Embed Size (px)
Citation preview

Sleep Research Online 2(3): 57-63, 1999http://www.sro.org/1999/Sakai/xx/ Printed in the USA. All rights reserved.
Correspondence:Kazuya Sakai, Ph.D., INSERM U480, Université Claude Bernard, Lyon 1, 8 Avenue Rockefeller, 69373 Lyon Cedex 08,France, Tel: 33-478-777-122, Fax: 33-478-777-172, E-mail: [email protected].
1096-214X© 1999 WebSciences
Single unit recording studies in freely moving cats havedemonstrated the existence of two distinct populations of state-specific neurons in the brainstem, one, termed PS-on neurons,showing a tonic discharge just prior to and throughoutparadoxical sleep (PS), and the other, PS-off neurons,exhibiting a marked reduction or complete cessation ofdischarge during PS (see Hobson et al., 1986; Sakai, 1988). Itis generally accepted that PS-on neurons are either cholinergicor cholinoceptive, while PS-off neurons are serotonergic,noradrenergic, histaminergic, and possibly adrenergic, but notdopaminergic (Jacobs, 1985; Steriade and McCarley, 1990;Sakai et al., 1990a; Jones, 1991). It has been proposed that PSis generated as a result of tonic excitation of PS-on neurons andinhibition of PS-off neurons (reciprocal interactionhypotheses) (Hobson et al., 1975, 1986; Sakai, 1984, 1988).
We have demonstrated recently the existence of twodifferent groups of PS-on neurons in the dorsal pontinetegmentum (Sakai and Koyama, 1996), one of which ischaracterized by a broad action potential, a slow conductionvelocity, and an inhibitory response to iontophoreticallyapplied carbachol (Carb-I PS-on neurons), and another whichis characterized by a short action potential, a fast conductionvelocity, and an excitatory response to applied carbachol(Carb-E PS-on neurons). Carb-I PS-on neurons are locatedexclusively in the mediodorsal pontine tegmentum which
contains cholinergic neurons, especially in the rostral part ofthe nucleus peri-locus coeruleus alpha (peri-LCalpha), whileCarb-E PS-on neurons are found in both the cholinergic(rostral) and non-cholinergic (caudal) regions of the peri-LCalpha. In light of these findings, we have suggested thecholinergic nature of Carb-I PS-on neurons and non-cholinergic nature of Carb-E PS-on neurons. Here we reportthe existence of presumed non-monoaminergic PS-off neuronsin the medulla. Differing from the classically describedpresumed monoaminergic PS-off neurons, which arecharacterized by a broad action potential, a slow and regulardischarge during quiet waking (QW), a slow conductionvelocity and their location in monoaminergic structures, thepresumed non-monoaminergic PS-off neurons arecharacterized by a short action potential, a high rate ofspontaneous discharge during QW, a fast conduction velocity,their location in non-monoaminergic structures, and theirinsensitivity to serotonergic or adrenergic autoreceptoragonists.
METHODS
Experiments were performed on eight adult cats withchronically implanted electrodes for standard polygraphicrecordings. Single units were recorded through chronically
Using extracellular single unit recording in the medulla of freely moving cats, we have found a population of PS-off ("TypeII") neurons that are distinct from the classically described monoaminergic PS-off ("Type I") neurons. The presumed non-monoaminergic Type II PS-off neurons (n=22) showed a relatively high rate of tonic discharge during both quiet waking andslow-wave sleep (10.4±4.1 and 9.3±3.1 spikes/sec, mean ± S.D., respectively) and a marked overall decrease in discharge rateduring PS (0.3±0.4 spikes/sec). In contrast to the presumed monoaminergic PS-off neurons (n=62), Type II PS-off neuronsshowed short-lasting phasic discharges during PS, often in association with rapid eye movement and PGO wave bursts. TheseType II neurons were all characterized by a short action potential which was significantly different from that of themonoaminergic PS-off neurons described so far. Five out of 22 neurons were identified antidromically by stimulation of theventrolateral reticulospinal tract (vlRST) at the caudal medulla, while 2 of the 22 were identified antidromically by stimulationof the peri-locus coeruleus alpha of the mediodorsal pontine tegmentum. Their mean conduction velocity (7.2±1.9 m/sec) wassignificantly higher than that (0.9±0.3 m/sec) of the presumed monoaminergic PS-off neurons which were identifiedexclusively by stimulation of the vlRST. In addition, when examined during the sleep-waking cycle, the antidromic responsesof Type II PS-off neurons were either completely blocked or reduced, with a prolongation of antidromic latency during PS.Most of these neurons were located in medullary structures containing no, or virtually no, monoaminergic neurons, and noneresponded by inhibition to systemic administration of serotonergic or adrenergic autoreceptor agonists. These findingsindicate the existence, in the medulla, of non-monoaminergic PS-off neurons that would play an important role in PSgeneration.
