Embed Size (px)
Citation preview

Research ArticleThe Compound of Mangiferin-Berberine Salt Has PotentActivities in Modulating Lipid and Glucose Metabolisms inHepG2 Cells
Can Wang1 Jian-Dong Jiang2 Wei Wu3 and Wei-Jia Kong1
1Department of Virology Institute of Medicinal Biotechnology Chinese Academy of Medical Sciences andPeking Union Medical College Beijing 100050 China2State Key Laboratory of Bioactive Natural Products and Function Institute of Materia Medica Chinese Academy ofMedical Sciences and Peking Union Medical College Beijing 100050 China3Changzhou Deze Medical Science Co Ltd Changzhou Jiangsu 213100 China
Correspondence should be addressed to Wei Wu weiwubrenda126com and Wei-Jia Kong wjkong894163com
Received 10 December 2015 Revised 28 February 2016 Accepted 7 March 2016
Academic Editor Adair Santos
Copyright copy 2016 Can Wang et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
The mangiferin-berberine (MB) salt was synthesized by ionic bonding of mangiferin (M) and berberine (B) at an equal molecularratio This study aimed to investigate the activities of MB salt in modulating lipid and glucose metabolisms in HepG2 cells After24 h treatment of the studying compounds cellular AMP-activated protein kinase 120572 (AMPK120572)acetyl-CoA carboxylase (ACC)protein levels and carnitine palmitoyltransferase (CPT) 1 activities intracellular lipid contents mRNA expression levels of targetgenes glucose consumption and glucose production amounts were determined Compound C (CC) was used in the blockingexperiments Our results showed thatMB salt increased p-AMPK120572 (Thr172)p-ACC (Ser79) levels andCPT1 activity and suppressedoleic acid- (OA-) induced lipid accumulation and upregulation of lipogenic genes potently in HepG2 cells The above activities ofMB salt were AMPK dependent and were superior to those of M or B when administered at an equal molar concentration MB saltenhanced basal and insulin-stimulated glucose consumption and suppressed gluconeogenesis more potently than M or B aloneThe inhibiting activity of MB salt on cellular gluconeogenesis was AMPK dependent Our results may support MB salt as a newkind of agent for the development of novel lipid or glucose-lowering drugs in the future
1 Introduction
The metabolism syndrome (MS) is characterized by dyslipi-demia glucose intolerance andor insulin resistance hyper-tension and obesity [1] If there are no proper interventionsMS may lead to diabetes and complications coronary heartdisease or even cancer eventually Currently chemical drugssuch as biguanide and thiazolidinedione are commonly usedin clinic for treatment of MS in order to improve metabolicdisorders [1]
In addition to the abovementioned chemical drugs nu-merous studies indicated that natural products isolated fromplants might have beneficial effects in modulating lipid andglucose metabolisms both in vitro and in animal models [2]Some natural products are now subjected to clinical studies
for the treatment of metabolic diseases among them a fewcompounds may have promising application prospects [3]
Mangiferin (M Figure 1(a)) a xanthone glycoside is anatural compound extracted from plants such as Mangiferaindica and Anemarrhena asphodeloides It was reported tohave hypolipidemic [4ndash7] hypoglycemic [8ndash12] insulin-sensitizing [13] antiobesity [8 9] antioxidative [14ndash16] andanti-inflammatory [14 17] activities in animal models as wellas in clinic The beneficial effects of M on lipid and glucosemetabolisms might be related to the activation of AMP-activated protein kinase (AMPK) [5 17 18] a key moleculethat controls energy balance and metabolism in organisms[19]
Berberine (B Figure 1(a)) an isoquinoline alkaloid isa natural compound isolated from plants such as Coptis
Hindawi Publishing CorporationBioMed Research InternationalVolume 2016 Article ID 8753436 14 pageshttpdxdoiorg10115520168753436
2 BioMed Research International
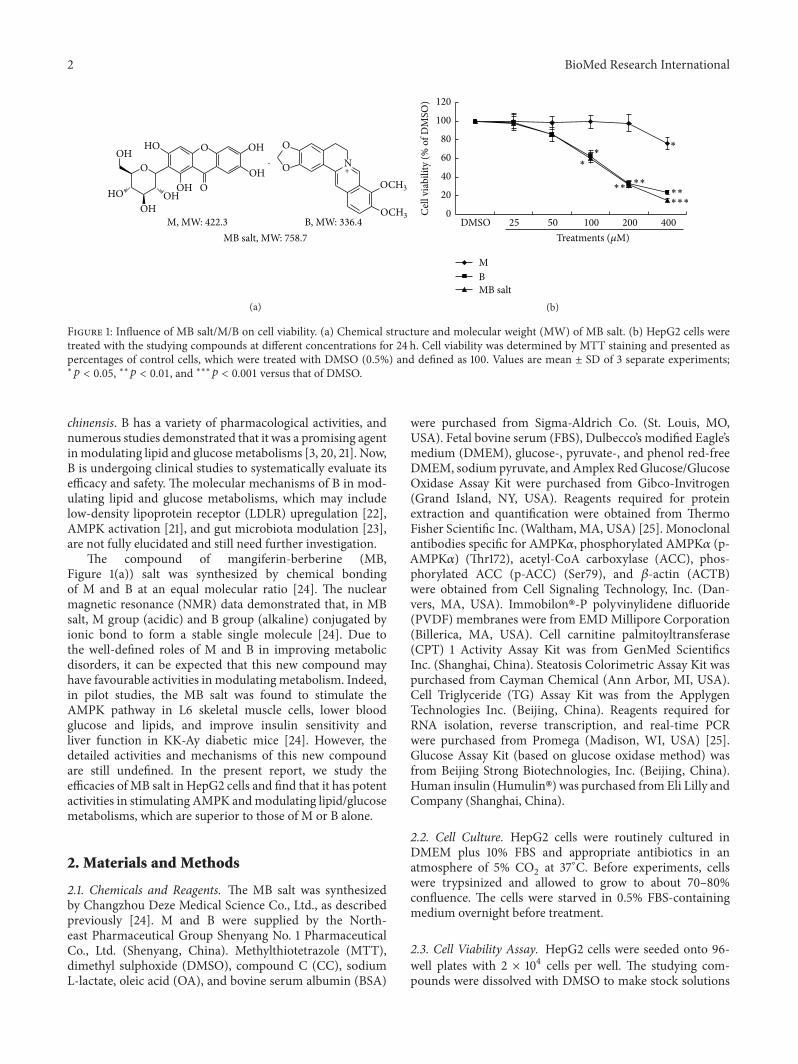
M MW 4223 B MW 3364MB salt MW 7587
N
O
O
O
O
O +
OCH3
OCH3
HO
HO OH
OHOH
OHOH
OHmiddot
(a)
DMSO 25 50 100 200 400
MBMB salt
lowast
lowastlowast
lowastlowastlowastlowast lowastlowastlowastlowastlowast
Treatments (120583M)
0
20
40
60
80
100
120
Cel
l via
bilit
y (
of D
MSO
)
(b)
Figure 1 Influence of MB saltMB on cell viability (a) Chemical structure and molecular weight (MW) of MB salt (b) HepG2 cells weretreated with the studying compounds at different concentrations for 24 h Cell viability was determined by MTT staining and presented aspercentages of control cells which were treated with DMSO (05) and defined as 100 Values are mean plusmn SD of 3 separate experimentslowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO
chinensis B has a variety of pharmacological activities andnumerous studies demonstrated that it was a promising agentinmodulating lipid and glucosemetabolisms [3 20 21] NowB is undergoing clinical studies to systematically evaluate itsefficacy and safety The molecular mechanisms of B in mod-ulating lipid and glucose metabolisms which may includelow-density lipoprotein receptor (LDLR) upregulation [22]AMPK activation [21] and gut microbiota modulation [23]are not fully elucidated and still need further investigation
The compound of mangiferin-berberine (MBFigure 1(a)) salt was synthesized by chemical bondingof M and B at an equal molecular ratio [24] The nuclearmagnetic resonance (NMR) data demonstrated that in MBsalt M group (acidic) and B group (alkaline) conjugated byionic bond to form a stable single molecule [24] Due tothe well-defined roles of M and B in improving metabolicdisorders it can be expected that this new compound mayhave favourable activities in modulatingmetabolism Indeedin pilot studies the MB salt was found to stimulate theAMPK pathway in L6 skeletal muscle cells lower bloodglucose and lipids and improve insulin sensitivity andliver function in KK-Ay diabetic mice [24] However thedetailed activities and mechanisms of this new compoundare still undefined In the present report we study theefficacies of MB salt in HepG2 cells and find that it has potentactivities in stimulating AMPK andmodulating lipidglucosemetabolisms which are superior to those of M or B alone
2 Materials and Methods
21 Chemicals and Reagents The MB salt was synthesizedby Changzhou Deze Medical Science Co Ltd as describedpreviously [24] M and B were supplied by the North-east Pharmaceutical Group Shenyang No 1 PharmaceuticalCo Ltd (Shenyang China) Methylthiotetrazole (MTT)dimethyl sulphoxide (DMSO) compound C (CC) sodiumL-lactate oleic acid (OA) and bovine serum albumin (BSA)
were purchased from Sigma-Aldrich Co (St Louis MOUSA) Fetal bovine serum (FBS) Dulbeccorsquos modified Eaglersquosmedium (DMEM) glucose- pyruvate- and phenol red-freeDMEM sodium pyruvate andAmplex RedGlucoseGlucoseOxidase Assay Kit were purchased from Gibco-Invitrogen(Grand Island NY USA) Reagents required for proteinextraction and quantification were obtained from ThermoFisher Scientific Inc (Waltham MA USA) [25] Monoclonalantibodies specific for AMPK120572 phosphorylated AMPK120572 (p-AMPK120572) (Thr172) acetyl-CoA carboxylase (ACC) phos-phorylated ACC (p-ACC) (Ser79) and 120573-actin (ACTB)were obtained from Cell Signaling Technology Inc (Dan-vers MA USA) Immobilon-P polyvinylidene difluoride(PVDF) membranes were from EMDMillipore Corporation(Billerica MA USA) Cell carnitine palmitoyltransferase(CPT) 1 Activity Assay Kit was from GenMed ScientificsInc (Shanghai China) Steatosis Colorimetric Assay Kit waspurchased from Cayman Chemical (Ann Arbor MI USA)Cell Triglyceride (TG) Assay Kit was from the ApplygenTechnologies Inc (Beijing China) Reagents required forRNA isolation reverse transcription and real-time PCRwere purchased from Promega (Madison WI USA) [25]Glucose Assay Kit (based on glucose oxidase method) wasfrom Beijing Strong Biotechnologies Inc (Beijing China)Human insulin (Humulin) was purchased from Eli Lilly andCompany (Shanghai China)
22 Cell Culture HepG2 cells were routinely cultured inDMEM plus 10 FBS and appropriate antibiotics in anatmosphere of 5 CO
2at 37∘C Before experiments cells
were trypsinized and allowed to grow to about 70ndash80confluence The cells were starved in 05 FBS-containingmedium overnight before treatment
23 Cell Viability Assay HepG2 cells were seeded onto 96-well plates with 2 times 104 cells per well The studying com-pounds were dissolved with DMSO to make stock solutions
BioMed Research International 3
at a concentration of 80mM which were stored at minus20∘Cin aliquots Before usage the stock solutions were thawedand serially diluted with DMEM + 05 FBS After 24 h ofincubation and serum starvation cells were treated with thestudying compounds for 24 h as indicated (Figure 1(b)) eachtreatment had 5 replicate wells Control cells were treatedwith 05 DMSO in DMEM + 05 FBS a concentrationwith no cytotoxicity [26] and equal to the concentration ofDMSO in cells treated with 400120583M of studying compoundsCell viability was determined by MTT staining as describedpreviously [27] the absorbance was read by a VICTOR X4Multilabel Plate Reader (PerkinElmer Inc Waltham MAUSA) at a wavelength of 595 nm The results were presentedas percentages of control cells which were defined as 100Thevalues of 50 inhibiting concentrations (IC
50values) of the
compounds were calculated as described before [27]
24 Western Blot After treatment with the studying com-pounds for 24 h cell total proteins were extracted andquantified Samples containing about 20 120583g of protein wereused for 10 sodium dodecyl sulfate polyacrylamide gelelectrophoresis (SDS-PAGE)The blots were then transferredfrom gels onto PVDF membranes as described previously[25] After blocking the protein levels of AMPK120572 ACC andACTBwere detectedwith specificmonoclonal antibodies andan appropriate secondary antibody signals were developedwith an ECL kit (EMD Millipore Corporation) p-AMPK120572(Thr172) and p-ACC (Ser79) levels were examined by phos-phospecific antibodies after removal of antibody bindingfrom the membranes After scanning and quantificationthe levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) werenormalized to those of AMPK120572 and ACC and plotted asindicated
25 Cellular CPT1 Activity Assay After treatment for 24 hcells were harvested samples containing 50 120583g of proteinwere used for CPT1 activity assay according to the supplierrsquosprotocol The CPT1 experiments were repeated for 3 timescellular CPT1 activities were presented as nmol of coenzymeA (CoA) produced in the assay system per minute
26 Induction of Steatosis Oil Red O (ORO) Staining andIntracellular TGAssay OAwas dissolved in sterile phosphatebuffered saline (PBS) + 5 BSA [28] to make a stock solutionof 6mM and was stored at minus20∘C in aliquots Before usagethe stock solution was thawed and diluted with DMEM +05 FBS for 10 times HepG2 cells were seeded onto 6-wellplates with 5 times 105 cells per well After 24 h of incubationand serum starvation cells were left untreated or treated with06mM of OA for 24 h MB saltMB were serially dilutedfrom their stock solutions with DMEM + 05 FBS at thesame time of OA administration they were added to the cellsat indicated concentrations except for some cells which wereused as OA control After treatment intracellular lipids werestained with ORO by the Steatosis Colorimetric Assay Kitaccording to the supplierrsquos protocol After staining the cellswere observed under a light microscope and photographedIn parallel experiments cells were harvested after treatment
Table 1 Primers for real-time PCR (51015840 to 31015840)
Gene Forward primer Reverse primerLDLR aggacggctacagctaccc ctccaggcagatgttcacgPEPCK gctctgaggaggagaatgg tgctcttgggtgacgataacG6Pase gtgaattaccaagactcccag gcccatggcatggccagagggSREBP1c cgacatcgaagacatgcttcag ggaaggcttcaagagaggagcChREBP agagacaagatccgcctgaa cttccagtagttccctccaFAS gacatcgtccattcgtttgtg cggatcaccttcttgagctccSCD1 ggtgatgttccagaggaggta ggcagagtagtcataggaaaggGAPDH agccacatcgctcagacac gcccaatacgaccaaatcc
intracellular TG contents were determined by the Cell TGAssay Kit and normalized to protein concentrations mRNAlevels of lipogenic transcription factors and their target genes(Table 1) were determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR)
