Embed Size (px)
Citation preview

1218
BIOLOGY OF REPRODUCTION 72, 1218–1223 (2005)Published online before print 19 January 2005.DOI 10.1095/biolreprod.104.038141
Role of Adenosine Triphosphate, Active Mitochondria, and Microtubulesin the Acquisition of Developmental Competence of ParthenogeneticallyActivated Pig Oocytes1
Tiziana A.L. Brevini, Rita Vassena, Chiara Francisci, and Fulvio Gandolfi2
Department of Anatomy of Domestic Animals, Faculty of Veterinary Medicine, University of Milan,10-20133 Milano, Italy
ABSTRACT
The purpose of this work was to determine the mechanismsregulating the acquisition of cytoplasmic maturation and em-bryonic developmental competence in pig oocytes. The presenceor the absence of porcine follicular fluid (pff; 25% or 0%) inthe maturation medium was used as a means to achieve com-plete nuclear maturation accompanied or not accompanied bycytoplasmic maturation. ATP content, active mitochondria re-location, and microtubule distribution were analyzed at differ-ent times during in vitro maturation (IVM). While nuclear mat-uration did not differ among the two groups, parthenogeneticembryonic development was significantly higher (41.5%) in the25% pff group than in the 0% pff group (19.0%) with blasto-cysts that had a significantly higher number of blastomeres (76.16 6.3, and 47.2 6 6.5, respectively). Oocyte ATP content in-creased significantly during IVM, but at the end of maturationno significant differences were observed between high- and low-competence oocytes. An extensive relocation of mitochondriato the inner cytoplasm during IVM together with the formationof a well-developed mesh of cytoplasmic microtubules was ob-served only in the high-competence oocyte group. However, nodifferences in the formation of microtubules associated with themeiotic spindles were observed between high- and low-compe-tence groups. We conclude that low developmental competenceis associated with the lack of a microtubule cytoplasmic net-work, which prevents correct relocation of mitochondria and islikely to reflect a more generally altered compartmentalizationof the ooplasm. This can be independent from the formation ofthe microtubule machinery required for the completion of chro-mosome disjunctions and does not affect the overall ATP con-tent.
ATP, early development, gamete biology, meiosis, microtubule,mitochondria, ovum, parthenogenesis, pig oocyte
INTRODUCTION
In vitro production of mammalian embryos has a widevariety of applications ranging from production of clonedanimals and genetic engineering, to treatment of infertilityby assisted reproductive technologies. Recently, there hasbeen an increasing interest in producing large quantities ofpig embryos through in vitro maturation (IVM)/in vitro fer-
1Supported by MIUR-COFIN 2002.074357.2Correspondence: Fulvio Gandolfi, Istituto di Anatomia degli Animali Do-mestici, via Celoria, 10-20133 Milano, Italy. FAX: 39 025 031 7980;e-mail: [email protected]
Received: 16 November 2004.First decision: 16 December 2004.Accepted: 17 January 2005.Q 2005 by the Society for the Study of Reproduction, Inc.ISSN: 0006-3363. http://www.biolreprod.org
tilization techniques because of the physiological similari-ties of pig oocytes to those of humans, which makes theseanimals a valuable experimental model for basic as well asbiomedical research [1].
While the only parameter that can reliably measure theefficiency of an in vitro system is the ability to produce aviable fetus from a cultured embryo, for practical and eco-nomical reasons, the most commonly used measure of ef-ficiency remains the proportion of immature oocytes de-veloping to the blastocyst stage.
Several studies have clearly demonstrated that oocytequality is the most crucial factor affecting blastocyst rate[2]; however, the essence of oocyte quality remains elusiveand it certainly includes many aspects [3]. We have previ-ously identified post-transcriptional modification of the ma-ternal mRNA molecules stored in the ooplasm as one ofthese aspects [4, 5]. In the present work we took into con-sideration the role of cytoplasmic compartmentalization,which has been suggested to play an important role in thecompletion of coordinate nuclear and cytoplasmic oocytematuration [6]. In particular, the activity and cytoplasmicdistribution of mitochondria was chosen as an easily de-tectable marker of cytoplasm compartmentalization; micro-tubules were also studied as the recognized responsiblepartner for the establishment and maintenance of cellularasymmetry during oocyte maturation [7].
The activity and organization of mitochondria are nec-essary features among the diverse events involved in cy-toplasmic maturation [8]. The primary function of mito-chondria is to generate ATP, which is necessary for severalfunctions, including motility, maintenance of cellular ho-meostasis, and regulation of cell survival [9]. The patternof mitochondria distribution and their metabolic activity un-dergo changes during oocyte maturation in many species,including mouse [8, 10], cow [11], human [12], and pig[13].
