Embed Size (px)
Citation preview

Mar. Drugs 2015, 13, 788-805; doi:10.3390/md13020788
marine drugs ISSN 1660-3397
www.mdpi.com/journal/marinedrugs
Article
Seaweed Supplements Normalise Metabolic, Cardiovascular and
Liver Responses in High-Carbohydrate, High-Fat Fed Rats
Senthil Arun Kumar 1, Marie Magnusson 2, Leigh C. Ward 3, Nicholas A. Paul 2 and
Lindsay Brown 4,*
1 Centre for Systems Biology, University of Southern Queensland, Toowoomba 4350, Australia;
E-Mail: [email protected] 2 MACRO—the Centre for Macroalgal Resources and Biotechnology, and College of Marine &
Environmental Sciences, James Cook University, Townsville, QLD 4811, Australia;
E-Mails: [email protected] (M.M.); [email protected] (N.A.P.) 3 School of Chemistry and Molecular Biosciences, The University of Queensland, Brisbane 4072,
Australia; E-Mail: [email protected] 4 School of Health and Wellbeing, The University of Southern Queensland,
Toowoomba 4350, Australia
* Author to whom correspondence should be addressed; E-Mail: [email protected];
Tel.: +61-7-4631-1319.
Academic Editor: Constantina Nasopoulou
Received: 2 October 2014 / Accepted: 21 January 2015 / Published: 2 February 2015
Abstract: Increased seaweed consumption may be linked to the lower incidence of
metabolic syndrome in eastern Asia. This study investigated the responses to two tropical
green seaweeds, Ulva ohnoi (UO) and Derbesia tenuissima (DT), in a rat model of human
metabolic syndrome. Male Wistar rats (330–340 g) were fed either a corn starch-rich diet or
a high-carbohydrate, high-fat diet with 25% fructose in drinking water, for 16 weeks.
High-carbohydrate, high-fat diet-fed rats showed the signs of metabolic syndrome leading
to abdominal obesity, cardiovascular remodelling and non-alcoholic fatty liver disease. Food
was supplemented with 5% dried UO or DT for the final 8 weeks only. UO lowered total
final body fat mass by 24%, systolic blood pressure by 29 mmHg, and improved glucose
utilisation and insulin sensitivity. In contrast, DT did not change total body fat mass but
decreased plasma triglycerides by 38% and total cholesterol by 17%. UO contained 18.1%
soluble fibre as part of 40.9% total fibre, and increased magnesium, while DT contained
23.4% total fibre, essentially as insoluble fibre. UO was more effective in reducing metabolic
syndrome than DT, possibly due to the increased intake of soluble fibre and magnesium.
OPEN ACCESS

Mar. Drugs 2015, 13 789
Keywords: obesity; hypertension; fatty liver; tropical seaweeds; soluble fibre
1. Introduction
Obesity, defined as excess body fat, is a major health-care problem that increases the risk of
cardiovascular and metabolic disorders such as hypertension, ischaemic stroke, insulin resistance, impaired
glucose tolerance, hyperinsulinaemia and dyslipidaemia [1,2]. Current treatment of obesity is aimed at
modifying dietary habits, lowering calorie and fat intake, and increasing exercise to increase calorie
expenditure [3,4], rather than drugs. Long-term drug treatment of obesity shows modest effects in many
patients, and there are potential health hazards when drug therapies are combined for obesity management [5].
Seaweeds are considered as part of a healthy diet, especially in Japan, Korea, China and the
Philippines [6,7]. Seaweeds possess anti-diabetic, antioxidant, anti-obesity, anti-hyperlipidaemic and
anti-inflammatory activities [8]. Seaweeds contain higher potassium, magnesium and calcium ion
concentrations than other foods [7]. Seaweeds may prevent diet-induced cardiovascular disease as an
alternative source of dietary fibre [9]. Fibre is the largest component of the seaweed biomass [10,11] and
therefore may be present in sufficient amounts when included in the diet to prevent metabolic syndrome
associated with obesity, type 2 diabetes and cardiovascular complications [12]. Seaweeds are typically
low in fat (<5%), with the omega-3 fatty acids as the major fat component, including α- and γ-linolenic
acids, eicosapentaenoic acid and docosahexaenoic acid [13].
The combination of fibre as polysaccharides, with antioxidants, omega-3 fatty acids and minerals
indicates that seaweeds could be targeted as functional foods for metabolic syndrome in westernised
diets [6,8]. Ulva species showed cholesterol-lowering and cardioprotective properties, as well as
anti-inflammatory potential [14,15]. Supplementation of a high-calorie diet with the edible green
seaweed Ulva linza lowered intra-abdominal fat pads by 35%, blood pressure by 35%, concentrations of
blood glucose by 31%, and serum cholesterol and triglycerides concentrations by 17% and 20% [16].
The fast growth rate and broad geographical distribution of tropical species of Ulva, such as Ulva ohnoi
(UO), could provide a sustainable source of these seaweeds for new applications as functional foods as
they are suitable for intensive aquaculture production [17]. Another tropical green seaweed with potential
for commercialisation is Derbesia tenuissima (DT), which has been targeted for its nutritional attributes,
especially a higher content of polyunsaturated fatty acids [18]. In contrast to Ulva, there is little evidence
for Derbesia species as a functional food in metabolic syndrome, although methanolic extracts of
Derbesia species showed high inhibitory activity in vitro against protein tyrosine phosphatase 1B [19],
a negative regulator of insulin receptors associated with signal transduction.
This study measured the responses to two tropical green seaweeds, Ulva ohnoi and Derbesia
tenuissima, on metabolic parameters and on the structure and function of the cardiovascular system and
liver in rats fed a high-carbohydrate, high-fat diet. These rats showed symptoms of metabolic syndrome
with metabolic abnormalities, cardiovascular remodelling, and non-alcoholic steatohepatitis [20]. Seaweed
supplementation was given for the final 8 weeks of a 16 week protocol. The structure and function of
the heart were characterised by echocardiography, isolated Langendorff heart preparation and
histopathology, while the structure and function of the liver were characterised by histopathology and

Mar. Drugs 2015, 13 790
plasma biochemical analyses. In addition, metabolic function was characterised by fat measurements, and
glucose and insulin tolerance tests. We show that tropical green seaweeds, in particular UO, improve
cardiovascular, liver and metabolic parameters in this rat model of human metabolic syndrome.
2. Results
2.1. Nutritional Composition of Seaweeds
The seaweed supplements differed in their concentrations of soluble and insoluble fibre, fatty acids
and minerals. The total dietary fibre content of UO was 40.9% of dry algae with 18.1% as soluble fibre
and 22.8% as insoluble fibre while DT contained 23.4% of dry algae, and this was only insoluble fibre.
DT had higher fatty acid content (4.9% of dry algae) than UO (1.2% of dry algae), with higher omega-3
fatty acid content (38.8% of total fatty acids) and omega-6 fatty acid content (12.7% of total fatty acids).
Mineral contents of UO and DT are given in Supplementary Table S1. The magnesium intake of high-
carbohydrate, high-fat + Ulva ohnoi 5% (HUO) rats (59.5 mg/day) was higher than high-fat + Derbesia
tenuissima 5% (HDT) (29.3 mg/day) and H (15.1 mg/day) rats (Supplementary Table S2). Neither UO
nor DT altered the total fatty acids intake in corn starch + Ulva ohnoi 5% (CUO) and corn starch +
Derbesia tenuissima 5% (CDT) compared to C rats, nor in HUO and HDT compared to H rats
(Supplementary Table S3). The intake of α-linolenic acid was higher in CDT (28.9 mg/day) than in HDT
(16.6 mg/day) treated rats. CDT rats had higher eicosapentaenoic acid intake (1.9 mg/day) than HDT
rats (1.3 mg/day).
2.2. Metabolic Variables
Consumption of food and water was higher in C rats than H rats (Table 1). Compared with C groups
(C and CUO), increased energy intake occurred with no change in body weight gain and energy
efficiency in H and HUO rats (Table 1). An increased body weight gain was observed in both CDT and
HDT rats compared to C rats, while only HDT rats had an increased energy intake (Table 1).