27 RNA Extraction and Real-Time RT-PCR Total RNA wasisolated from cells and reversely transcribed into cDNAsaccording to the supplierrsquos protocolsWe performed real-timePCR in an ABI Prism 7900 High-Throughput Real-TimePCR System (Applied Biosystems Foster City CA USA)with gene specific primers (Table 1) The reaction conditionwas the same as our previous report [25] For relative quan-tification of target genes the comparative threshold cycle(CT) method was used with glyceraldehyde-3-phosphatedehydrogenase (GAPDH) as an internal control The mRNAexpression levels of target geneswere plotted as fold of controlcells which were designated as 1
28 Glucose Consumption Assay HepG2 cells were seededonto 24-well plates with 2 times 105 cells per well For basalglucose consumption assay cells were treated with DMSO orthe studying compounds for 24 h as indicated with 4 replicatewells for each treatment For insulin-stimulated glucoseconsumption assay cells were treated with 005 nM of humaninsulin (diluted with DMEM + 05 FBS) together withDMSO or the studying compounds for 24 h Glucose levels inthe supernatant of media were assayed with a commerciallyavailable kit Glucose consumption was calculated as glucoselevel of the fresh mediumminus glucose level of the culturedmedium
29 Glucose Production Assay Cells were seeded and treatedas in the glucose consumption assay After treatment with thestudying compounds cells were washed twice with PBS Theglucose production medium was prepared by adding sodiumpyruvate and sodium L-lactate to the glucose- pyruvate-and phenol red-free DMEM to final concentrations of 2mMand 20mM respectively For one well of the 24-well platecells were loaded with 100120583L of the glucose productionmedium and incubated for 4 h at 37∘C Glucose concen-trations in the supernatant were analyzed by the AmplexRed GlucoseGlucose Oxidase Assay Kit according to thesupplierrsquos protocol The values were normalized to protein
4 BioMed Research International
concentrations and presented as percentages of DMSO con-trol which was defined as 100 In parallel experiments cellswere harvested for real-timeRT-PCRanalysis of the key genesinvolved in gluconeogenesis after 24 h treatment of studyingcompounds
210 Blocking Experiments After serum starvation cells werepretreatedwith 10120583MofCC (dissolved inDMSO) for 30minthen studying compounds or OA were added 24 h latercells were harvested for western blot CPT1 activity assay oilred O (ORO) staining intracellular TG assay basal glucoseconsumption assay glucose production assay or real-timeRT-PCR
211 Statistical Analysis Values are mean plusmn SD of 3-4repeated experiments After validation of the test for homo-geneity of variance differences among studying groups wereexamined by one-way ANOVA followed by the Newman-Keuls test for multiple comparisons 119901 lt 005was consideredto be statistically significant
3 Results
31 Cytotoxicities of Studying Compounds First we deter-mined the influences of MB saltMB on cell viability by theMTT method As shown in Figure 1(b) after 24 h treatmentM alone reduced the viability of HepG2 cells only when itsconcentration reached 400120583M (119901 lt 005 versus DMSO)TheIC50of M is larger than 400 120583M For B and MB salt cell via-
bilities declined slightly with no statistical significance whentheir concentrations reached 50120583M which was in agreementwith a previous report [29]When the concentrations of B andMB salt reached 100 120583M cell viabilities declined significantlyas compared to that of DMSO (119901 lt 005) And when theirconcentrations reached 400 120583M there were only averagely234 and 153 living cells left after 24 h treatment (119901 lt 001or 119901 lt 0001 versus DMSO)The IC
50values of B andMB salt
were 1339 plusmn 106 120583M and 1310 plusmn 94 120583M respectively Ourresult was close to a previous report inwhich the IC
50value of
B was 4233 120583gmL in HepG2 cells [30] We observed that thecells remained in good and healthy state if the concentrationsof B and MB salt did not exceed 50 120583M So the maximumconcentrations of the studying compounds we used in thefollowing experiments were 50 120583M
32 MB Salt Activates AMPK More Potently Than M or BAlone As shown in Figure 2(a) MB salt increased the levelsof p-AMPK120572 (Thr172) and p-ACC (Ser79) in dose-dependentmanners after 24 h of administration MB salt at 125120583Mcould stimulate the cellular AMPKpathway significantly (119901 lt005 versus DMSO) The stimulating activities of MB salt onthe phosphorylation of AMPK and ACC were completelyblocked by CC (Figure 2(b)) a specific inhibitor of AMPKTo compare the bioactivities of MB saltMB in stimulatingAMPK we used these compounds to treat HepG2 cells at anequal molar concentration As shown in Figure 2(c) whenadministered alone 25120583M of M and B could increase p-AMPK120572 (Thr172) and p-ACC (Ser79) levels by about 78ndash85 (119901 lt 005 versus DMSO) For comparison MB salt
at 25 120583M increased p-AMPK120572 (Thr172) and p-ACC (Ser79)levels averagely by 161- and 167-fold respectively (119901 lt 001versus DMSO) The efficacy of MB salt on the AMPKACCpathway was significantly superior to that of M or B alone(119901 lt 005)
We then determined the influences of the studying com-pounds on cellular CPT1 activity As shown in Figure 3(a)in HepG2 cells MB salt enhanced CPT1 activity in a mannersimilar to that of AMPK which could be abolished by CCpretreatment (Figure 3(b)) And again the activity of MB salton CPT1 was stronger than that of M or B when treated alone(119901 lt 005) (Figure 3(c)) Taken together the above resultsdemonstrate that MB salt is a potent AMPK activator and itsbioactivity is superior to that of M or B alone
33 MB Salt Suppresses Hepatic Steatosis More EffectivelyThan M or B Alone To investigate the influence of MB salton lipid metabolism HepG2 cells were challenged with OA[31] As shown in Figure 4 06mM of OA treatment for24 h induced steatosis and increased intracellular TG level(119901 lt 0001 versus untreated) dramatically in HepG2 cellsas determined by ORO staining and intracellular TG assayCoadministration ofMB salt prevented steatosis and reducedintracellular TG content in dose-dependent manners (datanot shown)The efficacies ofMB salt were completely blockedby CC pretreatment (Figure 4(a)) The MB salt suppressedhepatic steatosis and TG accumulation more effectively thanM or B when administered at an equal molar concentrationAs shown in Figure 4(b) 25120583MofMB salt treatment for 24 hreduced intracellular TG content averagely by 569 (119901 lt001 versus OA alone) which was superior to that of M orB alone (119901 lt 005)
In parallel with the development of steatosis the mRNAexpression levels of lipogenic transcription factors like sterolregulatory element-binding protein 1c (SREBP1c) and car-bohydrate responsive element-binding protein (ChREBP) aswell as their target genes like fatty acid synthase (FAS) andstearoyl-CoA desaturase 1 (SCD1) increased greatly after OAadministration (Figure 5 119901 lt 0001 versus untreated) TheMB salt suppressed the upregulation of the above genes indose-dependent (Figure 5(a)) and AMPK-dependent (Fig-ure 5(b)) manners In agreement with its stimulating activityon AMPK and inhibiting activity on steatosis the MBsalt reduced the expression levels of lipogenic genes moreeffectively than M or B alone (119901 lt 005 Figure 5(c))Taken together these results indicate that MB salt suppresseslipogenesis steatosis and TG accumulation in HpeG2 cellsthrough AMPK activation and its efficacies are more potentthan M or B alone
As B is able to upregulate LDLRmRNAexpression in livercells [22] we want to know whether or not M has the sameactivity As shown in Figure 6(a) M alone had no influenceon hepatic LDLR expression even when its concentrationreached 100 120583M On the contrary LDLR mRNA level couldbe upregulated by B at 125 120583M in HepG2 cells (119901 lt 005versusDMSO) (Figure 6(b))The activity ofMB salt on LDLRexpression was the same as that of B alone (Figure 6(b))
BioMed Research International 5
ACTB
ACC
p-ACC (Ser79)125DMSO 25 50
0
05
1
15
2
25
3
35
DMSO 125 25 50
p-ACC
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
lowastlowast
lowastlowast lowastlowast lowastlowast lowastlowast
MB salt (120583M)
MB salt (120583M)
p-AMPK120572 (Thr172)
AMPK120572
p-AMPK120572
(a)
ACC
p-ACC (Ser79)
ACTB
lowastlowast lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
MB salt (25 120583M)
CC (10120583M)
CC (10 120583M)
minus minus +
MB salt(25120583M) minus minus+ +
minus minus+ +
+
minus minus + +
0
05
1
15
2
25
3N
orm
aliz
ed le
vels
(fold
of D
MSO
)
(b)
DMSO M B MB salt
DMSO M B MB salt
ACTB
ACC
p-ACC (Ser79) lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
Treatments (25 120583M)
Treatments (25 120583M)
$lowastlowast$
lowastlowastlowastlowast
0
05
1
15
2
25
3
35
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
(c)
Figure 2 Stimulating effect of MB salt on the AMPK pathway After serum starvation cells were treated with different concentrations of MBsalt for 24 h (a) Alternatively cells were pretreated with CC for 30min thenMB salt was added and incubated for 24 h (b) In the comparisonexperiment an equal concentration ofMB saltMB was used to treat the cells for 24 h (c) DMSO (01) was used as control After treatmentcell total proteins were extracted the levels of p-AMPK120572 (Thr172) AMPK120572 p-ACC (Ser79) ACC and ACTB were determined by westernblot Representative blots are presented The protein levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) were normalized to those of AMPK120572and ACC respectively and plotted as fold of DMSO treated cells Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus those of M alone or B alone in (c)
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
2 BioMed Research International
M MW 4223 B MW 3364MB salt MW 7587
N
O
O
O
O
O +
OCH3
OCH3
HO
HO OH
OHOH
OHOH
OHmiddot
(a)
DMSO 25 50 100 200 400
MBMB salt
lowast
lowastlowast
lowastlowastlowastlowast lowastlowastlowastlowastlowast
Treatments (120583M)
0
20
40
60
80
100
120
Cel
l via
bilit
y (
of D
MSO
)
(b)
Figure 1 Influence of MB saltMB on cell viability (a) Chemical structure and molecular weight (MW) of MB salt (b) HepG2 cells weretreated with the studying compounds at different concentrations for 24 h Cell viability was determined by MTT staining and presented aspercentages of control cells which were treated with DMSO (05) and defined as 100 Values are mean plusmn SD of 3 separate experimentslowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO
chinensis B has a variety of pharmacological activities andnumerous studies demonstrated that it was a promising agentinmodulating lipid and glucosemetabolisms [3 20 21] NowB is undergoing clinical studies to systematically evaluate itsefficacy and safety The molecular mechanisms of B in mod-ulating lipid and glucose metabolisms which may includelow-density lipoprotein receptor (LDLR) upregulation [22]AMPK activation [21] and gut microbiota modulation [23]are not fully elucidated and still need further investigation
The compound of mangiferin-berberine (MBFigure 1(a)) salt was synthesized by chemical bondingof M and B at an equal molecular ratio [24] The nuclearmagnetic resonance (NMR) data demonstrated that in MBsalt M group (acidic) and B group (alkaline) conjugated byionic bond to form a stable single molecule [24] Due tothe well-defined roles of M and B in improving metabolicdisorders it can be expected that this new compound mayhave favourable activities in modulatingmetabolism Indeedin pilot studies the MB salt was found to stimulate theAMPK pathway in L6 skeletal muscle cells lower bloodglucose and lipids and improve insulin sensitivity andliver function in KK-Ay diabetic mice [24] However thedetailed activities and mechanisms of this new compoundare still undefined In the present report we study theefficacies of MB salt in HepG2 cells and find that it has potentactivities in stimulating AMPK andmodulating lipidglucosemetabolisms which are superior to those of M or B alone
2 Materials and Methods
