Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, JUlY 1989, p. 3040-3050 Vol. 63, No. 70022-538X/89/073040-11$02.00/0Copyright ©) 1989, American Society for Microbiology
Responsiveness of the Epstein-Barr Virus NotI Repeat Promoter tothe Z Transactivator Is Mediated in a Cell-Type-Specific Manner by
Two Independent Signal RegionsPAUL M. LIEBERMAN,' J. MARIE HARDWICK,12 AND S. DIANE HAYWARD' 2*
The Virology Laboratories, Department of Pharmacology and Molecular Sciences,' and Department of Neurology,2 TheJohns Hopkins University School of Medicine, Baltimore, Maryland 21205
Received 3 January 1989/Accepted 10 March 1989
Cells latently infected with Epstein-Barr virus (EBV) can be activated to express lytic-cycle polypeptides bythe introduction of the EBV-encoded Z transactivator, indicating that this protein has a pivotal role in virusreactivation. We examined the target specificity of the Z transactivator in short-term cotransfection assays andfound that the most responsive target to Z transactivation was the divergent NotI repeat promoter, locatedwithin the EBV BamHI H fragment. In contrast, target plasmids containing the cat gene linked to heterologousviral promoters were not activated by cotransfection with the Z gene. SI nuclease analysis of RNA fromchemically induced B95-8 cells and from Vero cells cotransfected with NotI repeat promoter-CAT and Zshowed that Z transactivation increased the level of correctly initiated, stable RNA transcripts. The NotI repeatgene (ntr) gives rise to a highly abundant mRNA species after chemical induction of lytic virus replication, butno protein product had been previously identified. Using monospecific antiserum raised against a syntheticpeptide from the BHLF1 open reading frame, we demonstrated that the ntr gene encodes a protein product thatis found in nuclear patches colocalizing with nucleoli. A series of deletions introduced into the upstreamsequences of the NotI-repeat-promoter revealed two separate Z-response regions. The minimal promoter regionbetween -7 and -155 of the leftward RNA cap site and an upstream region between -644 and -902 were bothindependently capable of conferring Z responsiveness. However, the minimal region, which was activated byZ cotransfection in Vero cells, was poorly responsive in lymphocytes, whereas the response of the far-upstreamregion to Z cotransfection was lymphocyte specific. In its human host, EBV infects both epithelial andlymphocyte populations. This dual lifestyle may have led to the evolution of multiple Z-response signals thatenable the Z transactivator to interact with both cell-specific promoter and enhancer factors.
Epstein-Barr virus (EBV) is a human herpesvirus whosegenome exists in B lymphocytes predominantly as a latent,multicopy episome. The latent state is characterized by apattern of gene expression limited to six nuclear antigens(14, 24-27, 31, 32, 41, 46, 47), two latency membraneantigens (16, 23, 28, 37), and two small polymerase IIItranscripts (1, 38). These genes encode functions that main-tain the latent state and provide a growth advantage for theinfected cell. Latency gene products presumably participatein the etiology of EBV-associated neoplasias, includingnasopharyngeal carcinoma and Burkitt's lymphoma, andboth primary and reactivated EBV infections complicate themanagement of immunosuppressed individuals (49).The lytic cycle of EBV is required for horizontal trans-
mission of the virus. The shift from latency to the lytic cycleappears to be governed by a complex interaction betweencellular and viral regulatory pathways. The EBV lytic cyclecan be induced through a variety of chemical manipulations,such as treatment with sodium butyrate (40), halogenatedpyrimidines (18, 21), 12-O-tetradecanoylphorbol-13-acetate(TPA) (60), calcium ionophores (15), or antiserum to humanimmunoglobulin M (57). Some of these agents are thought toact by stimulation of protein kinase C (4). In addition tochemical manipulations, superinfection with the EBVP3HR-1 isolate, which carries defective heterogenous ge-nomes, efficiently disrupts latency (42, 50).The coordinate regulation required for the sequential
synthesis of viral proteins during the productive cycle has
* Corresponding author.
led investigators to search for virus-encoded trans-actingregulatory factors. Classical genetics showed that T antigenof simian virus 40 (SV40), adenovirus ElA proteins, and theherpes simplex virus IE175 protein were essential for pro-ductive infection, and these gene products proved to betranscriptional activators. No genetic mutational informa-tion for EBV exists to implicate essential trans-acting genes.Consequently, we and others have used short-term-expressionassays with recombinant genomic fragments to identify andcharacterize potential EBV regulatory factors. The observa-tion by Countryman and Miller (13) that BamHI-WZ-de-fective DNA induced lytic-cycle polypeptides on transfec-tion into latently EBV-infected D98/HR-1 cells was a keyfirst step in the elucidation of the molecular events leading toEBV reactivation. Subsequently, Takada et al. (56) demon-strated that a long terminal repeat-directed BamHI-Z genefrom the standard virus genome induced expression from acotransfected EBV BamHI YHF DNA fragment. TheBamHI-Z-encoded transactivator was mapped to the BZLF1open reading frame by Chevallier-Greco et al. (7).
Previously we published evidence that the BSLF2-BMLF1 and BRLF1 open reading frames, each of which isretained in defective DNA genomes, also encode transacti-vators (9, 10, 22, 39, 59). In transient-expression assays theproduct of BSLF2-BMLF1, the MS protein, activatesexpression from a variety of heterologous target promotersincluding those derived from SV40, adenovirus, and herpessimplex virus (39). Kenney et al. (33) have suggested thatMS may function at the posttranscriptional level. The Rtransactivator has a more limited and specific target range,
3040
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

CELL-SPECIFIC Z-RESPONSE SIGNAL REGIONS 3041
with the rightward direction of the divergent promoter inBamHI-H being particularly responsive (22). The Z, MS,and R transactivators all behave as immediate-early genes insuperinfection experiments and are all transcribed as splicedmRNAs (6, 51-53).The BamHI H fragment contains a complex set of se-
quences (DSL) which are duplicated in a separate region ofthe genome and which contain the recently described originof lytic replication (20, 36). In addition, these sequencespartially overlap with a divergent promoter region control-ling the NotI repeat (ntr) transcript in the leftward directionand a transcript for a cytoplasmic early antigen in therightward direction. In this work, we present evidence thatthe Z transactivator can independently and specifically acti-vate the levels of correctly initiated stable RNA from theNotI repeat promoter (NRP) and that this repetitive ntrmRNA transcript encodes a detectable protein product.Furthermore, dissection of the NRP revealed two indepen-dent Z-response regions with differing properties. The pro-moter-proximal signal region was responsive to Z cotrans-fection in Vero cells, whereas the upstream signals werespecific for 1B4 cells.
MATERIALS AND METHODS
Cells and transfections. Vero and NIH 3T3 cells weregrown in Dulbecco modified Eagle medium containing 10%fetal calf serum and maintained at 37°C in a 5% CO2incubator. Raji, 1B4, and B95-8 lymphoblastoid cells weremaintained at 5 x 105 cells per ml in RPMI 1640 mediumsupplemented with 10% fetal calf serum. Vero and NIH 3T3cells were transfected by calcium phosphate precipitation, aspreviously reported (43). Cells (2.5 x 105) were plated insix-well cluster dishes the day before transfection. Themedium was changed 4 to 5 h prior to transfection. CaCl, (3pmol) was added dropwise to a DNA cocktail containing 42mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid; pH 7.05), 27 mM NaCl, 10 mM KCl, 1.4 mMNa2HPO4, and 12 mM glucose. After 25 min at roomtemperature, the DNA-calcium phosphate precipitate wasadded dropwise to cells. Four hours after addition of thetransfection mix, the cells were treated with 15% glycerol inDulbecco modified Eagle medium for 1 min, washed oncewith phosphate-buffered saline, and recultured in Dulbeccomodified Eagle medium-10% fetal calf serum for an addi-tional 48 h before harvesting. Lymphoblastoid cells weretransfected by the DEAE-dextran method essentially asdescribed by Takada et al. (56). Cells were suspended at107/ml in Tris-buffered saline (TBS; 0.1 g of MgCl2 6H,O,0.1 g of CaCl2, 8.0 g of NaCl, 0.38 g of KCl, 0.1 g ofNa2HPO4 12H20, and 3.0 g of Tris hydrochloride per liter[pH 7.5]). DNA was mixed with 300 Rl of TBS and 300 pIl ofTBS-DEAE-dextran (1 mg/ml; 500,000 g/mol; Pharmacia,Inc.). Approximately 106 cells in 100 Rl of TBS were layeredonto the DNA-DEAE-dextran solution and mixed at roomtemperature for 30 min. Cells were pelleted in a variable-speed Eppendorf microcentrifuge at 3,000 rpm for 3 min,washed twice with TBS, and then suspended in 1.5 ml ofRPMI 1640 medium containing 10% fetal calf serum. Cellswere harvested for CAT assays or fixed for immunofluores-cence at 48 h after transfection.