CURRENT CLAIM:
Are There Non-Monoaminergic Paradoxical Sleep-OffNeurons in the Brainstem?
Kazuya Sakai1 and Norio Kanamori2
1INSERM U480, Département de Médecine Expérimentale, Université Claude Bernard, Lyon 69373, France and2Department of Physiology, Tokushima University School of Dentistry, Tokushima 770, Japan
Non-monoaminergic PS-off neurons exist in the brainstem.
implanted flexible Formvar-coated stainless steel wires (32 µmdiameter) in the unrestrained, freely moving condition. Thesemicroelectrodes were assembled into bundles of 6 wires thatcould be lowered by means of an attached microdriveassembly. The microwires were inserted through the
cerebellum into the medulla at an angle of 72° to the horizontalplane. Recordings of the cortical, hippocampal, and dorsallateral geniculate EEG rhythms, EOG, and neck EMG werealso made using standard techniques. In addition, bipolarstimulation electrodes, consisting of two stainless-steel wires(200 µm diameter, 1.0-1.5 mm apart and bared 0.5 mm at thetip), were implanted stereotaxically into the lateral posteriorhypothalamus (A9.0, L3.0, HC -4.0), peri-locus coeruleusalpha (peri-LCalpha) (P2.0, L2.0, HC-3.5), and ventrolaterallateral reticulospinal tract (vlRST) (P15.0, L2.5, HC-10.0).Stimulation was performed with square pulses (0.05-0.5 ms,0.05-2.5 mA, 0.5-1.0 Hz) below the threshold for movementsusing a WPI 302 stimulator via a stimulus isolation unit. Themain criteria for recognizing antidromic responses were a fixedlatency, a collision test with spontaneous spikes, and the abilityto follow high frequency stimulation (Lipski, 1981).Conduction velocity was estimated from the straight linedistance between recording and stimulating sites and theshortest antidromic latency when the antidromic latency wasshortened in steps with increasing strength of stimulation.
The unit activities were amplified using a conventionalamplifier with low and high cut-off filters of 100 Hz and 10
58 SAKAI AND KANAMORI
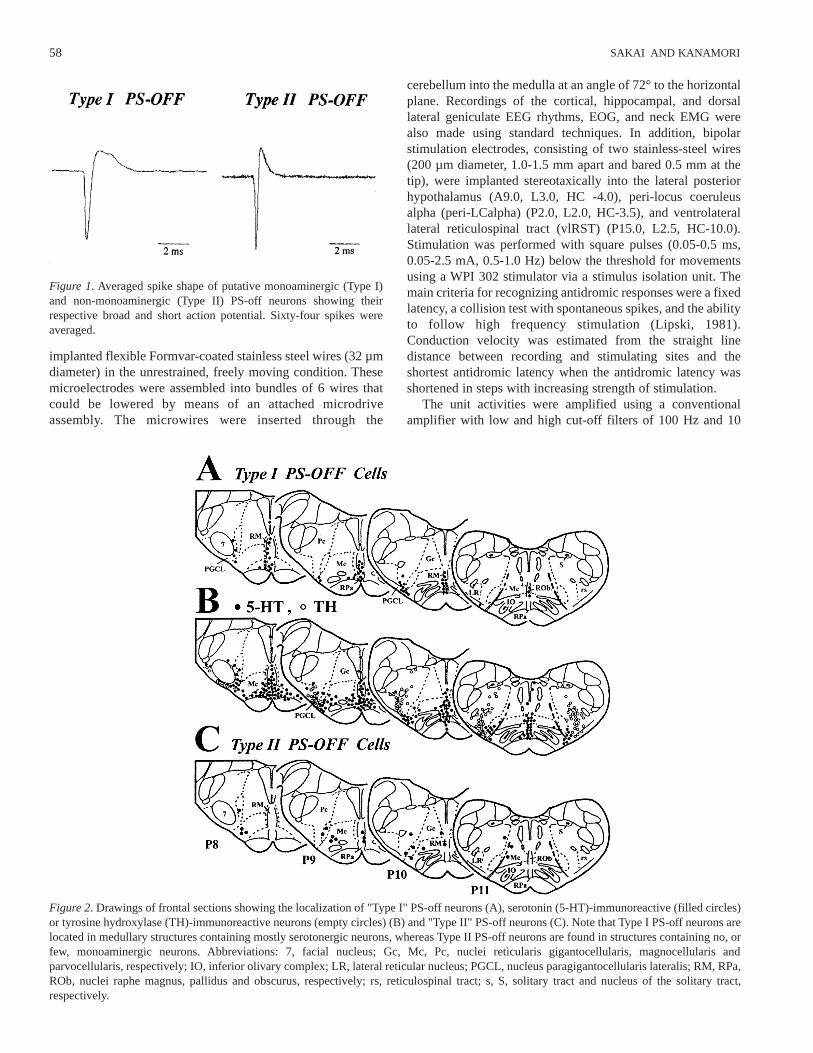
Figure 2. Drawings of frontal sections showing the localization of "Type I" PS-off neurons (A), serotonin (5-HT)-immunoreactive (filled circles)or tyrosine hydroxylase (TH)-immunoreactive neurons (empty circles) (B) and "Type II" PS-off neurons (C). Note that Type I PS-off neurons arelocated in medullary structures containing mostly serotonergic neurons, whereas Type II PS-off neurons are found in structures containing no, orfew, monoaminergic neurons. Abbreviations: 7, facial nucleus; Gc, Mc, Pc, nuclei reticularis gigantocellularis, magnocellularis andparvocellularis, respectively; IO, inferior olivary complex; LR, lateral reticular nucleus; PGCL, nucleus paragigantocellularis lateralis; RM, RPa,ROb, nuclei raphe magnus, pallidus and obscurus, respectively; rs, reticulospinal tract; s, S, solitary tract and nucleus of the solitary tract,respectively.