21 Chemicals and Reagents The MB salt was synthesizedby Changzhou Deze Medical Science Co Ltd as describedpreviously [24] M and B were supplied by the North-east Pharmaceutical Group Shenyang No 1 PharmaceuticalCo Ltd (Shenyang China) Methylthiotetrazole (MTT)dimethyl sulphoxide (DMSO) compound C (CC) sodiumL-lactate oleic acid (OA) and bovine serum albumin (BSA)
were purchased from Sigma-Aldrich Co (St Louis MOUSA) Fetal bovine serum (FBS) Dulbeccorsquos modified Eaglersquosmedium (DMEM) glucose- pyruvate- and phenol red-freeDMEM sodium pyruvate andAmplex RedGlucoseGlucoseOxidase Assay Kit were purchased from Gibco-Invitrogen(Grand Island NY USA) Reagents required for proteinextraction and quantification were obtained from ThermoFisher Scientific Inc (Waltham MA USA) [25] Monoclonalantibodies specific for AMPK120572 phosphorylated AMPK120572 (p-AMPK120572) (Thr172) acetyl-CoA carboxylase (ACC) phos-phorylated ACC (p-ACC) (Ser79) and 120573-actin (ACTB)were obtained from Cell Signaling Technology Inc (Dan-vers MA USA) Immobilon-P polyvinylidene difluoride(PVDF) membranes were from EMDMillipore Corporation(Billerica MA USA) Cell carnitine palmitoyltransferase(CPT) 1 Activity Assay Kit was from GenMed ScientificsInc (Shanghai China) Steatosis Colorimetric Assay Kit waspurchased from Cayman Chemical (Ann Arbor MI USA)Cell Triglyceride (TG) Assay Kit was from the ApplygenTechnologies Inc (Beijing China) Reagents required forRNA isolation reverse transcription and real-time PCRwere purchased from Promega (Madison WI USA) [25]Glucose Assay Kit (based on glucose oxidase method) wasfrom Beijing Strong Biotechnologies Inc (Beijing China)Human insulin (Humulin) was purchased from Eli Lilly andCompany (Shanghai China)
22 Cell Culture HepG2 cells were routinely cultured inDMEM plus 10 FBS and appropriate antibiotics in anatmosphere of 5 CO
2at 37∘C Before experiments cells
were trypsinized and allowed to grow to about 70ndash80confluence The cells were starved in 05 FBS-containingmedium overnight before treatment
23 Cell Viability Assay HepG2 cells were seeded onto 96-well plates with 2 times 104 cells per well The studying com-pounds were dissolved with DMSO to make stock solutions
BioMed Research International 3
at a concentration of 80mM which were stored at minus20∘Cin aliquots Before usage the stock solutions were thawedand serially diluted with DMEM + 05 FBS After 24 h ofincubation and serum starvation cells were treated with thestudying compounds for 24 h as indicated (Figure 1(b)) eachtreatment had 5 replicate wells Control cells were treatedwith 05 DMSO in DMEM + 05 FBS a concentrationwith no cytotoxicity [26] and equal to the concentration ofDMSO in cells treated with 400120583M of studying compoundsCell viability was determined by MTT staining as describedpreviously [27] the absorbance was read by a VICTOR X4Multilabel Plate Reader (PerkinElmer Inc Waltham MAUSA) at a wavelength of 595 nm The results were presentedas percentages of control cells which were defined as 100Thevalues of 50 inhibiting concentrations (IC
50values) of the
compounds were calculated as described before [27]
24 Western Blot After treatment with the studying com-pounds for 24 h cell total proteins were extracted andquantified Samples containing about 20 120583g of protein wereused for 10 sodium dodecyl sulfate polyacrylamide gelelectrophoresis (SDS-PAGE)The blots were then transferredfrom gels onto PVDF membranes as described previously[25] After blocking the protein levels of AMPK120572 ACC andACTBwere detectedwith specificmonoclonal antibodies andan appropriate secondary antibody signals were developedwith an ECL kit (EMD Millipore Corporation) p-AMPK120572(Thr172) and p-ACC (Ser79) levels were examined by phos-phospecific antibodies after removal of antibody bindingfrom the membranes After scanning and quantificationthe levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) werenormalized to those of AMPK120572 and ACC and plotted asindicated
25 Cellular CPT1 Activity Assay After treatment for 24 hcells were harvested samples containing 50 120583g of proteinwere used for CPT1 activity assay according to the supplierrsquosprotocol The CPT1 experiments were repeated for 3 timescellular CPT1 activities were presented as nmol of coenzymeA (CoA) produced in the assay system per minute
26 Induction of Steatosis Oil Red O (ORO) Staining andIntracellular TGAssay OAwas dissolved in sterile phosphatebuffered saline (PBS) + 5 BSA [28] to make a stock solutionof 6mM and was stored at minus20∘C in aliquots Before usagethe stock solution was thawed and diluted with DMEM +05 FBS for 10 times HepG2 cells were seeded onto 6-wellplates with 5 times 105 cells per well After 24 h of incubationand serum starvation cells were left untreated or treated with06mM of OA for 24 h MB saltMB were serially dilutedfrom their stock solutions with DMEM + 05 FBS at thesame time of OA administration they were added to the cellsat indicated concentrations except for some cells which wereused as OA control After treatment intracellular lipids werestained with ORO by the Steatosis Colorimetric Assay Kitaccording to the supplierrsquos protocol After staining the cellswere observed under a light microscope and photographedIn parallel experiments cells were harvested after treatment
Table 1 Primers for real-time PCR (51015840 to 31015840)
Gene Forward primer Reverse primerLDLR aggacggctacagctaccc ctccaggcagatgttcacgPEPCK gctctgaggaggagaatgg tgctcttgggtgacgataacG6Pase gtgaattaccaagactcccag gcccatggcatggccagagggSREBP1c cgacatcgaagacatgcttcag ggaaggcttcaagagaggagcChREBP agagacaagatccgcctgaa cttccagtagttccctccaFAS gacatcgtccattcgtttgtg cggatcaccttcttgagctccSCD1 ggtgatgttccagaggaggta ggcagagtagtcataggaaaggGAPDH agccacatcgctcagacac gcccaatacgaccaaatcc
intracellular TG contents were determined by the Cell TGAssay Kit and normalized to protein concentrations mRNAlevels of lipogenic transcription factors and their target genes(Table 1) were determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR)
27 RNA Extraction and Real-Time RT-PCR Total RNA wasisolated from cells and reversely transcribed into cDNAsaccording to the supplierrsquos protocolsWe performed real-timePCR in an ABI Prism 7900 High-Throughput Real-TimePCR System (Applied Biosystems Foster City CA USA)with gene specific primers (Table 1) The reaction conditionwas the same as our previous report [25] For relative quan-tification of target genes the comparative threshold cycle(CT) method was used with glyceraldehyde-3-phosphatedehydrogenase (GAPDH) as an internal control The mRNAexpression levels of target geneswere plotted as fold of controlcells which were designated as 1
28 Glucose Consumption Assay HepG2 cells were seededonto 24-well plates with 2 times 105 cells per well For basalglucose consumption assay cells were treated with DMSO orthe studying compounds for 24 h as indicated with 4 replicatewells for each treatment For insulin-stimulated glucoseconsumption assay cells were treated with 005 nM of humaninsulin (diluted with DMEM + 05 FBS) together withDMSO or the studying compounds for 24 h Glucose levels inthe supernatant of media were assayed with a commerciallyavailable kit Glucose consumption was calculated as glucoselevel of the fresh mediumminus glucose level of the culturedmedium
29 Glucose Production Assay Cells were seeded and treatedas in the glucose consumption assay After treatment with thestudying compounds cells were washed twice with PBS Theglucose production medium was prepared by adding sodiumpyruvate and sodium L-lactate to the glucose- pyruvate-and phenol red-free DMEM to final concentrations of 2mMand 20mM respectively For one well of the 24-well platecells were loaded with 100120583L of the glucose productionmedium and incubated for 4 h at 37∘C Glucose concen-trations in the supernatant were analyzed by the AmplexRed GlucoseGlucose Oxidase Assay Kit according to thesupplierrsquos protocol The values were normalized to protein
4 BioMed Research International
concentrations and presented as percentages of DMSO con-trol which was defined as 100 In parallel experiments cellswere harvested for real-timeRT-PCRanalysis of the key genesinvolved in gluconeogenesis after 24 h treatment of studyingcompounds
210 Blocking Experiments After serum starvation cells werepretreatedwith 10120583MofCC (dissolved inDMSO) for 30minthen studying compounds or OA were added 24 h latercells were harvested for western blot CPT1 activity assay oilred O (ORO) staining intracellular TG assay basal glucoseconsumption assay glucose production assay or real-timeRT-PCR
211 Statistical Analysis Values are mean plusmn SD of 3-4repeated experiments After validation of the test for homo-geneity of variance differences among studying groups wereexamined by one-way ANOVA followed by the Newman-Keuls test for multiple comparisons 119901 lt 005was consideredto be statistically significant
3 Results
31 Cytotoxicities of Studying Compounds First we deter-mined the influences of MB saltMB on cell viability by theMTT method As shown in Figure 1(b) after 24 h treatmentM alone reduced the viability of HepG2 cells only when itsconcentration reached 400120583M (119901 lt 005 versus DMSO)TheIC50of M is larger than 400 120583M For B and MB salt cell via-
bilities declined slightly with no statistical significance whentheir concentrations reached 50120583M which was in agreementwith a previous report [29]When the concentrations of B andMB salt reached 100 120583M cell viabilities declined significantlyas compared to that of DMSO (119901 lt 005) And when theirconcentrations reached 400 120583M there were only averagely234 and 153 living cells left after 24 h treatment (119901 lt 001or 119901 lt 0001 versus DMSO)The IC
50values of B andMB salt
were 1339 plusmn 106 120583M and 1310 plusmn 94 120583M respectively Ourresult was close to a previous report inwhich the IC
50value of
B was 4233 120583gmL in HepG2 cells [30] We observed that thecells remained in good and healthy state if the concentrationsof B and MB salt did not exceed 50 120583M So the maximumconcentrations of the studying compounds we used in thefollowing experiments were 50 120583M
32 MB Salt Activates AMPK More Potently Than M or BAlone As shown in Figure 2(a) MB salt increased the levelsof p-AMPK120572 (Thr172) and p-ACC (Ser79) in dose-dependentmanners after 24 h of administration MB salt at 125120583Mcould stimulate the cellular AMPKpathway significantly (119901 lt005 versus DMSO) The stimulating activities of MB salt onthe phosphorylation of AMPK and ACC were completelyblocked by CC (Figure 2(b)) a specific inhibitor of AMPKTo compare the bioactivities of MB saltMB in stimulatingAMPK we used these compounds to treat HepG2 cells at anequal molar concentration As shown in Figure 2(c) whenadministered alone 25120583M of M and B could increase p-AMPK120572 (Thr172) and p-ACC (Ser79) levels by about 78ndash85 (119901 lt 005 versus DMSO) For comparison MB salt
at 25 120583M increased p-AMPK120572 (Thr172) and p-ACC (Ser79)levels averagely by 161- and 167-fold respectively (119901 lt 001versus DMSO) The efficacy of MB salt on the AMPKACCpathway was significantly superior to that of M or B alone(119901 lt 005)
We then determined the influences of the studying com-pounds on cellular CPT1 activity As shown in Figure 3(a)in HepG2 cells MB salt enhanced CPT1 activity in a mannersimilar to that of AMPK which could be abolished by CCpretreatment (Figure 3(b)) And again the activity of MB salton CPT1 was stronger than that of M or B when treated alone(119901 lt 005) (Figure 3(c)) Taken together the above resultsdemonstrate that MB salt is a potent AMPK activator and itsbioactivity is superior to that of M or B alone