Metabolic parameters are given in Table 2. Compared with C rats, H rats had lower lean mass with
higher total body fat mass, abdominal fat mass, abdominal circumference and visceral adiposity index.
HUO but not HDT lowered total body fat mass. The glucose utilisation and insulin sensitivity measured
as AUC were improved in HUO and HDT rats, compared with H rats. The bone mineral content was
higher in H rats, compared with C rats with no effect of either seaweed treatment. Increased plasma
concentrations of NEFA, triglycerides and total cholesterol were observed in H rats, whereas no changes
were observed in C groups. Plasma NEFA concentrations were increased in HUO rats. DT treatment did
not change the plasma NEFA concentration but normalised plasma triglycerides and total cholesterol
concentrations in HDT rats. No changes were observed in plasma sodium, potassium and magnesium
ion concentrations in the treatment groups.
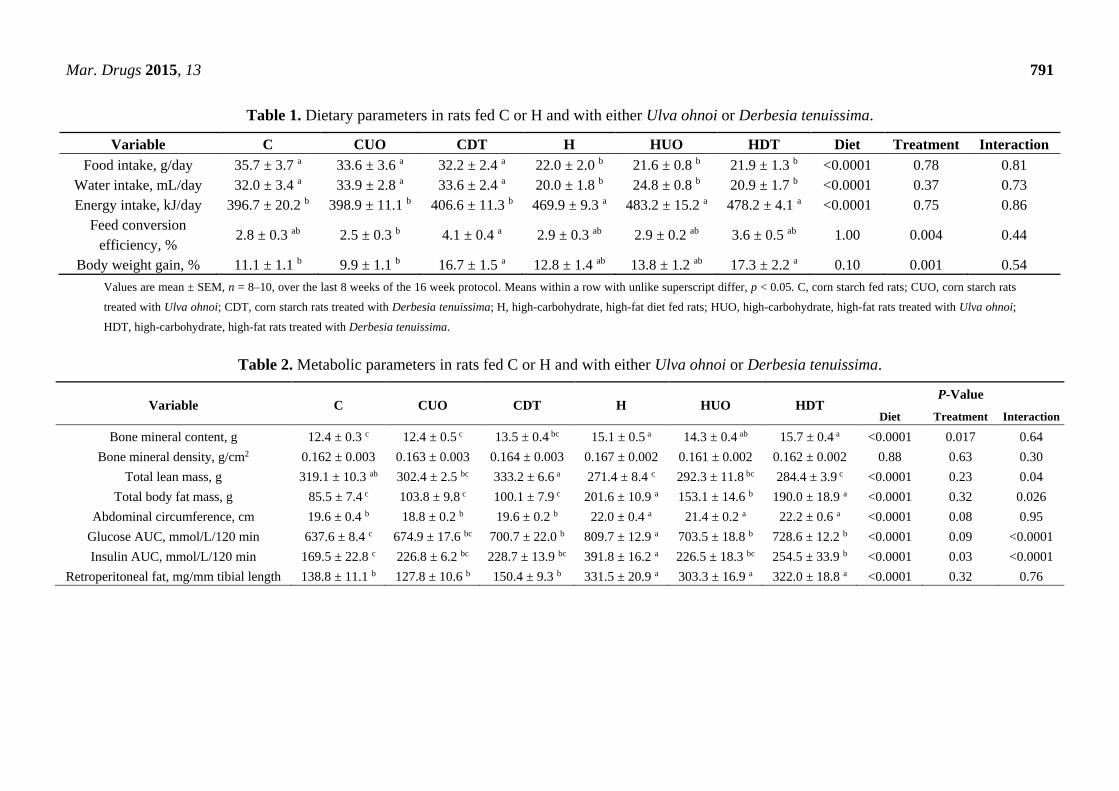
Mar. Drugs 2015, 13 791
Table 1. Dietary parameters in rats fed C or H and with either Ulva ohnoi or Derbesia tenuissima.
Variable C CUO CDT H HUO HDT Diet Treatment Interaction
Food intake, g/day 35.7 ± 3.7 a 33.6 ± 3.6 a 32.2 ± 2.4 a 22.0 ± 2.0 b 21.6 ± 0.8 b 21.9 ± 1.3 b <0.0001 0.78 0.81
Water intake, mL/day 32.0 ± 3.4 a 33.9 ± 2.8 a 33.6 ± 2.4 a 20.0 ± 1.8 b 24.8 ± 0.8 b 20.9 ± 1.7 b <0.0001 0.37 0.73
Energy intake, kJ/day 396.7 ± 20.2 b 398.9 ± 11.1 b 406.6 ± 11.3 b 469.9 ± 9.3 a 483.2 ± 15.2 a 478.2 ± 4.1 a <0.0001 0.75 0.86
Feed conversion
efficiency, % 2.8 ± 0.3 ab 2.5 ± 0.3 b 4.1 ± 0.4 a 2.9 ± 0.3 ab 2.9 ± 0.2 ab 3.6 ± 0.5 ab 1.00 0.004 0.44
Body weight gain, % 11.1 ± 1.1 b 9.9 ± 1.1 b 16.7 ± 1.5 a 12.8 ± 1.4 ab 13.8 ± 1.2 ab 17.3 ± 2.2 a 0.10 0.001 0.54
Values are mean ± SEM, n = 8–10, over the last 8 weeks of the 16 week protocol. Means within a row with unlike superscript differ, p < 0.05. C, corn starch fed rats; CUO, corn starch rats
treated with Ulva ohnoi; CDT, corn starch rats treated with Derbesia tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated with Ulva ohnoi;
HDT, high-carbohydrate, high-fat rats treated with Derbesia tenuissima.
Table 2. Metabolic parameters in rats fed C or H and with either Ulva ohnoi or Derbesia tenuissima.
Variable C CUO CDT H HUO HDT P-Value
Diet Treatment Interaction
Bone mineral content, g 12.4 ± 0.3 c 12.4 ± 0.5 c 13.5 ± 0.4 bc 15.1 ± 0.5 a 14.3 ± 0.4 ab 15.7 ± 0.4 a <0.0001 0.017 0.64
Bone mineral density, g/cm2 0.162 ± 0.003 0.163 ± 0.003 0.164 ± 0.003 0.167 ± 0.002 0.161 ± 0.002 0.162 ± 0.002 0.88 0.63 0.30
Total lean mass, g 319.1 ± 10.3 ab 302.4 ± 2.5 bc 333.2 ± 6.6 a 271.4 ± 8.4 c 292.3 ± 11.8 bc 284.4 ± 3.9 c <0.0001 0.23 0.04
Total body fat mass, g 85.5 ± 7.4 c 103.8 ± 9.8 c 100.1 ± 7.9 c 201.6 ± 10.9 a 153.1 ± 14.6 b 190.0 ± 18.9 a <0.0001 0.32 0.026
Abdominal circumference, cm 19.6 ± 0.4 b 18.8 ± 0.2 b 19.6 ± 0.2 b 22.0 ± 0.4 a 21.4 ± 0.2 a 22.2 ± 0.6 a <0.0001 0.08 0.95
Glucose AUC, mmol/L/120 min 637.6 ± 8.4 c 674.9 ± 17.6 bc 700.7 ± 22.0 b 809.7 ± 12.9 a 703.5 ± 18.8 b 728.6 ± 12.2 b <0.0001 0.09 <0.0001
Insulin AUC, mmol/L/120 min 169.5 ± 22.8 c 226.8 ± 6.2 bc 228.7 ± 13.9 bc 391.8 ± 16.2 a 226.5 ± 18.3 bc 254.5 ± 33.9 b <0.0001 0.03 <0.0001