Figure 1.Averaged spike shape of putative monoaminergic (Type I)and non-monoaminergic (Type II) PS-off neurons showing theirrespective broad and short action potential. Sixty-four spikes wereaveraged.
kHz, respectively. The action potentials were displayed on adigital memory oscilloscope equipped with a processor forspike waveform averaging, and 64 or 128 action potentialswere averaged for each neuron to determine the spike shape.Analysis of unitary activity, such as discharge rate, interspikeand post-stimulus time histograms, and auto- and cross-correlograms, was performed using the CED 1401 dataprocessor and Spike 2 software. The mean firing rates werecalculated from 100-sec recordings using 5- or 10-sec bins.Statistical analysis was carried out using Student's t-test oranalysis of variance (ANOVA). The drugs used, 5-methoxy-N,N-dimethyltryptamine (5-MeODMT) and clonidinehydrochloride (both from Sigma), were dissolved inphysiological saline immediately before use and injectedsystemically (i.m.) in a volume of 0.2-0.4 ml. 5-MeODMT wasused rather than 8-OH-DPAT, a selective 5-HT1A agonist,since our preliminary study showed that 8-OH-DPAT inducedmarked behavioral agitation in the animals when administeredat the same dose and by the same route.
At the end of the experiments, the location of the end andseveral passage points of the track of the microelectrodes wereexamined histologically using the Prussian blue reactiontechnique on Nissl-stained sections. The immunohistochemicalprocedures used for the localization of serotonin (5-HT),tyrosine hydroxylase (TH), phenylethanolamine N-methyltransferase (PNMT) or choline acetyltransferase(ChAT) to identify serotonergic, catecholaminergic, adrenergicand cholinergic neurons have previously been described indetail (Sakai et al., 1990b).
RESULTS
The results presented here were obtained from 84 PS-offneurons recorded in the rostral medulla. PS-off neurons weredefined as those showing the highest rate of tonic spontaneousdischarge during waking (W), a reduction of discharge rate
during slow-wave sleep (SWS), and a further reduction, oreven complete cessation, of discharge during paradoxical sleep(PS).
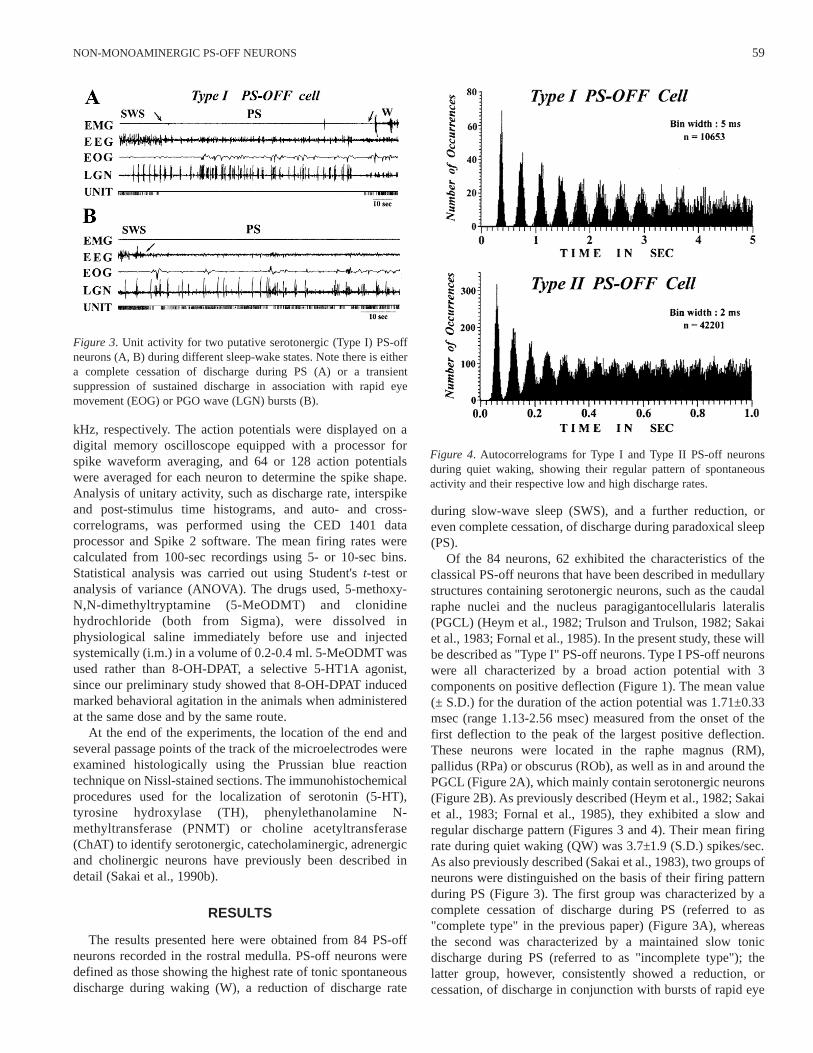
Of the 84 neurons, 62 exhibited the characteristics of theclassical PS-off neurons that have been described in medullarystructures containing serotonergic neurons, such as the caudalraphe nuclei and the nucleus paragigantocellularis lateralis(PGCL) (Heym et al., 1982; Trulson and Trulson, 1982; Sakaiet al., 1983; Fornal et al., 1985). In the present study, these willbe described as "Type I" PS-off neurons. Type I PS-off neuronswere all characterized by a broad