Plasmids. The following heterologous viral promoters con-trolling cat gene expression were used. pSV2CAT andpAlOCAT contain the SV40 early regulatory region with the72-base-pair (bp) repeat enhancer sequences (pSV2CAT) orwithout the enhancer region (pAlOCAT) (obtained from G.
Khoury, National Institutes of Health, Bethesda, Md. [35]).pDG3 (IE94-CAT) is a BAL 31 5' deletion mutant of thepTJ278 plasmid containing sequences to -130 bp from thecap site of simian cytomegalovirus (CMV) immediate-earlypromoter (30). E3-CAT (obtained from 0. Andrisani and N.Jones, Purdue University, West Lafayette, Ind.) containsthe adenovirus type 5 E3 control region linked to the CATgene (58). p175K-CAT (pPOH2) contains the herpes simplexvirus immediate-early gene IE175 control regions regulatingCAT expression (43).The EBV-derived targets included MS-CAT (pDH136), in
which the upstream sequences controlling the expression ofthe MS gene (51) were fused to CAT. Sequences from B95-8BamHI-S (pSL95) were subcloned as a 730-bp SphI-NheIfragment into a pBR322 derivative (pKP54) polylinker. Thesequences were then moved as a BamHI-HindIII fragmentinto BamHI-HindIII-cleaved pCAT-B'. The ribonucleotidereductase promoter CAT construction (RR-CAT) was de-rived from EBV sequences regulating the 38-kilodalton smallsubunit of ribonucleotide reductase in BamHI-a (3, 51). A750-bp HindIll fragment was isolated from BamHI-a andthen cleaved by Sau3AI. A 230-bp SauAI fragment wascloned in the correct orientation into the BamHI site ofpCAT-B' to form plasmid pPL23A. NRP-CAT has beendescribed previously (39) as pDH123. Briefly, it contains theduplicated region (DSL) of the B95-8 genome, within a1,070-bp NaeI-to-HincII fragment which was cloned intopCAT-B' with BamHI-HindIII linkers. In NRP-CAT, the catgene is regulated by promoter sequences from +40 to -1030of the ntr gene. The NRP-CAT deletion mutants weregenerated from convenient restriction site cleavages (seeFig. 5). Plasmids pPL52, pPL27, pPL28, and pPL29 were allconstructed from XmaI-KpnI partial digestions followed byT4 polymerase blunt-end formation and ligation. PlasmidpDH124 is a complete KpnI deletion, pDH125 is the com-plete XmaI deletion, and pDH140 is the complete deletion ofboth XmaI and KpnI subfragments, retaining 226 bp ofintervening sequence. In plasmid pPL98 (NRP -155 to -7)the EBV leader sequence was removed by BAL 31 deletionfrom the BamHI site of pPL52. This plasmid retains NRPsequences from -7 to -155. Plasmid pMH103 contains the258-bp XmaI fragment mapping between -644 and -902 ofthe NRP inserted at the BglII site of A10-CAT. In anintermediate cloning step, the XmaI fragment was firstintroduced into the XmaI site on the polylinker of pGH56and subsequently isolated as a BglII fragment. The BHLF1expression plasmid pMH39 contains the 2,196-bp SphI-StuIfragment of B95-8 BamHI-H cloned behind the SV40 earlypromoter of plasmid pGH52.
Plasmids used as effector genes included the human CMVmajor IE68 gene complex (IE1 plus IE2) encoded in plasmidpRL103 (48), the EBV MS gene product expressed from thesimian CMV IE94 promoter of pTJ278 in pPL12 (39), and theBZLF1 gene product in pPL17. This plasmid was con-structed by cloning the EBV BamHI Z fragment between theBglII and BamHI sites of pGH52, a pSV2neo derivative withBamHI-BgIII linkers surrounding the neomycin resistancegene cassette. In this construction, the putative BZLF1promoter is retained and is presumably activated by theSV40 enhancer.CAT assays. CAT assays were performed as described
previously (19, 39). Briefly, cells were harvested 48 h aftertransfection, and cell extracts were incubated with 0.2 ,uCi of[14C]chloramphenicol (60 mCi/mmol) for 1 h at 37°C. Theacetylated forms were separated by chromatography inchloroform-methanol (95:5) solvent. CAT activity was quan-
VOL. 63, 1989
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3042 LIEBERMAN ET AL.
titated by isolating the substrate and the mono- and diacety-lated products from the thin-layer chromatography platesand counting in liquid scintillation fluid in an LKB 1216liquid scintillation counter.RNA analysis. Poly(A)-containing RNA was isolated from
TPA-induced B958 cells by the method of Chirgwin et al. (8).The 30-[il hybridization reaction mixture consisted of 0.3 iLgof DNA probe, 5 VLg of mRNA, and 100 pLg of tRNA inhybridization buffer [40 mM piperazine-N,N'-bis(2-ethane-sulfonic acid) (PIPES; pH 6.4), 1 mM EDTA, 0.4 M NaCl,80% formamide]. The probe (12 pmol of the 2,298-bp NcoI-BamHI fragment from BamnHI-H) was treated with calfintestinal phosphatase before end labeling. Samples wereallowed to hybridize for 3 h at 57°C. Single-stranded probewas digested with 33, 100, or 300 U of S1 nuclease (BethesdaResearch Laboratories, Inc.) at 37°C for 45 min (5). Sampleswere ethanol precipitated twice to remove salt and loadedonto an 8% sequencing gel along with probe that wassequenced by chemical degradation. For the measurement ofCAT RNA, either 50 p.g of total cellular RNA or 10 Lg ofpoly(A) selected mRNA (2) was hybridized to a single-stranded, body-labeled DNA probe. Single-stranded probeswere generated by primer extension with 30 ng of [cx-32P]dATP end-labeled CAT gene primer (5'-TTGGGATATATCAACGGTNG-3'), annealed to 2 p.g of denatureddouble-stranded template (pPL52), and extended with Kle-now polymerase in the presence of [U-32P]dATP (800 Ci/mmol) for 30 min at 37°C. The reaction was then stopped byphenol extraction, and the mixture was purified over a G-50spin column and linearized with BglII. Single-stranded probewas isolated from denaturing acrylamide-urea gels, purified,and used for S1 protection assays as described above.
Preparation of monospecific antiserum. A potentially im-munogenic region in the predicted amino acid sequence ofthe BHLF1 open reading frame was identified by using amodified Chou and Fasman program (11). A synthetic pep-tide was made corresponding to amino acids 163-to 177 fromthe first repeat unit of the BHLF1 product (3). Serine-cysteine was added at the carboxy terminus to provide aspacer and a sulfhydryl group for linkage, and D-tyrosinewas added to the amino terminus to help prevent degradationand for quantitation. The peptide was linked to keyhole limpethemocyanin and used as an immunogen (55). Keyhole limpethemocyanin (10 mg) was dissolved in 1.5 ml buffer A (0.2 Msodium phosphate [pH 7.0], 1 mM EDTA, 0.02% sodiumazide). N-ot-Maleimidobutyryloxysuccinimide (GMBS) (1.4mg) in 25 ,ul of dimethylformamide was added dropwise tokeyhold limpet hemocyanin, and the mixture was incubatedfor 30 to 60 min with stirring. The synthetic peptide (1 to 10mg) in 1 ml of buffer B (0.01 M sodium phosphate [pH 6.0],1 mM EDTA, 0.02% sodium azide) was added to the keyholelimpet hemocyanin-GMBS mixture and incubated at 37°C for2 h with frequent agitation. The sample was prepared forinjection by mixing 1 ml of cross-linked peptide with 1 ml ofFreund complete adjuvant and sonicating the mixture togenerate a paste. Rabbits were immunized at 10 intradermalsites. Six weeks later they were boosted with 0.5 ml ofcross-linked peptide (stored at -20°C) plus 0.5 ml of Freundincomplete adjuvant. The mixture was sonicated as aboveand injected subcutaneously.
Immunofluorescence. At 48 h after transfection, cells werefixed with acetone at -20°C for 10 min. The acetone wasremoved by washing with phosphate-buffered saline, andnonspecific sites were blocked with 0.1% bovine serumalbumin. All antibodies were diluted in phosphate-bufferedsaline plus 0.1% bovine serum albumin. Samples were
incubated for 30 min at room temperature with 20 ,ul of rabbitanti-BHLF-peptide antiserum (1:50 dilution), washed, andincubated again with fluorescein isothiocyanate-conjugatedgoat anti-rabbit second antibody. Slides were washed andmounted in 1 mg of p-phenylenediamine per ml to retardfading of fluorescence.