The transfer of mitochondria within different areas ofthe cell is mediated by a cytoskeletal network of microtu-bules [14]. Microtubules, homologous polymers of a- andb-tubulin, are dynamic components of the cell cytoskeleton;they are ubiquitously present in mammalian cells and areinvolved in diverse functions such as determining cellshape and movement, transportation of molecules and or-ganelles, meiosis, and mitosis. In the mouse oocyte, twopopulations of microtubules can be identified: one linkedto the DNA, and another located in the cytoplasm [15].Despite the role that microtubules play during meiotic mat-uration and competence acquisition, relatively little infor-mation is available for species other than the mouse. Inparticular, no clear data are available on microtubule for-mation and distribution during pig oocyte maturation. In

1219ATP, MITOCHONDRIA, AND TUBULIN IN OOCYTE COMPETENCE
TABLE 1. Effect of pff during IVM on oocyte maturation, parthenogenetic activation, embryo development, and blastomere number.a
Parameter Oocytes
pff concentration
0% 25%
M IIActivated with 1 PN and 1 PBBlastocyst rateb
Blastomeres/BL 6 SEMc
6690
396
23/33 (81.8%)29/42 (69%)35/184 (19.0%)47.2 6 6.5(n 5 15)
28/33 (84.8%)36/48 (75%)88/212 (41.5%)76.1 6 6.3(n 5 20)
a Data were obtained through three replicates. M II 5 metaphase II; PN, pronucleus; PB, polar body; BL, blastocyst.b Statistically different distribution as determined with X2 analysis (P 5 0.009).c Values are statistically different as determined with one-way ANOVA (P 5 0.004).
fact, some studies described only a microtubule populationlinked to the DNA, while a cytoplasmic microtubule net-work has not been identified at all [16, 17]; in others, a finemesh of cytoplasmic microtubules has been reported onlyaround the time of germinal vesicle breakdown [18, 19].
To identify the cellular mechanisms that are associatedwith a reduced viability of pig oocytes we used an IVMsystem in which pig oocytes can complete the meiotic pro-cess at a high rate, but they predictably show either a lowor a high developmental competence to the blastocyst stagein vitro, following parthenogenetic activation. This system,therefore, uncouples nuclear and cytoplasmic maturation,making it possible to study the role of ATP content, mi-gration of active mitochondria, and microtubule dynamicsthroughout the entire process of IVM.
MATERIALS AND METHODSUnless otherwise indicated, chemicals were purchased from Sigma-
Aldrich (Milan, Italy).
Porcine Follicular Fluid PreparationOvaries were collected at the slaughterhouse. Porcine follicular fluid
(pff) was aspirated from follicles of prepuberal gilts with a diameter of 2–7mm in 15-ml tubes. Cellular debris was allowed to settle for 20 min, andtubes were centrifuged for 15 min at 400 3 g at 48C. The supernatant wasaspirated and filtered through a 0.22-mm membrane (Sterilin, Milan, Italy).Aliquots of 2.5 ml of pff were stored at 2208C until use.
Recovery and Maturation of Cumulus-Oocyte ComplexesCumulus-oocyte complexes (COCs) were obtained from ovaries of
slaughtered gilts weighing 160 6 15 kg and aged 240 6 15 days. Follicleswith a diameter of 2–7 mm were aspirated with an 18-gauge needle andvacuum pressure of 50 ml/min. The follicles aspirate was collected in 15-ml tubes, and COCs were washed twice in prewarmed (378C) aspirationmedium. The medium was composed of TCM-199 supplemented with 6.5mg/ml Hepes, 1.1 mg/ml sodium bicarbonate, 4 mg/ml BSA, 75 mg/mlpenicillin, and 50 mg/ml streptomycin (199D). Only COCs with large,compact cumulus and homogeneous oocyte cytoplasm were selected forIVM. IVM was performed as previously described [20] with minor mod-ifications. Briefly, COCs were cultured in TCM-199 supplemented with1.1 mg/ml sodium bicarbonate, 0.1 mg/ml sodium pyruvate, 0.5 mM cys-teamin, 8.2 mg/ml insulin, 10 ng/ml epidermal growth factor, 1 mM di-butryl cAMP, 0.5 IU/ml porcine FSH:LH (Pluset; Serono, Rome, Italy),1.0 mg/ml 17 b-estradiol, 75 mg/ml penicillin, and 50 mg/ml streptomycin(199-IVM). Based on previous preliminary data [21], 25% or no pff wasadded to 199-IVM medium. COCs were matured in this medium for 22 hat 38.58C in an atmosphere of 5% CO2 in air. COCs were washed in freshmedium and matured for an additional 24 h in 199-IVM without dibutrylcAMP.
Meiotic AssessmentAt the end of each IVM experiment, a representative sample of oocytes
(approximately 10% of the total 552 oocytes used for the developmentalstudies) was denuded by gently pipetting in medium 199D containing0.1% hyaluronidase at 38.58C, washed in medium 199D, and mounted on
microscope slides. Samples were fixed for 48 h in acetic acid:ethanol (1:3) and stained with 0.1% (v/v) acetic orcein for 30 min. They were de-stained in glycerol:acetic acid:water (1:1:3) and meiotic stage was evalu-ated using a Nikon Eclipse E600 microscope (Nikon Instruments, SestoFiorentino, Italy) at 2003 and 4003 magnification.
Other oocytes of each group were used for the measurement of ATPcontent or for the staining of active mitochondria and microtubules. Allthe remaining oocytes were parthenogenetically activated and cultured invitro. The relative procedures are described below.
Parthenogenetic Activation and In Vitro CultureAfter IVM, oocytes were denuded as described above, washed for 10
min in TCM-199 supplemented with 20% (v/v) fetal calf serum (FCS),and then incubated in TALP medium for 30 min at 38.58C. Parthenoge-netic activation was performed according to the method described by Bo-quest et al. [22] by sequentially exposing the oocytes to 5 mM ionomycinin TALP for 5 min at 38.58C in the dark; and to 2 mM 6-DMAP inmedium NCSU-23 [23] for 3 h at 38.58C in an atmosphere of 5% CO2,5% O2, and 90% N2. In a separate experiment, 16 h from the end of theincubation in 6-DMAP, the incidence of second polar body retention wasassessed by acetic orcein staining as described above and compared be-tween the two maturation treatments in a total of 90 oocytes.