action potential with 3components on positive deflection (Figure 1). The mean value(± S.D.) for the duration of the action potential was 1.71±0.33msec (range 1.13-2.56 msec) measured from the onset of thefirst deflection to the peak of the largest positive deflection.These neurons were located in the raphe magnus (RM),pallidus (RPa) or obscurus (ROb), as well as in and around thePGCL (Figure 2A), which mainly contain serotonergic neurons(Figure 2B). As previously described (Heym et al., 1982; Sakaiet al., 1983; Fornal et al., 1985), they exhibited a slow andregular discharge pattern (Figures 3 and 4). Their mean firingrate during quiet waking (QW) was 3.7±1.9 (S.D.) spikes/sec.As also previously described (Sakai et al., 1983), two groups ofneurons were distinguished on the basis of their firing patternduring PS (Figure 3). The first group was characterized by acomplete cessation of discharge during PS (referred to as"complete type" in the previous paper) (Figure 3A), whereasthe second was characterized by a maintained slow tonicdischarge during PS (referred to as "incomplete type"); thelatter group, however, consistently showed a reduction, orcessation, of discharge in conjunction with bursts of rapid eye
59NON-MONOAMINERGIC PS-OFF NEURONS
Figure 3. Unit activity for two putative serotonergic (Type I) PS-offneurons (A, B) during different sleep-wake states. Note there is eithera complete cessation of discharge during PS (A) or a transientsuppression of sustained discharge in association with rapid eyemovement (EOG) or PGO wave (LGN) bursts (B).
Figure 4. Autocorrelograms for Type I and Type II PS-off neuronsduring quiet waking, showing their regular pattern of spontaneousactivity and their respective low and high discharge rates.

movement (REM) and PGO waves occurring during PSepisodes (84% mean percent reduction in firing rate relative toQW) (Figure 3B), compared with a 62% mean percentreduction in firing rate in PS without REM and PGO wavebursts relative to QW. When examined using systemicadministration of 5-MeODMT (250 µg/kg, i.m.), all neuronsresponded to the serotonergic autoreceptor agonist with asignificant reduction (n=11) (Figure 5A), or cessation (n=10)(Figure 5C-1), of discharge, as previously reported by Jacobset al. (1983). Seven out of 62 Type I PS-off neurons wereidentified antidromically by stimulation of the ventrolateralreticulospinal tract (vlRST) at the caudal medulla. Their meanconduction velocity was 0.9±0.3 (S.D.) m/sec (range: 0.5-1.2m/sec). None of the 62 responded antidromically to stimuliapplied to the peri-locus coeruleus alpha (peri-LCalpha) of themediodorsal pontine tegmentum or posterior hypothalamus.
