Embed Size (px)
Citation preview

Hindawi Publishing CorporationInternational Journal of GenomicsVolume 2013 Article ID 383024 12 pageshttpdxdoiorg1011552013383024
Research ArticleA Complex Network of MicroRNAs Expressed in Brain andGenes Associated with Amyotrophic Lateral Sclerosis
Santosh Shinde Neelima Arora and Utpal Bhadra
Functional Genomics and Gene Silencing Group Centre for Cellular and Molecular Biology Hyderabad 500007 India
Correspondence should be addressed to Utpal Bhadra utpalccmbresin
Received 10 January 2013 Accepted 5 June 2013
Academic Editor John Parkinson
Copyright copy 2013 Santosh Shinde et alThis is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Amyotrophic Lateral Sclerosis (ALS) is a rare neurological disease affecting mainly motor neurons and often leads to paralysisand death in extreme cases For exploring the role of microRNAs in genes regulation in ALS disease miRanda was employed forprediction of target sites of miRNAs expressed in various parts of brain and CNS on 35 genes associated with ALS Similar searchwas conducted using TargetScan and PicTar for prediction of target sites in 31015840 UTR only 1456 target sites were predicted usingmiRanda and more target sites were found in 51015840 UTR and CDS region as compared to 31015840 UTR 11 target sites were predicted to becommon by all the algorithms and thus these represent the most significant sites Target site hotspots were identified and wererecognized as hotspots for multiple miRNAs action thus acting as favoured sites of action for the repression of gene expressionThe complex interplay of genes and miRNAs brought about by multiplicity and cooperativity was explored This investigation willaid in elucidating the mechanism of action of miRNAs for the considered genes The intrinsic network of miRNAs expressed innervous system and genes associated with ALSmay provide rapid and effective outcome for therapeutic applications and diagnosis
1 Introduction
A complete coordination of brain signals and their functionalexecution in different body parts is often found in multi-cellular organisms However mechanism underlying suchcoordination between brain and body is still a mystery Howa trivial defect in the coordinated communication can leadto a disease has been a subject of study for many yearsNeurological disorders are responsible for major proportionof the global burden of disease [1] The mammoth rise inincidence of neurological disorders has led to initiatives fordeclaration of global epidemic With the significant rise innumber of cases of neurological disorders across the globeit is postulated that no particular geographical region or agegroup is immune to these disorders Among these neurode-generative disorders that are triggered by environmental andgenetic factors are major cause of concern for aged popula-tion Neurodegenerative diseases such as Alzheimerrsquos diseaseParkinsonrsquos disease prion diseases Amyotrophic LateralSclerosis Huntingtonrsquos disease and various spinocerebellarataxias account for significant morbidity Progressive andslow degeneration of neurons as witnessed in these diseases
leads to neuron dysfunctionThe escalating cost of treatmentand financial burden in terms of loss in disability adjusted lifeyears (DALYs) along with personal trauma and social stigmawarrants the need for exploring new alternatives and novelways for combating these diseases
Since its description in 1869 by French neurologistCharcot [2] ALS has become a well-documented motorneuron disease (MND) Amyotrophic Lateral Sclerosis (ALS)is referred to by different names such as motor neurondisease (MND) Charcotrsquos disease and Lou Gehrigrsquos diseaseALS affects motor neurons that are involved in controllingmuscle movement and the disease is characterized by gener-alized weakness muscle atrophy and progressive muscularparalysis reflecting degeneration of motor neurons in theprimary motor cortex brain stem and spinal cord [3 4]Often sidelined as an orphan disease owing to a small numberof cases worldwide ALS has high lifetime risk despite its lowfrequency [5]
Many studies have established that the disease patternhas a gender bias as men are more prone to ALS [6ndash10] Itis also reported that several races are more susceptible toALS incidence [11] ALS is a fatal disorder often manifesting
2 International Journal of Genomics
itself at a late age Till date there is no tangible treatmentplan available for ALS [12] Despite extensive research onALS exact mechanism of ALS continues to be a mystery asmultiple processes like protein aggregation ex-cytotoxicityoxidative stress and mitochondrial malfunction inflamma-tion and defects in axonal transport are proposed [13] About90ofALS cases have got no genetic link (sporadic)while therest are familial [14]Though highly heterogeneous in natureclinical symptoms are often relied on for disease diagnosisand degree of severity depends on the number of motorneurons affected But more often it is regarded as a diseasewith obscure pathogenesis due to vast spectrum of clinicalindications Several genes expressed in different pathways areimplicated in manifestation of ALS
It has not been long since the discovery of involve-ment of a tiny stretch of 21ndash23 endogenous ribonucleotidescalled ldquomiRNArdquo in gene regulation It is predicted that aconsiderable proportion of human genes is controlled bythe miRNAs [15] MicroRNAs often propounded as beingldquomicro in size macro in functionrdquo [16] have been found tobe implicated in various vital processes The wide range ofprocesses which are reported to be regulated by miRNAsare cell differentiation development stress resistance [17]and several processes in neurodegenerative disorders [18ndash20] Growing evidence about the role of microRNAs inneurodegenerative disorders has incited innumerable studiesacross the globe Present era has witnessed a revolution ininformation technology and the avalanche of informationarising from high throughput methods has warranted theneed for employing in silico methods for obtaining fastsimple and accurate results Many computational algorithmsand tools available for prediction of miRNA and their targetsites have led to unfathomable progress in this field [21 22]Although computational analysis suffers from drawbacks likehigh signal to noise ratio yet high speed of prediction andanalysis makes it advantageous for initial screening Besidesbeing skill intensive and time consuming the conventionalmethods used for biochemical profiling ofmiRNA expressionare also marred by problems arising due to transient andlow level of microRNA expression specificity to tissue typeand complex interaction of miRNAs and their targets [23]Annotation and analysis of sites of action of miRNAs provideimportant clues with regard to their function and modeof action Comprehensive information about multifariousassociation of miRNAs with genes of ALS pathway is notreadily available Ample number of miRNAs expressed inbrain and central nervous system anticipated to play a partin regulation of coordination of nervous system adds a layerof complexity in gene regulation Here we identified targetsof microRNAs predominantly expressed in brain on thegene associated with ALS A diagram depicting a complexinteraction between microRNAs and genes was constructedwhich will aid in deciphering the role of miRNAs in ALSetiology This study offers sufficient information regardingthe regulation of ALS genes which represents first effort forsimplification of microRNA-based control of ALS genes Italso sheds ample light on the complex interplay of genes andtheir targeted miRNAs
2 Materials and Methods
In current study we emphasize on identification of fractionof miRNA targets specific to the pathway involved in ALSand propose an interaction network between these genes andmiRNA Figure 1 represents stepwise workflow of the study
21 Selection of Genes Genes involved in ALS manifestationcontribute in a unique pathway in a synchronized mannerand are referred to in KEGG (Kyoto Encyclopedia of Genesand Genomes) [24] pathway The KEGG pathway database isan excellent repository that provides biochemical and geneticinformation related to pathways in a sequential manner inform of diagrams depicting associations molecular interac-tions and reactions KEGG (httpwwwgenomejpkegg)was explored for information on genes involved in ALSThe ALS pathway represented in KEGG consists of 35 genespresent on different chromosomes
The genes considered were SLC1A2 solute carrier fam-ily 1 (glial high affinity glutamate transporter) member 2GRIA1 TNF tumor necrosis factor tumor necrosis factorreceptor superfamily member 1A caspase 1 superoxide dis-mutase 1 tumor protein p53 CHP calcium binding proteinP22 BID BH3 interacting domain death agonist TOMM40translocase of outer mitochondrial membrane 40 homologBCL2 B-cell CLLlymphoma 2 BCL2-like 1 BAX BCL2-associated X protein BAD BCL2-associated agonist of celldeath cytochrome c somatic apoptotic peptidase activatingfactor 1 caspase 9 caspase 3 Der1-like domain family member1 MAP3K5 DAXX death-domain associated protein MAP2K3 MAP2K6 MAPK14 nitric oxide synthase 1 copper chaper-one for superoxide dismutase glutathione peroxidase 1 cata-lase neurofilament light polypeptide NEFM neurofilamentmedium polypeptide NEFH neurofilament heavy polypeptideperipherin Amyotrophic Lateral Sclerosis 2 RAB5A and ras-related C3 botulinum toxin substrate 1GPX1 and RAB5Aare located on chromosome 3 while TNF MAP3K5 DAXXdeath-domain associated protein and MAPK14 are locatedon chromosome 6 CYCS and RAC1 are positioned onchromosome 7 and DERL1 NEFL and NEFM are sitedon chromosome 8 Five genes namely SLC1A2 CASP1BAD CCS and CAT are present on chromosome 11 whileTNFRSF1A APAF1NOS1 and PRPH reside on chromosome12 Detailed information of each gene is summarized inTable S1 (in Supplementary Material Available online athttpdxdoiorg1011552013383024) Gene sequences wereretrieved in Fasta format (a widely acceptable format) frompublic database NCBI
22 Selection of miRNA Dataset Information regarding spe-cific microRNAs expressed in midbrain cerebellum hip-pocampus and frontal cortex of human was collected fromthe Human microRNA and Disease Database (HMDD)http20238126151hmddmirnamd) (mir2disease database (httpwwwmir2diseaseorg) and other relevant ref-erences Mature miRNA sequences were collected frommiR-base (httpwwwmirbaseorg) [25]Thenumber ofmiRNAsexpressed in cerebellum hippocampusmidbrain and frontal
International Journal of Genomics 3
35 genes involved in ALS selected from KEGG pathway database
Exploration of databases for information on nervous system specific miRNA sequences
Retrieval of considered gene sequences from NCBI in Fasta format
Collection of mature miRNA sequences expressed in nervous system from miRbase
Prediction of target sites using miRanda TargetScan and PicTar
Comparison of results for finding common target sites
Calculation of multiplicity and cooperativity
Construction of interaction map
Figure 1 Schematic representation of the workflow
cortex available in public repository is 62 93 112 and 105respectively Out of these fewmiRNAs are not only limited toone region but are expressed in other parts also Expressionof 23 miRNAs is limited only to midbrain while 17 miRNAsare expressed in hippocampus The number of miRNAs thatare expressed only in frontal cortex and cerebellum is 14 and4 respectively
The concept of miRNA target sites finds applicationin functional annotation miRanda (version 10 b) avail-able at httpwwwmicrornaorgmicrornareleaseNotesdoa dynamic programming algorithm that relies on thesequence complementarity [26 27] was employed for predic-tion of target sites ofmiRNAs expressed in nervous system onthe selected genes miRanda allots higher weights to matchesat the 51015840 end of the mature miRNA while considering thefree energy of the RNA-RNA duplex [28] and the degree ofconservation of the miRNA targets across related genomesThe analysis was performed keeping the following cut-offvalues for prediction of target sites Gap Open Penalty 20Gap Extend 800 match score (119878) ge 15000 duplex freeenergy (Δ119866) = minus2500 kcalmol and scaling parameter(119908) = 300 The selected gene sequences and maturemiRNAs expressed in human midbrain cerebellum hip-pocampus and frontal cortex were used as reference andquery sequences respectively as input to miRanda