RESULTS
Specificity of the Z transactivator. Investigators in severallaboratories have reported that an enhancer-driven BZLF1gene can reactivate the EBV lytic cycle when transfectedinto latently infected cells (7, 13, 56). As an initial control,we examined the ability of the Z transactivator and of otherimmediate-early genes, also under the control of strong viralpromoters, to reactivate latent EBV genomes from Rajicells. Both the human CMV IE68-2 gene product and theEBV BMLF1 protein have been shown to be powerfulnonspecific activators of gene expression (39, 48). Raji cellswere transfected with these immediate-early genes (pPL12[MS], pPL17 [Z], and pRL103 [human CMV IE68], or withpBR322 control DNA) and assayed for activation of anabortive lytic cycle by indirect immunofluorescence screen-ing with the R3 monoclonal antibody directed against theBMRF1 early-gene product (45). The Raji cells showed<0.1% early-antigen-positive cells when untreated. Whenthe Raji cells were screened 48 h after transfection, only Zincreased early-antigen expression (36 of 1,500 [2.4%] posi-tive). There was no induction (O of 1,500) by the twononspecific transactivators. This indicates that there areZ-specific targets whose expression is essential for reactiva-tion.A variety of heterologous viral promoters were used as
targets to screen the range and specificity of promoterelements susceptible to Z transactivation. Since the back-ground of EBV genomes in EBV-positive cells might com-plicate the analysis of gene expression in short-term assaysin which Z was used, we performed the experiments by usingVero cells. In contrast to the efficient transactivation in-duced by the MS gene product, cotransfection with Z did notincrease expression from the SV40 minimal early promoter(AlOCAT), the herpes simplex virus IE175 promoter (175K-CAT), a deletion variant (pDG3) of the simian CMV imme-diate-early promoter (IE94-CAT), or the adenovirus earlygene 3 promoter (E3-CAT) (Fig. la).The results obtained with a set of EBV targets are shown
in Fig. lb. The NRP-CAT construction responded with a13-fold-higher level of activation when cotransfected with Z.None of the other EBV promoters tested responded with thesame efficiency as NRP-CAT. The MS-CAT construction, inwhich the CAT gene is driven by the promoter for the MStransactivator, gave a twofold induction on cotransfectionwith Z. The ribonucleotide reductase CAT recombinant didnot show a significant response to either the MS or Z geneproducts individually, although a 10-fold response was ob-tained when the MS and Z genes were cotransfected jointly.The RR-CAT construct was induced 80-fold by the strongnonspecific transactivator encoded by the human CMVimmediate-early IE68 gene complex, further emphasizingthat neither the MS nor the Z gene products producedmaximal activation of this promoter.To ensure that the relative specificities observed were not
merely a result of the particular effector-to-target-gene ratiosbeing used, we performed dose-response experiments ontwo EBV promoters that responded with different effi-ciencies to the Z and MS transactivators. The NRP-CAT and
J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
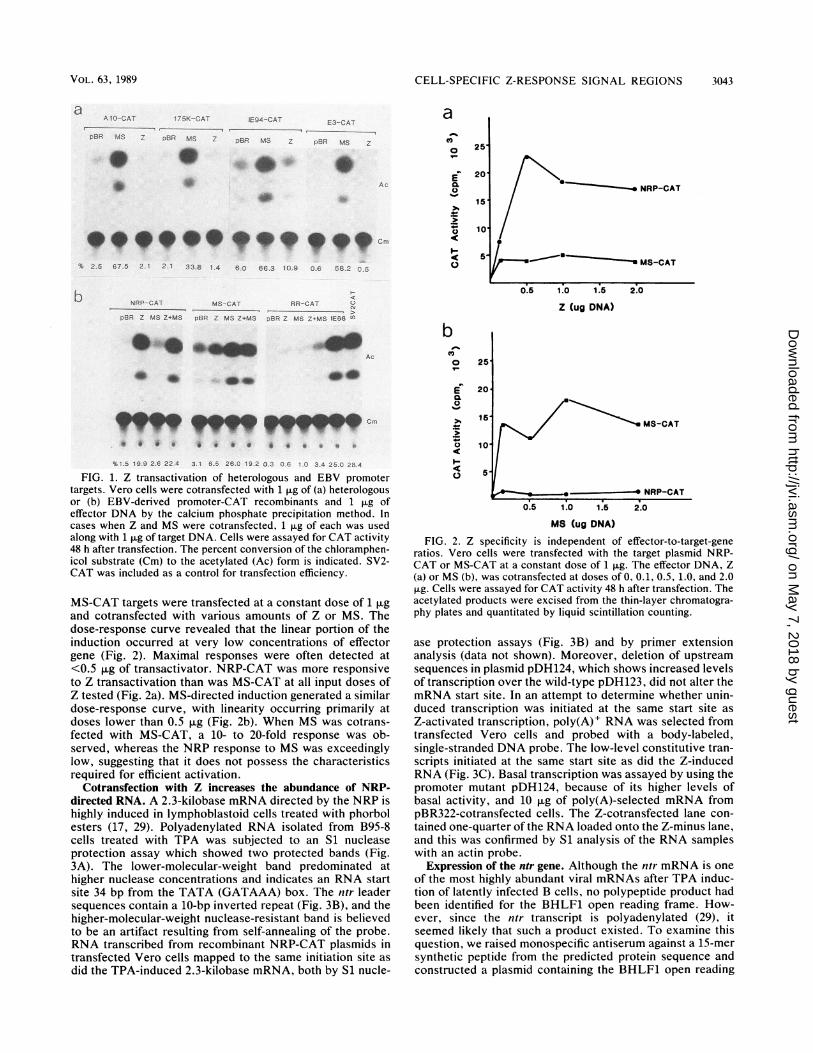
CELL-SPECIFIC Z-RESPONSE SIGNAL REGIONS 3043
aAI -CAT '7-5K-CAT IE94--CAT E3- CA T
pBR MS 7 )B MS 2 PBRS MS BBIR MS a
S a
% 2.5 67.5 2.1 121 33.8 1.4 6.0 66.3 10.9 0.6 5B S
b NRP-CAT MS-- CAT RR-CAT u
pBR Z MS Z.MS pBR Z MS Z4MS --BR Z MS 7MS IE68 r
Ac
4, _r
99y9qy9 Crs0 0 0 * * ea a * e a.
%1 5 19.9 2.6 22 4 3 1 6.5 26-0 19 2 0 3 06 1 0 3.4 25.2 28.4FIG. 1. Z transactivation of heterologous and EBV promoter
targets. Vero cells were cotransfected with 1 ,ug of (a) heterologousor (b) EBV-derived promoter-CAT recombinants and 1 ,ug ofeffector DNA by the calcium phosphate precipitation method. Incases when Z and MS were cotransfected, 1 ,ug of each was usedalong with 1 ,ug of target DNA. Cells were assayed for CAT activity48 h after transfection. The percent conversion of the chloramphen-icol substrate (Cm) to the acetylated (Ac) form is indicated. SV2-CAT was included as a control for transfection efficiency.
MS-CAT targets were transfected at a constant dose of 1 jigand cotransfected with various amounts of Z or MS. Thedose-response curve revealed that the linear portion of theinduction occurred at very low concentrations of effectorgene (Fig. 2). Maximal responses were often detected at<0.5 jig of transactivator. NRP-CAT was more responsiveto Z transactivation than was MS-CAT at all input doses ofZ tested (Fig. 2a). MS-directed induction generated a similardose-response curve, with linearity occurring primarily atdoses lower than 0.5 ,ug (Fig. 2b). When MS was cotrans-fected with MS-CAT, a 10- to 20-fold response was ob-served, whereas the NRP response to MS was exceedinglylow, suggesting that it does not possess the characteristicsrequired for efficient activation.
Cotransfection with Z increases the abundance of NRP-directed RNA. A 2.3-kilobase mRNA directed by the NRP ishighly induced in lymphoblastoid cells treated with phorbolesters (17, 29). Polyadenylated RNA isolated from B95-8cells treated with TPA was subjected to an S1 nucleaseprotection assay which showed two protected bands (Fig.3A). The lower-molecular-weight band predominated athigher nuclease concentrations and indicates an RNA startsite 34 bp from the TATA (GATAAA) box. The ntr leadersequences contain a 10-bp inverted repeat (Fig. 3B), and thehigher-molecular-weight nuclease-resistant band is believedto be an artifact resulting from self-annealing of the probe.RNA transcribed from recombinant NRP-CAT plasmids intransfected Vero cells tnapped to the same initiation site asdid the TPA-induced 2.3-kilobase mRNA, both by S1 nucle-
a0co0
EQ0
0
I-0
b0co't0
EQ0
-
01c:
251
201
15
10
5-1
20'
15'
10
NRP-CAT
I-MS-CAT
0.5 1.0 1.5 2.0
Z (ug DNA)
iMS-CAT
-. - NRP-CAT
1.0 1.5 2.0
MS (ug DNA)FIG. 2. Z specificity is independent of effector-to-target-gene
ratios. Vero cells were transfected with the target plasmid NRP-CAT or MS-CAT at a constant dose of 1 ,ug. The effector DNA, Z(a) or MS (b), was cotransfected at doses of 0, 0.1, 0.5, 1.0, and 2.0jig. Cells were assayed for CAT activity 48 h after transfection. Theacetylated products were excised from the thin-layer chromatogra-phy plates and quantitated by liquid scintillation counting.
ase protection assays (Fig. 3B) and by primer extensionanalysis (data not shown). Moreover, deletion of upstreamsequences in plasmid pDH124, which shows increased levelsof transcription over the wild-type pDH123, did not alter themRNA start site. In an attempt to determine whether unin-duced transcription was initiated at the same start site asZ-activated transcription, poly(A)+ RNA was selected fromtransfected Vero cells and probed with a body-labeled,single-stranded DNA probe. The low-level constitutive tran-scripts initiated at the same start site as did the Z-inducedRNA (Fig. 3C). Basal transcription was assayed by using thepromoter mutant pDH124, because of its higher levels ofbasal activity, and 10 jig of poly(A)-selected mRNA frompBR322-cotransfected cells. The Z-cotransfected lane con-tained one-quarter of the RNA loaded onto the Z-minus lane,and this was confirmed by S1 analysis of the RNA sampleswith an actin probe.