In contrast to the Type I PS-off neurons described above, 22of the 88 PS-off neurons, which we will designate as "Type IIPS-off neurons", were characterized by a short action potential,a fast conduction velocity, their location outsidemonoaminergic structures, short phasic discharges with REMand PGO wave bursts, and a lack of inhibitory response to 5-MeODMT or clonidine, an alpha2 adrenergic agonist. Asshown in Figure 1, the mean duration of the action potentialwas 0.60±0.14 msec (range 0.37-0.88 msec), distinct from, andsignificantly shorter (p<0.001, two-tailed t-test) than, that of
Type I PS-off neurons. Although some neurons were found inthe raphe nuclei, the majority were located in structures thatcontain no, or virtually no, monoaminergic neurons, e.g., thenuclei reticularis magnocellularis (Mc), gigantocellularis (Gc),and parvocellularis (Pc) (Figure 2C). As shown in Figures 4-6,they displayed high rates of tonic discharges as compared toType I PS-off neurons, e.g., their mean discharge rate duringQW was 10.4±4.1 spikes/sec, significantly different (p>0.001,two-tailed t-test) from the 3.7±1.9 spikes/sec seen in Type I PS-off neurons, while those during SWS without PGO waves, PSwithout REM and PGO wave bursts, and PS with REM andPGO wave bursts were 9.3±3.8, 0.3±0.4, and 4.4±3.9spikes/sec, respectively. Although Type II PS-off neuronsshowed significant phasic changes in discharge rates duringactive waking (AW) in relation to body movements, theydisplayed a regular discharge pattern during both QW andSWS, as shown in Figure 4. In sharp contrast to Type I PS-offneurons, Type II PS-off neurons exhibited short-lasting phasicfiring during PS, often in association with REM and PGO wavebursts (Figure 6); no specific correlation with phasic motoractivities, such as eye or pinnae movement, was observed. Asillustrated in Figure 5B and 5C-2, Type II PS-off neurons didnot respond with reduction of discharge to either 5-MeODMT(250 µg /kg, i.m., n=6) or clonidine (25 µg /kg, i.m., n=1; datanot shown).
Seven of the 22 Type II PS-off neurons were identifiedantidromically by stimulation of either the vlRST (n=5) or theperi-LCalpha (n=2). Their mean conduction velocity was
60 SAKAI AND KANAMORI
Figure 5. Effects of systemic administration of 5-methoxy-N,N-dimethyltryptamine (5-MeODMT) on an "incomplete" Type I (A), a"complete" Type I (C1) and two Type II (B, C2) PS-off neurons. Notethat the Type I PS-off cells are inhibited by 5-MeODMT, whereas theType II PS-off cells exhibit no inhibitory response to the serotonergicautoreceptor agonist. Note also that the Type I and Type II PS-off cellswere recorded simultaneously in C. Arrows indicate 5-MeODMTinjection points.
Figure 6. Discharge pattern of a Type II PS-off cell during the sleep-waking cycle. Note suppression of spontaneous tonic discharge duringparadoxical sleep (PS) and the occurrence of phasic spontaneousactivity, often in conjunction with rapid eye movement (EOG) andPGO wave (LGN) bursts.

7.3±1.9 m/sec (range 4.6-10.0 m/sec), significantly different(p<0.001, two-tailed t-test) from that of 0.9±0.3 m/sec seen forType I PS-off neurons. Although it was unsuccessful for TypeI PS-off neurons, the antidromic responses of two Type IIneurons could be examined in detail during sleep-wake statesusing the same stimulus intensity. As shown in Figure 7, onecell showed faithful antidromic responses during both QW andSWS, but, during PS displayed a complete blockage ofantidromic responses of both initial segment (IS) and somato-dendritic (SD) spikes, IS-SD break or a blockage of the SDspike alone (Figure 7B). The other cell exhibited a greater than10% decrease in antidromic responsiveness during PS tostimulation of the vlRST and displayed prolongation ofantidromic latencies from 0.80 msec in SWS to 0.90 msec inPS (data not shown), suggesting inhibitory synaptic volleysimpinging on non-monoaminergic PS-off neurons during PS(see Lipski, 1981).
DISCUSSION
The present findings strongly suggest the existence of non-monoaminergic PS-off neurons in the medulla, which are
characterized by a short action potential, a fast conductionvelocity, a high rate of spontaneous discharge during both Wand SWS, unresponsiveness to serotonergic or adrenergicautoreceptor agonists, and their location essentially in non-monoaminergic structures. Until now, all PS-off neuronsshowing PS-selective discharge cessation have been regardedas monoaminergic, usually serotonergic, noradrenergic orhistaminergic, being reported in the raphe nuclei andventrolateral medulla, containing serotonergic neurons(McGinty et al., 1973; Heym et al., 1982; Sakai et al., 1983;Rasmussen et al., 1984; Fornal et al., 1985), the dorsolateralpontine tegmentum, containing noradrenergic neurons(Hobson et al., 1973; McGinty and Sakai, 1973; Saito et al.,1977; Sakai, 1980; 1991; Aston-Jones and Bloom, 1981;Reiner, 1986), and in the posterior hypothalamus, containinghistaminergic neurons (Vanni-Mercier et al., 1984; Sakai et al.,1990a). Features common to these putative monoaminergicneurons are their long duration of action potential and a slowand fairly regular pattern of spontaneous activity during QW(see Jacobs, 1985; Steriade and McCarley, 1990; Sakai et al.,1990; Jacobs and Azmitia, 1992 for review). Unlike thesepresumed monoaminergic PS-off neurons, the presumed non-monoaminergic Type II PS-off neurons all displayed a shortaction potential and a higher rate of spontaneous dischargeduring QW. Type II PS-off neurons appear to be non-catecholaminergic, since they are not located in medullaryregions containing tyrosine hydroxylase (TH)-immunoreactivenoradrenergic or adrenergic neurons (Figure 2). In addition,they have higher conduction velocities (7.3±1.9 m/sec) thanputative adrenergic C1 neurons (0.9±0.1 m/sec; mean ±S.E.M.) (Haselton and Guyenet, 1989), which is in line withthe demonstration that most C1 neurons give rise tounmyelinated axons (Milner et al., 1987). Although theclonidine unresponsiveness found in the one Type II PS-off celltested is also in favor of the assumption of the non-adrenergicnature of Type II PS-off cells, further pharmacologicalexamination of a large sample of cells is needed for this issue.The activity of both adrenergic and noradrenergic medullaryneurons during the sleep-waking cycle is not yet known andwill be an important subject for future studies. Type II PS-offcells also appear to be non-serotonergic, since they weremainly located in medullary regions containing no, or few,serotonergic neurons; the present findings on their fastconduction velocity and insensitivity to the systemicadministration of 5-MeODMT, a serotonergic autoreceptoragonist, further strengthen this assumption. A basic questiontherefore arises as to the neurochemical nature and functionalroles of these putative non-monoaminergic Type II PS-offneurons.
