphere (dots on dashed curves) and water temperature (thin curve) along two transects of the upwelling zone off Oman during July-August, 1995 (from Körtzinger et al., 1997).
In the Arabian Sea, seasonal contrasts in pCO2 and in the air-sea flux are best marked in coastal waters, but large seasonality in the open ocean is also observed in the Southern Indian Ocean(Figure 2; Louanchi et al., 1996; Sabine et al., 2000). The amplitude of seasonal changes in the open Arabian Sea is somewhat uncertain. The U.S. JGOFS observations in the central andwestern Arabian Sea over a complete annual cycle, involving continuous, automatedmeasurements, revealed substantial seasonal changes in the offshore region, with the lowest∆pCO2 (<32 µatm) occurring during the non-Southwest Monsoon periods (Goyet et al., 1998).

15
Figure 2. Season-wise distribution of surface water pCO2 (µatm) in the Indian Ocean (fromSabine et al., 2000).
In comparison, the Indian JGOFS data, mostly based on pH-TCO2 or pH-alkalinitymeasurements (Sarma et al., 1998), indicated smaller variability and higher ∆pCO2 (up to 124µatm). Accordingly, estimates of annual CO2 flux to the atmosphere from the Arabian Sea alsodiffer significantly (7 TgC by Goyet et al., 1998, and 79 TgC by Sarma et al., 1998). The latter estimate is close to that [30-76 TgC (3 months)-1] based on observations during the SWMonsoon season as a part of the German JGOFS programme (Körtzinger et al., 1997). Whilesome of these differences may arise from different techniques used for measurements, datainterpolation and computations, they probably reflect the large temporal (both seasonal andinterannual) and spatial variations that characterize air-sea CO2 exchange in this region.
The WOCE-JGOFS data combined with earlier measurements made during the GEOSECS andINDIGO expeditions have been used to evaluate the extent of anthropogenic CO2 penetration in the Indian Ocean (Sabine et al., 1999). The highest concentrations and the deepest penetrations(>1 km) were found to occur in the region of the subtropical convergence (30-40°S) evidentlythrough the formation of intermediate waters, while these were the least in the Antarctic region. The northwestern Indian Ocean, which receives outflows from the Persian Gulf and the Red Sea, appears to hold more anthropogenic CO2 than the equatorial Indian Ocean (Sabine et al., 199;Figure 3A). The total inventory of anthropogenic CO2 in the Indian Ocean north of 35°S hasbeen estimated to be 13.6 PgC (1 Pg = 1015 g) while the increase over the 18-year intervalbetween the GEOSECS and WOCE-JGOFS expeditions works out to be 4.1 Pg C. An interesting aspect of these results is that the observed trends of anthropogenic CO2 distribution (Sabine etal., 199; Figure 3A) show some important differences from those predicted by the Princeton

16
ocean biogeochemistry model (Sabine et al., 1999; Figure 3B) with the databased penetrationestimate being higher in the northern Indian Ocean and lower in the Antarctic sector.
Figure 3. Distribution of vertically integrated anthropogenic CO2 (mol m-2) based on (A) dataand (B) model estimates (from Sabine et al., 1999).
Evaluation of anthropogenic CO2 penetration in the northern Indian Ocean has also been madefollowing another approach that takes into account contributions from various source watermasses. This work is documented in Goyet et al. (1999). The WOCE data used for this studywere collected along three transects, the first in the Gulf of Aden parallel to the Yemeni coast,the second across the Arabian Sea along 9°N latitude, and the third across the Bay of Bengal(including the Andaman Sea) along 10°N latitude. Surface waters in these regions werecomputed to contain over 40 µmol kg-1 of anthropogenic CO2. The depth of penetration of theanthropogenic signal was noticed to decrease from ~1500 m in the Gulf of Aden (which receives freshly-formed intermediate waters from the Red Sea) to <1000 m in the Bay of Bengal.Underway pCO2 analysis carried out during the same expedition (Figure 4) revealed fairly large supersaturations in the Gulf of Aden and in the western and central parts of the Arabian Sea.This was presumably due to the aforementioned upwelling and warming of CO2-rich deeperwaters. Lower pCO2 values in the easternmost portion of the Arabian Sea transect and alongmost of the Bay of Bengal transect were associated with low-salinity surface water masses. Itmay be noted that the ∆pCO2 reached a minimum of ~-30 µatm in the Andaman Sea thatreceives large runoff from the Irrawaddy and Salween rivers. This further reinforces the viewthat freshwater discharge in the northeastern Indian Ocean makes this region a net sink foratmospheric CO2 unlike the northwestern Indian Ocean (Kumar et al., 1996).Regional variability of pCO2 in surface waters and of the causative processes (thermodynamics,vertical mixing, biological activity and air-sea exchange) has also been demonstrated throughmodelling by Louanchi et al. (1996). The one-dimensional model employed by these authors indifferent provinces of the Indian Ocean north of 50°S latitude, yielded a spatio-temporaldistribution that closely matched with the observations. However, some discrepancies betweenthe model results and observations were noticed. For example, the model-predicted distributionfor the Arabian Sea showed two peaks separated by a minimum during the SW Monsoon (July-August) that is not consistent with the data as described above.

17
Figure 4. Distributions of pCO2 in surface water (circles) and in the atmosphere (triangles)during August-October, 1995, along three WOCE transects: (a) from Gulf of Aden to theArabian Sea; (b) along Lat. 9°N in the Arabian Sea; and (c) along Lat. 10°N in the Bay ofBengal and the Andaman Sea (from Goyet et al., 1999).
This indicates a possible overestimation of the loss (through the biological pump and air-seaexchange) or an underestimation of supply (entrapment from deeper water). Also, the seasonalcontrast in surface pCO2 was generally greater in the eastern equatorial Indian Ocean than in the northwest Indian Ocean. The model results enabled the identification of four different provinces based on their characteristic surface pCO2 fields. While the region south of 35°S latitude appears to be a net sink (~500 TgC y-1) for atmospheric CO2 during all months (with a maximum during the Austral winter), that north of 10°S latitude is a net source (~150 TgC y-1 with maximalemission during the SW Monsoon). Between these two extremes lie the southern subtropicalregion and the transition zone bounded by the latitudes 20°S and 10°S; the former serves as amoderate sink (~170 TgC y-1) whereas the latter is neither a sink nor a source on an annual basis.
Sabine et al. (2000) have recently utilized 240,000 surface pCO2 measurements made during1995-96 under JGOFS, WOCE and OACES to establish its relationships with other parameterssuch as the sea surface temperature (SST), salinity and location. These relationships are thenused with the NCEP/NCAR reanalysis, SSTs, winds and atmospheric pressures and the salinityvalues derived from Levitus et al., (1994) to compute basin-wide pCO2 and air-sea fluxes for

18
each month. Their results show a larger seasonal cycle for the tropical and subtropical IndianOcean than simulated by Louanchi et al. (1996). Also, unlike the Pacific and the AtlanticOceans, the equatorial Indian Ocean appears to be a relatively small source of CO2 to theatmosphere. The net annual flux of CO2 for the region lying north of 36°S latitude was 150 TgC, and it was directed from the atmosphere to the ocean. This is at variance with the results ofLouanchi et al. (1996) who concluded that the Indian Ocean north of 35°S latitude serves as aweak source of atmospheric CO2 (22 TgC y-1). The estimate of Sabine et al. (2000) is inagreement with that of Takahashi et al. (1999), although the regional breakdowns of theestimates are different because of different techniques used for data processing. As expected, the air-sea fluxes exhibit large geographical and seasonal changes. The Indian Ocean as a whole(north of 36°N) oscillates between a weak net source (22 Tg) during January-March and a netsink (87 Tg) during July-September. Interestingly the period of maximal net absorption is thesame as that of maximal SW Monsoon-forced emissions from the Arabian Sea reflecting thedominance of removal by CO2-undersaturated waters, largely due to biological processes, indetermining the net CO2 uptake/emission balance in the region. The study of Sabine et al. (2000)did not cover the frontal zone, which is an even stronger sink of atmospheric CO2. For example, Metzl et al. (1997) estimated that the sub-Antarctic waters (40-50°S) in the Indian sector absorb 420 TgC annually; this compares favourably with the model-derived estimate of Louanchi et al. (1996) for the region south of 35°S. Thus, it may be concluded that the Indian Ocean, includingthe sub-Antarctic Zone, on balance absorbs a minimum of 0.5 Pg CO2-C from the atmosphereannually, a quarter of the total oceanic CO2 uptake (Houghton et al., 1996). It may be noted that the average annual increase (4.1/18 = 0.23 Pg C y-1) in anthropogenic CO2 inventory north of35°S estimated by Sabine et al. (1999) is of the same order.
Molecular Nitrogen and Nitrous Oxide
The Arabian Sea is one of three major oceanic water-column denitrification sites. Currentestimates of denitrification rates in this region vary from 12 TgN y-1 (Mantoura et al., 1993) to33 TgN y-1 (Naqvi and Shailaja, 1993). The high-quality nutrient data collected during theJGOFS cruises were expected to narrow this gap. Howell et al. (1997) combined the nitrate and chlorofluorocarbon (CFC-11) data collected during the WOCE programme to arrive at adenitrification rate of 21 TgN y-1. This is slightly lower than most other estimates (the mostrecent one being 33 TgN y-1 by Bange et al., 2000). Thus, it would appear that the Arabian Sea accounts for at least one-third of the global oceanic water column denitrification (Naqvi, 1987).This high rate of molecular nitrogen (N2) production is countered by substantial N-fixation in the region as evident from recent measurements of natural abundance of nitrogen isotopes indissolved nitrate (NO3
-) (Brandes et al., 1998). Brandes et al. (1998) observed that near-surfaceNO3
- in the Arabian Sea suboxic zone has a nitrogen isotopic composition close to the oceanicaverage (δ15N ≈ 6 per mil) in spite of the huge denitrification-related enrichment of 15N (δ15N>11 per mil) in subsurface waters (>200 m). This has been attributed to the fixation of lightnitrogen at an estimated basin-wide rate of 6 Tg y-1. This implies a net loss of combined nitrogen and production of N2 at a rate of ~15-27 TgN y-1.
Apart from denitrification and nitrogen fixation, the other important term in the nitrogen budget to which the Arabian Sea makes a significant contribution, is the efflux of N2O from the seasurface. Of the total inputs of N2O to the atmosphere from all natural sources (~12 TgN-N2O y-
1), at least one-third seems to come from the oceans (Nevison et al., 1995). However, emissions of N2O are not uniformly distributed over the oceanic surface with the eastern-boundaryupwelling zones underlain by O2-deficient waters making disproportionately large contributions(Codispoti and Christensen, 1985; Suntharalingam et al., 2000). Studies carried out underJGOFS have led to a better quantification of this efflux. These studies (e.g., Figure 5) revealed

19
very high surface saturations (>300%) in the upwelling zones off Oman, Somalia andsouthwestern India (Bange et al., 1996; de Wilde and Helder, 1997; Patra et al., 1999).
Figure 5. N2O of equilibrated seawater along the same transect as in Figure 1 (lower panel)(from Bange et al., 1996).
The sea-to-air fluxes of N2O from these regions are also similarly high (maximum of ~500 µmolm-2 d-1). In the open ocean, surface N2O saturations and its fluxes to the atmosphere aregenerally lower (Figure 5) than those reported previously (Law and Owens, 1990; Naqvi andNoronha, 1991). These also show smaller temporal variability except for the zones of offshoreupwelling and winter mixing (Bange et al., 1996; de Wilde and Helder, 1997; Lal and Patra,1998). However, even in coastal upwelling zones, seasonal changes in the mixed layer N2Osaturations may sometimes be small, as observed by Upstill-Goddard et al. (1999) in the western Arabian Sea, indicating a balance between the processes of gas exchange, net production andsupply/removal by advection and vertical mixing. Nevertheless, large changes in air-seaexchange can still occur due to variations in the wind speed (Upstill-Goddard et al., 1999).However, the highest globally recorded N2O concentrations in surface water (up to 436 nM or8,250% saturation) were measured within the seasonally-suboxic zone off western India during a three-year (1997-99) study. This study was conducted as part of the Indian LOICZ (Land-OceanInteractions in the Coastal Zone) programme (Naqvi et al., 2000). These extreme concentrations are higher than those observed (62.5 nM or 953% saturation) during a more limited survey made during the SW Monsoon of 1995 (Naqvi et al., 1998). Interestingly, the highest concentrationswere found in waters that were apparently affected by denitrification, as suggested by highnitrite concentrations (see Figure 6), a process that generally results in a net consumption ofN2O (Codispoti and Christensen, 1985).
This is consistent with the results of incubation experiments (Naqvi et al., 2000), whichindicated that in shallow, denitrifying systems with high rates of denitrification and experiencing frequent aeration due to turbulence, the N2O-reductase activity may be suppressed to such adegree that a large fraction of the nitrogen undergoing bacterial reduction ends up as N2O (Naqvi et al., 2000).

20
Figure 6. Plot of nitrite (NO2-) versus nitrous oxide (N2O) in the shallow (open circles) and deep
(closed circles) suboxic zones of the Arabian Sea (from Naqvi et al., 2000).
Estimates of the annually integrated N2O-flux from the Arabian Sea, based on the JGOFS data,range between 0.16 and 1.5 Tg N2O y-1. These estimates are strongly dependent on the choice of model used to compute the air-sea exchange coefficient and the interpolation scheme. Recently, Bange et al. (submitted) have pooled all the data generated between 1977 and 1997 to computehigh resolution (1°x1°) seasonal and annual N2O concentration fields in the surface layer. Theannual emission rate, estimated to range between 0.33 and 0.70 Tg N2O, seems to be dominatedby fluxes from coastal regions during the SW Monsoon. However, this data set does not include the anomalously high N2O concentrations in the inner shelf off the Indian west coast. In thisregion high oxygen demand coupled with strong stratification causes severe oxygen depletionbelow the shallow pycnocline. But while the causative processes (upwelling of nutrient-richwaters and the outflow of freshwater from land resulting from intense rainfall in the coastalzone) are primarily natural, it is quite likely that the coastal hypoxic system has furtherintensified in recent years. Such intensification, evident from the previously unreportedoccurrence of hydrogen sulphide, might arise from an increase in productivity fuelled by ahigher flux of nutrients from land. This could have contributed to the enormous N2Oaccumulation observed in the region. But if the N2O concentrations in the region were high even before this intensification, then the above estimate of “natural” N2O emissions from this regionwould be too low. In any case, if we take into account the seasonal flux of N2O from the shelf(0.06-0.39 Tg N2O; Naqvi et al., 2000), the current sea-to-air flux of N2O from this region could be as much as 1 Tg N2O y-1, about one quarter of the total global oceanic emissions (Houghtonet al., 1996).
Ammonia and Methylamines
Under the UK JGOFS programme, measurements of ammonia/ammonium (NH3/NH4+) and
methylamines (MAs) [monomethylamine (MMA), dimethylamine (DMA) and trimethylamine(TMA)] were made in seawater, atmosphere and rainwater in the central and western ArabianSea during two seasons (August-October and November-December, 1994) (Gibb et al., 1999).
0 200 400 600N2O (nM)
0
4
8
12
16
NO
2-(µ
M)

21
Concentrations of all species were generally elevated in the coastal upwelling zone off Omanrelative to those measured in the central Arabian Sea, with NH3/NH4
+ levels being up to twoorders of magnitude higher than those of the MAs. MMA was the most abundant MA, beingpresent in concentrations reaching up to 66 nM, while TMA levels did not exceed 4 nM (see also Burkill, this volume). Both the coastal and offshore waters of the Arabian Sea appeared to emitNH3 to the atmosphere with large temporal variations. However, the air-sea exchange probablyrepresented a small proportion of the aquatic turnover of NH3 and MAs, which seem to becontrolled mainly by the structure, activity and seasonality of the microbiological cycle. Most of the removal of NH3 and MAs appeared to occur through wet deposition (Figure 7), contributing little (<1%) to the new production. However, by virtue of being the dominant atmospheric bases, NH3 and MAs may play an important role in tropospheric chemistry and climatic modification.
Figure 7. Schematic diagram of the cycling of NH3+NH4+ in the remote Arabian Sea in whichpools (boxes, nmol m-3) and processes (arrows, nmol m-3 d-1) are represented in broadapproximation to their relative magnitudes (from Gibb et al., 1999).
Nitrate and Lead-210
As a part of the Indian JGOFS programme, seasonal distributions of NO3- and lead-210 (210Pb)
have been studied in the aerosols over the central-eastern Arabian Sea for investigating thesources of NO3
- and quantifying its deposition flux (Sarin et al., 1999). Higher NO3-
concentrations (0.4 to 4.1 µg m-3) were observed during the Spring Intermonsoon and NEMonsoon as compared to the SW Monsoon (0.2-0.8 µg m-3). Using a NO3
-/210Pb ratio of 43 µgdpm-1 and the measured 210Pb deposition flux of 20 dpm m-2 d-1, the atmospheric deposition ofNO3
- was estimated to be 1 mg m-2 d-1. These results suggest that that the atmospheric NO3-
deposition plays an insignificant role in fuelling new production in the region.
Methane
Prior to the JGOFS programme, there had been only one report on CH4 distribution in theArabian Sea (Owens et al., 1991). High surface-water concentrations and consequently larger-than-the-oceanic-average CH4 fluxes observed in this study prompted additional investigationson CH4 by several groups, the results of which have been published recently (Patra et al., 1998;

22
Bange et al., 1998; Upstill-Goddard et al.¸ 1999; Jayakumar et al., 2001). In agreement with the findings of Owens et al. (1991), Patra et al. (1998) reported high surface saturations, averaging140, 173 and 200% during the Northeast Monsoon, Spring Intermonsoon and SW Monsoonperiods in the central and eastern Arabian Sea. Upstill-Goddard et al. (1999) observed moderate saturations in the Omani upwelling zone both during the late SW Monsoon (170±55%) andSpring Intermonsoon (179±15%) seasons; the corresponding open ocean values were 130±5%and 115±2%. In contrast to these data, Bange et al. (1998) showed near-saturation (103-107%)in the central Arabian Sea, rising to a maximum of 156% in the upwelled water near the coast of Oman (Figure 8). The annual emission rate of CH4 from the Arabian Sea was estimated at0.032-0.053 Tg CH4 y-1 by Patra et al. (1998), 0.011-0.020 Tg CH4 y-1 by Bange et al. (1998)and 0.1-0.18 Tg CH4 y-1 by Upstill-Goddard et al. (1999). Whether the order-of-magnitudedifferences in the published estimates of CH4 emissions from the Arabian Sea represent naturalvariations or they arise, at least in part, from systematic measurement errors is still not clear.However, even the highest of these estimates is probably still <2% of the estimated emission rate from the oceans as a whole (Upstill-Goddard et al., 1999), which itself is at the most 4% of the atmospheric flux of CH4 from all natural and anthropogenic sources (Crutzen, 1991). Evidently, unlike N2O, CH4 emissions from the Arabian Sea are not globally significant.
It should be noted that most of the CH4 data collected in the Arabian Sea have come fromoffshore areas. As pointed out by Bange et al. (1994), the contribution of the coastal zone to the oceanic CH4 budget may be disproportionately large. This is also supported by the results of arecent study (Jayakumar et al., 2001), which yielded CH4 saturations comparable to thoseobserved by other workers in the open ocean, but very large supersaturations (up to ~2500%) in the coastal waters affected by monsoonal runoff from land. Still higher supersaturations (up to~13,000%) were recorded in the estuarine surface water pointing to a source of CH4 in coastalwetlands. Elevated (CH4) concentrations (~50 nM) have also been found in the seasonal oxygen-depleted zone, when the sub-pycnocline waters turned fully anoxic (D.A. Jayakumar,unpublished data). However, the impact of these processes on the CH4 budget of the Arabian Sea still remains to be evaluated fully (see also Burkill, this volume).
Dimethylsulphide
As part of the UK and German JGOFS cruises and a related programme in India, measurements of dimethylsulphide (DMS) have been made for the first time in the Arabian Sea. Although thecomplete data sets generated during these studies have not yet been made available, large spatial and temporal changes in the concentrations of DMS, and its precursor and oxidation product,dimethylsulphoniopropionate (DMSP) and dimethylsulphoxide (DMSO), respectively, appear to occur in this region. For example, in the productive surface waters of the Omani upwelling zone, sampled by Hatton et al. (1999), marked increases in the concentrations of DMSP (on averagefrom 21.5 to 37.5 nM), DMSO (from 2.9 to 7.7 nM), and DMS (from 1.3 to 3.3 nM) wereobserved over an interval of three weeks during the transition from SW Monsoon to FallIntermonsoon, presumably caused by a shift in the dominant phytoplankton groups (fromdiatoms to prymnesiophytes). The concentrations were relatively low (21.0, 4.3 and 2.1 nM,respectively) in the oligotrophic surface waters of the central Arabian Sea. DMSO occurred infairly high and rather uniform concentrations throughout the water column, and although itsproduction through photo-oxidation in the surface layer probably represented an important sinkof DMS, significant amounts of the latter should be emitted to the atmosphere from this region.This flux remains to be quantified, and for this purpose representative data from differentbiogeochemical provinces (particularly those which support the growth of the DMS-producingorganisms such as coccolithophores and Phaeocystis) are needed.

23
Figure 8. CH4 saturations (solid curves) and sea surface temperature (dashed curves) in thewestern Arabian Sea during June-July 1997. The transect ABC was located offshore outside the upwelling zone while the drift stations experienced upwelling (from Bange et al., 1998).
Recommendations
Due to the delicate biogeochemical balance that exists in the Arabian Sea, minorclimatic/environmental perturbations can have potentially large impacts on chemical fluxes,especially of greenhouse gases in the region. It is for this reason that the Arabian Sea is regarded as a sensitive ocean scale barometer of global climate change (Mantoura et al., 1993). Recentresults suggest that the human activities may already be modifying such fluxes, even though itcannot be established whether the effects observed are due to local processes (eutrophication)alone or caused by slight modifications of physical forcing functions (circulation and freshwater runoff) (Naqvi et al., 2000). Nonetheless, since these fluxes are globally significant, we mustkeep a close watch on both the coastal and open ocean oxygen-deficient zones, fully understand the mechanisms of biogeochemical transformations using modern tools (for example, stableisotopes and molecular probes), identify the emerging trends and develop capability to predictfuture changes. In spite of the recent focus on this region under several internationalprogrammes, there still exist some serious deficiencies in the geographical and temporalcoverage that need to be remedied. For example, the Bay of Bengal still remains to beadequately sampled (especially along the Burmese and Bangladeshi coasts). Also, thecontribution of benthic cycling to greenhouse gas fluxes is still largely unknown. As anothermajor international programme is unlikely to be launched in this region in near future, seriousefforts should be made towards capacity building such that the less-developed coastal states can initiate studies in their coastal zones.

24
2.4 Primary Productivity in the Arabian Sea
John Marra
Lamont-Doherty Earth ObservatoryColumbia University, Palisades, NY 10964
USA
Introduction
Between 1994 and early 1996, the JGOFS programme conducted a comprehensive study of theArabian Sea not seen since the International Indian Ocean Expedition of the early 1960’s (seeKrey and Babenerd, 1976). Combining all the primary production observations, the Arabian Sea is now one of the better-sampled regions of the ocean in terms of organic productivity. The data collected during the various JGOFS programmes consist of photosynthesis versus irradianceresponse curves (Sathyendranath et al., 1999; Johnson et al., in review), standard primaryproductivity measurements (assimilation of 14C) (Mahdupratap et al., 1996; Barber et al., 2001;Savidge and Gilpin, 1999), community production measurements (light-dark bottle O2)(Robinson and Williams, 1999; Dickson et al., 2001), and gross production estimates (Dicksonet al., 2001). The data sets also include measurements that further support the productivitystudies. These include phytoplankton absorption cross-sections (Marra et al., 2000), the varietyof phytoplankton pigments (Latasa and Bidigare, 1998), the variety of phytoplankton taxa(Garrison et al., 1998, 2000), and on certain of the cruises, spectral irradiance as a function ofdepth (Pinkerton et al., 1999; Marra et al., 2000). All the observations are spatially extensive in the eastern and central parts of the Arabian Sea, and span August 1994 through December 1995 at nearly monthly time intervals.
The JGOFS programmes have uncovered new findings about the Arabian Sea, as well asreinforced some of the previous suppositions about the effects of the monsoons on thephytoplankton and their rates of production. In this short report I focus on the measurements ofproductivity from 14C assimilation, productivity as estimated from variables measured frommoored sensors, and the mixed layer depth as an indicator of the extent of mixing in the watercolumn. For the comparison of shipboard measurements, I compiled data that was readilyavailable in the published literature or tabulated on web sites.
Prior Work
Our sense of progress can be appreciated by examining what we thought we knew about theproductivity cycles in the Arabian Sea before embarking on the JGOFS programmes in 1994 and 1995. Sharon Smith compiled what was then known into a report (U.S. JGOFS, 1991). That the Arabian Sea is productive, by virtue of the semi-annual monsoons and important fisheries, wasnot in question. The theoretical basis (e.g., Bauer et al., 1991), supported by a small number ofcruises to the region, was that the summertime Southwest (SW) Monsoon, sometimes called the Findlater Jet (Findlater, 1971), blowing toward Asia, would generate upwelling near the coast of Oman. Further offshore, the changing curl of the wind-stress on either side of the Findlater Jetwould produce an Ekman divergence region and upwelling to the north of its axis. South of theJet axis, however, the curl of the wind-stress would be such as to produce a convergence,downwelling, and deep mixed layers. The effect of the winter Northeast (NE) Monsoon, coming off the Asian continent, was believed to be minor, perhaps driving upwelling near the coasts ofIndia and Pakistan.
According to the earlier literature, the SW Monsoon dominates seasonality in the Arabian Seaalmost entirely. What we knew of the biology seemed to reinforce this idea. The ocean colour

25
data from CZCS show large increases during August and September, a few months following the onset of the SW Monsoon in late May and June, (Banse, 1987; Brock et al., 1991). Because ofcloudiness (and perhaps dust), however, there are no satellite images for most of June and July,thus the oceanic response to the SW Monsoon was almost unknown. Brock et al. (1993) modelthe seasonal change in primary production, extrapolating both from a chlorophyll profile duringthe Spring Intermonsoon, and from the composite ocean colour images for each of the seasons.They predict a seasonal productivity cycle with a maximum in August and September, and atover 2 g C m-2 d-1, four- or five-fold greater than the rest of the year. Yoder et al. (1993)completed a more comprehensive analysis of the CZCS data for the tropics in general, and alsofound a peak in chlorophyll a biomass dominating the Indian Ocean at the end of summer (SWMonsoon). The primary productivity estimated by the satellite imagery appears to be a multipleof the seasonal variation in chlorophyll biomass (Brock et al., 1993; Howard and Yoder, 1997).
The geographic and seasonal variability in primary production is, to a large extent, driven byvariations in water column mixing, as indicated by the depth of the mixed layer. Water columnmixing controls the availability of irradiance and affects the flux nutrients to the phytoplanktonat the surface. Like other areas of the world ocean, the re-stratification and vertical mixing of the water column is central to the seasonal cycle in the Arabian Sea (Sathyendranath and Platt 1994,1999; McCreary et al., 1996). The relationship between productivity and mixed layer depth,however, is not direct. Deep mixed layers may not allow enough irradiance for phytoplankton to accumulate when they are mixed over great depth. In shallow mixed layers, on the other hand,nutrients are depleted more quickly than they can be replenished by mixing, and productivitywould subsequently decline. Conditions that provide marginal or transient stability to the watercolumn are optimal for phytoplankton productivity.
Very little hydrographic data for the Arabian Sea exist prior to the JGOFS effort. An offshoresection of density after the SW Monsoon shows an increase in mixed layer depth going offshore, expected from the changing sign of the wind-stress curl (Bauer et al., 1991). Seasonally,Sathyendranath and Platt (1994) suggest how chlorophyll biomass and mixed layer depth mayinteract, using data from the CZCS and climatologies of hydrography. Given upwelling to thenorth of the Findlater Jet, and downwelling to the south, however, the Arabian Sea might beexpected to resemble upwelling regimes in eastern boundary currents. The difference in theArabian Sea is, of course, the annual reversal of winds.
In summary, the historical data clearly indicated the importance of the monsoons to theoceanography of the Arabian Sea. The effect of the SW Monsoon is to divide the Arabian Seainto two zones, upwelling nearer the Arabian Peninsula, and downwelling offshore. The CZCSdata provided tantalizing glimpses of the potential for productivity in the Arabian Sea andimportant ideas, but little information on the means by which environmental forcing governedthe seasonal cycle, and less still of the actual magnitudes of productivity. In reviewing thefindings of the recent programmes, I highlight the seasonal variability and the changes on- andoffshore, both in the context of a changing mixed layer depth as the indicator of water columnmixing and re-stratification. I divide consideration of the data into temporal and spatialvariability. The analysis here suggests (1) that, seasonally, productivity is less dominated by the SW Monsoon period, and (2) that productivity is less variable going offshore from the ArabianPeninsula.
Temporal Variability
For temporal variability in the Arabian Sea, I rely on an estimate from the bio-optical sensorsmoored in the central Arabian Sea from October 1994 to October 1995 (see Dickey et al., 1998). The mooring serves as a context for comparison with other data over the seasonal cycle. The

26
optical estimate of productivity is based on moored observations of chlorophyll (Kinkade et al.,2000) and surface irradiance (Weller et al., 1998). The governing equation is,
P = φ(E)• a*ph•(z)• Chl-a •E(z)
where P is the rate of production (mol-C m-3 d-1), φ(E) is the quantum yield for photosyntheticcarbon fixation (mol-C/mol-photons), a*ph is the mean (over wavelength) chlorophyll-specificabsorption cross-section (m2 mgChl a-1), Chl-a is chlorophyll a (mg m-3), and E is the total daily irradiance (mol-photons m-2 d-1) at depth z. I plot the data regardless of the year in which thedata were collected in order to see patterns in the seasonality in the productivity. The danger ofthis, of course, is that there will be inter-annual variability in the magnitude and timing of themonsoons. Also, mesoscale phenomena can cause variability among the various estimates.
The variability in chlorophyll a and primary productivity, seasonally, shows large variations, but not what might have been expected based on prior work (Figure 1). First, there are mesoscalephenomena present in the mooring records (Dickey et al., 1998), and also in the shipboard data(Barber et al., 2001). The mooring recorded two major eddy-like features, one in December1994, and a second in August 1995. The December eddy, in particular, advected highchlorophyll a concentrations through the mooring site. Second, the productivity observed during the NE Monsoon was much higher than anticipated (Mahdupratap et al, 1996). Convectivemixing during the NE Monsoon (Weller et al., 1998; Wiggert et al., 2000) turned out to be animportant source of nutrients, which leads to enhanced rates of production over that expectedfrom prior analysis of CZCS imagery. The calculation in Marra et al. (1998) shows a steadyincrease in productivity with the wind-driven deepening (Weller et al., 1998) of the mixed layer with the progression of the SW Monsoon in June and July. The maximum in chlorophyll a andproductivity occurs in August and September following the cessation of mixing. There is not adirect relationship between mixed layer depth and the concentrations of chlorophyll a.Chlorophyll a increases after the water column re-stratifies following the cessation of monsoonwinds, and declines during the shallow mixed layers of the Intermonsoon periods. Interestingly,the data of Ryther et al. (1966) are consistent with the JGOFS results.
There is variability in the shipboard productivity data. However, when plotted againstchlorophyll a, nearly all the data fall within an acceptable range (Figure 2). The data fromOwens et al. (1993) are the exception. This is the closest data point to the position of themooring, but a nearby station shows almost twice the productivity for the same amount ofchlorophyll a, and agreement with the other data. Overall, I conclude that the magnitude ofcarbon assimilation is being consistently estimated, and that the pattern that has emerged reflects at least the seasonal variability in chlorophyll a biomass and primary productivity.
The mooring productivity estimates (Figure 1) were accomplished using a constant absorptioncoefficient for phytoplankton, a*ph (the mean value over wavelength) and a constant maximum(realizable) quantum yield for carbon assimilation, φmax. Thus, productivity in those estimatesonly varied as a function of chlorophyll concentration, irradiance (and consequently, the depth of the euphotic zone), and mixed layer depth (Marra et al., 1998). The reason for the constancy ina*ph and φmax was that the variability of these parameters is little understood, especially at theday-to-day time scale at which we can obtain estimates of chlorophyll and irradiance. Therefore, we decided to keep them constant. However, other data show that these parameters change,certainly between the monsoons (Sathyendranath et al., 1999; Johnson et al., in press), andperhaps over shorter time periods as well. It is possible that changing bio-optical properties ofthe phytoplankton are responsible for the differences between the mooring estimates and theshipboard data, especially at low biomass and productivity (Figure 2). There is a need in future studies to understand how phytoplankton absorption and quantum yields change in response toenvironmental conditions at greater resolution than has heretofore been measured.

27
(a)
(b)
Figure 1. (a) Chlorophyll a (mg m-2) as a function of day number for 1994 (after vertical line)and 1995 (before vertical line). Data shown are from the mooring (15° 30’ N, 61° 30’ E) (dots), the Arabian Sea Expedition (16° 12’N, 62° W) (squares), Arabesque (19° N, 62° E: circles; 19°N, 64° E, diamonds), and from Madhupratap et al. (1998) (15° N, 64° W: triangles). (b) Carbon assimilation (mmol-C m-2 d-1) as a function of day number [symbols as in (a)]. Mixed layerdepth (MLD) estimated from the mooring data is also shown.
Onshore-Offshore Variability
There are three features of the spatial variability in the Arabian Sea that result in large part from the monsoons. First, sea-surface temperature (SST) always increases offshore (Gundersen et al.,1998). During the SW Monsoon, there is a particularly large jump from the coastal stations,followed by smaller increases further offshore. Nevertheless, despite the season, SST alwaysincreases offshore. Second, mixed layer depths always increase offshore as well (Figure 3),indicating a downward tilt to the pycnocline going in the offshore direction. Third, surfacenutrient concentrations almost always decrease offshore (Morrison et al., 1998; Woodward etal., 1999), although during the Northeast Monsoon in 1995, there was a maximum in surfacenitrate at 16 °N (700 km offshore) (Dickson et al., 1999).
0
10
20
30
40
50
60
70
80
0 50 100 150 200 250 300 350
Day Number
Ch
l-a
19941995
-100
-50
0
50
100
150
200
250
0 50 100 150 200 250 300 350
Day Number
C A
ssim
ilati
on
19941995
MLD

28
Figure 2. Carbon assimilation as a function of chlorophyll a for the data from the Arabian Sea.
All of these observations point to the large-scale upwelling regime inshore of the geographicaxis of the Findlater Jet. Coastal upwelling occurs nearshore, and Ekman driven upwellingoccurs over the broad reaches of the offshore ocean. South of the Findlater Jet, and in the region where the curl of the wind-stress becomes negative, convergent circulation and downwellingmaintain deep mixed layers. While there is evidence for substantial eddy variability in theNorthern Arabian Sea (Flagg and Kim, 1998), the overall trends are preserved in their meanvalues in the transect data on the line offshore proceeding southeast from Ras Madraka, Oman.To be sure, there is considerable noise in the mixed layer depth data, but the downward trend is preserved in all seasons. In addition to mesoscale phenomena, the variability can also be caused by instrument or observational error, or by internal waves.
In contrast to what might be expected, productivity does not decrease in the offshore direction(Savidge and Gilpin, 1999; Barber et al., 2001) despite the increasing mixed layer depths and the decline in nutrient availability. The reason is most certainly the physiological adaptation in theoffshore communities. The suite of pigments is altered in the offshore direction toward thatindicative of greater dominance by picoplankton and other small forms (Latasa and Bidigare,1998; Barlow et al., 1999). Marra et al. (2000) noted that the pigment suite in the offshorepopulations also conferred an adaptation to higher irradiances, by having proportionately morepigments used for photoprotection. The advection of coastal waters in upwelled filaments(Latasa and Bidigare, 1998; Barber et al., 2001) is another factor that can complicate theinterpretation of the offshore trends. However, the larger effect is the changing photosyntheticcharacteristics in the offshore populations, which permits relatively high rates of production inthese well-lit, but relatively nutrient-poor areas.

29
Figure 3. Estimates of the mixed layer depth as a function of latitude (°N) in the Arabian Sea.The circles are the mean depth for all the cruises in the Arabian Sea Expedition in 1994-1995,and the error bars represent the standard deviation from the mean. The individual estimates areplotted as small diamonds.