33 MB Salt Suppresses Hepatic Steatosis More EffectivelyThan M or B Alone To investigate the influence of MB salton lipid metabolism HepG2 cells were challenged with OA[31] As shown in Figure 4 06mM of OA treatment for24 h induced steatosis and increased intracellular TG level(119901 lt 0001 versus untreated) dramatically in HepG2 cellsas determined by ORO staining and intracellular TG assayCoadministration ofMB salt prevented steatosis and reducedintracellular TG content in dose-dependent manners (datanot shown)The efficacies ofMB salt were completely blockedby CC pretreatment (Figure 4(a)) The MB salt suppressedhepatic steatosis and TG accumulation more effectively thanM or B when administered at an equal molar concentrationAs shown in Figure 4(b) 25120583MofMB salt treatment for 24 hreduced intracellular TG content averagely by 569 (119901 lt001 versus OA alone) which was superior to that of M orB alone (119901 lt 005)
In parallel with the development of steatosis the mRNAexpression levels of lipogenic transcription factors like sterolregulatory element-binding protein 1c (SREBP1c) and car-bohydrate responsive element-binding protein (ChREBP) aswell as their target genes like fatty acid synthase (FAS) andstearoyl-CoA desaturase 1 (SCD1) increased greatly after OAadministration (Figure 5 119901 lt 0001 versus untreated) TheMB salt suppressed the upregulation of the above genes indose-dependent (Figure 5(a)) and AMPK-dependent (Fig-ure 5(b)) manners In agreement with its stimulating activityon AMPK and inhibiting activity on steatosis the MBsalt reduced the expression levels of lipogenic genes moreeffectively than M or B alone (119901 lt 005 Figure 5(c))Taken together these results indicate that MB salt suppresseslipogenesis steatosis and TG accumulation in HpeG2 cellsthrough AMPK activation and its efficacies are more potentthan M or B alone
As B is able to upregulate LDLRmRNAexpression in livercells [22] we want to know whether or not M has the sameactivity As shown in Figure 6(a) M alone had no influenceon hepatic LDLR expression even when its concentrationreached 100 120583M On the contrary LDLR mRNA level couldbe upregulated by B at 125 120583M in HepG2 cells (119901 lt 005versusDMSO) (Figure 6(b))The activity ofMB salt on LDLRexpression was the same as that of B alone (Figure 6(b))
BioMed Research International 5
ACTB
ACC
p-ACC (Ser79)125DMSO 25 50
0
05
1
15
2
25
3
35
DMSO 125 25 50
p-ACC
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
lowastlowast
lowastlowast lowastlowast lowastlowast lowastlowast
MB salt (120583M)
MB salt (120583M)
p-AMPK120572 (Thr172)
AMPK120572
p-AMPK120572
(a)
ACC
p-ACC (Ser79)
ACTB
lowastlowast lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
MB salt (25 120583M)
CC (10120583M)
CC (10 120583M)
minus minus +
MB salt(25120583M) minus minus+ +
minus minus+ +
+
minus minus + +
0
05
1
15
2
25
3N
orm
aliz
ed le
vels
(fold
of D
MSO
)
(b)
DMSO M B MB salt
DMSO M B MB salt
ACTB
ACC
p-ACC (Ser79) lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
Treatments (25 120583M)
Treatments (25 120583M)
$lowastlowast$
lowastlowastlowastlowast
0
05
1
15
2
25
3
35
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
(c)
Figure 2 Stimulating effect of MB salt on the AMPK pathway After serum starvation cells were treated with different concentrations of MBsalt for 24 h (a) Alternatively cells were pretreated with CC for 30min thenMB salt was added and incubated for 24 h (b) In the comparisonexperiment an equal concentration ofMB saltMB was used to treat the cells for 24 h (c) DMSO (01) was used as control After treatmentcell total proteins were extracted the levels of p-AMPK120572 (Thr172) AMPK120572 p-ACC (Ser79) ACC and ACTB were determined by westernblot Representative blots are presented The protein levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) were normalized to those of AMPK120572and ACC respectively and plotted as fold of DMSO treated cells Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus those of M alone or B alone in (c)
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 3
at a concentration of 80mM which were stored at minus20∘Cin aliquots Before usage the stock solutions were thawedand serially diluted with DMEM + 05 FBS After 24 h ofincubation and serum starvation cells were treated with thestudying compounds for 24 h as indicated (Figure 1(b)) eachtreatment had 5 replicate wells Control cells were treatedwith 05 DMSO in DMEM + 05 FBS a concentrationwith no cytotoxicity [26] and equal to the concentration ofDMSO in cells treated with 400120583M of studying compoundsCell viability was determined by MTT staining as describedpreviously [27] the absorbance was read by a VICTOR X4Multilabel Plate Reader (PerkinElmer Inc Waltham MAUSA) at a wavelength of 595 nm The results were presentedas percentages of control cells which were defined as 100Thevalues of 50 inhibiting concentrations (IC
50values) of the
compounds were calculated as described before [27]
24 Western Blot After treatment with the studying com-pounds for 24 h cell total proteins were extracted andquantified Samples containing about 20 120583g of protein wereused for 10 sodium dodecyl sulfate polyacrylamide gelelectrophoresis (SDS-PAGE)The blots were then transferredfrom gels onto PVDF membranes as described previously[25] After blocking the protein levels of AMPK120572 ACC andACTBwere detectedwith specificmonoclonal antibodies andan appropriate secondary antibody signals were developedwith an ECL kit (EMD Millipore Corporation) p-AMPK120572(Thr172) and p-ACC (Ser79) levels were examined by phos-phospecific antibodies after removal of antibody bindingfrom the membranes After scanning and quantificationthe levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) werenormalized to those of AMPK120572 and ACC and plotted asindicated
25 Cellular CPT1 Activity Assay After treatment for 24 hcells were harvested samples containing 50 120583g of proteinwere used for CPT1 activity assay according to the supplierrsquosprotocol The CPT1 experiments were repeated for 3 timescellular CPT1 activities were presented as nmol of coenzymeA (CoA) produced in the assay system per minute
26 Induction of Steatosis Oil Red O (ORO) Staining andIntracellular TGAssay OAwas dissolved in sterile phosphatebuffered saline (PBS) + 5 BSA [28] to make a stock solutionof 6mM and was stored at minus20∘C in aliquots Before usagethe stock solution was thawed and diluted with DMEM +05 FBS for 10 times HepG2 cells were seeded onto 6-wellplates with 5 times 105 cells per well After 24 h of incubationand serum starvation cells were left untreated or treated with06mM of OA for 24 h MB saltMB were serially dilutedfrom their stock solutions with DMEM + 05 FBS at thesame time of OA administration they were added to the cellsat indicated concentrations except for some cells which wereused as OA control After treatment intracellular lipids werestained with ORO by the Steatosis Colorimetric Assay Kitaccording to the supplierrsquos protocol After staining the cellswere observed under a light microscope and photographedIn parallel experiments cells were harvested after treatment
Table 1 Primers for real-time PCR (51015840 to 31015840)
Gene Forward primer Reverse primerLDLR aggacggctacagctaccc ctccaggcagatgttcacgPEPCK gctctgaggaggagaatgg tgctcttgggtgacgataacG6Pase gtgaattaccaagactcccag gcccatggcatggccagagggSREBP1c cgacatcgaagacatgcttcag ggaaggcttcaagagaggagcChREBP agagacaagatccgcctgaa cttccagtagttccctccaFAS gacatcgtccattcgtttgtg cggatcaccttcttgagctccSCD1 ggtgatgttccagaggaggta ggcagagtagtcataggaaaggGAPDH agccacatcgctcagacac gcccaatacgaccaaatcc
intracellular TG contents were determined by the Cell TGAssay Kit and normalized to protein concentrations mRNAlevels of lipogenic transcription factors and their target genes(Table 1) were determined by real-time reverse transcriptase-polymerase chain reaction (RT-PCR)
27 RNA Extraction and Real-Time RT-PCR Total RNA wasisolated from cells and reversely transcribed into cDNAsaccording to the supplierrsquos protocolsWe performed real-timePCR in an ABI Prism 7900 High-Throughput Real-TimePCR System (Applied Biosystems Foster City CA USA)with gene specific primers (Table 1) The reaction conditionwas the same as our previous report [25] For relative quan-tification of target genes the comparative threshold cycle(CT) method was used with glyceraldehyde-3-phosphatedehydrogenase (GAPDH) as an internal control The mRNAexpression levels of target geneswere plotted as fold of controlcells which were designated as 1
28 Glucose Consumption Assay HepG2 cells were seededonto 24-well plates with 2 times 105 cells per well For basalglucose consumption assay cells were treated with DMSO orthe studying compounds for 24 h as indicated with 4 replicatewells for each treatment For insulin-stimulated glucoseconsumption assay cells were treated with 005 nM of humaninsulin (diluted with DMEM + 05 FBS) together withDMSO or the studying compounds for 24 h Glucose levels inthe supernatant of media were assayed with a commerciallyavailable kit Glucose consumption was calculated as glucoselevel of the fresh mediumminus glucose level of the culturedmedium
29 Glucose Production Assay Cells were seeded and treatedas in the glucose consumption assay After treatment with thestudying compounds cells were washed twice with PBS Theglucose production medium was prepared by adding sodiumpyruvate and sodium L-lactate to the glucose- pyruvate-and phenol red-free DMEM to final concentrations of 2mMand 20mM respectively For one well of the 24-well platecells were loaded with 100120583L of the glucose productionmedium and incubated for 4 h at 37∘C Glucose concen-trations in the supernatant were analyzed by the AmplexRed GlucoseGlucose Oxidase Assay Kit according to thesupplierrsquos protocol The values were normalized to protein
4 BioMed Research International
concentrations and presented as percentages of DMSO con-trol which was defined as 100 In parallel experiments cellswere harvested for real-timeRT-PCRanalysis of the key genesinvolved in gluconeogenesis after 24 h treatment of studyingcompounds
210 Blocking Experiments After serum starvation cells werepretreatedwith 10120583MofCC (dissolved inDMSO) for 30minthen studying compounds or OA were added 24 h latercells were harvested for western blot CPT1 activity assay oilred O (ORO) staining intracellular TG assay basal glucoseconsumption assay glucose production assay or real-timeRT-PCR
211 Statistical Analysis Values are mean plusmn SD of 3-4repeated experiments After validation of the test for homo-geneity of variance differences among studying groups wereexamined by one-way ANOVA followed by the Newman-Keuls test for multiple comparisons 119901 lt 005was consideredto be statistically significant
3 Results
31 Cytotoxicities of Studying Compounds First we deter-mined the influences of MB saltMB on cell viability by theMTT method As shown in Figure 1(b) after 24 h treatmentM alone reduced the viability of HepG2 cells only when itsconcentration reached 400120583M (119901 lt 005 versus DMSO)TheIC50of M is larger than 400 120583M For B and MB salt cell via-
bilities declined slightly with no statistical significance whentheir concentrations reached 50120583M which was in agreementwith a previous report [29]When the concentrations of B andMB salt reached 100 120583M cell viabilities declined significantlyas compared to that of DMSO (119901 lt 005) And when theirconcentrations reached 400 120583M there were only averagely234 and 153 living cells left after 24 h treatment (119901 lt 001or 119901 lt 0001 versus DMSO)The IC
50values of B andMB salt
were 1339 plusmn 106 120583M and 1310 plusmn 94 120583M respectively Ourresult was close to a previous report inwhich the IC
50value of
B was 4233 120583gmL in HepG2 cells [30] We observed that thecells remained in good and healthy state if the concentrationsof B and MB salt did not exceed 50 120583M So the maximumconcentrations of the studying compounds we used in thefollowing experiments were 50 120583M
32 MB Salt Activates AMPK More Potently Than M or BAlone As shown in Figure 2(a) MB salt increased the levelsof p-AMPK120572 (Thr172) and p-ACC (Ser79) in dose-dependentmanners after 24 h of administration MB salt at 125120583Mcould stimulate the cellular AMPKpathway significantly (119901 lt005 versus DMSO) The stimulating activities of MB salt onthe phosphorylation of AMPK and ACC were completelyblocked by CC (Figure 2(b)) a specific inhibitor of AMPKTo compare the bioactivities of MB saltMB in stimulatingAMPK we used these compounds to treat HepG2 cells at anequal molar concentration As shown in Figure 2(c) whenadministered alone 25120583M of M and B could increase p-AMPK120572 (Thr172) and p-ACC (Ser79) levels by about 78ndash85 (119901 lt 005 versus DMSO) For comparison MB salt