Retroperitoneal fat, mg/mm tibial length 138.8 ± 11.1 b 127.8 ± 10.6 b 150.4 ± 9.3 b 331.5 ± 20.9 a 303.3 ± 16.9 a 322.0 ± 18.8 a <0.0001 0.32 0.76
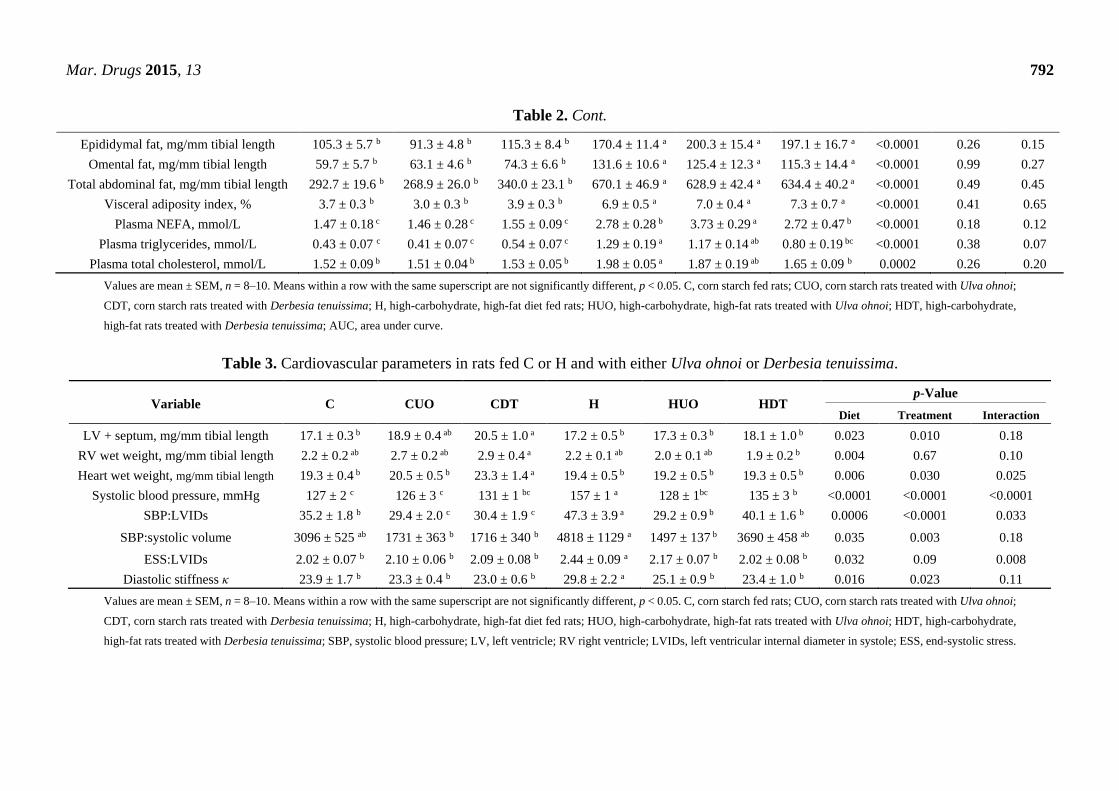
Mar. Drugs 2015, 13 792
Table 2. Cont.
Epididymal fat, mg/mm tibial length 105.3 ± 5.7 b 91.3 ± 4.8 b 115.3 ± 8.4 b 170.4 ± 11.4 a 200.3 ± 15.4 a 197.1 ± 16.7 a <0.0001 0.26 0.15
Omental fat, mg/mm tibial length 59.7 ± 5.7 b 63.1 ± 4.6 b 74.3 ± 6.6 b 131.6 ± 10.6 a 125.4 ± 12.3 a 115.3 ± 14.4 a <0.0001 0.99 0.27
Total abdominal fat, mg/mm tibial length 292.7 ± 19.6 b 268.9 ± 26.0 b 340.0 ± 23.1 b 670.1 ± 46.9 a 628.9 ± 42.4 a 634.4 ± 40.2 a <0.0001 0.49 0.45
Visceral adiposity index, % 3.7 ± 0.3 b 3.0 ± 0.3 b 3.9 ± 0.3 b 6.9 ± 0.5 a 7.0 ± 0.4 a 7.3 ± 0.7 a <0.0001 0.41 0.65
Plasma NEFA, mmol/L 1.47 ± 0.18 c 1.46 ± 0.28 c 1.55 ± 0.09 c 2.78 ± 0.28 b 3.73 ± 0.29 a 2.72 ± 0.47 b <0.0001 0.18 0.12
Plasma triglycerides, mmol/L 0.43 ± 0.07 c 0.41 ± 0.07 c 0.54 ± 0.07 c 1.29 ± 0.19 a 1.17 ± 0.14 ab 0.80 ± 0.19 bc <0.0001 0.38 0.07
Plasma total cholesterol, mmol/L 1.52 ± 0.09 b 1.51 ± 0.04 b 1.53 ± 0.05 b 1.98 ± 0.05 a 1.87 ± 0.19 ab 1.65 ± 0.09 b 0.0002 0.26 0.20
Values are mean ± SEM, n = 8–10. Means within a row with the same superscript are not significantly different, p < 0.05. C, corn starch fed rats; CUO, corn starch rats treated with Ulva ohnoi;
CDT, corn starch rats treated with Derbesia tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated with Ulva ohnoi; HDT, high-carbohydrate,
high-fat rats treated with Derbesia tenuissima; AUC, area under curve.
Table 3. Cardiovascular parameters in rats fed C or H and with either Ulva ohnoi or Derbesia tenuissima.
Variable C CUO CDT H HUO HDT p-Value
Diet Treatment Interaction
LV + septum, mg/mm tibial length 17.1 ± 0.3 b 18.9 ± 0.4 ab 20.5 ± 1.0 a 17.2 ± 0.5 b 17.3 ± 0.3 b 18.1 ± 1.0 b 0.023 0.010 0.18
RV wet weight, mg/mm tibial length 2.2 ± 0.2 ab 2.7 ± 0.2 ab 2.9 ± 0.4 a 2.2 ± 0.1 ab 2.0 ± 0.1 ab 1.9 ± 0.2 b 0.004 0.67 0.10
Heart wet weight, mg/mm tibial length 19.3 ± 0.4 b 20.5 ± 0.5 b 23.3 ± 1.4 a 19.4 ± 0.5 b 19.2 ± 0.5 b 19.3 ± 0.5 b 0.006 0.030 0.025
Systolic blood pressure, mmHg 127 ± 2 c 126 ± 3 c 131 ± 1 bc 157 ± 1 a 128 ± 1bc 135 ± 3 b <0.0001 <0.0001 <0.0001
SBP:LVIDs 35.2 ± 1.8 b 29.4 ± 2.0 c 30.4 ± 1.9 c 47.3 ± 3.9 a 29.2 ± 0.9 b 40.1 ± 1.6 b 0.0006 <0.0001 0.033
SBP:systolic volume 3096 ± 525 ab 1731 ± 363 b 1716 ± 340 b 4818 ± 1129 a 1497 ± 137 b 3690 ± 458 ab 0.035 0.003 0.18
ESS:LVIDs 2.02 ± 0.07 b 2.10 ± 0.06 b 2.09 ± 0.08 b 2.44 ± 0.09 a 2.17 ± 0.07 b 2.02 ± 0.08 b 0.032 0.09 0.008
Diastolic stiffness κ 23.9 ± 1.7 b 23.3 ± 0.4 b 23.0 ± 0.6 b 29.8 ± 2.2 a 25.1 ± 0.9 b 23.4 ± 1.0 b 0.016 0.023 0.11
Values are mean ± SEM, n = 8–10. Means within a row with the same superscript are not significantly different, p < 0.05. C, corn starch fed rats; CUO, corn starch rats treated with Ulva ohnoi;
CDT, corn starch rats treated with Derbesia tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated with Ulva ohnoi; HDT, high-carbohydrate,
high-fat rats treated with Derbesia tenuissima; SBP, systolic blood pressure; LV, left ventricle; RV right ventricle; LVIDs, left ventricular internal diameter in systole; ESS, end-systolic stress.