30
2.5 Remote Sensing of Ocean Colour in the Arabian Sea
Louisa Watts and Shubha Sathyendranath
Biological Oceanography Section, Bedford Institute of OceanographyP.O. Box 1006, Dartmouth, Nova Scotia, B2Y 4A2
Canada
Introduction
It is no exaggeration to state that the availability of remotely sensed data on surface-oceanproperties from satellites has revolutionised oceanography. The vantage point from space, which earth-orbiting satellites provide, has allowed spatially detailed measurements to be made almostinstantaneously over wide areas and at regular frequency allowing monitoring of the ocean on a global scale. Sea-surface colour, temperature, height and roughness measurements can becollected by remote-sensing techniques, thus providing synoptic information on biological andphysical processes and opening up the possibility of a multidisciplinary approach to marineecosystem studies. This chapter considers the advances made in remote-sensing of the ArabianSea in a biogeochemical context, so that we are mainly focussing on advances within the remote sensing of ocean colour and its applications (but see Capone et al., 1998; Böhm et al., 1999; Shi et al., 1999 for applications of remotely-sensed sea-surface temperature and sea-surface height in the Arabian Sea).
Biological oceanography has been revolutionised by the provision of ocean colour data thatprovide us with a two-dimensional window into the synoptic state of the pelagic ecosystem (Platt et al., 1995), and an opportunity to calculate ocean-basin scale primary production from biomass fields. At the time of writing JGOFS Report 17 (1995), the only available ocean colour data had been collected by the Coastal Zone Colour Scanner (CZCS) for the period 1978-1986. Thelaunch of the state-of-the-art, ocean colour satellite sensor, SeaWiFS (Sea Wide Field-of-ViewSensor), had been delayed until mid-1995 but the ocean colour data collected thereafter were tobe available for the JGOFS Arabian Sea Process Study. This Process Study was thus planned to be “the first test of the JGOFS goal to relate remotely-sensed variability to oceanic processes of the carbon cycle” (JGOFS Report 17, 1995). Unfortunately, the SeaWiFS sensor was notlaunched until August 1997, leaving the JGOFS programme without remotely sensed oceancolour data.
Despite the lack of SeaWiFS ocean colour data during the JGOFS cruises to the Arabian Sea,significant advances have been made within the JGOFS framework, both in the area of algorithm development and in the area of validation of the quality of the retrieval of phytoplanktonpigment data from SeaWiFS. Advances have also been made in protocol-development for thecomputation of primary production using satellite-observed phytoplankton biomass fields andbio-optical properties of the surface and near-surface waters. In this chapter, the progress madein these areas is reviewed. Furthermore, some recommendations are made for building on theJGOFS experience for remote sensing in the Arabian Sea.
Remote-sensing Algorithms: The Retrieval of Chlorophyll a Biomass and FluorometricPigment Concentration
Standard protocols for processing remotely sensed ocean colour data to derive the water-leavingradiance signal at several wavebands involve a correction for atmospheric noise. These multi-waveband data are then processed to obtain a measure of pigment concentration in the surfacewaters. The accuracy of this information depends on the suitability of algorithms used to convert the water-leaving radiance values into estimates of chlorophyll a and fluorometric pigment

31
concentration. These algorithms are based on the influence of pigments on phytoplanktonabsorption and hence on ocean colour.
As part of the U.S. contribution to JGOFS, Kinkade et al. (2001) have developed an empiricalalgorithm relating the ratio of downwelling irradiances at 443 nm to 550 nm (blue to greenwavelength ratio) with the integrated chlorophyll a concentration through the photic zone (r2 =0.73). Data were determined from moored irradiance and fluorescence measurements, collectedover a year in the Arabian Sea.
Using an essentially empirical approach, Aiken et al. (1995) have developed a number of multi-band algorithms for the retrieval of chlorophyll a and total fluorometric pigment concentrationfrom SeaWiFS data. These algorithms are based on two ratios of water-leaving radiances: the443:555 ratio and the 490:555 ratio and were an attempt to produce global chlorophyll a andfluorometric pigment algorithms using two band ratios. As part of the ARABESQUEprogramme (the UK contribution to JGOFS Arabian Sea Process Studies), Pinkerton et al.(1999) tested the performance of the Aiken et al. (1995) algorithms in the northwestern IndianOcean. Shipboard measurements were made of the depth-varying upwelling and downwellingirradiances in parallel with phytoplankton pigment concentrations during a SW Monsoon and an Intermonsoon period. This data set was used to test the validity of the algorithms described byAiken et al. (1995). The algorithms based on the 443:555 band-ratio estimated near-surfacechlorophyll a and fluorometric pigment concentrations satisfactorily, i.e., with an accuracy of ±35% of the actual value, in accordance with the goal set within the SeaWiFS mission statement.In contrast, the mean errors from the algorithms based on the 490:555 band-ratio were greaterthan 35% over both seasons. This was attributed to the sensitivity of these algorithms to seasonal and regional changes in the proportions and composition of photosynthetic and photoprotectivecarotenoid pigments present in the phytoplankton. Such variability can arise from seasonal andregional changes in ambient light levels (Bricaud et al., 1995; Johnsen and Sakshaug, 1996;Culver and Perry, 1999; Stuart et al., 2000; Marra et al., 2000) and from changes in the physical forcing, with subsequent changes in phytoplankton population composition.
Indeed, prior to the JGOFS programme, little was known about the absorption characteristics of phytoplankton in the northwestern Indian Ocean. As a result of the Netherlands Indian OceanProgramme, the ARABESQUE and U.S. JGOFS cruises, this database has been substantiallyincreased: light absorption by phytoplankton has been measured during the NE Monsoon(Sathyendranath et al., 1996); SW Monsoon and November (Fall) Intermonsoon(Sathyendranath et al, 1999); and Spring Intermonsoon and NE (winter) Monsoon (Marra et al., 2000). Examination of these data sets has taught us that the intra-annual variability of thephysical forcing in the Arabian Sea, brought about by the monsoon system, causes changes inthe phytoplankton composition and cell size (Campbell et al., 1998; Garrison et al., 1998; Brown et al., 1999; Savidge and Gilpin, 1999; Tarran et al., 1999; Banse and English, 2000;Shalapyonok et al., 2001) and in the pigment make-up of these cells (Latasa and Bidigare, 1998; Barlow et al., 1999; Banse and English, 2000). This, in turn, gives rise to changes in theabsorption properties of the phytoplankton and consequently in ocean-colour (Sathyendranath etal., 1996; Sathyendranath et al., 1999; Marra et al., 2000; Stuart et al., 2000). Present universalalgorithms for the retrieval of chlorophyll a biomass and fluorometric pigment concentration, as described above (Aiken et al., 1995), are inadequate to incorporate such time- and space-variantbio-optical properties of the surface and near-surface waters found in the Arabian Sea. It istherefore desirable that these algorithms be tuned to match the optical properties of thephytoplankton present in the area and to incorporate their geographical and seasonal variability(see Recommendations). The practical implementation of a patchwork of regional and seasonal algorithms would, however, pose some problems that merit more attention.

32
Retrieval of Diffuse Attenuation Coefficient
Kinkade et al. (2001) developed an empirical algorithm relating the ratio of downwellingirradiances at 443 nm to 550 nm with the average vertical attenuation coefficient forphotosynthetically active radiation (PAR; r2 = 0.845). Pinkerton et al. (1999) have shown thatthe diffuse attenuation coefficient at 490 nm may be estimated robustly from remotely senseddata using the ratio of normalised water-leaving radiance at 443 nm to that at 555 nm and thealgorithm developed by Austin and Petzold (1981), with the 555 nm band used instead of the550 nm band.
Primary Production Estimates by Remote-sensing of Biogeochemical Properties
Another application of remotely sensed ocean colour data is in the computation of primaryproduction on large horizontal scales: maps of surface ocean biomass can be derived fromremotely sensed data on ocean colour. These maps can then be combined with a spectral andangular model of submarine light coupled with a model of the spectral response of algalphotosynthesis to compute oceanic primary production (Platt and Sathyendranath, 1988;Sathyendranath et al., 1989; Sathyendranath et al., 1995). In addition to satellite-derivedinformation on phytoplankton biomass, this processing of maps requires specification of thephotosynthetic characteristics of the phytoplankton: namely, the parameters of thephotosynthesis-irradiance (P-I) curve and information on the vertical distribution of biomass.These data are accumulated by observations from ships and so tend to be sporadic in both space and time. Prior to the JGOFS initiative, little was known about the magnitude and variation ofthe P-I parameters in the northwestern Indian Ocean. As a result of the ARABESQUE and U.S. JGOFS cruises, the database for this area has been supplemented substantially. Observationsshow that there are temporal differences in the assimilation number and the slope of the P-Icurve, between the Monsoon and the Intermonsoon seasons, although these differences are notas strong as those observed for the chlorophyll-specific absorption coefficient (Sathyendranath etal., 1996; Sathyendranath et al., 1999). Spatial differences in the P-I parameters have also beenobserved between filaments and adjacent waters during the Fall Intermonsoon (Toon et al.,2000), which were associated with differences in the phytoplankton community structure.Photosynthesis-Irradiance parameters measured during the Netherlands Indian OceanProgramme, SW Monsoon and Spring Intermonsoon 1992, showed both spatial and temporalvariability (Kromcamp et al., 1997).
Watts et al. (1999) have used such shipboard data collected during the ARABESQUE cruises(Sathyendranath et al., 1999) to successfully compute integrated primary production and newproduction over a Monsoon and Intermonsoon period for the northwest Indian Ocean basin.They combined archived ocean colour data (CZCS), the P-I and absorption data (Sathyendranath et al., 1999), vertical biomass data (Burkill, 1999), nitrogen assimilation data (Watts and Owens, 1999) and the models as described above. The protocols developed by Watts et al. (1999) for the computation of primary and new production incorporate the intra-annual variability ofphytoplankton growth within the Arabian Sea by partitioning the area into a system of “dynamic biogeochemical provinces” (see below).
The Development of “Biogeochemical Provinces” in the Arabian Sea
The decomposition of an ocean basin into a suite of regions where the area enclosed by theboundary of any region is under common physical forcing with respect to the factors thatinfluence the growth of the phytoplankton is called a partition into a system of “dynamicbiogeochemical provinces” (Longhurst et al., 1995; Longhurst, 1998; Platt and Sathyendranath,1999). The underlying concept is that provinces can be used as a template for the assignment of

33
the parameters of the P-I curve, and of the function describing the vertical distribution ofphytoplankton biomass, for the modelling of oceanic primary production. According to thisconcept, every pixel on an ocean colour image will be assigned parameter values according tothe province to which they are assigned. Primary production can then be computed for that pixel, allowing horizontal extrapolation of shipboard data to ocean-basin scales. Some desirableproperties for a suite of provinces, as defined by Platt and Sathyendranath (1999), are as follows: (1) they should be few in number; (2) the boundary of any province should be elastic, to account for seasonal and interannual variations in the physical forcing; (3) the boundaries should beaccessible to remote-sensing; (4) within seasons the province should have predictable verticalstructure and biogeochemical rate parameters; and (5) they should provide templates forprovince-specific rules. Although it is widely recognised that robust methods for theclassification of such provinces are required for global estimates of oceanic primary production, this work is still in its infancy. The wide range of biogeochemical conditions encountered in the Arabian Sea, however, have made it an ideal setting for the development of such protocols andsignificant advances in this area have been made within the JGOFS programme.
As part of the Canadian contribution to JGOFS, Longhurst et al. (1995) and Longhurst (1998)have made the first formal partition of the entire World Ocean into 57 biogeochemicalprovinces, and used this partitioning for the computation of global primary production.Longhurst et al. (1995) classified provinces according to available knowledge of the regionaloceanographic characteristics of the ocean basins. Combinations of the following factors weretaken into account: latitude, depth of water, proximity of coastline, seasonal irradiance cycle,local wind regime, local precipitation regime, distant forcing of pycnocline depth, and nutrientsupply (described in Longhurst, 1998). Longhurst et al. (1995) identified two provinces for thenorthwestern Indian Ocean (as compared with six by Watts et al., 1999; and four by Brock et al.,1998; see below). It should be noted that the significant step forward made by Longhurst et al.(1995) had a disadvantage in the sense that, in their implementation, the boundaries of theprovinces were static.
Following on from the Longhurst et al. (1995) study and also as part of the Canadiancontribution to JGOFS, Brock et al. (1998) have classified the northwestern Indian Ocean intoprovinces. Their approach is different from that of Longhurst et al. (1995) in that they use acombination of climatologies of mixed layer depths and archived ocean colour (CZCS) data topartition the Arabian Sea into four distinct provinces, based on the modelled attenuation of light through the photic zone and the subsequent relation of the depth of the photic zone to the depthof the mixed layer. This approach is referred to as biohydro-optical classification.
As part of the UK and Canadian contribution to JGOFS, Watts et al. (1999) have used datacollected during the ARABESQUE cruises to partition the northwestern Indian Ocean into asuite of six distinct biogeochemical provinces (Figure 1), characterised by a unique combination of bathymetry, sea-surface temperature (SST) and sea-surface chlorophyll a data. The provinceproperties satisfy at least (1) to (4) of the criteria described by Platt and Sathyendranath (1999). An advantage of the Watts et al. (1999) protocol is that the boundaries of the provinces aredynamic, since their positions are determined from sea-surface properties that can be measuredsynoptically by a satellite. Given that there were no satellites monitoring ocean colour at the time of the ARABESQUE cruises, Watts et al. (1999) tested their protocol with archived oceancolour (CZCS) and SST (AVHRR) data. The protocol was found to be statistically robust,facilitating the computation of near real-time estimates of primary production and newproduction on large spatial scales.

34
Figure 1. A summary of the work carried out by Watts et al. (1999) as part of the UK andCanadian contributions to the JGOFS programme in the Arabian Sea (ARABESQUE cruises,1994).
CZCS, AVHRR, Bathymetry data + Observed sea-surface properties (ARABESQUE 1994)
6 Biogeochemical Provinces (September 1994)
Shipboard data (ARABESQUE 1994) + LocalAlgorithm Model (Platt and Sathyendranath)
Predicted Integrated Primary Production (September 1994)
Empirically derived algorithm
Computed vs. Observed Primary Production:R2 = 0.89 Slope = 0.72 n=10
Computed vs. Observed F-ratio:R2 = 0.81 Slope = 0.55 n=10
Predicted Integrated New Production (September 1994)

35
Recommendations
1. The retrieval of chlorophyll a biomass and fluorometric pigment concentrations
The optical data collected during the ARABESQUE cruises, as described in Pinkerton et al.(1999), clearly indicate that the global remote sensing algorithms presently in use for pigmentretrieval require some regional and seasonal adjustments. These must allow for the bio-opticalchanges observed in near-surface and surface waters brought about by changes in phytoplankton composition in response to changes in oceanographic conditions caused by the monsoon-reversals in the Arabian Sea. It is therefore recommended that new algorithms be developed toallow the retrieval of information on particular taxonomic groups from remote-sensingmeasurements. This will require thorough investigation of the advanced spectral and radiometric properties offered by the new generation of satellites (IOCCG, 1998) in relation to shipboardmeasurements of the light absorption characteristics of different phytoplankton species and their regional and seasonal variability. This can be carried out using the existing databases for theArabian Sea but there is no doubt that building on these databases through future Arabian Seacruises will facilitate this optimisation and increase the accuracy of the remote-sensingalgorithms.
We therefore recommend further ship-time in the Arabian Sea to cover all seasons, maintainingat least the same spatial coverage used in the ARABESQUE and U.S. JGOFS cruises.
2. Primary production estimates by remote sensing of biogeochemical properties
We have available a spectrally resolved model for the computation of primary production (Plattand Sathyendranath, 1988; Sathyendranath et al., 1989; Sathyendranath et al., 1995), given P-Iand vertical biomass information. Additional effort is required as follows.
(i) In the collection of more shipboard measurements of P-I parameters, light absorptionand pigment data over all seasons and on at least the same spatial scale as the U.S. JGOFS andARABESQUE cruises. This can be achieved with more sea-time in the Arabian Sea;
(ii) In the development of protocols for the assignment of the parameters that drive themodel to every pixel on the SeaWiFS image. It seems inevitable that the method applied will be within the context of some scheme of biogeochemical provinces (Platt et al., 1995) so that theprinciple question for the future is about which criteria are used to classify provinces. Is itpossible to develop algorithms that define the boundaries between provinces in real-time at theglobal scale and would these be appropriate for an area such as the Arabian Sea, which exhibitssuch spatial and temporal variability? The discussion on the retrieval of phytoplankton pigmentusing a universal SeaWiFS algorithm (Aiken et al., 1995) would suggest not. Therefore, regional and seasonal algorithms to take account of changing phytoplankton compositions arerecommended. Whilst the approach taken by Watts et al. (1999) for defining provinces appearspromising, the idea of dynamic biogeochemical partitions to facilitate large-scale analyses ofocean-basins is still young and the necessary methodology is still being refined. Such approaches require testing of shipboard data simultaneously with real-time remotely sensed ocean colourdata. Clearly, this is only possible with future sea-time in the Arabian Sea.

36
2.6 Microbial Dynamics2
Peter H Burkill
Plymouth Marine Laboratory, Natural Environment Research CouncilProspect Place, West Hoe, Plymouth, PL1 3DH
UK
Introduction
Considerable recent progress has been made in understanding the role played by microbes in the biogeochemistry of the Arabian Sea. Almost one hundred papers have been published on thistopic in the last decade, mostly stimulated by the JGOFS Process Studies that were planned bySCOR (1995). This chapter focuses on the microbial communities, principally bacteria andprotozoa, in pelagic and benthic biogeochemistry. Recent findings on primary production anddeep benthos are excluded, since these topics are covered elsewhere in this report.
Seasonality
A characteristic of the Arabian Sea is the strong pattern of seasonality caused by monsoonalforcing of the basin. The response of microbial community structure to monsoonal forcing in the Arabian Sea has been studied over an annual cycle (Campbell et al., 1998). Picoplanktonpopulations of heterotrophic bacteria (HBac), Prochlorococcus spp. (Pro), Synechococcus spp.(Syn) and picoeucaryotic algae (Peuc) were shown to vary considerably along two transectsfrom the coast of Oman to 1500 km offshore. HBac were numerically dominant in surfacewaters in all regions (<3 x 106 cells ml-1), with higher maximum abundances in coastal watersthan at offshore stations. Conversely, Pro were most abundant at the oligotrophic offshorestations, and 100-fold lower, or absent, at coastal stations, except during the SpringIntermonsoon when abundances were extremely high along the entire southern transect. Synabundances were highly variable, with no consistent trend between coastal and offshore stations. This variability may be explained by the prevalence of mesoscale eddies, but also could be dueto overlapping distributions of multiple Syn populations distinguished by pigment type. Syn with a low phycourobilin (PUB) to phycoerythrobilin (PEB) ratio pigment type were more abundantat coastal stations, whereas Syn with a high PUB:PEB ratio increased in abundance offshore.Average depth profiles for Pro, Syn, and HBac displayed uniform abundance in the surfacemixed layer, with a rapid decrease below; however, during the Spring Intermonsoon mostprofiles had a peak at the base of the surface mixed layer. Distributions of Peuc typicallydisplayed a subsurface maximum near the base of the surface mixed layer, except during the SW Monsoon when abundance peaked near the surface. This report is the first to describe theseasonal and spatial variation in microbial community structure in the Arabian Sea over acomplete monsoon cycle. Overall, eukaryotes were more important at coastal stations, whileprokaryotes predominant at offshore stations. Pro abundance was restricted to warm oligotrophic waters and was inversely related to surface nitrate concentrations; thus, an increase in the % Pro as a fraction of total prokaryote abundance was also indicative of oligotrophic conditions. Theeffects of SW Monsoonal forcing on microbial community structure resulted in an increase ofPeuc, but this response was limited to coastal stations. Pro and Syn remained dominant atoffshore stations.
2MSS published since 1992 extracted from the ISI database using ‘Arabian Sea’, ‘microbial’, ‘bacteria’ and ‘microzooplankton’

37
The role of such forcing during the NE Monsoon has been studied by Kumar et al. (2001)through time-series observations at a station situated at 21°N, 64°E. At this location, sea-surfacetemperature cooled and the mixed layer deepened due to winter convection. Co-variation ofnitrate concentrations in the surface layers and concentrations of chlorophyll a and primaryproduction in the euphotic zone with mixed-layer depth (MLD) and wind suggests that carbonfixation was controlled primarily by physical forcing. Cooler waters during winter (NEMonsoon) 1997 relative to winter 1995 were associated with deeper MLDs, higher nitrateconcentrations, elevated primary productivity, and higher chlorophyll a concentrations, leadingto the inference that even a 1°C decrease in SST could lead to significantly higher primaryproductivity. Satellite data on sea-surface temperature (advanced very high-resolutionradiometer; AVHRR) and TOPEX/POSEIDON altimeter data suggest that this interannualvariation is of basin-wide spatial scale. After the termination of winter cooling and subsequentwarming during the Spring Intermonsoon, when primary production drops, bacteria andmicrozooplankton proliferate. This suggests that the organisms of the microbial loop may be animportant food source that sustains the mesozooplankton throughout this period (Madhupratap etal., 1996).
The seasonal dynamics of the microbial activity in the euphotic zone has been modelled(Ryabchenko et al., 1998). A nitrogen-based, seven-component ecosystem model has beencoupled to a three-dimensional quasi- geostrophic ocean general circulation model and appliedto a simulation of the seasonal variability of physical and ecosystem variables in thenorthwestern Indian Ocean. Comparison with available data has highlighted many similaritiesand some disagreements between the model and observations. The Ryabchenko model produces an entrainment bloom in May-June in the central Arabian Sea for which there is no evidence in the CZCS data and does not reproduce the observed August - September bloom in the northernArabian Sea. In the latter case, analysis showed that grazing control by zooplankton prevents the development of the model bloom. A comparison of our solution with the results obtained byMcCreary et al. (1996) using a 2.5-layer physical-ecosystem model showed that, despitenumerous differences between the models, the period and location for most blooms in thesesolutions coincide closely. The model reproduced the differences in primary productivitybetween the NE and SW Monsoon periods and showed that the model phytoplankton in the main upwelling areas were only seriously nutrient limited during the Spring Intermonsoon period. For most of the year, the model predicts that phytoplankton production is closely coupled tozooplankton grazing with blooms only occurring when there are rapid changes in phytoplankton growth rate due either to the entrainment of nitrate into the mixed layer or decreased lightlimitation (when the mixed layer is shallowing thereby allowing the phytoplankton to escapefrom grazing control). The predicted particle flux from the euphotic zone was of the order of40% of the primary production and lagged the latter quantity by 8-10 days. This temporal lagmeant that the correlation between daily values of the two quantities was low, and this maypartly explain the difficulties of finding good correlations between observations of these twoquantities.
Bacterial Biogeochemistry
Studies of bacteria in the Arabian Sea (Azam et al., 1994), suggest that temporal contiguity ofhighly oligotrophic and eutrophic periods, along with high water temperatures, may result inunique features of bacteria-organic matter coupling, nutrient cycling, and sedimentation, whichare unlike those in classical oligotrophic and eutrophic waters. Bacteria- phytoplanktoninteractions may influence phytoplankton aggregation and its timing. Azam et al. (1994)hypothesise that, within aggregates, hydrolytic ectoenzyme activity, together with condensationreactions between the hydrolysis products, produce molecular species that are not readilydegraded by pelagic bacteria. Accumulation of a reservoir of such slow-to-degrade dissolved

38
organic carbon (DOC) is proposed to be a carbon flux and energy buffer, which moderates theresponse of bacteria to the dramatic variations in primary production in the Arabian Sea. Use of the slow-to-degrade DOC pool during the Intermonsoons could temporarily render the ArabianSea net-heterotrophic and a source of CO2 to the atmosphere.
DOC is an important resource for bacteria. While DOC is generally considered to beconservative in marine waters, Kumar et al. (1990) have shown DOC to be non-conservative in the basin. Concentrations of DOC were shown to be 3-4 times higher than those reported earlier, and decreased northward from the equator. Total carbon dioxide (TCO2) increases in proportion of the oxygen utilized, thereby revealing the dominant biological role in the carbon turnover.The CO2 added through dissolution of biogenic debris is found to decrease southward.Decomposition of organic material contributes at least 64% to the CO2 addition that increasessouthward, the rest being from dissolution of skeletal material. Evidence is provided for theutilization of oxygen and nitrate for DOC oxidative decomposition. Accumulation of DOCwithout its complete oxidation to CO2 could be the main reason for the TCO2 decrease insouthern Arabian Sea. Relationships of DOC with nitrification and denitrification processesshow that the microbial population plays a major role in regulating the DOC contents in theseawater of this region. Consumption/decomposition by denitrifying bacteria and othermicroorganisms responsible for nitrogen cycling in the sea are found to be intimately related tothe DOC dynamics and are responsible for decreased DOC concentrations in the north. DOCaccumulation in the southern Arabian Sea seems to facilitate bacterio-particulate aggregateformation and consequent nitrification, which results in excess nitrate.
Bacteria often proliferate in regions of high organic matter and may become sites for elevatedmicrobial activity. One source of the latter are transport exopolymer particles (TEP). Kumar etal. (1998) have studied the behaviour of TEP in the Arabian Sea and found that this differs from that of the Bay of Bengal. TEP concentrations were lower in the Bay of Bengal due to fasterscavenging from water column because of interaction with mineral particles. TEP concentrations were higher and occurred even in intense suboxic layers in the Arabian Sea. These resultssupport the mineral ballast theory advocated for the Bay of Bengal and suggest that there is asurplus of organic matter over and above that required to meet the higher carbon demand bybacteria in the denitrifying waters of the Arabian Sea. The situation in the Arabian Sea duringthe late SW Monsoon would seem to be different. Bacterioplankton distributions and production, studied by Ducklow (1993) show abundances and production to be high (>1 x 109 cells l-1 and30-92% of primary production, respectively) along the oceanic portions of the transects. Theseelevated levels may indicate a response to the decline of summer phytoplankton bloomsstimulated by monsoonal deepening of the mixed layer. The bacterial production estimates ofthis study, which used very conservative conversion factors, suggest that the typical carbonsources used to support bacterial production (e.g., phytoplankton exudation, particle breakdown) could supply only a fraction of the bacterial demand. A more recent study on the relationshipbetween TEP and bacteria has been carried out by Ramaiah et al. (2000) who found highregional variability in the vertical distribution of TOC with high concentrations in surface waters at 18-20°N along 64°E due to upwelling-induced productivity. TEP concentrations decreasedwith depth and were lower between 200 and 500 m. Bacterial concentrations were up to 1.99 x108 l-1 in the surface waters and decreased by an order of magnitude or more at depths below 500 m. A better relationship has been found between bacterial abundance and concentrations of TEP than between bacteria and TOC, suggesting that bacterial metabolism is fuelled by TEPavailability. Assuming a carbon assimilation of 33%, bacterial carbon demand (BCD) wasestimated to be 1- 4 g C m-2 d-1 in the surface waters. The observed TEP concentrations appearto be sufficient in meeting the surface and subsurface BCD in the northern Arabian Sea.

39
Bacteria use extracellular enzymes to degrade particulate material. Activity of enzymes such asphosphatase (P-ase) was determined together with bacterial abundance and production ratesduring two SW Monsoons (Hoppe and Ullrich 1999). Samples were collected along the cruisetracks from the equator to the upwelling region at the shelf edge off Oman. Depth profiles of P-ase activity were found to be strikingly different from those of the other enzymes. While valuesof aminopeptidase and beta-glucosidase generally decreased below the euphotic zone, P-aseincreased by factors of 1 to 7. The ratio of peptidase- to P-ase activity was from 4 to 21 at thesurface and from 3 to 5 at 800 m depth. Because P-ase production (dissolved and cell-bound) indeep waters is mainly dependent on bacteria, P-ase activities per bacterial cell were calculated:these were, on average, 37 times higher at 800 m than at the surface. A positive correlation of P-ase activity with phosphate concentrations was observed in depth profiles below the euphoticzone, but this relationship was much more variable in the mixed surface layer. Theseobservations suggest that C-limited bacteria in the deep strata did not primarily focus on thephosphate generated by their P-ase activity but on the organic carbon compounds which weresimultaneously produced and which could probably not be taken up prior to the hydrolyticdetachment of phosphate. It is hypothesised that a considerable part of the measured P-aseactivity was dissolved and may be important for the slow, but steady regeneration of phosphateand organic C in mesopelagic waters.
Bacterioplankton activity in surface waters during and after the 1994 SW Monsoon (Pomroy and Joint, 1999) showed that while phytoplankton production was significantly higher during the SW Monsoon, bacterial numbers showed little difference. Bacteria were most abundant in theeuphotic zone with highest numbers (<1.6 x 109 bacterial-1) found in the shelf waters. Mostbacteria were small cocci, which were larger (diameter 0.40 µm) during the SW Monsoon period than in the Spring Intermonsoon (diameter 0.36 µm). Bacterial production, measured bythymidine and leucine incorporation, confirmed highest rates on the shelf, and decreasedoffshore. In the central Arabian Sea, thymidine incorporation rates were similar in the SWMonsoon and Spring Intermonsoon periods, but higher rates of leucine incorporation weremeasured during the SW Monsoon season. Bacterial production was a relatively smallproportion of phytoplankton production in both periods sampled, estimated to be equivalent tobetween 10 and 30% of the daily primary production.
Bacterial abundance and production in the central and eastern Arabian Sea, studied by Ramaiahet al. (1996), have confirmed the results of Pomroy and Joint (1999) from the western side of the basin. They found higher bacterial concentrations (~1 x 109 cells L-1) during September andApril/May compared to the SW Monsoon period of July/August and the winter period ofFebruary/March. Although primary production was low during April/May, bacterial productionwas much higher during this period, unlike July/August. Bacterial concentrations also showed an increase from the northern to the southern Arabian Sea, suggesting the presence of high amounts of dissolved organic carbon in this area. High picoplankton concentrations (<45 x 106 cells L-1)were observed during February, particularly in the northern Arabian Sea. Rapid turnover ofbacteria during the Spring Intermonsoon period of April/May suggests the predominance of a'microbial loop' in the foodweb and a prevailing source of dissolved organic carbon in theoceanic waters.
The response of bacteria to year-round high primary productivity has shown that bacterialproductivity, both in the surface layer and integrated over the euphotic zone, was elevated lessthan 2-fold during the SW Monsoon (Ducklow et al., 2001). This suggests that bacterialpopulations are closer to steady state than phytoplankton that typically increase in concentrationduring this period. Surprisingly, bacterial abundance and production were low during the NEMonsoon, but then increased in March during the Spring Intermonsoon (Ducklow et al., 2001).

40
Bacterial production seems to respond differently to the seasonal pattern in different regions.Wiebinga et al. (1997) found that during the SW Monsoon, the Somali Current showed highestbacterial production (<849 mgC m-2 d-1) in regions with enrichment of the surface waters byupwelling of cold, nutrient-rich, deep water. In contrast, the Gulf of Aden and the Red Sea were most productive during the NE Monsoon (225 mgC m-2 d-1). Depth profiles of the upper 300 m in general showed a subsurface maximum in bacterial abundance and production at 20-70–mdepth. Heterotrophic activity and primary production were closely correlated, indicating thedependence of bacterioplankton on local phytoplankton-derived organic carbon and their abilityto adapt quickly to changes in the environment. The bacterial carbon demand in the upper 300 mof the water column was largely supplied by phytoplankton production in the euphotic zone.Bacterial production was 18 ± 7% (average ± S.D.) of primary production. Assuming anassimilation efficiency of 50% for marine bacteria, they consumed up to half of the carbonproduced by the phytoplankton.
A comparison of the bacterial activity in the Arabian Sea with other ocean basins has been made (Ducklow, 1999). While euphotic zone bacterial stocks in the Arabian Sea were remarkablysimilar, averaging about 1 gC m-2, compared to subarctic and oligotrophic gyres of the northAtlantic and north Pacific oceans and the upwelling zone of the equatorial Pacific Ocean,bacterial production and growth rates varied more widely. This suggests that regulation ofbiomass and production may occur independently.
While most of the focus in the JGOFS studies has been on carbon cycling, it has not beenentirely so. The role of bacteria in iodine cycling has been greatly facilitated by shipboardexperimental studies in the Arabian Sea. Using cultures of the ubiquitous marine bacteriumShewanella putrefaciens, the chemical reduction of iodate in seawater has been shown to bebiologically mediated (Farrenkopf et al., 1997). Shipboard incubations of filtered seawatercollected from the oxygen minimum zone (OMZ) with this bacterium, showed evidence ofiodate reduction. Inorganic chemical reduction of iodate in these samples was ruled out since no free sulphide was measurable and concentrations of ammonia and nitrite were found to be lessthan 5 µM. To confirm that bacterial metabolism was involved, iodate reduction to iodide byShewanella putrefaciens strain MR-4 was studied in artificial seawater using electrochemicalmethods. It is thought that MR-4 is a facultative anaerobe that may switch amongst a suite ofelectron accepters to support metabolism. In all experiments, MR-4 reduced all iodate to iodide. The rate of formation of [I-] in the culture followed pseudo-first order kinetics. This is the firstreport of the marine bacterial reduction of iodate where the concentrations of iodide and iodatewere measured directly. These results may help to explain the depth distribution of iodinespeciation reported in productive waters like the Arabian Sea.
The composition of the bacterial community was studied using denaturing gradient gelelectrophoresis (DGGE) of rRNA genes (Riemann et al., 1999). During the NE Monsoon, thetotal bacterial community’s genomic DNA was analysed by PCR amplification of 16S rRNAgene fragments, followed by DGGE. In total, 20 DGGE bands reflecting unique or varyingphylotypes were excised, cloned, and sequenced. Bacterial groups commonly found in oceanicwaters (e.g., the SAR11 cluster of alpha-Proteobacteria and cyanobacteria) were present, butsurprisingly none of the sequenced amplicons were related to gamma-Proteobacteria or tomembers of the Cytophaga-Flavobacter-Bacteroides phylum. Amplicons related tomagnetotactic bacteria were found for the first time in pelagic oceanic waters. In the mixed layer the bacterial community was dominated by the same set of approximately 15 phylotypes at allstations, but unique phylotypes were found with increasing depth. Except for cyanobacteria,comparison of the bacterial community composition in surface waters from January andDecember 1995 showed only minor differences, despite significant differences in environmental

41
parameters. These data suggest a horizontal homogeneity and some degree of seasonalpredictability of bacterial community composition in the Arabian Sea.
Microbial Role in the Vertical Flux of Particulates
Microbes play key roles in the transformation of material as it fluxes vertically through the water column. Using NMR spectroscopy, Hedges et al. (2001) have shown evidence that there is non-selective preservation of organic matter in sinking marine particles in the Arabian Sea. Almostall the particulate organic matter exported from ocean surface waters into the ocean interior isrecycled. The fraction of this organic matter remaining becomes buried in ocean sediments, thus sequestering carbon and so influencing atmospheric carbon dioxide concentrations. Theprocesses controlling the extensive biodegradation of sinking particles remain unclear, partlybecause of the difficulty in resolving the composition of the residual organic matter at depth with existing chromatographic techniques. The chemical structure of organic carbon in both surfaceplankton and sinking particulate matter has been analysed. Minimal changes occur in bulkorganic composition, despite extensive (>98%) biodegradation, and amino acid-like materialpredominates throughout the water column. The compositional similarity between phytoplankton biomass and the small remnant of organic matter reaching the ocean interior indicates that theformation of unusual biochemicals, either by chemical recombination or microbial biosynthesis,is not the main process controlling the preservation of particulate organic carbon within thewater column. It seems that the organic matter might be protected from degradation by theinorganic matrix of sinking particles.
Bacterial activity has been suggested as an agent in the concentration and vertical transport ofexcess manganese (Nair et al., 1999) as well as the fatty acid composition of settling particles(Reemtsma and Ittekkot 1992).
Microbial Role in Methane Production
The Arabian Sea is a globally significant ocean basin for methanogenesis (Owens et al., 1991). Jayakumar et al. (2001) have described the patterns of methane found in the eastern and centralbasin in April - May and August - September when surface waters showed 110-2521%saturation. The highest values were observed during the SW Monsoon in the inner shelf wherecoastal upwelling combined with freshwater runoff to produce very strong near-surfacestratification. Methane inputs from coastal wetlands through seasonal runoff may have occurred as abnormally high saturations (up to similar to 13,000%) were recorded in the estuarine surface water. In situ production, favoured by very high biological production in conjunction with theprevalence of suboxic conditions in the upwelled water, could be the other major source. Incomparison, sedimentary inputs seemed to be of lesser importance in spite of previously reported occurrence of gas-charged sediments in this region. Methane profiles in the open central Arabian Sea showed two maxima. The more pronounced deeper maximum, occurring at 150-200 mdepth, was similar to the feature seen elsewhere in the oceans, but was probably intensified here due to an acute oxygen deficiency. It showed some correlation with the subsurface particlemaximum characteristic of the denitrifying layer. The dominant mechanism of its formationmight be in situ production within particles rather than advection from the continental shelf asconcluded by previous workers. The less pronounced and previously unreported shallowermaximum, occurring in the well-oxygenated upper 50 m of the water column, was moredynamic probably as a result of variability of the balance between methane production due tobiological activity and its losses through microbial oxidation and air-sea exchange (see alsoChapter 2.3).