Positions of the target sites for these miRNAs on 35genes were explored and sites prone to multiple miRNAswere identified as hot spots Multiplicity and cooperativitywere determined on the basis of miRNA and targeted geneinteractions After assembling the data for all genes miRNAswere selected based on high multiplicity A complex inter-action network among selected genes and top miRNAs (aselucidated by the algorithm) was constructed
23 Prediction of Common miRNA Targets by MultiplePrograms Target sites of microRNA predicted solely bymiRanda were further verified using TargetScan (Release 51)[29] The predicted miRNA target binding sites in the 31015840UTR were also compared using PicTar [30] which predictsmiRNA target sites based on the conservationmiRNA targetsonly in 31015840 UTR that are common in these miRNA predictionalgorithms and their comparative analysis are provided inTable 4 Based on the intersection of results microRNAswereselected and multiplicity and cooperativity were determined
3 Results and Discussion
Delicate balance of gene expression involved in a trait ismaintained by employing different regulatory mechanismsOne of the important processes is microRNA-mediated generegulation which basically fine-tunes the expression of genesessential for the pathway Prediction of miRNA target sitesusing bioinformatics approach offers noteworthy advantagessuch as sequence specificity rapid selection and screeningproceduresThismethodology can be exploited for the recog-nition of molecular hallmarks of different disease traits But amajorweakness of in silico analyses is the lack of experimentalvalidation For circumventing such problem a combinatorialapproach can be implementedwhere target sites are predictedusing multiple programs This methodology can verify thepotential targets of microRNAs Further the sites of miRNAaction will aid in identifying the stage of regulation ofgenes associated with ALS that can be effectively utilized fortherapeutic applications
Out of miRNAs that are expressed in midbrain cere-bellum hippocampus and frontal cortex majority of them
4 International Journal of Genomics
are from midbrain (Figure 2) while nearly equal proportionsof microRNAs are expressed in hippocampus and frontalcortex The microRNAs in cerebellum are less in numbercompared to miRNAs expressed in other regions considered(Figure 2) The target sites of the microRNA expressed indifferent parts of the brain were predicted on the 35 genesrelated to ALS for initial screening The result obtained frommiRanda (version 10) indicates that 477 target sites werepredicted for miRNAs expressed in midbrain as comparedto the target sites (411) predicted for miRNAs expressedin hippocampus However total target sites predicted formiRNAs expressed in cerebellum and frontal cortexes were175 and 395 respectively The number of targets for miRNAsexpressed in cerebellum is considerably less as compared tobinding sites for miRNAs expressed in other areas of thebrain Surprisingly microRNA targets were distributed in thedifferent regions of the genes and are not limited to only 31015840UTR Total 1456 target sites were predicted using miRandafor miRNAs considered in the study Majority of them werefound in CDS Target sites in the 31015840 untranslated region iscomparatively less However a significant number of targetswere also found in the 51015840 UTR of the genes (Figure 3) Earlierthe occurrence of miRNA target sites in coding region and 51015840UTR of genes was considered as exception in animals Butnumerous recent studies have established that microRNAscan target different regions of the gene and microRNA-based regulation is not confined only to 31015840 UTR [31ndash35]Experiments aimed at identifying specific parameter or factorfor effective targeting of 31015840 UTR by miRNAs have failedconsiderably Conversely the existence of miRNA target sitesin ORF of Nanog OCT4 and SOX1 during induction ofstem cell pluripotency [36] supports the existence of miRNAbinding sites in other sites of the genes in animals Theseresults established the notion that microRNA target sitesare not restricted to 31015840 UTR in animals Such variability inregions also adds cues for anticipating the mode of actionspecially their role in transcriptional posttranscriptionaland translation inhibition As the majority of the algorithmsdeveloped for miRNA target site prediction considers only31015840 UTR verification of 31015840 UTR target site prediction using acombinatorial approach will be useful
Target sites on ALS genes were classified according tothe microRNAs expressed in four different regions of brainMaximum target sites were predicted in the CDS of all thegenes considered However for the miRNAs expressed inmidbrain hippocampus and frontal cortex the number oftargets in the coding region is threefold greater than thenumber of miRNA targets found in 51015840 UTR and 31015840 UTRbut in miRNAs expressed in cerebellum the number oftargets in CDS was about twofold levels as compared to otherregions of the genes As there were less miRNAs expressedin cerebellum this can explain small number of target sitespredicted on genes associated to ALS Figure 4 shows thedistribution of collective target sites of miRNAs expressed inbrain in the various regions of the selected genes
Next we analyzed the microRNA targets in each geneassociated with the ALS pathways No target sites could bedetected in 31015840 UTR for miRNAs expressed in midbrain forCASP1 GRIA1 GPX1 DAXX CAT SOD1 NEFM NEFL and
Midbrain 33
Frontal cortex 27
Hippocampus 28
Cerebellum 12
MidbrainFrontal cortex
HippocampusCerebellum
Figure 2 Percentage of miRNAs expressed in various parts of thebrain
0100200300400500600700800900
CDS 3998400UTR5998400UTR
Figure 3 Distribution of target sites in different regions of selectedgenes for the selected miRNAs
85
46
83 85
277
83
222 222
115
46
10488
0
50
100
150
200
250
300
Midbrain Cerebellum Hippocampus Frontal cortex
CDS3998400UTR
5998400UTR
Figure 4 Distribution of target sites in 51015840 UTR CDS and 31015840 UTRfor miRNAs expressed in various parts of nervous system
International Journal of Genomics 5
NEFH genes Similarly CDS of two genesCYCS and RAB5Ahave no potential targets Conversely CDS of MAP3K5have maximum number of targets for miRNAs expressedin midbrain On the other hand 51015840 UTR lack any targetsites of same miRNAs in CASP1 GRIA1 GPX1 DAXX CATSOD1NEFMNEFLNEFHRAC1 andTNF genesThe geneshaving maximum and minimum number of target sites werealso identified The miRNAs expressed in midbrain showedthe least number of target sites (3) in RAC1 and CASP1genes while 29 target sites were predicted in NOS1 Similarresults were obtained for miRNAs expressed in cerebellumfor the same genes For instance RAC1 has only 1 targetsite whereas NOS1 has 11 target sites For miRNAs expressedin hippocampus CASP1 and GRIA1 genes have minimumnumber of predicted sites (3) and MAP3K5 was predictedto possess maximum target sites (33) Similarly CASP1 andSOD1 showed 2 target sites each while 23 target sites werepredicted for TNFRSF1A for miRNAs expressed in frontalcortex On the contrary target sites could not be predictedin CDS for 6 genes namely GPX1 CYCS CHP CASP3SOD1 and RAB5A for miRNAs expressed in cerebellumNo target sites could be predicted in 31015840 UTR of ALS2 BIDCASP1 GRIA1 DAXX CCS CAT CASP9 TOMM40 TNFSOD1 RAC1 NEFM and NEFL genes No target site couldbe predicted in BCL2 CASP1 GRIA1 DAXX CYCS CASP3TNF RAC1 NEFM and NEFH genes for miRNAs expressedin cerebellum These results indicate that the majority ofthe ALS genes lack any target site for miRNAs expressed incerebellum Target sites predicted using miRanda are sum-marized in Table S2 (in Supplementary Material) miRNAtargets were found in most of the genes but surprisingly fewgenesCASP1GRIA1DAXXCAT SOD1 andNEFM lack anymiRNA target site in their 31015840 UTR
31 Hotspot Identification MicroRNAs either share the samecomplementary target sites or proximally located sites adja-cent to other miRNA target site A hot spot can be defined asa stretch of sequence that is prone to action of a group ofmiR-NAs Many studies have revealed that usually microRNAshaving the same origin are regulated and expressed togetherin time and space [37] Such miRNAs play extensive rolein many vital processes and carry out important biologicalfunctions in different pathways [38ndash40] It is quite possiblethat all the miRNA occupying position in ldquohotspotrdquo maynot be equally effective for controlling the same gene Basedon competitive selection predominant miRNA may outcastother miRNAs and get potentially involved in repression ofother microRNA or in similar events But exact mechanismbehind such selection remains unknown It will be interestingto elucidate the factors responsible for variable effectivenessof microRNAs Various researchers have used different stan-dards andparameters such as the same chromosomal locationfor more than two miRNAs the same orientation phyloge-netic relationships and absence of interfering transcriptionunit [41 42] As miRNAs are presumed to be implicatedin important biological processes miRNA targets predictedusing miRanda were then further analyzed for identifyingmiRNA-prone regions in all selected genes We defined
a region as hotspot or region prone tomultiplemiRNA actionif the region showed minimum 10 nucleotides overlap fromthe starting position and concurrence of 3 miRNA targetsTotal 11 miRNAs-prone regions were identified in 35 genesassociated to ALS (Figure SF1-37 in SupplementaryMaterial)CYCS and MAP2K3 showed 1 hotspot each in 31015840 UTR whilesingle target site hotspot was found in 51015840 UTR in 4 genesthat is BAX DERL1 SLC1A2 and RAB5A Two such regionswere found in CDS of CASP9 and 3 other genes namelyTNF MAPK14 and MAP2K3 showed the occurrence ofsingle region vulnerable to multiple miRNAs (Table S7 inSupplementary Material)
In miRNA cluster prediction 2 hot spots were identifiedin MAP2K3 The hotspot in CDS was found to have targetsites for 3 miRNAsthat is hsa-mir-323-5p hsa-mir-654-5p and hsa-mir-433 (Figure 5) Target sites for 3 miRNAsnamely hsa-mir-107 hsa-mir-423-5p and hsa-mir-103 arecongregated in another hotspot found in 31015840 UTR
MAP2K3 having 2 such hotspots is expected to showmore sensitivity towards gene regulation by microRNAsOccurrence of such small number of hot spots indicates thepossible specificity of miRNAs and stringency of regulationThe variability in existence of such target site hotspotsenhances the complexity in gene regulation These resultssuggest that any region in a gene namely 51015840 UTR CDS and31015840 UTR can be targeted by the microRNAs as opposed tothe earlier belief which postulated 31015840 UTR as the only site ofaction of microRNAs
32 Multifarious Relationships among Gene and miRNAsMicroRNA-mediated regulation offers a whole new dimen-sion and often researchers are left confounded by sheercomplexity of its level Not akin to the earlier incidencesof one miRNA targeting one gene distribution of targetsites for the miRNAs points out various possibilities wheremultiplicity and cooperativity come into picture In principleone miRNA can target more than one gene (multiplicity)and one gene can be controlled by more than one miRNA(cooperativity) [26] Multiplicity is a property arising fromrelaxed base pairing As reported earlier single miRNAcan control hundreds of genes Distribution of target siteson genes is rarely uniform and these target sites are oftenfound scattered across a region and range of occurrenceis also known to vary in numbers in a gene [27] Oftenpresence of more target sites per gene indicates efficientregulation In this study we found multiple target sites inmost of the genes After assembling all data multiplicity andcooperativity were calculated From these analyses selectivemiRNAs that displayed maximum number of interactionswith the genes associated with manifestation of ALS wereused for generation of an interaction network (Table 1) Weobserved that hsa-miR-370 showed maximum multiplicityas it showed 65 interactions with 30 gene sequences whileno target site could be predicted in 5 genes namely CASP1GPX1DERL1CCS and SOD1Thus it is expected that degreeof repression by hsa-mir-370 will be considerably high Nexthsa-miR-874 displayed 52 interactions with 29 genes but didnot show any target in CASP1 CHP SOD1 RAC1 RAB5A
6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 International Journal of Genomics