Presumptive parthenotes were washed thoroughly in NCSU-23 andcultured in groups of 25–35 in 50-ml NCSU-23 drops under mineral oilat 38.58C in the same atmosphere as above. Embryonic cleavage wasrecorded 48 h postactivation. On Day 5 postactivation, half of the mediumwas replaced with fresh NCSU-23 containing 20% (v/v) FCS to reach afinal FCS concentration of 10% (v/v) in the in vitro culture drop. At Day7 postactivation, in vitro culture was interrupted and embryonic develop-ment was evaluated. The experiment was replicated five times.
Measurement of the ATP Content of OocytesATP content of denuded, nonmatured and matured oocytes (n 5 282)
was assessed using a commercial assay based on a luciferin-luciferasereaction (Bioluminescent Somatic Cell Assay Kit; Sigma) as previouslydescribed [11]. Briefly, samples were rinsed twice in PBS and then trans-ferred individually in 50 ml of PBS (sample buffer) into plastic tubes onmelting ice. Then, 100 ml of somatic cell reagent (FL-SAR) was added toeach tube. Samples were incubated on melting ice for exactly 5 min inthe dark, and then 100 ml of assay buffer (dilution 1:25 with ATP assaymix dilution buffer, FL-AAB) was added, and the tubes were kept forexactly 5 min at room temperature in the dark. ATP content was measuredwith a luminometer (Stratec Biomedical Systems AG, Birkenfeld, Ger-many). An eight-point standard curve (0–7 pmol/tube) was included ineach experiment (n 5 3), and the content of ATP was determined fromthe formula for the standard curve.
Staining of Active Mitochondria and MicrotubulesOocytes at different stages of maturation were stained with Mito-
Tracker Orange CMTM-Ros (Molecular Probes Europe, Leiden, The Neth-erlands). A stock solution of the dye was prepared according to manufac-turer’s specifications. MitoTracker Orange CMTM-Ros stains selectivelyactive mitochondria and is well retained after fixation; the dye was usedat a concentration of 250 nM in medium 199D for 30 min at 38.58C. Afterstaining, oocytes were briefly rinsed in PBS and fixed in 3.7% parafor-maldehyde in PBS for 30 min at 378C. DNA was stained with 49,69-diamidino-2-phenylidole (DAPI; 0.2 mg/ml), and samples were mountedon glass slides in a glycerol-based mounting medium. Oocytes were storedbelow 08C until confocal microscopy was performed.
1220 BREVINI ET AL.
TABLE 2. ATP content of single oocytes at collection and after differentIVM treatments.
Stage of IVM Treatment Oocytes (n)pmol ATP/oocyte
6 SD
At collectionM IIM II
—0% pff
25% pff
366972
1.21 6 0.10a
3.08 6 0.42b
3.60 6 0.51b
a,b Different superscripts within the same column indicate statistically dif-ferent values (P , 0.05).
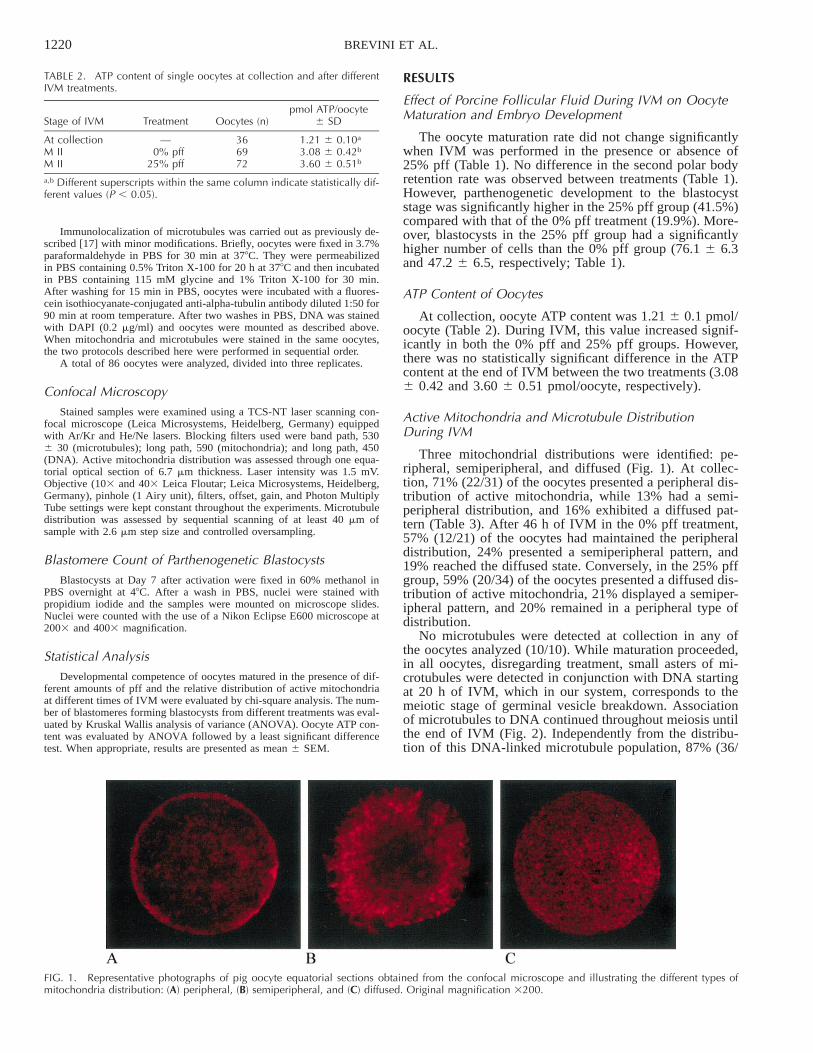
FIG. 1. Representative photographs of pig oocyte equatorial sections obtained from the confocal microscope and illustrating the different types ofmitochondria distribution: (A) peripheral, (B) semiperipheral, and (C) diffused. Original magnification 3200.