42
The importance of the upper ocean in methanogenic activity has been elucidated using isotopes(Faber et al., 1994) at 24°N, 65°E. This investigation has shown that surface waters of theArabian Sea are enriched in methane, confirming that this is a source for the atmosphere. Carbon isotope ratios point to a bacterial origin of methane that is generated in the surface waters.Concentration changes and variations of carbon isotope ratios also suggest that methane seepingfrom sea floor sediments of the Arabian Sea is oxidized by bacterial activity and does not reachthe atmosphere.
Microzooplankton Trophodynamics
Microbial herbivory has been studied extensively in recent years, and now we have a completeseasonal picture. During the SW Monsoon, microzooplankton grazing utilized 81% ofphytoplankton growth at offshore oligotrophic stations and 54% at upwelling coastal stations(Brown et al., 1999). Similar results have been reported by Edwards et al. (1999) who found that microzooplankton herbivory was greater than mesozooplankton herbivory both nearshore andoffshore. While mesozooplankton cropped ~12% of the daily primary production, themicrozooplankton grazed up to 60% of the daily algal production. At this time, themicrozooplankton community was dominated by aloricate ciliates and heterotrophicdinoflagellates. Their concentrations reached 850 cm-3 in nearshore waters, with highestconcentrations beneath the mixed layer while offshore, concentrations peaked in surface waters(Stelfox et al., 1999). Microzooplankton biomass varied between 1 and 12 µgC l-1, andnanoflagellates, dinoflagellates, and ciliates reached maximum biomass at different locations and depths. Heterotrophs comprised 18-27% of the biomass over most of the U.S. JGOFS transect(Garrison et al., 1998).
Trophic losses of bacteria, studied during the Fall Intermonsoon period, were found to be high(4.1% h-1) in the shallow waters of the Gulf of Oman. Lower values were found in the ArabianSea, with higher (1.7% h-1) rates inshore than in the central gyre. Heterotrophic nanoflagellateswere the primary bacterivores; larger microzooplankton accounted for 33% of total bacterivoryin the Gulf of Oman but only 16% in the central Arabian Sea. Time-course experimentsindicated substantial diel changes in bacterivory making extrapolation to daily rates difficult(Weisse, 1999). In the Fall Intermonsoon, microzooplankton concentrations increasedthroughout the basin reaching standing stocks in excess of 300 mgC m-2 (Stelfox et al., 1999).Microzooplankton biomass reached a maximum of 36 µgC l-1 during the Fall Intermonsoon and was higher than that of mesozooplankton (Gauns et al., 1996), suggesting that the microbialcommunities may dominate the plankton in this season. This is in keeping with the finding thatsmaller algae such as cyanobacteria predominate in the flora, and form a major food source formicrozooplankton (Burkill et al., 1993). The stocks of Synechococcus comprise up to 40% of the POC, and up to 71% of the cyanobacteria could be grazed daily. High turnover by grazing wasmirrored by high growth rates suggesting close coupling between growth and loss processes. It is clear that cyanobacteria are a major component of a dynamic but well-balanced microbialfoodweb present during the Fall Intermonsoon period.
During the NE Monsoon, trophic coupling seems to be much less pronounced, despite similarstocks of microzooplankton (2.0 and 1.8 gC m-2 respectively) in both monsoon seasons (Dennett et al., 1999). Algal growth rates average 0.8 d-1 during the NE Monsoon while microzooplankton grazing crops ca. 0.3 d-1 (Caron and Dennett, 1999). Such values complement those of Landry etal. (1998) who found microzooplankton grazing rates of 0.6 d-1 for surface waters. Herbivorywas estimated to account for 38% d-1 of the algal stock and 67% of the primary production(Reckermann and Veldhuis 1997). Experimental removal of the grazers >10µm in size enhanced algal growth and mortality suggesting a trophic cascade within the microbial community.

43
Microbial Nitrogen Cycling
Microbially driven aspects of the nitrogen cycle have been described and modelled. A studyduring the NE Monsoon by McCarthy et al. (1999) revealed that only the southern most stations(10° and 12° N) and one shallow coastal station were as nutrient-depleted as had been expectedfrom the few relevant prior studies in this region. Experiments were conducted to ascertain therelative importance of different nitrogenous nutrients and the sufficiency of local regenerationprocesses in supplying nitrogenous nutrients utilized in primary production. Except for thesouthern oligotrophic stations, the euphotic zone concentrations of nitrate were typically 5- to10-fold greater than those of nitrite and ammonium. There was considerable variation (20- to 40-fold) in nutrient concentration both within and between the two sections on each cruise. Allnitrogenous nutrients were more abundant (2- to 4-fold) later in the NE Monsoon. Strongvertical gradients in euphotic zone ammonium concentration, with higher concentrations atdepth, were common. This was in contrast to the nearly uniform euphotic zone concentrationsfor both nitrate and nitrite. Half-saturation constants for uptake were higher for nitrate (1.7 µMkg-1 than for ammonium (0.47 µM kg-1). Evidence for the suppressing effect of ammonium onnitrate uptake was widespread, although not as severe as has been noted for some other regions.Both the degree of sensitivity of nitrate uptake to ammonium concentration and the half-saturation constant for nitrate uptake were correlated with ambient nitrate concentration. Thecombined effect of high affinity for low concentrations of ammonium and the effect ofammonium concentration on nitrate uptake resulted in similarly low f-ratios, 0.15 and 0.13, forearly and late observations in the NE Monsoon, respectively. Stations with high f-ratios had thelowest euphotic zone ammonium concentrations, and these stations were either very near shoreor far from shore in the most oligotrophic waters. At several stations, particularly early in the NE Monsoon, the utilization rates for nitrite were up to 50% the utilization rates for nitrate. Whenconverted with a Redfield C:N value of 6.7, the total N uptake rates measured in this study were commensurate with measurements of C productivity. While nutrient concentrations at somestations approached levels low enough to limit phytoplankton growth, light was shown to bevery important in regulating N uptake at all stations in this study. Diel periodicity was observed for uptake of all nitrogenous nutrients at all stations. The amplitude of this periodicity waspositively correlated with nutrient concentration. The strongest of these relationships occurredwith nitrate. Ammonium concentration strongly influenced the vertical profiles for nitrate andammonium uptake. Both nitrite and ammonium were regenerated within the euphotic zone atrates comparable to rates of uptake of these nutrients, and thus maintenance of mixed layerconcentrations did not require diffusive or advective fluxes from other sources. Observedturnover times for ammonium were typically less than one day. Rapid turnover and the stronglight regulation of ammonium uptake allowed the development and maintenance of verticalstructure in ammonium concentration within the euphotic zone. In spite of the strong positiveeffect of light on nitrite uptake and its strong negative effect on NO2 production, the combinedeffects of much longer turnover times for this nutrient and mixed layer dynamics resulted innearly uniform nitrite concentrations within the euphotic zone. Responses of the NE Monsoonplanktonic community to light and nutrients, in conjunction with mixed layer dynamics, allowed for efficient recycling of N within the mixed layer. As the NE Monsoon evolved and the mixedlayer deepened convectively, nitrite and nitrate concentrations increased correspondingly withthe entrainment of deeper water. Planktonic N productivity increased 2-fold, but without asignificant change in the new to recycled N proportionality. Consequently, nitrate turnover time increased from about 1 month to greater than 3 months. This reflected the over-ridingimportance of recycling processes in supplying nitrogenous nutrients for primary productionthroughout the duration of the NE Monsoon. As a result, nitrate supplied to the euphotic zoneduring the NE Monsoon is, for the most part, conserved for utilization during the subsequentSpring Intermonsoon season.

44
The role of bacteria in denitrification has been studied by Naqvi et al. (1993) who found anintermediate nepheloid layer (INL) associated with high microbial metabolic rates anddenitrification. A wide range of simultaneous optical, physical, chemical, and biochemicalmeasurements showed that the INL was invariably associated with the secondary nitritemaximum. Maxima in particulate protein and the activity of the respiratory electron transportsystem (ETS) are also found within the INL. Since the INL persisted far from the continentalmargin and intensified offshore, it may not be related to the transport of material resuspendedalong the continental margin. An apparent correlation of the INL with the previously reportedsubsurface maximum in bacterial abundance suggests that a local increase in the abundance ofbacteria may be responsible for the increased turbidity. Positive correlations of the beamattenuation anomaly with nitrite and nitrate deficit suggest that most of these bacteria may bedenitrifiers. The organic carbon demand within the denitrifying layer, computed from theobserved ETS activity, appears to be several fold higher than the sinking carbon fluxes to thedenitrifying layer, requiring additional modes of supply of biodegradable organic matter. It isproposed that a bacterial maximum could be maintained with efficient utilization of thedissolved organic matter within the denitrifying waters (see also Chapter 2.3).
Another aspect of the role played by microbes in the nitrogen cycle concerns methylamineproduction. The distributions and biogeochemistry of methylamines and ammonium in theArabian Sea have been reported by Gibb et al. (1999). They found concentrations of NH4
+ up to two orders of magnitude greater than those of methylamines throughout the region.Monomethylamine was generally the most abundant MA, whilst trimethylamine was found onlyat concentrations < 4 nM. Low concentrations in open ocean meso- and oligo-trophic regionscontrasted with the elevated levels recorded in the highly productive coastal upwelling waters of the NW Arabian Sea. In total, methylamines contributed < 1% dissolved organic nitrogen(DON). Depth maxima of methylamines were generally associated with those of chlorophyll a,and in offshore regions, also with those of ammonium (above the thermo-, oxy- and nitra-clines).Trimethylamine maxima were recorded at the base of the thermo- and oxyclines, resolved fromthe other analytes. Through correlation studies, a degree of diatom specific MMA productionwas inferred and microzooplankton grazing found to influence significantly all aqueous MAconcentrations. Enhanced correlation of monomethylamine concentrations withmesozooplankton abundance was attributed to their ability to graze diatoms. We postulate thatthe nitrogen taken up in nutrient-rich, diatom-dominated regions of the Arabian Sea will be used both biosynthetically and anabolically. This may be accompanied by introduction ofmethylamines into the aqueous phase through enzymatic precursor degradation, nitrogendetoxification, senescence or lysis and be accelerated through grazing pressure, particularly thatof mesozooplankton on diatoms. In contrast, under the more oligotrophic conditions recorded in the remote Arabian Sea, those species of phytoplankton with a lower nitrogen demand arefavoured, e.g., prymnesiophytes and dinoflagellates. Correspondingly, lower methylamineconcentrations are recorded in these regions (see also Chapter 2.3).
Nitrogen cycling in suboxic and anoxic conditions in the Arabian Sea has been modelled(Yakushev and Neretin, 1997). The vertical distributions of nitrogen compounds (total organicnitrogen, ammonium, nitrate, nitrite) as well as dissolved oxygen agree reasonably well with the observations. Bacterially mediated processes are described by first-order equations using O2
concentration. Denitrification is estimated to be 34 TgN y-1 in the Arabian Sea.
Oxygen Minimum Zone
A key oceanographic characteristic of the Arabian Sea is the presence of low oxygenconcentration in mid water. This oxygen minimum zone (OMZ) is spatially variable, showinggreatest oxygen deficiency in the northern basin. Its vertical expression is also variable showing

45
greatest deficiency over the depth horizon of ca. 200 to 2000 metres, although exact horizon ofthe OMZ depends on the location and season. A summary of the seasonal interaction of themonsoon with the OMZ has been reported by Morrison et al. (1999) for 1995. They considered that dissolved oxygen, nitrite, nitrate and density values are all required to delineate the OMZ,and the regions of denitrification. The suboxic conditions within the northern Arabian Sea aredocumented, as well as biological and chemical consequences of this phenomenon. Overall, theconditions found in the suboxic portion of the water column in the Arabian Sea were not greatlydifferent from what has been reported in the literature with respect to oxygen, nitrate and nitritedistributions. Within the main thermocline, portions of the OMZ were found that were suboxic(oxygen less than similar to 4.5 µM) and contained secondary nitrite maxima withconcentrations that sometimes exceeded 6.0 µM, suggesting active nitrate reduction anddenitrification. Although there may have been a reduction in the degree of suboxia during theSW Monsoon, strong seasonality was not observed, as has been suggested by some previouswork. There was little evidence for the occurrence of secondary nitrite maxima in waters withoxygen concentrations greater than 4.5 µM. The northern basin appeared to accumulate largernitrate deficits due to longer residence times even though the denitrification rates, shown byreduced nitrite concentrations, there might be lower. Organism distributions showed strongrelationships to the oxygen profiles, especially in locations where the OMZ was pronounced, but the biological responses to the OMZ varied with type of organism. The regional extent ofintermediate nepheloid layers in our data corresponds well with the region of the secondarynitrite maximum. This is a region of denitrification, and the presence and activities of bacteriaare assumed to cause the increase in particles.
The importance of the OMZ for denitrification has long been known. However, measurements of respiration associated with denitrification have been hampered by the difficulty of makingaccurate measurements of O2 in the OMZ. One way around this problem is to use cellularmeasurements of the electron-transport system (ETS) to quantify respiration in the OMZ (Naqvi and Shailaja, 1993). Measurements of ETS activity at 15 stations in the Arabian Sea during theNE Monsoon yielded high respiration rates that do not correlate with the trends in primaryproductivity. The rates are unlikely to be sustained by the supply of carbon associated with thesinking particles alone, and seem to suggest a major role for dissolved and/or suspended organic matter in fuelling oxygen consumption and denitrification. Naqvi and Shailaja (1993) havecomputed a denitrification rate of 24 - 33 TgN y-1 in the Arabian Sea. This estimate agrees with the estimate based on the exports of nitrate deficits outside the denitrification zone. Theventilation time of the denitrifying layer is calculated as approximately 1 year.
The effect of the OMZ on the benthos has also been studied (Schmaljohann et al., 2001) in thenortheastern Arabian Sea off Pakistan. Here the upper continental slope between 350 and 850 m water depth, which is in the centre of the oxygen-minimum zone, is characterized by numeroussites of small-scale seeps of methane- and sulphide-charged porewater. White bacterial matswith diameters <1 m of filamentous sulphur-oxidizing bacteria were discovered. Seep sediments, as well as non-seep sediments, in the vicinity were characterized by the occurrence of thebacterium Thioploca in near-surface layers between 0 and 13 cm depth. Thioploca bundles were up to 20 mm in length and contained up to 20 filaments of varying diameters, between 3 and 75 µm. Up to 169 bundles cm-2 were counted. Maximum numbers occurred in the top 9 cm ofsediment, which contained very low concentrations of soluble sulphide (<0.2 µM) and highamounts of elemental sulphur (up to 10 µmol cm-3). Moderate sulphate reduction activity(between 20 and 190 nM cm-3 d-1) was detected in the top 10 cm of these sediments, resulting in a gradual downcore decrease of sulphate concentrations. CO2 fixation rates had distinct maximaat the sediment surface and declined to background values below 5-cm depth.

46
Preservation and composition of organic matter in surface sediments were investigated on threedifferent transects on the Pakistan continental margin across the OMZ (Schulte et al., 2000). To assess whether the OMZ has an effect on the total organic carbon (TOC), the extractable lipidswere quantified by gas chromatography, combined gas chromatography/mass spectrometry, andcompound-specific stable carbon isotope measurements. These lipids were dominated by marine organic matter as indicated by the abundance of lipids typical of marine biota and by the bulkand molecular isotopic composition. Sediments from within the OMZ are enriched in organiccarbon and in several extractable lipids such as phytol, n-alcohols, total sterols, n-C-35 alkanes, relative to stations above and below this zone. Other lipid concentrations, such as those of totaln-fatty acids and total n-alkanes fail to show any relation to the OMZ. Only a weak correlationof TOC with mineral surface area was found in sediments deposited within the OMZ. Incontrast, sediments from outside the OMZ do not show any relationship between TOC andsurface area. Among the extractable lipids, only the n-alkane concentration is highly correlatedwith surface area in sediments from the Hab and Makran areas. In sediments from outside theOMZ, the phytol and sterol concentrations are also weakly correlated with mineral surface area. The depositional environment of the Indus Fan offers the best conditions for an enhancedpreservation of organic matter. The OMZ, together with the undisturbed sedimentation atmoderate rates, seems to be mainly responsible for the high TOC values in this area. Overall, the type of organic matter and its lability toward oxic degradation, the mineral surface area, themineral composition, and possibly the secondary productivity by sedimentary bacteria alsoappear to have an influence on organic matter accumulation and composition.
The biogenic origins of organic matter in sediments underlying the OMZ have been investigated using lipids (Smallwood and Wolff, 2000) and amino acids (Suthhof et al., 2000). The highabundance of compounds that can be ascribed to primary producers, namely diatoms,dinoflagellates, coccolithophores, Eustigmatophycae and cyanobacteria, reflects the highproductivity of surface waters, but there is little direct molecular evidence of zooplanktonreworking of organic material, except at the deepest water site (similar to 1250 m). There is,however, clear evidence of benthic re-working, probably by detritivorous megafauna, at one ofthe sites near the base of the oxygen minimum zone (similar to 1000 m). The relative abundance of biological markers that can be ascribed to aerobic and anaerobic bacteria in the sediments,namely the hopanoids and branched fatty acids, respectively, mirrors the concentrations ofoxygen in overlying water. Considerable intra- and inter-site variability probably results fromthe spatial and temporal variability in production and from the complexity of sedimentationprocesses. There is little clear evidence that bottom water oxygenation affects quantitativepreservation of organic matter.
Benthic Microbial Biogeochemistry
Denitrification is the process by which nitrate is reduced to gaseous nitrogen species, typicallyN2 or N2O, and is the dominant mechanism for removal of fixed nitrogen from the biosphere. In the oceans, denitrification is mediated by bacteria in suboxic environments and, by controllingthe supply of fixed nitrogen, is an important limiting factor for marine productivity.Denitrification produces substantial 15N enrichment in subsurface nitrate, which is reflected inthe isotopic composition of sinking particulate nitrogen; sediment 15N/14N ratios in regions with suboxic water columns may therefore provide a record of past changes in denitrificationintensity. Altabet et al. (1995) report nitrogen isotope data for sediment cores from three sites in the Arabian Sea. At all three sites, large, near-synchronous downcore variations in 15N/14N were found, which are best explained by regional changes in the isotopic composition of subsurfacenitrate, and hence denitrification. Moreover, these variations are synchronous with Milankovitch cycles, thereby establishing a link with climate. These large, climate-linked variations, in a

47
region that contributes significantly to global marine denitrification, are likely to have perturbed marine biogeochemical cycles during the Late Quaternary period.
Benthic bacterial and nanoflagellate concentrations in the sediments and benthic boundary layer have been studied during upwelling and non-upwelling seasons, in the Arabian Sea (Bak andNieuwland, 1997). There was a significant decrease with depth in benthic bacteria and benthicnanoflagellates. Bacterial concentration and biomass were significantly larger in the bottomwater during the upwelling season, whereas benthic flagellates were significantly morenumerous in the sediments in the non-upwelling season.
The distribution of microbial biomass in sediments at eight sites in the Arabian Sea has beenstudied using phospholipids (Boetius and Lochte, 2000). Surface sediment phospholipidconcentrations at 4 abyssal sites ranged from 7 nM cm-3 at the southern site (SAST; 10 degreesN 65 degrees E, 4425 m) to 29 nM cm-3 at the western site (WAST; 16 degrees N 60 degrees E, 4045 m). The high values detected at the abyssal station WAST exceeded those in the literaturefor other abyssal sites and were comparable to values from the upper continental slope of the NE Atlantic and Arctic oceans. At the four continental slope sites in the Arabian Sea, phospholipidconcentrations ranged from 9 to 53 nmol cm-3 with the maximum values at stations A (2314 m)and D (3142 m) close to the Omani coast. Records of particulate organic carbon flux to the deep sea are available for four of the investigated locations, allowing a test of the hypothesis that the standing stock of benthic microorganisms in the deep sea is controlled by substrate availability,i.e. particle sedimentation. Total microbial biomass in the surface sediments of the Arabian Sea was positively correlated with sedimentation rates, consistent with previous studies of otheroceans.
The microbiology of the benthic nepheloid layer (BNL) has been studied (Boetius et al., 2000a) at six stations in the deep Arabian Sea during May and February. Suspended particulates andbacterial activity were found to be more variable vertically than regionally or temporally.Compared to the deep-water column, bacterial concentrations increased towards the seafloorwith highest concentrations of suspended particulates, bacterial leucine incorporation and theactivity of different enzymes found in the near-bottom water. Suspended particulates in the BNL had a high chlorophyll a-to-POC ratio and were of higher organic carbon content than thesinking particles or the particulate matter at the sediment surface. The carbon demand of thebacterial community in the BNL, estimated by leucine incorporation experiments, exceeded thevertical POC flux. Thus, it seems that the enhanced microbial activity and biomass close to theseafloor is mainly supported by the resuspension of small phytodetrital particles and by the DOC flux.
Substantial regional variation in benthic microbial biomass and activity has been found in theArabian Sea with highest values in the western Arabian Sea (station WAST), decreasingapproximately three-fold to the south (station SAST). Benthic microbial biomass and activityduring the NE Monsoon was as high or higher than subsequent to the SW Monsoon, indicating a very rapid turnover of POC in the surface sediments. This variation in the biomass and activityof the microbial assemblages in the deep Arabian Sea can largely be explained by the regionaland temporal variation in POC flux. Compared to other abyssal regions, the substantially higher benthic microbial biomasses and activities in the Arabian Sea reflect the extremely highproductivity of this tropical basin (Boetius et al., 2000b).
Sulphate reduction in near-surface sediments of the deep Arabian Sea has been studied usingstable isotopes (Bottcher et al., 2000). Depth profiles of the stable sulphur isotopic compositionof dissolved sulphate in near-surface sediments at five stations of the deep Arabian Sea indicate that net microbial sulphate reduction can take place in the sediments. At stations WAST and

48
NAST this occurred at depths below about 12 cm. At WAST this was more pronounced than at station NAST, most likely due to a higher organic carbon content in turbiditic sediments.Sulphate reduction did not take place within the upper 10 cm of the surface sediments at anystations, and no significant isotopic indication for sulphate reduction was found down to 30 cmbelow the surface (bsf) at station SAST. Results are in accordance with accumulation of reduced, isotopically light sulphur species below about 6 cm bsf at station WAST. Bottcher et al. (2000) concluded that the sulphur isotopic composition of remaining sulphate is more sensitive to netsulphate reduction than the [SO4]/[Cl] ratio. The sulphur isotopic composition of a verticalprofile for dissolved sulphate through the water column at station WAST was essentiallyconstant (250 - 4047 m: δ34S = +20.49 ± 0.008 parts per thousand, versus V-CDT, n = 8). Asimilar constancy (20 - 4565 m: δ34S = +20.57 ± 0.06 parts per thousand versus V-CDT, n = 15) was found for the station BIOTRANS in the northeastern Atlantic Ocean (47°N 19°W),indicating that the oxygen minimum zone in the Arabian Sea has no influence on the sulphurisotopic composition of dissolved sulphate.
A possible microbial origin of phosphorites in the Arabian Sea has been proposed (Rao et al.,1992). Petrographical, mineralogical and microprobe analyses of phosphorites from ErrorSeamount, in the northwestern Arabian Sea, demonstrate that microbial processes played animportant role in the early diagenetic formation of phosphorites during subaerial exposure of the seamount. The phosphorites on Error Seamount occur as laminated crusts and massive slabs.Low-magnesium calcite and carbonate fluorapatite are the major minerals of the phosphorites,with goethite being important only in the massive slab phosphorites. Elemental sulphur,pyrrhotite, gypsum and chlorite are the mineral phases present in the acid-insoluble residues ofthe phosphorites. Elemental sulphur, which is exclusively formed by microbial processes, occurs as submicron-sized granules on gypsum surfaces. The laminated crust phosphorites consist oforganic-rich and organic-poor Laminae interlayered with ghost pellets, index fossils ofOligocene to lower Miocene age, peloids and coated grains. Massive slab phosphorites are fine-grained, and consist of goethitic and non-goethitic zones. Non-goethitic zones comprise mainlymicrosphorite with Oligocene to lower Miocene fossils and microfossils. Goethitic zones containabundant Quaternary skeletons. Microprobe analyses of the Error Seamount phosphoritesindicate that the geochemistry of these phosphorites is similar to that of the Tertiary Seamountphosphorites in the Atlantic and Pacific oceans and that they are enriched in Ca and depleted in P compared to other seamount and island phosphorites in the Indian Ocean. Microbial activity is evident in both types of phosphorites and occurs in the form of microborings in peloids and onskeletal fragments, encrustation of phosphate crystals on hollow globular microstructures andphosphate sheaths enclosing organic filaments. The presence of calcite rim cements, microspar-filled patches and recrystallised grains indicates exposure of the seamount under subaerialconditions, during which phosphatisation took place.
Microbial Respiration
Respiration rates in the Arabian Sea have been reported for both the monsoon seasons (Dickson et al., 2001). They have investigated the relationships among oxygen, carbon and nitrogenproduction and respiration rate measurements made in the Arabian Sea. Increased biologicalproduction characterized the SW Monsoon, with rates 12-53% higher than the NE Monsoon. Inmost cases, there was remarkable similarity in production rates during the two monsoons and an absence of strong spatial gradients in production between nearshore and offshore waters,especially during the SW Monsoon. Daily 14C and total 15N production underestimated gross Cproduction, and at the majority of stations 14C and total 15N production were either the same asnet C production or between gross and net C production. Moreover, new production (15N-nitrate), scaled to carbon, was substantially less than net C production. Approximately 50% ofthe POC-14C was metabolised during the photoperiod, with smaller losses (7-11%) overnight.

49
The simplest explanation for the discrepancy between gross and total 15N production andbetween net C and new production was the loss of 15N-labeled particulate matter as dissolvedorganic matter. Partitioning of metabolised gross C production into respiratory and dissolvedpools showed distinct onshore-offshore distributions that appeared to be related to thecomposition of the phytoplankton assemblage and probably reflected the trophodynamics of theecosystem. The percentage of gross C production released as dissolved organic carbon (DOC)was highest in the nearshore waters where diatoms dominated the phytoplankton assemblage,while community respiration was a more important fate for production further offshore wherepicoplankton prevailed. In general, stations that retained more gross C production as netproduction (i.e., high net C/gross C ratios) had higher rates of DOC production relative tocommunity respiration. Locations where community respiration exceeded DOC production were characterized by low rates of net C production and had low net C/gross C ratios. In thoseecosystems, less net C production was retained because higher metabolic losses reduced gross C production to a greater extent than at the more productive sites.
The Electron Transport System (ETS) is a biochemical proxy for respiration and has been usedby Naqvi et al. (1996) to compare respiration in the Arabian Sea and the Bay of Bengal. Theyfound much lower ETS activities in subsurface waters of the Bay of Bengal than those measured in the Arabian Sea. Lower respiration rates in the Bay of Bengal are corroborated by the muchweaker north-south gradients in oxygen and total carbon dioxide. These are, however, in conflict with the higher sinking fluxes of organic carbon measured with sediment traps. The observations support the view that particulate organic matter may undergo a lesser degree of oxidation in thewater column through its incorporation into rapidly sinking matter, perhaps because of themassive inputs of terrigenous matter in the Bay of Bengal. Differential respiration rates maycause changes in the distribution of suspended particulate matter and may also explain why theBay of Bengal is not a site of water-column denitrification in spite of an apparently slowerrenewal of the intermediate waters as compared to the Arabian Sea.
Synthesis Studies
Few attempts have been made at synthesising the recently advanced understanding of microbialbiogeochemistry of the Arabian Sea. One of these is the synthesis of microbial dynamics fromthe U.S. study made by Garrison et al. (2000). In this, the biomass in the upper 100 metres from a number of sources are combined, the structure of the food web evaluated, and changes in food web structure are related to carbon cycling processes. Microbial biomass ranged fromapproximately 1.5 to > 5.2 gC m-2, with heterotrophic bacteria contributing 16 and 44%;autotrophs contributing 43-64%, with the remainder made up of nano- and microheterotrophs.Autotrophs and nano- and microheterotrophs showed a general pattern of higher values atcoastal stations, with the lowest values offshore. Heterotrophic bacteria (Hbac) showed nosignificant spatial variations. The Spring Intermonsoon and early NE Monsoon were dominatedby autotrophic picoplankton, Prochlorococcus and Synechococcus. The late NE Monsoon andlate SW Monsoon periods showed an increase in the larger size fractions of the primaryproducers. At several stations during the SW Monsoon, autotrophic microplankton, primarilydiatoms and Phaeocystis colonies, predominated. Increases in the size of autotrophs were alsoreflected in increasing sizes of nano- and microheterotrophs. The biomass estimates based oncytometry and microscopy are consistent with measurement of pigments, POC and PON.Changes in community structure were assessed using the percent similarity index (PSI) inconjunction with multidimensional scaling (MDS) or single-linkage clustering analysis to showhow assemblages differed among cruises and stations. Station clustering reflected environmental heterogeneity, and many of the conspicuous changes could be associated with changes intemperature, salinity and nutrient concentrations. Despite inherent problems in combining datafrom a variety of sources, the present community biomass estimates were well constrained by

50
bulk measurements such as chlorophyll a, POC and PON, and by comparisons with otherquantitative and qualitative studies. The most striking correlation between food web structureand carbon cycling was the dominance of large phytoplankton, primarily diatoms, and theseasonal maxima of mass flux during the SW Monsoon. High nutrient conditions associated with upwelling during the SW Monsoon would explain the predominance of diatoms during thisseason. The sinking of large, ungrazed diatom cells is one possible explanation for the fluxobservations, but may not be consistent with the observation of concurrent increases in largermicrozooplankton consumers (heterotrophic dinoflagellates and ciliates) and mesozooplanktonduring this season. Food-web structure during the early NE Monsoon and Intermonsoonssuggests carbon cycling by the microbial community predominated in those seasons.
Recommendations
1. Almost one hundred microbial papers have been published in this field since 1992. Half ofthese were in the last 3 years. The report shows how rich the literature on the Arabian Seahas become, and how recent much of this is.
2. There have been few attempts at synthesis: a full synthesis of microbial dynamics is nowwarranted.

51
2.7 Zooplankton
Sharon L. Smith
The Rosenstiel School, University of Miami4600 Rickenbacker Causeway
Miami, FL 33149USA
Martien Baars
Netherlands Institute for Sea ResearchP.O. Box 59, 1790 AB Den Burg, Texel
The Netherlands
Introduction
The Arabian Sea is unique in the world for its upwelling areas that seem to support large pelagic fisheries (Bakun et al., 1998). Until recently, zooplankton of the Arabian Sea region were knownprimarily from three oceanographic expeditions in this century: The John Murray Expedition(JME; September 1933 - May 1934), the International Indian Ocean Expedition (IIOE; 1959-1965), and the Indian Ocean Expedition (INDEX; 1979). In the 1990's, investigations by theNetherlands (Netherlands Indian Ocean Programme; NIOP; 1992-1993), the United Kingdom(Arabesque; 1994), Germany (German JGOFS; 1995, 1997), Pakistan (North Arabian SeaEnvironmental and Ecological Research; NASEER, 1992-1994), India (Indian JGOFS, 1995-1997) and the United States (U.S. JGOFS, 1994-1996) expanded our knowledge of the region.
The Arabian Sea’s Paradox of the Plankton
Close scrutiny of zooplankton biomass data for the northwestern Indian Ocean, including theArabian Sea, has shown that seasonal oscillations thought to arise from seasonal variability inphytoplankton abundance actually do not occur (Baars and Brummer, 1995; Madhupratap et al.,1996; Baars and Oosterhuis, 1998; Baars, 1999). Prior to the investigations of JGOFS, there was no explanation of the IIOE result that phytoplankton biomass varied widely while zooplanktonbiomass remained reasonably constant throughout the year. With the thorough seasonalobservations of JGOFS, it is now apparent that large fluctuations in primary productivity andsurface chlorophyll a do not occur (Barber et al., 2001). Convection during the NE Monsoonresults in substantial primary production that was not apparent in the IIOE atlases. Therefore,there is no seasonal food limitation of zooplankton productivity in the Arabian Sea. There are,however, seasonal variations in the zooplankton community, most notably the ontogeneticallymigrating Calanoides carinatus that is at the surface in upwelling areas during the SW Monsoon season (Smith, 1995; Smith et al., 1998). Detailed seasonal comparisons of other taxa remain to be done for the JGOFS study area. Prior seasonal investigation of the Somali segment of the NW Indian Ocean showed shifts in abundances of other taxa in both the NE and SW Monsoonseasons (Smith, 1988).
The Oxygen Minimum Zone (OMZ)
The sharp subsurface gradient between oxic and suboxic waters, which generally occurs between 100 and 150 metres in the JGOFS study area, is an impenetrable boundary for many zooplankton (Morrison et al., 1999). At the station with the most intense oxygen deficiency in 1995, 85% of the zooplankton biomass in the upper 300 meters was in the upper 100 meters day and night; diel vertical migration was suppressed in the area of strong and persistent oxygen depletion (Smith et

52
al., 1998). Earlier studies have shown that the copepod Pleuromamma indica has its highestabundances at low oxygen and that of seven species of this genus in the Indian Ocean only P.indica has the ability to survive in low oxygen waters (Saraswathy and Iyer, 1986; Goswami etal., 1992). Although Pleuromamma indica migrates daily to the surface layer (Saraswathy andIyer, 1986), it must have physiological adaptations to withstand long exposure to suboxicconditions. These adaptations have not been investigated. Between 400 and 500 meters, theabundances of many taxa fall nearly to zero (Morrison et al., 1999). Earlier studies whichcompared zooplankton biomass in areas of the Arabian Sea containing an OMZ with those thatdid not have shown that even in the oxic upper 100 meters biomass in an area with an OMZ was half that in areas without an OMZ (Vinogradov and Voronina, 1962). At the lower boundary of the OMZ (600-1000 metres), biomass in the area of the OMZ is not different from biomass in an area that does not contain an OMZ. The copepod Lucicutia grandis is an indicator of this habitat, and in the Arabian Sea it was found feeding and reproducing at depth year-round (Wishner et al.,2000). The material in its guts revealed feeding on a wide range of particles (Gowing andWishner, 1998; Wishner et al., 2000).
Seasonal Differences in Biomass and Species Composition
Surveys of the west coast of India, extending up to 700 kilometres offshore, have shown clearlythat both zooplankton biomass (330-µm mesh) and euphausiid biomass are elevated during theSW Monsoon season (Mathew et al., 1990). The maximum epipelagic biomass in the upper 150 meters of the eastern Arabian Sea (287 mmolC m-2; Mathew et al., 1990) is similar to thatobserved in the upper 200 meters off Somalia in the SW Monsoon season (363 mmolC m-2;Baars and Oosterhuis, 1997). The highest biomass of epipelagic zooplankton measured in theentire Indian Ocean during the IIOE was 444 mmolC m-2 (0-200 meters), and this was located in the coastal upwelling area off Oman (Qasim, 1977). The onshore-offshore gradient inzooplankton biomass off Oman was strong, from 444 mmolC m-2 nearshore (20°N, 58°E) to 110 mmolC m-2 offshore at 10°N, 65°E (Qasim, 1977; his displacement volumes converted to carbon using Wiebe, 1988, for dry weight and Ikeda, 1974, for carbon).
Although the zooplankton biomass of the Arabian Sea is largely comprised of copepods, basin-scale distribution and abundance of individual species are generally poorly known. DuringApril-October (SW Monsoon), peak copepod abundances occur in the upwelling regions offSomalia and Arabia, but high abundances also occur far offshore of the Arabian Peninsula.During October-April (NE Monsoon), copepods are abundant off the Arabian Peninsula and inthe Gulf of Oman, but are somewhat less abundant (compared to the SW Monsoon) off Somalia. Unfortunately, the IIOE plankton collections have not been exploited extensively at the specieslevel; consequently, the reported results from the IIOE plankton investigations leave us ill-prepared to specify with any confidence which copepod species are dominant regionally orseasonally (GLOBEC, 1993).
In the northwestern Indian Ocean, large calanoid copepods of the family Eucalanidae dominate with mean abundances of 3000 and 2250 per standard haul (0-200 m) in the SW and NEMonsoons, respectively (Stephen et al., 1992). The genera Euchaeta (1500/1900 for SW/NEMonsoons) and Pleuromamma (270/1100 for SW/NE Monsoons) are also abundant. In the upperlayer of the Arabian Sea, at least 231 species of copepod have been identified (Lane et al.,1998).
One of the assets of the U.S. JGOFS investigation of the Arabian Sea was sampling of the entire annual cycle of zooplankton. Highest biomass in the upper 300 meters was measured within 350 kilometres of the coast during the SW Monsoon (Smith et al., 1998), and highest biomass in the 0-1000–m layer also was measured during the SW Monsoon (Wishner et al., 1998). In the whole
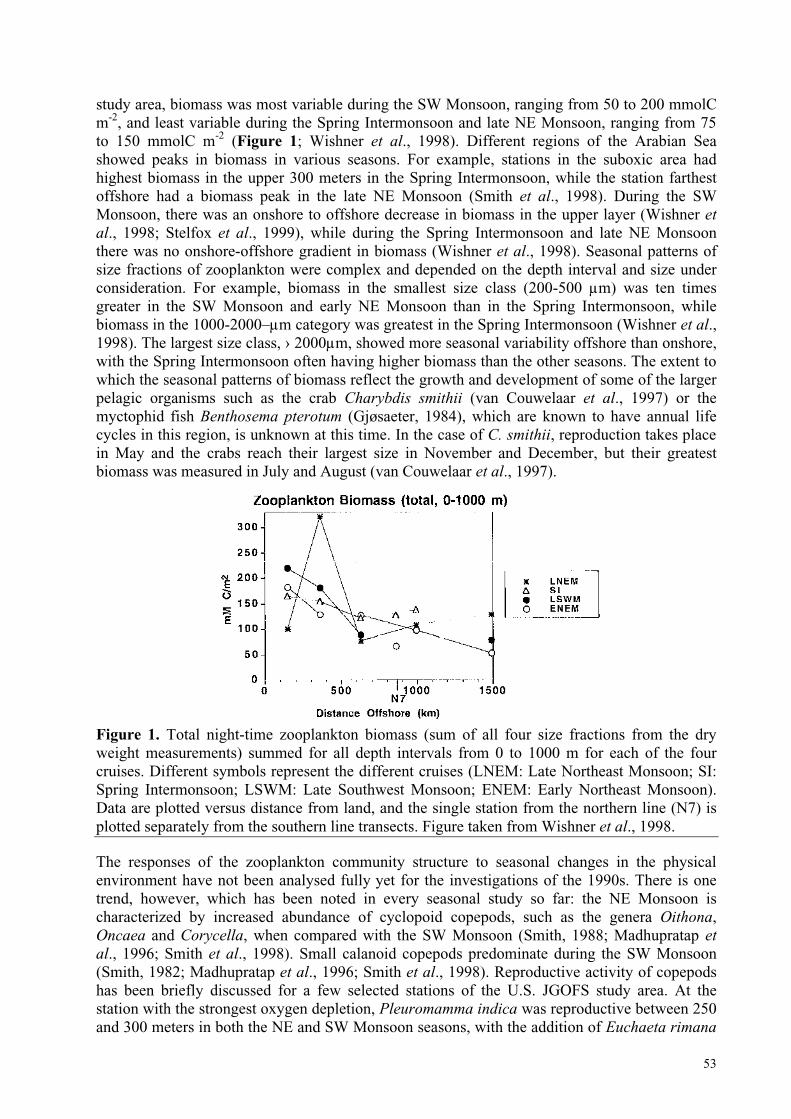
53
study area, biomass was most variable during the SW Monsoon, ranging from 50 to 200 mmolCm-2, and least variable during the Spring Intermonsoon and late NE Monsoon, ranging from 75to 150 mmolC m-2 (Figure 1; Wishner et al., 1998). Different regions of the Arabian Seashowed peaks in biomass in various seasons. For example, stations in the suboxic area hadhighest biomass in the upper 300 meters in the Spring Intermonsoon, while the station farthestoffshore had a biomass peak in the late NE Monsoon (Smith et al., 1998). During the SWMonsoon, there was an onshore to offshore decrease in biomass in the upper layer (Wishner etal., 1998; Stelfox et al., 1999), while during the Spring Intermonsoon and late NE Monsoonthere was no onshore-offshore gradient in biomass (Wishner et al., 1998). Seasonal patterns ofsize fractions of zooplankton were complex and depended on the depth interval and size underconsideration. For example, biomass in the smallest size class (200-500 µm) was ten timesgreater in the SW Monsoon and early NE Monsoon than in the Spring Intermonsoon, whilebiomass in the 1000-2000–µm category was greatest in the Spring Intermonsoon (Wishner et al.,1998). The largest size class, › 2000µm, showed more seasonal variability offshore than onshore, with the Spring Intermonsoon often having higher biomass than the other seasons. The extent to which the seasonal patterns of biomass reflect the growth and development of some of the larger pelagic organisms such as the crab Charybdis smithii (van Couwelaar et al., 1997) or themyctophid fish Benthosema pterotum (Gjøsaeter, 1984), which are known to have annual lifecycles in this region, is unknown at this time. In the case of C. smithii, reproduction takes place in May and the crabs reach their largest size in November and December, but their greatestbiomass was measured in July and August (van Couwelaar et al., 1997).
Figure 1. Total night-time zooplankton biomass (sum of all four size fractions from the dryweight measurements) summed for all depth intervals from 0 to 1000 m for each of the fourcruises. Different symbols represent the different cruises (LNEM: Late Northeast Monsoon; SI:Spring Intermonsoon; LSWM: Late Southwest Monsoon; ENEM: Early Northeast Monsoon).Data are plotted versus distance from land, and the single station from the northern line (N7) isplotted separately from the southern line transects. Figure taken from Wishner et al., 1998.
The responses of the zooplankton community structure to seasonal changes in the physicalenvironment have not been analysed fully yet for the investigations of the 1990s. There is onetrend, however, which has been noted in every seasonal study so far: the NE Monsoon ischaracterized by increased abundance of cyclopoid copepods, such as the genera Oithona,Oncaea and Corycella, when compared with the SW Monsoon (Smith, 1988; Madhupratap etal., 1996; Smith et al., 1998). Small calanoid copepods predominate during the SW Monsoon(Smith, 1982; Madhupratap et al., 1996; Smith et al., 1998). Reproductive activity of copepodshas been briefly discussed for a few selected stations of the U.S. JGOFS study area. At thestation with the strongest oxygen depletion, Pleuromamma indica was reproductive between 250 and 300 meters in both the NE and SW Monsoon seasons, with the addition of Euchaeta rimana