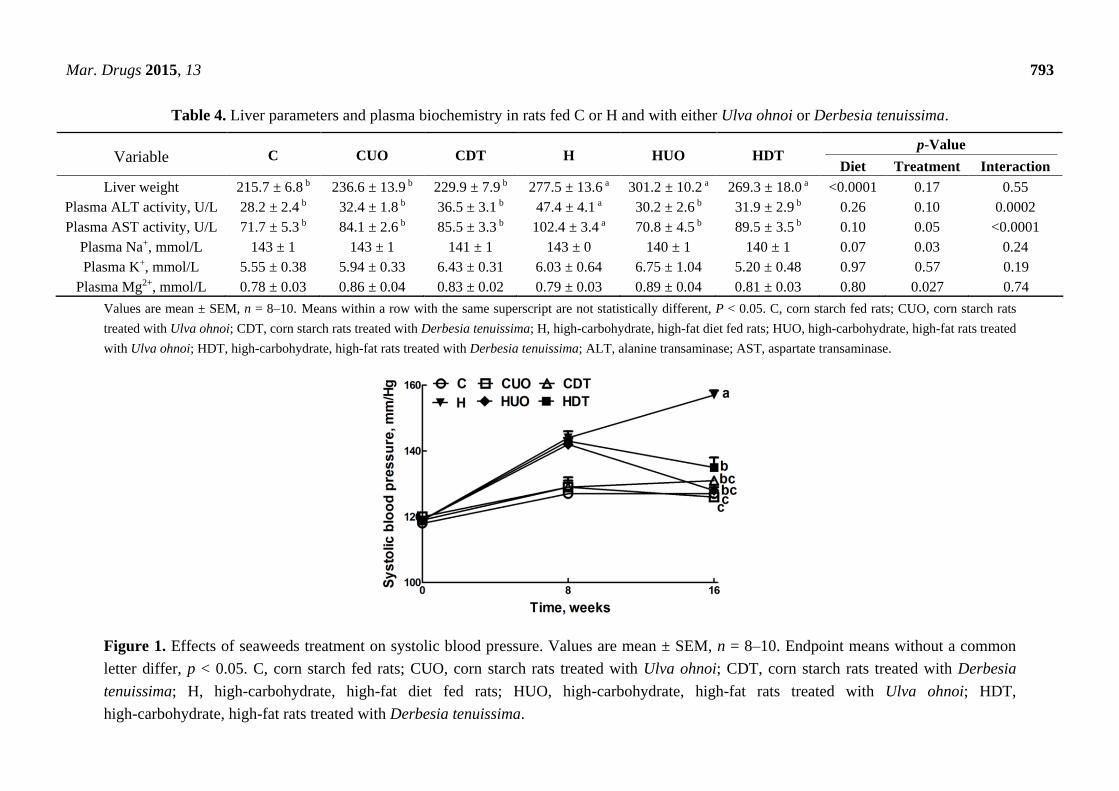
Mar. Drugs 2015, 13 793
Table 4. Liver parameters and plasma biochemistry in rats fed C or H and with either Ulva ohnoi or Derbesia tenuissima.
Variable C CUO CDT H HUO HDT p-Value
Diet Treatment Interaction
Liver weight 215.7 ± 6.8 b 236.6 ± 13.9 b 229.9 ± 7.9 b 277.5 ± 13.6 a 301.2 ± 10.2 a 269.3 ± 18.0 a <0.0001 0.17 0.55
Plasma ALT activity, U/L 28.2 ± 2.4 b 32.4 ± 1.8 b 36.5 ± 3.1 b 47.4 ± 4.1 a 30.2 ± 2.6 b 31.9 ± 2.9 b 0.26 0.10 0.0002
Plasma AST activity, U/L 71.7 ± 5.3 b 84.1 ± 2.6 b 85.5 ± 3.3 b 102.4 ± 3.4 a 70.8 ± 4.5 b 89.5 ± 3.5 b 0.10 0.05 <0.0001
Plasma Na+, mmol/L 143 ± 1 143 ± 1 141 ± 1 143 ± 0 140 ± 1 140 ± 1 0.07 0.03 0.24
Plasma K+, mmol/L 5.55 ± 0.38 5.94 ± 0.33 6.43 ± 0.31 6.03 ± 0.64 6.75 ± 1.04 5.20 ± 0.48 0.97 0.57 0.19
Plasma Mg2+, mmol/L 0.78 ± 0.03 0.86 ± 0.04 0.83 ± 0.02 0.79 ± 0.03 0.89 ± 0.04 0.81 ± 0.03 0.80 0.027 0.74
Values are mean ± SEM, n = 8–10. Means within a row with the same superscript are not statistically different, P < 0.05. C, corn starch fed rats; CUO, corn starch rats
treated with Ulva ohnoi; CDT, corn starch rats treated with Derbesia tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated
with Ulva ohnoi; HDT, high-carbohydrate, high-fat rats treated with Derbesia tenuissima; ALT, alanine transaminase; AST, aspartate transaminase.
Figure 1. Effects of seaweeds treatment on systolic blood pressure. Values are mean ± SEM, n = 8–10. Endpoint means without a common
letter differ, p < 0.05. C, corn starch fed rats; CUO, corn starch rats treated with Ulva ohnoi; CDT, corn starch rats treated with Derbesia
tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated with Ulva ohnoi; HDT,
high-carbohydrate, high-fat rats treated with Derbesia tenuissima.
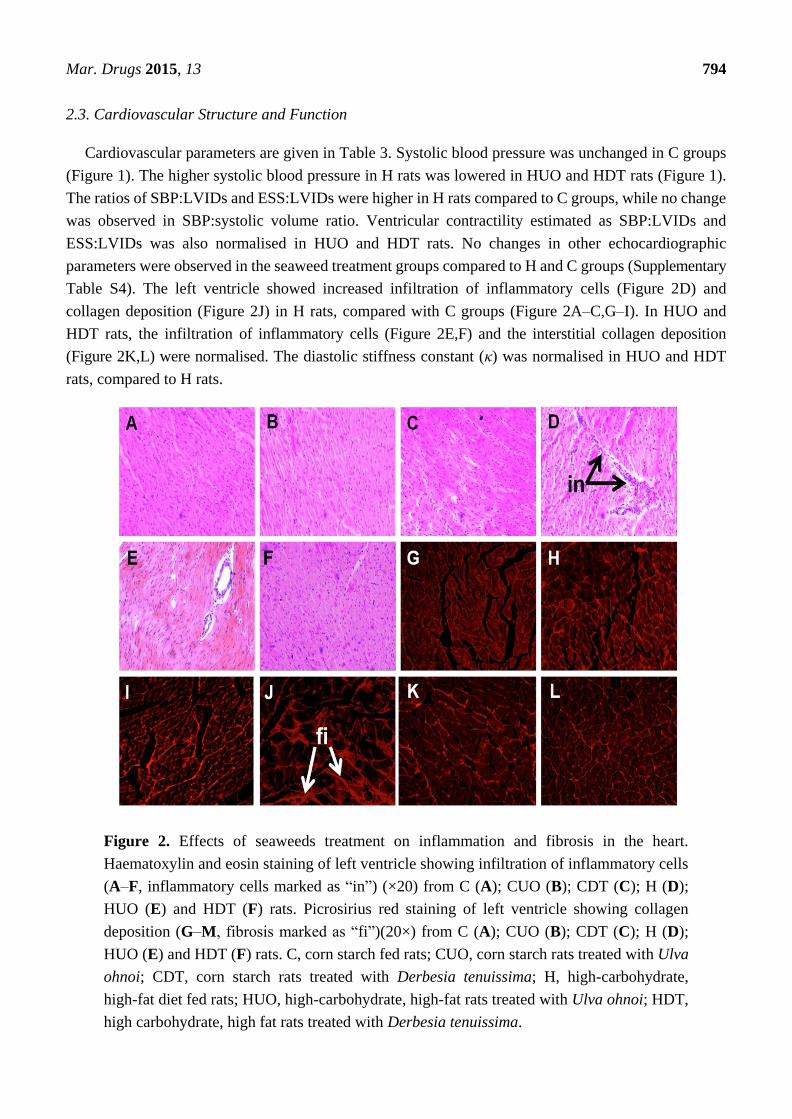
Mar. Drugs 2015, 13 794
2.3. Cardiovascular Structure and Function
Cardiovascular parameters are given in Table 3. Systolic blood pressure was unchanged in C groups
(Figure 1). The higher systolic blood pressure in H rats was lowered in HUO and HDT rats (Figure 1).