Immunolocalization of microtubules was carried out as previously de-scribed [17] with minor modifications. Briefly, oocytes were fixed in 3.7%paraformaldehyde in PBS for 30 min at 378C. They were permeabilizedin PBS containing 0.5% Triton X-100 for 20 h at 378C and then incubatedin PBS containing 115 mM glycine and 1% Triton X-100 for 30 min.After washing for 15 min in PBS, oocytes were incubated with a fluores-cein isothiocyanate-conjugated anti-alpha-tubulin antibody diluted 1:50 for90 min at room temperature. After two washes in PBS, DNA was stainedwith DAPI (0.2 mg/ml) and oocytes were mounted as described above.When mitochondria and microtubules were stained in the same oocytes,the two protocols described here were performed in sequential order.
A total of 86 oocytes were analyzed, divided into three replicates.
Confocal MicroscopyStained samples were examined using a TCS-NT laser scanning con-
focal microscope (Leica Microsystems, Heidelberg, Germany) equippedwith Ar/Kr and He/Ne lasers. Blocking filters used were band path, 5306 30 (microtubules); long path, 590 (mitochondria); and long path, 450(DNA). Active mitochondria distribution was assessed through one equa-torial optical section of 6.7 mm thickness. Laser intensity was 1.5 mV.Objective (103 and 403 Leica Floutar; Leica Microsystems, Heidelberg,Germany), pinhole (1 Airy unit), filters, offset, gain, and Photon MultiplyTube settings were kept constant throughout the experiments. Microtubuledistribution was assessed by sequential scanning of at least 40 mm ofsample with 2.6 mm step size and controlled oversampling.
Blastomere Count of Parthenogenetic BlastocystsBlastocysts at Day 7 after activation were fixed in 60% methanol in
PBS overnight at 48C. After a wash in PBS, nuclei were stained withpropidium iodide and the samples were mounted on microscope slides.Nuclei were counted with the use of a Nikon Eclipse E600 microscope at2003 and 4003 magnification.
Statistical AnalysisDevelopmental competence of oocytes matured in the presence of dif-
ferent amounts of pff and the relative distribution of active mitochondriaat different times of IVM were evaluated by chi-square analysis. The num-ber of blastomeres forming blastocysts from different treatments was eval-uated by Kruskal Wallis analysis of variance (ANOVA). Oocyte ATP con-tent was evaluated by ANOVA followed by a least significant differencetest. When appropriate, results are presented as mean 6 SEM.
RESULTS
Effect of Porcine Follicular Fluid During IVM on OocyteMaturation and Embryo Development
The oocyte maturation rate did not change significantlywhen IVM was performed in the presence or absence of25% pff (Table 1). No difference in the second polar bodyretention rate was observed between treatments (Table 1).However, parthenogenetic development to the blastocyststage was significantly higher in the 25% pff group (41.5%)compared with that of the 0% pff treatment (19.9%). More-over, blastocysts in the 25% pff group had a significantlyhigher number of cells than the 0% pff group (76.1 6 6.3and 47.2 6 6.5, respectively; Table 1).
ATP Content of Oocytes
At collection, oocyte ATP content was 1.21 6 0.1 pmol/oocyte (Table 2). During IVM, this value increased signif-icantly in both the 0% pff and 25% pff groups. However,there was no statistically significant difference in the ATPcontent at the end of IVM between the two treatments (3.086 0.42 and 3.60 6 0.51 pmol/oocyte, respectively).
Active Mitochondria and Microtubule DistributionDuring IVM
Three mitochondrial distributions were identified: pe-ripheral, semiperipheral, and diffused (Fig. 1). At collec-tion, 71% (22/31) of the oocytes presented a peripheral dis-tribution of active mitochondria, while 13% had a semi-peripheral distribution, and 16% exhibited a diffused pat-tern (Table 3). After 46 h of IVM in the 0% pff treatment,57% (12/21) of the oocytes had maintained the peripheraldistribution, 24% presented a semiperipheral pattern, and19% reached the diffused state. Conversely, in the 25% pffgroup, 59% (20/34) of the oocytes presented a diffused dis-tribution of active mitochondria, 21% displayed a semiper-ipheral pattern, and 20% remained in a peripheral type ofdistribution.
No microtubules were detected at collection in any ofthe oocytes analyzed (10/10). While maturation proceeded,in all oocytes, disregarding treatment, small asters of mi-crotubules were detected in conjunction with DNA startingat 20 h of IVM, which in our system, corresponds to themeiotic stage of germinal vesicle breakdown. Associationof microtubules to DNA continued throughout meiosis untilthe end of IVM (Fig. 2). Independently from the distribu-tion of this DNA-linked microtubule population, 87% (36/
1221ATP, MITOCHONDRIA, AND TUBULIN IN OOCYTE COMPETENCE
TABLE 3. Relative distribution of active mitochondria in oocytes at collection and after different IVM treatments.