54
and Amallothrix indica in the SW Monsoon and Lucicutia grandis in the NE Monsoon (Smith etal., 1998). In the upper 25 meters at this station, Undinula vulgaris and Paracalanus aculeatuswere reproductive in both the NE and SW Monsoons, with the addition of Cosmocalanusdarwini and Calocalanus plumulosus in the SW Monsoon and Acrocalanus longicornis, E.rimana and Candacia pachydactila in the NE Monsoon (Smith et al., 1998).
At a station which is within the upwelling area during the SW Monsoon, the zooplankton at 250-300 meters were dominated by P. indica, Oncaea subtilis and Clytemnestra scutellata in the NE Monsoon and by P. indica, Eucalanus crassus, Eucalanus elongatus and Heterostyliteslongicornis in the SW Monsoon. All of the SW Monsoon species have adult body lengths ofapproximately 2 mm and larger; the shift to larger body size nearshore (in the upwelling area)during the SW Monsoon is a feature not seen in other parts of the Arabian Sea. In the upper 25meters at this station, the species most abundant during the NE Monsoon are smaller sized andless abundant during the SW Monsoon. The most abundant species of the upper 25 meters in the upwelling area during the SW Monsoon are Paracalanus aculeatus, Paracalanus denudatus and Paracalanus dubius (Smith et al., 1998). Analyses of changes in the zooplankton communities at various stations explained by seasonal shifts in the phytoplankton communities and mixed layerdynamics is an area rich with possibilities for future study.
The Upwelling Area off Oman in the SW Monsoon
The upwelling areas of Oman and Somalia both contain at least one highly successful copepod(35% of total copepod biomass, Smith, 1982), Calanoides carinatus, which probably evolvedduring past periods of intensified and widespread upwelling (Fleminger, 1986). Its life historyevolved to exploit a seasonally predictable food supply (Smith, 1982). Another calanoidcopepod, Eucalanus crassus, has been observed co-occurring with C. carinatus off Somaliasuggesting a similar, yet undescribed, life history. Re-examination of samples of the upper 200meters collected in the International Indian Ocean Expedition (IIOE) of 1964-1965 have shownthat C. carinatus was collected from 24°N to 29°S in the western Indian Ocean, and from thecoast of Somalia and Mozambique to Java (Stephen et al., 1992). These two species possess high ingestion rates, and are probably capable of significantly impacting the phytoplanktonconcentration and microplankton populations in the upwelling areas off Somalia and Oman.Although the presence of C. carinatus and E. crassus at the surface is transitory and seasonal(Figure 2; June-September), when they are present, they dominate the system, and are thuspotentially important prey items for macro-invertebrates and pelagic fishes during the upwelling season.
The demographics of Calanoides carinatus have been described most fully in areas of upwelling off the western coast of Africa, particularly off Ghana and the Ivory Coast. The species is insurface layers during the upwelling season and spends the remainder of the year at mesopelagicdepths (Mensah, 1974; Binet and Suisse de St. Claire, 1975) where its respiration is fuelled bystored lipids. The lipid that can be accumulated by copepodite stage V during feeding upon theupwelling assemblage of the Benguela Current can be considerable, up to 71% of dry bodyweight (Borchers and Hutchings, 1986). The evidence from the Somali coast is consistent withpatterns established for this species on the west coast of Africa. Secondary production, primarily reproduction, is highest in cool, recently upwelled waters nearshore. As juveniles move offshore, somatic growth becomes a larger fraction of total potential secondary production (Smith, 1992).The life cycle of C. carinatus, wherever it is found, has distinctive characteristics that allow it to exploit a predictably intermittent food supply (Smith, 1982). These characteristics are rapidgrowth rate (Peterson and Painting, 1990), herbivorous feeding particularly on diatoms(Schnack, 1982), massive lipid storage by subadult stages (Borchers and Hutchings, 1986), anddiapause at mesopelagic depths (Mensah, 1974; Binet and Suisse de St. Claire, 1975).

55
The distribution of Eucalanus crassus extends throughout seasonal upwelling regions within the Indian Ocean (Tranter, 1977; data of S. Smith and M. Baars). Eucalanus crassus was mostabundant off the southern coast of Java during the SW Monsoon upwelling season (June-July;Tranter, 1977). In the Arabian Sea, E. crassus co-occurred with C. carinatus off the coast ofOman and Somalia (Smith, unpublished data). These species were present rarely at the surfaceoff the Omani coast during the non-upwelling season. During the non-upwelling season, C.carinatus is found at depths of 200–1500 meters in the central Arabian Sea (Smith and Baars,unpublished data).
What role do these large-bodied, migrating copepods play in the upwelling ecosystem? Doontogenetically migrating predators force ecosystem restructuring in the upwelling area?
Figure 2. Arabian Sea: annual (1995) cycle of wind speed (light blue line) measured by amooring positioned at 15°30’N, 61°30’E (redrawn from Weller et al., 1998); of total particulateflux (green bars) measured by a sediment trap at 858 meters positioned at 17°13’N, 59°36’E(redrawn from Honjo et al., 1999); of vertical distribution of the copepod Calanoides carinatusstage V (yellow circles) collected throughout the northern Arabian Sea from Somalia to Oman(Somali data provided by M. Baars; June data for Oman derived from samples collected by M.Wood; remainder collected by S. Smith). Insets at the top show sea-surface temperature offOman obtained by the advanced very high-resolution radiometer (AVHRR) at various timesduring the upwelling season in 1995. Darker blue depicts the area of cool sea-surfacetemperatures (redrawn from Smith et al., 1998). Note how quickly the sea-surface warmsfollowing cessation of upwelling-favourable winds in September.

56
Recommendations
1) Detailed comparisons of the species composition data from the 1960s and 1990s need to beconducted to determine climate change effects on the zooplankton communities of theArabian Sea.
2) Detailed study of seasonal successions and life cycles of dominant zooplankton species inthis region where annual life cycles are common need to be conducted.
3) Seasonal successions in zooplankton communities need to be analysed in terms of seasonalchanges in the phytoplankton and microplankton communities and the physical environment.
4) Physiological adaptations of species adapted for life in the oxygen minimum zone needinvestigation.

57
2.8 The Role of the Oxygen Minimum Zone in Biogeochemical Cycles
Sharon L. Smith
The Rosenstiel School, University of Miami4600 Rickenbacker Causeway
Miami, FL 33149USA
John M. Morrison
Department of Marine, Earth and Atmospheric SciencesNorth Carolina State University, Raleigh, NC 27695
USA
Louis A. Codispoti
Horn Point Environmental LaboratoriesUniversity of Maryland, Cambridge, MD 21613
USA
Background
The semi-annual reversal in wind stress associated with the monsoon cycle, water massintrusions from marginal seas and the other oceans, and the fact that this basin has no opening to the north and therefore no subtropical convergence or deep water formation, give the upperwaters of the Arabian Sea a unique thermohaline structure and circulation (Wyrtki, 1971;Morrison and Olson, 1992; Morrison et al., 1998). The complex water mass structure is due inpart to advection and interleaving of water masses, and in part to formation of high-salinitywaters in the Red Sea, Persian Gulf and northern portion of the basin (Arabian Sea Water),which sink to moderate depths in the central basin. For a summary of the physical domain of the Arabian Sea, see the hydrography section (Chapter 2.2) of this report. Overall, the conditionsfound in the suboxic portion of the water column in the Arabian Sea during JGOFS were notgreatly different from what has been reported in the literature with respect to oxygen, nitrate and nitrite distributions (e.g., Naqvi, 1992).
Physical Characteristics
The horizontal extent of the oxygen minimum zone (OMZ) in the Arabian Sea may varyseasonally, with its areal extent in the NE Monsoon exceeding that during the SW Monsoon(Figure 1; Eremeev et al., 1994). The upper boundary of the OMZ often occurs at sigma-thetavalues (~ 24.8 σθ) that are characteristic of the Arabian Sea Water (ASW) salinity maximum.This feature has a maximum depth of ~150 m in the northern Arabian Sea. During the NEMonsoon, this water mass can be replenished by convective processes in the northern ArabianSea (Morrison et al., 1998). Thus, the upper boundary of the OMZ receives some direct re-oxygenation as a consequence of evaporative cooling and convection during the NE Monsoon.Lower salinity water with σθ values in the range of ASW reach the sea surface in coastalupwelling that occurs during the SW Monsoon along the coast of Oman (Morrison et al., 1998), and mixing of these waters under the strong winds and intense eddy field of the SW Monsoonalso contributes to re-oxygenation of the upper waters of the OMZ.

58
Figure 1. The Arabian Sea showing the geographical position of the oxygen minimum zone(dark blue) and the cruise tracks of the U.S. observational programme (red, yellow), U.S.sediment traps (green triangles) and the air-sea interaction mooring (black bowtie).
The “core” of the suboxic layers with elevated nitrite values is more or less coincident with thePersian Gulf Water (PGW) salinity maximum layer that has its “core” at a density of ~26.6 σθ.Outflow from the Persian Gulf is only a minor component of the ambient waters in the northern Arabian Sea, but it gives it a salinity marker that is closely associated with the suboxic portionsof the OMZ (Morrison et al., 1998).
The bottom of the suboxic portions of the OMZ are roughly coincident with the characteristicdensity (~ 27.2 σθ) of Red Sea Water (RSW). This high-salinity outflow from the Red Sea sinks to depths of ~ 750 m and forms a salinity maximum in portions of the Indian Ocean, but thismaximum is weak or absent from our stations in the northern Arabian Sea suggesting that RSWmixes quickly with other water masses found at this density within the Arabian Sea, such asIndian Central Water (You and Tomczak, 1993) and North Indian Intermediate Water (Kumarand Li, 1996). The residence time of OMZ waters is relatively short, ca. 10 years (Olson et al.,1993).
The combination of source waters impoverished in oxygen entering the Arabian Sea around thetip of India (Swallow, 1984) or from the Southern Ocean (You and Tomczak, 1993), weakaeration arising from a of lack of an opening to the north, and productivity year-round driven by upwelling or convection delivering organic material to the already depleted subsurface watersare sufficient to explain the OMZ. It is also possible that lateral transfer of organic material

59
settled on the seabed of the shelf and shelf break off India intensify decomposition processesusing oxygen (Somayajulu et al., 1996).
Kim and Flagg (2001) using TOPEX/Poseidon sea-level anomalies (SLA) and shipboardacoustic Doppler current profiler (ADCP) data show that in the central Arabian Sea there is alarge increase in eddy kinetic energy in the west and southwest and to a lesser extent to the south and a large area of relatively reduced eddy activity over much of the eastern and northernArabian Sea. The area of reduced eddy activity coincides with the location of the most intenseportions of the oxygen minimum zone found in the northern Arabian Sea. They show that notonly is the level of kinetic energy lower in this area but the area also shows reduced verticalpenetration of the eddy field. This suggests that at least part of the reason for the existence of the OMZ in the area, or at least its proximity to the surface, may be the lack of mixing and verticalmotion associated with a vigorous eddy field.
During the SW Monsoon, the only organized circulation off the Arabian Peninsula is anortheastward coastal jet that develops along the south coast of Oman in early May and persiststhroughout the SW (summer) Monsoon. This Ras al Hadd Jet (Böhm et al., 1999) flows past Ras al Hadd at the southwest corner of the Gulf of Oman and meanders into the interior, transporting cooler, coastally upwelled waters with high biogenic content into the warmer environment of the Gulf of Oman and northern Arabian Sea. This is the most plausible way to transport biologicalmaterial from the upwelling region into the eastern portion of the OMZ. The distribution of theOMZ seems to match this hypothesis in that it is closest to the surface with the largest verticalextent in the northeast and gets progressively deeper and thinner to the south and to the west(Kim and Flagg, 2001).
Chemical Characteristics
The standard U.S. JGOFS Arabian Sea station sampling grid in relation to the oxygen minimum zone (Figure 1; shaded area) where the work of Naqvi (e.g., Naqvi, 1991a; Naqvi, 1991b;Naqvi, et al., 1992; Naqvi, 1994) suggests that average secondary nitrite maximum values are =1 µM. These features are called secondary nitrite maxima because they normally occur deeper in the water column than the primary nitrite maxima that are a typical feature near the base of theeuphotic zone and which arise from phytoplankton reduction of nitrate, nitrification or both(Codispoti and Christensen, 1985). Such nitrite concentrations in suboxic water (dissolvedoxygen < ~0.1 ml l-1 or <~4.5 µM or <~4.5 µmol kg-1) indicate that denitrification is a prominent respiratory process. Naqvi et al. (1992) point out that, unlike the denitrification zone off Peru(Codispoti and Packard, 1980), the zones of lowest oxygen and highest primary production aregeographically separated in the Arabian Sea. The shaded region in Figure 1 indicates thehorizontal extent of the main denitrification zone, and it is obvious that it lies offshore of theupwelling that is most intense adjacent to the Arabian Peninsula. This situation has led tospeculation about how organic matter is supplied to the denitrification zone, and the assumption is that horizontal processes are important in supplying such material (Naqvi et al., 1992). Itshould also be noted that transient secondary nitrite maxima occur outside of the shaded zone(Morrison et al., 1998).
The U.S. JGOFS programme included measurements of trace metals and their alteration by theoxygen minimum zone (Lewis and Luther, 2000; Witter et al., 2000). In the OMZ, two distinctdissolved manganese (d-Mn) maxima were observed (Lewis and Luther, 2000) at depths of 200-300 m and 600 m. The latter peak was largely confined to stations north of 19°N. This mid-depthmaximum was found in the low oxygen core of the OMZ in the region where denitrification was observed and appears to be maintained by a southward, horizontal, advective-diffusive flux of

60
dissolved manganese from highly reducing Pakistan margin sediments. Nepheloid layers,presumably bacterial, were also found at depths of 150 - 350 m (Gardner et al., 1999; Gundersenet al., 1998; Morrison et al., 1998). Particulate Mn profiles displayed corresponding minima forthis same depth interval, suggesting reductive dissolution of Mn-oxyhydroxides. Theseobservations imply active, in situ, microbially mediated redox cycling of manganese in the upper OMZ, as observed in other suboxic environments. Witter et al. (2000) measured organic Fe (III) speciation at the surface and in the OMZ during the Spring Intermonsoon season, and found that total ligand concentrations increased by a factor of 2-3 from the surface to the oxygen minimum zone. Ligand concentrations were always higher than total iron concentrations. This suggeststhat in the OMZ, ligands are produced during organic matter degradation. In addition, "excess"ligand concentrations in the OMZ were 2 to 20 times higher than surface waters (upper 100 m). Although stability constant values are comparable in magnitude to those reported in the Pacificand northwestern Atlantic oceans, measured total ligand concentrations in the Arabian Sea arehigher. This suggests that in areas that receive high Fe inputs through upwelling and/oratmospheric deposition, marine organisms produce "excess" ligands to keep Fe soluble inseawater for extended intervals.
Biological Characteristics
The oxygen minimum zone, where it occurs, occupies a large segment of the water columnextending from ~100 meters to depths of 1000-1200 meters. Earlier work has noted that forsome species assemblages in the northwestern Indian Ocean, the numbers of species declineprogressing northward from the equator: copepods of the genus Pleuromamma (Saraswathy andIyer, 1986) and myctophid fish (Kinzer et al., 1993). The abundances of those few species thathave adaptations for the OMZ increase in the northern Arabian Sea. When the OMZ impinges on the shelf break or shelf regions, catastrophic mortality occurs in the benthic communities(Vinogradov and Voronina, 1962) and demersal fish populations (Banse, 1959). Large expanses of dead fish at the surface in the open Arabian Sea have also been attributed to upwardmovement of the oxygen-depleted waters (Vinogradov and Voronina, 1962; Foxton, 1965).Vertical profiles of zooplankton biomass often show a large decrease of biomass within theOMZ layer (Vinogradov, 1970).
The Pelagic Organisms. In the Arabian Sea, several distributional patterns show the impact ofthe OMZ (Morrison et al., 1998; Herring et al., 1998): (1) exclusion from the suboxic (< 4.5µM) core of the OMZ of most zooplankton biomass and many zooplankton and nekton speciesand groups (Figure 2), but, paradoxically, (2) the occurrence of extremely high abundances of a few species of diel vertical migrators at depth during the daytime, well within the suboxic zone(Figure 2), (3) organism-specific (and probably species-specific) distribution boundaries at theupper and lower edges of the OMZ, probably associated with particular oxygen concentrations,and (4) the appearance of a subsurface abundance peak for many organisms and particles in thelower OMZ associated with the oxygen gradient between 2.3 - 4.5 µM (0.05 - 0.1 ml l-1).
Oxygen values below about 4.5 µM that extend over vertical ranges of hundreds of meters areassociated with major distributional responses of the mesozooplankton and nekton, as seen inother regions of the world’s ocean such as the Eastern Tropical Pacific Ocean. In thesesituations, most zooplankton are absent from the suboxic part of the water column, andbiomasses at depth (except for the daytime vertical migrators) are extremely low (Wishner et al.,1998). Presumably, most organisms were unable to tolerate the extremely low oxygen conditions and were excluded from the OMZ by physiological constraints. The total water column biomass (0 - 1000 m) in areas where the OMZ existed showed consistent vertical pattern. Biomass washigh in the surface layer, markedly reduced in the range of the OMZ, and increased again

61
beneath the OMZ, but not to levels observed in the surface layer (Wishner et al., 1998). Atstations away from the OMZ, the vertical profile lacked the marked reduction of biomass within the OMZ. Investigation of the distribution of the swimming crab Charybdis smithii showed thatin its one year life cycle it remained above the OMZ at all times of the year, and moved inshore over the continental shelves to breed during the NE Monsoon (van Couwelaar et al., 1997).Occasionally high concentrations of dead C. smithii (1 m-2) are observed on the seafloor;whether this is caused by fluctuations in the OMZ is unknown but when this occurs, the carbonflux via C. smithii can be equal to 30% of the annual POC flux (Christiansen and Boetius, 2000). The exclusion of larger organisms from OMZ depths suggests that sinking material transits thiszone relatively rapidly, unaltered by consumers, and arrives at depth as a relatively fresh foodsource for deeper-living organisms.
The presence of a very high biomass of diel vertical migrators that moved between the surfacewaters at night and the suboxic waters during the day was a surprising finding from the JGOFSsampling (Morrison et al., 1998; Herring et al., 1998; Luo et al., 2000; Ashjian et al., 2002).Many of these animals spent the day at depths where the oxygen was less than 0.1 ml l-1 (or 4.5 µM). It is notable that the depth to which vertical migrators descend (150-400 m) is similar tothe depths at which the intermediate nepheloid layers occur. Material transported and excretedby migrators may form a substrate for denitrifying processes. Prominent diel vertical migrators

62
in samples analysed so far were euphausiids (especially Euphausia diomedaea) and severalspecies of myctophid fish (L. Madin, pers. comm.).
The ADCP showed the diel migration and the movement into and out of the OMZ of largespecies, such as myctophid fish and euphausiids (Morrison et al., 1998; Luo et al., 2000), but asimilar migration of smaller species, such as copepods was not observed (Smith et al., 1998;Wishner et al., 1998). Daytime acoustic returns from depth were strong, and the dawn sinkingand dusk rise of the fauna were obvious. However, at night the biomass remaining in the suboxic zone was so low that no ADCP signal was detectable at these depths. ADCP records alsoshowed that there were at least two groups of plankton (used loosely), one that stayed in theupper mixed layer and another that made daily excursions. As the gaps between the mixed layer and the deep scattering layer indicate, the near-surface and deep plankton populations arecompletely separate vertically, with few plankton found in between. This was also seen in theMOCNESS profiles (Smith et al., 1998; Wishner et al., 1998). The acoustic records show thatthe vertical migration was composed of separate groups that began their migrations at slightlydifferent times. These could be different size classes of myctophids as has been observed in the Gulf of Oman (Valinassab, 2001).
An increase in zooplankton biomass and abundance typically occurred in the lower OMZ within the depth range of the oxygen gradient between 0.05 - 0.1 ml l-1 (2.3 - 4.5 µM) (where oxygenstarted to increase again) at locations with a strong and extensive OMZ (Wishner et al., 1998).This subsurface biomass feature is also known to be a location of relatively high zooplanktonfeeding rates and potential food resources (POC) (Wishner et al., 1998). In the Arabian Sea atthe depth of the subsurface biomass peak, a peak in aggregates was sometimes observed with acamera profiler (Walsh, pers. comm.). The occurrence of high plankton and microbial biomassand a potentially short food chain suggest the possibility of intense utilization and biologicalalteration of suspended and sinking particles within a narrow midwater depth zone, definedphysically by the oxygen gradients of the OMZ. The copepod Lucicutia grandis is an indicatorof the lower OMZ interface; L. grandis occurs where the oxygen gradient is 0.07-0.15 ml l-1
(Wishner et al., 2000). In this habitat at 600-1000 metres and within the OMZ, L. grandis isreproductive year-round and feeds on a wide range of material (surface–derived, deep-seadetritus, zooplankton and aggregates; Wishner et al., 2000). Results from the JGOFS sedimenttrap programme also suggest that substantial changes in the quantity and quality of the sinkingflux, possibly due to zooplankton processes, occurred within this depth zone (Lee et al., 1998).
Various taxa in the subsurface biomass peak showed different distributional responses to theoxygen gradients, probably because of physiological differences in their ability to cope with low oxygen. For example, the abundance peak of gelatinous taxa such as salps and doliolids wasrelatively shallow, where oxygen was still very low in the OMZ. Crustaceans (copepods, shrimp) and fish (Cyclothone) apparently required more oxygen and increased deeper in the watercolumn where oxygen was somewhat higher.
Another group of organisms, the seasonal migrants such as the copepod Calanoides carinatus,spent a portion of their life cycle in the suboxic zone and in the more oxygenated water beneath it, mostly in a non-feeding, diapausing stage. They rose into the surface waters in large numbers, where they fed and reproduced (Smith et al., 1998) during the SW Monsoon. Thus, although the resident zooplankton biomass in the core of the OMZ was extremely low, there were severalassemblages of organisms that inhabited parts of the suboxic zone, especially its upper and lower edges, at different times of the day or year.
Zooplankton biomass profiles showed substantial geographic variability across the Arabian Sea, but comparatively little change between seasons at particular stations, except in the surface water

63
and nearshore (Wishner et al., 1998). The consistency of the biomass profiles through timeimplies long-term stability in the structure and function of the mid-water ecosystem over a broad geographic region and shows the strong influence of the OMZ on the biology of the deepArabian Sea.
A deep chlorophyll a maximum showing high primary productivity was identified in the SpringIntermonsoon of 1987 (Pollehne et al., 1993) but the cells composing the layer could not beidentified. The position of this layer at the bottom of the euphotic zone but in high nutrientssuggested it was a major component of new production and sub-euphotic zone flux. Itsrelationship to the OMZ was not identified. In the Fall Intermonsoon of 1986, again subsurfacemaxima of cyanobacteria were identified but the role of the OMZ was not discussed (Burkill etal., 1993). During the U.S. JGOFS cruises of 1995, details of the oxic-suboxic interfaces of theOMZ were investigated. Johnson et al. (1999) found secondary fluorescence maxima (SFM)near the oxic-suboxic interface at the base of the euphotic zone. “These secondary peaks weremost prominent during the early NE Monsoons in the central oligotrophic portion of the Arabian Sea…Based on high performance liquid chromatography…and flow cytometry analyses, SFMwere determined to be populated almost exclusively by the marine cyanobacteriumProchlorococcus spp…. SFM were about half the magnitude of primary fluorescencepeaks…Photosynthesis versus irradiance response curves revealed an efficient populationadapted to extremely low light (~0.02-0.05% surface irradiance) largely through increased lightabsorption capabilities…Deck-incubated C-14 uptake as well as dilution growth experimentsrevealed instantaneous growth rates on the order of µ = 0.01/d…” In another study (Goericke etal., 2000), the OMZ is implicated in the distribution and physiology of Prochlorococcus in theArabian Sea but not conclusively. They state, “…we report discovery of populations ofProchlorococcus in layers below the oxyclines of the oxygen minimum zones of the Arabian Sea and the Eastern Tropical Pacific off Mexico. The unusual aspects of these populations are thatthese were at times virtual monoalgal cultures found at a depth of 80 to 140 m…As onlyProchlorococcus thrives in these layers but Prochlorococcus and eukaryotic picoplanktoncoexist in and below subsurface chlorophyll maxima, we conclude that the low oxygenconcentrations at the deep Prochlorococcus maxima are the determining factor, but we are notable to identify any specific physiological functions that are affected by low oxygenconcentrations in eukaryotes but not Prochlorococcus.” The last fifteen years have demonstrated the huge impact of new instrumentation that permits routine identification and sorting ofplanktonic individuals and new methods for describing plant pigments.
The Benthic Organisms. Coincident investigations addressed the effect of the OMZ on thebenthic biota. Where the OMZ impinges on the continental slope off Oman, spionid wormsshowed significantly increased surface area of their gas-exchange organs (size and number ofBranchiae), which is presumably an adaptive response to the low oxygen (Lamont and Gage,2000). The diversity and evenness of the benthic community impacted by the OMZ was reduced compared to similar communities unaffected by suboxic conditions, but abundance and biomass of the community in the OMZ were greater than outside the OMZ (Levin et al., 2000). Thecommunity living in contact with the OMZ was composed primarily of worms and an ampeliscid amphipod (Levin et al., 2000). Food availability (surface pigment concentration) and oxygenconcentration explained most of the variance in species richness, diversity and eveness.Bioturbation is another aspect of the benthic habitat and geochemistry that is altered by an OMZ impinging on a shelf or slope. Bioturbation in sediments of the Oman margin was reduced incomparison with sediments impacted by oxygenated waters in the Atlantic and Pacific oceans(Smith et al., 2000). It is noteworthy that in both the pelagic and benthic ecosystems, the OMZof the Arabian Sea occurs in association with reduced diversity, reduced evenness but increasedbiomass of the few species which can withstand suboxic conditions.

64
Recommendations
1) Since the OMZ is a source of nitrogen to the atmosphere, and rates of denitrification heremay be much higher than previously estimated, the oceanic nitrogen cycle and its impact on climate change need to be studied.
2) Daily migration of large biomass into and out of the OMZ, probably mostly myctophids butalso Euphausiids and copepods, suggests physiological adaptations that have not beeninvestigated yet.
3) The potential role of physical forcing in determining the position and strength of the Arabian Sea’s OMZ needs explicit investigation.
4) The OMZ may be an indicator of climate change exhibiting rapid response to perturbationand therefore merits monitoring in this context.

65
2.9 Sedimentation
T. Rixen*, M.V.S. Guptha+ and V. Ittekkot*
* Zentrum für Marine Tropenökologie, Universität BremenFahrenheitstraße 6, D-28359 Bremen
Germany
+ National Institute of OceanographyDona Paula, Goa 403 004
India
Introduction
One of the major JGOFS tasks is to quantify the export of carbon from the ocean’s surface intothe ocean's interior, and to shed light on processes coupling carbon flux and climate. Sedimenttraps are at present the only tools that can intercept directly and continuously particles settlingthrough the water column but their trapping efficiency, and thus, their accuracy can be biased by hydrodynamic and biological effects (Lee et al., 1988; Gust et al., 1992; 1994). Scholten et al.(2001) showed that the 230Thorium trapping efficiency of sediment traps moored at differentsites and water depths in eastern North Atlantic Ocean varies between 9 and 143%. In general,these trapping efficiencies increase with increasing water-depth (Scholten et al., 2001). Bycomparing data from various ocean basins, Yu et al., (2001), found that sediment trap dataobtained at water depths above 1200 m are problematic whereas traps deployed below that depth exhibit acceptable trapping efficiencies. The latter is supported by the Indo-German sedimenttrap experiment that showed co-variations between surface ocean processes and deep oceanfluxes measured between 1986 and 1997 in the western, central and eastern Arabian Sea (e.g.,Nair et al., 1989; Haake et al., 1993; Rixen et al., 1996; 2000).
During the Dutch, German, Indian and the U.S. JGOFS field experiments between 1993 and1995 approximately 40 moored sediment traps were deployed at different water depths in theArabian Sea (Figure 1). The U.S. sediment trap moorings were arranged along a transect fromthe highly productive upwelling area off Oman past the long-term Indo-German sediment trapsites in the western and central Arabian Sea into the oligotrophic southern Arabian Sea. TheDutch activities included sediment trap experiments in the upwelling area off Somalia and inaddition, one trap was deployed in the northern Arabian Sea in collaboration with Pakistan andGermany.
The Monsoonal Impact on Deep Ocean Fluxes
The seasonal reversing monsoon winds over the Arabian Sea are stronger during summer (SWMonsoon) and winter (NE Monsoon) than during the Intermonsoon periods. This is reflected byincreased particle fluxes in the deep western, central and eastern Arabian Sea pointing to a close coupling between the monsoon and the deep sea (Nair et al., 1989). During the JGOFS fieldexperiments this bimodal, monsoon-driven flux pattern, was also observed in the northernArabian Sea off Pakistan (Suthhof et al., 1999) and along the U.S. JGOFS sediment trap transect in the western Arabian Sea (Lee et al. 1998; Honjo et al., 1999) and off Somalia (Broerse et al.,2000). In the southern open Arabian Sea this pattern is only weakly evident and the peak organic carbon fluxes are even lower than those during the Intermonsoon periods at the other sites(Honjo et al., 1999; Rixen et al., submitted). This implies that the southern boundary of themonsoon-driven high flux region is located somewhere between 10 and 15°N in the openArabian Sea.

66
Within the monsoon-driven, high flux region, the organic carbon transport into the deep sea isgenerally lower during the NE than during the SW Monsoon. This difference is mostpronounced off Somalia (Broerse et al., 2000), in the western Arabian Sea and disappears almost entirely in the northern Arabian Sea off Pakistan where the NE and SW Monsoon fluxes arealmost equal (Suthhof et al., 1999). However, these results demand interdisciplinaryinvestigations such as those carried out during the international JGOFS to elucidate theprocesses that transfer monsoonal signals into to the deep sea.
The Southwest (SW) Monsoon
The SW Monsoon is characterised by a low level Jet (Findlater Jet) blowing almost parallel tothe Arabian coast over the Arabian Sea (Findlater, 1966, 1969; Rixen et al., 1996). In thewestern Arabian Sea north of the axis of the Findlater Jet, the monsoon winds induce upwelling, which is strongest along the Oman coast (e.g., Luther et al., 1990; Brock et al., 1991; Rixen etal., 2000). South of the Findlater Jet axis, downwelling occurs due to the wind stress curl.During the U.S. JGOFS field experiments, the ONR Arabian Sea Central Mooring (ASCM) was deployed almost below the axis of the Findlater Jet between the western and central Arabian Sea trap sites (Dickey et al., 1998). It revealed a deepening of the mixed layer during the initial onset of the SW Monsoon in 1995, while in the course of the SW Monsoon the mixed layer becameshallower (Figure 2). The initial mixed layer deepening was due to downwelling or windstirring, whereas the decreasing mixed layer depth can result from upwelled water propagatingoffshore. This agrees with the mean SW Monsoon organic carbon fluxes that generally decrease with increasing distance from the coast (Figure 1). The deviation from this trend close to thecoast is probably caused by marine organisms which are assumed to be exported into the deep-sea during their later resting stage and not during their earlier growing stage (Smetacek, 1985).On the other hand, spatial and temporal variations during the advection of upwelled water(Manghnani et al., 1998) can also contribute to the observed deviation.
However, during the early onset of the SW Monsoon, increasing organic carbon fluxes areassociated with enhanced carbonate to biogenic opal ratios at all long-term trap sites (Figure 3;Rixen et al., submitted). This indicates an increasing share of carbonate-bearing organisms in the organic matter exported into the deep sea. During the SW Monsoon, the carbonate-to-biogenic
Figure 1. Sediment trap sites in the Arabian Sea.Black dots show the long-term Indo-Germanssediment trap locations (deployed since 1986),black squares indicate the Dutch sediment trap sitesoff Somalia (deployed: 1992/1993; e.g., Koning etal., 1997), light grey circles reveal the U.S.locations (deployed: 1994/1995; Lee et al., 1998,Honjo et al., 1999) and the open circle shows theGerman/ Pakistan site (deployed: 1994/1996;Suthhof et al., 1999). The lower panel shows themean SW Monsoon fluxes obtained by trapsdeployed at a water-depth of approximately 3000 mversus the distance between the trap site and thecoast. Data are from Lee et al. (1998; grey circles)and from Rixen et al. (submitted, black circles).