itself at a late age Till date there is no tangible treatmentplan available for ALS [12] Despite extensive research onALS exact mechanism of ALS continues to be a mystery asmultiple processes like protein aggregation ex-cytotoxicityoxidative stress and mitochondrial malfunction inflamma-tion and defects in axonal transport are proposed [13] About90ofALS cases have got no genetic link (sporadic)while therest are familial [14]Though highly heterogeneous in natureclinical symptoms are often relied on for disease diagnosisand degree of severity depends on the number of motorneurons affected But more often it is regarded as a diseasewith obscure pathogenesis due to vast spectrum of clinicalindications Several genes expressed in different pathways areimplicated in manifestation of ALS
It has not been long since the discovery of involve-ment of a tiny stretch of 21ndash23 endogenous ribonucleotidescalled ldquomiRNArdquo in gene regulation It is predicted that aconsiderable proportion of human genes is controlled bythe miRNAs [15] MicroRNAs often propounded as beingldquomicro in size macro in functionrdquo [16] have been found tobe implicated in various vital processes The wide range ofprocesses which are reported to be regulated by miRNAsare cell differentiation development stress resistance [17]and several processes in neurodegenerative disorders [18ndash20] Growing evidence about the role of microRNAs inneurodegenerative disorders has incited innumerable studiesacross the globe Present era has witnessed a revolution ininformation technology and the avalanche of informationarising from high throughput methods has warranted theneed for employing in silico methods for obtaining fastsimple and accurate results Many computational algorithmsand tools available for prediction of miRNA and their targetsites have led to unfathomable progress in this field [21 22]Although computational analysis suffers from drawbacks likehigh signal to noise ratio yet high speed of prediction andanalysis makes it advantageous for initial screening Besidesbeing skill intensive and time consuming the conventionalmethods used for biochemical profiling ofmiRNA expressionare also marred by problems arising due to transient andlow level of microRNA expression specificity to tissue typeand complex interaction of miRNAs and their targets [23]Annotation and analysis of sites of action of miRNAs provideimportant clues with regard to their function and modeof action Comprehensive information about multifariousassociation of miRNAs with genes of ALS pathway is notreadily available Ample number of miRNAs expressed inbrain and central nervous system anticipated to play a partin regulation of coordination of nervous system adds a layerof complexity in gene regulation Here we identified targetsof microRNAs predominantly expressed in brain on thegene associated with ALS A diagram depicting a complexinteraction between microRNAs and genes was constructedwhich will aid in deciphering the role of miRNAs in ALSetiology This study offers sufficient information regardingthe regulation of ALS genes which represents first effort forsimplification of microRNA-based control of ALS genes Italso sheds ample light on the complex interplay of genes andtheir targeted miRNAs
2 Materials and Methods
In current study we emphasize on identification of fractionof miRNA targets specific to the pathway involved in ALSand propose an interaction network between these genes andmiRNA Figure 1 represents stepwise workflow of the study
21 Selection of Genes Genes involved in ALS manifestationcontribute in a unique pathway in a synchronized mannerand are referred to in KEGG (Kyoto Encyclopedia of Genesand Genomes) [24] pathway The KEGG pathway database isan excellent repository that provides biochemical and geneticinformation related to pathways in a sequential manner inform of diagrams depicting associations molecular interac-tions and reactions KEGG (httpwwwgenomejpkegg)was explored for information on genes involved in ALSThe ALS pathway represented in KEGG consists of 35 genespresent on different chromosomes
The genes considered were SLC1A2 solute carrier fam-ily 1 (glial high affinity glutamate transporter) member 2GRIA1 TNF tumor necrosis factor tumor necrosis factorreceptor superfamily member 1A caspase 1 superoxide dis-mutase 1 tumor protein p53 CHP calcium binding proteinP22 BID BH3 interacting domain death agonist TOMM40translocase of outer mitochondrial membrane 40 homologBCL2 B-cell CLLlymphoma 2 BCL2-like 1 BAX BCL2-associated X protein BAD BCL2-associated agonist of celldeath cytochrome c somatic apoptotic peptidase activatingfactor 1 caspase 9 caspase 3 Der1-like domain family member1 MAP3K5 DAXX death-domain associated protein MAP2K3 MAP2K6 MAPK14 nitric oxide synthase 1 copper chaper-one for superoxide dismutase glutathione peroxidase 1 cata-lase neurofilament light polypeptide NEFM neurofilamentmedium polypeptide NEFH neurofilament heavy polypeptideperipherin Amyotrophic Lateral Sclerosis 2 RAB5A and ras-related C3 botulinum toxin substrate 1GPX1 and RAB5Aare located on chromosome 3 while TNF MAP3K5 DAXXdeath-domain associated protein and MAPK14 are locatedon chromosome 6 CYCS and RAC1 are positioned onchromosome 7 and DERL1 NEFL and NEFM are sitedon chromosome 8 Five genes namely SLC1A2 CASP1BAD CCS and CAT are present on chromosome 11 whileTNFRSF1A APAF1NOS1 and PRPH reside on chromosome12 Detailed information of each gene is summarized inTable S1 (in Supplementary Material Available online athttpdxdoiorg1011552013383024) Gene sequences wereretrieved in Fasta format (a widely acceptable format) frompublic database NCBI
22 Selection of miRNA Dataset Information regarding spe-cific microRNAs expressed in midbrain cerebellum hip-pocampus and frontal cortex of human was collected fromthe Human microRNA and Disease Database (HMDD)http20238126151hmddmirnamd) (mir2disease database (httpwwwmir2diseaseorg) and other relevant ref-erences Mature miRNA sequences were collected frommiR-base (httpwwwmirbaseorg) [25]Thenumber ofmiRNAsexpressed in cerebellum hippocampusmidbrain and frontal
International Journal of Genomics 3
35 genes involved in ALS selected from KEGG pathway database
Exploration of databases for information on nervous system specific miRNA sequences
Retrieval of considered gene sequences from NCBI in Fasta format
Collection of mature miRNA sequences expressed in nervous system from miRbase
Prediction of target sites using miRanda TargetScan and PicTar
Comparison of results for finding common target sites
Calculation of multiplicity and cooperativity
Construction of interaction map
Figure 1 Schematic representation of the workflow
cortex available in public repository is 62 93 112 and 105respectively Out of these fewmiRNAs are not only limited toone region but are expressed in other parts also Expressionof 23 miRNAs is limited only to midbrain while 17 miRNAsare expressed in hippocampus The number of miRNAs thatare expressed only in frontal cortex and cerebellum is 14 and4 respectively
The concept of miRNA target sites finds applicationin functional annotation miRanda (version 10 b) avail-able at httpwwwmicrornaorgmicrornareleaseNotesdoa dynamic programming algorithm that relies on thesequence complementarity [26 27] was employed for predic-tion of target sites ofmiRNAs expressed in nervous system onthe selected genes miRanda allots higher weights to matchesat the 51015840 end of the mature miRNA while considering thefree energy of the RNA-RNA duplex [28] and the degree ofconservation of the miRNA targets across related genomesThe analysis was performed keeping the following cut-offvalues for prediction of target sites Gap Open Penalty 20Gap Extend 800 match score (119878) ge 15000 duplex freeenergy (Δ119866) = minus2500 kcalmol and scaling parameter(119908) = 300 The selected gene sequences and maturemiRNAs expressed in human midbrain cerebellum hip-pocampus and frontal cortex were used as reference andquery sequences respectively as input to miRanda
Positions of the target sites for these miRNAs on 35genes were explored and sites prone to multiple miRNAswere identified as hot spots Multiplicity and cooperativitywere determined on the basis of miRNA and targeted geneinteractions After assembling the data for all genes miRNAswere selected based on high multiplicity A complex inter-action network among selected genes and top miRNAs (aselucidated by the algorithm) was constructed
23 Prediction of Common miRNA Targets by MultiplePrograms Target sites of microRNA predicted solely bymiRanda were further verified using TargetScan (Release 51)[29] The predicted miRNA target binding sites in the 31015840UTR were also compared using PicTar [30] which predictsmiRNA target sites based on the conservationmiRNA targetsonly in 31015840 UTR that are common in these miRNA predictionalgorithms and their comparative analysis are provided inTable 4 Based on the intersection of results microRNAswereselected and multiplicity and cooperativity were determined
3 Results and Discussion
Delicate balance of gene expression involved in a trait ismaintained by employing different regulatory mechanismsOne of the important processes is microRNA-mediated generegulation which basically fine-tunes the expression of genesessential for the pathway Prediction of miRNA target sitesusing bioinformatics approach offers noteworthy advantagessuch as sequence specificity rapid selection and screeningproceduresThismethodology can be exploited for the recog-nition of molecular hallmarks of different disease traits But amajorweakness of in silico analyses is the lack of experimentalvalidation For circumventing such problem a combinatorialapproach can be implementedwhere target sites are predictedusing multiple programs This methodology can verify thepotential targets of microRNAs Further the sites of miRNAaction will aid in identifying the stage of regulation ofgenes associated with ALS that can be effectively utilized fortherapeutic applications
Out of miRNAs that are expressed in midbrain cere-bellum hippocampus and frontal cortex majority of them
4 International Journal of Genomics
are from midbrain (Figure 2) while nearly equal proportionsof microRNAs are expressed in hippocampus and frontalcortex The microRNAs in cerebellum are less in numbercompared to miRNAs expressed in other regions considered(Figure 2) The target sites of the microRNA expressed indifferent parts of the brain were predicted on the 35 genesrelated to ALS for initial screening The result obtained frommiRanda (version 10) indicates that 477 target sites werepredicted for miRNAs expressed in midbrain as comparedto the target sites (411) predicted for miRNAs expressedin hippocampus However total target sites predicted formiRNAs expressed in cerebellum and frontal cortexes were175 and 395 respectively The number of targets for miRNAsexpressed in cerebellum is considerably less as compared tobinding sites for miRNAs expressed in other areas of thebrain Surprisingly microRNA targets were distributed in thedifferent regions of the genes and are not limited to only 31015840UTR Total 1456 target sites were predicted using miRandafor miRNAs considered in the study Majority of them werefound in CDS Target sites in the 31015840 untranslated region iscomparatively less However a significant number of targetswere also found in the 51015840 UTR of the genes (Figure 3) Earlierthe occurrence of miRNA target sites in coding region and 51015840UTR of genes was considered as exception in animals Butnumerous recent studies have established that microRNAscan target different regions of the gene and microRNA-based regulation is not confined only to 31015840 UTR [31ndash35]Experiments aimed at identifying specific parameter or factorfor effective targeting of 31015840 UTR by miRNAs have failedconsiderably Conversely the existence of miRNA target sitesin ORF of Nanog OCT4 and SOX1 during induction ofstem cell pluripotency [36] supports the existence of miRNAbinding sites in other sites of the genes in animals Theseresults established the notion that microRNA target sitesare not restricted to 31015840 UTR in animals Such variability inregions also adds cues for anticipating the mode of actionspecially their role in transcriptional posttranscriptionaland translation inhibition As the majority of the algorithmsdeveloped for miRNA target site prediction considers only31015840 UTR verification of 31015840 UTR target site prediction using acombinatorial approach will be useful