Expression of the ntr gene. Although the ntr mRNA is oneof the most highly abundant viral mRNAs after TPA induc-tion of latently infected B cells, no polypeptide product hadbeen identified for the BHLF1 open reading frame. How-ever, since the ntr transcript is polyadenylated (29), itseemed likely that such a product existed. To examine thisquestion, we raised monospecific antiserum against a 15-mersynthetic peptide from the predicted protein sequence andconstructed a plasmid containing the BHLF1 open reading
VOL. 63, 1989
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3044 LIEBERMAN ET AL.
A B Cp 2J4-i p i:)4i- 23
II1 I' j 7 + "'
k. .1
VW2
;;'-GW
-/. +..
_/._ 4*I ..
_s
p
(-)
01C)
C)
_m 4
CA'T Probe
I
A zlIir bt:it:L
FIG. 3. Z increases the abundance of correctly initiated stable mRNA by Si nuclease protection assays. (A) Identification of the initiationsite for RNA directed by the NRP in TPA-treated B95-8 cells as determined. (B) Induction of NRP-CAT (pDH123 and pDH124) RNA inZ-cotransfected Vero cells and (C) comparison of the start sites for uninduced and Z-induced NRP-CAT mRNAs by using oligo(dT)-selectedRNA. To verify the RNA quantitation, we subjected a duplicate sample to S1 analysis with an actin probe.
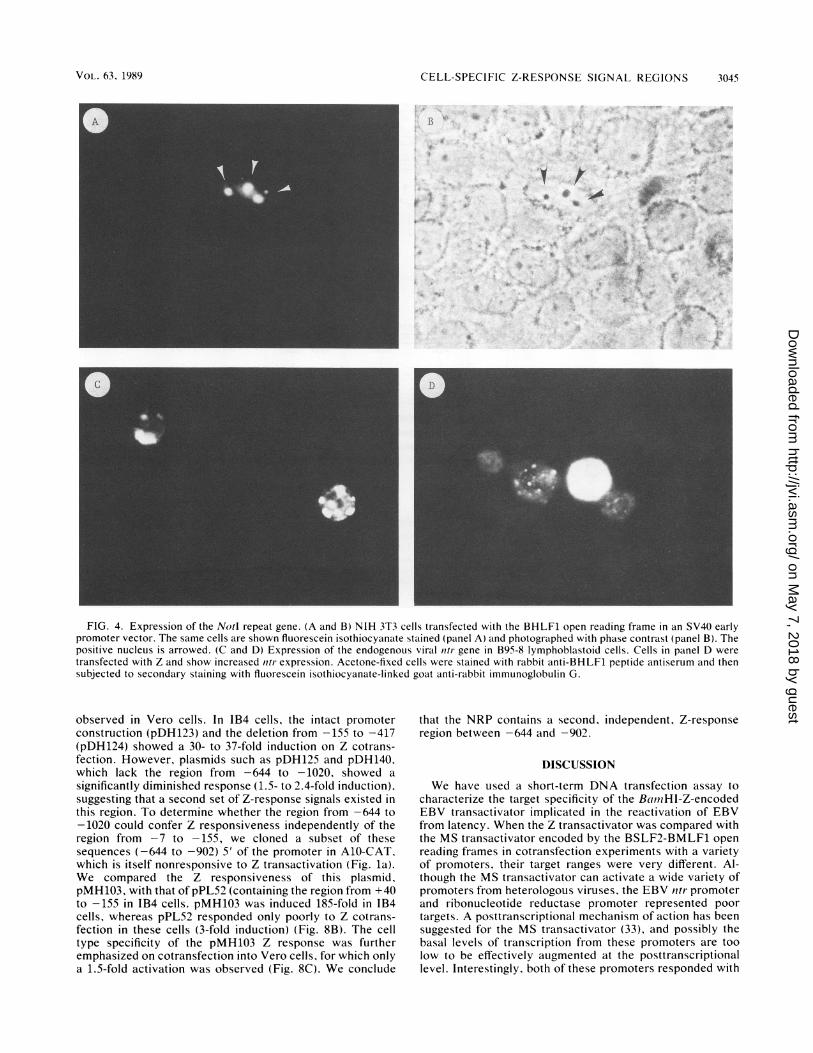
frame under the control of the SV40 early promoter. NIH3T3 cells were transfected with this BHLF1 plasmid, and 48h after transfection, the cells were stained with the anti-BHLF1 antiserum in an indirect immunofluorescence assay.The transfected cells showed discrete foci of staining withinthe nucleus, and these fluorescent foci colocalized with thecell nucleoli, as shown by phase microscopy (Fig. 4A and B).No staining was observed on control (pBR322) transfectedcells (data not shown). To determine whether the Ntr proteinwas synthesized in EBV-infected cells, we also used the Ntrantibody to stain B95-8 cells (Fig. 4C). Approximately 5% ofthe B95-8 cells exhibited a staining pattern of discretenuclear foci similar to that observed in the transfected NIH3T3 cells. The proportion of cells staining with the Ntrantibody was the same as that staining with the R3 anti-earlyantigen antibody and is consistent with expression of a
lytic-cycle gene product. When B95-8 cells were transfectedwith Z, a two- to threefold increase in the number of cellsstaining with the anti-Ntr antibody was observed (Fig. 4D).This indicates that the endogenous ntr gene can also respondto Z.