The ratios of SBP:LVIDs and ESS:LVIDs were higher in H rats compared to C groups, while no change
was observed in SBP:systolic volume ratio. Ventricular contractility estimated as SBP:LVIDs and
ESS:LVIDs was also normalised in HUO and HDT rats. No changes in other echocardiographic
parameters were observed in the seaweed treatment groups compared to H and C groups (Supplementary
Table S4). The left ventricle showed increased infiltration of inflammatory cells (Figure 2D) and
collagen deposition (Figure 2J) in H rats, compared with C groups (Figure 2A–C,G–I). In HUO and
HDT rats, the infiltration of inflammatory cells (Figure 2E,F) and the interstitial collagen deposition
(Figure 2K,L) were normalised. The diastolic stiffness constant (κ) was normalised in HUO and HDT
rats, compared to H rats.
Figure 2. Effects of seaweeds treatment on inflammation and fibrosis in the heart.
Haematoxylin and eosin staining of left ventricle showing infiltration of inflammatory cells
(A–F, inflammatory cells marked as “in”) (×20) from C (A); CUO (B); CDT (C); H (D);
HUO (E) and HDT (F) rats. Picrosirius red staining of left ventricle showing collagen
deposition (G–M, fibrosis marked as “fi”)(20×) from C (A); CUO (B); CDT (C); H (D);
HUO (E) and HDT (F) rats. C, corn starch fed rats; CUO, corn starch rats treated with Ulva
ohnoi; CDT, corn starch rats treated with Derbesia tenuissima; H, high-carbohydrate,
high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated with Ulva ohnoi; HDT,
high carbohydrate, high fat rats treated with Derbesia tenuissima.
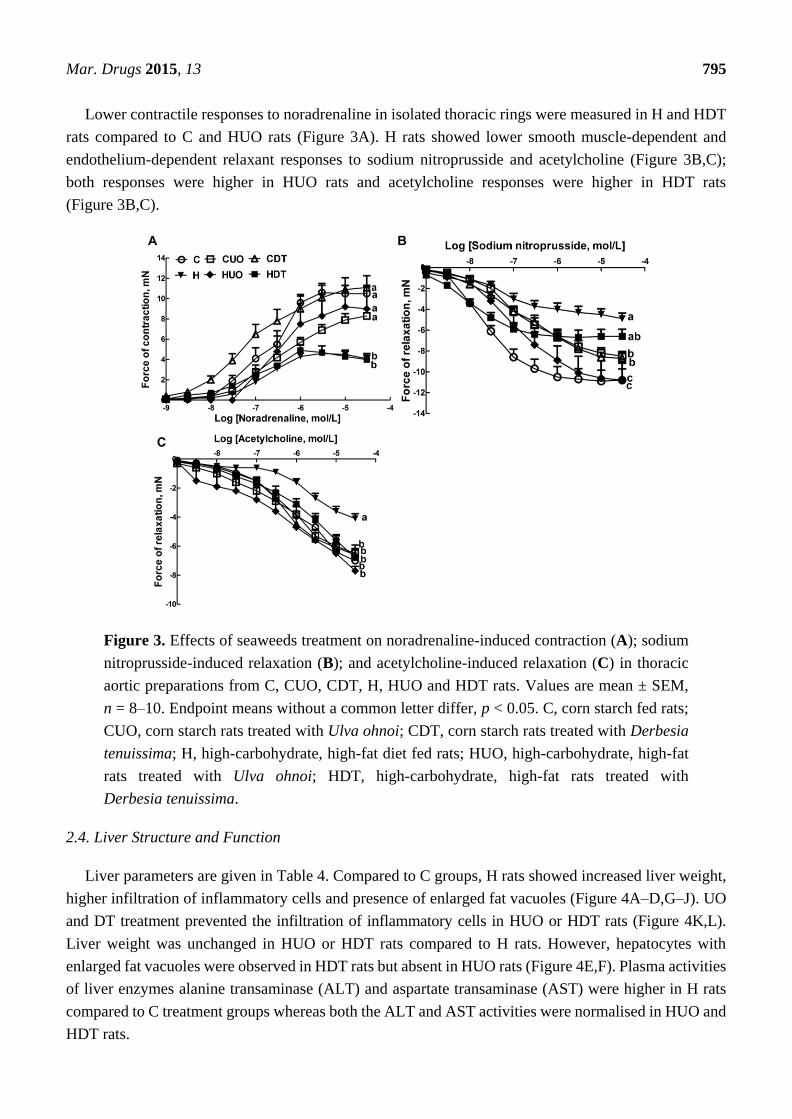
Mar. Drugs 2015, 13 795
Lower contractile responses to noradrenaline in isolated thoracic rings were measured in H and HDT
rats compared to C and HUO rats (Figure 3A). H rats showed lower smooth muscle-dependent and
endothelium-dependent relaxant responses to sodium nitroprusside and acetylcholine (Figure 3B,C);
both responses were higher in HUO rats and acetylcholine responses were higher in HDT rats
(Figure 3B,C).
Figure 3. Effects of seaweeds treatment on noradrenaline-induced contraction (A); sodium
nitroprusside-induced relaxation (B); and acetylcholine-induced relaxation (C) in thoracic
aortic preparations from C, CUO, CDT, H, HUO and HDT rats. Values are mean ± SEM,
n = 8–10. Endpoint means without a common letter differ, p < 0.05. C, corn starch fed rats;
CUO, corn starch rats treated with Ulva ohnoi; CDT, corn starch rats treated with Derbesia
tenuissima; H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat
rats treated with Ulva ohnoi; HDT, high-carbohydrate, high-fat rats treated with
Derbesia tenuissima.
2.4. Liver Structure and Function
Liver parameters are given in Table 4. Compared to C groups, H rats showed increased liver weight,
higher infiltration of inflammatory cells and presence of enlarged fat vacuoles (Figure 4A–D,G–J). UO
and DT treatment prevented the infiltration of inflammatory cells in HUO or HDT rats (Figure 4K,L).
Liver weight was unchanged in HUO or HDT rats compared to H rats. However, hepatocytes with
enlarged fat vacuoles were observed in HDT rats but absent in HUO rats (Figure 4E,F). Plasma activities
of liver enzymes alanine transaminase (ALT) and aspartate transaminase (AST) were higher in H rats
compared to C treatment groups whereas both the ALT and AST activities were normalised in HUO and
HDT rats.
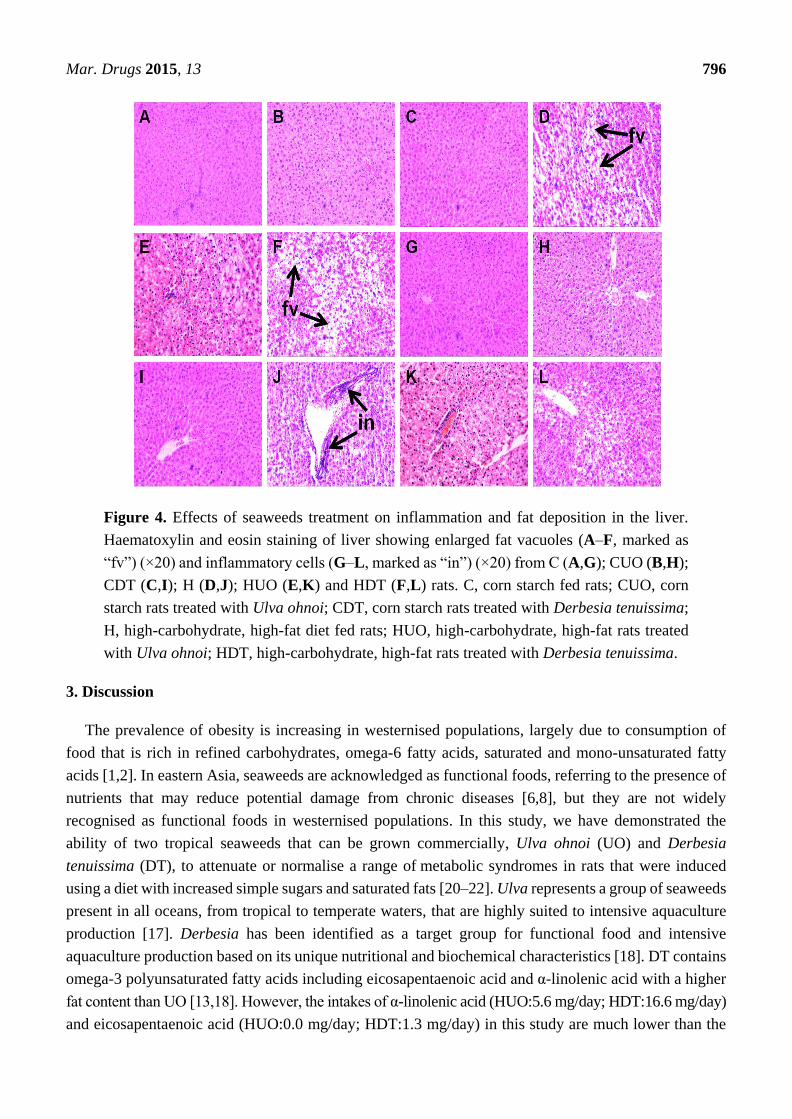
Mar. Drugs 2015, 13 796
Figure 4. Effects of seaweeds treatment on inflammation and fat deposition in the liver.