Target sites on ALS genes were classified according tothe microRNAs expressed in four different regions of brainMaximum target sites were predicted in the CDS of all thegenes considered However for the miRNAs expressed inmidbrain hippocampus and frontal cortex the number oftargets in the coding region is threefold greater than thenumber of miRNA targets found in 51015840 UTR and 31015840 UTRbut in miRNAs expressed in cerebellum the number oftargets in CDS was about twofold levels as compared to otherregions of the genes As there were less miRNAs expressedin cerebellum this can explain small number of target sitespredicted on genes associated to ALS Figure 4 shows thedistribution of collective target sites of miRNAs expressed inbrain in the various regions of the selected genes
Next we analyzed the microRNA targets in each geneassociated with the ALS pathways No target sites could bedetected in 31015840 UTR for miRNAs expressed in midbrain forCASP1 GRIA1 GPX1 DAXX CAT SOD1 NEFM NEFL and
Midbrain 33
Frontal cortex 27
Hippocampus 28
Cerebellum 12
MidbrainFrontal cortex
HippocampusCerebellum
Figure 2 Percentage of miRNAs expressed in various parts of thebrain
0100200300400500600700800900
CDS 3998400UTR5998400UTR
Figure 3 Distribution of target sites in different regions of selectedgenes for the selected miRNAs
85
46
83 85
277
83
222 222
115
46
10488
0
50
100
150
200
250
300
Midbrain Cerebellum Hippocampus Frontal cortex
CDS3998400UTR
5998400UTR
Figure 4 Distribution of target sites in 51015840 UTR CDS and 31015840 UTRfor miRNAs expressed in various parts of nervous system
International Journal of Genomics 5
NEFH genes Similarly CDS of two genesCYCS and RAB5Ahave no potential targets Conversely CDS of MAP3K5have maximum number of targets for miRNAs expressedin midbrain On the other hand 51015840 UTR lack any targetsites of same miRNAs in CASP1 GRIA1 GPX1 DAXX CATSOD1NEFMNEFLNEFHRAC1 andTNF genesThe geneshaving maximum and minimum number of target sites werealso identified The miRNAs expressed in midbrain showedthe least number of target sites (3) in RAC1 and CASP1genes while 29 target sites were predicted in NOS1 Similarresults were obtained for miRNAs expressed in cerebellumfor the same genes For instance RAC1 has only 1 targetsite whereas NOS1 has 11 target sites For miRNAs expressedin hippocampus CASP1 and GRIA1 genes have minimumnumber of predicted sites (3) and MAP3K5 was predictedto possess maximum target sites (33) Similarly CASP1 andSOD1 showed 2 target sites each while 23 target sites werepredicted for TNFRSF1A for miRNAs expressed in frontalcortex On the contrary target sites could not be predictedin CDS for 6 genes namely GPX1 CYCS CHP CASP3SOD1 and RAB5A for miRNAs expressed in cerebellumNo target sites could be predicted in 31015840 UTR of ALS2 BIDCASP1 GRIA1 DAXX CCS CAT CASP9 TOMM40 TNFSOD1 RAC1 NEFM and NEFL genes No target site couldbe predicted in BCL2 CASP1 GRIA1 DAXX CYCS CASP3TNF RAC1 NEFM and NEFH genes for miRNAs expressedin cerebellum These results indicate that the majority ofthe ALS genes lack any target site for miRNAs expressed incerebellum Target sites predicted using miRanda are sum-marized in Table S2 (in Supplementary Material) miRNAtargets were found in most of the genes but surprisingly fewgenesCASP1GRIA1DAXXCAT SOD1 andNEFM lack anymiRNA target site in their 31015840 UTR
31 Hotspot Identification MicroRNAs either share the samecomplementary target sites or proximally located sites adja-cent to other miRNA target site A hot spot can be defined asa stretch of sequence that is prone to action of a group ofmiR-NAs Many studies have revealed that usually microRNAshaving the same origin are regulated and expressed togetherin time and space [37] Such miRNAs play extensive rolein many vital processes and carry out important biologicalfunctions in different pathways [38ndash40] It is quite possiblethat all the miRNA occupying position in ldquohotspotrdquo maynot be equally effective for controlling the same gene Basedon competitive selection predominant miRNA may outcastother miRNAs and get potentially involved in repression ofother microRNA or in similar events But exact mechanismbehind such selection remains unknown It will be interestingto elucidate the factors responsible for variable effectivenessof microRNAs Various researchers have used different stan-dards andparameters such as the same chromosomal locationfor more than two miRNAs the same orientation phyloge-netic relationships and absence of interfering transcriptionunit [41 42] As miRNAs are presumed to be implicatedin important biological processes miRNA targets predictedusing miRanda were then further analyzed for identifyingmiRNA-prone regions in all selected genes We defined
a region as hotspot or region prone tomultiplemiRNA actionif the region showed minimum 10 nucleotides overlap fromthe starting position and concurrence of 3 miRNA targetsTotal 11 miRNAs-prone regions were identified in 35 genesassociated to ALS (Figure SF1-37 in SupplementaryMaterial)CYCS and MAP2K3 showed 1 hotspot each in 31015840 UTR whilesingle target site hotspot was found in 51015840 UTR in 4 genesthat is BAX DERL1 SLC1A2 and RAB5A Two such regionswere found in CDS of CASP9 and 3 other genes namelyTNF MAPK14 and MAP2K3 showed the occurrence ofsingle region vulnerable to multiple miRNAs (Table S7 inSupplementary Material)
In miRNA cluster prediction 2 hot spots were identifiedin MAP2K3 The hotspot in CDS was found to have targetsites for 3 miRNAsthat is hsa-mir-323-5p hsa-mir-654-5p and hsa-mir-433 (Figure 5) Target sites for 3 miRNAsnamely hsa-mir-107 hsa-mir-423-5p and hsa-mir-103 arecongregated in another hotspot found in 31015840 UTR
MAP2K3 having 2 such hotspots is expected to showmore sensitivity towards gene regulation by microRNAsOccurrence of such small number of hot spots indicates thepossible specificity of miRNAs and stringency of regulationThe variability in existence of such target site hotspotsenhances the complexity in gene regulation These resultssuggest that any region in a gene namely 51015840 UTR CDS and31015840 UTR can be targeted by the microRNAs as opposed tothe earlier belief which postulated 31015840 UTR as the only site ofaction of microRNAs
32 Multifarious Relationships among Gene and miRNAsMicroRNA-mediated regulation offers a whole new dimen-sion and often researchers are left confounded by sheercomplexity of its level Not akin to the earlier incidencesof one miRNA targeting one gene distribution of targetsites for the miRNAs points out various possibilities wheremultiplicity and cooperativity come into picture In principleone miRNA can target more than one gene (multiplicity)and one gene can be controlled by more than one miRNA(cooperativity) [26] Multiplicity is a property arising fromrelaxed base pairing As reported earlier single miRNAcan control hundreds of genes Distribution of target siteson genes is rarely uniform and these target sites are oftenfound scattered across a region and range of occurrenceis also known to vary in numbers in a gene [27] Oftenpresence of more target sites per gene indicates efficientregulation In this study we found multiple target sites inmost of the genes After assembling all data multiplicity andcooperativity were calculated From these analyses selectivemiRNAs that displayed maximum number of interactionswith the genes associated with manifestation of ALS wereused for generation of an interaction network (Table 1) Weobserved that hsa-miR-370 showed maximum multiplicityas it showed 65 interactions with 30 gene sequences whileno target site could be predicted in 5 genes namely CASP1GPX1DERL1CCS and SOD1Thus it is expected that degreeof repression by hsa-mir-370 will be considerably high Nexthsa-miR-874 displayed 52 interactions with 29 genes but didnot show any target in CASP1 CHP SOD1 RAC1 RAB5A
6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 3
35 genes involved in ALS selected from KEGG pathway database
Exploration of databases for information on nervous system specific miRNA sequences
Retrieval of considered gene sequences from NCBI in Fasta format
Collection of mature miRNA sequences expressed in nervous system from miRbase
Prediction of target sites using miRanda TargetScan and PicTar
Comparison of results for finding common target sites
Calculation of multiplicity and cooperativity
Construction of interaction map
Figure 1 Schematic representation of the workflow
cortex available in public repository is 62 93 112 and 105respectively Out of these fewmiRNAs are not only limited toone region but are expressed in other parts also Expressionof 23 miRNAs is limited only to midbrain while 17 miRNAsare expressed in hippocampus The number of miRNAs thatare expressed only in frontal cortex and cerebellum is 14 and4 respectively
The concept of miRNA target sites finds applicationin functional annotation miRanda (version 10 b) avail-able at httpwwwmicrornaorgmicrornareleaseNotesdoa dynamic programming algorithm that relies on thesequence complementarity [26 27] was employed for predic-tion of target sites ofmiRNAs expressed in nervous system onthe selected genes miRanda allots higher weights to matchesat the 51015840 end of the mature miRNA while considering thefree energy of the RNA-RNA duplex [28] and the degree ofconservation of the miRNA targets across related genomesThe analysis was performed keeping the following cut-offvalues for prediction of target sites Gap Open Penalty 20Gap Extend 800 match score (119878) ge 15000 duplex freeenergy (Δ119866) = minus2500 kcalmol and scaling parameter(119908) = 300 The selected gene sequences and maturemiRNAs expressed in human midbrain cerebellum hip-pocampus and frontal cortex were used as reference andquery sequences respectively as input to miRanda
Positions of the target sites for these miRNAs on 35genes were explored and sites prone to multiple miRNAswere identified as hot spots Multiplicity and cooperativitywere determined on the basis of miRNA and targeted geneinteractions After assembling the data for all genes miRNAswere selected based on high multiplicity A complex inter-action network among selected genes and top miRNAs (aselucidated by the algorithm) was constructed
23 Prediction of Common miRNA Targets by MultiplePrograms Target sites of microRNA predicted solely bymiRanda were further verified using TargetScan (Release 51)[29] The predicted miRNA target binding sites in the 31015840UTR were also compared using PicTar [30] which predictsmiRNA target sites based on the conservationmiRNA targetsonly in 31015840 UTR that are common in these miRNA predictionalgorithms and their comparative analysis are provided inTable 4 Based on the intersection of results microRNAswereselected and multiplicity and cooperativity were determined
3 Results and Discussion
Delicate balance of gene expression involved in a trait ismaintained by employing different regulatory mechanismsOne of the important processes is microRNA-mediated generegulation which basically fine-tunes the expression of genesessential for the pathway Prediction of miRNA target sitesusing bioinformatics approach offers noteworthy advantagessuch as sequence specificity rapid selection and screeningproceduresThismethodology can be exploited for the recog-nition of molecular hallmarks of different disease traits But amajorweakness of in silico analyses is the lack of experimentalvalidation For circumventing such problem a combinatorialapproach can be implementedwhere target sites are predictedusing multiple programs This methodology can verify thepotential targets of microRNAs Further the sites of miRNAaction will aid in identifying the stage of regulation ofgenes associated with ALS that can be effectively utilized fortherapeutic applications
Out of miRNAs that are expressed in midbrain cere-bellum hippocampus and frontal cortex majority of them
4 International Journal of Genomics