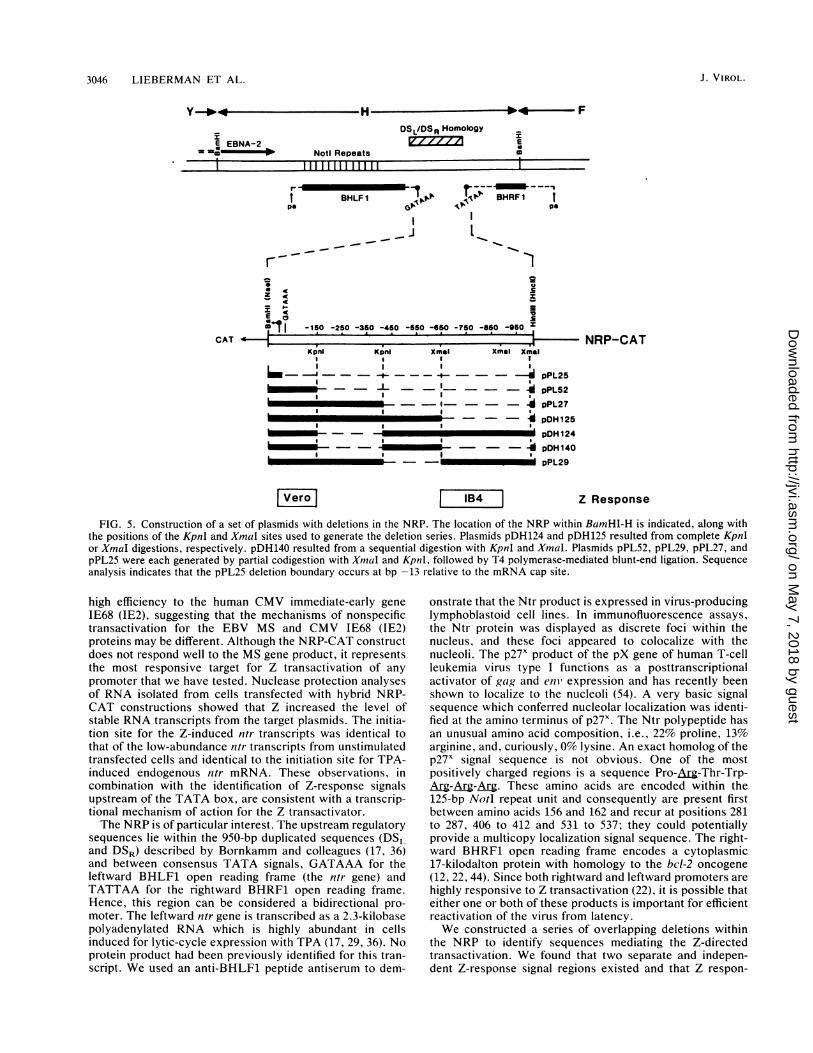
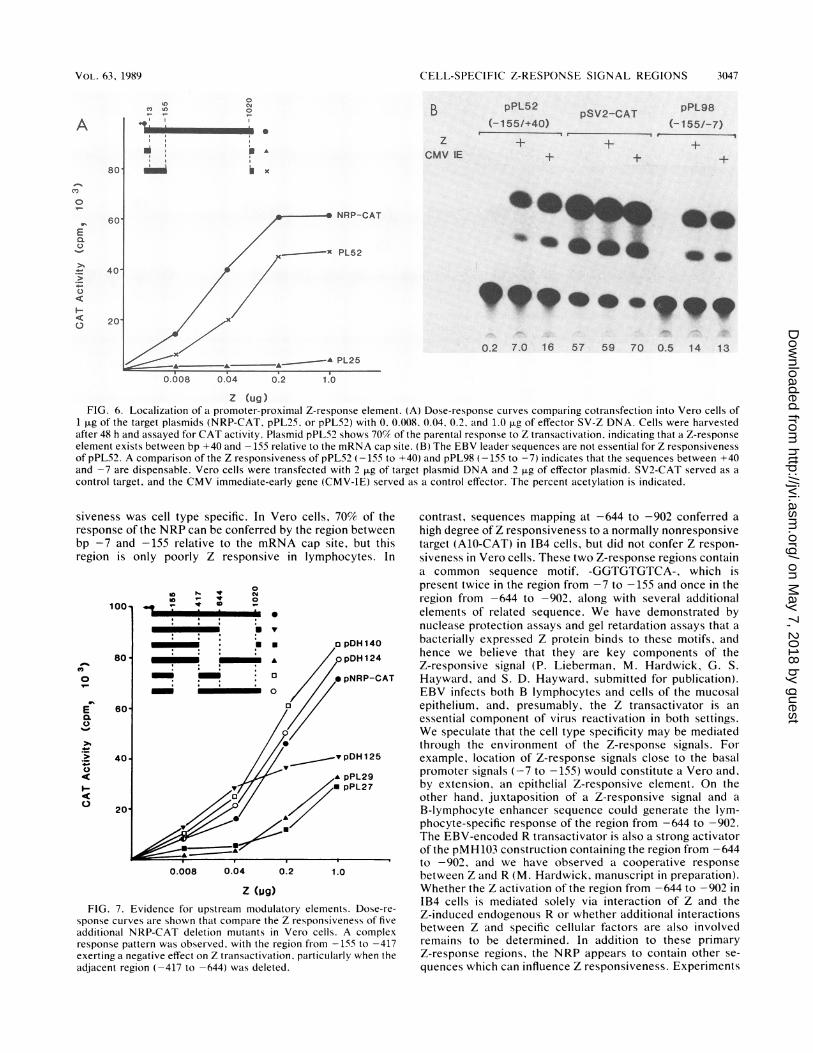
Localization of a Z response element between -7 and -155of the NRP. Since the NRP-CAT construct responded withhigh efficiency to Z-directed transactivation, we constructeda series of overlapping KpnI and XmnaI deletions in the1,030-bp promoter region (Fig. 5) to map the responseelements in NRP. This promoter region is bounded byfunctional leftward- and rightward-directed TATA boxes(22). In these experiments we are examining expression fromthe leftward TATA box. The following experiments with theNRP deletion mutants were performed with Vero cells in a
dose-response protocol with target DNA kept at a constantinput level of 1 pLg while the level of Z-gene activatorincreased from 0.008 to 1.0 ,ug. Figure 6A shows evidencefor a strong response element that yields 70% of the activityof the complete promoter in Vero cells. Plasmid pPL52,which contains sequences from +40 to -155 of the NRP,responded almost as well as did the intact 1,020-bp pro-
moter, indicating that this region contains Z-response ele-ments. Plasmid pPL25, which contains sequences from +40to -13 and lacks a TATA box, was used as a control target.No response was detected. The NRP-CAT constructionscontain a 40-bp EBV leader sequence. To determine whetherthese sequences were required for Z responsiveness, theEBV leader was removed by BAL 31 digestion to create a-155 to -7 NRP-CAT construction (pPL98). This -155 to-7 variant responded to Z transactivation just as well as didpPL52 (-155 to +40) (Fig. 6B).
Evidence for additional upstream modulatory elements.Attempts to evaluate the effects of the presence or absenceof blocks of upstream sequence delineated by the KpnI andXmaI sites revealed a complicated pattern of Z responsive-ness. When the 5' upstream sequences were extended from-155 to the KpnI site at -417 (pPL27), an unusual behaviorpattern was observed. The additional 260 bp partially sup-pressed the induction mediated by the primary Z-responseelement at all doses of transactivator (Fig. 7). Plasmidswhich were deleted in this region (-155 to -417) andretained the adjacent region (-417 to -644), e.g., pDH124and pDH140, were consistently more Z responsive than theparental NRP-CAT, whereas plasmids with the complemen-tary arrangement in which -155 to -417 was present and-417 to -644 was absent, e.g., pPL27 and pPL29, showed adiminished Z responsiveness. These results suggest that a
negative regulatory region may be present within the se-
quences from -155 to -417, with compensatory positiveelements in the adjacent region. These regions may beimportant in determining basal levels of expression from theNRP. Plasmid pDH125, which is deleted in the region -644to -1020, responded to Z cotransfection at 50% of the levelof the complete promoter (Fig. 7).
Location of an IB4 cell-specific Z-responsive region between-644 and -902. To further assess the contribution of theupstream sequences, we also performed cotransfection ex-
periments with latently EBV-infected IB4 lymphocytes (Fig.8A). The results obtained differed dramatically from those
[4t
L.§
J. VIROL.
04.4itt-A
k
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
CELL-SPECIFIC Z-RESPONSE SIGNAL REGIONS 3045
P;
Ip.
Y l
FIG. 4. Expression of the Notd repeat gene. (A and B) NIH 3T3 cells transfected with the BHLF1 open reading frame in an SV40 earlypromoter vector. The same cells are shown fluorescein isothiocyanate stained (panel A) and photographed with phase contrast (panel B). Thepositive nucleus is arrowed. (C and D) Expression of the endogenous viral Itli- gene in B95-8 lymphoblastoid cells. Cells in panel D weretransfected with Z and show increased titr expression. Acetone-fixed cells were stained with rabbit anti-BHLF1 peptide antiserum and thensubjected to secondary staining with fluorescein isothiocyanate-linked goat anti-rabbit immunoglobulin G.
observed in Vero cells. In 1B4 cells, the intact promoterconstruction (pDH123) and the deletion from -155 to -417(pDH124) showed a 30- to 37-fold induction on Z cotrans-fection. However, plasmids such as pDH125 and pDH140,which lack the region from -644 to -1020, showed asignificantly diminished response (1.5- to 2.4-fold induction),suggesting that a second set of Z-response signals existed inthis region. To determine whether the region from -644 to-1020 could confer Z responsiveness independently of theregion from -7 to -155, we cloned a subset of thesesequences (-644 to -902) 5' of the promoter in A10-CAT,which is itself nonresponsive to Z transactivation (Fig. la).We compared the Z responsiveness of this plasmid,pMH103, with that of pPL52 (containing the region from +40to -155 in 1B4 cells. pMH103 was induced 185-fold in 1B4cells, whereas pPL52 responded only poorly to Z cotrans-fection in these cells (3-fold induction) (Fig. 8B). The celltype specificity of the pMH103 Z response was furtheremphasized on cotransfection into Vero cells, for which onlya 1.5-fold activation was observed (Fig. 8C). We conclude
that the NRP contains a second, independent, Z-responseregion between -644 and -902.