at 25 120583M increased p-AMPK120572 (Thr172) and p-ACC (Ser79)levels averagely by 161- and 167-fold respectively (119901 lt 001versus DMSO) The efficacy of MB salt on the AMPKACCpathway was significantly superior to that of M or B alone(119901 lt 005)
We then determined the influences of the studying com-pounds on cellular CPT1 activity As shown in Figure 3(a)in HepG2 cells MB salt enhanced CPT1 activity in a mannersimilar to that of AMPK which could be abolished by CCpretreatment (Figure 3(b)) And again the activity of MB salton CPT1 was stronger than that of M or B when treated alone(119901 lt 005) (Figure 3(c)) Taken together the above resultsdemonstrate that MB salt is a potent AMPK activator and itsbioactivity is superior to that of M or B alone
33 MB Salt Suppresses Hepatic Steatosis More EffectivelyThan M or B Alone To investigate the influence of MB salton lipid metabolism HepG2 cells were challenged with OA[31] As shown in Figure 4 06mM of OA treatment for24 h induced steatosis and increased intracellular TG level(119901 lt 0001 versus untreated) dramatically in HepG2 cellsas determined by ORO staining and intracellular TG assayCoadministration ofMB salt prevented steatosis and reducedintracellular TG content in dose-dependent manners (datanot shown)The efficacies ofMB salt were completely blockedby CC pretreatment (Figure 4(a)) The MB salt suppressedhepatic steatosis and TG accumulation more effectively thanM or B when administered at an equal molar concentrationAs shown in Figure 4(b) 25120583MofMB salt treatment for 24 hreduced intracellular TG content averagely by 569 (119901 lt001 versus OA alone) which was superior to that of M orB alone (119901 lt 005)
In parallel with the development of steatosis the mRNAexpression levels of lipogenic transcription factors like sterolregulatory element-binding protein 1c (SREBP1c) and car-bohydrate responsive element-binding protein (ChREBP) aswell as their target genes like fatty acid synthase (FAS) andstearoyl-CoA desaturase 1 (SCD1) increased greatly after OAadministration (Figure 5 119901 lt 0001 versus untreated) TheMB salt suppressed the upregulation of the above genes indose-dependent (Figure 5(a)) and AMPK-dependent (Fig-ure 5(b)) manners In agreement with its stimulating activityon AMPK and inhibiting activity on steatosis the MBsalt reduced the expression levels of lipogenic genes moreeffectively than M or B alone (119901 lt 005 Figure 5(c))Taken together these results indicate that MB salt suppresseslipogenesis steatosis and TG accumulation in HpeG2 cellsthrough AMPK activation and its efficacies are more potentthan M or B alone
As B is able to upregulate LDLRmRNAexpression in livercells [22] we want to know whether or not M has the sameactivity As shown in Figure 6(a) M alone had no influenceon hepatic LDLR expression even when its concentrationreached 100 120583M On the contrary LDLR mRNA level couldbe upregulated by B at 125 120583M in HepG2 cells (119901 lt 005versusDMSO) (Figure 6(b))The activity ofMB salt on LDLRexpression was the same as that of B alone (Figure 6(b))
BioMed Research International 5
ACTB
ACC
p-ACC (Ser79)125DMSO 25 50
0
05
1
15
2
25
3
35
DMSO 125 25 50
p-ACC
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
lowastlowast
lowastlowast lowastlowast lowastlowast lowastlowast
MB salt (120583M)
MB salt (120583M)
p-AMPK120572 (Thr172)
AMPK120572
p-AMPK120572
(a)
ACC
p-ACC (Ser79)
ACTB
lowastlowast lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
MB salt (25 120583M)
CC (10120583M)
CC (10 120583M)
minus minus +
MB salt(25120583M) minus minus+ +
minus minus+ +
+
minus minus + +
0
05
1
15
2
25
3N
orm
aliz
ed le
vels
(fold
of D
MSO
)
(b)
DMSO M B MB salt
DMSO M B MB salt
ACTB
ACC
p-ACC (Ser79) lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
Treatments (25 120583M)
Treatments (25 120583M)
$lowastlowast$
lowastlowastlowastlowast
0
05
1
15
2
25
3
35
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
(c)
Figure 2 Stimulating effect of MB salt on the AMPK pathway After serum starvation cells were treated with different concentrations of MBsalt for 24 h (a) Alternatively cells were pretreated with CC for 30min thenMB salt was added and incubated for 24 h (b) In the comparisonexperiment an equal concentration ofMB saltMB was used to treat the cells for 24 h (c) DMSO (01) was used as control After treatmentcell total proteins were extracted the levels of p-AMPK120572 (Thr172) AMPK120572 p-ACC (Ser79) ACC and ACTB were determined by westernblot Representative blots are presented The protein levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) were normalized to those of AMPK120572and ACC respectively and plotted as fold of DMSO treated cells Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus those of M alone or B alone in (c)
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

4 BioMed Research International
concentrations and presented as percentages of DMSO con-trol which was defined as 100 In parallel experiments cellswere harvested for real-timeRT-PCRanalysis of the key genesinvolved in gluconeogenesis after 24 h treatment of studyingcompounds
210 Blocking Experiments After serum starvation cells werepretreatedwith 10120583MofCC (dissolved inDMSO) for 30minthen studying compounds or OA were added 24 h latercells were harvested for western blot CPT1 activity assay oilred O (ORO) staining intracellular TG assay basal glucoseconsumption assay glucose production assay or real-timeRT-PCR
211 Statistical Analysis Values are mean plusmn SD of 3-4repeated experiments After validation of the test for homo-geneity of variance differences among studying groups wereexamined by one-way ANOVA followed by the Newman-Keuls test for multiple comparisons 119901 lt 005was consideredto be statistically significant
3 Results
31 Cytotoxicities of Studying Compounds First we deter-mined the influences of MB saltMB on cell viability by theMTT method As shown in Figure 1(b) after 24 h treatmentM alone reduced the viability of HepG2 cells only when itsconcentration reached 400120583M (119901 lt 005 versus DMSO)TheIC50of M is larger than 400 120583M For B and MB salt cell via-
bilities declined slightly with no statistical significance whentheir concentrations reached 50120583M which was in agreementwith a previous report [29]When the concentrations of B andMB salt reached 100 120583M cell viabilities declined significantlyas compared to that of DMSO (119901 lt 005) And when theirconcentrations reached 400 120583M there were only averagely234 and 153 living cells left after 24 h treatment (119901 lt 001or 119901 lt 0001 versus DMSO)The IC
50values of B andMB salt
were 1339 plusmn 106 120583M and 1310 plusmn 94 120583M respectively Ourresult was close to a previous report inwhich the IC
50value of
B was 4233 120583gmL in HepG2 cells [30] We observed that thecells remained in good and healthy state if the concentrationsof B and MB salt did not exceed 50 120583M So the maximumconcentrations of the studying compounds we used in thefollowing experiments were 50 120583M
32 MB Salt Activates AMPK More Potently Than M or BAlone As shown in Figure 2(a) MB salt increased the levelsof p-AMPK120572 (Thr172) and p-ACC (Ser79) in dose-dependentmanners after 24 h of administration MB salt at 125120583Mcould stimulate the cellular AMPKpathway significantly (119901 lt005 versus DMSO) The stimulating activities of MB salt onthe phosphorylation of AMPK and ACC were completelyblocked by CC (Figure 2(b)) a specific inhibitor of AMPKTo compare the bioactivities of MB saltMB in stimulatingAMPK we used these compounds to treat HepG2 cells at anequal molar concentration As shown in Figure 2(c) whenadministered alone 25120583M of M and B could increase p-AMPK120572 (Thr172) and p-ACC (Ser79) levels by about 78ndash85 (119901 lt 005 versus DMSO) For comparison MB salt
at 25 120583M increased p-AMPK120572 (Thr172) and p-ACC (Ser79)levels averagely by 161- and 167-fold respectively (119901 lt 001versus DMSO) The efficacy of MB salt on the AMPKACCpathway was significantly superior to that of M or B alone(119901 lt 005)
We then determined the influences of the studying com-pounds on cellular CPT1 activity As shown in Figure 3(a)in HepG2 cells MB salt enhanced CPT1 activity in a mannersimilar to that of AMPK which could be abolished by CCpretreatment (Figure 3(b)) And again the activity of MB salton CPT1 was stronger than that of M or B when treated alone(119901 lt 005) (Figure 3(c)) Taken together the above resultsdemonstrate that MB salt is a potent AMPK activator and itsbioactivity is superior to that of M or B alone
33 MB Salt Suppresses Hepatic Steatosis More EffectivelyThan M or B Alone To investigate the influence of MB salton lipid metabolism HepG2 cells were challenged with OA[31] As shown in Figure 4 06mM of OA treatment for24 h induced steatosis and increased intracellular TG level(119901 lt 0001 versus untreated) dramatically in HepG2 cellsas determined by ORO staining and intracellular TG assayCoadministration ofMB salt prevented steatosis and reducedintracellular TG content in dose-dependent manners (datanot shown)The efficacies ofMB salt were completely blockedby CC pretreatment (Figure 4(a)) The MB salt suppressedhepatic steatosis and TG accumulation more effectively thanM or B when administered at an equal molar concentrationAs shown in Figure 4(b) 25120583MofMB salt treatment for 24 hreduced intracellular TG content averagely by 569 (119901 lt001 versus OA alone) which was superior to that of M orB alone (119901 lt 005)
In parallel with the development of steatosis the mRNAexpression levels of lipogenic transcription factors like sterolregulatory element-binding protein 1c (SREBP1c) and car-bohydrate responsive element-binding protein (ChREBP) aswell as their target genes like fatty acid synthase (FAS) andstearoyl-CoA desaturase 1 (SCD1) increased greatly after OAadministration (Figure 5 119901 lt 0001 versus untreated) TheMB salt suppressed the upregulation of the above genes indose-dependent (Figure 5(a)) and AMPK-dependent (Fig-ure 5(b)) manners In agreement with its stimulating activityon AMPK and inhibiting activity on steatosis the MBsalt reduced the expression levels of lipogenic genes moreeffectively than M or B alone (119901 lt 005 Figure 5(c))Taken together these results indicate that MB salt suppresseslipogenesis steatosis and TG accumulation in HpeG2 cellsthrough AMPK activation and its efficacies are more potentthan M or B alone
As B is able to upregulate LDLRmRNAexpression in livercells [22] we want to know whether or not M has the sameactivity As shown in Figure 6(a) M alone had no influenceon hepatic LDLR expression even when its concentrationreached 100 120583M On the contrary LDLR mRNA level couldbe upregulated by B at 125 120583M in HepG2 cells (119901 lt 005versusDMSO) (Figure 6(b))The activity ofMB salt on LDLRexpression was the same as that of B alone (Figure 6(b))
BioMed Research International 5
ACTB
ACC
p-ACC (Ser79)125DMSO 25 50
0
05
1
15
2
25
3
35
DMSO 125 25 50
p-ACC
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
lowastlowast
lowastlowast lowastlowast lowastlowast lowastlowast
MB salt (120583M)
MB salt (120583M)
p-AMPK120572 (Thr172)
AMPK120572
p-AMPK120572
(a)
ACC
p-ACC (Ser79)
ACTB
lowastlowast lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
MB salt (25 120583M)
CC (10120583M)
CC (10 120583M)
minus minus +
MB salt(25120583M) minus minus+ +
minus minus+ +
+
minus minus + +
0
05
1
15
2
25
3N
orm
aliz
ed le
vels
(fold
of D
MSO
)
(b)
DMSO M B MB salt
DMSO M B MB salt
ACTB
ACC
p-ACC (Ser79) lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
Treatments (25 120583M)
Treatments (25 120583M)
$lowastlowast$
lowastlowastlowastlowast
0
05
1
15
2
25
3
35
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
(c)