are from midbrain (Figure 2) while nearly equal proportionsof microRNAs are expressed in hippocampus and frontalcortex The microRNAs in cerebellum are less in numbercompared to miRNAs expressed in other regions considered(Figure 2) The target sites of the microRNA expressed indifferent parts of the brain were predicted on the 35 genesrelated to ALS for initial screening The result obtained frommiRanda (version 10) indicates that 477 target sites werepredicted for miRNAs expressed in midbrain as comparedto the target sites (411) predicted for miRNAs expressedin hippocampus However total target sites predicted formiRNAs expressed in cerebellum and frontal cortexes were175 and 395 respectively The number of targets for miRNAsexpressed in cerebellum is considerably less as compared tobinding sites for miRNAs expressed in other areas of thebrain Surprisingly microRNA targets were distributed in thedifferent regions of the genes and are not limited to only 31015840UTR Total 1456 target sites were predicted using miRandafor miRNAs considered in the study Majority of them werefound in CDS Target sites in the 31015840 untranslated region iscomparatively less However a significant number of targetswere also found in the 51015840 UTR of the genes (Figure 3) Earlierthe occurrence of miRNA target sites in coding region and 51015840UTR of genes was considered as exception in animals Butnumerous recent studies have established that microRNAscan target different regions of the gene and microRNA-based regulation is not confined only to 31015840 UTR [31ndash35]Experiments aimed at identifying specific parameter or factorfor effective targeting of 31015840 UTR by miRNAs have failedconsiderably Conversely the existence of miRNA target sitesin ORF of Nanog OCT4 and SOX1 during induction ofstem cell pluripotency [36] supports the existence of miRNAbinding sites in other sites of the genes in animals Theseresults established the notion that microRNA target sitesare not restricted to 31015840 UTR in animals Such variability inregions also adds cues for anticipating the mode of actionspecially their role in transcriptional posttranscriptionaland translation inhibition As the majority of the algorithmsdeveloped for miRNA target site prediction considers only31015840 UTR verification of 31015840 UTR target site prediction using acombinatorial approach will be useful
Target sites on ALS genes were classified according tothe microRNAs expressed in four different regions of brainMaximum target sites were predicted in the CDS of all thegenes considered However for the miRNAs expressed inmidbrain hippocampus and frontal cortex the number oftargets in the coding region is threefold greater than thenumber of miRNA targets found in 51015840 UTR and 31015840 UTRbut in miRNAs expressed in cerebellum the number oftargets in CDS was about twofold levels as compared to otherregions of the genes As there were less miRNAs expressedin cerebellum this can explain small number of target sitespredicted on genes associated to ALS Figure 4 shows thedistribution of collective target sites of miRNAs expressed inbrain in the various regions of the selected genes
Next we analyzed the microRNA targets in each geneassociated with the ALS pathways No target sites could bedetected in 31015840 UTR for miRNAs expressed in midbrain forCASP1 GRIA1 GPX1 DAXX CAT SOD1 NEFM NEFL and
Midbrain 33
Frontal cortex 27
Hippocampus 28
Cerebellum 12
MidbrainFrontal cortex
HippocampusCerebellum
Figure 2 Percentage of miRNAs expressed in various parts of thebrain
0100200300400500600700800900
CDS 3998400UTR5998400UTR
Figure 3 Distribution of target sites in different regions of selectedgenes for the selected miRNAs
85
46
83 85
277
83
222 222
115
46
10488
0
50
100
150
200
250
300
Midbrain Cerebellum Hippocampus Frontal cortex
CDS3998400UTR
5998400UTR
Figure 4 Distribution of target sites in 51015840 UTR CDS and 31015840 UTRfor miRNAs expressed in various parts of nervous system
International Journal of Genomics 5
NEFH genes Similarly CDS of two genesCYCS and RAB5Ahave no potential targets Conversely CDS of MAP3K5have maximum number of targets for miRNAs expressedin midbrain On the other hand 51015840 UTR lack any targetsites of same miRNAs in CASP1 GRIA1 GPX1 DAXX CATSOD1NEFMNEFLNEFHRAC1 andTNF genesThe geneshaving maximum and minimum number of target sites werealso identified The miRNAs expressed in midbrain showedthe least number of target sites (3) in RAC1 and CASP1genes while 29 target sites were predicted in NOS1 Similarresults were obtained for miRNAs expressed in cerebellumfor the same genes For instance RAC1 has only 1 targetsite whereas NOS1 has 11 target sites For miRNAs expressedin hippocampus CASP1 and GRIA1 genes have minimumnumber of predicted sites (3) and MAP3K5 was predictedto possess maximum target sites (33) Similarly CASP1 andSOD1 showed 2 target sites each while 23 target sites werepredicted for TNFRSF1A for miRNAs expressed in frontalcortex On the contrary target sites could not be predictedin CDS for 6 genes namely GPX1 CYCS CHP CASP3SOD1 and RAB5A for miRNAs expressed in cerebellumNo target sites could be predicted in 31015840 UTR of ALS2 BIDCASP1 GRIA1 DAXX CCS CAT CASP9 TOMM40 TNFSOD1 RAC1 NEFM and NEFL genes No target site couldbe predicted in BCL2 CASP1 GRIA1 DAXX CYCS CASP3TNF RAC1 NEFM and NEFH genes for miRNAs expressedin cerebellum These results indicate that the majority ofthe ALS genes lack any target site for miRNAs expressed incerebellum Target sites predicted using miRanda are sum-marized in Table S2 (in Supplementary Material) miRNAtargets were found in most of the genes but surprisingly fewgenesCASP1GRIA1DAXXCAT SOD1 andNEFM lack anymiRNA target site in their 31015840 UTR
31 Hotspot Identification MicroRNAs either share the samecomplementary target sites or proximally located sites adja-cent to other miRNA target site A hot spot can be defined asa stretch of sequence that is prone to action of a group ofmiR-NAs Many studies have revealed that usually microRNAshaving the same origin are regulated and expressed togetherin time and space [37] Such miRNAs play extensive rolein many vital processes and carry out important biologicalfunctions in different pathways [38ndash40] It is quite possiblethat all the miRNA occupying position in ldquohotspotrdquo maynot be equally effective for controlling the same gene Basedon competitive selection predominant miRNA may outcastother miRNAs and get potentially involved in repression ofother microRNA or in similar events But exact mechanismbehind such selection remains unknown It will be interestingto elucidate the factors responsible for variable effectivenessof microRNAs Various researchers have used different stan-dards andparameters such as the same chromosomal locationfor more than two miRNAs the same orientation phyloge-netic relationships and absence of interfering transcriptionunit [41 42] As miRNAs are presumed to be implicatedin important biological processes miRNA targets predictedusing miRanda were then further analyzed for identifyingmiRNA-prone regions in all selected genes We defined
a region as hotspot or region prone tomultiplemiRNA actionif the region showed minimum 10 nucleotides overlap fromthe starting position and concurrence of 3 miRNA targetsTotal 11 miRNAs-prone regions were identified in 35 genesassociated to ALS (Figure SF1-37 in SupplementaryMaterial)CYCS and MAP2K3 showed 1 hotspot each in 31015840 UTR whilesingle target site hotspot was found in 51015840 UTR in 4 genesthat is BAX DERL1 SLC1A2 and RAB5A Two such regionswere found in CDS of CASP9 and 3 other genes namelyTNF MAPK14 and MAP2K3 showed the occurrence ofsingle region vulnerable to multiple miRNAs (Table S7 inSupplementary Material)
In miRNA cluster prediction 2 hot spots were identifiedin MAP2K3 The hotspot in CDS was found to have targetsites for 3 miRNAsthat is hsa-mir-323-5p hsa-mir-654-5p and hsa-mir-433 (Figure 5) Target sites for 3 miRNAsnamely hsa-mir-107 hsa-mir-423-5p and hsa-mir-103 arecongregated in another hotspot found in 31015840 UTR
MAP2K3 having 2 such hotspots is expected to showmore sensitivity towards gene regulation by microRNAsOccurrence of such small number of hot spots indicates thepossible specificity of miRNAs and stringency of regulationThe variability in existence of such target site hotspotsenhances the complexity in gene regulation These resultssuggest that any region in a gene namely 51015840 UTR CDS and31015840 UTR can be targeted by the microRNAs as opposed tothe earlier belief which postulated 31015840 UTR as the only site ofaction of microRNAs
32 Multifarious Relationships among Gene and miRNAsMicroRNA-mediated regulation offers a whole new dimen-sion and often researchers are left confounded by sheercomplexity of its level Not akin to the earlier incidencesof one miRNA targeting one gene distribution of targetsites for the miRNAs points out various possibilities wheremultiplicity and cooperativity come into picture In principleone miRNA can target more than one gene (multiplicity)and one gene can be controlled by more than one miRNA(cooperativity) [26] Multiplicity is a property arising fromrelaxed base pairing As reported earlier single miRNAcan control hundreds of genes Distribution of target siteson genes is rarely uniform and these target sites are oftenfound scattered across a region and range of occurrenceis also known to vary in numbers in a gene [27] Oftenpresence of more target sites per gene indicates efficientregulation In this study we found multiple target sites inmost of the genes After assembling all data multiplicity andcooperativity were calculated From these analyses selectivemiRNAs that displayed maximum number of interactionswith the genes associated with manifestation of ALS wereused for generation of an interaction network (Table 1) Weobserved that hsa-miR-370 showed maximum multiplicityas it showed 65 interactions with 30 gene sequences whileno target site could be predicted in 5 genes namely CASP1GPX1DERL1CCS and SOD1Thus it is expected that degreeof repression by hsa-mir-370 will be considerably high Nexthsa-miR-874 displayed 52 interactions with 29 genes but didnot show any target in CASP1 CHP SOD1 RAC1 RAB5A
6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
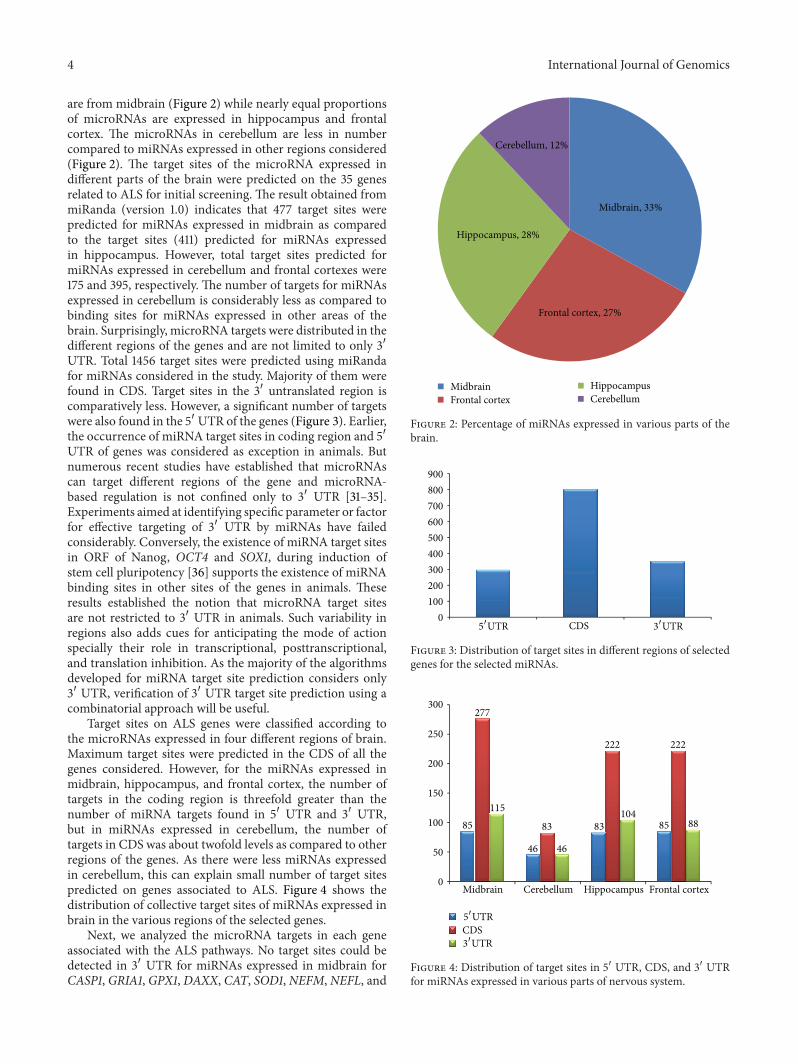
4 International Journal of Genomics
are from midbrain (Figure 2) while nearly equal proportionsof microRNAs are expressed in hippocampus and frontalcortex The microRNAs in cerebellum are less in numbercompared to miRNAs expressed in other regions considered(Figure 2) The target sites of the microRNA expressed indifferent parts of the brain were predicted on the 35 genesrelated to ALS for initial screening The result obtained frommiRanda (version 10) indicates that 477 target sites werepredicted for miRNAs expressed in midbrain as comparedto the target sites (411) predicted for miRNAs expressedin hippocampus However total target sites predicted formiRNAs expressed in cerebellum and frontal cortexes were175 and 395 respectively The number of targets for miRNAsexpressed in cerebellum is considerably less as compared tobinding sites for miRNAs expressed in other areas of thebrain Surprisingly microRNA targets were distributed in thedifferent regions of the genes and are not limited to only 31015840UTR Total 1456 target sites were predicted using miRandafor miRNAs considered in the study Majority of them werefound in CDS Target sites in the 31015840 untranslated region iscomparatively less However a significant number of targetswere also found in the 51015840 UTR of the genes (Figure 3) Earlierthe occurrence of miRNA target sites in coding region and 51015840UTR of genes was considered as exception in animals Butnumerous recent studies have established that microRNAscan target different regions of the gene and microRNA-based regulation is not confined only to 31015840 UTR [31ndash35]Experiments aimed at identifying specific parameter or factorfor effective targeting of 31015840 UTR by miRNAs have failedconsiderably Conversely the existence of miRNA target sitesin ORF of Nanog OCT4 and SOX1 during induction ofstem cell pluripotency [36] supports the existence of miRNAbinding sites in other sites of the genes in animals Theseresults established the notion that microRNA target sitesare not restricted to 31015840 UTR in animals Such variability inregions also adds cues for anticipating the mode of actionspecially their role in transcriptional posttranscriptionaland translation inhibition As the majority of the algorithmsdeveloped for miRNA target site prediction considers only31015840 UTR verification of 31015840 UTR target site prediction using acombinatorial approach will be useful
Target sites on ALS genes were classified according tothe microRNAs expressed in four different regions of brainMaximum target sites were predicted in the CDS of all thegenes considered However for the miRNAs expressed inmidbrain hippocampus and frontal cortex the number oftargets in the coding region is threefold greater than thenumber of miRNA targets found in 51015840 UTR and 31015840 UTRbut in miRNAs expressed in cerebellum the number oftargets in CDS was about twofold levels as compared to otherregions of the genes As there were less miRNAs expressedin cerebellum this can explain small number of target sitespredicted on genes associated to ALS Figure 4 shows thedistribution of collective target sites of miRNAs expressed inbrain in the various regions of the selected genes