DISCUSSION
We have used a short-term DNA transfection assay tocharacterize the target specificity of the BafmHI-Z-encodedEBV transactivator implicated in the reactivation of EBVfrom latency. When the Z transactivator was compared withthe MS transactivator encoded by the BSLF2-BMLF1 openreading frames in cotransfection experiments with a varietyof promoters, their target ranges were very different. Al-though the MS transactivator can activate a wide variety ofpromoters from heterologous viruses, the EBV ntt promoterand ribonucleotide reductase promoter represented poortargets. A posttranscriptional mechanism of action has beensuggested for the MS transactivator (33), and possibly thebasal levels of transcription from these promoters are toolow to be effectively augmented at the posttranscriptionallevel. Interestingly, both of these promoters responded with
VOL. 63. 1989
8- I' 1.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
3046 LIEBERMAN ET AL.
Y-44 H 4 FDS L/DSR Homology
E EBNA-2 Esw* Notl Repeats X
i II1111II11111111 1r
pa
Ps
CAT 4-
m y ~~~~~~~~~~---BHLF 1 J, .>Q 1BHRF1 t
6E s pa
K_~ ~ ~ ,:'p _v" p
-. a
an | -150 -250 -350 -460 -550 -650 -750 -860 -960I a
-L }--Kpnl Kpni Xmal Xmal Xmal
| I I ~ ~ ~ ~~~~~I I__ _ _ __ e--_ pPL25
- - - 1- - -- id pPL52
- -I - - - Xd pPL27_IS I a pDH125
- - pDH124-m-oos - - - -d pDH 140
I I______________-- pPL29
Ivero I I IB4 ]
NRP-CAT
Z Response
FIG. 5. Construction of a set of plasmids with deletions in the NRP. The location of the NRP within BamHI-H is indicated, along withthe positions of the KpnI and XmaI sites used to generate the deletion series. Plasmids pDH124 and pDH125 resulted from complete Kpnlor XmaI digestions, respectively. pDH140 resulted from a sequential digestion with KpnI and Xmal. Plasmids pPL52, pPL29, pPL27, andpPL25 were each generated by partial codigestion with Xmal and Kpnl, followed by T4 polymerase-mediated blunt-end ligation. Sequenceanalysis indicates that the pPL25 deletion boundary occurs at bp -13 relative to the mRNA cap site.
high efficiency to the human CMV immediate-early geneIE68 (IE2), suggesting that the mechanisms of nonspecifictransactivation for the EBV MS and CMV IE68 (IE2)proteins may be different. Although the NRP-CAT constructdoes not respond well to the MS gene product, it representsthe most responsive target for Z transactivation of anypromoter that we have tested. Nuclease protection analysesof RNA isolated from cells transfected with hybrid NRP-CAT constructions showed that Z increased the level ofstable RNA transcripts from the target plasmids. The initia-tion site for the Z-induced ntr transcripts was identical tothat of the low-abundance ntr transcripts from unstimulatedtransfected cells and identical to the initiation site for TPA-induced endogenous ntr mRNA. These observations, incombination with the identification of Z-response signalsupstream of the TATA box, are consistent with a transcrip-tional mechanism of action for the Z transactivator.The NRP is of particular interest. The upstream regulatory
sequences lie within the 950-bp duplicated sequences (DSLand DSR) described by Bornkamm and colleagues (17, 36)and between consensus TATA signals, GATAAA for theleftward BHLF1 open reading frame (the ntr gene) andTATTAA for the rightward BHRF1 open reading frame.Hence, this region can be considered a bidirectional pro-moter. The leftward ntr gene is transcribed as a 2.3-kilobasepolyadenylated RNA which is highly abundant in cellsinduced for lytic-cycle expression with TPA (17, 29, 36). Noprotein product had been previously identified for this tran-script. We used an anti-BHLF1 peptide antiserum to dem-
onstrate that the Ntr product is expressed in virus-producinglymphoblastoid cell lines. In immunofluorescence assays,the Ntr protein was displayed as discrete foci within thenucleus, and these foci appeared to colocalize with thenucleoli. The p27X product of the pX gene of human T-cellleukemia virus type I functions as a posttranscriptionalactivator of gag and eni' expression and has recently beenshown to localize to the nucleoli (54). A very basic signalsequence which conferred nucleolar localization was identi-fied at the amino terminus of p27x. The Ntr polypeptide hasan unusual amino acid composition, i.e., 22% proline, 13%arginine, and, curiously, 0% lysine. An exact homolog of thep27X signal sequence is not obvious. One of the mostpositively charged regions is a sequence Pro-Arg-Thr-Trp-Arg-Arg-Arg. These amino acids are encoded within the125-bp Notl repeat unit and consequently are present firstbetween amino acids 156 and 162 and recur at positions 281to 287, 406 to 412 and 531 to 537; they could potentiallyprovide a multicopy localization signal sequence. The right-ward BHRF1 open reading frame encodes a cytoplasmic17-kilodalton protein with homology to the bcl-2 oncogene(12, 22, 44). Since both rightward and leftward promoters arehighly responsive to Z transactivation (22), it is possible thateither one or both of these products is important for efficientreactivation of the virus from latency.We constructed a series of overlapping deletions within
the NRP to identify sequences mediating the Z-directedtransactivation. We found that two separate and indepen-dent Z-response signal regions existed and that Z respon-
J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from
CELL-SPECIFIC Z-RESPONSE SIGNAL REGIONS 3047
C 0
Axi
80i E6
CO
E
C.
0._
2C
0
0
.
B
zCMV IE
NRP-CAT
PL52
PL25
pPL52PSV2-CAT PPL98
(-155/+40) (-155/-7).
±+
++ +
*;.
0.2 7.0 16 57 59 70 0.5 14 13
Z (ug)FIG. 6. Localization of a promoter-proximal Z-response element. (A) Dose-response curves comparing cotransfection into Vero cells of
1 ,ug of the target plasmids (NRP-CAT, pPL25. or pPL52) with 0. 0.008, 0.04. 0.2. and 1.0 Fg of effector SV-Z DNA. Cells were harvestedafter 48 h and assayed for CAT activity. Plasmid pPL52 shows 70% of the parental response to Z transactivation, indicating that a Z-responseelement exists between bp +40 and -155 relative to the mRNA cap site. (B) The EBV leader sequences are not essential for Z responsivenessof pPL52. A comparison of the Z responsiveness of pPL52 (-155 to +40) and pPL98 (-155 to -7) indicates that the sequences between +40and -7 are dispensable. Vero cells were transfected with 2 ,ug of target plasmid DNA and 2 p.g of effector plasmid. SV2-CAT served as acontrol target, and the CMV immediate-early gene (CMV-IE) served as a control effector. The percent acetylation is indicated.
siveness was cell type specific. In Vero cells. 70% of theresponse of the NRP can be conferred by the region betweenbp -7 and -155 relative to the mRNA cap site, but thisregion is only poorly Z responsive in lymphocytes. In
0
1001 _.q - qioo
_ * n O pDH14080 pDH 124
e. . . .
o=.0 .
pNRP-CAT0
E 60 o
. 40- /pDH125
A pPL29CpPL27
0.008 0.04 0.2 1.0
Z (9g)
FIG. 7. Evidence for upstream modulatory elements. Dose-re-sponse curves are shown that compare the Z responsiveness of fiveadditional NRP-CAT deletion mutants in Vero cells. A complexresponse pattern was observed, with the region from -155 to -417exerting a negative effect on Z transactivation. particularly when theadjacent region (-417 to -644) was deleted.
contrast, sequences mapping at -644 to -902 conferred ahigh degree of Z responsiveness to a normally nonresponsivetarget (A10-CAT) in IB4 cells, but did not confer Z respon-siveness in Vero cells. These two Z-response regions containa common sequence motif, -GGTGTGTCA-, which ispresent twice in the region from -7 to -155 and once in theregion from -644 to -902, along with several additionalelements of related sequence. We have demonstrated bynuclease protection assays and gel retardation assays that abacterially expressed Z protein binds to these motifs, andhence we believe that they are key components of theZ-responsive signal (P. Lieberman, M. Hardwick, G. S.Hayward, and S. D. Hayward, submitted for publication).EBV infects both B lymphocytes and cells of the mucosalepithelium, and, presumably, the Z transactivator is anessential component of virus reactivation in both settings.We speculate that the cell type specificity may be mediatedthrough the environment of the Z-response signals. Forexample, location of Z-response signals close to the basalpromoter signals (-7 to -155) would constitute a Vero and,by extension, an epithelial Z-responsive element. On theother hand, juxtaposition of a Z-responsive signal and aB-lymphocyte enhancer sequence could generate the lym-phocyte-specific response of the region from -644 to -902.The EBV-encoded R transactivator is also a strong activatorof the pMH103 construction containing the region from -644to -902, and we have observed a cooperative responsebetween Z and R (M. Hardwick, manuscript in preparation).Whether the Z activation of the region from -644 to -902 inIB4 cells is mediated solely via interaction of Z and theZ-induced endogenous R or whether additional interactionsbetween Z and specific cellular factors are also involvedremains to be determined. In addition to these primaryZ-response regions, the NRP appears to contain other se-quences which can influence Z responsiveness. Experiments
-
VOL. 63, 1989
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3048 LIEBERMAN ET AL.