Haematoxylin and eosin staining of liver showing enlarged fat vacuoles (A–F, marked as
“fv”) (×20) and inflammatory cells (G–L, marked as “in”) (×20) from C (A,G); CUO (B,H);
CDT (C,I); H (D,J); HUO (E,K) and HDT (F,L) rats. C, corn starch fed rats; CUO, corn
starch rats treated with Ulva ohnoi; CDT, corn starch rats treated with Derbesia tenuissima;
H, high-carbohydrate, high-fat diet fed rats; HUO, high-carbohydrate, high-fat rats treated
with Ulva ohnoi; HDT, high-carbohydrate, high-fat rats treated with Derbesia tenuissima.
3. Discussion
The prevalence of obesity is increasing in westernised populations, largely due to consumption of
food that is rich in refined carbohydrates, omega-6 fatty acids, saturated and mono-unsaturated fatty
acids [1,2]. In eastern Asia, seaweeds are acknowledged as functional foods, referring to the presence of
nutrients that may reduce potential damage from chronic diseases [6,8], but they are not widely
recognised as functional foods in westernised populations. In this study, we have demonstrated the
ability of two tropical seaweeds that can be grown commercially, Ulva ohnoi (UO) and Derbesia
tenuissima (DT), to attenuate or normalise a range of metabolic syndromes in rats that were induced
using a diet with increased simple sugars and saturated fats [20–22]. Ulva represents a group of seaweeds
present in all oceans, from tropical to temperate waters, that are highly suited to intensive aquaculture
production [17]. Derbesia has been identified as a target group for functional food and intensive
aquaculture production based on its unique nutritional and biochemical characteristics [18]. DT contains
omega-3 polyunsaturated fatty acids including eicosapentaenoic acid and α-linolenic acid with a higher
fat content than UO [13,18]. However, the intakes of α-linolenic acid (HUO:5.6 mg/day; HDT:16.6 mg/day)
and eicosapentaenoic acid (HUO:0.0 mg/day; HDT:1.3 mg/day) in this study are much lower than the

Mar. Drugs 2015, 13 797
intake of α-linolenic acid (350 mg/day) and eicosapentaenoic acid (550 mg/day) that attenuated diet-induced
metabolic syndrome in previous studies using the same model [22]. Thus, the intakes of omega-3
polyunsaturated fatty acids in this study are unlikely to explain the differences between seaweeds or be
sufficient on their own to reverse metabolic changes in the H rats. Therefore, we now focus on three
possible mechanisms whereby increased intake of fibre or magnesium ions could improve metabolic
syndrome by acting as prebiotics, improving cardiovascular function or decreasing inflammation.
Differing fibre contents with UO containing similar amounts of both soluble and insoluble fibre and
DT containing only insoluble fibre could partially explain the different responses to these two seaweeds.
Dietary fibre, including both soluble (ß-glucans, pectins, gums, mucilages and some hemicelluloses) and
insoluble fibre (celluloses and hemicelluloses) from whole grains, vegetables and fruits, was directly
associated with reducing the risk factors of cardiovascular disease, and the management of obesity,
hypertension, hyperlipidaemia and diabetes [23]. The American Dietetic Association recommends a daily
dietary fibre intake of 25 g for adult females and 38 g for adult men [24]. Based on body surface area,
conversion of the seaweed intake in the current study in rats to humans [25] would provide a fibre intake
of between 5.8 and 15.6 g/day, lower than the recommendations, but sufficient to double current intake
in the USA. The physicochemical properties of seaweed fibre, such as the ability to absorb and hold
water, ion exchange capacity and viscosity, will slow down absorption of compounds in the gut [26,27].
We suggest that the high soluble fibre content of UO could increase gastrointestinal viscosity and
therefore inhibit intestinal absorption of fatty acids more effectively than DT, leading to the decreased
total body fat mass with UO but not DT. Both UO and DT treatments improved glucose utilisation and
insulin sensitivity, potentially through similar mechanisms. This has been shown with viscous soluble
dietary fibre from linseed in young male Wistar rats [28]. Further, the fibre present in green seaweed is
more similar to terrestrial crops than the fibre present in red and brown seaweed [29]. The fibre may
work as a prebiotic, defined as polysaccharides that are not broken down in the stomach but are
fermented in the intestine to improve gut bacteria [30]; these changes in intestinal bacteria could prevent
obesity [31].
In addition to fibre, the mineral ion contents of the seaweed supplements were up to 3-fold higher
than the control diets. The increased magnesium content in UO could delay the onset of diabetes as
reported in OLETF rats where an increased magnesium intake of 16 mg/day [32] improved insulin
sensitivity and glucose utilisation [33]. In contrast, magnesium deficiency aggravated the insulin
resistance produced by high fat diets given to growing rats [34]. None of the other major minerals in the
seaweeds such as potassium, calcium and zinc were present in sufficient quantities to have bioactive
effects on the metabolic syndrome [35–37].
The potential influence of fibre and minerals on metabolic syndrome extended to other physiological
changes in the high-carbohydrate, high-fat fed rats with attenuation or normalisation of the increases in
blood pressure, ventricular diastolic stiffness, fibrosis and liver damage. High dietary fibre supplements
in hypertensive subjects reduced systolic and diastolic blood pressures compared to subjects with low
dietary fibre [38]. Increased dietary fibre may reduce blood pressure by reducing insulin resistance, by
reducing fat stores [6,39] or by increasing magnesium intake [33], or all three as in this study. The
decreased infiltration of inflammatory cells probably precedes the decreased collagen synthesis and
deposition in both the heart and liver. Both UO and DT supplementation improved liver structure and
function, with multiple potential mechanisms such as improved insulin sensitivity and glucose tolerance,

Mar. Drugs 2015, 13 798
decreased blood pressure, decreased fat deposition and decreased infiltration of inflammatory cells.
Further, supplementation of DT to H rats reduced the plasma concentrations of triglycerides and total
cholesterol. We suggest that the bioactive polysaccharides present in DT and UO may be responsible for
these cardiovascular and liver responses [6,9] and deserve further study.
Inflammation is critically important in the development of obesity [40]; we showed increased
infiltration of inflammatory cells in the heart and liver of H rats that was markedly attenuated by both
UO and DT. In human adults with an increased fat intake of 78.8–84.2 g/day, increased dietary fibre
intake of 14.3–16.6 g/day decreased serum C-reactive protein concentrations, a non-specific marker of
inflammation [41]. Bioactive polysaccharides from many different seaweeds have demonstrated
anti-inflammatory activity [42]. Furthermore, in vivo and in vitro studies observed that dietary fibre and
short-chain carboxylic acids, such as propionate and butyrate, released during the bacterial fermentation
of dietary fibre as prebiotics attenuated the production of pro-inflammatory cytokines including
interleukin-8, interleukin-6 and tumour necrosis factor-α [43]. Similar modes of action are likely with
green seaweed polysaccharides [29].
4. Experimental Section
4.1. Resources: Algae, Diet Components, Rats
Two species of green algae (Chlorophyta), Ulva ohnoi (UO) and Derbesia tenuissima (DT), were
cultured in the aquaculture facilities of James Cook University, Townsville, Australia. All biomass was
produced in large outdoor tanks with capacities of 2500 L for Derbesia and >10,000 L for Ulva. Biomass
was harvested on two occasions separated by ~12 months in 2011–2012. The biomass was rinsed in
freshwater and freeze-dried, after which sub-samples were taken for analyses of fibre components (100
g dry-weight), and minerals and fatty acid concentrations (200 mg dry-weight).