Although the firing pattern of Type II PS-off neurons wasvariable during AW, they showed a relatively high rate (about10 Hz) of regular discharge pattern during QW and SWS(Figure 4), during which multiple phasic synaptic inputsshould be considerably reduced. The majority of Type II PS-offneurons were found in the rostral and ventrolateral medullaryregion. In this regard, it is worth mentioning the presence ofnon-monoaminergic neurons showing a "pacemaker-like"discharge pattern that have been reported in the rat rostral
61NON-MONOAMINERGIC PS-OFF NEURONS
Figure 7. Antidromic responses of a Type II PS-off cell followingstimulation of the peri-locus coeruleus alpha (Peri-LCalpha) of themediodorsal pontine tegmentum during waking (A) and sleep (B).Their fixed latency, collision with a spontaneous spike(s), and abilityto follow high frequency stimulation are shown, respectively, in A1,A2 and A3. In A4, the cell displays a blockade of the somatodendriticspike (indicated by an arrow) at a high rate of repetitive stimulation.Note also that the cell responds faithfully to each stimulus during bothquiet waking and slow-wave sleep (SWS) (B1), whereas duringparadoxical sleep (PS), the same stimulus results in completeblockade of antidromic responses of both the initial segment andsomatodendritic spikes (B2), a break of IS and SD spikes (B3,indicated by an arrow), or a blockade of the SD spike (B4, indicatedby an arrow).

ventrolateral medulla, both in vitro (Sun et al., 1988a) and invivo, particularly in the presence of a glutamate receptorantagonist (Sun et al., 1988b; see also Granata and Kitai,1992). These neurons have been supposed to be glutamatergicin nature and sympathoexcitatory in function via directprojections to the spinal cord (Sun et al., 1988a,b). Their firingrate (9 spikes/sec in vitro; Sun et al., 1988a) and regularactivity, as well as their projection to the spinal cord arereminiscent of the characteristics of descending Type II PS-offneurons, as described in the present study. Further studies areneeded to determine whether these neurons are implicated inchanges in autonomic events during the sleep-waking cycle.
Although once supposed to be glutamatergic, the exactneurochemical identity of putative sympathoexcitatorybulbospinal neurons is not yet known. Histochemical studies ofthe rostral ventrolateral medulla have demonstrated thepresence of many putative neurotransmitters, includingbiogenic amines, acetylcholine, neuropeptides and bothexcitatory and inhibitory amino acids, within the cell bodies(see Chalmers and Pilowsky, 1991 for review). Recently, theexistence in the rostral ventrolateral medulla of GABAergicpacemaker neurons having a sympathoinhibitory function hasbeen suggested (Hayer et al., 1996). It is especially worthspeculating about the GABAergic neurochemical identity ofType II PS-off neurons when we consider their roles in themechanisms underlying PS generation. In previous reports ofstudies in freely moving cats, PS-on neurons showing a highrate of tonic discharge just prior to and throughout PS weredescribed in the nuclei reticularis magnocellularis (Mc),parvocellularis (Pc) and paragigantocellularis lateralis (PGCL)(Sakai et al., 1979; Kanamori et al., 1980; Sakai, 1988), areasin which Type II PS-off neurons were recorded in the presentstudy. It seems likely that Type II PS-off neurons exert directinhibition on medullary PS-on neurons through a local circuit.The present finding on the ascending projection of some TypeII PS-off neurons to the peri-LCalpha, which also contains PS-on neurons (Sakai, 1980; 1988), allows further speculation ona direct inhibition of pontine PS-on neurons via medullaryGABAergic PS-on neurons, an idea suggested by the facts thationtophoretically applied bicuculline, a GABAA receptorantagonist, produces excitation of PS-on neurons andsubsequently induces PS (Sakai and Koyama, 1996) and that,conversely, microdialysis application of muscimol (0.5-1.0mM), a potent GABAA receptor agonist, to the peri-LCalpharesults in inhibition of PS (Sakai, Onoe and Crochet,unpublished data). Although GABAergic neurons are widelydistributed in the brainstem, the existence in the brain of non-monoaminergic PS-off neurons having a PS-inhibitoryfunction has not been reported until the present study whichshowed their presence in the medulla.