A 1B4 Cells B IB4 Cells C Vero Cells
9
N
9 9Q 9
Z - + - + +_
0.5 1.2 0.7 1.1 0.9 27 1.3 48 30
z - + + -+i
* ~ 0@I 4 1_ o
0.4 74 0.5 1.8 0.9 42 70
900 AA0
z - + - +
0
0 .6 3 2 1 0 1 .5
FIG. 8. Identification of a separate 1B4-specific Z-response region in the NRP. (A) Cotransfection of 1B4 lymphoblastoid cells with Z andthe NRP-CAT deletion derivatives revealed that plasmids pDH125 and pDH140 were poorly responsive in these cells. This suggests that theregion (-644 to - 1020) deleted from these plasmids contains essential sequences. (B) Transfer of the region from -644 to -902 into A10-CAT(pMH103) conferred Z responsiveness in 1B4 cells. This contrasts with the inability of the NRP-CAT (pPL52) (-155 to +40) to respond inthese cells. (C) Transfection of Vero cells with pMH103 confirmed the B-cell specificity of the region from -644 to -902, since pMH103 wasnot activated by Z in Vero cells. In these experiments, cells were transfected with 2.0 ,ug of target DNA and 2.0 ,ug of pBR322 or Z effectorDNA, and the percent acetylation is indicated.
with the deletion variants suggested the existence of anegative upstream regulatory region between -155 and-417. The constitutive activity of the NRP-CAT series ofconstructions was too low for us to assess the effect of theregion from -155 to -417 on basal promoter strength.However, in a similar series of constructions linked to therightward-directed promoter, removal of the region from-155 to -417 resulted in a three- to fivefold increase in basallevels of expression (Hardwick, in preparation). The regionbetween -417 and -644 had a positive effect on Z respon-siveness. Since no Z-binding sites are detectable in thisregion (Lieberman et al., submitted), it is assumed that thisregion also modifies intrinsic promoter strength and there-fore that these two regions are affecting Z responsivenessindirectly rather than directly.The divergent NRP region overlaps with the recently
identified origin of the lytic replication, ori-lyt (20). Both ofthe Z-response regions identified here are located within theori-lyt boundaries. The region from -644 to -902 fallswithin the segment of ori-lyt which can be replaced byenhancer sequences from the CMV immediate-early gene.This enhancer function would be consistent with our abilityto move this region into A10-CAT and transfer Z responsive-ness. The inclusion of the Z-response region from -7 to-155 within ori-lyt may reflect a requirement for transcrip-tional activation to maximize ori-lyt function, although amore direct role for Z in the replication process has not yetbeen eliminated. The presence of ori-lyt sequences withinthe NRP constructions would not have influenced the out-come of our transfection assays, since the majority of theexperiments were performed with EBV-negative Vero cellsand, for the Z-response region from -644 to -902, thesequences transferred to A10-CAT do not themselves con-stitute a functional origin.
ACKNOWLEDGMENTS
We thank Gary S. Hayward for helpful discussions; Mario Amzelfor guidance in the selection of a suitable BHLF1 synthetic peptide;Mabel Chiu, Barbara Daily, and Marilyn Cox for technical assis-tance; and Pamela Wright and Sarah Heaggans for assistance withmanuscript preparation.
This work was funded by Public Health Service grants CA30356(to S.D.H.) and CA43532 (to J.M.H.) from the National Cancer
Institute. Support for P.M.L. came in part from Training Programgrant CA09243 from the National Cancer Institute.
LITERATURE CITED1. Arrand, J. R., and L. Rymo. 1982. Characterization of the major
Epstein-Barr virus-specific RNA in Burkitt lymphoma-derivedcells. J. Virol. 41:376-379.
2. Aviv, H., and P. Leder. 1972. Purification of biologically activeglobin messenger RNA by chromatography on oligothymidylicacid-cellulose. Proc. Natl. Acad. Sci. USA 69:1408-1412.
3. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J.Farrell, T. J. Gibson, G. Hatfull, G. S. Hudson, S. C. Satchwell,C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNAsequence and expression of the B95-8 Epstein-Barr virus ge-nome. Nature (London) 310:207-211.
4. Bell, R. M. 1986. Protein kinase C activation by diacylglycerolsecond messengers. Cell 45:631-632.
5. Berk, A. J., and P. A. Sharp. 1977. Sizing and mapping of earlyadenovirus mRNA's by gel electrophoresis of S1 endonuclease-digested hybrids. Cell 12:721-732.
6. Biggin, M., M. Bodescot, M. Perricaudet, and P. Farrell. 1987.Epstein-Barr virus gene expression in P3HRl-superinfectedRaji cells. J. Virol. 61:3120-3132.
7. Chevallier-Greco, A., E. Manet, P. Chavrier, C. Mosnier, J.Daillie, and A. Sergeant. 1986. Both Epstein-Barr virus (EBV)-encoded trans-acting factors, EB1 and EB2, are required toactivate transcription from the EBV early promoter. EMBO J.5:3243-3249.
8. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J,Rutter. 1979. Isolation of biologically active ribonucleic acidfrom sources enriched in ribonuclease. Biochemistry 18:5294-5299.
9. Cho, M.-S., L. Gissmann, and S. D. Hayward. 1984. Epstein-Barr virus (P3HR-1) defective DNA codes for components ofboth the early antigen and viral capsid antigen complexes.Virology 137:9-19.
10. Cho, M.-S., K.-T. Jeang, and S. D. Hayward. 1985. Localizationof the coding region for an Epstein-Barr virus early antigen andinducible expression of this 60-kilodalton nuclear protein intransfected fibroblast cell lines. J. Virol. 56:852-859.
11. Chou, P. Y., and G. D. Fasman. 1974. Conformational parame-ters for amino-acids in helical P-sheet and random coil regions.Biochemistry 13:211-222.
12. Cleary, M. L., S. D. Smith, and J. Sklar. 1986. Cloning andstructural analysis of cDNAs for bcl-2/immunoglobulin tran-script resulting from the t(14:18) translocation. Cell 47:19-28.
13. Countryman, J., and G. Miller. 1985. Activation of expression
J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

CELL-SPECIFIC Z-RESPONSE SIGNAL REGIONS 3049
of latent Epstein-Barr herpesvirus after gene transfer with asmall cloned subfragment of heterogeneous viral DNA. Proc.Natl. Acad. Sci. USA 82:4085-4089.
14. Dillner, J., B. Kallin, H. Alexander, I. Ernberg, M. Uno, Y. Ono,G. Klein, and R. Lerner. 1986. An Epstein-Barr virus (EBV)-determined nuclear antigen (EBNA5) partly encoded by thetransformation-associated Bam-WYH region of EBV DNA:preferential expression in lymphoblastoid cell lines. Proc. Natl.Acad. Sci. USA 83:6641-6645.
15. Faggioni, A., C. Zampetta, S. Grimaldi, G. Barile, L. Frati, andJ. Lazdins. 1986. Calcium modulation activates Epstein-Barrvirus genome in latently infected cells. Science 232:1554-1556.
16. Fennewald, S., V. van Santen, and E. Kieff. 1984. Nucleotidesequence of an mRNA transcribed in latent growth-transforming virus infection indicates that it may encode amembrane protein. J. Virol. 51:411-419.
17. Freese, U. K., G. Laux, J. Hudewentz, E. Schwarz, and G. W.Bornkamm. 1983. Two distant clusters of partially homologoussmall repeats of Epstein-Barr virus are transcribed upon induc-tion of an abortive or lytic cycle of the virus. J. Virol. 48:731-743.
18. Gerber, P. 1972. Activation of Epstein-Barr virus by 5-bro-modeoxy-uridine in "virus-free" human cells. Proc. NatI.Acad. Sci. USA 69:83-85.
19. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
20. Hammerschmidt, W., and B. Sugden. 1988. Identification andcharacterization of ori-Lyt, a lytic origin of DNA replication ofEpstein-Barr virus. Cell 55:427-433.
21. Hampar, B., J. G. Derge, L. M. Martos, and J. L. Walker. 1972.Synthesis of Epstein-Barr virus after activation of the viralgenome in a "virus-negative' human lymphoblastoid cell (Raji)made resistant to 5-bromodeoxyuridine. Proc. Natl. Acad. Sci.USA 69:78-82.
22. Hardwick, J. M., P. M. Lieberman, and S. D. Hayward. 1988. Anew Epstein-Barr virus transactivator, R. induces expression ofa cytoplasmic early antigen. J. Virol. 62:2274-2284.
23. Hennessy, K., S. Fennewald, M. Hummel, T. Cole, and E. Kieff.1984. A membrane protein encoded by Epstein-Barr virus inlatent growth transforming infection. Proc. Natl. Acad. Sci.USA 81:7209-7211.
24. Hennessy, K., S. Fennewald, and E. Kieff. 1985. A third viralnuclear protein in lymphoblasts immortalized by Epstein-Barrvirus. Proc. Natl. Acad. Sci. USA 82:5944-5948.
25. Hennessy, K., and E. Kieff. 1983. One of two Epstein-Barr virusnuclear antigens contains a glycine-alanine copolymer domain.Proc. NatI. Acad. Sci. USA 80:5665-5669.
26. Hennessy, K., and E. Kieff. 1985. A second nuclear protein isencoded by Epstein-Barr virus in latent infection. Science227:1238-1240.
27. Hennessy, K., F. Wang, E. Woodland-Bushman, and E. Kieff.1986. Definitive identification of a member of the Epstein-Barrvirus nuclear protein 3 family. Proc. Natl. Acad. Sci. USA83:5693-5697.