67
opal ratios decrease due to a higher contribution of silicate-bearing organisms such as diatoms to the exported matter. This feature is well known in the western Arabian Sea and has been related to the nutrient distribution in the water column showing an increase in silicate concentrationsthat is steeper than that of nitrate and phosphate (Haake et al., 1993). The initial upwelling ormixed layer deepening seems, thus, to be too weak to carry sufficient silicate into the euphoticzone to favour diatom blooms. This situation changes during the later phase of the SW Monsoon when an increasing influence of coastal upwelling leads to diatom blooms at the trap site in thewestern Arabian Sea (Rixen et al., 2000). Detailed plankton studies during this period revealedthat flagellates succeed diatoms between the western and central Arabian Sea (Garrison et al.,2000) probably due to the lack of dissolved silicate. This might be responsible for the relativelyhigh carbonate/biogenic opal ratios during the later phase of the SW Monsoon in the centralArabian Sea compared to those measured at the other sites (Figure 3).
Figure 2. Depth of the mixed layer (MLD black line) and euphotic zone (1% PAR light level; green line) measured at the ONR central Arabian Sea mooring between October 1994 and 1995. The data are redrawn from Dickey et al. (1998). MLD represents the water-depth at which the temperature is 1°C lower than at the surface. Additionally, the organic carbon fluxes (blue line) and the carbonate biogenic opal ratio (red line) determined at the sediment trap site (WAST) during the year 1995 are shown.
Tracer experiments carried out with a 3-D coupled ocean-atmosphere model (Maier-Reimer,submitted) indicate that nutrients upwelled off Oman can propagate along with the coastalcurrent into the northern Arabian Sea, and further along the Indian coast into the eastern Arabian Sea. Thus, the enhanced fluxes at these sites might also be influenced by the upwelling offOman. This implies that the western Arabian Sea upwelling influences the peak organic carbonexport throughout the open Arabian Sea north of at least 15°N during the SW Monsoon and also causes the low carbonate/biogenic opal ratios in eastern Arabian Sea. Off Somalia, the high SW Monsoon fluxes are linked to upwelling occurring at the western exteriors of the Great Whirl(e.g., Conan and Brummer, 2000).
To gain more quantitative information on the link between deep ocean fluxes and upwelling,biweekly averaged wind fields derived from satellite measurements for the period between 1987 and 1995 were converted into up- and downwelling velocities. Subsequently, upwellingvelocities off Oman were correlated with deep ocean fluxes obtained at the western Arabian Sea trap site (Rixen et al., 2000). This analysis revealed that changes in the upwelling velocitiescaused by variations in the strength of the Findlater Jet exert the main control of organic carbon

68
fluxes into the deep western, (Rixen et al., 2000b), and most likely also into the central, eastern and northern Arabian Sea, during the SW Monsoon.
Figure 3. The mean annual organic carbon fluxes and carbonate to biogenic opal ratios at thetrap sites in the western (WAST), central (CAST) and eastern Arabian Sea (EAST).
The Northeast (NE) Monsoon
During the NE Monsoon, mixed layer deepening caused by winter cooling and wind mixingentrains nutrient-enriched subsurface waters into the euphotic zone (e.g., Banse and McClain,1986; Madhupratap et al., 1996). The resulting phytoplankton bloom has been assumed to cause the enhanced organic carbon fluxes measured in the deep western, central and eastern ArabianSea (Nair et al., 1989; Haake et al., 1993). During the NE Monsoon of 1994/95, the ONRASCM buoy recorded the deepest mixed layer approximately between December and February(Figure 2). The respective euphotic zone was almost half as deep as the mixed layer and thecorresponding organic carbon fluxes in the western Arabian Sea were even lower than during the following Spring Intermonsoon period (May/June; Figure 2; Rixen et al., submitted). The peakorganic carbon flux occurred at the end of the NE Monsoon as the mixed layer became shallower

69
than the euphotic zone. High organic carbon fluxes at the end and low fluxes during the peak of the NE Monsoon are a regular, repeating feature that have been observed not only in the western but also in the central and eastern Arabian Sea. A second organic carbon flux peak was generally measured at the beginning of the NE Monsoon in December during the initial deepening of themixed layer (Rixen et al., submitted). A similar bimodal flux-pattern during the NE Monsoonwas also observed at the U.S. and the Dutch sediment trap sites, which shows that it is acharacteristic feature in the NE Monsoon season in the Arabian Sea (Prahl et al., 2000; Broerse, et al., 2000). The interrelation between mixed layer depth and depth of the euphotic zone leading to the early and late NE Monsoon blooms is similar to those causing autumn and spring bloomsin the temperate open oceans (Kiefer and Kremer, 1981).
In the Arabian Sea, the bloom of the early NE Monsoon is associated with enhanced carbonate/biogenic opal ratios, whereas the sinking particles during the bloom of the late NE Monsoonreveal low carbonate/biogenic opal ratios (Figure 3). This indicates that in contrast to the earlyNE Monsoon bloom, the late NE Monsoon organic carbon flux peak is mainly driven by diatom blooms. Since the early NE Monsoon, bloom is caused by the initial deepening of the mixedlayer and the late NE Monsoon bloom succeeds the maximum mixed layer depth, the shift in the phytoplankton composition can be related to the depth from which the water in the euphoticzone originates.
Organic Carbon Export into the Ocean’s Interior
The organic carbon flux from the euphotic zone into the deep-sea and its remineralisation in the water column is assumed to be associated with primary production (Deuser and Ross, 1980).Primary production and sediment trap data are used to develop models describing this link byincluding an additional term accounting for the influence of water-depth on organic carbonfluxes (e.g., Suess, 1980; Betzer, 1984; Pace et al., 1987; Berger; 1988). These models havebeen applied for global analysis (e.g., Jahnke, 1996) and are often integrated into largecirculation models utilised for climate predictions (e.g., Maier-Reimer, 1993). The organiccarbon export derived from the 234Th-method reveal, in contrast to the assumption within thecarbon flux models, a decoupling between primary and export production (Buesseler, 1998;Buesseler et al., 1998). However, the annual averaged primary production rates (Buesseler et al.,1998) converted into deep ocean organic carbon fluxes by using the models, correlate with theorganic carbon fluxes measured by sediment traps at water-depths > 1000 m (Table 1). Inaddition to this, the model introduced by Pace et al. (1987), shows the best quantitativeagreement between measured and modelled data (Figure 4). The correlation can be improved by running the models with primary production rates derived from the CZCS between 1978 and1986 (e.g., Antoine et al., 1996; Behrenfeld and Falkowski, 1997; Table 1). Here, the primaryproduction rates were averaged for an area of one degree around the trap sites and the U.S. flux data were reduced by 11% since the 1994 and 1995 data are 11% above the mean annual flux as indicated by the long-term record in the western Arabian Sea (Rixen et al., 2000b). Despite theimproved correlations, the data derived from the Pace et al. (1987) model are slightly lower than the measured ones (Figure 4). This might be attributed to an underestimation of the satellite-derived mean annual primary production rates due to insufficient data coverage during thehighly productive SW Monsoon (Banse and English, 2000). Nevertheless, the agreementbetween modelled and measured data can be improved by modifying the coefficients or by using the coefficients originally introduced by Betzer et al. (1984) (Table 1; Figure 5).
The ratios between primary and export production implied by Betzer et al. (1984) are higherthan those reported by Buesseler (1998; generally < 0.1) and Buesseler et al. (1998; Figure 6).
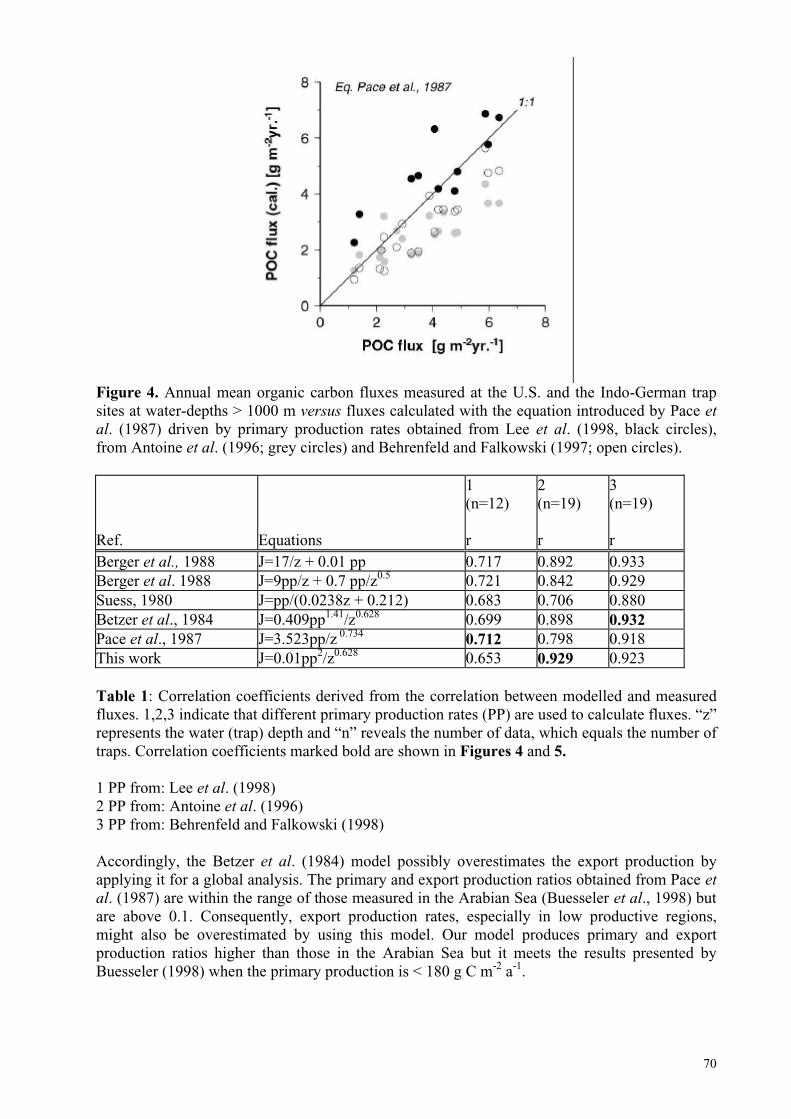
70
Figure 4. Annual mean organic carbon fluxes measured at the U.S. and the Indo-German trapsites at water-depths > 1000 m versus fluxes calculated with the equation introduced by Pace etal. (1987) driven by primary production rates obtained from Lee et al. (1998, black circles),from Antoine et al. (1996; grey circles) and Behrenfeld and Falkowski (1997; open circles).
Table 1: Correlation coefficients derived from the correlation between modelled and measuredfluxes. 1,2,3 indicate that different primary production rates (PP) are used to calculate fluxes. “z” represents the water (trap) depth and “n” reveals the number of data, which equals the number of traps. Correlation coefficients marked bold are shown in Figures 4 and 5.
1 PP from: Lee et al. (1998)2 PP from: Antoine et al. (1996)3 PP from: Behrenfeld and Falkowski (1998)
Accordingly, the Betzer et al. (1984) model possibly overestimates the export production byapplying it for a global analysis. The primary and export production ratios obtained from Pace etal. (1987) are within the range of those measured in the Arabian Sea (Buesseler et al., 1998) but are above 0.1. Consequently, export production rates, especially in low productive regions,might also be overestimated by using this model. Our model produces primary and exportproduction ratios higher than those in the Arabian Sea but it meets the results presented byBuesseler (1998) when the primary production is < 180 g C m-2 a-1.
Ref. Equations
1(n=12)
r
2(n=19)
r
3(n=19)
rBerger et al., 1988 J=17/z + 0.01 pp 0.717 0.892 0.933Berger et al. 1988 J=9pp/z + 0.7 pp/z0.5 0.721 0.842 0.929Suess, 1980 J=pp/(0.0238z + 0.212) 0.683 0.706 0.880Betzer et al., 1984 J=0.409pp1.41/z0.628 0.699 0.898 0.932Pace et al., 1987 J=3.523pp/z 0.734 0.712 0.798 0.918This work J=0.01pp2/z0.628 0.653 0.929 0.923

71
Figure 5. Annual mean organic carbon fluxes measured at the U.S. and the Indo-German trapsites at water-depths > 1000 m versus modelled fluxes. “PP”=Primary production; “TP”=sediment trap depth.
Figure 6. Primary versus export production as derived from various equations and from Arabian Sea data (circles; data are from Buesseler et al., 1998).
To quantify carbon export into the ocean interior we used the model introduced by Pace et al.(1987) and our equation. The primary production rates (Antoine et al., 1996) were interpolatedon a one-degree grid. The resulting export production into the Arabian Sea north of 6°N (3.52 X 1012 m2 excluding shelf and slope regions; water-depths > 2000 m), ranges between 84 to 91 X1012 g C a-1. This is approximately 50% lower than the export production rates derived fromnitrate assimilation measurements (Watts et al., 1999; Bange et al., 2000). However, the globalorganic carbon flux at a water-depths of 2000 m resulting from our equation and from Pace et al.(1987) agrees quite well with the mean annual fluxes derived from a global summary of

72
sediment trap data (340 X 1012 g C a-1; Lampitt and Antia, 1997; Table 2). Overall, these datashow that the Arabian Sea accounts for approximately 3% of the global organic carbon transport into the ocean’s interior.
Table 2. Annual mean carbon fluxes derived from primary production rates (Antoine et al.,1996; Behrenfeld and Falkowski, 1997). PP = primary production; Exp100 = export production;Exp2000 = organic carbon flux at 2000 m water-depth; Surf. Sed. = the organic carbon flux which reaches the sediment surface (water-depths were obtained from:http://ingrid.ldgo.columbia.edu/SOURCES/WORLDBATH).
PP[1012 C g]
Exp.100
[1012 C g]Exp.2000
[1012 C g]Surf. Sed.[1012 C g]
Arabian Sea (area: 3.5 1012 m2)J=0.01pp2/z0.628; PP=1 700.1 86.3 13.2 9.8J=3.523pp/z 0.734; PP=1 700.1 84.0 9.3 6.4J=3.523pp/z 0.734; PP=2 761.7 91.4 10.1 7.1
global (area: 302.1 012 m2)J=0.01pp2/z0.628; PP=1 29924.9 1890.7 288.1 188.1J=3.523pp/z 0.734; PP=1 29924.9 3588.7 398.1 238.0J=3.523pp/z 0.734; PP=2 34484.6 4135.6 458.8 274.0
1 Antoine et al. (1996); 2 Behrenfeld and Falkowski (1997)
Deep-sea Fluxes and Sedimentary Organic Carbon Burial
The difference between the deep-sea organic carbon fluxes and the organic carbon burial rates in the underlying sediments allows us to estimate the remineralisation rates of organic carbon at the sediment-water interface. Nevertheless, it should be considered that the top 1 cm of the surfacesediment should have an age of several hundred years according to the sedimentation ratespublished by Paropkari et al. (1992) and the longest record of deep-sea fluxes is approximately 9 years (Rixen et al., submitted). However, Jahnke (1996) used calcium carbonate-correctedorganic carbon burial rates and data on oxygen consumption measured in the Atlantic andPacific oceans to study the relation between organic carbon fluxes and burial rates. Heintroduced a regression equation that describes the relationship between oxygen consumptionand calcium carbonate-corrected organic carbon burial. After converting oxygen consumptioninto organic carbon remineralisation, this equation could be transformed so that it allows thecalculation of the organic carbon fluxes at the sediment surface. This transformed equation fitsthe data derived from the Arabian Sea (Rixen et al., 2000b), which supports the global validityof the equation introduced by Jahnke (1996). However, Jahnke (1996) suggested that globally396 X 1012 gC y-1 reaches the surface sediments, whereas our analysis results in an organiccarbon flux of 188 to 274 X 1012 gC y-1 (Table 2). Although this, in addition to the resultsdiscussed previously, indicates an agreement between the different global approaches forestimating the deep ocean carbon budget, within the order of a factor of two, there are significantlocal mismatches. For example, our analysis shows higher organic carbon fluxes along sea floor spreading zones compared to Jahnke (1996) (Figure 7).

73
Figure 7. Annual mean organic carbon fluxes reaching the sea floor (water-depths > 2000 m)derived from primary production rates (Antoine et al., 1996), water-depth(http://ingrid.ldgo.columbia.edu/SOURCES/WORLDBATH) and the equation: POC flux =0.01pp2/z0.628 (pp=primary production, and z= water depth).
Conclusion
Similar to temperate open ocean environments, variations of the depth of the mixed layer and the euphotic zone control the organic carbon export in the Arabian Sea except in regionsinfluenced by upwelling during the SW Monsoon. The strength of the upwelling and the heightof the subsequent export of organic matter are mainly determined by the intensity of theFindlater Jet.
Moreover, the strength of the physical forcing mechanism, also influences the planktoncomposition whereby stronger forcing favours diatom blooms and lowers the carbonate/biogenicopal ratios. Such changes, so far attributed to an enhanced dust supply, are assumed to havelowered the atmospheric CO2 concentration significantly during glacial times (Harrison, 2000;Tréguer and Pondaven, 2000). The marine silica cycle, however, is still poorly understood andnew investigations are needed to adequately integrate the silica cycle into global models (Dittert et al., 2001).
Satellite-derived primary production rates can be used to estimate deep ocean fluxes onannual time-scales by using organic carbon models such as those introduced in this study and by Pace et al. (1987). In the Arabian Sea, the differences between modelled and measured deepfluxes are lower than by a factor two. Extending this approach globally results in a mean deep-sea flux, which meets those derived from sediment trap data within the same error range. Theimplied primary/export production ratios are close to those reported by Buesseler (1998).
The export production rates for the Arabian Sea range between 83 and 91 gC y-1 and account for approximately 3% of the global open ocean export production (excluding shelf and sloperegions). This suggests that the export production in the Arabian Sea is close to the estimatedannual emissions of CO2 from the Arabian Sea into the atmosphere (74-94 X 1012 gC;Somasundar et al., 1990; George et al., 1994; Körtzinger et al., 1997; Ewald, 1998) and morethan twice as high as those suggested by Goyet et al. (1998). Accordingly, small changes in themarine biosphere could have the potential to influence the CO2 emissions significantly.
In the Arabian Sea, organic carbon burial rates can be related to primary production andcarbonate accumulation rates as suggested by Jahnke (1996). Jahnke’s and our budgets reveal an acceptable agreement on a global scale. Problems, however, occur on regional scales in, forexample, sea floor spreading zones and continental margins.

74
2.10 Arabian Sea Benthic Dynamics
Greg Cowie
Marine and Environmental Geoscience Group, Department of Geology and GeophysicsUniversity of Edinburgh, West Mains Road, Edinburgh EH9 3JW
UK
Chemical and biological processes occurring in the benthic boundary layer (BBL) and surficialsediments are of central importance to the biogeochemical cycling and fluxes of numerous keyelements, and thus were an important focus of the international JGOFS research programme.The Arabian Sea was recognised as an exceptional site for studies of these processes (cf. JGOFS Report 17). Strongly seasonal and spatially variable productivity results in a correspondinglyvariable supply of organic material to the BBL. The different margins (e.g., Africa/Arabia versusIndia/Pakistan) experience widely different oceanographic conditions (circulation, upwelling,lithogenic input, seasonality, and so forth). These differences, combined with an intense, basin-wide oxygen minimum zone (OMZ) that impinges on the continental margins, result in anexceptional range of BBL and sedimentary environments, with respect to critical factorsincluding food supply, redox conditions, and the nature and activity of benthic communities.Therefore, different regions of the Arabian Sea, and locations across its margins, offer strikingcontrasts likely to provide important and generally applicable insights on the nature of benthicprocesses and on their significance to the global cycles of key elements and to the makeup ofsedimentary records.
Because of the unusual conditions that prevail, the sediments and benthic processes of theArabian Sea are also of major significance in their own right. For example, high accumulationrates, particularly on the margins, produce detailed records of past changes in climatic andoceanographic conditions (e.g., in the intensity of the monsoons and associated upwelling andproductivity, and in the extent and intensity of the OMZ). In some locations (e.g., the Indusmargin), surficial sediments from within the OMZ are undisturbed by macrofauna, resulting inlaminated (varved) sediments offering particularly detailed Holocene records (ca. 1 mm y-1).Furthermore, margin sediments in the Arabian Sea are, in general, unusually rich in organicmatter. This may represent a globally significant long-term carbon sink. Indeed, such marginsettings, with high productivity and severe oxygen depletion, are often cited as the type ofdepositional environment for organic-rich facies found in the geological record. Benthic studiesunder the strongly contrasting depositional conditions across the Arabian Sea are likely toprovide new, comparatively unambiguous, information on the relative importance ofcontributing factors (e.g., productivity versus oxygen depletion), which remains the subject ofconsiderable debate.
Benthic processes within the OMZ of the Arabian Sea are also likely to be important to theglobal cycles of N and P and to the generation of the mid-water OMZ. The relative importanceof sedimentary denitrification in the Arabian Sea remains unclear, but, given the areal extent ofthe OMZ and the amount of organic matter remineralised across the BBL, it may be significantrelative to rates occurring in the water column, which account for ca. 30% of the global pelagic total. Consequently, the sediments may also make a significant contribution to the net flux of the greenhouse gas N2O that occurs from the Arabian Sea. Similarly, burial of authigenic P in thereducing margin sediments of the Arabian Sea has potential global significance for oceanic Pinventories and productivity. The contrasting redox conditions across the margins of the Arabian Sea also potentially offer valuable information on the cycling of redox-sensitive metals and onthe nature and importance of suboxic microbial processes that use Fe and Mn oxyhydroxides as

75
electron acceptors. Studies of these and other biogeochemical processes should clarify the role of the sediments in determining the oxygen depletion and other unusual features of Arabian Seawater-column chemistry.
In recognition of the broad significance of Arabian Sea benthic processes, benthic studiesbecame an important objective of the international JGOFS research programme (see JGOFSReport 17). The proposed studies built on various investigations of the physical, biological andgeochemical characteristics of surficial deep-sea and marginal Arabian Sea sediments carried out over the last century during several past expeditions to the Arabian Sea. The U.S. JGOFSprogramme (1994-1996), involving some 17 cruise legs, was largely centred on water-columnprocesses and was carried out mainly in the western basin of the Arabian Sea and on the Omanmargin. However, in addition to some sediment coring, the programme included sediment trapdeployments in various locations and with differing sampling strategies (depths, durations,sampling intervals), which are providing important information on the magnitude, compositionand seasonality of particle fluxes to the underlying BBL, and the degree of alteration that occurs in the water column.
Benthic processes were a more central focus of the Dutch, UK, German, and Indianprogrammes. The Dutch programme involved 11 RV Tyro cruises in 1992-1993 and included an important benthic component (geological and biogeochemical) focused on both deep-sea andmarginal sediments (both Oman and Pakistan margins). Eight German JGOFS cruises werecarried out in 1995-1997 and included sediment trap deployments as well as a scientificprogramme that included a major sedimentary component, focused on benthic geochemistry and biology (microbial and macrofaunal). The major study sites were at five abyssal stations wherelong-term sediment trap deployments were made, in the northern, southern, eastern, central andwestern basins, but ancillary FS Sonne cruises (PAKOMIN, 1993) also involved trapdeployments and detailed sediment studies on transects across the Pakistan margin. The UKJGOFS programme (ARABESQUE) involved four RRS Discovery cruises in 1994, one of which was dedicated to linked studies of the biology and biogeochemical processes of sediments on the southern Oman margin (biological surveys, sediment accumulation and bioturbation rates,benthic respiration and fluxes, organic geochemistry). Finally, the Indian JGOFS programmeinvolved 5 cruises on ORV Sagar Kanya (1992-1995) in the eastern Arabian Sea, and includedstudies of active sedimentary processes (sediment trap deployments and sediment coring), withthe objectives being to ascertain burial rates of biogenic elements in deep-sea and marginalregions and the role of the continental margins in the removal/supply of material to the Arabian Sea.
The Arabian Sea JGOFS studies outlined above are at various stages of completion and in manycases, the results have yet to be published. Nonetheless, some important new findings thatconcern benthic processes in the Arabian Sea have appeared recently, as a result of JGOFS andancillary research projects. The subject areas of both geochemical and biological studies arediverse.
First, as outlined above, a number of programmes (German, U.S. and Indian), in both deep-seaand margin settings, have included sediment trap deployments at various depths and durations(e.g., Lee et al., 1998). These are yielding valuable information on the magnitude, seasonalityand composition of the pulsed particle inputs to the BBL, how these vary with region, and howthey impact benthic communities, fluxes and C cycling (e.g., Andruleit et al., 2000; Boetius and Lochte, 2000; Boetius et al., 2000a,b; Broerse et al., 2000; Conan and Brummer, 2000; Grandel et al., 2000; Honjo et al., 1999; Koppelmann et al., 2000; Pfannkuche et al., 2000; Rixen et al.,2000a,b; Sommer and Pfannkuche, 2000; Turnewitsch et al., 2000; Witte and Pfannkuche, 2000; Witte, 2000). Further information is provided on the nature and extent of particle alteration

76
within the water-column or across the BBL (e.g., Hedges et al., 2001), and implications forinterpreting sedimentary records (e.g., Prahl et al., 2000), and on the relative importance ofbenthic processes in generating the OMZ and other features of water-column chemistry. Therecent trap deployments build on near-continuous deployments made by German and Indiangroups at open-ocean sites in the eastern, central and western Arabian Sea since the 1980s (e.g.,Ramaswamy and Nair, 1994; Rixen et al., 1996).
Studies of benthic biogeochemical processes during the Dutch, UK and German expeditionsincluded various down-core investigations of the solid-phase and pore-water chemistry ofsurficial sediments from both marginal and deep-sea sites. Detailed profiling of oxygen,nutrients and trace metals in the porewaters was carried out, coupled with determinations ofsolute fluxes across the benthic interface (via benthic chambers) and uptake rates of differentelectron acceptors. Along with assessments of microbial community structure and activity, these results provide key information on the role of benthic processes in C cycling in the Arabian Sea, and how C burial and the contributions of aerobic and anaerobic processes vary with location.Comparatively little of the work on process rates is published yet. Somayajulu et al. (1996) used Ra isotopes to estimate the contribution of margin sediments to total denitrification in theArabian Sea, and Schenau et al., (2000) recently provided evidence for active phosphoriteformation in surficial OMZ sediments. Schmaljohann et al. (2001) also have reported on theoccurrence of sulphur oxidizing bacteria in OMZ sediments, which, along with biomarker andstable isotopic data (e.g., Cowie et al., 1999; Schulte et al., 2000), may indicate an importantrole for chemoautotrophic C sources within the OMZ. Other published studies on benthicprocesses have mainly been limited to abyssal sites from contrasting regions of the Arabian Sea. These have included studies on near-bottom and sedimentary bacterial activity (Boetius andLochte, 2000; Boetius et al., 2000a, b), benthic respiration rates (Witte and Pfannkuche, 2000)and nutrient fluxes (Grandel et al., 2000), bioturbation rates (Turnewitsch et al., 2000), sulphurisotopes as indices of sulphate reduction rates (Bottcher et al., 2000) and distributions ofbiogenic compounds as indices of benthic community activity (Pfannkuche et al., 2000), andhow these respond to temporal and spatial variability in particle fluxes. Luff et al. (2000) used a biogeochemical modelling exercise to compare C cycling and benthic process rates at the samesites, based on observed porewater and solid-phase compositions. Other published compositional information on surficial Arabian Sea sediments includes assessments of the extent of sulphatereduction (e.g., Littke et al., 1997; Luckge et al., 1999), clay mineral distributions (e.g., Rao and Rao, 1995; Chauhan and Gujar, 1996) and the biochemical, inorganic and stable-isotopiccompositions of Pakistan, Indian and Oman margin sediments (e.g., Satyanarana and Ramana,1994; Calvert et al., 1995; Crusius et al., 1996; Nath et al., 1997; Passier et al., 1997; van derWeijden et al., 1998; Babu et al., 1999; Cowie et al., 1999; Keil and Cowie, 1999; Smallwoodand Wolff, 2000; Schulte et al., 2000; Suthhof et al., 2000). Organic geochemical studies havealso included determination of radiocarbon ages of individual organic compounds (Eglinton etal., 1997), and calibration of unsaturation indices in long-chain ketones (Uk’
37) in surficialsediments against measured sea-surface temperatures (SST; Sonzogni et al., 1997a, b) forsubsequent use in paleo-SST interpretations (e.g., Rostek et al., 1997).
Many of these geochemical characterisations and process rate determinations also will contribute to ongoing debates on the processes that control organic matter distributions in marine sediments and, specifically, on the factors that lead to the organic-rich sediments found on the continentalmargins of the Arabian Sea. The central issue is whether oxygen depletion is a primary,independent cause of organic matter enrichment. As examples, Pedersen et al. (1992) andCalvert et al. (1995) found no clear physical relationship between the depths of the sedimentaryorganic matter maximum and the mid-water oxygen minimum, and no consistent difference inbulk organic matter composition (C/N ratios, hydrocarbon content, δ13Corg) for surficial

77
sediments within and outside the OMZ, on the Oman and Indian margins, respectively. Theyconcluded that factors including winnowing and offshore changes in productivity, depth andsedimentation rate are responsible for mid-slope maxima in organic matter concentrations, along with sediment texture and dilution effects. Similar conclusions were drawn by Thamban et al.(1997), Rao and Veerayya (2000) and Babu et al. (1999), but also invoking an effect of bottomtopography (enhanced accumulation on marginal highs). In contrast (also as examples),Paropkari et al. (1992) concluded that the overall distribution of organic matter in Arabian Seasediments could only be explained by preferential preservation in the absence of oxygen as thedominant influence. More recent comparisons of the organic matter contents of surficialsediments (van der Weijden et al., 1998) and longer sediment records (e.g., Reichart, 1998) from the Pakistan margin and Murray Ridge also conclude that oxygen depletion is of primaryimportance, possibly because the OMZ is more intense in this area (undisturbed, varvedsediments within the OMZ, in contrast to bioturbation across the entire Oman margin).
However, more detailed organic geochemical studies suggest that the controls are complex.Comparisons with faunal abundances and bioturbation indicate that benthic macrofauna mayform a major control on the distributions of organic matter content and quality across the Oman margin (Smallwood et al., 1999; Smallwood and Wolff, 2000), not clearly linked with bottom-water oxygen concentrations. Other biochemical analyses also show compositional differencesin surface sediments across the Pakistan margin (e.g., Cowie et al., 1999; Suthhof et al., 2000;Schulte et al., 2000), but, in general, these are subtle and organic-rich sediments occurring under more oxidizing conditions below the strict OMZ indicate that factors other than oxygendepletion strongly influence mid-slope organic matter enrichments. If cross-margin differencesin organic matter content are due to differences in preservation, one of the more enigmaticresults of recent studies of surficial Arabian Sea sediments is that there is little indication ofdown-core sedimentary organic matter decay. Moreover, although net sulphate reduction isevident even in abyssal sediments, particularly at depth (e.g., Luckge et al., 1999; Bottcher et al.,2000), it is at most moderate and insufficient to significantly deplete porewater sulphateconcentrations within the surficial 50 cm, even in the core of the OMZ (Crusius et al., 1996;Passier et al., 1997; van der Weijden et al., 1998; Cowie et al., 1999) even in the core of theOMZ. This contrasts strongly with organic-rich littoral sediments and even with much lessorganic-rich sediments on other ocean margins with oxygenated bottom waters. It suggests onlymoderately reducing redox conditions, even on the Pakistan margin where the OMZ is mostpronounced, and that the organic matter present is unusually refractory and/or that post-oxicorganic matter decay is inhibited.
Biological investigations of surficial Arabian Sea sediments on several JGOFS (and other)programmes have included studies of the distributions of foraminifera (e.g., Naidu andMalmgren, 1995; Gooday et al., 1998; Jannink et al., 1998) and dinoflagellate cysts (Zonneveld, 1997) as a function of depth, redox conditions, productivity and other oceanographic parameters (with comparisons to distributions in other oceans). These provide important tools for paleo-environmental interpretations of nanofossil assemblages in Arabian Sea sediment records (e.g.,den Dulk et al., 1998). The UK JGOFS expedition and the recent German JGOFS programmehave included the most comprehensive investigation of benthic microbial and faunalabundances, diversity, community structure and activity carried out across the Oman margin and at abyssal sites across the Arabian Sea. Some of the results of these studies have recentlyappeared in dedicated volumes of Deep-Sea Research (Gage et al., 2000; Pfannkuche andLochte, 2000).
The UK JGOFS studies included further work on foraminiferan communities and theirrelationships to organic matter enrichments, oxygen availability and metazoan abundances(Gooday et al., 2000). In another paper, Gooday et al. (2000) reported on a new species of giant

78
protozoan from the Oman margin below the OMZ, which complements a previous report byLevin and Edesa (1997) of a first finding of mudball-building cirratulid polychaetes within theOMZ on the Oman margin. Also, Lamont and Gage (2000) examined morphological adaptations in polychaetes to hypoxia, while Cook et al. (2000) test its influence on nematode abundance,and Creasey et al. (2000) examine the population biology, parasite pressure and populationgenetics of galatheid crabs inhabiting the lower boundary of the OMZ. Meadows et al. (2000)and Murray et al. (2000) investigate relationships between bioturbation, sediment geochemistryand geotechnical parameters. Levin et al. (2000) reported on relationships betweenenvironmental conditions and macrofaunal assemblages and lifestyles across the OMZ at theOman margin, with a notable observation that abundance maxima occur well within the OMZ.Smith et al. (2000) examined the nature and extent of bioturbation across the OMZ, andSmallwood and Wolff (2000) reported on spatial and depth distributions of organic matterquality, and related these to distributions in biological abundances and activity. Biologicalstudies from the German JGOFS programme include reports on sedimentary and BBL microbial populations and activity (Boetius and Lochte, 2000; Boetius et al., 2000a, b), bioturbation rates(Turnewitsch et al; 2000), and distributions, abundances and diversity of foraminifera(Kurbjeweit et al., 2000), metazoan meio- (Sommer and Pfannkuche, 2000) and macrofauna(Witte, 2000), and benthopelagic nekton (Christiansen and Martin, 2000) in relation toregionally and seasonally differing particle fluxes.
Recommendations
The published articles outlined above provide a sampling of the wealth of information that willcome from the variety of ongoing studies on Arabian Sea benthic dynamics and processes,which are exceptional in nature and of great biogeochemical importance. Suggestions for futureresearch would include:• Systematic studies of the structure, function and trophic interactions of Arabian Sea benthic
communities (microbes to megafauna), and how these vary with region, with depth, and inrelation to the OMZ, and in response to seasonal variation in organic matter supply.
• Linked to these should be a comprehensive assessment of the nature and rates of sedimentary biogeochemical processes, and how these vary seasonally and in relation to the OMZ. Thiswould combine: detailed geochemical characterisation of the sediments (both solid-phase and
porewaters) and their redox status measurements of microbial process rates (both aerobic and anaerobic), characterisation
and quantification of biological mixing and irrigation processes, and an assessment ofthe importance of chemoautotrophic processes within the OMZ in situ determinations of the directions and magnitudes of benthic fluxes of oxygen,
nutrients, trace metals and dissolved organic matter.Together with a concerted modelling effort, these studies would combine to provide animportant and broadly relevant view of benthic biogeochemistry in the Arabian Sea. Specificadvances would include new information on:• the global significance of benthic bio-element fluxes to/from one of the planet’s largest
expanses of suboxic, organic-rich sediments• the mechanisms of C cycling, and differences in C burial efficiency, in contrasting redox
settings and, perhaps, a conclusive answer as to the primary controls on organic Cpreservation and distribution
• the environmental factors that control benthic community structure and function, and howthe benthic community contributes to C cycling and burial.

79
2.11 Modelling Physical-Biogeochemical Interactions
Jerry Wiggert
Earth System Science Interdisciplinary CenterUniversity of Maryland
College Park, MD 20742-2465USA
Introduction
The various Arabian Sea JGOFS field campaigns that took place from 1990 to 1997 haveprovided a significant expansion of the observational database for this region (van Weering etal., 1997; Smith et al., 1998b; Burkill, 1999). The ongoing analysis of these data has led to aseminal advancement in our understanding of the physical and biogeochemical processes atwork in this complex region (Smith, 2001 and references within). The in situ biogeochemicalobservations collected over this time period provide a comprehensive sampling of theconstituents of the primary and secondary ecosystem (Landry et al., 1998; Smith et al., 1998a;Ducklow et al., 2001), along with distributions of chemical species and fluxes of particulatematter (Lee et al., 1998; Morrison et al., 1998; Honjo et al., 1999). Measurements of the region's physical characteristics have provided new insights into how the ocean's dynamical response tothe monsoon's annual cycle of meteorological forcing impacts biogeochemical processes(Dickey et al., 1998; Weller et al., 1998; Lee et al., 2000). These physical and biogeochemicaldata sets have prompted several modelling studies that have already appeared in the recentliterature (Wiggert et al., 2000; McCreary et al., 2001). These data, as both solution validationand as a basis for ecosystem model parameter optimisation (Ryabchenko et al., 1998; Fashamand Evans, 2000), are also an integral component of ongoing research employing coupledphysical-biological models that cover the full range of spatial domains (i.e., 1-D to Arabian Sea to Indian Ocean) as part of the U.S. JGOFS Synthesis and Modelling Project (SMP). The SW(summer) Monsoon has received significant attention in modelling studies in the past, due to the elevated primary production that occurs during this period (Bauer et al., 1991; Brock andMcClain, 1992; Keen et al., 1997). More recently, it has become apparent that primaryproductivity during the NE (winter) Monsoon is also relatively high and plays a prominent rolein defining the ecosystem’s annual cycle (Smith et al., 1998; Barber et al., 2001). A shortsummary of the pre-JGOFS paradigm, followed by a synopsis of the fresh insights garneredfrom modelling investigations stimulated by the recent observations, is presented for eachmonsoon period. This is followed by some closing remarks, which include some suggested focus areas for future analyses that would benefit from the JGOFS observations collected over the last decade.
The Southwest Monsoon
Oceanic response to large-magnitude wind forcing dominates the circulation pattern of theArabian Sea during the SW Monsoon (Schott and McCreary, 2001). The so-called Findlater Jethugs the coast of Somalia before heading offshore as strong southwesterlies that parallel theArabian Peninsula (Shetye et al., 1994). Notable current features resulting from the annual onset of the SW Monsoon winds include the Great Whirl off of the Somali coast and the offshoreadvection of waters upwelled along the southern coast of Oman (Schott, 1993; Flagg and Kim,1998). The synoptic distributions of surface chlorophyll a first revealed by the Coastal ZoneColor Scanner (CZCS) stimulated a number of modelling studies that looked to identify thespecific monsoon-driven physical processes that caused these prominent SW Monsoon

80
phytoplankton blooms. Clearly, upwelling along the Omani coast plays a major role during thisperiod (Brock and McClain, 1992; Kindle, this volume). Though hampered by excessivecloudiness, the CZCS observations also indicated that elevated chlorophyll a concentrationsextended offshore at least an order of magnitude beyond the internal Rossby radius of O(50 km) (Brock et al., 1991). A region of persistent, large-magnitude Ekman pumping, resulting from thehorizontal distribution of the monsoon winds, extends ~ 450 km offshore of the ArabianPeninsula during the SW Monsoon (Brock and McClain, 1992). This led to a debate in theliterature as to whether Ekman pumping (Brock et al., 1991; Bauer et al., 1992) or a combination of horizontal advection and wind-forced vertical entrainment (Young and Kindle, 1994;McCreary et al., 1996; Keen et al., 1997) leads to the nutrient enrichment that supports theelevated, offshore phytoplankton biomass.
High levels of eddy kinetic energy, and current filaments that transported distinct physical andbio-optical properties more than 700 km offshore, were observed during the JGOFS cruises(Flagg and Kim, 1998; Lee et al., 2000). In addition, HPLC (high-performance liquidchromatography) observations and primary productivity measurements obtained by JGOFSresearchers during the 1995 SW Monsoon extended the CZCS-suggested offshore distributionpattern by revealing elevated chlorophyll a concentrations and phytoplankton growth rates atleast 1000 km offshore (Latasa and Bidigare, 1998; Barber et al., 2001). These offshorephytoplankton distributions may be linked to the eastward advection of upwelled waters withinthe Somali Current (Young and Kindle, 1994; McCreary et al., 1996; Murtugudde et al., 1999).Furthermore, new results from a U.S. JGOFS SMP-funded modelling study demonstrate that aseries of detrainment/entrainment phytoplankton blooms result from a shoaling/deepening mixed layer responding to high-frequency wind forcing observed by a meteorological buoy locatedalong the climatological axis of the Findlater Jet (R. Hood, pers. comm.). The JGOFSobservations and the noted model study belie the impact of Ekman pumping as the mechanismthat defines the distribution of the SW Monsoon phytoplankton bloom, since these results arefrom areas well offshore of the region of positive wind curl (Manghnani et al., 1998).
The Northeast Monsoon
The cool, dry northeasterly winds that characterize the NE Monsoon result in a typicalwintertime convection/nutrient enrichment scenario in the northern Arabian Sea (Banse andMcClain, 1986; Madhupratap et al., 1996). Evaporative cooling increases the salinity of thesurface waters, which further promotes the convective mixing. However, this convection isinhibited during the day by solar heating of the surface waters. These competing surface heatfluxes result in a persistent, large-magnitude diurnal oscillation in mixed layer depth (e.g.,Wiggert et al., 2000). In comparison to previously reported values, surprisingly high rates ofprimary productivity were measured by JGOFS researchers in concert with this convectivemixing (Barber et al., 2001; Smith, 2001) and an accumulation of phytoplankton biomass wasapparent over the course of the NE Monsoon (Wiggert et al., 2000). Values of the f-ratiodetermined by nutrient assimilation experiments during the NE Monsoon showed thatphytoplankton growth was primarily supported by ammonium uptake (McCarthy et al., 1999;Watts and Owens, 1999). Indeed, McCarthy et al. (1999) note that rates of primary productivityincreased 2-fold while nitrate slowly accumulated in the surface layer. These observationsindicate that some physical and/or biological control limits the full utilization of availablenutrients, which runs counter to the previously tendered supposition that variations in wintertime nitrate entrainment lead to the interannual variability noted in distributions of surfacechlorophyll a concentrations (Banse and McClain, 1986; Banse and English, 1993).
Results from the JGOFS observations suggest that this incomplete utilization of nutrients is aresult of the combined impact of zooplankton grazing and the mixed layer's diurnal cycling.