Stage of IVM Treatment Peripheral (n) Semi-peripheral (n) Diffused (n)
At collectionMIIMII
—0% pff
25% pffa
71% (22/31)57% (12/21)20% (7/34)
13% (4/31)24% (5/21)20% (7/34)
16% (5/31)19% (4/21)60% (20/34)
a Overall distribution is different (P , 0.05) from the other two groups as determined with X2 analysis.
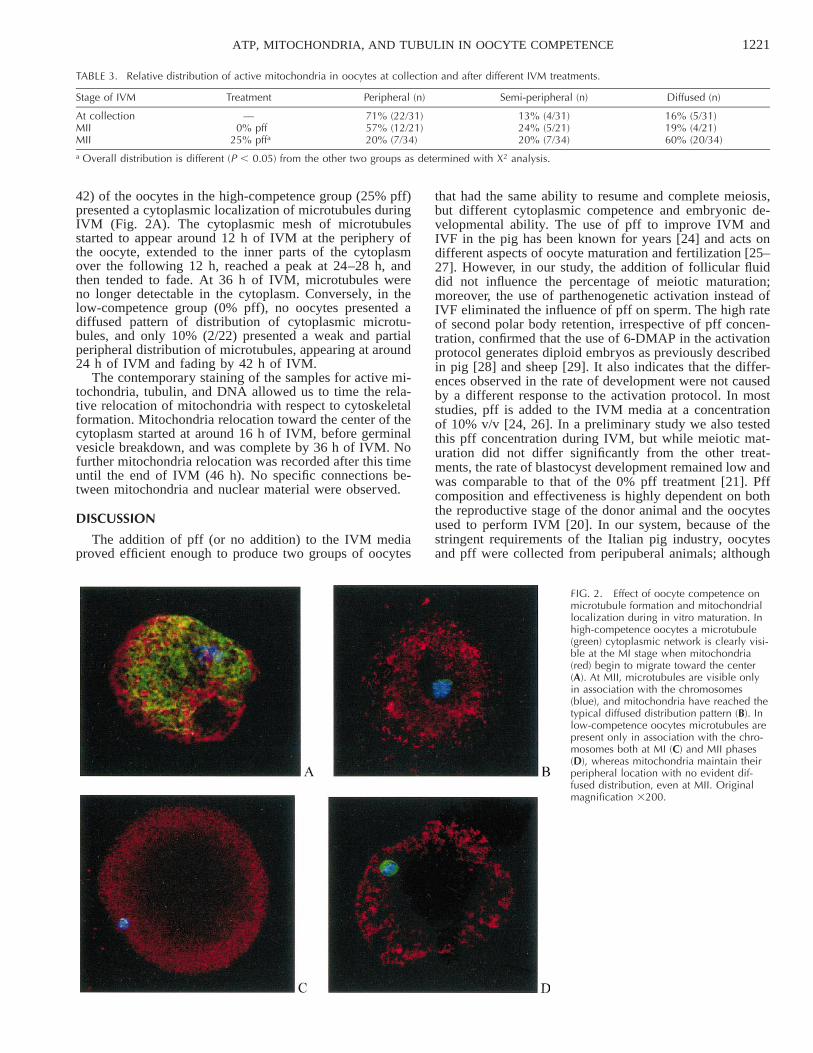
FIG. 2. Effect of oocyte competence onmicrotubule formation and mitochondriallocalization during in vitro maturation. Inhigh-competence oocytes a microtubule(green) cytoplasmic network is clearly visi-ble at the MI stage when mitochondria(red) begin to migrate toward the center(A). At MII, microtubules are visible onlyin association with the chromosomes(blue), and mitochondria have reached thetypical diffused distribution pattern (B). Inlow-competence oocytes microtubules arepresent only in association with the chro-mosomes both at MI (C) and MII phases(D), whereas mitochondria maintain theirperipheral location with no evident dif-fused distribution, even at MII. Originalmagnification 3200.
42) of the oocytes in the high-competence group (25% pff)presented a cytoplasmic localization of microtubules duringIVM (Fig. 2A). The cytoplasmic mesh of microtubulesstarted to appear around 12 h of IVM at the periphery ofthe oocyte, extended to the inner parts of the cytoplasmover the following 12 h, reached a peak at 24–28 h, andthen tended to fade. At 36 h of IVM, microtubules wereno longer detectable in the cytoplasm. Conversely, in thelow-competence group (0% pff), no oocytes presented adiffused pattern of distribution of cytoplasmic microtu-bules, and only 10% (2/22) presented a weak and partialperipheral distribution of microtubules, appearing at around24 h of IVM and fading by 42 h of IVM.
The contemporary staining of the samples for active mi-tochondria, tubulin, and DNA allowed us to time the rela-tive relocation of mitochondria with respect to cytoskeletalformation. Mitochondria relocation toward the center of thecytoplasm started at around 16 h of IVM, before germinalvesicle breakdown, and was complete by 36 h of IVM. Nofurther mitochondria relocation was recorded after this timeuntil the end of IVM (46 h). No specific connections be-tween mitochondria and nuclear material were observed.