28. Hudson, G. S., P. Farrell, and B. G. Barrell. 1985. Two relatedbut differentially expressed potential membrane proteins en-coded by the EcoRI Dhet region of Epstein-Barr virus B95-8. J.Virol. 53:528-535.
29. Jeang, K.-T., and S. D. Hayward. 1983. Organization of theEpstein-Barr virus DNA molecule. 111. Location of the P3HR-1deletion junction and characterization of the Notl repeat unitsthat form part of the template for an abundant 12-0-tetrade-canoylphorbol-13-acetate-induced mRNA transcript. J. Virol.48:135-148.
30. Jeang, K.-T., D. R. Rawlins, P. J. Rosenfeld, J. H. Shero, T. J.Kelly, and G. S. Hayward. 1987. Multiple tandemly repeatedbinding sites for cellular nuclear factor 1 that surround the majorimmediate-early promoters of simian and human cytomegalovi-rus. J. Virol. 61:1559-1570.
31. Joab, I., D. T. Rowe, M. Bodescot, J. C. Nicolas, P. J. Farrell,and M. Perricaudet. 1987. Mapping of the gene coding forEpstein-Barr virus determined nuclear antigen EBNA-3 and its
transient overexpression in a human cell line by using anadenovirus expression vector. J. Virol. 61:3340-3344.
32. Kallin, B., J. Dillner, 1. Ernberg, B. R. Ehlin-Henriksson, A.Rosen, W. Henle, G. Henle, and G. Klein. 1986. Four virallydetermined nuclear antigens are expressed in Epstein-Barr virustransformed cells. Proc. Natl. Acad. Sci. USA 83:1499-1503.
33. Kenney, S., J. Kamine, D. Markovitz, R. Fenrick, and J. Pagano.1988. An Epstein-Barr virus immediate-early gene producttrans-activates gene expression from the human immunodefi-ciency virus long terminal repeat. Proc. Natl. Acad. Sci. USA85:1652-1656.
34. Kieff, E., K. Hennessy, S. Fennewald, T. Matsuo, T. Dambaugh,M. Heller, and M. Hummel. 1985. Biochemistry of latent Ep-stein-Barr virus infection and associated cell growth transfor-mation, p. 323-339. In G. M. Lenoir, G. T. O'Conor, andC. L. M. Olweny (ed.). Burkitt's lymphoma: a human cancermodel. International Agency for Research on Cancer, Lyon,France.
35. Laimins, L. A., P. Gruss, R. Pozzatti, and G. Khoury. 1984.Characterization of enhancer elements in the long terminalrepeat of Moloney murine sarcoma virus. J. Virol. 49:183-189.
36. Laux, G., U. K. Freese, and G. W. Bornkamm. 1985. Structureand evolution of two related transcription units of Epstein-Barrvirus carrying small tandem repeats. J. Virol. 56:987-995.
37. Laux, G., M. Perricaudet, and P. J. Farrell. 1988. A splicedEpstein-Barr virus gene expressed in immortalized lymphocytesis created by circularization of the linear viral genome. EMBOJ. 7:769-774.
38. Lerner, M., N. Andrews, G. Miller, and J. Steitz. 1981. Twosmall RNAs encoded by Epstein-Barr virus and complexed withprotein are precipitated by antibody from patients with systemiclupus erythematosus. Proc. Natl. Acad. Sci. USA 78:805-809.
39. Lieberman, P. M., P. O'Hare, G. S. Hayward, and S. D.Hayward. 1986. Promiscuous tr{anis-activation of gene expres-sion by an Epstein-Barr virus-encoded early nuclear protein. J.Virol. 60:140-148.
40. Luka, J., B. Kallin, and G. Klein. 1979. Induction of theEpstein-Barr virus (EBV) cycle in latently infected cells byn-butyrate. Virology 94:228-231.
41. Miller, G. 1985. Epstein-Barr virus. p. 563-589. In B. N. Fields(ed.). Virology. Raven Press, New York.
42. Miller, G., M. Rabson, and L. Heston. 1984. Epstein-Barr viruswith heterogeneous DNA disrupts latency. J. Virol. 50:174-182.
43. O'Hare, P., and G. S. Hayward. 1984. Expression of recombi-nant genes containing herpes simplex virus delayed-early andimmediate-early regulatory regions and trans-activation by her-pesvirus infection. J. Virol. 52:522-531.
44. Pearson, G. R., J. Luka, L. Petti, J. Sample, M. Birkenbach, D.Braun, and E. Kieff. 1987. Identification of an Epstein-Barrvirus early gene encoding a second component of the restrictedearly anitigen complex. Virology 160:151-161.
45. Pearson, G. R., B. Vroman, B. Chase, T. Sculley, M. Hummel,and E. Kieff. 1983. Identification of polypeptide components ofthe Epstein-Barr virus early antigen complex using monoclonalantibodies. J. Virol. 47:193-201.
46. Petti, L., and E. Kieff. 1988. A sixth Epstein-Barr virus nuclearprotein (EBNA3B) is expressed in latently infected growth-transformed lymphocytes. J. Virol. 62:2173-2178.
47. Petti, L., J. Sample, F. Wang, and E. Kieff. 1988. A fifthEpstein-Barr virus nuclear protein (EBNA3C) is expressed inlatently infected growth-transformed lymphocytes. J. Virol.62:1330-1338.
48. Pizzorno, M., P. O'Hare, L. Sha, R. LaFemina, and G. S.Hayward. 1988. tratis-Activation and autoregulation of geneexpression by the immediate-early region 2 gene products ofhuman cytomegalovirus. J. Virol. 62:1167-1179.
49. Purtilo, D. T. 1985. Association of Epstein-Barr virus andlymphoproliferative diseases in immune deficient persons. p.3-17. Ini P. H. Levine. D. V. Ablashi, G. R. Pearson. and S. D.Kotteridis (ed.). EBV and associated diseases. Martinus NijhoffPublishing. The Hague. The Netherlands.
50. Rabson, M., L. Heston, and G. Miller. 1983. Identification of a
rare Epstein-Barr virus that enhances early antigen expression.
VOL. 63. 1989
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from

3050 LIEBERMAN ET AL.
Proc. Natl. Acad. Sci. USA 80:2762-2766.51. Sample, J., G. Lancz, and M. Nonoyama. 1986. Mapping of
genes in Ba,inHI-M of Epstein-Barr virus DNA that may deter-mine the fate of viral infection. J. Virol. 57:145-154.
52. Sample, J., A. Tanaka, G. Lancz, and M. Nonoyama. 1984.Identification of Epstein-Barr virus genes expressed during theearly phase of virus replication and during lymphocyte immor-talization. Virology 139:1-10.
53. Seibl, R., M. Motz, and H. Wolf. 1986. Strain-specific transcrip-tion and translation of the BalmHl-Z area of Epstein-Barr virus.
J. Virol. 60:902-909.54. Siomi, H., H. Shida, S. H. Nam, T. Nosaka, M. Maki, and M.
Hatanaka. 1988. Sequence requirements for nucleolar localiza-tion of human T cell leukemia virus type 1 pX protein, whichregulates viral RNA processing. Cell 55:197-209.
55. Sutcliffe, J. G., T. M. Shinnick, N. Green, F.-T. Liu, H. L.Niman, and R. A. Lerner. 1980. Chemical synthesis of a
polypeptide predicted from nucleotide sequence allows detec-
tion of a new retroviral gene product. Nature (London) 287:801-805.
56. Takada, K., N. Shimizu, S. Sakuma, and Y. Ono. 1986. fians-
Activation of the latent Epstein-Barr virus (EBV) genome aftertransfection of the EBV DNA fragment. J. Virol. 57:1016-1022.
57. Tovey, M. G., G. Lenoir, and J. Begon-Louis. 1978. Activationof latent Epstein-Barr virus by antibody to human IgM. Nature(London) 276:270-272.
58. Weeks, D. L., and N. C. Jones. 1983. EIA control of gene
expression is mediated by sequences 5' to the transcriptionalstarts of the early viral genes. Mol. Cell. Biol. 3:1222-1234.
59. Wong, K.-M., and A. J. Levine. 1986. Identification and map-
ping of Epstein-Barr virus early antigens and demonstration of a
viral gene activator that functions in trans. J. Virol. 60:149-156.60. zur Hausen, H., F. J. O'Neill, U. K. Freese, and E. Hecker. 1978.
Persisting oncogenic herpesvirus induced by the tumor pro-
moter TPA. Nature (London) 272:373-375.
J. VIROL.
on May 7, 2018 by guest
http://jvi.asm.org/
Dow
nloaded from










![Cianciana luoghi noti e meno noti [mobile]](https://img.pdfslide.net/doc/110x75/568ca92b1a28ab186d9c5c3a/cianciana-luoghi-noti-e-meno-noti-mobile.jpg)


