Figure 2 Stimulating effect of MB salt on the AMPK pathway After serum starvation cells were treated with different concentrations of MBsalt for 24 h (a) Alternatively cells were pretreated with CC for 30min thenMB salt was added and incubated for 24 h (b) In the comparisonexperiment an equal concentration ofMB saltMB was used to treat the cells for 24 h (c) DMSO (01) was used as control After treatmentcell total proteins were extracted the levels of p-AMPK120572 (Thr172) AMPK120572 p-ACC (Ser79) ACC and ACTB were determined by westernblot Representative blots are presented The protein levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) were normalized to those of AMPK120572and ACC respectively and plotted as fold of DMSO treated cells Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus those of M alone or B alone in (c)
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
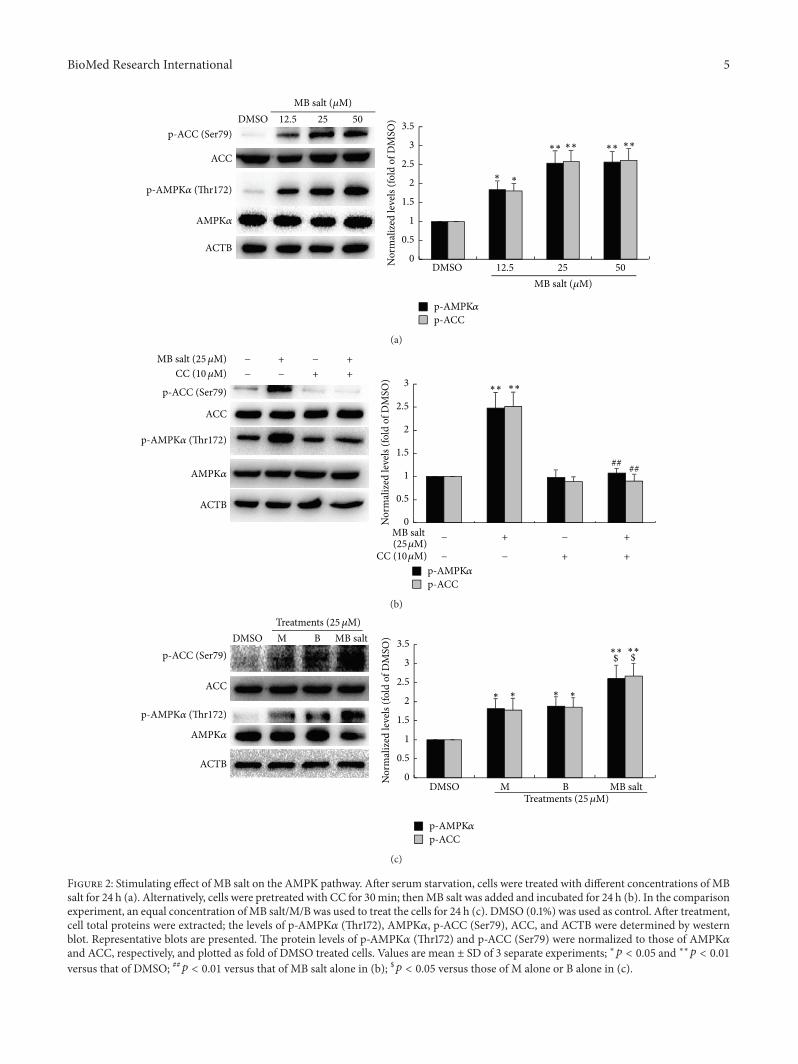
BioMed Research International 5
ACTB
ACC
p-ACC (Ser79)125DMSO 25 50
0
05
1
15
2
25
3
35
DMSO 125 25 50
p-ACC
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
lowastlowast
lowastlowast lowastlowast lowastlowast lowastlowast
MB salt (120583M)
MB salt (120583M)
p-AMPK120572 (Thr172)
AMPK120572
p-AMPK120572
(a)
ACC
p-ACC (Ser79)
ACTB
lowastlowast lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
MB salt (25 120583M)
CC (10120583M)
CC (10 120583M)
minus minus +
MB salt(25120583M) minus minus+ +
minus minus+ +
+
minus minus + +
0
05
1
15
2
25
3N
orm
aliz
ed le
vels
(fold
of D
MSO
)
(b)
DMSO M B MB salt
DMSO M B MB salt
ACTB
ACC
p-ACC (Ser79) lowastlowast
p-AMPK120572 (Thr172)
AMPK120572
p-ACCp-AMPK120572
Treatments (25 120583M)
Treatments (25 120583M)
$lowastlowast$
lowastlowastlowastlowast
0
05
1
15
2
25
3
35
Nor
mal
ized
leve
ls (fo
ld o
f DM
SO)
(c)
Figure 2 Stimulating effect of MB salt on the AMPK pathway After serum starvation cells were treated with different concentrations of MBsalt for 24 h (a) Alternatively cells were pretreated with CC for 30min thenMB salt was added and incubated for 24 h (b) In the comparisonexperiment an equal concentration ofMB saltMB was used to treat the cells for 24 h (c) DMSO (01) was used as control After treatmentcell total proteins were extracted the levels of p-AMPK120572 (Thr172) AMPK120572 p-ACC (Ser79) ACC and ACTB were determined by westernblot Representative blots are presented The protein levels of p-AMPK120572 (Thr172) and p-ACC (Ser79) were normalized to those of AMPK120572and ACC respectively and plotted as fold of DMSO treated cells Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus those of M alone or B alone in (c)
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
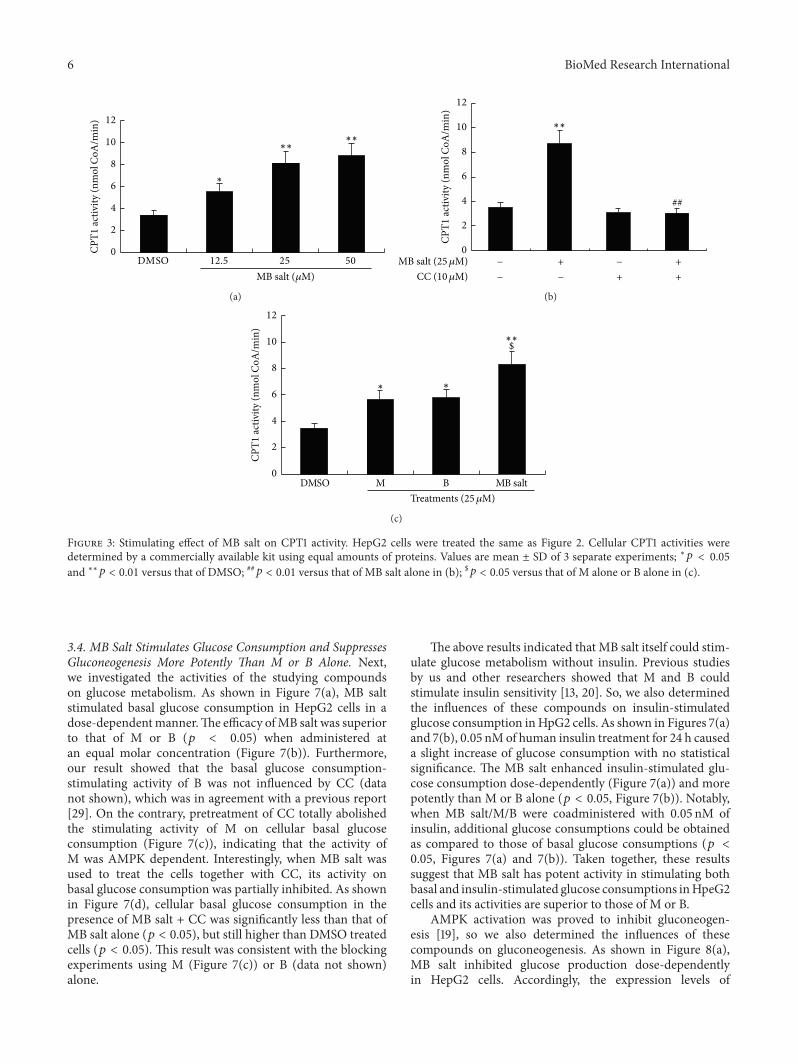
6 BioMed Research International
DMSO 125 25 50
lowast
lowastlowastlowastlowast
MB salt (120583M)
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(a)
lowastlowast
MB salt (25 120583M)CC (10 120583M)
minus
minusminus
minus +
+
+
+
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
(b)
DMSO M B MB salt
lowastlowast$
lowast lowast
0
2
4
6
8
10
12
CPT1
activ
ity (n
mol
CoA
min
)
Treatments (25 120583M)
(c)
Figure 3 Stimulating effect of MB salt on CPT1 activity HepG2 cells were treated the same as Figure 2 Cellular CPT1 activities weredetermined by a commercially available kit using equal amounts of proteins Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 001 versus that of MB salt alone in (b) $119901 lt 005 versus that of M alone or B alone in (c)
34 MB Salt Stimulates Glucose Consumption and SuppressesGluconeogenesis More Potently Than M or B Alone Nextwe investigated the activities of the studying compoundson glucose metabolism As shown in Figure 7(a) MB saltstimulated basal glucose consumption in HepG2 cells in adose-dependentmannerThe efficacy ofMB salt was superiorto that of M or B (119901 lt 005) when administered atan equal molar concentration (Figure 7(b)) Furthermoreour result showed that the basal glucose consumption-stimulating activity of B was not influenced by CC (datanot shown) which was in agreement with a previous report[29] On the contrary pretreatment of CC totally abolishedthe stimulating activity of M on cellular basal glucoseconsumption (Figure 7(c)) indicating that the activity ofM was AMPK dependent Interestingly when MB salt wasused to treat the cells together with CC its activity onbasal glucose consumption was partially inhibited As shownin Figure 7(d) cellular basal glucose consumption in thepresence of MB salt + CC was significantly less than that ofMB salt alone (119901 lt 005) but still higher than DMSO treatedcells (119901 lt 005) This result was consistent with the blockingexperiments using M (Figure 7(c)) or B (data not shown)alone
The above results indicated that MB salt itself could stim-ulate glucose metabolism without insulin Previous studiesby us and other researchers showed that M and B couldstimulate insulin sensitivity [13 20] So we also determinedthe influences of these compounds on insulin-stimulatedglucose consumption inHpG2 cells As shown in Figures 7(a)and 7(b) 005 nMof human insulin treatment for 24 h causeda slight increase of glucose consumption with no statisticalsignificance The MB salt enhanced insulin-stimulated glu-cose consumption dose-dependently (Figure 7(a)) and morepotently than M or B alone (119901 lt 005 Figure 7(b)) Notablywhen MB saltMB were coadministered with 005 nM ofinsulin additional glucose consumptions could be obtainedas compared to those of basal glucose consumptions (119901 lt005 Figures 7(a) and 7(b)) Taken together these resultssuggest that MB salt has potent activity in stimulating bothbasal and insulin-stimulated glucose consumptions inHpeG2cells and its activities are superior to those of M or B
AMPK activation was proved to inhibit gluconeogen-esis [19] so we also determined the influences of thesecompounds on gluconeogenesis As shown in Figure 8(a)MB salt inhibited glucose production dose-dependentlyin HepG2 cells Accordingly the expression levels of
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
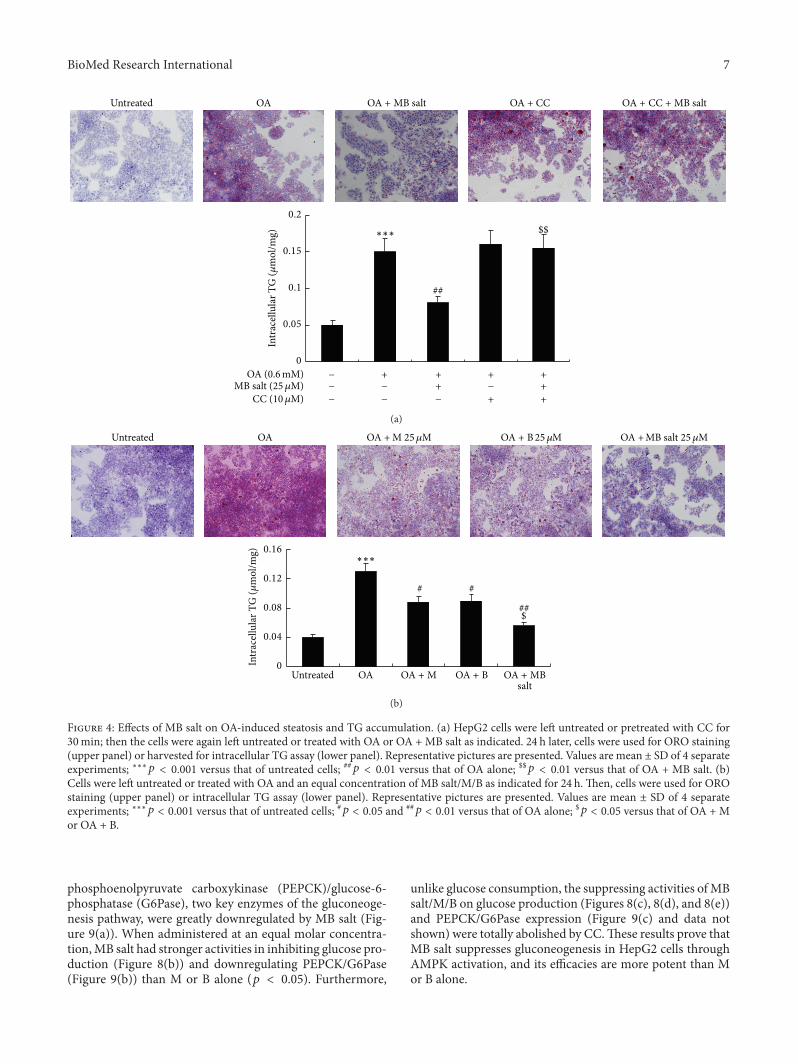
BioMed Research International 7
Untreated OA OA + MB salt OA + CC OA + CC + MB salt
$$lowastlowastlowast
Intr
acel
lula
r TG
(120583m
olm
g)
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
+ + + +++
+ +
minusminusminus
minus minus minus
minus
0
005
01
015
02
(a)
Untreated OA OA + M 25 120583M OA + B 25 120583M OA + MB salt 25 120583M
$
lowastlowastlowast
Untreated OA OA + M OA + B OA + MBsalt
Intr
acel
lula
r TG
(120583m
olm
g)
0
004
008
012
016
(b)
Figure 4 Effects of MB salt on OA-induced steatosis and TG accumulation (a) HepG2 cells were left untreated or pretreated with CC for30min then the cells were again left untreated or treated with OA or OA + MB salt as indicated 24 h later cells were used for ORO staining(upper panel) or harvested for intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001 versus that of OA + MB salt (b)Cells were left untreated or treated with OA and an equal concentration of MB saltMB as indicated for 24 h Then cells were used for OROstaining (upper panel) or intracellular TG assay (lower panel) Representative pictures are presented Values are mean plusmn SD of 4 separateexperiments lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + Mor OA + B
phosphoenolpyruvate carboxykinase (PEPCK)glucose-6-phosphatase (G6Pase) two key enzymes of the gluconeoge-nesis pathway were greatly downregulated by MB salt (Fig-ure 9(a)) When administered at an equal molar concentra-tionMB salt had stronger activities in inhibiting glucose pro-duction (Figure 8(b)) and downregulating PEPCKG6Pase(Figure 9(b)) than M or B alone (119901 lt 005) Furthermore
unlike glucose consumption the suppressing activities ofMBsaltMB on glucose production (Figures 8(c) 8(d) and 8(e))and PEPCKG6Pase expression (Figure 9(c) and data notshown) were totally abolished by CCThese results prove thatMB salt suppresses gluconeogenesis in HepG2 cells throughAMPK activation and its efficacies are more potent than Mor B alone
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
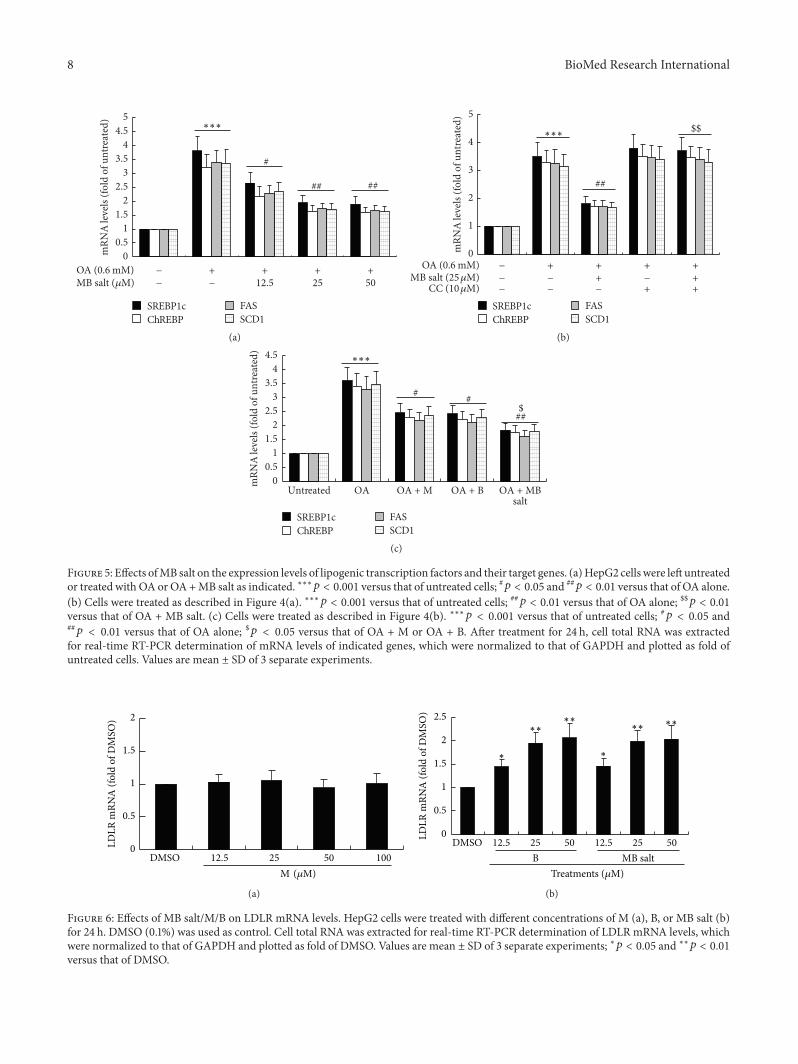
8 BioMed Research International
SREBP1cChREBP
FASSCD1
125 25 50
lowastlowastlowast
minusminus minus
++++OA (06 mM)MB salt (120583M)
005
115
225
335
445
5m
RNA
leve
ls (fo
ld o
f unt
reat
ed)
(a)
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$$
minusminusminus minus
minusminus
minus++++
+ +++
OA (06 mM)MB salt (25 120583M)
CC (10 120583M)
0
1
2
3
4
5
mRN
A le
vels
(fold
of u
ntre
ated
)
(b)
Untreated OA OA + M OA + B OA + MBsalt
SREBP1cChREBP
FASSCD1
lowastlowastlowast
$
005
115
225
335
445
mRN
A le
vels
(fold
of u
ntre
ated
)
(c)
Figure 5 Effects ofMB salt on the expression levels of lipogenic transcription factors and their target genes (a)HepG2 cells were left untreatedor treated with OA or OA+MB salt as indicated lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and
119901 lt 001 versus that of OA alone(b) Cells were treated as described in Figure 4(a) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 001 versus that of OA alone $$119901 lt 001versus that of OA + MB salt (c) Cells were treated as described in Figure 4(b) lowastlowastlowast119901 lt 0001 versus that of untreated cells 119901 lt 005 and119901 lt 001 versus that of OA alone $119901 lt 005 versus that of OA + M or OA + B After treatment for 24 h cell total RNA was extracted
for real-time RT-PCR determination of mRNA levels of indicated genes which were normalized to that of GAPDH and plotted as fold ofuntreated cells Values are mean plusmn SD of 3 separate experiments
DMSO 125 25 50 1000
05
1
15
2
LDLR
mRN
A (f
old
of D
MSO
)
M (120583M)
(a)
DMSO 125 25 50 125 25 50B MB salt
0
05
1
15
2
25
LDLR
mRN
A (f
old
of D
MSO
)
Treatments (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowast
(b)
Figure 6 Effects of MB saltMB on LDLR mRNA levels HepG2 cells were treated with different concentrations of M (a) B or MB salt (b)for 24 h DMSO (01) was used as control Cell total RNA was extracted for real-time RT-PCR determination of LDLR mRNA levels whichwere normalized to that of GAPDH and plotted as fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001versus that of DMSO
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
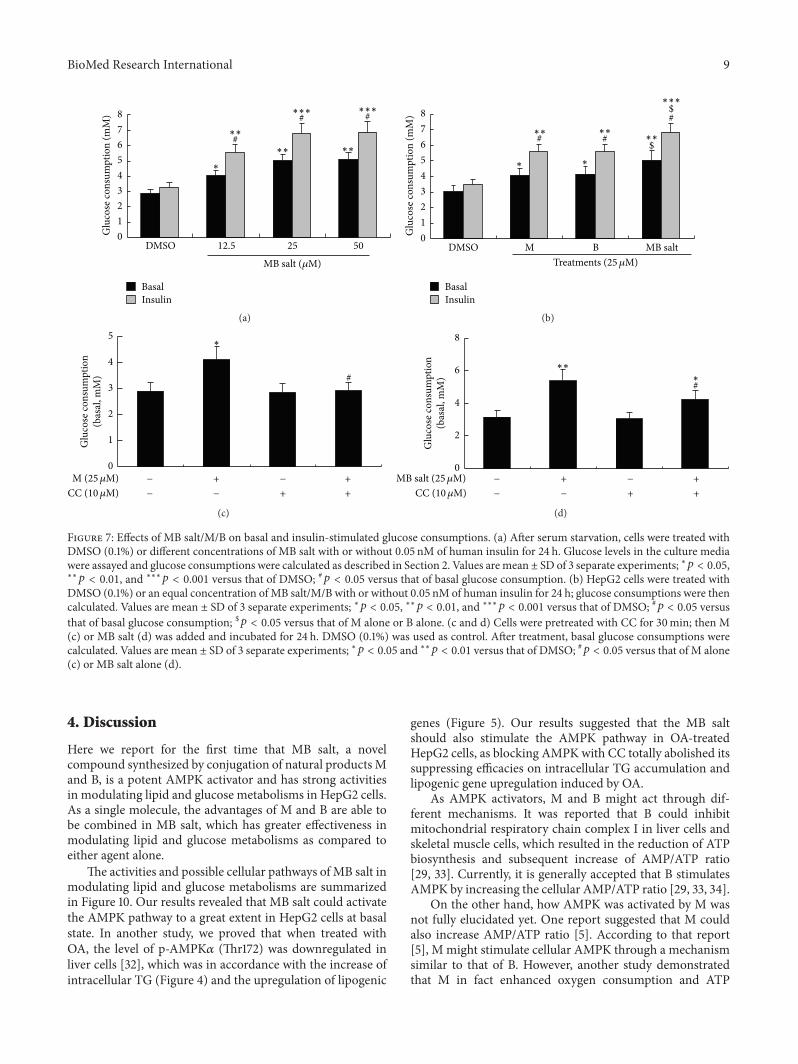
BioMed Research International 9
DMSO 125 25 50
BasalInsulin
MB salt (120583M)
lowastlowastlowastlowast
lowastlowastlowast
lowastlowast
lowast
lowastlowastlowast
012345678
Glu
cose
cons
umpt
ion
(mM
)
(a)
BasalInsulin
DMSO M B MB saltTreatments (25 120583M)
lowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
$
$
012345678
Glu
cose
cons
umpt
ion
(mM
)
(b)
lowast
+ +minusminus
++ minusminusM (25 120583M)CC (10 120583M)
0
1
2
3
4
5
Glu
cose
cons
umpt
ion
(bas
al m
M)
(c)
lowastlowastlowast
++
+ +
minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
0
2
4
6
8
Glu
cose
cons
umpt
ion
(bas
al m
M)
(d)
Figure 7 Effects of MB saltMB on basal and insulin-stimulated glucose consumptions (a) After serum starvation cells were treated withDMSO (01) or different concentrations of MB salt with or without 005 nM of human insulin for 24 h Glucose levels in the culture mediawere assayed and glucose consumptions were calculated as described in Section 2 Values aremean plusmn SD of 3 separate experiments lowast119901 lt 005lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versus that of basal glucose consumption (b) HepG2 cells were treated with
DMSO (01) or an equal concentration of MB saltMB with or without 005 nM of human insulin for 24 h glucose consumptions were thencalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 lowastlowast119901 lt 001 and lowastlowastlowast119901 lt 0001 versus that of DMSO 119901 lt 005 versusthat of basal glucose consumption $119901 lt 005 versus that of M alone or B alone (c and d) Cells were pretreated with CC for 30min then M(c) or MB salt (d) was added and incubated for 24 h DMSO (01) was used as control After treatment basal glucose consumptions werecalculated Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO 119901 lt 005 versus that of M alone(c) or MB salt alone (d)
4 Discussion