DISCUSSION
The addition of pff (or no addition) to the IVM mediaproved efficient enough to produce two groups of oocytes
that had the same ability to resume and complete meiosis,but different cytoplasmic competence and embryonic de-velopmental ability. The use of pff to improve IVM andIVF in the pig has been known for years [24] and acts ondifferent aspects of oocyte maturation and fertilization [25–27]. However, in our study, the addition of follicular fluiddid not influence the percentage of meiotic maturation;moreover, the use of parthenogenetic activation instead ofIVF eliminated the influence of pff on sperm. The high rateof second polar body retention, irrespective of pff concen-tration, confirmed that the use of 6-DMAP in the activationprotocol generates diploid embryos as previously describedin pig [28] and sheep [29]. It also indicates that the differ-ences observed in the rate of development were not causedby a different response to the activation protocol. In moststudies, pff is added to the IVM media at a concentrationof 10% v/v [24, 26]. In a preliminary study we also testedthis pff concentration during IVM, but while meiotic mat-uration did not differ significantly from the other treat-ments, the rate of blastocyst development remained low andwas comparable to that of the 0% pff treatment [21]. Pffcomposition and effectiveness is highly dependent on boththe reproductive stage of the donor animal and the oocytesused to perform IVM [20]. In our system, because of thestringent requirements of the Italian pig industry, oocytesand pff were collected from peripuberal animals; although

1222 BREVINI ET AL.
no direct tests have been performed in this regard, it ispossible that this combination requires a higher pff concen-tration to be effective.
The role played by ATP in the acquisition of meioticcompetence has been studied in many species, but it re-mains difficult to define in detail [9]. Our results indicatea statistically significant increase during IVM of ATP con-tent in pig oocytes similar to what has been reported in thecow [11]. Recently, an increased oxidative activity of mi-tochondria has also been reported during IVM of pig oo-cytes [30]. To the contrary, no ATP increase was observedduring mouse oocyte maturation [31]. In cattle, COCs pre-senting a progressively worsening morphology displayed adecreasing average content of ATP, both at the beginning(germinal vesicle) and at the end of IVM (MII). Becauseoocyte morphology was linked to blastocyst rate, a relationbetween ATP content and developmental competence wasevident in this species [11]. This relation was not observedin our experiments because the increase in ATP during mat-uration was not influenced by the amount of pff employedduring IVM and its concentration was not related to oocytedevelopmental competence. Our observations agree with arecent study performed in pigs [32] in which it was ob-served that no significant differences in ATP content werepresent between oocytes matured in vivo or in vitro. There-fore, pigs may be different from other species in that ahigher rate of embryonic development had been associatedwith higher concentrations of ATP not only in cattle [11],but also in humans and mice [31].
Changes in mitochondrial organization are believed tobe a faithful indicator of oocyte capacity to sustain embry-onic development [33] and to occur during IVM in manyspecies including mouse [8, 10], cow [11], and human [12].Pig mitochondria migrate during maturation from the pe-ripheral part of the oocyte to the inner region of the cellboth in vitro [13] and in vivo [30], even though informationis scarce on the correlation between this phenomenon andsubsequent oocyte development. Our results indicate thatrelocation of active mitochondria is significantly more fre-quent in the oocyte group with high developmental com-petence. Conversely, little or no relocation was observedafter IVM in the low-competence group, suggesting that thelack of mitochondrial distribution through the ooplasm is amarker of cytoplasm immaturity and is strongly linked tolow developmental ability.
The transfer of mitochondria within different areas ofthe cytoplasm is mediated by the cytoskeletal network ofmicrotubules in the oocytes of mice [14] as well as pigs[13]. While there is general agreement on the involvementof microtubules in forming the meiotic spindle, the dynam-ics of cytoplasm-associated microtubules in pig oocytes re-mains to be elucidated. Some studies failed to identify acytoplasmic microtubule network altogether [16, 34], whileother reports describe only a finely distributed mesh of cy-toplasmic microtubules surrounding the germinal vesiclenucleus [18, 19, 35]. Therefore, the question remains openwhether the missed distribution of mitochondria during thematuration of defective oocytes was due to the lack of for-mation of an appropriate microtubule network or wascaused by an uncoupling between mitochondria and micro-tubule, possibly related to the lack of specific motor mol-ecules such as dynein and kinesin [36, 37]. Here we de-scribed the formation of a microtubule network in the oo-plasm of the high-competence group that began to formbefore germinal vesicle breakdown at 12 h of IVM fromthe periphery of the oocyte, extended to the inner parts of
the cytoplasm over the following 12 h, reached a peak at24–28 h, and then tended to fade. At 36 h of IVM, micro-tubules were no longer detectable in the cytoplasm, butpersisted until the end of maturation in correspondence withthe DNA. The contemporary staining of active mitochon-dria, DNA, and microtubules allowed us to demonstratethat the lack of relocation of active mitochondria to theinner part of the oocyte is related to the absence of anappropriate and timely formation of the microtubule net-work in the cytoplasm. In the oocytes belonging to the low-competence group we did not observe a diffused pattern ofdistribution of cytoplasmic microtubules, and only 10% ofthem presented a weak and partial peripheral distributionof microtubules, appearing at around 24 h of IVM and fad-ing by 42 h of IVM. This suggests that the lack of mito-chondria relocation observed in the defective oocyte groupmay be due to their inability to form a cytoplasmic micro-tubule network rather than to the inability of mitochondriato migrate along the tubules. The formation of a normalmeiotic spindle, observed in both groups of oocytes, ex-plains why the rate of meiotic progression to the secondmetaphase was not altered in the low-competence group.This observation clearly illustrates the hypothesis that de-fective oocyte developmental competence is due to the un-coupling of nuclear and cytoplasmic maturation. While thenormal formation of nucleus-associated microtubules al-lows the correct segregation of chromosomes during thereductive meiotic divisions, the lack of a microtubule cy-toplasmic network prevents a correct relocation of mito-chondria, which is likely to reflect a more generally alteredcompartmentalization of the ooplasm. It is not clear, how-ever, whether the missing relocation of mitochondria alsohas a direct effect because it was not linked to differencesin the total ATP content of the oocytes. However mito-chondria are also involved in Ca21 signaling during eggactivation [38], and the recent observation that estrogen re-ceptor beta is localized to mitochondria [39] suggests thata specific distribution of these organelles may be necessaryfor a correct oocyte response to estradiol-17b, which ispresent at high levels in preovulatory follicles.