Both pontine and medullary PS-on neurons dischargetonically and selectively just prior to, and throughout, periodsof PS, satisfying the selectivity, tonicity, and tonic PS-latencycriteria necessary for being PS-generator neurons (Sakai,1988). There is a mirror image, or exactly inverse relationshipin terms of cellular discharge between PS-on and putativemonoaminergic PS-off neurons that cease firing selectivelyduring PS, suggesting a mutual "inhibitory" interaction
between the two distinct neuronal populations (Sakai, 1984;1988). In keeping with the mutual inhibitory interactionhypothesis, presumed serotonergic medullary PS-off neuronscease firing during PS, especially during PS episodes markedby the presence of REM and PGO wave bursts (Sakai et al.,1983; and the present study), in which PS-on neurons exhibitan increase in tonic discharge rate (Sakai, 1984; 1988).However, during PS, Type II PS-off neurons exhibited phasicdischarges in association with the bursts of REMs and PGOwaves accompanying an increase in activity of PS-on neurons,suggesting a possible excitatory action of PS-on neurons onnon-monoaminergic PS-off neurons, and thereby supporting,in part, the reciprocal "excitatory-inhibitory" interactionhypothesis between REM-on and REM-off cells, as originallyproposed by Hobson and McCarley between cholinergic REM-on and monoaminergic REM-off cells (Hobson et al., 1975;1986).
In conclusion, the present study demonstrates the existenceof non-monoaminergic PS-off neurons in the medulla and thusopens up the possibility of multiple interactions between PS-onand monoaminergic and non-monoaminergic PS-off neurons.
ACKNOWLEDGMENTS
This work was supported by INSERM U480.
REFERENCES
1. Aston-Jones G, Bloom FE. Activity of norepinephrine-containing locus coeruleus neurons in behaving ratsanticipates fluctuations in the sleep-waking cycle. JNeurosci 1981; 1: 876-86.
2. Chalmers J, Pilowsky P. Brainstem and bulbospinalneurotransmitter systems in the control of blood pressure. JHypertension 1991; 9: 675-94.
3. Fornal C, Auerbach S, Jacobs BL. Activity of serotonin-containing neurons in nucleus raphe magnus in freelymoving cats. Exp Neurol 1985; 88: 590-608.
4. Granata AR, Kitai ST. Intracellular analysis in vivo ofdifferent barosensitive bulbospinal neurons in the rat rostralventrolateral medulla. J Neurosci 1992; 12: 1-20.
5. Haselton JR, Guyenet PG. Electrophysiologicalcharacterization of putative C1 adrenergic neurons in therat. Neuroscience 1989; 30: 199-214.
6. Hayer A, Piguet P, Feltz P. GABA-induced responses in electro-physiologically characterized neurons within the rat rostro-ventrolateral medulla in vitro. Brain Res 1996; 709: 173-83.
7. Heym J, Steinfels GF, Jacobs BL. Activity of serotonin-containing neurons in the nucleus raphe pallidus of freelymoving cats. Brain Research 1982; 251: 259-76.
8. Hobson JA, McCarley RW, Wyzinsky PW, Pivik RT.Reciprocal tonic firing by FTG and LC neurons during thesleep-waking cycle. Sleep Res 1973; 2: 29.
9. Hobson JA, McCarley RW, Wyzinsky PW. Sleep cycleoscillation: reciprocal discharge by two brainstem neurons.Science 1975; 189: 55-8.
10. Hobson JA, Lydic R, Baghdoyan HA. Evolving concepts of sleepcycle generations. Behav Brain Science 1986; 9: 371-448.
11. Jacobs BL, Heym J, Rasmussen K. Raphe neurons: firing ratecorrelates with size of drug response. Eur J Pharmacol1983; 90: 275-8.
62 SAKAI AND KANAMORI

12. Jacobs BL. Overview of the activity of brain monoaminergicneurons across the sleep-wake cycle. In: Wauquier A,Gaillard V, Monti JM, Radulovacki M, eds. Sleep:Neurotransmitters and Neuromodulators. New York: RavenPress, 1985, pp. 1-14.