In each sub-sample, 24 trace elements were quantified and mean values are reported (n = 2
sub-samples per species). Al, Ca, K, Na, S and P were analysed by Inductively Coupled Plasma Optical
Emission Spectrometry, while metals and metalloids (As, B, Ba, Cd, Co, Cr, Cu, Fe, Hg, Mg, Mn, Mo,
Ni, Pb, Se, Sr, V and Zn) were analysed by Inductively Coupled Plasma Mass Spectrometry at the
Advanced Analytical Centre, James Cook University, Townsville. Insoluble and soluble fibre was
analysed on a combined sample of 100 g for each species containing 50 g from each harvest time. Fibre
analyses were run using enzymatic-gravimetric methods by National Measurement Institute, Sydney,
Australia (AOAC Official Method 985.29 for insoluble fibre; AOAC Official Method 993.19 for soluble
fibre). Total crude lipids and fatty acids were extracted and analysed [13]. All remaining freeze-dried
biomass was stored in vacuum-sealed bags under refrigeration until preparation of the food.
The experimental groups consisted of 72 male Wistar rats (9–10 weeks-old; 336 ± 2 g) individually
housed in a temperature-controlled (20 ± 2 °C), 12 h light/dark cycle environment with unrestricted
access to water and food. The CS diet contained 570 g cornstarch, 155 g powdered rat food (Specialty
Feeds, Glen Forest, WA, Australia), 25 g Hubble, Mendel and Wakeman (HMW) salt mixture and
250 g water per kilogram of diet. The HCHF diet consisted of 175 g fructose, 395 g sweetened condensed
milk, 200 g beef tallow, 155 g powdered rat food, 25 g HMW salt mixture and 50 g water per kilogram
of diet. In addition, the drinking water for the HCHF group was supplemented with 25% fructose. Energy

Mar. Drugs 2015, 13 799
intake was calculated from the following values in kJ/g: fructose, 15.40; corn starch, 15.94; condensed
milk, 13.80; beef tallow, 37.70 and powdered rat food, 13.80. The energy densities of the CS diet and
the HCHF diet were 11.23 kJ/g and 17.83 kJ/g of food respectively and an additional 3.85 kJ/mL in the
drinking water for the HCHF diet-fed rats [20,21]. The rats were randomly divided into 6 separate groups
(n = 12 each) and fed with corn starch (C), corn starch + Ulva ohnoi 5% (CUO), corn starch + Derbesia
tenuissima 5% (CDT), high-carbohydrate, high-fat (H), high-carbohydrate, high-fat + Ulva ohnoi 5%
(HUO), or high-carbohydrate, high-fat + Derbesia tenuissima 5% (HDT). The seaweed-supplemented
diets were prepared by adding 5% of seaweeds to replace an equivalent amount of water in the diet. The
drinking water in all H groups included 25% fructose. The seaweed-supplemented diets were
administered for 8 weeks starting 8 weeks after the initiation of the C or H diets.
All experimentation was approved by the Animal Ethics Committees of The University of
Queensland and University of Southern Queensland under the guidelines of the National Health and
Medical Research Council of Australia. Rats were monitored daily for body weight, and food and water
intakes. Daily seaweed intake was calculated from the daily food intake. The fatty acid concentrations
of both C and H control diets used for the calculation of mean daily fatty acids intake were obtained
from our previous study [22].
Abdominal circumference of rats was measured using a standard measuring tape during the period of
anaesthesia for systolic blood pressure (SBP) measurements. SBP measurements, and oral glucose and
insulin tolerance tests were conducted at 0, 8 and 16 weeks. Oral glucose tolerance tests were performed
following determination of overnight fasting blood glucose concentrations in tail vein blood using
Medisense Precision Q.I.D glucose meters (Abbott Laboratories, Bedford, MA, USA). For overnight
fasting, rats were deprived of food for 12 h. Fructose-supplemented drinking water in HCHF groups was
replaced with normal drinking water for the overnight food-deprivation period. Rats were given a
glucose load of 2 g/kg body weight as 40% glucose solution via oral gavage and blood glucose
concentrations were measured again 30, 60, 90 and 120 min after oral glucose administration. For insulin
tolerance, basal blood glucose concentrations were measured after 4–5 h of food deprivation as above.
The rats were injected ip 0.33 IU/kg insulin-R (Eli Lilly Australia, West Ryde, NSW, Australia), and
tail vein blood samples were taken at 0, 30, 60, 90 and 120 min. Rats were withdrawn from the test if
the blood glucose dropped below 1.1 mmol/L, and 4 g/kg glucose was immediately administered by oral
gavage to prevent hypoglycemia [20–22]. Dual energy X-ray absorptiometric (DXA) measurements
were performed on rats after 16 weeks of feeding, 2 days before euthanasia using a Norland XR36 DXA
instrument (Norland Corp, Fort Atkinson, WI, USA). DXA scans were analysed using the manufacturer’s
recommended software for use in laboratory animals (Small Subject Analysis Software, version 2.5.3/1.3.1;
Norland Corp). The precision error of lean mass for replicate measurements, with repositioning,
was 3.2% [20–22]. Feed conversion efficiency was calculated as [mean body weight gain (in grams)/
daily energy intake (in kilojoules)]. Visceral adiposity index (%) was calculated [20–22].
The experimental protocol is summarised in Figure 5.

Mar. Drugs 2015, 13 800
Figure 5. Outline of the experimental protocol for measurements on rats. SBP, systolic blood
pressure; OGTT, oral glucose tolerance test; AC, abdominal circumference, ITT, insulin
tolerance test; DEXA, dual energy X-ray absorptiometry; UO, Ulva ohnoi; DT,
Derbesia tenuissima.
4.2. Cardiovascular Measurements
Systolic blood pressure was measured under light sedation following intraperitoneal injection of
Zoletil (tiletamine 15 mg/kg, zolazepam 15 mg/kg; Virbac, Milperra, Australia) using a MLT1010
Piezo-Electric Pulse Transducer (ADInstruments, Sydney, Australia) and inflatable tail-cuff connected
to a MLT844 Physiological Pressure Transducer (ADInstruments, Sydney, Australia) using PowerLab
data acquisition unit (ADInstruments, Sydney, Australia) [20–22].
Echocardiographic examinations (Phillips iE33, 12MHz transducer, Best, The Netherlands) were
performed to assess the cardiovascular structure and function in all rats at 16 weeks [20–22]. Rats were
anaesthetised using Zoletil (tiletamine 25 mg/kg and zolazepam 25 mg/kg, intraperitoneally (i.p);
Virbac, Peakhurst, Australia) and Ilium Xylazil (xylazine 15 mg/kg, i.p.; Troy Laboratories, Smithfield,
Australia). The ventricular contractility indexes were calculated including ratio of systolic blood
pressure (SBP) to left ventricular internal diameter in systole (LVIDs), ratio of SBP to systolic volume,
and ratio of end-systolic stress (ESS) to LVIDs [21,44].
The left ventricular function of the rats was assessed using the Langendorff heart preparation [20–22].
Terminal anaesthesia was induced via i.p. injection of pentobarbitone sodium (Lethabarb®, 100 mg/kg,
Virbac, Milperra, Australia). Blood (~5 mL) was taken from the abdominal aorta after heparin (200 IU;
Sigma-Aldrich Australia, Sydney, Australia) administration through the right femoral vein.
Isovolumetric ventricular function was measured by inserting a latex balloon catheter into the left
ventricle of the isolated heart connected to a Capto SP844 MLT844 physiological pressure transducer
and Chart software on a MacLab system (ADInstruments, Sydney, Australia).
Thoracic aortic rings (~4 mm in length) were suspended in an organ bath chamber with a resting
tension of approximately 10 mN. Cumulative concentration-response (contraction) curves were
measured for noradrenaline (Sigma-Aldrich Australia, Sydney, Australia); concentration-response

Mar. Drugs 2015, 13 801
(relaxation) curves were measured for acetylcholine (Sigma-Aldrich Australia, Sydney, Australia) and
sodium nitroprusside (Sigma-Aldrich Australia, Sydney, Australia) in the presence of a submaximal
(70%) contraction to noradrenaline [20–22].