We conclude that low developmental competence is as-sociated with the lack of a microtubule cytoplasmic net-work, which prevents the correct relocation of mitochon-dria, and is likely to reflect a more generally altered com-partmentalization of the ooplasm. This can occur indepen-dently from the formation of the microtubule machineryrequired for the completion of chromosome disjunctionsand does not affect the overall ATP content.
ACKNOWLEDGMENTS
We are grateful to the members of the Department of Obstetrics andGynaecology, University of Adelaide; Prof. David Armstrong for helpfuldiscussion and suggestions; Dr. Chris Grupen for sharing with us his vastknowledge of pig in vitro production; Ms. Melanie Bagg for technicalhelp in some preliminary experiments; Dr. Umberto Fascio, Director ofCIMA (Interdepartmental Center for Advanced Microscopy), Universityof Milan, for assistance with confocal microscopy data acquisition; and toProf. Daniela Ghisotti, Department of Biology, University of Milan, forhelp with ATP measurements.
REFERENCES
1. Prather RS, Hawley RJ, Carter DB, Lai L, Greenstein JL. Transgenicswine for biomedicine and agriculture. Theriogenology 2003; 59:115–123.
2. Sirard MA. Resumption of meiosis: mechanism involved in meioticprogression and its relation with developmental competence. Therio-genology 2001; 55:1241–1254.

1223ATP, MITOCHONDRIA, AND TUBULIN IN OOCYTE COMPETENCE
3. Duranthon V, Renard JP. The developmental competence of mam-malian oocytes: a convenient but biologically fuzzy concept. Therio-genology 2001; 55:1277–1289.
4. Brevini-Gandolfi TAL, Gandolfi F. The maternal legacy to the em-bryo: cytoplasmic components and their effects on early development.Theriogenology 2001; 55:1255–1276.
5. Brevini TAL, Cillo F, Francisci C, Gandolfi F. Polyadenylation of oo-cyte transcripts as marker of developmental competence. In: WatsonAJ (ed.), Mammalian Embryo Genomics. Paris: OECD Publications;2003:7–18.
6. Combelles CM, Albertini DF. Microtubule patterning during meioticmaturation in mouse oocytes is determined by cell cycle-specific sort-ing and redistribution of gamma-tubulin. Dev Biol 2001; 239:281–294.
7. Gard DL, Cha BJ, King E. The organization and animal-vegetal asym-metry of cytokeratin filaments in stage VI Xenopus oocytes is depen-dent upon F-actin and microtubules. Dev Biol 1997; 184:95–114.
8. Van Blerkom J, Runner MN. Mitochondrial reorganization during re-sumption of arrested meiosis in the mouse oocyte. Am J Anat 1984;171:335–355.
9. St John JC. The transmission of mitochondrial DNA following as-sisted reproductive techniques. Theriogenology 2002; 57:109–123.
10. Calarco PG. Polarization of mitochondria in the unfertilized mouseoocyte. Dev Genet 1995; 16:36–43.
11. Stojkovic M, Machado SA, Stojkovic P, Zakhartchenko V, Hutzler P,Goncalves PB, Wolf E. Mitochondrial distribution and adenosine tri-phosphate content of bovine oocytes before and after in vitro matu-ration: correlation with morphological criteria and developmental ca-pacity after in vitro fertilization and culture. Biol Reprod 2001; 64:904–909.
12. Wilding M, Dale B, Marino M, di Matteo L, Alviggi C, Pisaturo ML,Lombardi L, De Placido G. Mitochondrial aggregation patterns andactivity in human oocytes and preimplantation embryos. Hum Reprod2001; 16:909–917.
13. Sun QY, Wu GM, Lai L, Park KW, Cabot R, Cheong HT, Day BN,Prather RS, Schatten H. Translocation of active mitochondria duringpig oocyte maturation, fertilization and early embryo development invitro. Reproduction 2001; 122:155–163.
14. Van Blerkom J. Microtubule mediation of cytoplasmic and nuclearmaturation during the early stages of resumed meiosis in culturedmouse oocytes. Proc Natl Acad Sci U S A 1991; 88:5031–5035.
15. Maro B, Howlett SK, Webb M. Non-spindle microtubule organizingcenters in metaphase II-arrested mouse oocytes. J Cell Biol 1985; 101:1665–1672.
16. Kim NH, Funahashi H, Prather RS, Schatten G, Day BN. Microtubuleand microfilament dynamics in porcine oocytes during meiotic mat-uration. Mol Reprod Dev 1996; 43:248–255.