Next we analyzed the microRNA targets in each geneassociated with the ALS pathways No target sites could bedetected in 31015840 UTR for miRNAs expressed in midbrain forCASP1 GRIA1 GPX1 DAXX CAT SOD1 NEFM NEFL and
Midbrain 33
Frontal cortex 27
Hippocampus 28
Cerebellum 12
MidbrainFrontal cortex
HippocampusCerebellum
Figure 2 Percentage of miRNAs expressed in various parts of thebrain
0100200300400500600700800900
CDS 3998400UTR5998400UTR
Figure 3 Distribution of target sites in different regions of selectedgenes for the selected miRNAs
85
46
83 85
277
83
222 222
115
46
10488
0
50
100
150
200
250
300
Midbrain Cerebellum Hippocampus Frontal cortex
CDS3998400UTR
5998400UTR
Figure 4 Distribution of target sites in 51015840 UTR CDS and 31015840 UTRfor miRNAs expressed in various parts of nervous system
International Journal of Genomics 5
NEFH genes Similarly CDS of two genesCYCS and RAB5Ahave no potential targets Conversely CDS of MAP3K5have maximum number of targets for miRNAs expressedin midbrain On the other hand 51015840 UTR lack any targetsites of same miRNAs in CASP1 GRIA1 GPX1 DAXX CATSOD1NEFMNEFLNEFHRAC1 andTNF genesThe geneshaving maximum and minimum number of target sites werealso identified The miRNAs expressed in midbrain showedthe least number of target sites (3) in RAC1 and CASP1genes while 29 target sites were predicted in NOS1 Similarresults were obtained for miRNAs expressed in cerebellumfor the same genes For instance RAC1 has only 1 targetsite whereas NOS1 has 11 target sites For miRNAs expressedin hippocampus CASP1 and GRIA1 genes have minimumnumber of predicted sites (3) and MAP3K5 was predictedto possess maximum target sites (33) Similarly CASP1 andSOD1 showed 2 target sites each while 23 target sites werepredicted for TNFRSF1A for miRNAs expressed in frontalcortex On the contrary target sites could not be predictedin CDS for 6 genes namely GPX1 CYCS CHP CASP3SOD1 and RAB5A for miRNAs expressed in cerebellumNo target sites could be predicted in 31015840 UTR of ALS2 BIDCASP1 GRIA1 DAXX CCS CAT CASP9 TOMM40 TNFSOD1 RAC1 NEFM and NEFL genes No target site couldbe predicted in BCL2 CASP1 GRIA1 DAXX CYCS CASP3TNF RAC1 NEFM and NEFH genes for miRNAs expressedin cerebellum These results indicate that the majority ofthe ALS genes lack any target site for miRNAs expressed incerebellum Target sites predicted using miRanda are sum-marized in Table S2 (in Supplementary Material) miRNAtargets were found in most of the genes but surprisingly fewgenesCASP1GRIA1DAXXCAT SOD1 andNEFM lack anymiRNA target site in their 31015840 UTR
31 Hotspot Identification MicroRNAs either share the samecomplementary target sites or proximally located sites adja-cent to other miRNA target site A hot spot can be defined asa stretch of sequence that is prone to action of a group ofmiR-NAs Many studies have revealed that usually microRNAshaving the same origin are regulated and expressed togetherin time and space [37] Such miRNAs play extensive rolein many vital processes and carry out important biologicalfunctions in different pathways [38ndash40] It is quite possiblethat all the miRNA occupying position in ldquohotspotrdquo maynot be equally effective for controlling the same gene Basedon competitive selection predominant miRNA may outcastother miRNAs and get potentially involved in repression ofother microRNA or in similar events But exact mechanismbehind such selection remains unknown It will be interestingto elucidate the factors responsible for variable effectivenessof microRNAs Various researchers have used different stan-dards andparameters such as the same chromosomal locationfor more than two miRNAs the same orientation phyloge-netic relationships and absence of interfering transcriptionunit [41 42] As miRNAs are presumed to be implicatedin important biological processes miRNA targets predictedusing miRanda were then further analyzed for identifyingmiRNA-prone regions in all selected genes We defined
a region as hotspot or region prone tomultiplemiRNA actionif the region showed minimum 10 nucleotides overlap fromthe starting position and concurrence of 3 miRNA targetsTotal 11 miRNAs-prone regions were identified in 35 genesassociated to ALS (Figure SF1-37 in SupplementaryMaterial)CYCS and MAP2K3 showed 1 hotspot each in 31015840 UTR whilesingle target site hotspot was found in 51015840 UTR in 4 genesthat is BAX DERL1 SLC1A2 and RAB5A Two such regionswere found in CDS of CASP9 and 3 other genes namelyTNF MAPK14 and MAP2K3 showed the occurrence ofsingle region vulnerable to multiple miRNAs (Table S7 inSupplementary Material)
In miRNA cluster prediction 2 hot spots were identifiedin MAP2K3 The hotspot in CDS was found to have targetsites for 3 miRNAsthat is hsa-mir-323-5p hsa-mir-654-5p and hsa-mir-433 (Figure 5) Target sites for 3 miRNAsnamely hsa-mir-107 hsa-mir-423-5p and hsa-mir-103 arecongregated in another hotspot found in 31015840 UTR
MAP2K3 having 2 such hotspots is expected to showmore sensitivity towards gene regulation by microRNAsOccurrence of such small number of hot spots indicates thepossible specificity of miRNAs and stringency of regulationThe variability in existence of such target site hotspotsenhances the complexity in gene regulation These resultssuggest that any region in a gene namely 51015840 UTR CDS and31015840 UTR can be targeted by the microRNAs as opposed tothe earlier belief which postulated 31015840 UTR as the only site ofaction of microRNAs
32 Multifarious Relationships among Gene and miRNAsMicroRNA-mediated regulation offers a whole new dimen-sion and often researchers are left confounded by sheercomplexity of its level Not akin to the earlier incidencesof one miRNA targeting one gene distribution of targetsites for the miRNAs points out various possibilities wheremultiplicity and cooperativity come into picture In principleone miRNA can target more than one gene (multiplicity)and one gene can be controlled by more than one miRNA(cooperativity) [26] Multiplicity is a property arising fromrelaxed base pairing As reported earlier single miRNAcan control hundreds of genes Distribution of target siteson genes is rarely uniform and these target sites are oftenfound scattered across a region and range of occurrenceis also known to vary in numbers in a gene [27] Oftenpresence of more target sites per gene indicates efficientregulation In this study we found multiple target sites inmost of the genes After assembling all data multiplicity andcooperativity were calculated From these analyses selectivemiRNAs that displayed maximum number of interactionswith the genes associated with manifestation of ALS wereused for generation of an interaction network (Table 1) Weobserved that hsa-miR-370 showed maximum multiplicityas it showed 65 interactions with 30 gene sequences whileno target site could be predicted in 5 genes namely CASP1GPX1DERL1CCS and SOD1Thus it is expected that degreeof repression by hsa-mir-370 will be considerably high Nexthsa-miR-874 displayed 52 interactions with 29 genes but didnot show any target in CASP1 CHP SOD1 RAC1 RAB5A
6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

International Journal of Genomics 5
NEFH genes Similarly CDS of two genesCYCS and RAB5Ahave no potential targets Conversely CDS of MAP3K5have maximum number of targets for miRNAs expressedin midbrain On the other hand 51015840 UTR lack any targetsites of same miRNAs in CASP1 GRIA1 GPX1 DAXX CATSOD1NEFMNEFLNEFHRAC1 andTNF genesThe geneshaving maximum and minimum number of target sites werealso identified The miRNAs expressed in midbrain showedthe least number of target sites (3) in RAC1 and CASP1genes while 29 target sites were predicted in NOS1 Similarresults were obtained for miRNAs expressed in cerebellumfor the same genes For instance RAC1 has only 1 targetsite whereas NOS1 has 11 target sites For miRNAs expressedin hippocampus CASP1 and GRIA1 genes have minimumnumber of predicted sites (3) and MAP3K5 was predictedto possess maximum target sites (33) Similarly CASP1 andSOD1 showed 2 target sites each while 23 target sites werepredicted for TNFRSF1A for miRNAs expressed in frontalcortex On the contrary target sites could not be predictedin CDS for 6 genes namely GPX1 CYCS CHP CASP3SOD1 and RAB5A for miRNAs expressed in cerebellumNo target sites could be predicted in 31015840 UTR of ALS2 BIDCASP1 GRIA1 DAXX CCS CAT CASP9 TOMM40 TNFSOD1 RAC1 NEFM and NEFL genes No target site couldbe predicted in BCL2 CASP1 GRIA1 DAXX CYCS CASP3TNF RAC1 NEFM and NEFH genes for miRNAs expressedin cerebellum These results indicate that the majority ofthe ALS genes lack any target site for miRNAs expressed incerebellum Target sites predicted using miRanda are sum-marized in Table S2 (in Supplementary Material) miRNAtargets were found in most of the genes but surprisingly fewgenesCASP1GRIA1DAXXCAT SOD1 andNEFM lack anymiRNA target site in their 31015840 UTR
31 Hotspot Identification MicroRNAs either share the samecomplementary target sites or proximally located sites adja-cent to other miRNA target site A hot spot can be defined asa stretch of sequence that is prone to action of a group ofmiR-NAs Many studies have revealed that usually microRNAshaving the same origin are regulated and expressed togetherin time and space [37] Such miRNAs play extensive rolein many vital processes and carry out important biologicalfunctions in different pathways [38ndash40] It is quite possiblethat all the miRNA occupying position in ldquohotspotrdquo maynot be equally effective for controlling the same gene Basedon competitive selection predominant miRNA may outcastother miRNAs and get potentially involved in repression ofother microRNA or in similar events But exact mechanismbehind such selection remains unknown It will be interestingto elucidate the factors responsible for variable effectivenessof microRNAs Various researchers have used different stan-dards andparameters such as the same chromosomal locationfor more than two miRNAs the same orientation phyloge-netic relationships and absence of interfering transcriptionunit [41 42] As miRNAs are presumed to be implicatedin important biological processes miRNA targets predictedusing miRanda were then further analyzed for identifyingmiRNA-prone regions in all selected genes We defined
a region as hotspot or region prone tomultiplemiRNA actionif the region showed minimum 10 nucleotides overlap fromthe starting position and concurrence of 3 miRNA targetsTotal 11 miRNAs-prone regions were identified in 35 genesassociated to ALS (Figure SF1-37 in SupplementaryMaterial)CYCS and MAP2K3 showed 1 hotspot each in 31015840 UTR whilesingle target site hotspot was found in 51015840 UTR in 4 genesthat is BAX DERL1 SLC1A2 and RAB5A Two such regionswere found in CDS of CASP9 and 3 other genes namelyTNF MAPK14 and MAP2K3 showed the occurrence ofsingle region vulnerable to multiple miRNAs (Table S7 inSupplementary Material)
In miRNA cluster prediction 2 hot spots were identifiedin MAP2K3 The hotspot in CDS was found to have targetsites for 3 miRNAsthat is hsa-mir-323-5p hsa-mir-654-5p and hsa-mir-433 (Figure 5) Target sites for 3 miRNAsnamely hsa-mir-107 hsa-mir-423-5p and hsa-mir-103 arecongregated in another hotspot found in 31015840 UTR
MAP2K3 having 2 such hotspots is expected to showmore sensitivity towards gene regulation by microRNAsOccurrence of such small number of hot spots indicates thepossible specificity of miRNAs and stringency of regulationThe variability in existence of such target site hotspotsenhances the complexity in gene regulation These resultssuggest that any region in a gene namely 51015840 UTR CDS and31015840 UTR can be targeted by the microRNAs as opposed tothe earlier belief which postulated 31015840 UTR as the only site ofaction of microRNAs
32 Multifarious Relationships among Gene and miRNAsMicroRNA-mediated regulation offers a whole new dimen-sion and often researchers are left confounded by sheercomplexity of its level Not akin to the earlier incidencesof one miRNA targeting one gene distribution of targetsites for the miRNAs points out various possibilities wheremultiplicity and cooperativity come into picture In principleone miRNA can target more than one gene (multiplicity)and one gene can be controlled by more than one miRNA(cooperativity) [26] Multiplicity is a property arising fromrelaxed base pairing As reported earlier single miRNAcan control hundreds of genes Distribution of target siteson genes is rarely uniform and these target sites are oftenfound scattered across a region and range of occurrenceis also known to vary in numbers in a gene [27] Oftenpresence of more target sites per gene indicates efficientregulation In this study we found multiple target sites inmost of the genes After assembling all data multiplicity andcooperativity were calculated From these analyses selectivemiRNAs that displayed maximum number of interactionswith the genes associated with manifestation of ALS wereused for generation of an interaction network (Table 1) Weobserved that hsa-miR-370 showed maximum multiplicityas it showed 65 interactions with 30 gene sequences whileno target site could be predicted in 5 genes namely CASP1GPX1DERL1CCS and SOD1Thus it is expected that degreeof repression by hsa-mir-370 will be considerably high Nexthsa-miR-874 displayed 52 interactions with 29 genes but didnot show any target in CASP1 CHP SOD1 RAC1 RAB5A