81
Microzooplankton grazing accounted for 86% of phytoplankton growth (Landry et al., 1998).Furthermore, estimated rates of fecal pellet production by mesozooplankton were significantlyhigher than the observed rates of export production (Roman et al., 2000). These studies suggestsignificant grazing control and recycling of biogenic material within the upper layer, which isconsistent with the f-ratios determined in the nutrient cycling studies already noted. The diurnalmixing homogenizes the surface layer's physical and biogeochemical properties down to theseasonal thermocline, nominally on a daily basis (Gardner et al., 1999). This restrains theaccumulation of phytoplankton biomass in the euphotic zone and provides a continual supply of plant material subject to remineralisation in the sub-euphotic zone (Wiggert et al., 2000;McCreary et al., 2001). By incorporating the diurnal irradiance cycle within their coupled bio-physical model, McCreary et al., (2001) introduced a diurnal mixed layer which generatedphytoplankton blooms that exhibited more realistic magnitude, timing, and temporal extent thanobserved in a previous solution (McCreary et al., 1996). Through the further inclusion of dailyforcing, the best correspondence to a phytoplankton production time-series (Marra et al., 1998)was achieved. In another modelling study focused on diurnal mixing, Wiggert et al., (2000)suggested interannual variations in the depth of the permanent mixing barrier (i.e., the seasonalthermocline), in conjunction with a diurnally cycling mixed layer, could account for theinterannual variability in surface chlorophyll a distribution noted previously.
This hypothesis has been tested with notable success (Wiggert et al., 2001). The analysis madeuse of the first three years of NE Monsoon-period ocean colour distributions observed bySeaWiFS (Sea-viewing Wide Field of view Sensor) and the corresponding thermocline depthsextracted from an interannually forced ocean general circulation model (OGCM) solution(Murtugudde and Busalacchi, 1999). SeaWiFS ocean colour distributions (8-day composites)from the same period for three successive NE Monsoons are shown (Figure 1). These revealsignificantly lower pigment concentrations during the 97/98 NE Monsoon. The boxed regiondepicted here defines the area over which spatial averages were made for the subsequentanalysis.
The time-series (of the spatial mean within the boxed region) of thermocline depth for the three NE Monsoon periods is shown (Figure 2a), revealing that for the 97/98 NE Monsoon, theOGCM-predicted thermocline was consistently at least 2σ below the 35-year mean. Thedistribution of chlorophyll a concentration as a function of thermocline depth for the fullsequence of SeaWiFS 8-day composite imagery for the three NE Monsoon periods is shown(Figure 2b). The two solid lines represent the surface chlorophyll a concentration (at noon anddusk) as predicted by a simple, 1-D growth/mixing scheme (complete details are provided inWiggert et al., 2001). This result suggests that the convolution of a diurnally cycling mixed layer and an interannually varying thermocline depth results in the observed interannual variation inpigment concentration.
Closing Remarks and Suggestions for Future Modelling Efforts
The extensive in situ observational database that is now available as a result of the JGOFSactivities in the Arabian Sea has already altered our understanding of the spatial extent of theSW Monsoon’s phytoplankton bloom and the magnitude of primary production during the NEMonsoon. These data are certain to provide additional new insights for years to come. Themodelling studies that have so far resulted from the JGOFS expeditions in the Arabian Seaconsistently demonstrate that including the full spectrum of physical variability, especially highfrequency processes, is vital to developing an accurate simulation of the biogeochemical system(Wiggert et al., 2000; McCreary et al., 2001). Modelling efforts aimed at achieving insight intothe impact of mesoscale current structures on the region's biogeochemical processes would also

82
be timely, given the eddy and filament structures that manifest during the SW Monsoon (Kindle, this volume).
The synthesis of the JGOFS observations with an eye toward providing the modellingcommunity with conceptualisations that lead to more robust representations of ecosystembehaviour without overwhelming computational resources will be an ongoing challenge thatshould be vigorously pursued. For example, the food web structures developed in Garrison et al.(2000), which evolved from the analysis of Campbell et al. (1998), provide genuine insight intothe seasonal/spatial variations/transitions among, and between, various ecosystem components.The conceptual food webs developed in these analyses will be invaluable for assessing how well ecosystem models can reproduce the observed spatial and temporal structures. Furthermore, such conceptualisations will help illuminate the present shortcomings of the various ecosystemmodels as well as aid the continuing exploration for model forms that best capture the fulldynamic range of the natural variability (e.g., Yajnik and Sharada, 1992; Sharada and Yajnik,1997; Fasham and Evans, 2000).
Figure 1. Surface chlorophyll a distribution for 11-18 December is shown for three NEMonsoon periods observed by SeaWiFS (97/98, 98/99, 99/00). We have defined a focus region(15-18°N, 60-65°E) away from coastal influences. This is superimposed on each image, and aids in making consistent comparisons between images. During 97/98, pigment concentrations range from 0.3 to 0.6 mg Chl a m-3. During 98/99 and 99/00, the typical range is 0.7 to 1.0 mg Chl am-3, with values ranging up to 2.0 mg Chl a m-3.

83
Figure 2(a). The individual time-series of 21°C-isotherm depth for the three years of SeaWiFScoverage are shown. The 35-year mean is also shown, with (± s) depicted by the grey shadedregion. This figure indicates that the thermocline during the 97/98 NE Monsoon is consistentlyat least 2σ below the mean. (b). Chlorophyll a versus thermocline depth. The blue and browncurves are chlorophyll a at noon and sundown, respectively, from the 1-D model. The SeaWiFSand CZCS chlorophyll a observations, matched up with their corresponding OGCM-predictedisotherm depths, have been superimposed. The dashed black line is a linear fit to the SeaWiFSdata. The SeaWiFS data and the model result show an excellent correspondence.
A similar distillation of the observations of biogeochemical processes associated with theArabian Sea’s extensive oxygen minimum zone (OMZ) would facilitate the incorporation of this feature within future ecosystem modelling efforts. In addition to a full chapter devoted todescribing the OMZ's impact on biogeochemical cycling (Smith et al., this volume), no fewerthan four of the other chapters presented herein call for further analysis of, and insight into, itssignificance in this region. The hydrographic surveys performed during the U.S. JGOFS cruiseshave resulted in a new appreciation of the extent of the OMZ (Morrison et al., 1999).Furthermore, it has become apparent that the OMZ significantly reduces the diel verticalmigrations of the zooplankton community (Smith et al., 1998; Wishner et al., 1998). Sincezooplankton significantly modify carbon fluxes through grazing, excretion, repackaging andvertical motility, their absence from the mid-depths of the water column could have profoundeffects on regional carbon cycling (Wishner et al., 1998). These processes need to be betterunderstood so enhancements to carbon cycle modelling efforts (e.g., Swathi et al., 2000) can be

84
implemented, which should, in turn, further our understanding of whether the Arabian Sea is anet source or sink of atmospheric CO2 (Lal, 1994; Smith, 2001). It is also worth noting thatdenitrification within the OMZ produces the nitrous oxide (N2O) that subsequently effluxes tothe atmosphere at an estimated rate of 0.4 Tg N2O y-1 (Bange et al., 2000). This rate does notinclude the significant contribution made by waters from the West Indian continental shelf(Naqvi, this volume). Nitrous oxide is a potent greenhouse gas that merits attention in futureclimate studies since denitrification rates appear to be increasing, which may be indicative of aglobal warming-monsoon intensification feedback scenario (Naqvi, et al., 2000).
Finally, the anoxic conditions of the OMZ result in greater iron solubility and higherconcentrations of bio-available iron below the ocean's surface layer (Measures and Vink, 1999;Witter et al., 2000). Iron is a requirement for phytoplankton photosynthesis (Martin et al., 1991)and the surface waters of the Arabian Sea are characterized by non-limiting iron concentrationsthroughout the year (Measures and Vink, 1999). While significant aeolian iron deposition isdocumented (Tindale and Pease, 1999), questions remain as to whether this deposition issufficient to maintain the observed surface iron concentrations or if entrainment/upwelling ofiron-rich OMZ waters must also be taken into account. Applying models that include explicitiron biogeochemistry (e.g., Christian et al., 2001) to the Arabian Sea would help characterize the OMZ's possible role as a source of iron to the euphotic zone.
Acknowledgements
The author gratefully acknowledges the support of the NASA Oceanography Program (NAG58595). This chapter has benefited significantly from the comments of (in alphabetical order)Raleigh Hood, John Kindle and Ragu Murtugudde. I would also like to thank Sharon Smith forinviting this contribution and Louisa Watts for her tireless assistance, which was very muchappreciated.

85
3. MAJOR ACHIEVEMENTS, RECOMMENDATIONS AND FUTURE TIME-LINE
3.1 Major Achievements
At the time that the JGOFS Arabian Sea studies were organized, first-order issues such aswhether or not the Arabian Sea was a sink or a source for atmospheric carbon dioxide were stillunclear. Did high rates of primary productivity and large concentrations of sedimentary carbonmake it a sink or were these processes overwhelmed by outgassing of carbon dioxide arisingfrom the warming of deeper waters reaching the sea surface via upwelling and mixing? The wide range of climatic variability revealed in the Arabian Sea’s sedimentary record, the inter-annualvariability in the strength of monsoons that has been documented for the recent past, andsuggestions that the mean strength of the monsoon would change under plausible scenarios ofclimate change, all suggested that the Arabian Sea is an excellent environment in which to study biogeochemical cycling and climate change. Since the goals of JGOFS are (1) to determine andunderstand on a global scale the processes controlling the time-varying fluxes of carbon andassociated biogenic elements in the ocean and to evaluate the related exchanges with theatmosphere, sea floor and continental boundaries, and (2) to develop a capability to predict on a global scale the response of oceanic biogeochemical processes to anthropogenic perturbations, in particular those relating to climate change (SCOR, 1990), it was possible to make a compellingcase that JGOFS should support a major international research effort in the Arabian Sea. TheJGOFS studies were designed to provide a seasonally and spatially resolved carbon budget for a basin exhibiting some of the highest and lowest concentrations of plant biomass observedanywhere in the world’s ocean. To date, disciplinary papers have given us extraordinary newunderstanding of the components of the carbon cycle in the Arabian Sea, and these have beenconstrained by emerging interdisciplinary studies and modelling. Some of our new knowledge is summarized below.
Understanding the Physical Forcing
• Unlike currents depicted from ship-drift data, currents in the upper 400 meters did not follow the winds
• The velocity field was dominated by eddies• The only organized flow was the coastal current and Ras al Hadd Jet• Heat was lost in December/January and gained in August• Highest rates of heating occurred in the Intermonsoon seasons• Upwelled water in the SW Monsoon season is geostrophically advected offshore in filaments
due towind stress curl and sea level height
• Deepest mixed layers occur in the NE Monsoon season• Upwelling comes from as deep as 200 meters• In the NE Monsoon, surface cooling intensified and mixed layer deepens dramatically with
distanceoffshore due to convective overturning
• In the SW Monsoon, cooling offshore is due to filaments and wind driven entrainment andnot open
ocean upwelling (Ekman pumping)• Eddy kinetic energy is high in the west and southwest of the Arabian Sea, but much reduced
in theeastern and northern Arabian Sea

86
Understanding the Biogeochemical Response
• Flux of organic material to the seabed is dominated by an event at the end of the SWMonsoon
• Areally and annually the Arabian Sea is a slight sink for CO2 from the atmosphere• Primary production in the SW Monsoon is equal to that during the NE Monsoon• Iron is delivered from the atmosphere to the sea surface during SW Monsoon and exceeds
phytoplankton needs• Microbial loop is balanced except during the SW Monsoon• Diatom phytoplankton blooms develop late during the SW Monsoon• Substantial biomass inhabits the OMZ and vertically migrates• Zooplankton biomass is high all year round• A deep, active Prochlorococcus spp. layer exists during the NE Monsoon• Complex seasonal variability: TOC is lowest in the fall and highest in July, but bacteria
exhibit littleseasonality
• Mesozooplankton seem to exert top-down control of the microbial loop during the SWMonsoon
• The spatial area of surface chlorophyll, as observed by remotely-sensed ocean colour(CZCS), is
larger than that suggested by observations (possible effects of dust)• Both benthos and pelagos respond similarly to the OMZ with decreased diversity and
increasedbiomass of those few taxa able to exploit suboxic conditions
• The oceanic nitrogen cycle is far from steady-state; denitrification is increasing• Nutrient ratios suggest nitrogen is limiting and denitrification dominates nitrogen fixation• Mixed layer is deeper in the NE Monsoon than in the SW Monsoon
3.2 Recommendations
1) The JGOFS emphasis on basin-scale carbon budgets forced observational schemes that didnot allow the study of mesoscale spatial and temporal variability in the Arabian Sea. It istherefore recommended that investigations be made in the following areas:
• Coastal upwelling: physiology of phytoplankton (spin up) and washout• Scales of advection and ecological processes• Source and physiology of diapause zooplankton• Top-down control of the microbial loop in upwelling areas• Taxonomy and the microbial loop• The role of filaments in biogeochemistry• The role of eddies in biogeochemistry• Mechanisms generating filaments• Sediments as a source of Fe for phytoplankton• Biogeochemical responses• Role of larger predators such as crabs, myctophids, squid

87
2) The JGOFS emphasis on carbon cycling forced observational strategies that minimised thestudy of the nitrogen cycle and OMZ of the Arabian Sea. Therefore, investigations into thefollowing areas are recommended:
• Nitrogen cycle of the OMZ• Sediment biogeochemistry; denitrification• Chemical transformations• Physiological adaptation of all organisms in the OMZ
3) Understanding processes in the Arabian Sea would benefit from a comprehensive approach(models + data assimilation + remote sensing) to integrated problems. In this respect thefollowing integrations should be highlighted for future research:
• Sedimentation flux, surface forcing, and carbon burial rates• Diapause zooplankton, phytoplankton blooms and physical forcing• Potential linkage of Oman and Somalia upwellings• Aerosols, delivery of micronutrients (Fe), phytoplankton blooms, surface forcing• Onset of SW Monsoon• Food web structure adjustments during upwelling and convection
4) Monitoring of the OMZ should be made over the long term, as it is a sensitive barometer ofclimate change
5) Rigorous floristic and faunistic comparisons between the 1960’s and 1990’s should be madewith respect to species shifts, as these provide evidence for ecosystem adjustment to climatechange
3.3 Future Time-line and Research
Synthesis and modelling of the JGOFS data sets generated from the Arabian Sea campaign willcontinue until 2003. Major milestones envisaged are as follows:
2002In February, Indian Ocean Synthesis Group (IOSG) committee members (Smith, Naqvi andBurkill) will host a Special Session on Arabian Sea Synthesis at the ASLO meeting in Hawaii.Those presenting papers at this open meeting will be encouraged to focus on internationaldisciplinary syntheses. If there is sufficient interest, we may consider a further Special Issue ofDeep-Sea Research with an intended publication date of spring 2003.
2003We intend one or more multidisciplinary syntheses to be organised by the Indian OceanSynthesis Group (IOSG) for presentation at the Final JGOFS Open Science Conference. This isbeing organised for Washington, USA in April 2003. In addition, there may be a variablenumber of other syntheses, probably with a disciplinary focus. Publication would depend on the Symposium Organising Committee.

88
4. APPENDICES
APPENDIX 4.1
Membership of JGOFS Indian Ocean Synthesis Group (July 2001)
Professor Sharon Smith (Chair), University of Miami, The Rosenstiel School (RSMAS),Division of Marine Biology and Fisheries, 4600 Rickenbacker Causeway, Miami, Florida33149-1098, USA. [email protected]
Dr. Shahid Amjad, National Institute of Oceanography, ST-47, Block 1 Clifton, 75600 Karachi, PAKISTAN. [email protected]
Dr. Martien Baars, Netherlands Institute of Sea Research (NIOZ), P.O. Box 59, 1790 AB Den Burg, Texel, THE NETHERLANDS. [email protected]
Professor Karl Banse, University of Washington, Dept. of Oceanography, Box 355351,Seattle, Washington 98195-5351, USA. [email protected]
Professor Peter Burkill, Plymouth Marine Laboratory, Natural Environment Research Council, Prospect Place, West Hoe, Plymouth, PL1 3DH, UNITED KINGDOM. [email protected]
Dr. John Kindle, Oceanography Division, Naval Research Laboratory, Stennis Space Center,MS 39529-5004, USA. [email protected]
Dr. Wajih Naqvi, National Institute of Oceanography, Dona Paula, Goa 403 004, [email protected]
Dr. Tim Rixen, Zentrum für Marine Tropenökologie, Universität Bremen, Fahrenheitstraße 6,D-28359 Bremen, GERMANY. [email protected]
Dr. Shubha Sathyendranath (Vice Chair), Bedford Institute of Oceanography, BiologicalOceanography Section, P.O. Box 1006, Dartmouth, Nova Scotia, B2Y 4A2, [email protected]
Dr. Kirit S. Yajnik, Centre for Mathematical Modelling and Computer Simulation(CMMACS), Council of Scientific and Industrial Research (CSIR), F2, Regal Manor, 2/1 BrideSt., Belur Campus, Longford Town, Bangalore 560037, INDIA. [email protected]

89
APPENDIX 4.2
Reference List from Section 2
Abbott M.R. and Barksdale B. (1995) Variability in upwelling systems as observed by satelliteremote sensing. In: Upwelling in the Ocean: Modern Processes and Ancient Records, C.P.Summerhayes, K.C. Emeis, M.V. Angel, R.L. Smith and B. Zeitzschel (Eds.) p. 221-238. John Wiley and Sons, New York.
Aiken J., Moore G.F., Trees C.C., Hooker S.B., and Clark D.K. (1995) The SeaWiFS CZCS-type pigment algorithm. In: S.B. Hooker and E.R. Firestone (Eds.), NASA Technical Memo104566, vol. 29, SeaWiFS Technical Report Series, 34pp.
Altabet M.A., Francois R., Murray D.W., and Prell W.L. (1995) Climate-related variations indenitrification in the Arabian Sea from sediment N15/N14 ratios. Nature, 373(6514): 506-509.
Andruleit H.A., von Rad U., Bruns A., and Ittekkot V. (2000) Coccolithophore fluxes fromsediment traps in the northeastern Arabian Sea off Pakistan. Marine Micropaleontology, 38:285-308.
Antoine D., André J.M., and Morel A. (1996) Oceanic primary production 2. Estimation atglobal scale from satellite (coastal zone colour scanner) chlorophyll. Global BiogeochemicalCycles, 10: 57-69.
Arnone R.A., Martinolich P., Kindle J., Brink K.H., and Lee C.M. (2001) Characteristics of thecoastal filaments along the Oman coast. Journal Geophysical Research, submitted.
Ashjian C.J., Smith S.L., Flagg C.N. and Idrisi N. (2002) Distribution, annual cycle, and vertical migration of acoustically derived biomass in the Arabian Sea during 1994-1995. Deep-SeaResearch II, in press.
Austin R.W. and Pretzold T.J. (1981) The determination of the diffuse attenuation coefficient of seawater using the Coastal Zone Colour Scanner. In: Gower J.F.R. (Ed.), Oceanography fromSpace. Plenum Press, New York, p. 239-256.
Azam F., Steward G.F., Smith D.C., and Ducklow H.W. (1994) Significance of Bacteria inCarbon Fluxes in the Arabian Sea. In: D. Lal (Ed.), Biogeochemistry of the Arabian Sea,Proceedings of the Indian Academy of Sciences, Earth and Planetary Sciences, 103(2): 341-351.
Baars M. (1999) On the paradox of high mesozooplankton biomass, throughout the year in thewestern Arabian Sea: Re-analysis of IIOE data and comparison with newer data. Indian Journal of Marine Sciences, 28: 125-137.
Baars M. and Brummer G.J. (1995) Workshop reviews results from Netherlands Indian OceanProgramme. U.S. JGOFS News, 6: 10-11.
Baars M. and Oosterhuis S. (1998) Zooplankton biomass in the upper 200 m in and outside theseasonal upwelling areas of the western Arabian Sea. In: A. Pierrot-Bults and S. van der Spoel (Eds.), Pelagic Biogeography ICoPBII. Proceedings of the Second International Conference.IOC/UNESCO, Paris, p. 39-52.
Babu C.P., Brumsack H.J., Schnetger B. (1999) Distribution of organic carbon in surfacesediments along the eastern Arabian Sea: a revisit. Marine Geology, 162: 91-103.
Bak R.P.M. and Nieuwland G. (1997) Seasonal variation in bacterial and flagellate communities of deep-sea sediments in a monsoonal upwelling system. Deep-Sea Research II, 44: 1281-1292.

90
Baker S., Arnone R., and Sheres D. (1996) Characteristics of double vortices in the northeastern Arabian Sea and the North Atlantic Gulf Stream region. EOS, 77: 382.
Bakun A., Roy C., and Lluch-Cota S. (1998) Coastal upwelling and other processes regulatingecosystem productivity and fish production in the western Indian Ocean. In: K. Sherman, E.Okemwa and M. Ntiba (Eds.), Large Marine Ecosystems of the Indian Ocean: assessment,sustainability and management. Blackwell Science, Malden, MA, pp.103-141.
Bange H.W., Bartell U.H., Rapsomanikis S., and Andreae M.O. (1994) Methane in the BalticSea and North Sea and a reassessment of the marine emissions of methane. GlobalBiogeochemical Cycles, 8: 465-480.
Bange H.W., Rapsomanikis S., and Andreae M.O. (1996) Nitrous oxide emissions from theArabian Sea. Geophysical Research Letters, 23: 3175-3178.
Bange H.W., Ramesh R., Rapsomanikis S., and Andreae M.O. (1998) Methane in surface waters of the Arabian Sea. Geophysical Research Letters, 25: 3547-3550.
Bange H.W., Rixen T., Johansen A.M., Siefert R.L, Ramesh R., Ittekkot V., Hoffmann M.R. and Andreae M.O. (2000) A revised nitrogen budget for the Arabian Sea. Global BiogeochemicalCycles, 14: 1283-1297.
Bange H.W., Andreae M.O., Lal S., Law C.S., Naqvi S.W.A., Patra P.K., Rixen T., and Upstill-Goddard R.C. Nitrous oxide emissions from the Arabian Sea: A synthesis. GlobalBiogeochemical Cycles, submitted.
Banse K. (1959) On upwelling and bottom-trawling off the southwest coast of India. Journal ofthe Marine Biological Association of India, 1: 33-49.
Banse K. and McClain C.R. (1986) Winter blooms of phytoplankton in the Arabian Sea asobserved by the Coastal Zone Colour Scanner. Marine Ecology Progress Series, 34: 201- 211.
Banse K. (1987) Seasonality of phytoplankton chlorophyll in the central and northern ArabianSea. Deep-Sea Research, 34: 713-723.
Banse K. and English D.C. (1993) Revision of satellite-based phytoplankton pigment data fromthe Arabian Sea during the Northeast Monsoon. Marine Research (Pakistan), 2: 83-103.
Banse K. and English D.C. (2000) Geographical differences in seasonality of CZCS-derivedphytoplankton pigment in the Arabian Sea for 1978-1986. Deep-Sea Research, 47: 1623-1677.
Barber R.T., Marra J., Bidigare R.R., Codispoti L.A., Halpern D., Johnson Z., Latasa M.,Goericke R., and Smith S.L. (2001) Primary productivity and its regulation in the Arabian Sea during 1995. Deep-Sea Research II, 48: 1127-1172.
Barlow R.G., Mantoura R.F.C., and Cummings D.G. (1999) Monsoonal influence on thedistribution of phytoplankton pigments in the Arabian Sea. Deep Sea Research II, 46: 677-699.
Bauer S., Hitchcock G.L., and Olson D.B. (1991) Influence of monsoonally forced Ekmandynamics upon surface layer depth and plankton biomass distribution in the Arabian Sea.Deep-Sea Research, 38: 531-553.
Bauer S., Hitchcock G.L., and Olson D.B. (1992) Response of the Arabian Sea surface layer tomonsoon forcing. In: B.N. Desai (Ed.) Oceanography of the Indian Ocean. Oxford and IBHPublishing Co. Ltd., New Delhi, p. 659-672.
Beal L.M., Field A., and Gordon A.L. (2000a) Spreading of Red Sea overflow waters in theIndian Ocean. Journal Geophysical Research, 105: 8549-8564.
Beal L.M., Molinari R.L., Robbins P.E., and Chereskin R.K. (2000b) Reversing bottomcirculation in the Somali Basin. Geophysical Research Letters, 27: 2565-2568.

91
Behrenfeld M.J. and Falkowski P.G. (1997) Photosynthetic rates derived from satellite-basedchlorophyll concentrations. Limnology and Oceanography, 42: 1-20.
Berger W.H., Fisher K., Lai C., and Wu G. (1988) Ocean carbon flux: Global maps of primaryproduction and export production. In: C. Agegian (Ed.), Biogeochemical Cycling and FluxesBetween the Deep Euphotic Zone and Other Oceanic Realms. NOAA Symposium Series onUndersea Research, NOAA Undersea Research Program 3/2: 131-176.
Betzer P.B., Showers W.J., Laws E.A., Winn C.D., DiTullio G.R., and Kroopnick P.M. (1984)Primary productivity and particle fluxes on a transect of the equator at 153°W in the PacificOcean. Deep-Sea Research, 31: 1-11.
Binet, D. and Suisse de Sainte Claire E. (1975) Le copépode planctonique Calanoides carinatusrépartition et cycle biologique au large de la Côte d’Ivoire. Cahiers O.R.S.T.O.M. SérieOceéanographique, 13: 15-30.
Boetius A. and Lochte K. (2000) Regional variations of total microbial biomass in sediments ofthe deep Arabian Sea. Deep-Sea Research II, 47: 149-168.
Boetius A., Springer B., and Petry C. (2000a) Microbial activity and particulate matter in thebenthic nepheloid layer (BNL) of the deep Arabian Sea. Deep-Sea Research II, 47(14): 2687-2706.
Boetius A., Ferdelman T., and Lochte K. (2000b) Bacterial activity in sediments of the deepArabian Sea in relation to vertical flux. Deep-Sea Research II, 47(14): 2835-2875.
Böhm E., Morrison J.M., Manghnani V. Kim H.S., and Flagg C.N. (1999) The Ras al Hadd Jet:Remotely sensed and acoustic Doppler current profiler observations in 1994-1995. Deep SeaResearch II, 46: 1531-1550.
Borchers, P. and Hutchings L. (1986) Starvation tolerance, development time and eggproduction of Calanoides carinatus in the southern Benguela Current. Journal of PlanktonResearch, 8: 855-874.
Bottcher M.E., Schale H., Schnetger B., Wallmann K., and Brumsack H.J. (2000) Stable sulfurisotopes indicate net sulfate reduction in near-surface sediments of the deep Arabian Sea.Deep-Sea Research II, 47: 2769-2784.
Brandes J.A., Devol A.H., Jayakumar D.A., Yoshinari T., and Naqvi S.W.A. (1998) Isotopiccomposition of nitrate in the central Arabian Sea and eastern tropical North Pacific: a tracer for mixing and nitrogen cycles. Limnology and Oceanography, 43: 1680-1689.
Bricaud A., Babin M., Morel A., and Claustre H. (1995) Variability in the chlorophyll-specificabsorption coefficients of natural phytoplankton: analysis and parameterization. JournalGeophysical Research, 100: 13,321-13,332.
Brink K., Arnone R., Coble P., Flagg C., Jones B., Kindle J., Lee C., Phinney D., Wood M.,Yentsch C., and Young D. (1998) Monsoons boost biological productivity in the Arabian Sea.EOS, Transactions, American Geophysical Union, 79: 168-169.
Brock J.C., McClain C.R., Luther M.E., and Hay W.W. (1991) The phytoplankton bloom in thenorthwest Arabian Sea during the southwest monsoon of 1979. Journal of GeophysicalResearch, 96: 20,623-20,642.
Brock J.C. and McClain C.R. (1992) Interannual variability in phytoplankton blooms observedin the Northwestern Arabian Sea during the Southwest Monsoon. Journal GeophysicalResearch, 97: 733-750.
Brock J., Sathyendranath S., and Platt T. (1993) Modelling the seasonality of subsurface lightand primary production in the Arabian Sea. Marine Ecology Progress Series, 101: 209-221.

92
Brock J.C., Sathyendranath S., and Platt T. (1998) Biohydro-optical classification of thenorthwestern Indian Ocean. Marine Ecology Progress Series, 165: 1-15.
Broerse A.T.C., Brummer G.J.A., and Van Hinte J.E. (2000) Coccolithophore export production in response to monsoonal upwelling off Somalia (northwestern Indian Ocean). Deep-SeaResearch, 47: 2179-2205.
Brown S.L., Landry M.R., Barber R.T., Campbell L., Garrison D.L., and Gowing M.M. (1999)Picophytoplankton dynamics and production in the Arabian Sea during the 1995 southwestmonsoon. Deep-Sea Research II, 46: 1745-1768.
Buesseler K.O. (1998) The decoupling of production and particulate export in the surface ocean. Global Biogeochemical Cycles, 12: 297-310.
Buesseler K.O., Ball L., Andrews J., Benitez-Nelson C., Belastock R., Chai F., and Chao Y.(1998) Upper ocean export of particulate organic carbon in the Arabian Sea derived fromthorium-234. Deep-Sea Research I, 45: 2461-2487.
Burkill P.H., Leakey R.J.G., Owens N.J.P., and Mantoura R. (1993) Synechococcus and itsimportance to the microbial foodweb of the northwestern Indian Ocean. Deep-Sea Research II, 40: 773-782.
Burkill P.H. (1999) ARABESQUE: An overview. Deep-Sea Research II, 46: 529-547.
Burkill P.H. (2002) Microbial dynamics. In: L. Watts, P. Burkill and S. Smith (Eds.), ArabianSea Process Study. JGOFS International Planning Office, Bergen, Norway, this volume.
Calvert S.E., Pedersen T.F., Naidu P.D., and von Stackelberg U. (1995) On the organic carbonmaximum on the continental slope of the eastern Arabian Sea. Journal Marine Research, 53:269-296.
Campbell L., Landry M.R., Constantinou J., Nolla H.A., Brown S.L., Liu H., and Caron D.A.(1998) Response of microbial community structure to environmental forcing in the ArabianSea. Deep Sea Research II, 45: 2301-2325.
Capone D.G., Subramaniam A., Montoya J.P., Voss M., Humborg C., Johansen A.M., SiefertR.L., and Carpenter E.J. (1998) An extensive bloom of the N2 – fixing cyanobacteriumTrichodesmium erythraeum in the central Arabian Sea. Marine Ecology Progress Series, 172:281-292.
Caron D.A. and Dennett M.R. (1999) Phytoplankton growth and mortality during the 1995northeast monsoon and spring intermonsoon in the Arabian Sea. Deep-Sea Research Part II,46: 1665-1690.
Chandramohan D. (1997) Recent advances in marine microbiology: The Indian scenario. Journal of Marine Biotechnology, 5: 73-81.
Chauhan O.S. and Gujar A.R. (1996) Surficial clay mineral distribution on the southwesterncontinental margin of India: Evidence of input from the Bay of Bengal. Continental ShelfResearch, 16: 321-333.
Christian J.R., Verschell M.A., Murtugudde R., Busalacchi A.J. and McClain C.R. (2002)Biogeochemical modelling of the tropical Pacific Ocean. II. Iron biogeochemistry. Deep-SeaResearch II, in press.
Christiansen B. and Boetius A. (2000) Mass sedimentation of the swimming crab Charybdissmithii (Crustacea: Decapoda) in the deep Arabian Sea. Deep-Sea Research II, 47: 2673-2685.

93
Christiansen B. and Martin B. (2000) Observations on deep-sea benthopelagic nekton at twostations in the northern Arabian Sea: links to organic matter supply? Deep-Sea Research II, 47: 3027-3038.
Codispoti L. and Packard T. (1980) Denitrification rates in the eastern tropical South Pacific.Journal of Marine Research, 38: 453-477.
Codispoti L.A. and Christensen J.P. (1985) Nitrification, denitrification and nitrous oxidecycling in the eastern tropical South Pacific Ocean. Marine Chemistry, 16: 277-300.
Conan S.M.H. and Brummer G.J.A. (2000) Fluxes of planktic foraminifera in response tomonsoonal upwelling on the Somalia Basin margin. Deep-Sea Research II, 47: 2207-2227.
Cook A.A., Lambshead P.J.D., Hawkins L.E., Mitchell N., and Levin L.A. (2000) Nematodeabundance at the oxygen minimum zone in the Arabian Sea. Deep-Sea Research II, 47: 75-85.
Cowie G.L., Calvert S.E., Pedersen T.F., Schulz H., von Rad U. (1999) Organic content andpreservational controls in surfical shelf and slope sediment from the Arabian Sea (Pakistanmargin). Marine Geology, 161: 23-38.
Creasey S., Rogers A., Tyler P., Gage J., and Jollivet D. (2000) Genetic and morphometriccomparisons of squat lobster, Munidopsis scobina (Decapoda : Anomura : Galatheidae)populations, with notes on the phylogeny of the genus Munidopsis. Deep-Sea Research II, 47:87-118.
Crusius J., Calvert S., Pedersen T., and Sage D. (1996) Rhenium and molybdenum enrichmentsin sediments as indicators of oxic, suboxic and sulfidic conditions of deposition. Earth andPlanetary Science Letters, 145: 65-78.
Crutzen P.J. (1991) Methane’s sinks and sources. Nature, 350: 380-381.
Culver M.E. and Perry M.J. (1999) The response of photosynthetic absorption coefficients toirradiance in culture and in tidally mixed estuarine waters. Limnology and Oceanography, 44:24-36.
Cutler A.N., and Swallow J.C. (1984) Surface currents of the Indian Ocean (to 25S, 100E).Intitute of Ocean Science (Wormley), Technical Report 187.
de Wilde H.P.J., and Helder W. (1997) Nitrous oxide in the Somali Basin: the role of upwelling. Deep-Sea Research II, 44: 1319-1340.
den Dulk M., Reichart G.J., Memon G.M., Roelofs E.M.P., Zachariasse W.J., van der ZwaanG.J. (1998) Benthic foraminiferal response to variations in surface water productivity andoxygenation in the northern Arabian Sea. Marine Micropalaeontology, 35: 43-66.
Dennett M.R., Caron D.A., Murzov S.A., Polikarpov I.G., Gavrilova N.A., Georgieva L.V., and Kuzmenko L.V. (1999) Abundance and biomass of nano- and microplankton during the 1995northeast monsoon and spring intermonsoon in the Arabian Sea. Deep-Sea Research II, 46:1691-1717.
Deuser W.G. and Ross E.H. (1980) Seasonal change in the flux of organic carbon to the deepSargasso Sea. Nature, 283: 364-365.
Dickey T.D., Marra J., Sigurdson D.E., Weller R.A., Kinkade C.S., Zedler S.E., Wiggert J.D.and Langdon. C. (1998) Seasonal variability of bio-optical and physical properties in theArabian Sea: October 1994 – October 1995. Deep-Sea Research II, 45: 2001-2025.
Dickson M.L., Orchardo J., Barber R.T., Marra J., McCarthy J.J., and R.N. Sambrotto. (2001)Production and respiration rates in the Arabian Sea during the 1995 northeast and southwestmonsoons. Deep-Sea Research II, 48: 1199-1230.