17. Sun QY, Lai L, Park KW, Kuhholzer B, Prather RS, Schatten H. Dy-namic events are differently mediated by microfilaments, microtu-bules, and mitogen-activated protein kinase during porcine oocytematuration and fertilization in vitro. Biol Reprod 2001; 64:879–889.
18. Lee J, Miyano T, Moor RM. Spindle formation and dynamics of gam-ma-tubulin and nuclear mitotic apparatus protein distribution duringmeiosis in pig and mouse oocytes. Biol Reprod 2000; 62:1184–1192.
19. Ju JC, Tsay C, Ruan CW. Alterations and reversibility in the chro-matin, cytoskeleton and development of pig oocytes treated with ros-covitine. Mol Reprod Dev 2003; 64:482–491.
20. Grupen CG, McIlfatrick SM, Ashman RJ, Boquest AC, ArmstrongDT, Nottle MB. Relationship between donor animal age, follicularfluid steroid content and oocyte developmental competence in the pig.Reprod Fertil Dev 2003; 15:81–87.
21. Brevini TAL, Francisci C, Vassena R, Bagg MA, Grupen CG, Arm-strong DT, Gandolfi F. Follicular fluid concentration during pig IVMaffects oocyte parthenogenetic development and mitochondria distri-bution. Theriogenology 2003; 59:440.
22. Boquest AC, Grupen CG, Harrison SJ, McIlfatrick SM, Ashman RJ,d’Apice AJ, Nottle MB. Production of cloned pigs from cultured fetalfibroblast cells. Biol Reprod 2002; 66:1283–1287.
23. Petters RM, Wells KD. Culture of pig embryos. J Reprod Fertil 1993;48:61–73.
24. Yoshida M, Ishizaki Y, Kawagishi H, Bamba K, Kojima Y. Effects ofpig follicular fluid on maturation of pig oocytes in vitro and on theirsubsequent fertilizing and developmental capacity in vitro. J ReprodFertil 1992; 95:481–488.
25. Daen FP, Sato E, Naito K, Toyoda Y. The effect of pig follicular fluidfractions on cumulus expansion and male pronucleus formation inporcine oocytes matured and fertilized in vitro. J Reprod Fertil 1994;101:667–673.
26. Vatzias G, Hagen DR. Effects of porcine follicular fluid and oviduct-conditioned media on maturation and fertilization of porcine oocytesin vitro. Biol Reprod 1999; 60:42–48.
27. Funahashi H, Day BN. Effects of follicular fluid at fertilization invitro on sperm penetration in pig oocytes. J Reprod Fertil 1993; 99:97–103.
28. Grupen CG, Mau JC, McIlfatrick SM, Maddocks S, Nottle MB. Effectof 6-dimethylaminopurine on electrically activated in vitro maturedporcine oocytes. Mol Reprod Dev 2002; 62:387–396.
29. Loi P, Ledda S, Fulka J, Cappai P, Moor RM. Development of par-thenogenetic and cloned ovine embryos—effect of activation proto-cols. Biol Reprod 1998; 58:1177–1187.
30. Torner H, Brussow KP, Alm H, Ratky J, Pohland R, Tuchscherer A,Kanitz W. Mitochondrial aggregation patterns and activity in porcineoocytes and apoptosis in surrounding cumulus cells depends on thestage of pre-ovulatory maturation. Theriogenology 2004; 61:1675–1689.
31. Van Blerkom J, Davis PW, Lee J. ATP content of human oocytes anddevelopmental potential and outcome after in-vitro fertilization andembryo transfer. Hum Reprod 1995; 10:415–424.
32. Brad AM, Bormann CL, Swain JE, Durkin RE, Johnson AE, CliffordAL, Krisher RL. Glutathione and adenosine triphosphate content ofin vivo and in vitro matured porcine oocytes. Mol Reprod Dev 2003;64:492–498.
33. Bavister BD, Squirrell JM. Mitochondrial distribution and function inoocytes and early embryos. Hum Reprod 2000; 15(Suppl 2):189–198.
34. Sun QY, Lai L, Wu GM, Park KW, Day BN, Prather RS, Schatten H.Microtubule assembly after treatment of pig oocytes with taxol: cor-relation with chromosomes, gamma-tubulin, and MAP kinase. MolReprod Dev 2001; 60:481–490.
35. Rozinek J, Petr J, Grocholova R, Jilek F. Microtubule rearrangementduring in vitro maturation of pig oocytes. Effect of cycloheximide.Reprod Nutr Dev 1995; 35:685–694.
36. Schliwa M, Woehlke G. Molecular motors. Nature 2003; 422:759–765.
37. Cohen RS. Oocyte patterning: dynein and kinesin, inc. Curr Biol2002; 12:R797–R799.
38. Liu L, Hammar K, Smith PJ, Inoue S, Keefe DL. Mitochondrial mod-ulation of calcium signaling at the initiation of development. CellCalcium 2001; 30:423–433.
39. Yang SH, Liu R, Perez EJ, Wen Y, Stevens SM Jr, Valencia T, Brun-Zinkernagel AM, Prokai L, Will Y, Dykens J, Koulen P, Simpkins JW.Mitochondrial localization of estrogen receptor beta. Proc Natl AcadSci U S A 2004; 101:4130–4135.





