13. Jacobs BL, Azmitia EC. The structure and function of thebrain serotonin system. Physiol Rev 1992; 72: 165-229.
14. Jones BE. Paradoxical sleep and its chemical/structuralsubstrates in the brain. Neuroscience 1991; 40: 637-56.
15. Kanamori N, Sakai K, Jouvet M. Neuronal activity specific toparadoxical sleep in the ventromedial medullary reticularformation of unrestrained cats. Brain Res 1980; 189: 251-5.
16. Lipski J. Antidromic activation of neurons as an analytic toolin the study of the central nervous system. J Neurosci Meth1981; 4: 1-32.
17. McGinty DJ, Harper RM, Fairbanks MK. 5HT-containingneurons: Unit activity in behaving cats. In: Barchas J, UsdinE, eds. Serotonin and Behavior. New York: Academic Press,1973, pp. 267-79.
18. McGinty DJ, Sakai K. Unit activity of the dorsal pontinereticular formation in the cat. Sleep Res 1973; 2: 33.
19. Milner TA, Pickel VM, Park DH, Joh TH, Reis DJ.Phenylethanolamine N-methyltransferase-containingneurons in the rostral ventrolateral medulla of the rat. I.Normal ultrastructure. Brain Res 1987; 411: 28-45.
20. Rasmussen K, Heym J, Jacobs BL. Activity of serotonin-containing neurons in nucleus centralis superior of freelymoving cats. Exp Neurol 1984; 83: 302-17.
21. Reiner PB. Correlation analysis of central noradrenergicneurons in unanesthetized cats. Brain Res 1986; 378: 86-96.
22. Saito H, Sakai K, Jouvet M. Discharge patterns of the nucleusparabrachialis lateralis neurons of the cat during sleep andwaking. Brain Res 1977; 134: 59-72.
23. Sakai K, Kanamori N, Jouvet M. Activités unitairesspécifiques du sommeil paradoxal dans la formationréticulée bulbaire chez le chat non-restreint. CR Acad Sci(Paris) 1979; 289: 557-61.
24. Sakai K. Some anatomical and physiological properties ofponto-mesencephalic tegmental neurons with specialreference to the PGO waves and postural atonia duringparadoxical sleep in the cat. In: Hobson JA, Brazier MB,
eds. The Reticular Formation Revisited. New York: RavenPress, 1980, pp. 427-47.
25. Sakai K, Vanni-Mercier G, Jouvet M. Evidence for the presenceof PS-off neurons in the ventromedial medulla oblongata offreely moving cats. Exp Brain Res 1983; 49: 311-4.
26. Sakai K. Central mechanisms of paradoxical sleep. Exp BrainRes, supp 1984; 8: 3-18.
27. Sakai K. Executive mechanisms of paradoxical sleep. ArchItal Biol 1988; 479: 225-40.
28. Sakai K, El Mansari M, Lin JS, Zhang JG, Vanni-Mercier G.Posterior hypothalamus in the regulation of wakefulness andparadoxical sleep. In: Mancia M, Marini G, eds. The Diencephalonand Sleep. New York: Raven Press, 1990a, pp. 171-98.
29. Sakai K, Yoshimoto Y, Luppi PH, Fort P, El Mansari M,Salvert D, Jouvet M. Lower brainstem afferents to the catposterior hypothalamus: a double-labeling study. Brain ResBull 1990b; 24: 437-55.
30. Sakai K. Physiological properties and afferent connections ofthe locus coeruleus and adjacent tegmental neuronsinvolved in the generation of paradoxical sleep in the cat.In: Barnes CD, Pompeiano O, eds. Progress in BrainResearch. Amsterdam: Elsevier, 1991, Vol. 88, pp. 31-45.
31. Sakai K, Koyama Y. Are there cholinergic and non-cholinergic paradoxical sleep-on neurons in the pons?NeuroReport 1996; 7: 2449-53.
32. Steriade M, McCarley RW. Brainstem Control of Wakefulnessand Sleep. New York: Plenum Press, 1990.
33. Sun MK, Young BS, Hackett JT, Guyenet PG. Reticulospinalpacemaker neurons of the rat rostral ventrolateral medullawith putative sympathoexcitatory function: an intracellularstudy in vitro. Brain Res 1988a; 442: 229-39.
34. Sun MK, Hackett JT, Guyenet PG. Sympathoexcitatoryneurons of rostral ventrolateral medulla exhibit pacemakerproperties in the presence of a glutamate-receptorantagonist. Brain Res 1988b; 438: 23-40.
35. Trulson ME, Trulson VM. Activity of nucleus raphe pallidusneurons across the sleep-waking cycle in freely movingcats. Brain Res 1982; 237: 232-7.
36. Vanni-Mercier G, Sakai K, Jouvet M. 'Waking-state specific'neurons in the caudal hypothalamus of the cat. C R Acad Sci1984; 298: 195-200.
63NON-MONOAMINERGIC PS-OFF NEURONS





