94
Dittert N., Leynaert A., Ragueneau O., and Heinze C. (2001) Hunting and gathering silicon data to tackle climate forecasting. EOS, 113-117.
Donguy J.R. and Meyers G. (1996) Seasonal variations of sea-surface salinity and temperature in the tropical Indian Ocean. Deep-Sea Research I, 43: 117-138.
Ducklow H.W. (1993) Bacterioplankton distributions and production in the northwestern IndianOcean and Gulf of Oman, September 1986. Deep-Sea Research II, 40: 753-771.
Ducklow H.W. (1999) The bacterial component of the oceanic euphotic zone. FemsMicrobiology Ecology, 30: 1-10.
Ducklow H.W., Smith D.C., Campbell L., Landry M.R., Quinby H.L., Steward G.F., and AzamF. (2001) Heterotrophic bacterioplankton in the Arabian Sea: Basinwide response to year-round high primary productivity. Deep-Sea Research II, 48(6-7): 1303-1323.
Düing W. (1970) The Monsoon Regime of the Currents in the Indian Ocean. East-West CenterPress, Honolulu, 68pp.
Edwards E.S., Burkill P.H., and Stelfox C.E. (1999) Zooplankton herbivory in the Arabian Seaduring and after the SW monsoon, 1994. Deep-Sea Research II, 46: 843-863.
Eglinton T.I., Benitez-Nelson B.C., Pearson A., McNichol A.P., Bauer J.E., and Druffel E.R.M. (1997) Variability in radiocarbon ages of individual organic compounds from marinesediments. Science, 277: 796-799.
Elliot A.J. and Savidge G. (1990) Some features of the upwelling off Oman. Journal MarineResearch, 48: 319-333.
Eremeev V.N., Ivanov V.A., Kosarev A.N. and Polyakova A.V. (1994) Natural peculiarities ofthe Arabian Sea. In: Biogeochemical Processes in the Arabian Sea, U.S.-C.I.S. Arabian SeaWorkshop. Sevastopol, Ukraine, p. 26-34.
Ewald S. (1998) Satellitengestützte Abschätzung der Einflüsse von Kühler haut undSchaumbedeckung des Ozeans auf den globalen CO2-Fluß zwischen Ozean und Atmosphäre.Dissertation, Universität Hamburg.
Faber E., Gerling P., Berner U. and Sohns E. (1994) Methane in ocean waters - concentrationand carbon-isotope variability at East Pacific Rise and in the Arabian Sea. EnvironmentalMonitoring and Assessment. 31(1-2): 139-144.
Farrenkopf A.M., Dollhopf M.E., Chadhain S.N., Luther III G.W., and Nealson K.H. (1997)Reduction of iodate in seawater during Arabian Sea shipboard incubations and in laboratorycultures of the marine bacterium Shewanella putrefaciens strain MR-4. Marine Chemistry,57(3-4): 347-354.
Fasham M.J.R. and Evans G.T. (2000) Advances in ecosystem modelling within JGOFS. In:R.B. Hanson, H.W. Ducklow and J.G. Field (Eds.). The Changing Ocean Carbon Cycle: Amidterm synthesis of the Joint Global Ocean Flux Study. Cambridge University Press,Cambridge, 514pp.
Fieux M. and Stommel H. (1977) Onset of southwest monsoon over the Arabian Sea frommarine reports of surface winds: structure and variability. Monthly Weather Review, 105: 231-236.
Findlater J. (1966) Cross-equatorial jet stream at low level over Kenya. MeteorologicalMagazine, 95: 353-364.
Findlater J. (1969) A major low-level air current near the Indian Ocean during the northernsummer. Quarterly Journal of the Royal Meteorological Society, 95: 362-380.

95
Findlater J. (1971) Mean monthly air flow at low levels over the western Indian Ocean.Geophysical Memoirs, London, 16: 1-53.
Fine R.A. (1993) Circulation of Antarctic Intermediate Water in the South Indian Ocean. Deep-Sea Research I, 40: 2021-2042.
Fischer A.S. (2000) The Upper Ocean Response to the Monsoon in the Arabian Sea. Ph.D.Dissertation, Massachusetts Institute of Technology, Woods Hole Oceanographic Institution,222pp.
Flagg C.N. and Kim H.S. (1998) Upper ocean currents in the northern Arabian Sea fromshipboard ADCP measurements collected during the 1994-1996 U.S. JGOFS and ONRprograms. Deep Sea Research II, 45: 1917-1959.
Fleminger A. (1986) The pleistocene equatorial barrier between the Indian and Pacific Oceansand a likely cause of Wallace’s line. UNESCO Technical Papers in Marine Science, 49: 84-97.
Foxton P. (1965) A mass fish mortality on the Somali Coast. Deep-Sea Research, 12: 17-19.
Gage J.D., Levin L.A. and Wolff G.A. (2000) Benthic processes in the Arabian Sea: introduction and overview. Deep-Sea Research II, 47: 1-7.
Gardner W.D., Gundersen J.S., Richardson M.J., and Walsh I.D. (1999) The role of seasonal and diel changes in mixed-layer depth on carbon and chlorophyll distributions in the Arabian Sea.Deep-Sea Research II, 46: 1833-1858.
Garrison D.L., Gowing M.M., and Hughes M.P. (1998) Nano- and microplankton in the northern Arabian Sea during the southwest monsoon, August-September 1995 - A US-JGOFS study.Deep-Sea Research II, 45: 2269-2299.
Garrison D.L., Gowing M.M., Hughes M.P., Campbell L., Caron D.A., Dennett M.R.,Shalapyonok A., Olson R.J., Landry M.R., Brown S.L., Liu H.B., Azam F., Steward G.F.,Ducklow H.W., and Smith D.C. (2000) Microbial food web structure in the Arabian Sea: a US JGOFS study. Deep-Sea Research II, 47: 1387-1422.
Gauns M., Mohanraju R., and Madhupratap M. (1996) Studies on the microzooplankton fromthe central and eastern Arabian Sea. Current Science, 71: 874-877.
George M.D., Dileep Kumar M., Naqvi S.W.A., Banerjee S., Narekar P.V., de Sousa S.N., andJayakumar D.A. (1994) Study of the carbon dioxide system in the northern Indian Oceanduring premonsoon. Marine Chemistry, 47: 243-254.
Gibb S.W., Mantoura R.F.C., Liss P.S., and Barlow R.G. (1999a) Distributions andbiogeochemistries of methylamines and ammonium in the Arabian Sea. Deep-Sea Research II, 46: 593-615.
Gibb S.W., Mantoura R.F.C. and Liss P.S. (1999b) Ocean-atmosphere exchange andatmospheric speciation of ammonia and methylamines in the region of the NW Arabian Sea.Global Biogeochemical Cycles, 13: 161-178.
Gjøsaeter J. (1984) Mesopelagic fish, a large potential resource in the Arabian Sea. Deep-SeaResearch, 31: 1019-1035.
GLOBEC (1993) Implementation plan and workshop report for U.S. GLOBEC studies in theArabian Sea. U.S. Global Ocean Ecosystem Dynamics Report 9, University of California,Berkeley, CA, 105pp.
Goericke R., Olson R. and Shalpyonok A. (2000) A novel niche for Prochlorococcus sp. in low-light suboxic environments in the Arabian Sea and the Eastern Tropical North Pacific. Deep-Sea Research I, 47: 1183-1205.

96
Gooday A.J., Bett B.J., Shires R., and Lambshead P.J.D. (1998) Deep-sea benthic foraminiferalspecies diversity in the NE Atlantic and NW Arabian Sea: a synthesis. Deep-Sea Research II,45: 165-201.
Gooday A.J., Bernhard J.M., Levin L.A., and Suhr S.B. (2000a) Foraminifera in the Arabian Sea oxygen minimum zone and other oxygen-deficient settings: taxonomic composition, diversity,and relation to metazoan faunas. Deep-Sea Research II, 47: 25-54.
Gooday A.J., Bowser S.S., Bett B.J., and Smith C.R. (2000b) A large testate protist, Gromiaspaerica sp. nov. (Order Filosea), from the bathyal Arabian Sea. Deep-Sea Research II, 47: 55-73.
Goswami S., Gajbhiye S. and Padmavati G. (1992) Distribution of Pleuromamma (Copepoda:Metridiidae) along a north-south transect in the Indian Ocean. In: B.N. Desai (Ed.),Oceanography of the Indian Ocean, Oxford and IBH Publishing Co. Ltd., New Delhi, p. 157-166.
Gowing M.M. and Wishner K.F. (1998) Feeding ecology of the copepod Lucicutia aff. L-grandis near the lower interface of the Arabian Sea oxygen minimum zone. Deep-Sea Research II, 45: 2433-2459.
Goyet C., Millero F.J., O’Sullivan D.W., Eischeid G., McCue S.J. and Bellerby R.G.J. (1998)Temporal variations of pCO2 in surface seawater of the Arabian Sea in 1995. Deep-SeaResearch I, 45: 609-623.
Goyet C., Coatanoan C., Eischeid G., Amaoka T., Okuda K., Healy R. and Tsunogai S. (1999)Spatial variations of total CO2 and total alkalinity in the northern Indian Ocean: A novelapproach for the quantification of anthropogenic CO2 in seawater. Journal Marine Research,57: 135-163.
Grandel S., Rickert D., Schlüter M., and Wallmann K. (2000) Pore-water distribution andquantification of diffusive benthic fluxes of silicic acid, nitrate and phosphate in surfacesediments of the deep Arabian Sea. Deep-Sea Research II, 47: 2707-2734.
Gundersen J., Gardner W.D., Richardson M.J., and Walsh I.D. (1998) Effects of monsoons onthe seasonal and spatial distribution of POC and chlorophyll in the Arabian Sea. Deep-SeaResearch II, 45: 2103-2132.
Gust G., Byrne R.H., Bernstein R.E., Betzer P.R., and Bowles W. (1992) Particle fluxes andmoving fluids: Experience from synchronous trap collections in the Sargasso Sea. Deep- SeaResearch, 39: 1071-1083.
Gust G., Michaels A.F., Johnson R., Deuser W.G. and Bowles W. (1994) Mooring line motionsand sediment trap hydromechanics: In situ intercomparison of three common deploymentdesigns. Deep-Sea Research, 41: 831-857.
Haake B., Ittekkot V., Rixen T., Ramaswamy V., Nair R.R., and Curry W.B. (1993) Seasonalityand interannual variability of particle fluxes to the deep Arabian Sea. Deep-Sea Research, 40:1323-1344.
Halpern D., Freilich M.H., and Weller R.A. (1998) Arabian Sea Surface winds and oceantransports determined from ERS-1 scatterometer. Journal Geophysical Research-Oceans, 103:7799-7806.
Harrison K.G. (2000) Role of increased marine silica input on paleo-pCO2 levels.Paleoceanography, 15(3): 292-298.
Hatton A.D., Malin G., and Liss P.S. (1999) Distribution of biogenic sulphur compounds during and just after the SW monsoon in the Arabian Sea. Deep-Sea Research II, 46: 617-632.

97
Hedges J.I., Baldock J.A., Gélinas Y., Lee C., Peterson M., and Wakeham S. (2001) Evidencefor non-selective preservation of organic matter in sinking marine particles. Nature, 409(6822): 801-804.
Herring P., Fasham M, Weeks A., Hemmings J., Roe H., Pugh P., Holley S., Crisp N., and Angel M. (1998) Across-slope relations between the biological populations, the euphotic zone and the oxygen minimum layer off the coast of Oman during the southwest monsoon (August, 1994).Progress in Oceanography, 41: 69-109.
Honjo S., Dymond J., Prell W., and Ittekkot V. (1999) Monsoon-controlled export fluxes to the interior of the Arabian Sea. Deep-Sea Research, 46: 1859-1902.
Hoppe H.G. and Ullrich S. (1999) Profiles of ectoenzymes in the Indian Ocean: phenomena ofphosphatase activity in the mesopelagic zone. Aquatic Microbial Ecology, 19: 139-148.
Houghton J.T., Filho L.G.M, Callander A.A., Harris N., Kattenberg A., and Maskell K. (1996)Climate Change 1995 - The Science of Climate Change. Cambridge University Press,Cambridge, 572pp.
Howard K.L. and Yoder J.A. (1997) Contribution of the subtropical oceans to global primaryproduction. In: Liu C.T. (Ed.), Space Remote Sensing of Subtropical Oceans. Pergamon Press, London, p. 157-167.
Howell E.A., Doney S.C., Fine R.A., and Olson D.B. (1997) Geochemical estimates ofdenitrification in the Arabian Sea and the Bay of Bengal. Geophysical Research Letters, 24:3549-2552.
Ikeda T. (1974) Nutritional ecology of marine zooplankton. Memoirs of the Faculty of Fisheries, Hokkaido University, 22: 1-97.
IOCCG (1998) Minimum requirements for an operational ocean-colour sensor for the openocean. International Ocean Colour Coordinating Group, Dartmouth, Canada (Rep No. 1,IOCCG).
Jahnke R.A. (1996) The global ocean flux of particulate organic carbon: Areal distribution andmagnitude. Global Biogeochemical Cycles, 10: 71-88.
Jannink N.T., Zachariasse W.J., and van der Zwaan G.J. (1998) Living (Rose Bengal stained)benthic foraminifera from the Pakistan continental margin (northern Arabian Sea). Deep-SeaResearch I, 45: 1483-1513.
Jayakumar D.A., Naqvi S.W.A., Narvekar P.V., and George M.D. (2001) Methane in coastal and offshore waters of the Arabian Sea. Marine Chemistry, 74: 1-13.
Johns W.E. and Olson D.B. (1999) Observations of seasonal exchange through the Straight ofHormuz. Oceanography, 11, p. 58.
Johnson G.C., Warren, B.A., and Olson D.B. (1991a) A deep boundary current in the ArabianBasin. Deep-Sea Research, 38: 653-661.
Johnson G.C., Warren, B.A. and Olson D.B. (1991b) Flow of bottom water in the Somali Basin. Deep-Sea Research, 38: 637-652.
Johnsen G and Sakshaug E. (1993) Bio-optical characteristics and photoadaptive responses inthe toxic and bloom-forming dinoflagellates Gyrodinium aureolum, Gymnodiniumgalatheanum and two strains of Prorocentrum minimum. Journal Phycology, 29: 627-642.
Johnson Z., Landry M., Bidigare R., Brown S., Campbell L., Gunderson J., Marra J. and TreesC. (1999) Energetics and growth kinetics of a deep Prochlorococcus spp. population in theArabian Sea. Deep-Sea Research II, 46: 1719-1743.

98
Johnson Z., Bidigare R., Goericke R., Marra J., Trees C., and Barber R. (2002) Photosyntheticphysiology and physicochemical forcing in the Arabian Sea, 1995. Deep-Sea Research I, 49, in press.
Keen T.R., Kindle J.C., and Young D.K. (1997) The interaction of southwest monsoonupwelling, advection and primary production in the northwest Arabian Sea. Journal MarineSystems, 13: 61-82.
Keil R.G. and Cowie G.L. (1999) Organic matter preservation through the oxygen minimumzone of the NE Arabian Sea as discerned by mineral-organic interactions. Marine Geology,161: 13-22.
Kiefer D.A. and Kremer J.N. (1981) Origins of vertical patterns of phytoplankton and nutrientsin the temperate, open ocean: a stratigraphic hypothesis. Deep-Sea Research, 28: 1087-1105.
Kim H.-S., Flagg C.N., and Howden S.D. (2001) Northern Arabian Sea variability fromTOPEX/Poseidon altimetry data: an extension of the U.S. JGOFS/ONR shipboard ADCPstudy. Deep Sea Research, II, 48: 1069-1096.
Kindle J.C. (2002) The surface circulation of the northern Arabian Sea. In: L. Watts, P. Burkilland S. Smith (Eds.), Arabian Sea Process Study. JGOFS International Planning Office, Bergen, Norway, this volume.
Kinkade C., Marra J., Dickey T., and Weller R. (2000) An annual cycle of phytoplanktonbiomass in the Arabian Sea, 1994-1995, as determined by moored optical sensors. Deep-SeaResearch II, 48: 1285-1301.
Kinzer J., Böttger-Schnack R., and Schulz K. (1993) Aspects of horizontal distribution and dietof myctophid fish in the Arabian Sea with reference to the deep water oxygen deficiency.Deep-Sea Research II, 40: 783-800.
Koning E., Brummer G.J., van Raaphorst W., van Bennekom J., Helder W., and van Iperen J.(1997) Settling dissolution and burial of biogenic silica in the sediments off Somalia(northwestern Indian Ocean). Deep-Sea Research II, 44: 1341-1360.
Koppelmann R., Schäfer P., and Schiebel R. (2000) Organic carbon losses measured byheterotrophic activity of mesozooplankton and CaCO3 flux in the bathypelagic zone of theArabian Sea. Deep-Sea Research II, 47: 169-187.
Körtzinger A., Duinker J.C., and Mintrop L. (1997) Strong CO2 emissions from the Arabian Sea during the southwest monsoon. Geophysical Research Letters, 24: 1763-1766.
Krey J. and B. Babenerd. (1976) Phytoplankton Production Atlas of the International IndianOcean Expedition. Intergovernmental Oceanographic Commission, Paris, 70pp.
Kromcamp J., De Bie M, Goosen N., Peene J., Van Rijswijk P., Sinke J., and Duineveld G.C.A. (1997) Primary production by phytoplankton along the Kenyan coast during the SE monsoonand November intermonsoon 1992, and the occurrence of Trichodesmium. Deep Sea Research II, 44: 1195-1212.
Kumar M.D., Rajendran A., Somasundar K., Haake B., Jenisch A., Shuo Z., Ittekkot V., andDesai B.N. (1990) Dynamics of Dissolved Organic-Carbon in the Northwestern Indian-Ocean.Marine Chemistry, 31: 299-316.
Kumar M.D. and Li Y.H. (1996) Spreading of water masses and regeneration of silica and 226Rain the Indian Ocean. Deep-Sea Research I, 43: 83-110.
Kumar M.D., Naqvi S.W.A., George M.D., and Jayakumar D.A. (1996) A sink for atmosphericcarbon dioxide in the northern Bay of Bengal. Journal Geophysical Research, 101: 18121-18125.

99
Kumar M.D., Sarma V., Ramaiah N., Gauns M., and de Sousa S.M. (1998) Biogeochemicalsignificance of transport exopolymer particles in the Indian Ocean. Geophysical ResearchLetters, 25: 81-84.
Kumar S.P. and Prasad, T.G. (1996) Winter cooling in the northern Arabian Sea. CurrentScience, 71: 834-841.
Kumar S.P., Ramaiah N., Gauns M., Sarma V.V.S.S., Muraleedharan P.M., Raghukumar S.,Dileep Kumar M., and Madhupratap M. (2001) Physical forcing of biological productivity inthe northern Arabian Sea during the northeast monsoon. Deep Sea Research II, 48: 1115-1126.
Kurbjeweit F., Schmiedl G., Shiebel R., Hemleben C., Pfannkuche O., Wallman K. and SchafeP. (2000) Distribution, biomass and diversity of benthic foraminifera in relation to sedimentgeochemistry in the Arabian Sea. Deep-Sea Research II, 47: 2913-2955.
Lal D. (1994) Biogeochemistry of the Arabian Sea: Present information and gaps. Proceedingsof the Indian Academy of Sciences, Earth and Planetary Sciences, 103: 99-106.
Lal S. and Patra P.K. (1998) Variabilities in the fluxes and annual emissions of nitrous oxidefrom the Arabian Sea. Global Biogeochemical Cycles, 12: 321-327.
Lamont P.A. and Gage J.D. (2000) Morphological responses of macrobenthic polychaetes to low oxygen on the Oman continental slope, NW Arabian Sea. Deep-Sea Research II, 47: 9-24.
Lampitt R.S. and Antia A.N. (1997) Particle flux in deep seas: regional characteristics andtemporal variability. Deep-Sea Research I, 44: 1377-1403.
Landry M.R., Brown S.L., Campbell L., Constantinou J., and Liu H. (1998) Spatial patterns inphytoplankton growth and microzooplankton grazing in the Arabian Sea during monsoonforcing. Deep-Sea Research II, 45: 2353-2368.
Lane P., Smith S., Zaragoza J., Prusova I., and Roman M. (1998) Copepod taxonomy,abundance and biomass in the upper 300 meters of the Arabian Sea during the southwestmonsoon (August/September) of 1995. RSMAS Technical Report 98-007, University ofMiami, Miami, FL, 409pp.
Latasa M. and Bidigare R.R. (1998) A comparison of phytoplankton populations of the Arabian Sea during the Spring Intermonsoon and Southwest Monsoon of 1995 as described by HPLC-analyzed pigments. Deep Sea Research II, 45:2133-2170.
Law C.S. and Owens N.J.P. (1990) Significant flux of atmospheric nitrous oxide from thenorthwest Indian Ocean. Nature, 346: 826-828.
Lee C., Wakeham S.G., and Hedges J.I. (1988) The measurements of oceanic particle flux - Are swimmers a problem? Oceanography, 1: 24-36.
Lee C., Murray D.W., Barber R.T., Buesseler K.O., Dymond J.D., Hedges J.I., Honjo S.,Manganini S.J., Marra J., Moser C., Peterson M.L., Prell W.L., and Wakeham S.G. (1998)Particulate organic carbon fluxes: compilation of results from the 1995 US JGOFS Arabian Sea Process study. Deep-Sea Research II, 45: 2489-2501.
Lee C.M., Jones B.H. and Brink K.H. (2000) The upper ocean response to monsoonal forcing in the Arabian Sea: seasonal and spatial variability. Deep Sea Research II, 47: 1177-1226.
Levin L.A. and Edesa S. (1997) The ecology of cirratulid mudballs on the Oman margin,northwest Arabian Sea. Marine Biology, 128: 671-678.
Levin L.A., Gage J.D., Martin C., and Lamont P.A. (2000) Macrobenthic community structurewithin and beneath the oxygen minimum zone, NW Arabian Sea, Deep-Sea Research II, 47:189-226.

100
Lewis B. and Luther III G. (2000) Processes controlling the distribution and cycling ofmanganese in the oxygen minimum zone of the Arabian Sea. Deep-Sea Research II, 47: 1541-1562.
Littke R., Lückge A., and Welte D.H. (1997) Quantification of organic matter degradation bymicrobial sulphate reduction for quaternary sediments from the northern Arabian Sea.Naturwissenschaften, 84(7): 312-315.
Longhurst A.R., Sathyendranath S., Platt T. and Caverhill C. (1995) An estimate of globalprimary production in the ocean from satellite radiometer data. Journal Plankton Research,17(6): 1245-1271.
Longhurst A.R. (1998) Ecological Geography of the Sea. Academic Press, San Diego, 398pp.
Louanchi F., Metzl N., and Poisson A. (1996) Modelling of monthly sea surface fCO2 in theIndian Ocean. Marine Chemistry, 55: 265-279.
Lückge A., Ercegovac M., Strauss H., and Littke R. (1999) Early diagenetic alteration of organic matter by sulfate reduction in Quaternary sediments from the northeastern Arabian Sea. Marine Geology, 158: 1-13.
Luff R., Wallmann K., Grandel S., and Schlüter M. (2000) Numerical modeling of benthicprocesses in the deep Arabian Sea. Deep-Sea Research II, 47: 3039-3072.
Luo J., Ortner P., Forcucci D., and Cummings S. (2000) Diel vertical migration of zooplanktonand mesopelagic fish in the Arabian Sea. Deep-Sea Research II, 47: 1451-1474.
Luther M.E., O’Brien J.J., and Prell W.L. (1990) Variability in upwelling fields in thenorthwestern Indian Ocean, 1, Model experiments for the last 18,000 years. Paleoceanography, 5: 433-445.
Madhupratap M., Gopalakrishnan T., Haridas P., Nair K., Aravindakshan P., Padmavati G., andPaul S. (1996a) Lack of seasonal and geographic variation in mesozooplankton biomass in the Arabian Sea and its structure in the mixed layer. Current Science, 71: 863-868.
Madhupratap M., Prasanna Kumar S., Bhattahiri P.M.A., Dileep Kumar M., Raghukumar S.,Nair K.K.C., and Ramaiah N. (1996b) Mechanisms of the biological response to winter coolingin the northeastern Arabian Sea. Nature, 384: 549-552.
Maier-Reimer E. (1993) Geochemical cycles in an ocean general circulation model. Preindustrial tracer distribution. Global Biogeochemical Cycles, 7: 645-677.
Maier-Reimer E. (2001) Nutrient supply to the Arabian Sea - A model study. Deep-Sea Research I, submitted.
Manghnani V., Morrison J.M., Hopkins T.S. and Böhm E. (1998) Advection of upwelled waters in the form of plumes off Oman during the southwest monsoon. Deep-Sea Research II, 45:2027-2052.
Mantoura R.F.C., Law C.S., Owens N.J.P, Burkill P.H., Woodward E.M.S., Howland R.J.M.,and Llewellyn C.A. (1993) Nitrogen biogeochemical cycling in the northwestern Indian Ocean. Deep-Sea Research II, 40: 651-671.
Marra J., Dickey, T.D., Kinkade, C.S., Ho. C., Sigurdson, D.E., Weller, R. and Barber R.T.(1998) Variability in primary production as observed from moored sensors in the centralArabian Sea in 1995. Deep-Sea Research II, 45: 2253-2268.
Marra J., Trees C.C., Bidigare R.R., and Barber R.T. (2000) Pigment absorption and quantumyields in the Arabian Sea. Deep Sea Research II, 47: 1279-1299.

101
Martin J.H., Gordon R.M. and Fitzwater S.F. (1991) The case for iron. Limnology andOceanography, 36: 1793-1803.
Mathew K., Naomi T., Antony G., and Scariah K. (1990) Distribution of euphausiacea in spaceand time in the Indian EEZ and contiguous seas. In: K. Mathew (Ed.), Proceedings of FirstWorkshop of Scientific Results, FORV Sagar Sampada, Central Marine Fisheries ResearchInstitute, Cochin, India, p. 121-127.
McCarthy J.J., Garside C., and Nevins J.L. (1999) Nitrogen dynamics during the Arabian Seanortheast monsoon. Deep-Sea Research II, 46: 1623-1664.
McCreary J.P. Jr., Kohler K.E., Hood R.R. and Olson D.B. (1996) A four-component ecosystem model of biological activity in the Arabian Sea. Progress in Oceanography, 37: 193-240.
McCreary J.P., Kohler K.E., Hood R.R., Smith S., Kindle J., Fischer A. and Weller R.A. (2001)Influences of diurnal and intraseasonal forcing on mixed-layer and biological variability in the central Arabian Sea. Journal Geophysical Research, 106: 7139-7155.
Meadows A., Meadows P.S., West F.J.C., and Murray J.M.H. (2000) Bioturbation, geochemistry and geotechnics of sediments affected by the oxygen minimum zone on the Oman continentalslope and abyssal plain, Arabian Sea. Deep-Sea Research II, 47: 259-280.
Measures C.I. and Vink S. (1999) Seasonal variations in the distribution of Fe and Al in thesurface waters of the Arabian Sea. Deep-Sea Research II, 46: 1597-1622.
Mensah M. (1974) The reproduction and feeding of the marine copepod Calanoides carinatus(Krøyer) in Ghanaian waters. Ghana Journal of Science, 14: 167-191.
Metzl N., Tilbrook B., and Poisson A. (1999) The annual fCO2 and the air-sea CO2 flux in thesub-Antarctic Ocean. Tellus, 51B: 849-861.
Mintrop L., Körtzinger A. and Duinker J.C. (1999) The carbon dioxide system in the northwestIndian Ocean during the southwest monsoon. Marine Chemistry, 64: 315-336.
Morrison J.M. and Olson D.B. (1992) Seasonal basinwide extremes in T-S characteristics in the near surface waters of the Arabian Sea and Somali Basin. In: B.N. Desai (Ed.), Oceanographyof the Indian Ocean. Oxford and IBH Publishing Co. Ltd., New Delhi, p. 605-616.
Morrison J.M. (1997) Inter-monsoonal changes in the TS properties of the near-surface waters of the northern Arabian Sea. Geophysical Research Letters, 24: 2553-2556.
Morrison J.M., Codispoti L.A., Gaurin S., Jones B., Manghnani V. and Zheng Z. (1998)Seasonal variation of hydrographic and nutrient fields during the US JGOFS Arabian SeaProcess Study. Deep-Sea Research II, 45: 2053-2102.
Morrison J.M., Codispoti L.A., Smith S.L., Wishner K, Flagg C., Gardner W.D., Gaurin S.,Naqvi S., Manghnani V., Prosperie L. and Gundersen J.S. (1999) The oxygen minimum zonein the Arabian Sea during 1995. Deep-Sea Research Part II-Topical Studies in Oceanography,46(8-9): 1903-1931.
Muraleedharan P.S. and Kumar S.P. (1996) Arabian Sea upwelling - A comparison betweencoastal and open ocean regions. Current Science, 71: 842-846.
Murray J.M.H., Meadows P.S., Meadows A., West F.J.C., and Wood D.M. (2000) Microscalegeotechnical variability in continental slope and abyssal sediments influenced by the oxygenminimum zone in the Arabian Sea. Deep-Sea Research II, 47: 281-301.
Murray S.P. and Johns W. (1997) Direct observations of seasonal exchange through the Bab elMandab Strait. Geophysical Research Letters, 24: 2557-2560.

102
Murtugudde, R. and Busalacchi, A.J. (1999) Interannual variability of the dynamics andthermodynamics of the tropical Indian Ocean. Journal of Climate, 12: 2300-2326.
Murtugudde R.G., Signorini S.R., Christian J.R., Busalacchi A.J., McClain C.R. and Picaut J.(1999) Ocean color variability of the tropical Indo-Pacific basin observed by SeaWiFS during1997-1998. Journal Geophysical Research, 104: 18351-18366.
Naidu P.D. and Malmgren B.A. (1995) Do benthic foraminifer records represent a productivityindex in oxygen minimum zone areas? An evaluation from the Oman margin, Arabian Sea.Marine Micropaleontology, 26: 49-55.
Nair R.R., Ittekkot V., Manganini S.J., Ramaswamy V., Haake B., Degens E.T., Desai B.N. and Honjo S. (1989) Increased particle flux to the deep ocean related to monsoons. Nature, 338:749-751.
Nair T.M.B., Ramaswamy V., Shankar R., and Ittekkot V. (1999) Seasonal and spatial variations in settling manganese fluxes in the northern Arabian Sea. Deep-Sea Research I, 46: 1827-1839.
Naqvi S.W.A. (1987) Some aspects of the oxygen-deficient conditions and denitrification in the Arabian Sea. Journal Marine Research, 49: 1049-1072.
Naqvi S.W.A. (1991) Denitrification processes in the Arabian Sea. In: Lal D. (Ed.)Biogeochemistry of the Arabian Sea. Indian Academy of Sciences, Bangalore, p. 181-202.
Naqvi S.W.A. and R.J. Noronha. (1991) Nitrous oxide in the Arabian Sea. Deep-Sea Research,38: 871-890.
Naqvi S., Noronha R, Shailaja M., Somasundar K., and Sen Gupta R. (1992) Some aspects of the nitrogen cycling in the Arabian Sea. In: B. Desai (Ed.), Oceanography of the Indian Ocean.Oxford and IBH Publishing Co., New Delhi, p. 285-311.
Naqvi S.W.A. and Shailaja M.S. (1993) Activity of the respiratory electron-transport system and respiration rates within the oxygen minimum layer of the Arabian Sea. Deep-Sea Research II,40: 687-695.
Naqvi S.W.A., Kumar M.D., Narvekar P.V., de Sousa S.N., George M.D., and D’Silva C. (1993) An intermediate nepheloid layer associated with high microbial metabolic rates anddenitrification in the northwest Indian Ocean. Journal of Geophysical Research, 98(C9):16469-16479.
Naqvi S.W.A., Shailaja M.S., Kumar M.D., and Sen Gupta R. (1996) Respiration rates insubsurface waters of the northern Indian Ocean: Evidence for low decomposition rates oforganic matter within the water column in the Bay of Bengal. Deep-Sea Research II, 43: 73-81.
Naqvi S.W.A., Yoshinari T., Jayakumar D.A., Altabet M.A., Narvekar P.V., Devol A.H.,Brandes J.A. and Codispoti L.A. (1998) Budgetary and biogeochemical implications of N2Oisotope signatures in the Arabian Sea. Nature, 394: 462-464.
Naqvi S.W.A., Jayakumar D.A., Narvekar P.V., Naik H., Sarma V.V.S.S., D’Souza W., JosephT. and George M.D. (2000) Increased marine production of N2O due to intensifying anoxia on the Indian continental shelf. Nature, 408: 346-349.
Naqvi S.W.A. (2002) Air-sea interactions and exchanges. In: L. Watts, P. Burkill and S. Smith(Eds.), Arabian Sea Process Study. JGOFS International Planning Office, Bergen, Norway, this volume.
Nath B.N., Bau M., Rao B.R., and Rao C.M. (1997) Trace and rare earth elemental variation inArabian Sea sediments through a transect across the oxygen minimum zone. GeochimicaCosmochimica Acta, 61: 2375-2388.

103
Nevison C.D., Weiss R.F., and Erickson D.J. (1995) Global oceanic emissions of nitrous oxide.Journal of Geophysical Research, 100: 15809-15820.
Olson D.B., Hitchcock, G.L., Fine, R.A., and Warren B.A. (1993) Maintenance of the low-oxygen layer in the central Arabian Sea. Deep-Sea Research, 40: 673-685.
Owens N.J.P., Law C.S., Mantoura R.F.C., Burkill P.H., and Llewellyn C.A. (1991) Methaneflux to the atmosphere from the Arabian Sea. Nature, 354: 293-295.
Owens N.J.P., Burkill P.H., Mantoura R.F.C., Woodward E.M.S., Bellan I.E., Aiken J., andLlewellyn R.J.M. (1993) Size-fractionated primary production and nitrogen assimilation in the northwestern Indian Ocean. Deep-Sea Research II, 40: 711-736.
Pace M.L., Knauer G.A., Karl D.M., and Martin J.H. (1987) Primary production, newproduction and vertical flux in the eastern Pacific Ocean. Nature, 325: 803-804.
Paropkari A.L., Babu C.P., and Mascarenhas A. (1992) A critical evaluation of depositionalparameters controlling the variability of organic carbon in the Arabian Sea sediments. MarineGeology, 107: 213-226.
Passier H.F., Luther G.W. III, DeLange G.J. (1997) Early diagenesis and sulphur speciation insediments of the Oman margin, northwestern Arabian Sea. Deep-Sea Research II, 44: 1361-1380.
Patra P.K., Lal S., Venkataramani S., Gauns M., and Sarma V.V.S.S. (1998) Seasonal variability in distribution and fluxes of methane in the Arabian Sea. Journal Geophysical Research, 103:1167-1176.
Patra P.K., Lal S., Venkataramani S., de Dousa S.N., Sarma V.V.S.S., and Sardesai S. (1999)Seasonal and spatial variability in N2O distribution in the Arabian Sea. Deep-Sea Research I,46: 529-543.
Pedersen T.F., Shimmield G.B., and Price N.B. (1992) Lack of enhanced preservation of organic matter in sediments under the oxygen minimum zone on the Oman margin. GeochimicaCosmochimica Acta, 56: 545-551.
Pfannkuche O., Sommer S., and Kähler A. (2000) Coupling between phytodetritus depositionand the small-sized benthic biota in the deep Arabian Sea: analyses of biogenic sedimentcompounds. Deep-Sea Research II, 47: 2805-2833.
Pinkerton M.H., Trees C.C., Aiken J., Bale A.J., Moore G.F., Barlow R.G., and Cummings D.G. (1999) Retrieval of near-surface bio-optical properties of the Arabian Sea from remotelysensed ocean colour. Deep Sea Research II, 46: 549-569.
Platt T. and Sathyendranath S. (1988) Oceanic primary production: Estimation by remotesensing at local and regional scales. Science, 241: 1613 – 1620.
Platt T., Sathyendranath S., and Longhurst A. (1995) Remote sensing of primary production inthe ocean: promise and fulfilment. Philosophical Transactions Royal Society London B, 348:191-202.
Platt T. and Sathyendranath S. (1999) Spatial structure of pelagic ecosystem processes in theglobal ocean. Ecosystems, 2: 384-394.
Pollehne F., Klein B., and Zeitzschel B. (1993) Low light adaptation and export production inthe deep chlorophyll maximum layer in the northern Indian Ocean. Deep-Sea Research II, 40:737-752.
Pomroy, A. and Joint I. (1999) Bacterioplankton activity in the surface waters of the Arabian Sea during and after the 1994 SW monsoon. Deep-Sea Research II, 46: 767-794.