6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 International Journal of Genomics
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-139-3p
hsa-miR-323-5phsa-miR-574-3p
hsa-miR-154hsa-miR-149
hsa-miR-433hsa-miR-654-5p
hsa-miR-323-5p
hsa-miR-127-3phsa-miR-323-5phsa-miR-27a
hsa-miR-125a-5p hsa-miR-127-3p hsa-miR-324-3p
hsa-miR-874
hsa-miR-103hsa-miR-423-5p
hsa-miR-107
hsa-miR-760
hsa-miR-324-5p
0 bp
0 bp 2319 bp
Figure 5 Schematic representation of miRNA targets on MAP2K3 gene
and MAP2K6 genes None of the top miRNAs display anyinteraction with SOD1 Several evidences implicated thatSOD1 is one of the core factor for ALS that encodes the freeradical scavenging enzyme copper zinc superoxide dismutasein ALS pathway It can be interpreted that these miRNAsmaynot be involved in regulation of SOD1 activity
In a similar approach the data was analyzed for esti-mating cooperativity Highest cooperativity was observedfor PRPH followed by TNFRSF1A and TOMM40 towardstop 10 microRNAs The complex picture presented by theseinteractions is difficult to interpret For instance PRPHis regulated by 10 miRNAs at 23 positions so these top10 miRNAs demonstrate high cooperativity towards PRPHTop 10 miRNAs exhibited 22 and 20 targets for TNFRSF1Aand TOMM40 genes Relatively very low cooperativity wasobserved in BID MAP2K6 and DERL1 genes It can beinferred that there exists a low sensitivity of these genestowards microRNA mediated gene regulation Seeminglysimple process of selection of microRNA targets is endowedwith inherent complexity and this led to the development ofa complex network by two phenomena that is multiplicityand cooperativityThese two propertiesmay serve as decidingfactors for themode of miRNA action which have significantrole in chromatin organization rapid RNA turnover ortranslational inhibition A complete and detailed analysis isrequired to unravelmicroRNA-based gene regulation Table 1shows the miRNAs with high multiplicity and cooperativity
33 A Simplistic Approach for Complex Interaction Weemployed a simple approach to derive a complex microRNA-ALS gene integrated network for envisaging the relationshipsshared by top 10 miRNA targeting ALS genes (Figure 6) Themajor advantage underlying this approach is its simplicity
in identifying complete sets of microRNA and their targetson a particular disease at a glance The highest number ofinteractions with genes (30) was shown by hsa-miR-370 Incontrast only 4 interactions were observed in MAP2K6 forhsa-miR-370 hsa-miR-760 and hsa-miR-744 miRNAs Ondetermining targets on an individual gene in considerationit was noted that the maximum number of interactions forselective topmicroRNAs was observed between hsa-miR-370and TNFRSF1A whereas SOD1 did not show any conclusiveinteraction with top miRNAs It can be presumed that SOD1may not be under control of the microRNAs These findingsprovide an important clue that few genes are devoid ofmicroRNA-based regulation Itmight appear that some of thecore components of ALS pathway may not be under directcontrol of miRNAs
34 Prediction of Common Targets To eliminate noise fromcomputational prediction other algorithms were used forverification of the data obtained from miRanda programHowever major drawback of such verification underlies inthe differences in number of predicted targets It can beattributed to the fact that miRNA target sites in animals arequite small with limited complementarity A slight variationin prediction algorithms is capable of producing diverseand variable results As a measure the convergence orintersection of the results was obtained for ensuring thereliability Secondmost of the algorithms restrict their searchonly to 31015840 UTR therefore there is no option available forvalidating themiRNA targets present in the 51015840 UTR andCDSA comparative analysis of three algorithms miRanda Tar-getScan and PicTar is summarized in Figure 7 TargetScanis a well-established tool for identifying complementary sitesfor miRNA binding which has low false positive rate owing
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
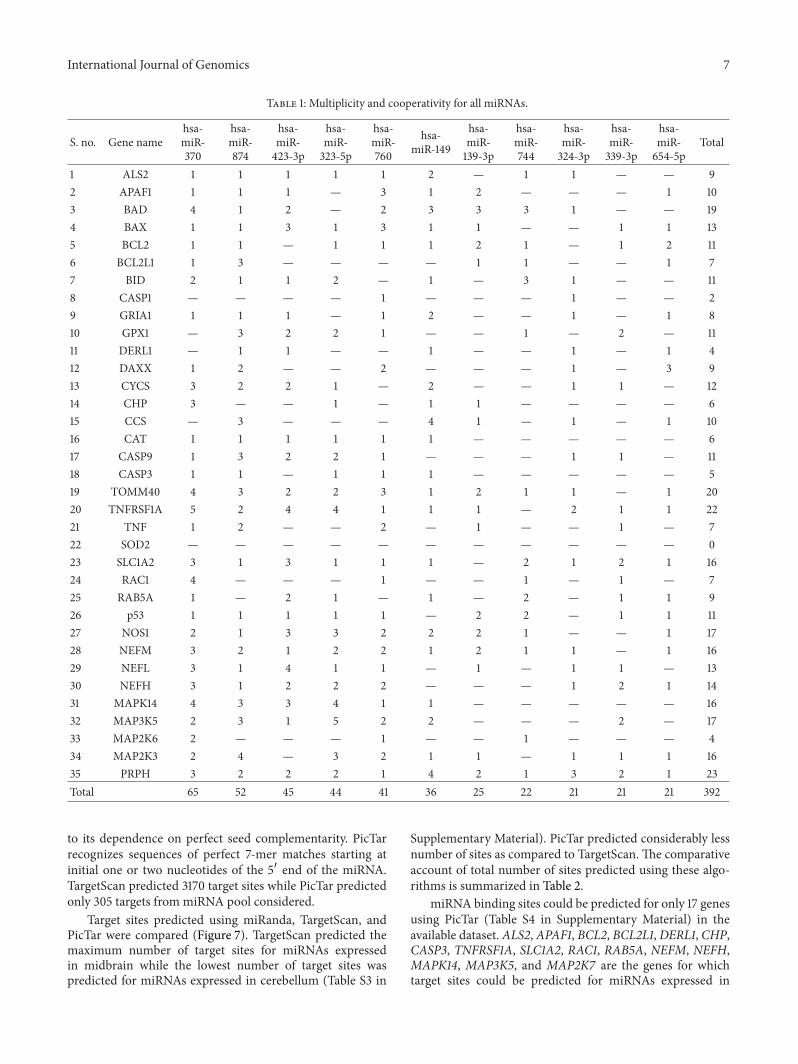
International Journal of Genomics 7
Table 1 Multiplicity and cooperativity for all miRNAs
S no Gene namehsa-miR-370
hsa-miR-874
hsa-miR-423-3p
hsa-miR-323-5p
hsa-miR-760
hsa-miR-149
hsa-miR-139-3p
hsa-miR-744
hsa-miR-324-3p
hsa-miR-339-3p
hsa-miR-654-5p
Total
1 ALS2 1 1 1 1 1 2 mdash 1 1 mdash mdash 92 APAF1 1 1 1 mdash 3 1 2 mdash mdash mdash 1 103 BAD 4 1 2 mdash 2 3 3 3 1 mdash mdash 194 BAX 1 1 3 1 3 1 1 mdash mdash 1 1 135 BCL2 1 1 mdash 1 1 1 2 1 mdash 1 2 116 BCL2L1 1 3 mdash mdash mdash mdash 1 1 mdash mdash 1 77 BID 2 1 1 2 mdash 1 mdash 3 1 mdash mdash 118 CASP1 mdash mdash mdash mdash 1 mdash mdash mdash 1 mdash mdash 29 GRIA1 1 1 1 mdash 1 2 mdash mdash 1 mdash 1 810 GPX1 mdash 3 2 2 1 mdash mdash 1 mdash 2 mdash 1111 DERL1 mdash 1 1 mdash mdash 1 mdash mdash 1 mdash 1 412 DAXX 1 2 mdash mdash 2 mdash mdash mdash 1 mdash 3 913 CYCS 3 2 2 1 mdash 2 mdash mdash 1 1 mdash 1214 CHP 3 mdash mdash 1 mdash 1 1 mdash mdash mdash mdash 615 CCS mdash 3 mdash mdash mdash 4 1 mdash 1 mdash 1 1016 CAT 1 1 1 1 1 1 mdash mdash mdash mdash mdash 617 CASP9 1 3 2 2 1 mdash mdash mdash 1 1 mdash 1118 CASP3 1 1 mdash 1 1 1 mdash mdash mdash mdash mdash 519 TOMM40 4 3 2 2 3 1 2 1 1 mdash 1 2020 TNFRSF1A 5 2 4 4 1 1 1 mdash 2 1 1 2221 TNF 1 2 mdash mdash 2 mdash 1 mdash mdash 1 mdash 722 SOD2 mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash mdash 023 SLC1A2 3 1 3 1 1 1 mdash 2 1 2 1 1624 RAC1 4 mdash mdash mdash 1 mdash mdash 1 mdash 1 mdash 725 RAB5A 1 mdash 2 1 mdash 1 mdash 2 mdash 1 1 926 p53 1 1 1 1 1 mdash 2 2 mdash 1 1 1127 NOS1 2 1 3 3 2 2 2 1 mdash mdash 1 1728 NEFM 3 2 1 2 2 1 2 1 1 mdash 1 1629 NEFL 3 1 4 1 1 mdash 1 mdash 1 1 mdash 1330 NEFH 3 1 2 2 2 mdash mdash mdash 1 2 1 1431 MAPK14 4 3 3 4 1 1 mdash mdash mdash mdash mdash 1632 MAP3K5 2 3 1 5 2 2 mdash mdash mdash 2 mdash 1733 MAP2K6 2 mdash mdash mdash 1 mdash mdash 1 mdash mdash mdash 434 MAP2K3 2 4 mdash 3 2 1 1 mdash 1 1 1 1635 PRPH 3 2 2 2 1 4 2 1 3 2 1 23Total 65 52 45 44 41 36 25 22 21 21 21 392
to its dependence on perfect seed complementarity PicTarrecognizes sequences of perfect 7-mer matches starting atinitial one or two nucleotides of the 51015840 end of the miRNATargetScan predicted 3170 target sites while PicTar predictedonly 305 targets from miRNA pool considered
Target sites predicted using miRanda TargetScan andPicTar were compared (Figure 7) TargetScan predicted themaximum number of target sites for miRNAs expressedin midbrain while the lowest number of target sites waspredicted for miRNAs expressed in cerebellum (Table S3 in
Supplementary Material) PicTar predicted considerably lessnumber of sites as compared to TargetScan The comparativeaccount of total number of sites predicted using these algo-rithms is summarized in Table 2
miRNA binding sites could be predicted for only 17 genesusing PicTar (Table S4 in Supplementary Material) in theavailable dataset ALS2 APAF1 BCL2 BCL2L1DERL1 CHPCASP3 TNFRSF1A SLC1A2 RAC1 RAB5A NEFM NEFHMAPK14 MAP3K5 and MAP2K7 are the genes for whichtarget sites could be predicted for miRNAs expressed in
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
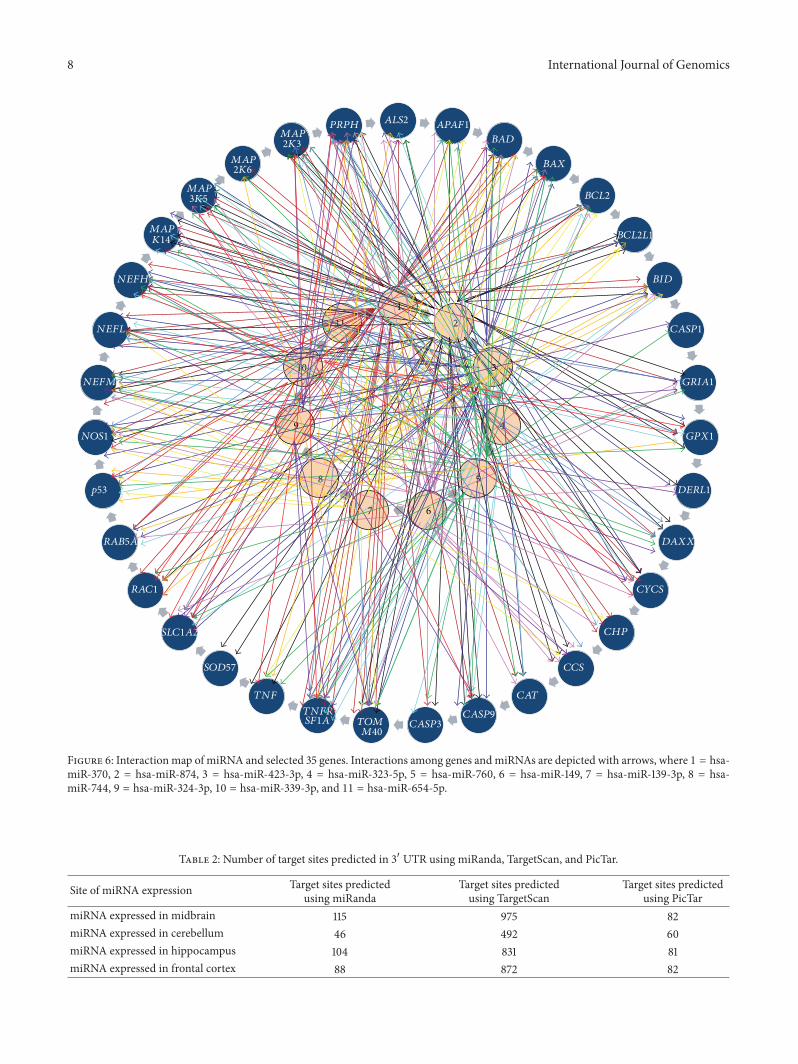
8 International Journal of Genomics
ALS2 APAF1
BAD
BAX
BCL2
BCL2L1
BID
CASP1
GRIA1
GPX1
DERL1
DAXX
CYCS
CHP
CCS
CAT
CASP9CASP3TOM
TNFRSF1A
TNF
SOD57
SLC1A2
RAC1
RAB5A
p53
NOS1
NEFM
NEFL
NEFH
MAP
MAP
MAP
MAPPRPH
1
2
3
4
5
67
8
9
10
11
M40
2K3
2K6
3K5
K14
Figure 6 Interaction map of miRNA and selected 35 genes Interactions among genes andmiRNAs are depicted with arrows where 1 = hsa-miR-370 2 = hsa-miR-874 3 = hsa-miR-423-3p 4 = hsa-miR-323-5p 5 = hsa-miR-760 6 = hsa-miR-149 7 = hsa-miR-139-3p 8 = hsa-miR-744 9 = hsa-miR-324-3p 10 = hsa-miR-339-3p and 11 = hsa-miR-654-5p
Table 2 Number of target sites predicted in 31015840 UTR using miRanda TargetScan and PicTar
Site of miRNA expression Target sites predictedusing miRanda
Target sites predictedusing TargetScan
Target sites predictedusing PicTar
miRNA expressed in midbrain 115 975 82miRNA expressed in cerebellum 46 492 60miRNA expressed in hippocampus 104 831 81miRNA expressed in frontal cortex 88 872 82
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
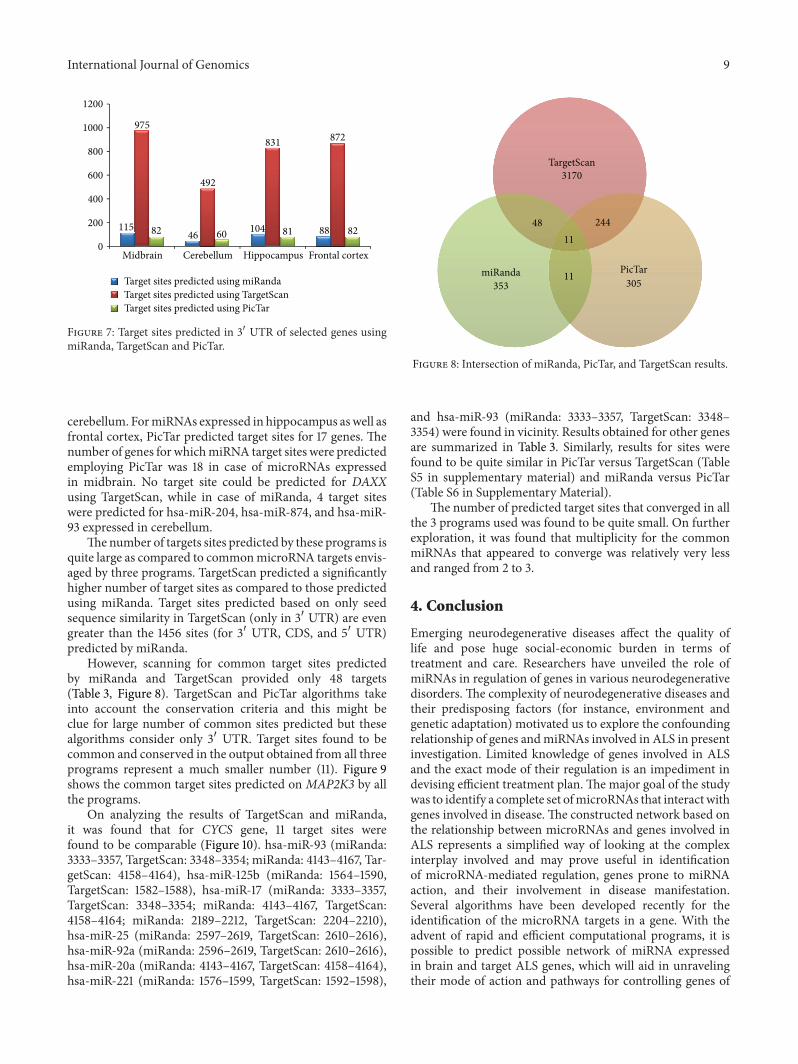
International Journal of Genomics 9
11546 104 88
975
492
831 872
82 60 81 820
200
400
600
800
1000
1200
Midbrain Cerebellum Hippocampus Frontal cortex
Target sites predicted using miRandaTarget sites predicted using TargetScanTarget sites predicted using PicTar
Figure 7 Target sites predicted in 31015840 UTR of selected genes usingmiRanda TargetScan and PicTar
cerebellum FormiRNAs expressed in hippocampus aswell asfrontal cortex PicTar predicted target sites for 17 genes Thenumber of genes for whichmiRNA target sites were predictedemploying PicTar was 18 in case of microRNAs expressedin midbrain No target site could be predicted for DAXXusing TargetScan while in case of miRanda 4 target siteswere predicted for hsa-miR-204 hsa-miR-874 and hsa-miR-93 expressed in cerebellum
Thenumber of targets sites predicted by these programs isquite large as compared to commonmicroRNA targets envis-aged by three programs TargetScan predicted a significantlyhigher number of target sites as compared to those predictedusing miRanda Target sites predicted based on only seedsequence similarity in TargetScan (only in 31015840 UTR) are evengreater than the 1456 sites (for 31015840 UTR CDS and 51015840 UTR)predicted by miRanda
However scanning for common target sites predictedby miRanda and TargetScan provided only 48 targets(Table 3 Figure 8) TargetScan and PicTar algorithms takeinto account the conservation criteria and this might beclue for large number of common sites predicted but thesealgorithms consider only 31015840 UTR Target sites found to becommon and conserved in the output obtained from all threeprograms represent a much smaller number (11) Figure 9shows the common target sites predicted on MAP2K3 by allthe programs
On analyzing the results of TargetScan and miRandait was found that for CYCS gene 11 target sites werefound to be comparable (Figure 10) hsa-miR-93 (miRanda3333ndash3357 TargetScan 3348ndash3354 miRanda 4143ndash4167 Tar-getScan 4158ndash4164) hsa-miR-125b (miRanda 1564ndash1590TargetScan 1582ndash1588) hsa-miR-17 (miRanda 3333ndash3357TargetScan 3348ndash3354 miRanda 4143ndash4167 TargetScan4158ndash4164 miRanda 2189ndash2212 TargetScan 2204ndash2210)hsa-miR-25 (miRanda 2597ndash2619 TargetScan 2610ndash2616)hsa-miR-92a (miRanda 2596ndash2619 TargetScan 2610ndash2616)hsa-miR-20a (miRanda 4143ndash4167 TargetScan 4158ndash4164)hsa-miR-221 (miRanda 1576ndash1599 TargetScan 1592ndash1598)
TargetScan3170
PicTar305
miRanda353
48 244
11
11
Figure 8 Intersection of miRanda PicTar and TargetScan results
and hsa-miR-93 (miRanda 3333ndash3357 TargetScan 3348ndash3354) were found in vicinity Results obtained for other genesare summarized in Table 3 Similarly results for sites werefound to be quite similar in PicTar versus TargetScan (TableS5 in supplementary material) and miRanda versus PicTar(Table S6 in Supplementary Material)
The number of predicted target sites that converged in allthe 3 programs used was found to be quite small On furtherexploration it was found that multiplicity for the commonmiRNAs that appeared to converge was relatively very lessand ranged from 2 to 3
4 Conclusion