Here we report for the first time that MB salt a novelcompound synthesized by conjugation of natural productsMand B is a potent AMPK activator and has strong activitiesin modulating lipid and glucose metabolisms in HepG2 cellsAs a single molecule the advantages of M and B are able tobe combined in MB salt which has greater effectiveness inmodulating lipid and glucose metabolisms as compared toeither agent alone
The activities and possible cellular pathways of MB salt inmodulating lipid and glucose metabolisms are summarizedin Figure 10 Our results revealed that MB salt could activatethe AMPK pathway to a great extent in HepG2 cells at basalstate In another study we proved that when treated withOA the level of p-AMPK120572 (Thr172) was downregulated inliver cells [32] which was in accordance with the increase ofintracellular TG (Figure 4) and the upregulation of lipogenic
genes (Figure 5) Our results suggested that the MB saltshould also stimulate the AMPK pathway in OA-treatedHepG2 cells as blocking AMPKwith CC totally abolished itssuppressing efficacies on intracellular TG accumulation andlipogenic gene upregulation induced by OA
As AMPK activators M and B might act through dif-ferent mechanisms It was reported that B could inhibitmitochondrial respiratory chain complex I in liver cells andskeletal muscle cells which resulted in the reduction of ATPbiosynthesis and subsequent increase of AMPATP ratio[29 33] Currently it is generally accepted that B stimulatesAMPK by increasing the cellular AMPATP ratio [29 33 34]
On the other hand how AMPK was activated by M wasnot fully elucidated yet One report suggested that M couldalso increase AMPATP ratio [5] According to that report[5] M might stimulate cellular AMPK through a mechanismsimilar to that of B However another study demonstratedthat M in fact enhanced oxygen consumption and ATP
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
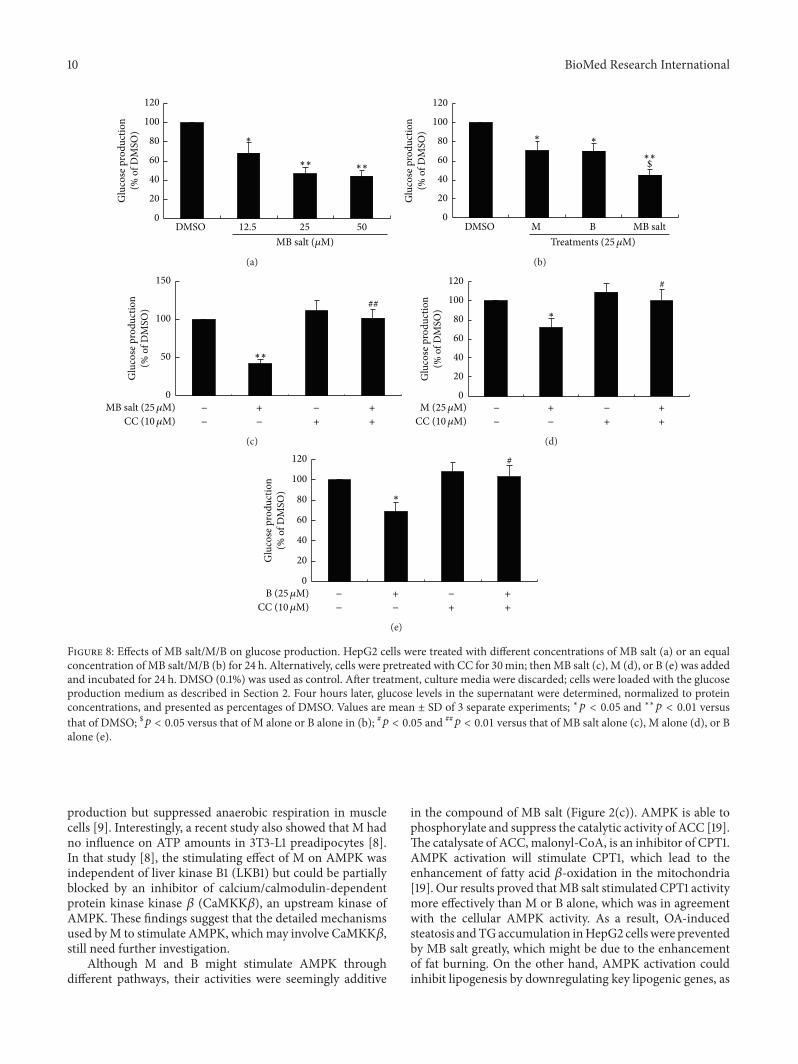
10 BioMed Research International
DMSO 125 25 500
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
MB salt (120583M)
lowast
lowastlowast lowastlowast
(a)
DMSO M B MB saltTreatments (25 120583M)
lowast lowast
lowastlowast$
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
(b)
0
50
100
150
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowastlowast
+
++
+ minus
minusminus
minusMB salt (25 120583M)CC (10 120583M)
(c)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO) lowast
+
++
+ minus
minusminus
minusM (25 120583M)CC (10 120583M)
(d)
0
20
40
60
80
100
120
Glu
cose
pro
duct
ion
( o
f DM
SO)
lowast
+
++
+ minus
minusminus
minusB (25 120583M)CC (10 120583M)
(e)
Figure 8 Effects of MB saltMB on glucose production HepG2 cells were treated with different concentrations of MB salt (a) or an equalconcentration of MB saltMB (b) for 24 h Alternatively cells were pretreated with CC for 30min thenMB salt (c) M (d) or B (e) was addedand incubated for 24 h DMSO (01) was used as control After treatment culture media were discarded cells were loaded with the glucoseproduction medium as described in Section 2 Four hours later glucose levels in the supernatant were determined normalized to proteinconcentrations and presented as percentages of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versusthat of DMSO $119901 lt 005 versus that of M alone or B alone in (b) 119901 lt 005 and
119901 lt 001 versus that of MB salt alone (c) M alone (d) or Balone (e)
production but suppressed anaerobic respiration in musclecells [9] Interestingly a recent study also showed that M hadno influence on ATP amounts in 3T3-L1 preadipocytes [8]In that study [8] the stimulating effect of M on AMPK wasindependent of liver kinase B1 (LKB1) but could be partiallyblocked by an inhibitor of calciumcalmodulin-dependentprotein kinase kinase 120573 (CaMKK120573) an upstream kinase ofAMPK These findings suggest that the detailed mechanismsused byM to stimulate AMPK whichmay involve CaMKK120573still need further investigation
Although M and B might stimulate AMPK throughdifferent pathways their activities were seemingly additive
in the compound of MB salt (Figure 2(c)) AMPK is able tophosphorylate and suppress the catalytic activity of ACC [19]The catalysate of ACC malonyl-CoA is an inhibitor of CPT1AMPK activation will stimulate CPT1 which lead to theenhancement of fatty acid 120573-oxidation in the mitochondria[19] Our results proved thatMB salt stimulated CPT1 activitymore effectively than M or B alone which was in agreementwith the cellular AMPK activity As a result OA-inducedsteatosis andTGaccumulation inHepG2 cells were preventedby MB salt greatly which might be due to the enhancementof fat burning On the other hand AMPK activation couldinhibit lipogenesis by downregulating key lipogenic genes as
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
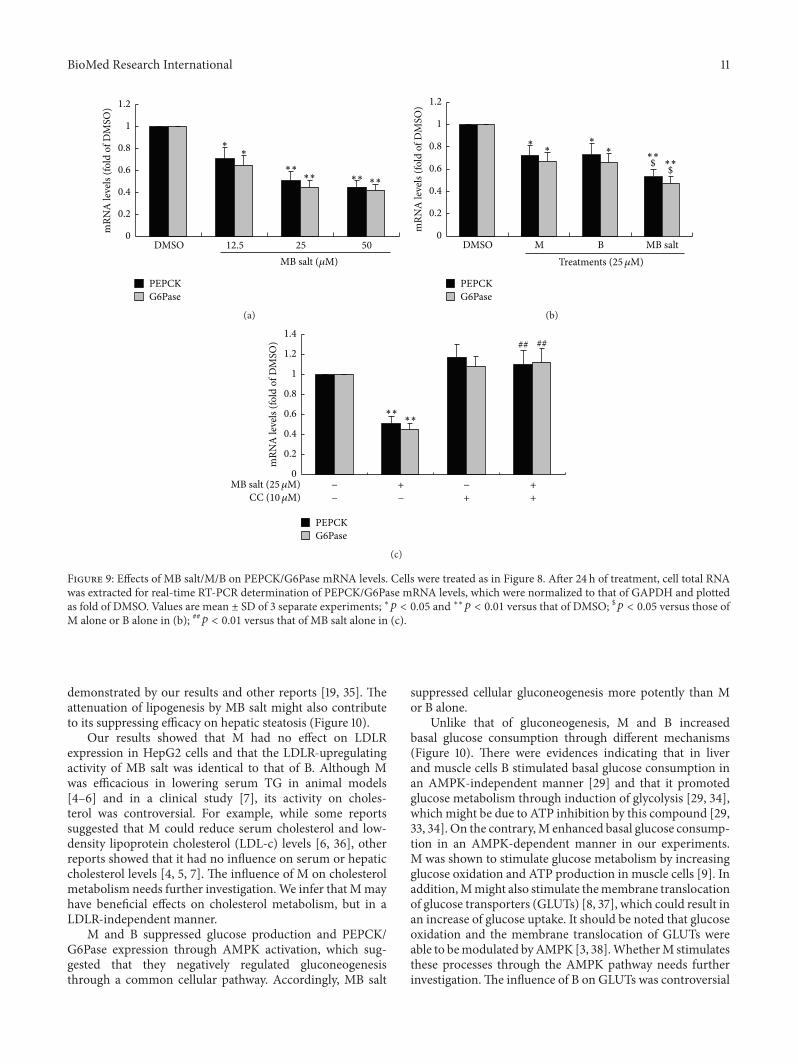
BioMed Research International 11
DMSO 125 25 50
PEPCKG6Pase
0
02
04
06
08
1
12m
RNA
leve
ls (fo
ld o
f DM
SO)
MB salt (120583M)
lowastlowast
lowastlowastlowastlowastlowastlowastlowastlowast
(a)
DMSO M B MB salt
PEPCKG6Pase
0
02
04
06
08
1
12
mRN
A le
vels
(fold
of D
MSO
)
Treatments (25 120583M)
lowastlowast
lowastlowast
$lowastlowast$
lowastlowast
(b)
PEPCKG6Pase
0
02
04
06
08
1
12
14
mRN
A le
vels
(fold
of D
MSO
)
lowastlowastlowastlowast
+
++
+
minusminus
minusminusMB salt (25 120583M)CC (10 120583M)
(c)
Figure 9 Effects of MB saltMB on PEPCKG6Pase mRNA levels Cells were treated as in Figure 8 After 24 h of treatment cell total RNAwas extracted for real-time RT-PCR determination of PEPCKG6Pase mRNA levels which were normalized to that of GAPDH and plottedas fold of DMSO Values are mean plusmn SD of 3 separate experiments lowast119901 lt 005 and lowastlowast119901 lt 001 versus that of DMSO $119901 lt 005 versus those ofM alone or B alone in (b) 119901 lt 001 versus that of MB salt alone in (c)
demonstrated by our results and other reports [19 35] Theattenuation of lipogenesis by MB salt might also contributeto its suppressing efficacy on hepatic steatosis (Figure 10)
Our results showed that M had no effect on LDLRexpression in HepG2 cells and that the LDLR-upregulatingactivity of MB salt was identical to that of B Although Mwas efficacious in lowering serum TG in animal models[4ndash6] and in a clinical study [7] its activity on choles-terol was controversial For example while some reportssuggested that M could reduce serum cholesterol and low-density lipoprotein cholesterol (LDL-c) levels [6 36] otherreports showed that it had no influence on serum or hepaticcholesterol levels [4 5 7] The influence of M on cholesterolmetabolism needs further investigationWe infer that Mmayhave beneficial effects on cholesterol metabolism but in aLDLR-independent manner
M and B suppressed glucose production and PEPCKG6Pase expression through AMPK activation which sug-gested that they negatively regulated gluconeogenesisthrough a common cellular pathway Accordingly MB salt
suppressed cellular gluconeogenesis more potently than Mor B alone
Unlike that of gluconeogenesis M and B increasedbasal glucose consumption through different mechanisms(Figure 10) There were evidences indicating that in liverand muscle cells B stimulated basal glucose consumption inan AMPK-independent manner [29] and that it promotedglucose metabolism through induction of glycolysis [29 34]which might be due to ATP inhibition by this compound [2933 34] On the contraryM enhanced basal glucose consump-tion in an AMPK-dependent manner in our experimentsM was shown to stimulate glucose metabolism by increasingglucose oxidation and ATP production in muscle cells [9] InadditionMmight also stimulate themembrane translocationof glucose transporters (GLUTs) [8 37] which could result inan increase of glucose uptake It should be noted that glucoseoxidation and the membrane translocation of GLUTs wereable to bemodulated byAMPK [3 38]WhetherM stimulatesthese processes through the AMPK pathway needs furtherinvestigationThe influence of B on GLUTs was controversial
12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

12 BioMed Research International
MBsalt
AMPK
Activation
p-ACC uarr
CPT1 activity uarr
TG accumulation darr
PEPCK mRNA darrG6Pase mRNA darr
Gluconeogenesis darr
Glycolysis uarr
Basal glucoseconsumption uarr
Glucoseoxidation uarr
Membranetranslocationof GLUTs uarr
Mitochondriainhibition
Insulin sensitivity uarr
InsR uarr
Lipogenic genes darr
Lipogenesis darr
CaMKK120573
Figure 10 A schematic figure summarizes the activities and possible pathways of MB salt in modulating lipid and glucose metabolismsIn HepG2 cells MB salt could activate AMPK potently during which M (in red lines) and B (in green lines) might have different modesof actions AMPK activation could cause the phosphorylation and inhibition of ACC which in turn stimulated cellular CPT1 activity andsuppressed TG accumulation On the other hand AMPK activation could result in the downregulation of lipogenic genes which could reduceintracellular TG as well PEPCK and G6Pase mRNA expression levels cloud also be downregulated by AMPK activation which could causeinhibition of gluconeogenesis M and B might enhance basal glucose consumption through different mechanisms While B was proved toincrease glycolysis and basal glucose consumption in an AMPK-independent manner the activity of M on basal glucose consumption wasAMPK dependent M was shown to stimulate glucose metabolism by increasing membrane translocation of GLUTs and enhancing glucoseoxidation two processes that were able to be modulated by AMPK In addition MB salt could enhance insulin sensitivity potently althoughM and B might act through different mechanisms Dashed lines and question marks represent pathways and mechanisms that need furtherinvestigation and validation
and some researchers considered that modulation of GLUTsmight not be the major mechanism of B to stimulate glucoseutilization [21]
Our results proved that all three studying compoundscould enhance the insulin-stimulated glucose consumptionin HpeG2 cells When coadministered with insulin the glu-cose consumption-stimulating efficacies ofMB saltMBweresignificantly increased Considering the low concentrationof insulin used in our experiment it could be inferred thatMB saltMB revealed insulin-sensitizing effectsThe detailedmechanisms ofM and B to increase insulin sensitivity are notfully elucidated Our previous results showed that B couldenhance insulin signaling through increasing the expressionof insulin receptor (InsR) in liver cells [39] M had noinfluence on the Akt pathway [18 37] but was shown toinduce CD36 redistribution [13] and activate peroxisomeproliferator-activated receptor-120574 (PPAR-120574) [37] in muscleswhich might have beneficial effects on insulin sensitivityHowever the influences ofMon insulin signaling in liver cellsneed further investigation
Although M and B might promote glucose consumptionand stimulate insulin sensitivity through different mecha-nisms when they were conjugated in the compound of MBsalt significantly improved activities were obtained Together
with the results of gluconeogenesis our findings demon-strated that MB salt had potent activities in modulatingglucose metabolism in HepG2 cells which were superior tothat of M or B alone
When M and B were administered at the same time totreat HepG2 cells they stimulated AMPK reduced intracel-lular TG promoted glucose consumption and suppressedglucose production similar to the MB salt at an equal molarconcentration (data not shown in this study) However the invivo situation may be different When administered alone Mand B are poorly absorbed as single molecules [40 41] whichmay have unfavourable influences on their pharmacologicalactivities in vivo However it was found that MB salt couldbe easily dissolved in strong acids [24] So in vivo in thestomach it is possible that MB salt can be fully dissolvedin gastric acid As a single molecule it is possible thatMB salt may have improved pharmacokinetic parameters ascompared to M and B when administered together whichdeserves further study
In a previous report [42] M and B were simply mixedtogether at a molecular ratio of 1 1 and were used to treatdiabeticmice Although themixture ofM andBwas shown toreduce blood glucose there was no comparison between themixture and M or B alone in that report [42] Furthermore
BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

BioMed Research International 13
that composition was proved to be unstable [24 42] whichmight limit its potential clinical application The MB saltis a single molecule it is stable and can be easily stored[24] which may provide convenience for future clinicalapplication
M andB are natural compoundswhich belong to differentclasses and their chemical skeletons are distinct As a resultthey exhibit different toxicities in HepG2 cells and have dif-ferent mechanisms in modulating metabolisms as discussedabove As a single molecule theMB salt contains anM groupand a B group It can be speculated that the mechanismsand pathways of MB salt in stimulating lipid and glucosemetabolisms may have combined features as compared tothose of M or B alone as illustrated in Figure 10 Indeedour results in the present study prove that MB salt stimulatesTG metabolism and inhibits glucose production through theAMPK pathway However the upstream signals recruited byMB salt to activate AMPK still need further investigationIn addition the influences of MB salt on glycolysis glucoseoxidationmembrane translocation ofGLUTs and the insulinsignaling pathway are not clear they need detailed investiga-tion and comparison with M or B when administered alone
In conclusion the new compound of MB salt has potentactivities in stimulating AMPK reducing intracellular lipidaccumulation enhancing glucose consumption and sup-pressing gluconeogenesis in HepG2 cells Our results maysupportMB salt as a new kind of agent for the development ofnovel lipid or glucose-lowering drugs Future studies shouldfocus on animal experiments to validate the metabolism-modulating activities of MB salt in vivo
Competing Interests
The authors declare no competing interests in this paper
Acknowledgments
This work was supported by the National Natural ScienceFoundation of China (81321004) and National Mega-Projectfor Innovation Drugs (2014ZX09101005-008)
References
[1] S OrsquoNeill and L OrsquoDriscoll ldquoMetabolic syndrome a closer lookat the growing epidemic and its associated pathologiesrdquoObesityReviews vol 16 no 1 pp 1ndash12 2015
[2] Y Gao Y Zhang J Zhu et al ldquoRecent progress in naturalproducts as DPP-4 inhibitorsrdquo Future Medicinal Chemistry vol7 no 8 pp 1079ndash1089 2015
[3] W Chang L Chen and M Hatch ldquoBerberine as a therapy fortype 2 diabetes and its complications frommechanismof actionto clinical studiesrdquo Biochemistry and Cell Biology vol 93 no 5pp 479ndash486 2015
[4] F Guo C Huang X Liao et al ldquoBeneficial effects of mangiferinon hyperlipidemia in high-fat-fed hamstersrdquo Molecular Nutri-tion and Food Research vol 55 no 12 pp 1809ndash1818 2011
[5] Y Niu S Li L Na et al ldquoMangiferin decreases plasma free fattyacids through promoting its catabolism in liver by activation ofAMPKrdquo PLoS ONE vol 7 no 1 Article ID e30782 2012
[6] J Lim Z Liu P Apontes et al ldquoDualmode action ofmangiferinin mouse liver under high fat dietrdquo PLoS ONE vol 9 no 3Article ID e90137 2014
[7] L Na Q Zhang S Jiang et al ldquoMangiferin supplementationimproves serum lipid profiles in overweight patients withhyperlipidemia a double-blind randomized controlled trialrdquoScientific Reports vol 5 article 10344 2015
[8] J Han J Yi F Liang et al ldquoX-3 a mangiferin derivativestimulates AMP-activated protein kinase and reduces hyper-glycemia and obesity in dbdb micerdquo Molecular and CellularEndocrinology vol 405 pp 63ndash73 2015
[9] P Apontes Z Liu K Su et al ldquoMangiferin stimulates car-bohydrate oxidation and protects against metabolic disordersinduced by high-fat dietsrdquo Diabetes vol 63 no 11 pp 3626ndash3636 2014
[10] P S Sellamuthu P Arulselvan S Fakurazi andM KandasamyldquoBeneficial effects of mangiferin isolated from Salacia chinensison biochemical and hematological parameters in rats withstreptozotocininduced diabetesrdquo Pakistan Journal of Pharma-ceutical Sciences vol 27 no 1 pp 161ndash167 2014
[11] X Li X Cui X Sun X Li Q Zhu and W Li ldquoMangiferinprevents diabetic nephropathy progression in streptozotocin-induced diabetic ratsrdquo Phytotherapy Research vol 24 no 6 pp893ndash899 2010
[12] T Miura H Ichiki I Hashimoto et al ldquoAntidiabetic activity ofa xanthone compound mangiferinrdquo Phytomedicine vol 8 no2 pp 85ndash87 2001
[13] L Zhou Y Pan R Chonan et al ldquoMitigation of insulinresistance by mangiferin in a rat model of fructose-inducedmetabolic syndrome is associated with modulation of CD36redistribution in the skeletal musclerdquo The Journal of Pharma-cology and ExperimentalTherapeutics vol 356 no 1 pp 74ndash842016
[14] P B Pal K Sinha and P C Sil ldquoMangiferin attenuates diabeticnephropathy by inhibiting oxidative stress mediated signalingcascade TNF120572 related and mitochondrial dependent apoptoticpathways in streptozotocin-induced diabetic ratsrdquo PLoS ONEvol 9 no 9 Article ID e107220 2014
[15] P S Sellamuthu P Arulselvan S Kamalraj S Fakurazi and MKandasamy ldquoProtective nature ofmangiferin on oxidative stressand antioxidant status in tissues of streptozotocin-induceddiabetic ratsrdquo ISRN Pharmacology vol 2013 Article ID 75010910 pages 2013
[16] P S Sellamuthu P Arulselvan B P Muniappan S Faku-razi and M Kandasamy ldquoMangiferin from salacia chinen-sis prevents oxidative stress and protects pancreatic 120573-cellsin streptozotocin-induced diabetic ratsrdquo Journal of MedicinalFood vol 16 no 8 pp 719ndash727 2013
[17] J Song J Li F Hou X Wang and B Liu ldquoMangiferininhibits endoplasmic reticulum stress-associated thioredoxin-interacting proteinNLRP3 inflammasome activation with reg-ulation of AMPK in endothelial cellsrdquoMetabolism Clinical andExperimental vol 64 no 3 pp 428ndash437 2015
[18] F Wang J Yan Y Niu et al ldquoMangiferin and its aglyconenorathyriol improve glucosemetabolismby activation ofAMP-activated protein kinaserdquo Pharmaceutical Biology vol 52 no 1pp 68ndash73 2014
[19] G R Steinberg and B E Kemp ldquoAMPK in health and diseaserdquoPhysiological Reviews vol 89 no 3 pp 1025ndash1078 2009
[20] J YaoW Kong and J Jiang ldquoLearning from berberine treatingchronic diseases through multiple targetsrdquo Science China LifeSciences vol 58 no 9 pp 854ndash859 2015
14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of

14 BioMed Research International
[21] J Yin J Ye andW Jia ldquoEffects and mechanisms of berberine indiabetes treatmentrdquo Acta Pharmaceutica Sinica B vol 2 no 4pp 327ndash334 2012
[22] W Kong JWei P Abidi et al ldquoBerberine is a novel cholesterol-lowering drug working through a unique mechanism distinctfrom statinsrdquo Nature Medicine vol 10 no 12 pp 1344ndash13512004
[23] X Zhang Y Zhao M Zhang et al ldquoStructural changes of gutmicrobiota during berberine-mediated prevention of obesityand insulin resistance in high-fat diet-fed ratsrdquo PLoS ONE vol7 no 8 Article ID e42529 2012
[24] H Teng andWWu ldquoMangiferin-berberine salt manufacturingmethod and use thereofrdquo US Patent 8673864 B2 2014
[25] Z Li J-D Jiang and W-J Kong ldquoBerberine up-regulates hep-atic low-density lipoprotein receptor through Ras-independentbut AMP-activated protein kinase-dependent Raf-1 activationrdquoBiological amp Pharmaceutical Bulletin vol 37 no 11 pp 1766ndash1775 2014
[26] W J Wang X N Luo X J Ma X Chao and Q Liu ldquoCytotoxiceffects of eight organic solvents in HepG2 and SMMC7721human hepatoma cellsrdquo Modern Traditional Chinese Medicinevol 33 no 4 pp 85ndash86 2013
[27] J-N Li D-Q Song Y-H Lin et al ldquoInhibition of micro-tubule polymerization by 3-bromopropionylamino benzoylurea(JIMB01) a new cancericidal tubulin ligandrdquo Biochemical Phar-macology vol 65 no 10 pp 1691ndash1699 2003
[28] Y Okamoto S Tanaka and Y Haga ldquoEnhanced GLUT2 geneexpression in an oleic acid-induced in vitro fatty liver modelrdquoHepatology Research vol 23 no 2 pp 138ndash144 2002
[29] M Xu Y Xiao J Yin et al ldquoBerberine promotes glucoseconsumption independently of AMP-activated protein kinaseactivationrdquo PLoS ONE vol 9 no 7 Article ID e103702 2014
[30] L-L Zhang L-N Ma D Yan et al ldquoDynamic monitoring ofthe cytotoxic effects of protoberberine alkaloids from RhizomaCoptidis on HepG2 cells using the xCELLigence systemrdquoChinese Journal of Natural Medicines vol 12 no 6 pp 428ndash4352014
[31] Y-N Geng B Yu andW-J Kong ldquoGastrodin ameliorates oleicacid-induced fat accumulation through activation of AMPKpathway in HL-7702 cellsrdquo Chinese Pharmacological Bulletinvol 31 no 1 pp 39ndash44 2015
[32] L Qu B Yu Z Li W Jiang J Jiang and W Kong ldquoGastrodinameliorates oxidative stress and proinflammatory responsein nonalcoholic fatty liver disease through the AMPKNrf2pathwayrdquo Phytotherapy Research vol 30 no 3 pp 402ndash4112016
[33] N Turner J-Y Li A Gosby et al ldquoBerberine and its more bio-logically available derivative dihydroberberine inhibit mito-chondrial respiratory complex I a mechanism for the actionof berberine to activate AMP-activated protein kinase andimprove insulin actionrdquo Diabetes vol 57 no 5 pp 1414ndash14182008
[34] J Yin Z Gao D Liu Z Liu and J Ye ldquoBerberine improvesglucose metabolism through induction of glycolysisrdquo AmericanJournal of PhysiologymdashEndocrinology and Metabolism vol 294no 1 pp E148ndashE156 2008
[35] C-C Liao T-T Ou H-P Huang and C-J Wang ldquoTheinhibition of oleic acid induced hepatic lipogenesis and thepromotion of lipolysis by caffeic acid via up-regulation of AMP-activated kinaserdquo Journal of the Science of Food and Agriculturevol 94 no 6 pp 1154ndash1162 2014
[36] S Muruganandan K Srinivasan S Gupta P K Gupta and JLal ldquoEffect of mangiferin on hyperglycemia and atherogenicityin streptozotocin diabetic ratsrdquo Journal of Ethnopharmacologyvol 97 no 3 pp 497ndash501 2005
[37] MDGironN Sevillano R Salto et al ldquoSalacia oblonga extractincreases glucose transporter 4-mediated glucose uptake in L6rat myotubes role of mangiferinrdquo Clinical Nutrition vol 28 no5 pp 565ndash574 2009
[38] M R Fernandez M D Henry and R E Lewis ldquoKinasesuppressor of Ras 2 (KSR2) regulates tumor cell transformationvia AMPKrdquo Molecular and Cellular Biology vol 32 no 18 pp3718ndash3731 2012
[39] W-J Kong H Zhang D-Q Song et al ldquoBerberine reducesinsulin resistance through protein kinase C-dependent up-regulation of insulin receptor expressionrdquoMetabolism Clinicaland Experimental vol 58 no 1 pp 109ndash119 2009
[40] H Ma H Chen L Sun L Tong and T Zhang ldquoImprovingpermeability and oral absorption ofmangiferin by phospholipidcomplexationrdquo Fitoterapia vol 93 pp 54ndash61 2014
[41] F Qiu Z Zhu N Kang S Piao G Qin and X Yao ldquoIsolationand identification of urinary metabolites of berberine in ratsand humansrdquo Drug Metabolism and Disposition vol 36 no 11pp 2159ndash2165 2008
[42] X J Li J G Deng Z L Tan N Liang and D Y LiaoldquoExperimental study on hypoglycemic effect of the mixture ofmangiferin and berberinerdquo Modern Chinese Medicine vol 10no 12 pp 26ndash28 2008
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Submit your manuscripts athttpwwwhindawicom
PainResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom
Volume 2014
ToxinsJournal of
VaccinesJournal of
Hindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AntibioticsInternational Journal of
ToxicologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
StrokeResearch and TreatmentHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Drug DeliveryJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in Pharmacological Sciences
Tropical MedicineJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Medicinal ChemistryInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
AddictionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Emergency Medicine InternationalHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Autoimmune Diseases
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anesthesiology Research and Practice
ScientificaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Pharmaceutics
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of





