104
Prahl F., Dymond J., and Sparrow M.A. (2000) Annual biomarker record for export productionin the central Arabian Sea. Deep-Sea Research II, 47: 1581-1604.
Prospero J.M. (1996) The atmospheric transport of Particles to the Ocean. In: V. Ittekkot, P.Schafer, S. Honjo and P.J. Depetris (Eds.), Particle Flux in the Ocean, SCOPE 57. John Wiley and Sons, New York, p. 19-52.
Qasim S. (1977) Contribution of zooplankton in the food chains of some warm waterenvironments. Proceedings of the Symposium on Warm Water Zooplankton. National Instituteof Oceanography, Goa, p. 700-708.
Quadfasel D., and Schott F. (1982) Water mass distribution at intermediate layers off the Somali coast during the onset of the southwest monsoon, 1979. Journal of Physical Oceanography, 12: 1358-1372.
Raghukumar C. and Raghukumar S. (1998) Barotolerance of fungi isolated from deep-seasediments of the Indian Ocean. Aquatic Microbial Ecology, 15(2): 153-163.
Raghukumar S. and Raghukumar C. (1999) Thraustochytrid fungoid protists in faecal pellets ofthe tunicate Pegea confoederata, their tolerance to deep-sea conditions and implication indegradation processes. Marine Ecology-Progress Series, 190: 133-140.
Ramaiah N., Raghukumar S., and Gauns M. (1996) Bacterial abundance and production in thecentral and eastern Arabian Sea. Current Science, 71: 878-882.
Ramaiah N., Sarma A., Gauns M., Kumar D.M., and Madhupratap M. (2000) Abundance andrelationship of bacteria with transparent exopolymer particles during the 1996 summermonsoon in the Arabian Sea. Proceedings of the Indian Academy of Sciences, Earth andPlanetary Sciences, 109: 443-451.
Ramaswamy V. and Nair R.R. (1994) Fluxes of material in the Arabian Sea and Bay of Bengal –Sediment trap studies. Proceedings of the Indian Academy of Sciences, Earth PlanetarySciences, 103: 189-210.
Rao B.R. and Veerayya M. (2000) Influence of marginal highs on the accumulation of organiccarbon along the continental slope off western India. Deep-Sea Research II, 47: 303-327.
Rao T.S.S. (1973) Zooplankton studies in the Indian Ocean. In: B. Zeitzschel and S. Gerlach(Eds.), The Biology of the Indian Ocean. Springer-Verlag, New York, p. 243-255.
Rao V.P., Lamboy M., and Natarajan R. (1992) Possible microbial origin of phosphorites onError Seamount, northwestern Arabian Sea. Marine Geology, 106: 149-164.
Rao V.P. and Rao B.R. (1995) Provenance and distribution of clay minerals in the sediments ofthe western continental shelf and slope of India. Continental Shelf Research, 15: 1757-1771.
Reckermann M. and Veldhuis M.J.W. (1997) Trophic interactions between picophytoplanktonand micro- and nanozooplankton in the western Arabian Sea during the NE monsoon 1993.Aquatic Microbial Ecology, 12: 263-273.
Reemtsma T. and Ittekkot V. (1992) Determination of factors controlling the fatty-acidcomposition of settling particles in the water column by principal-component analysis and their quantitative assessment by multiple-regression. Organic Geochemistry, 18: 121-129.
Reichart G. (1998) Late Quaternary variability of the Arabian Sea monsoon and oxygenminimum zone. PhD Dissertation, University of Utrecht, 173pp.
Riemann L., Steward G.F., Fandino L.B., Campbell L., Landry M.R., and Azam F. (1999)Bacterial community composition during two consecutive NE Monsoon periods in the Arabian Sea studied by denaturing gradient gel electrophoresis (DGGE) of rRNA genes. Deep-SeaResearch II, 46: 1791-1811.

105
Rixen T., Haake B., Ittekkot V., Guptha M.V.S., Nair R.R., and Schlüssel P. (1996) Couplingbetween SW monsoon-related surface and deep ocean processes as discerned from continuousparticle flux measurements and correlated satellite data. Journal Geophysical Research, 101:28,569-28,582.
Rixen T., Haake B., and Ittekkot V. (2000a) Sedimentation in the western Arabian Sea: The role of coastal and open-ocean upwelling. Deep-Sea Research I, 47: 2155-2178.
Rixen T., Ittekkot V., Haake-Gaye B., and Schäfer P. (2000b) The SW monsoon and itsinfluence on the deep- sea carbon cycle. Deep-Sea Research I, 47: 2629-2651.
Rixen T., Ittekkot V., Haake-Gaye B., and Schäfer P. (2001) Characteristics of the NE and SWmonsoon blooms as indicated by results from long- and short-term sediment trap experimentsin the Arabian Sea. Deep-Sea Research, submitted.
Robinson C. and Williams P.J.LeB. (1999) Plankton and net community production and darkrespiration in the Arabian Sea during and after the 1994 SW monsoon. Deep-Sea Research II,46: 745-766.
Rochford P.A., Kindle J.C., and Gallacher P.A. (2000) Sensitivity of the Arabian Sea mixedlayer to 1994-1995 operational wind products. Journal Geophysical Research, 105(C6): 14141-14162.
Roman M., Smith, S., Wishner, K., Zhang, X.S., and Gowing M. (2000) Mesozooplanktonproduction and grazing in the Arabian Sea. Deep-Sea Research II, 47: 1423-1450.
Rostek F., Bard E., Beaufort L., Sonzogni C., and Ganssen G. (1997) Sea surface temperatureand productivity records for the past 240 kyr in the Arabian Sea. Deep-Sea Research II, 44:1461-1480.
Rudnick D.L., Weller R.A., Eriksen C.C., Dickey T.D., Marra J., and Langdon C. (1997)Moored instruments weather the Arabian Sea, yield data. EOS, Transactions of the AmericanGeophysical Union, 78: 117-121.
Ryabchenko V.A., Gorchakov V.A., and Fasham M.J.R. (1998) Seasonal dynamics andbiological productivity in the Arabian Sea euphotic zone as simulated by a three-dimensionalecosystem model. Global Biogeochemical Cycles, 12: 501-530.
Ryther J.H., Hall J.R., Pease A.K., Bakun A., and Jones M.M. (1966) Primary organicproduction in relation to the chemistry and hydrography of the western Indian Ocean.Limnology and Oceanography, 11: 371-380.
Sabine C.L., Key R.M., Johnson K.M., Millero F.J., Poisson A., Sarmiento J.L., WallaceD.W.R., and Wynn C.D. (1999) Anthropogenic CO2 inventory of the Indian Ocean. GlobalBiogeochemical Cycles, 13: 179-198.
Sabine C.L., Wanninkhof R., Key R.M., Goyet C., and Millero F.J. (2000) Seasonal CO2 fluxes in the tropical and subtropical Indian Ocean. Marine Chemistry, 72: 33-53.
Saraswathy M. and Iyer H. (1986) Ecology of Pleuromamma indica Wolfenden (Copepoda-Calanoida) in the Indian Ocean. Indian Journal of Marine Sciences, 15: 219-222.
Sarin M.M., Rengarajan R., and Krishnaswami S. (1999) Aerosol NO3- and 210Pb distribution
over the central-eastern Arabian Sea and their air-sea deposition fluxes. Tellus, 51B: 749-758.
Sarma V.V.S.S., Dileep Kumar M., and George M.D. (1998) The central and eastern ArabianSea as a perennial source of atmospheric carbon dioxide. Tellus, 50B: 179-184.

106
Sarma V.V.S.S., Dileep Kumar M., Gauns M., and Madhupratap M. (2000) Seasonal controls on surface pCO2 in the central and eastern Arabian Sea. Proceedings Indian Academy of Science, Earth and Planetary Science, 109: 471-479.
Sarma V.V.S.S., Naqvi S.W.A., and Jayakumar D.A. Short-term variability in dissolvedinorganic carbon components in the northern Arabian Sea during NE monsoon: A time seriesapproach. Journal Marine Research, submitted.
Sathyendranath S., Platt T., Caverhill C.M., Warnock R.E., and Lewis M.R. (1989) Remotesensing of oceanic primary production: computations using a spectral model. Deep SeaResearch, 36: 431-453.
Sathyendranath S. and Platt T. (1994) New production and mixed layer dynamics. Proceedingsof the Indian Academy Science, 103: 177-188.
Sathyendranath S., Longhurst A., Caverhill C.M., and Platt T. (1995) Regionally and seasonallydifferentiated primary production in the North Atlantic. Deep Sea Research I, 42: 1773-1802.
Sathyendranath S., Platt T., Stuart V., Irwin B.D., Veldhuis M.J.W., Kraay G.W., and HarrisonW.G. (1996) Some bio-optical characteristics of phytoplankton in the NW Indian Ocean.Marine Ecology Progress Series, 132: 299-311.
Sathyendranath S., Stuart V., Irwin I.R., Maass H., Savidge G., Gilpin L., and Platt T. (1999)Seasonal variations in the bio-optical properties of phytoplankton in the Arabian Sea. Deep-SeaResearch II, 46: 633-653.
Sathyendranath S. and Platt T. (2000) Mixed layer dynamics and primary production in theArabian Sea. In: R. Hanson, H.W. Ducklow and J. Field (Eds.), The Changing Ocean CarbonCycle: A midterm synthesis of the Joint Global Ocean Flux Study. Cambridge University Press, Cambridge, 514pp.
Satyanarana D. and Ramana Y.V. (1994) Distribution of minor elements in the shelf and deep-sea sediments of the northern Arabian Sea. Marine Chemistry, 47: 215-226.
Savidge G. and Gilpin L. (1999) Seasonal influences on size-fractionated chlorophyll aconcentrations and primary production in the northwest Indian Ocean. Deep Sea Research II,46: 701-723.
Schenau S., Slomp C.P., and de Lange G.J. (2000) Phosphogenesis and active phosporiteformation in sediments from the Arabian Sea oxygen minimum zone. Marine Geology, 169: 1-20.
Schmaljohann R., Drews M., Walter S., Linke P., von Rad U., and Imhoff J.F. (2001) Oxygen-minimum zone sediments in the northeastern Arabian Sea off Pakistan: a habitat for thebacterium Thioploca. Marine Ecology-Progress Series, 211: 27-42.
Schnack S. (1982) Feeding habits of Calanoides carinatus (Krøyer) in the upwelling area offnorthwest Africa. Rapports et procès-verbaux des réunions. Conseil permanent internationalpour l’exploration de la Mer, 180: 303-306.
Scholten J.C., Fietzke J., Vogler S., Rutgers van der Loeff M.M., Mangini A., Koeve W.,Waniek J., Stoffers P., Antia A., Kus J. and Neuer S. (2001) Trapping efficiencies of sedimenttraps from the deep eastern Atlantic: the 230Th calibration. Deep-Sea Research, 48: 2383-2408.
Schott F. (1983) Monsoon response of the Somali Current and associated upwelling. Progress in Oceanography, 12: 357-389.
Schott F. and Fischer, J. (2000) Winter monsoon circulation of the northern Arabian Sea andSomali Current. Journal of Geophysical Research, 105: 6359-6376.

107
Schott F. and McCreary J.P. (2001) The monsoon circulation of the Indian Ocean. Progress inOceanography, 51: 1-123.
Schulte S., Mangelsdorf K., and Rullkötter J. (2000) Organic matter preservation on the Pakistan continental margin as revealed by biomarker geochemistry. Organic Geochemistry, 31(10):1005-1022.
SCOR (1995) Arabian Sea Process Study. Scientific Committee on Ocean Research, JGOFSReport 17, 64pp.
Shalapyonok A., Olson R.J., and Shalapyonok L.S. (2001) Arabian Sea phytoplankton duringthe southwest and northeast monsoons 1995: composition, size structure and biomass fromindividual cell properties measured by flow cytometry. Deep Sea Research II, 48: 1231-1261.
Sharada M.K. and Yajnik K.S. (1997) Seasonal variation of chlorophyll and primaryproductivity in central Arabian Sea: A macrocalibrated upper ocean ecosystem model.Proceedings of the Indian Academy of Science, Earth and Planetary Science, 106: 33-42.
Shetye S.R., Gouveia A.D. and Shenoi S.S.C. (1994) Circulation and water masses in theArabian Sea. In: D. Lal (Ed.), Biogeochemistry of the Arabian Sea, Proceedings of the IndianAcademy of Sciences, Earth and Planetary Sciences, 103(2): 9-25.
Shi W., Morrison J.M., Böhm E., and Manghnani V. (1999) Remotely sensed features in the US JGOFS Arabian Sea Process Study. Deep-Sea Research II, 46: 1551-1575.
Shi W., Morrison J.M., Böhm E., and Manghnani V. (2000) The Oman upwelling zone during1993, 1994 and 1995. Deep Sea Research II, 47: 1227-1247.
Smallwood B.J., Wolff G.A., Bett B.J., Smith C.R., Hoover D., Gage J.D., and Patience A.(1999) Megafauna can control the quality of organic matter in marine sediments.Naturwissenschaften, 86: 320-324.
Smallwood B.J. and Wolff G.A. (2000) Molecular characterisation of organic matter insediments underlying the oxygen minimum zone at the Oman Margin, Arabian Sea. Deep-SeaResearch II, 47: 353-375.
Smetacek V.S. (1985) Role of sinking in diatom life-history cycles: Ecological, evolutionary and geological significance. Marine Biology, 84: 239-251.
Smith C., Levin L., Hoover D., McMurtry G., and Gage J. (2000) Variations in bioturbationacross the oxygen minimum zone in the northwest Arabian Sea. Deep-Sea Research II, 47:227-257.
Smith R.L. and Bottero J.S. (1977) On upwelling in the Arabian Sea. In: M. Angel (Ed.), AVoyage of Discovery. Supplement to Deep-Sea Research, Pergamon Press, Oxford, p. 291-304.
Smith S.L. (1982) The northwestern Indian Ocean during the monsoons of 1979: distribution,abundance and feeding of zooplankton. Deep-Sea Research, 29: 1331-1353.
Smith S.L. (1988) Is the Somali Current a biological river in the northwestern Indian Ocean? In: M. Thompson and N. Tirmizi (Eds.), Proceedings of Symposium on Marine Science of theArabian Sea. American Institute of Biological Sciences, Washington, D.C., p. 37-45.
Smith S.L., Banse K., Cochran J.K., Codispoti L.A., Ducklow H.W., Luther M.E., Olson D.B.,Peterson W.T., Prell W.L., Surgi N., Swallow J.C. and Wishner K. (1991) U.S. JGOFS:Arabian Sea Process Study, U.S. JGOFS Planning Report No. 13, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts, 164pp.
Smith S.L. (1992) Secondary production in waters influenced by upwelling off the coast ofSomalia. In: B.N. Desai (Ed.), Oceanography of the Indian Ocean. Oxford and IBH Publishing Co. Ltd., New Delhi, p. 191-200.

108
Smith S.L. (1995) The Arabian Sea: mesozooplankton response to seasonal climate in a tropicalocean. ICES Journal of Marine Science, 52: 427-438.
Smith S.L., Codispoti L.A., Morrison J.M., and Barber R.T. (1998a) The 1994-1996 Arabian Sea Expedition: An integrated, interdisciplinary investigation of the response of the northwesternIndian Ocean to monsoonal forcing. Deep-Sea Research II, 45: 1905-1915.
Smith S., Roman M., Prusova I., Wishner K., Gowing M., Codispoti L., Barber R., Marra J. and Flagg C. (1998b) Seasonal response of zooplankton to monsoonal reversals in the Arabian Sea. Deep-Sea Research II, 45: 2369-2403.
Smith S.L. (2001) Understanding the Arabian Sea: Reflections on the 1994-1996 Arabian SeaExpedition. Deep-Sea Research II, 48: 1385-1402.
Smith S.L., Morrison J.M. and Codispoti L.A. (2002) The role of the oxygen minimum zone inbiogeochemical cycles. In: L. Watts, P. Burkill and S. Smith (Eds.), Arabian Sea ProcessStudy. JGOFS International Planning Office, Bergen, Norway, this volume.
Somasundar K., Rajendran A., Dileep Kumar M., and Sen Gupta R. (1990) Carbon and nitrogen budget of the Arabian Sea. Marine Chemistry, 30: 363-377.
Somayajulu B., Sarin M., and R. Ramesh. (1996) Denitrification in the eastern Arabian Sea:Evaluation of the role of continental margins using Ra isotopes. Deep-Sea Research II, 43:111-117.
Sommer S. and Pfannkuche O. (2000) Metazoan meiofauna of the deep Arabian Sea: standingstocks, size spectra, and regional variability in relation to monsoon induced enhancedsedimentation regimes of particulate organic matter. Deep-Sea Research II, 47: 2957-2977.
Sonzogni C., Bard E., Rostek F., Lafont R., Rosell-Melé A., and Eglinton G. (1997a) Core-topcalibration of the alkenone index vs. sea surface temperature in the Indian Ocean. Deep-SeaResearch II, 44: 1445-1460.
Sonzogni C., Bard E., Rostek F., Dollfus D., Rosell-Melé A., and Eglinton G. (1997b)Temperature and salinity effects on alkenone ratios measured in surface sediments from theIndian Ocean. Quaternary Research, 47: 344-345.
Stelfox C., Burkill P., Edwards E., Harris R., and Sleigh M. (1999) The structure of zooplankton communities, in the 2 to 2000 µm size range, in the Arabian Sea during and after the SWmonsoon, 1994. Deep-Sea Research II, 46: 815-842.
Stephen R. (1992) Copepod composition along southwest and southeast coasts of India. In: B.N. Desai (Ed.), Oceanography of the Indian Ocean. Oxford and IBH Publishing Co. Ltd., NewDelhi, p. 121-127.
Stephen R., Saraladevi K., Meenakshikunjamma P., Gopalakrishnan T., and Saraswathy M.(1992) Calanoid copepods of the International Indian Ocean Expedition collections. In: B.N.Desai (Ed.), Oceanography of the Indian Ocean. Oxford and IBH Publishing Co. Ltd., NewDelhi, p. 143-156.
Stuart V., Sathyendranath S., Head E.J.H., Platt T., Irwin B., and Maass H. (2000) Bio-opticalcharacteristics of diatom and prymnesiophyte populations in the Labrador Sea. Marine Ecology Progress Series, 201: 91-106.
Suess E. (1980) Particulate organic carbon flux in the oceans - surface productivity and oxygen utilization. Nature, 288: 260-263.
Suntharalingam P., Sarmiento J.L., and Toggweiler J.R. (2000) Global significance of nitrous-oxide production and transport from oceanic low-oxygen zones: A modelling study. GlobalBiogeochemical Cycles, 14: 1353-1370.

109
Suthhof A., Jennerjahn T., Rixen T., Schäfer P., Tiemann J., Haake B. and Ittekkot V. (1999)Time-varying fluxes and burial of organic carbon along the continental margins in thenortheastern Arabian Sea. In: Y. Saito, K. Ikehara and H. Katayama (Eds.), Land-Sea Link inAsia, Proceedings of an International Workshop on Sediment Transport and Storage inCoastal Sea-ocean Systems. JAMSTEC and Geological Survey of Japan, Tsukuba, Japan, p.275-280.
Suthhof A., Jennerjahn T.C., Schäfer P., and Ittekkot V. (2000) Nature of organic matter insurface sediments from the Pakistan continental margin and the deep Arabian Sea: amino acids. Deep-Sea Research II, 47(1-2): 329-351.
Swallow, J. (1984) Some aspects of the physical oceanography of the Indian Ocean. Deep-SeaResearch, 31: 639-650.
Swathi P.S., Sharada M.K., and Yajnik, K.S. (2000) A coupled physical-biological-chemicalmodel for the Indian Ocean. Proceedings of the Indian Academy of Science, Earth andPlanetary Science, 109: 503-537.
Tarran G.A., Burkill P.H., Edwards E.S., and Woodward E.M.S. (1999) Phytoplanktoncommunity structure in the Arabian Sea during and after the SW monsoon, 1994. Deep SeaResearch II, 46: 655-676.
Thamban M., Rao V.P., and Raju S.V. (1997) Controls on organic carbon distribution insediments from the eastern Arabian Sea margin. Geo-Marine Letters, 17: 220-227.
Tindale N.W. and Pease P.P. (1999) Aerosols over the Arabian Sea: Atmospheric transportpathways and concentrations of dust and sea salt. Deep-Sea Research II, 46: 1577-1595.
Toole J.M., and Warren B. (1993) A hydrographic section across the subtropical South IndianOcean. Deep-Sea Research I, 40: 1973-2019.
Toon R.K., Lohrenz S.E., Rathburn C.E., Wood A.M., Arnone R.A., Jones B.H., Kindle J.C.,and Weidemann A.D. (2000) Photosynthesis-irradiance parameters and community structureassociated with coastal filaments and adjacent waters in the northern Arabian Sea. Deep SeaResearch II, 47: 1249-1277.
Tranter D. (1977) Further studies of plankton ecosystems in the eastern Indian Ocean. V.Ecology of the Copepoda. Australian Journal of Marine and Freshwater Research, 28: 593-625.
Tréguer P. and Pondaven P. (2000) Global change: Silica control of carbon dioxide. Nature, 406: 358-359.
Turnewitsch R., Witte U., and Graf G. (2000) Bioturbation in the abyssal Arabian Sea: influence of fauna and food supply. Deep-Sea Research II, 47: 2877-2911.
Upstill-Goddard R.C., Barnes J. and Owens N.J.P. (1999) Nitrous oxide and methane during the 1994 SW monsoon in the Arabian Sea/northwestern Indian Ocean. Journal GeophysicalResearch, 104: 30,067-30,084.
Van Couwelaar, M., Angel M. and Madin L. (1997) The distribution and biology of theswimming crab Charybdis smithii McLeay, 1838 (Crustacea; Brachyura: Portunidae) in theNW Indian Ocean. Deep-Sea Research II, 44: 1251-1280.
Van der Weijden C.H., Reichart G.J., and Visser H.J. (1998) Enhanced preservation of organicmatter in sediments deposited within the oxygen minimum zone in the northeastern ArabianSea. Deep-Sea Research I, 46: 807-830.
Van Weering T.C.E., Helder, W., and Schalk P. (1997) The Netherlands Indian Oceanexpedition 1992-1993, first results and an introduction. Deep-Sea Research II, 44: 1177-1193.

110
Vinogradov M. and Voronina N. (1962) Influence of the oxygen deficit on the distribution ofplankton in the Arabian Sea. Okeanologiya, 1: 670-678 (English translation in Deep-SeaResearch, 9: 523-530).
Vinogradov M.E. (1970) Vertical distribution of the oceanic zooplankton. English translation,Israel Program of Scientific Publication 5513, Ketter Press, Jerusalem, 339pp.
Von Rad U., Rösch H, Berner U., Geyh M., Marchig V., and Schulz H. (1996) Authigeniccarbonates derived from oxidized methane vented from the Makran accretionary prism offPakistan. Marine Geology, 136(1-2): 55-77.
Warren B.A., Stommel, H., and Swallow J.C. (1966) Water masses and patterns of flow in theSomali Basin during the southwest monsoon. Deep-Sea Research, 13: 825-860.
Warren B.A. and Johnson G.C. (1992) Deep currents in the Arabian Sea in 1987. MarineGeology, 104: 179-188.
Watts L.J. and Owens N.J.P. (1999) Nitrogen assimilation and the f-ratio in the northwesternIndian Ocean during an intermonsoon period. Deep-Sea Research II, 46: 725-743.
Watts L.J., Sathyendranath S., Caverhill C., Maass H., Platt T., and Owens N.J.P. (1999)Modelling new production in the north west Indian Ocean region. Marine Ecology Progressseries, 183: 1-12.
Weisse T. (1999) Bacterivory in the northwestern Indian Ocean during the intermonsoonnortheast monsoon period. Deep-Sea Research II, 46: 795-814.
Weller R.A., Baumgartner M.A., Josey S.A., Fischer A.S. and Kindle J.C. (1998) Atmosphericforcing in the Arabian Sea during 1994-1995: observations and comparisons with climatologyand models. Deep Sea Research II, 45: 1961-1999.
Wiebe P. (1988) Functional regression equations for zooplankton displacement volume, wetweight, dry weight, and carbon: a correction. Fishery Bulletin, 86: 833-835.
Wiebinga C.J. and Veldhuis M.J.W. (1997) Abundance and productivity of bacterioplankton inrelation to seasonal upwelling in the northwest Indian Ocean. Deep-Sea Research I, 44: 451-476.
Wiggert J.D., Jones B.H., Dickey T.D., Brink K.H., Weller R.A., Marra J., and Codispoti L.A.(2000) The northeast monsoon’s impact on mixing, phytoplankton biomass and nutrientcycling in the Arabian Sea. Deep-Sea Research II, 47: 1353-1385.
Wiggert J.D., Murtugudde R.G., and McClain C.R. (2002) Processes controlling interannualvariations in wintertime primary productivity in the central Arabian Sea. Deep-Sea Research II, in press.
Wishner K.F., Gowing M.M., and Gelfman C. (1998) Mesozooplankton biomass in the upper1000 m in the Arabian Sea: overall seasonal and geographic patterns, and relationship tooxygen gradients. Deep-Sea Research II, 45: 2405-2432.
Wishner K., M. Gowing and Gelfman C. (2000) Living in suboxia: Ecology of an Arabian Seaoxygen minimum zone copepod. Limnology and Oceanography, 45: 1576-1593.
Witte U. (2000) Vertical distribution of metazoan macrofauna within the sediment at four siteswith contrasting food supply in the deep Arabian Sea. Deep-Sea Research II, 47: 2979-2997.
Witte U. and Pfannkuche O. (2000) High rates of benthic carbon remineralisation in the abyssal Arabian Sea. Deep-Sea Research II, 47: 2785-2804.
Witter E., Lewis B., and Luther III G. (2000) Iron speciation in the Arabian Sea. Deep-SeaResearch II, 47: 1517-1539.

111
Woodward E.M.S., Rees A.P. and Stephen J.A. (1999) The influence of the SW monsoon uponthe nutrient biogeochemistry of the Arabian Sea. Deep-Sea Research II, 46: 571-591.
Wyrtki K. (1971) Oceanographic Atlas of the International Indian Ocean Expedition. NationalScience Foundation, Washington, D.C., 531pp.
Yajnik K.S. and Sharada M.K. (1992) Dynamics of a basic phytoplankton model. In: B.N. Desai (Ed.), Oceanography of the Indian Ocean. Oxford and IBH Publishing Co. Ltd., New Delhi, p. 91-98.
Yakushev E.V. and Neretin L.N. (1997) One-dimensional modeling of nitrogen and sulfur cycles in the aphotic zones of the Black and Arabian Seas. Global Biogeochemical Cycles, 11(3):401-414.
Yoder J.A., McClain C.R., Feldman G.C. and Esaias W.E. (1993) Annual cycles ofphytoplankton chlorophyll concentrations in the global ocean: a satellite view. GlobalBiogeochemical Cycles, 7: 181-194.
You Y. and Tomczak M. (1993) Thermocline circulation and ventilation in the Indian Oceanderived from water mass analysis. Deep-Sea Research, 40: 13-56.
You Y. (1997) Seasonal variations of thermocline circulation and ventilation in the IndianOcean. Journal of Geophysical Research, 103: 10,391-10,422.
Young D.K. and Kindle J.C. (1994) Physical processes affecting availability of dissolved silicate for diatom production in the Arabian Sea. Journal of Geophysical Research, 99: 22619-22632.
Yu E.F., Francois R., Bacon M.P., Honjo S., Fleer A.P., Manganini S.J., Rutgers van der LoeffM.M. and Ittekkot V. (2001) Trapping efficiency of bottom-tethered sediment traps estimatedfrom the intercepted fluxes of 230 Th and 231Pa. Deep-Sea Research I, 48: 865-889.
Zonneveld K.A.F. (1997) Dynoflagellate cyst distribution in surface sediment from the ArabianSea (northwestern Indian Ocean) in relation to temperature and salinity gradients in the upperwater column. Deep-Sea Research II, 44: 1411-1443.

112
APPENDIX 4.3
Seminal Literature:1971-Present
1971. Wyrtki K. Oceanographic Atlas of the International Indian Ocean Expedition. NationalScience Foundation, Washington, D.C., 531pp.
1973. Zeitzschel B. (Ed.) The Biology of the Indian Ocean, Ecological Studies 3. Proceedingsfrom symposium held at Kiel University, Germany, March 31 – April 6, 1971. Published bySpringer-Verlag, Berlin, Heidelberg, New York, 549pp. ISBN 3-540-06004-9.
1976. Krey J. and Babenerd B. Phytoplankton Production Atlas of the International IndianOcean Expedition. Intergovernmental Oceanographic Commission, Paris, 70pp.
1977. Proceedings of the Symposium on Warm Water Zooplankton. Published by the NationalInstitute of Oceanography and Casa J.D. Fernandes, Goa, India, 722pp.
1984. Angel M. (Ed.) Marine Science of the North-west Indian Ocean and Adjacent Waters.Deep-Sea Research Volume 31, No.6-8. Published by Pergamon Press, London. ISBN 0 08030285 8.
1988. Thompson M.F. and Tirmizi N. Marine Science of the Arabian Sea: Proceedings of anInternational Conference. Published by the American Institute of Biological Sciences,Washington, D.C., 658pp.
1991. Smith S.L., K. Banse, J.K. Cochran, L.A. Codispoti, H.W. Ducklow, M.E. Luther, D.B.Olson, W.T. Peterson, W.L. Prell, N. Surgi, J.C. Swallow and K. Wishner. U.S. JGOFS: ArabianSea Process Study. U.S. JGOFS Planning Report No.13, Woods Hole Oceanographic Institution, Woods Hole, Massachusetts, 164pp.
1992. Desai B.N. (Ed.) Oceanography of the Indian Ocean. A collection of peer-reviewedpapers presented at the International Symposium on the Oceanography of The Indian Ocean,National Institute of Oceanography, Goa, India, January 1991. Published by Oxford and IBHPublishing Co. PVT. Ltd., New Delhi, India, 772pp. ISBN 81-204-0735-0.
1993. Ittekkot V. and Nair R.R. Monsoon Biogeochemistry. SCOPE/UNEP Sonderband Heft 76, 192pp. ISSN 0072-1115.
1993. Topical Studies in Oceanography. Biogeochemical Cycling in the Northwestern IndianOcean. Deep Sea Research II Volume 40, No. 3, 1993. Ed. J.D. Milliman, Guest Eds. P.H.Burkill, R.F.C. Mantoura and N.J.P. Owens. Published by Pergamon Press, London. ISSN 0967-0645.
1994. Baars M. (Ed.) Monsoons and Pelagic Systems. Cruise Reports from the NetherlandsIndian Ocean Programme Volume 1. Published by the National Museum of Natural History,Leiden. ISBN 90-73239-24-9.
1994. Lal D. (Ed.) Biogeochemistry of the Arabian Sea. Published by the Indian Academy ofScience, Bangalore, 560 080, India, 253pp.

113
1994. Biogeochemical Processes in the Arabian Sea. Proceedings from the US-CIS Arabian Sea Workshop, Sevastopol, Crimea, Ukraine, September 20-25, 1993. Published by Woods HoleOceanographic Institution, Marine Hydrophysical Institute, U.S. National Science Foundation.Edited by The Marine Hydrophysical Institute of National Academy of Sciences of the Ukraine.
1995. Thompson M.F. and Tirmizi N. (Eds.). The Arabian Sea: Living Marine Resources andthe Environment. Published by A.A. Balkema, Rotterdam, Netherlands, 730pp.
1997. Topical Studies in Oceanography. Netherlands Indian Ocean Program. (1992-1993) FirstResults. Deep Sea Research II Volume 44, No. 6-7. Ed. J.D. Milliman, Guest Eds. T.C.E. VanWeering, W. Helder and P. Schalk. Published by Pergamon Press, London. ISSN 0967-0645.
1998. Sherman K., Okemwa E.N., and Ntiba M.J. (Eds.). Large Marine Ecosystems of the IndianOcean, Assessment, Sustainability and Management. Published by Blackwell Science Inc.,Malden, MA, 394pp. ISBN 0 632 04318 0.
1998. Topical Studies in Oceanography. The 1994-1996 Arabian Sea Expedition: OceanicResponse to Monsoonal Forcing, Part 1. Deep Sea Research II Volume 45, No. 10-11. Ed. J.D.Milliman, Guest Ed. S.L. Smith. Published by Pergamon Press, London. ISSN 0967-0645.
1999. Topical Studies in Oceanography. ARABESQUE: UK JGOFS Process Studies in theArabian Sea. Deep Sea Research II Volume 46, No. 3-4, 1999. Ed. J.D. Milliman, Guest Ed.P.H. Burkill. Published by Pergamon Press, London. ISSN 0967-0645.
1999. Topical Studies in Oceanography. The 1994-1996 Arabian Sea Expedition: OceanicResponse to Monsoonal Forcing, Part 2. Deep Sea Research II Volume 46, No. 8-9. Ed. J.D.Milliman, Guest Ed. S.L. Smith. Published by Pergamon Press, London. ISSN 0967-0645.
2000. Gaur V.K. (Ed.). Proceedings of the Indian Academy of Sciences. Earth and PlanetarySciences. Special Issue on the International JGOFS Symposium on Biogeochemistry of theArabian Sea. Vol. 109, No.4. Published by Brilliant Printers Pvt. Ltd., Bangalore, India. ISSN0253-4126.
2000. Topical Studies in Oceanography. Benthic Processes in the Deep Arabian Sea. Deep SeaResearch II Volume 47, No. 1-2. Ed. J.D. Milliman, Guest Eds. J.D. Gage, L.A. Levin and G.A. Wolff. Published by Pergamon Press, London. ISSN 0967-0645.
2000. Topical Studies in Oceanography. The 1994-1996 Arabian Sea Expedition: OceanicResponse to Monsoonal Forcing, Part 3. Deep Sea Research II Volume 47, No. 7-8. Ed. J.D.Milliman, Guest Ed. S.L. Smith. Published by Pergamon Press, London. ISSN 0967-0645.
2000. Topical Studies in Oceanography. The Biogeochemistry of the Deep Arabian Sea. DeepSea Research II Volume 47, No. 14. Ed. J.D. Milliman, Guest Eds. O. Pfannkuche and K.Lochte. Published by Pergamon Press, London. ISSN 0967-0645.
2001. Topical Studies in Oceanography. The 1994-1996 Arabian Sea Expedition: OceanicResponse to Monsoonal Forcing, Part 4. Deep Sea Research II Volume 48, No. 6-7. Ed. J.D.Milliman, Guest Ed. S.L. Smith. Published by Pergamon Press, London. ISSN 0967-0645.

The JGOFS Report Series includes the following:
1 Report of the Second Session of the SCOR Committee for JGOFS. The Hague, September 19882 Report of the Third Session of the SCOR Committee for JGOFS. Honolulu, September 19893 Report of the JGOFS Pacific Planning Workshop. Honolulu, September 19894 JGOFS North Atlantic Bloom Experiment: Report of the First Data Workshop. Kiel, March 19905 Science Plan. August 19906 JGOFS Core Measurement Protocols: Reports of the Core Measurement Working Groups7 JGOFS North Atlantic Bloom Experiment, International Scientific Symposium Abstracts. Washington, November 19908 Report of the International Workshop on Equatorial Pacific Process Studies. Tokyo, April 19909 JGOFS Implementation Plan. (also published as IGBP Report No. 23) September 1992
10 The JGOFS Southern Ocean Study11 The Reports of JGOFS meetings held in Taipei, October 1992: Seventh Meeting of the JGOFS Scientific Steering
Committee; Global Synthesis in JGOFS - A Round Table Discussion; JGOFS Scientific and Organizational Issues in the Asian Region - Report of a Workshop; JGOFS/LOICZ Continental Margins Task Team - Report of the First Meeting. March 1993
12 Report of the Second Meeting of the JGOFS North Atlantic Planning Group13 The Reports of JGOFS meetings held in Carqueiranne, France, September 1993: Eighth Meeting of the JGOFS
Scientific Steering Committee; JGOFS Southern Ocean Planning Group - Report for 1992/93; Measurement of the Parameters of Photosynthesis - A Report from the JGOFS Photosynthesis Measurement Task Team. March 1994
14 Biogeochemical Ocean-Atmosphere Transfers. A paper for JGOFS and IGAC by Ronald Prinn, Peter Liss and Patrick Buat-Ménard. March 1994
15 Report of the JGOFS/LOICZ Task Team on Continental Margin Studies. April 199416 Report of the Ninth Meeting of the JGOFS Scientific Steering Committee, Victoria, B.C. Canada, October 1994 and The
Report of the JGOFS Southern Ocean Planning Group for 1993/9417 JGOFS Arabian Sea Process Study. March 199518 Joint Global Ocean Flux Study: Publications, 1988-1995. April 199519 Protocols for the Joint Global Ocean Flux studies (JGOFS) core measurements (reprint). June, 199620 Remote Sensing in the JGOFS programme. September 199621 First report of the JGOFS/LOICZ Continental Margins Task Team. October 199622 Report on the International Workshop on Continental Shelf Fluxes of Carbon, Nitrogen and Phosphorus. 199623 One-Dimensional models of water column biogeochemistry. Report of a workshop held in Toulouse, France, November-
December 1995. February 199724 Joint Global Ocean Flux Study: Publications, 1988-1996. October 199725 JGOFS/LOICZ Workshop on Non-Conservative Fluxes in the Continental Margins. October 1997.26 Report of the JGOFS/LOICZ Continental Margins Task Team Meeting, No 2. October 199727 Parameters of photosynthesis: definitions, theory and interpretation of results. August 199828 Eleventh meeting of the JGOFS SSC; Twelfth meeting of the JGOFS SSC; and the Second meeting of the North Pacific
Task Team. November 199829 JGOFS Data Management and Synthesis Workshop, 25-27 Sept. 1998, Bergen, Norway. Meeting Minutes. January 199930 Publications 1988-1999. January 200031 Thirteenth meeting of the JGOFS Scientific Steering Committee. Fourteenth meeting of the JGOFS Scientific Steering
Committee. Fifteenth meeting of the JGOFS Scientific Steering Committee. October 200132 Meeting of the Southern Ocean Synthesis Group, Year 1998. October 2001.33 Joint IGBP EU-US Meeting on the Ocean Component of an integrated Carbon Cycle Science Framework. October 200134 First Meeting of the North Atlantic Synthesis Group, 1998; Second Meeting of the North Atlantic Synthesis Group,
1999; Third Meeting of the North Atlantic Synthesis Group, 2001. October 2001
The following reports were published by SCOR in 1987 - 1989 prior to the establishment of the JGOFS Report Series:
• The Joint Global Ocean Flux Study: Background, Goals, Organizations, and Next Steps. Report of the International Scientific Planning and Coordination Meeting for Global Ocean Flux Studies. Sponsored by SCOR. Held at ICSU Headquarters, Paris, 17-19 February 1987
• North Atlantic Planning Workshop. Paris, 7-11 September 1987• SCOR Committee for the Joint Global Ocean Flux Study. Report of the First Session. Miami, January 1988• Report of the First Meeting of the JGOFS Pilot Study Cruise Coordinating Committee. Plymouth, UK, April 1988• Report of the JGOFS Working Group on Data Management. Bedford Institute of Oceanography, September 1988





