Emerging neurodegenerative diseases affect the quality oflife and pose huge social-economic burden in terms oftreatment and care Researchers have unveiled the role ofmiRNAs in regulation of genes in various neurodegenerativedisorders The complexity of neurodegenerative diseases andtheir predisposing factors (for instance environment andgenetic adaptation) motivated us to explore the confoundingrelationship of genes andmiRNAs involved in ALS in presentinvestigation Limited knowledge of genes involved in ALSand the exact mode of their regulation is an impediment indevising efficient treatment plan The major goal of the studywas to identify a complete set ofmicroRNAs that interactwithgenes involved in disease The constructed network based onthe relationship between microRNAs and genes involved inALS represents a simplified way of looking at the complexinterplay involved and may prove useful in identificationof microRNA-mediated regulation genes prone to miRNAaction and their involvement in disease manifestationSeveral algorithms have been developed recently for theidentification of the microRNA targets in a gene With theadvent of rapid and efficient computational programs it ispossible to predict possible network of miRNA expressedin brain and target ALS genes which will aid in unravelingtheir mode of action and pathways for controlling genes of
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
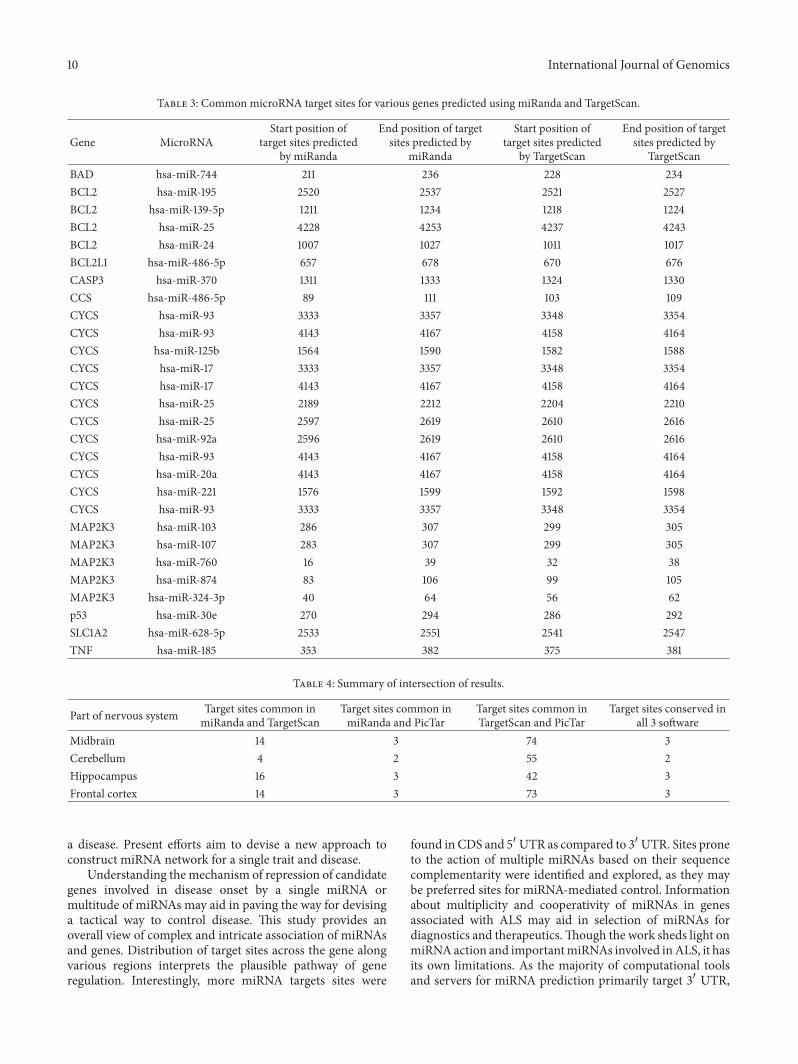
10 International Journal of Genomics
Table 3 Common microRNA target sites for various genes predicted using miRanda and TargetScan
Gene MicroRNAStart position of
target sites predictedby miRanda
End position of targetsites predicted by
miRanda
Start position oftarget sites predicted
by TargetScan
End position of targetsites predicted by
TargetScanBAD hsa-miR-744 211 236 228 234BCL2 hsa-miR-195 2520 2537 2521 2527BCL2 hsa-miR-139-5p 1211 1234 1218 1224BCL2 hsa-miR-25 4228 4253 4237 4243BCL2 hsa-miR-24 1007 1027 1011 1017BCL2L1 hsa-miR-486-5p 657 678 670 676CASP3 hsa-miR-370 1311 1333 1324 1330CCS hsa-miR-486-5p 89 111 103 109CYCS hsa-miR-93 3333 3357 3348 3354CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-125b 1564 1590 1582 1588CYCS hsa-miR-17 3333 3357 3348 3354CYCS hsa-miR-17 4143 4167 4158 4164CYCS hsa-miR-25 2189 2212 2204 2210CYCS hsa-miR-25 2597 2619 2610 2616CYCS hsa-miR-92a 2596 2619 2610 2616CYCS hsa-miR-93 4143 4167 4158 4164CYCS hsa-miR-20a 4143 4167 4158 4164CYCS hsa-miR-221 1576 1599 1592 1598CYCS hsa-miR-93 3333 3357 3348 3354MAP2K3 hsa-miR-103 286 307 299 305MAP2K3 hsa-miR-107 283 307 299 305MAP2K3 hsa-miR-760 16 39 32 38MAP2K3 hsa-miR-874 83 106 99 105MAP2K3 hsa-miR-324-3p 40 64 56 62p53 hsa-miR-30e 270 294 286 292SLC1A2 hsa-miR-628-5p 2533 2551 2541 2547TNF hsa-miR-185 353 382 375 381
Table 4 Summary of intersection of results
Part of nervous system Target sites common inmiRanda and TargetScan
Target sites common inmiRanda and PicTar
Target sites common inTargetScan and PicTar
Target sites conserved inall 3 software
Midbrain 14 3 74 3Cerebellum 4 2 55 2Hippocampus 16 3 42 3Frontal cortex 14 3 73 3
a disease Present efforts aim to devise a new approach toconstruct miRNA network for a single trait and disease
Understanding themechanism of repression of candidategenes involved in disease onset by a single miRNA ormultitude of miRNAs may aid in paving the way for devisinga tactical way to control disease This study provides anoverall view of complex and intricate association of miRNAsand genes Distribution of target sites across the gene alongvarious regions interprets the plausible pathway of generegulation Interestingly more miRNA targets sites were
found in CDS and 51015840UTR as compared to 31015840 UTR Sites proneto the action of multiple miRNAs based on their sequencecomplementarity were identified and explored as they maybe preferred sites for miRNA-mediated control Informationabout multiplicity and cooperativity of miRNAs in genesassociated with ALS may aid in selection of miRNAs fordiagnostics and therapeuticsThough the work sheds light onmiRNAaction and importantmiRNAs involved inALS it hasits own limitations As the majority of computational toolsand servers for miRNA prediction primarily target 31015840 UTR
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
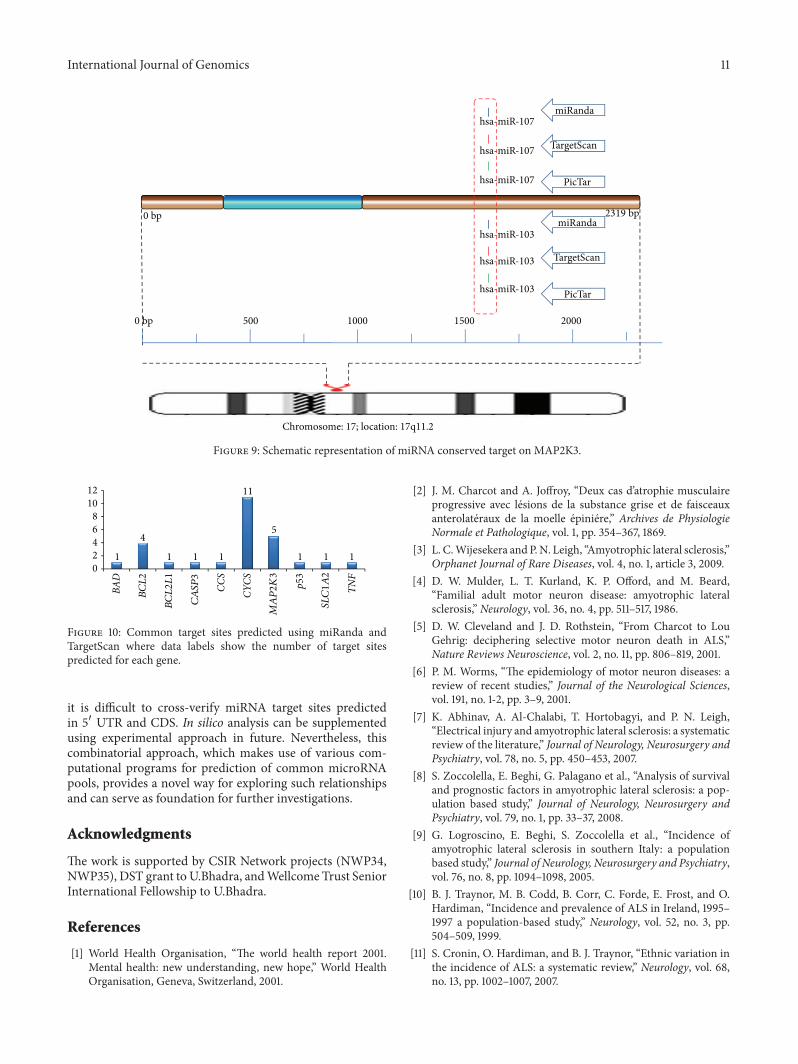
International Journal of Genomics 11
Chromosome 17 location 17q112
500 1000 1500 2000
hsa-miR-107
hsa-miR-107
hsa-miR-107
hsa-miR-103
hsa-miR-103
hsa-miR-103
miRanda
TargetScan
PicTar
miRanda
TargetScan
PicTar
0 bp
0 bp 2319 bp
Figure 9 Schematic representation of miRNA conserved target on MAP2K3
1
4
1 1 1
11
5
1 1 102468
1012
BAD
CCS
CYCS
TNF
BCL2
BCL2L1
CASP
3
MAP2K3
p53
SLC1A2
Figure 10 Common target sites predicted using miRanda andTargetScan where data labels show the number of target sitespredicted for each gene
it is difficult to cross-verify miRNA target sites predictedin 51015840 UTR and CDS In silico analysis can be supplementedusing experimental approach in future Nevertheless thiscombinatorial approach which makes use of various com-putational programs for prediction of common microRNApools provides a novel way for exploring such relationshipsand can serve as foundation for further investigations
Acknowledgments
The work is supported by CSIR Network projects (NWP34NWP35) DST grant to UBhadra andWellcome Trust SeniorInternational Fellowship to UBhadra
References
[1] World Health Organisation ldquoThe world health report 2001Mental health new understanding new hoperdquo World HealthOrganisation Geneva Switzerland 2001
[2] J M Charcot and A Joffroy ldquoDeux cas drsquoatrophie musculaireprogressive avec lesions de la substance grise et de faisceauxanterolateraux de la moelle epiniererdquo Archives de PhysiologieNormale et Pathologique vol 1 pp 354ndash367 1869
[3] L CWijesekera and PN Leigh ldquoAmyotrophic lateral sclerosisrdquoOrphanet Journal of Rare Diseases vol 4 no 1 article 3 2009
[4] D W Mulder L T Kurland K P Offord and M BeardldquoFamilial adult motor neuron disease amyotrophic lateralsclerosisrdquo Neurology vol 36 no 4 pp 511ndash517 1986
[5] D W Cleveland and J D Rothstein ldquoFrom Charcot to LouGehrig deciphering selective motor neuron death in ALSrdquoNature Reviews Neuroscience vol 2 no 11 pp 806ndash819 2001
[6] P M Worms ldquoThe epidemiology of motor neuron diseases areview of recent studiesrdquo Journal of the Neurological Sciencesvol 191 no 1-2 pp 3ndash9 2001
[7] K Abhinav A Al-Chalabi T Hortobagyi and P N LeighldquoElectrical injury and amyotrophic lateral sclerosis a systematicreview of the literaturerdquo Journal of Neurology Neurosurgery andPsychiatry vol 78 no 5 pp 450ndash453 2007
[8] S Zoccolella E Beghi G Palagano et al ldquoAnalysis of survivaland prognostic factors in amyotrophic lateral sclerosis a pop-ulation based studyrdquo Journal of Neurology Neurosurgery andPsychiatry vol 79 no 1 pp 33ndash37 2008
[9] G Logroscino E Beghi S Zoccolella et al ldquoIncidence ofamyotrophic lateral sclerosis in southern Italy a populationbased studyrdquo Journal of Neurology Neurosurgery and Psychiatryvol 76 no 8 pp 1094ndash1098 2005
[10] B J Traynor M B Codd B Corr C Forde E Frost and OHardiman ldquoIncidence and prevalence of ALS in Ireland 1995ndash1997 a population-based studyrdquo Neurology vol 52 no 3 pp504ndash509 1999
[11] S Cronin O Hardiman and B J Traynor ldquoEthnic variation inthe incidence of ALS a systematic reviewrdquo Neurology vol 68no 13 pp 1002ndash1007 2007
12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 International Journal of Genomics
[12] T G Lesnick E J Sorenson J E Ahlskog et al ldquoBeyondParkinson disease amyotrophic lateral sclerosis and the Axonguidance pathwayrdquo PLoS ONE vol 3 no 1 Article ID e14492008
[13] E Chevalier-Larsen and E L F Holzbaur ldquoAxonal transportand neurodegenerative diseaserdquo Biochimica et Biophysica Actavol 1762 no 11-12 pp 1094ndash1108 2006
[14] S Boillee C Vande Velde and D Cleveland ldquoALS a disease ofmotor neurons and their nonneuronal neighborsrdquo Neuron vol52 no 1 pp 39ndash59 2006
[15] N Rajewsky ldquomicroRNA target predictions in animalsrdquoNatureGenetics vol 38 no 1 pp S8ndashS13 2006
[16] S K Singh M Pal Bhadra H J Girschick and U BhadraldquoMicroRNAsmdashmicro in size but macro in functionrdquo FEBSJournal vol 275 no 20 pp 4929ndash4944 2008
[17] S N Bhattacharyya R Habermacher U Martine E I ClossandW Filipowicz ldquoRelief of microRNA-mediated translationalrepression in human cells subjected to stressrdquo Cell vol 125 no6 pp 1111ndash1124 2006
[18] W J Lukiw Y Zhao and G C Jian ldquoAn NF-120581B-sensitivemicro RNA-146a-mediated inflammatory circuit in alzheimerdisease and in stressed human brain cellsrdquo Journal of BiologicalChemistry vol 283 no 46 pp 31315ndash31322 2008
[19] R Saba C D Goodman R L C H Huzarewich C Robertsonand SA Booth ldquoAmiRNAsignature of prion inducedneurode-generationrdquo PLoS ONE vol 3 no 11 Article ID e3652 2008
[20] N Bushati and S M Cohen ldquomicroRNAs in neurodegenera-tionrdquo Current Opinion in Neurobiology vol 18 no 3 pp 292ndash296 2008
[21] J R Brown and P Sanseau ldquoA computational view of microR-NAs and their targetsrdquo Drug Discovery Today vol 10 no 8 pp595ndash601 2005
[22] I Bentwich ldquoPrediction and validation ofmicroRNAs and theirtargetsrdquo FEBS Letters vol 579 no 26 pp 5904ndash5910 2005
[23] E Berezikov V Guryev J Van De Belt E Wienholds RH A Plasterk and E Cuppen ldquoPhylogenetic shadowing andcomputational identification of human microRNA genesrdquo Cellvol 120 no 1 pp 21ndash24 2005
[24] M Kanehisa M Araki S Goto et al ldquoKEGG for linkinggenomes to life and the environmentrdquo Nucleic Acids Researchvol 36 no 1 pp D480ndashD484 2008
[25] S Griffiths-Jones H K Saini S Van Dongen and A JEnright ldquomiRBase tools for microRNA genomicsrdquo NucleicAcids Research vol 36 no 1 pp D154ndashD158 2008
[26] A J Enright B John U Gaul T Tuschl C Sander and D SMarks ldquoMicroRNA targets inDrosophilardquoGenome Biology vol5 pp 1ndash12 2003
[27] B John A J Enright A Aravin T Tuschl C Sander and D SMarks ldquoHumanmicroRNA targetsrdquo PLoS Biology vol 2 no 11article e363 2004
[28] S Wuchty W Fontana I L Hofacker and P Schuster ldquoCom-plete suboptimal folding of RNA and the stability of secondarystructuresrdquo Biopolymers vol 49 pp 145ndash165 1999
[29] B P Lewis C B Burge and D P Bartel ldquoConserved seedpairing often flanked by adenosines indicates that thousandsof human genes are microRNA targetsrdquo Cell vol 120 no 1 pp15ndash20 2005
[30] A Krek D Grun M N Poy et al ldquoCombinatorial microRNAtarget predictionsrdquo Nature Genetics vol 37 no 5 pp 495ndash5002005
[31] X Zhou X Duan J Qian and F Li ldquoAbundant conservedmicroRNA target sites in the 51015840-untranslated region and codingsequencerdquo Genetica vol 137 no 2 pp 159ndash164 2009
[32] A M Duursma M Kedde M Schrier C Le Sage andR Agami ldquomiR-148 targets human DNMT3b protein codingregionrdquo RNA vol 14 no 5 pp 872ndash877 2008
[33] J J Forman A Legesse-Miller and H A Coller ldquoA searchfor conserved sequences in coding regions reveals that thelet-7 microRNA targets Dicer within its coding sequencerdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 39 pp 14879ndash14884 2008
[34] A LalHHKimKAbdelmohsen et al ldquop16INK4a translationsuppressed bymiR-24rdquoPLoSONE vol 3 no 3 Article ID e18642008
[35] I Rigoutsos ldquoNew tricks for animal micrornas targeting ofamino acid coding regions at conserved and nonconservedsitesrdquo Cancer Research vol 69 no 8 pp 3245ndash3248 2009
[36] Y Tay J Zhang A M Thomson B Lim and I RigoutsosldquoMicroRNAs toNanogOct4 and Sox2 coding regionsmodulateembryonic stem cell differentiationrdquo Nature vol 455 no 7216pp 1124ndash1128 2008
[37] A Tanzer and P F Stadler ldquoMolecular evolution of amicroRNAclusterrdquo Journal ofMolecular Biology vol 339 no 2 pp 327ndash3352004
[38] Y Hayashita H Osada Y Tatematsu et al ldquoA polycistronicMicroRNA cluster miR-17-92 is overexpressed in human lungcancers and enhances cell proliferationrdquo Cancer Research vol65 no 21 pp 9628ndash9632 2005
[39] M A Samols J Hu R L Skalsky and R Renne ldquoCloningand identification of a MicroRNA cluster within the latency-associated region of Kaposirsquos sarcoma-associated herpesvirusrdquoJournal of Virology vol 79 no 14 pp 9301ndash9305 2005
[40] R Zhang Y-Q Wang and B Su ldquoMolecular evolution ofa primate-specific microRNA familyrdquo Molecular Biology andEvolution vol 25 no 7 pp 1493ndash1502 2008
[41] A Tranzer and P F Stadler ldquoEvolution ofmicroRNAsrdquoMethodsin Molecular Biology vol 342 pp 335ndash350 2006
[42] J R Lytle T A Yario and J A Steitz ldquoTarget mRNAs arerepressed as efficiently by microRNA-binding sites in the 51015840UTR as in the 31015840 UTRrdquo Proceedings of the National Academyof Sciences of the United States of America vol 104 no 23 pp9667ndash9672 2007
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology





