4.3. Organ Weights
The right and left ventricles were separated after Langendorff experiments and weighed. Liver, and
retroperitoneal, epididymal and omental fat pads were removed following heart removal and blotted
dry for weighing. Organ weights were normalised relative to the tibial length at the time of their removal
(in mg/mm) [20–22].
4.4. Histology
Two rats per group were taken exclusively for histological analysis. Two slides were prepared per
tissue specimen and two random, non-overlapping fields per slide were photographed. Immediately after
removal, heart and liver tissues were fixed in 10% neutral buffered formalin for 3 days and then
dehydrated and embedded in paraffin wax [20–22]. Thin sections (7 μm) of left ventricle and the liver
were cut and stained with haematoxylin and eosin stain for determination of inflammatory cell
infiltration with 20X and fat vacuole enlargement with 40× objectives using a Olympus BX51
microscope (Olympus, Melville, NY, USA). Collagen distribution was measured in the left ventricle
with picrosirius red stain. Laser confocal microscopy (Zeiss LSM 510 upright Confocal Microscope,
Carl Zeiss, North Ryde, Australia) was used to visualize collagen deposition [20–22].
4.5. Plasma Biochemistry
Blood was centrifuged within 30 min of collection into heparinised tubes at 5000 g for 15 min. Plasma
was separated and transferred to Eppendorf tubes (Thermo Fisher Scientific, Scoresby, Australia) for
storage at −20 °C before analysis. Activities of plasma enzymes and analyte concentrations were
determined using kits and controls supplied by Olympus using an Olympus analyser (AU 400, Tokyo,
Japan) [20–22].
4.6. Statistical Analysis
All data are presented as mean ± SEM. Data from C, CUO, CDT, H, HUO and HDT groups were
compared in a series of two-way ANOVAs (Analysis of Variance) with two types of “Diet”, high
carbohydrate and high fat diet or control cornstarch diet, and three types of “Treatment”, a control and
treatments supplemented with each seaweed, as the two fixed factors in the analyses; C was compared
with H, CUO or CDT; H was compared with HUO or HDT. Homogeneity of variance for ANOVA was
assessed using Bartlett’s test and variables that were not normally distributed were log-transformed
prior to analysis. Where the main effects were significant (p < 0.05), means were compared using
Newman-Keuls multiple comparisons. Where transformations did not result in normality or
homogeneity of variance, a Kruskal-Wallis non-parametric test was performed. All statistical analyses
were run using GraphPad Prism version 5.00 for Windows (GraphPad Software, La Jolla, CA, USA).

Mar. Drugs 2015, 13 802
5. Conclusions
Our findings suggest that soluble dietary fibre as a major component in UO could play a key role in
attenuating the signs of the metabolic syndrome such as hypertension, endothelial dysfunction,
diminished insulin sensitivity and glucose utilisation, increased cardiac stiffness, increased collagen
deposition, increased liver damage and increased fat mass in diet-induced obese rats. Further, we suggest
that the insoluble fibre of both DT and UO may improve glucose metabolism and that the increased
magnesium intake in UO could delay the onset of diabetes. Thus, these tropical seaweeds serve as a
commercially viable source of dietary fibre as a functional food, as they can be produced in large
quantities by aquaculture to attenuate the signs of metabolic syndrome.
Acknowledgments
Seaweed biomass was produced at James Cook University, supported by the MBD Energy
Research and Development program for Biological Carbon Capture and Storage and the Advanced
Manufacturing Cooperative Research Centre (AM-CRC), funded through the Australian Government’s
Cooperative Research Centre Scheme. SAK was partly supported by a scholarship from the University
of Southern Queensland.
Author Contributions
S.A.K., M.M., L.C.W. and N.A.P. performed the experiments; N.A.P. and L.B. designed the
experiments; all co-authors contributed to the manuscript. This manuscript forms part of the Ph.D. thesis
to USQ of SAK under the supervision of LB.
Conflicts of Interest
The authors declare no conflict of interest.
References
1. Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H. Obesity and
cardiovascular disease: Pathophysiology, evaluation, and effect of weight loss. Arterioscler.
Thromb. Vasc. Biol. 2006, 26, 968–976.
2. Lavie, C.J.; Milani, R.V.; Ventura, H.O. Obesity and cardiovascular disease: Risk factor, paradox,
and impact of weight loss. J. Am. Coll. Cardiol. 2009, 53, 1925–1932.
3. Munsters, M.J.; Saris, W.H. Body weight regulation and obesity: Dietary strategies to improve the
metabolic profile. Annu. Rev. Food Sci. Technol. 2014, 5, 39–51.
4. Chatzigeorgiou, A.; Kandaraki, E.; Papavassiliou, A.G.; Koutsilieris, M. Peripheral targets in
obesity treatment: A comprehensive update. Obes. Rev. 2014, 15, 487–503.
5. Gadde, K.M. Current pharmacotherapy for obesity: Extrapolation of clinical trials data to practice.
Expert Opin. Pharmacother. 2014, 15, 809–822.
6. Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to
reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100.

Mar. Drugs 2015, 13 803
7. MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible
seaweeds. Nutr. Rev. 2007, 65, 535–543.
8. Mohamed, S.; Hashim, S.N.; Rahman, H.A. Seaweeds: A sustainable functional food for
complementary and alternative therapy. Trends Food Sci. Tech. 2012, 23, 83–96.
9. Mayakrishnan, V.; Kannappan, P.; Abdullah, N.; Ahmed, A.B.A. Cardioprotective activity of
polysaccharides derived from marine algae: An overview. Trends Food Sci. Technol. 2013, 30,
98–104.
10. Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibre
contents in some ‘sea vegetables’. J. Sci. Food Agric. 1991, 54, 587–594.
11. Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible
seaweed products. Food Chem. 2007, 103, 891–899.
12. Jakobsdottir, G.; Nyman, M.; Fak, F. Designing future prebiotic fiber to target metabolic syndrome.
Nutrition 2014, 30, 497–502.
13. Gosch, B.J.; Magnusson, M.; Paul, N.A.; de Nys, R. Total lipid and fatty acid composition of
seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012,
4, 919–930.
14. Pengzhan, Y.; Ning, L.; Xiguang, L.; Gefei, Z.; Quanbin, Z.; Pengcheng, L. Antihyperlipidemic
effects of different molecular weight sulfated polysaccharides from Ulva pertusa (Chlorophyta).
Pharmacol. Res. 2003, 48, 543–549.
15. Wang, R.; Paul, V.J.; Luesch, H. Seaweed extracts and unsaturated fatty acid constituents from the
green alga Ulva lactuca as activators of the cytoprotective Nrf2-ARE pathway. Biol. Med. 2013,
57, 141–153.
16. Ramirez-Higuera, A.; Quevedo-Corona, L.; Paniagua-Castro, N.; Chamorro-Ceballos, G.;
Milliar-Garcia, A.; Jaramillo-Flores, M. Antioxidant enzymes gene expression and antihypertensive
effects of seaweeds Ulva linza and Lessonia trabeculata in rats fed a high-fat and high-sucrose diet.
J. Appl. Phycol. 2014, 26, 597–605.
17. Lawton, R.J.; Mata, L.; de Nys, R.; Paul, N.A. Algal bioremediation of waste waters from
land-based aquaculture using Ulva: Selecting target species and strains. PLoS One 2013, 8, e77344.
18. Magnusson, M.; Mata, L.; de Nys, R.; Paul, N.A. Biomass, lipid and fatty acid production in
large-scale cultures of the marine macroalga Derbesia tenuissima (Chlorophyta). Mar. Biotechnol. (NY)
2014, 16, 456–464.
19. Lee, H.J.; Kim, Y.A.; Lee, J.I.; Lee, B.J.; Seo, Y. Screening of Korean marine plants extracts for
inhibitory activity on protein tyrosine phosphatase 1B. J. Appl. Biol. Chem. 2007, 57, 74–77.
20. Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.;
Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and
cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624.
21. Panchal, S.K.; Poudyal, H.; Waanders, J.; Brown, L. Coffee extract attenuates changes in
cardiovascular and hepatic structure and function without decreasing obesity in high-carbohydrate,
high-fat diet-fed male rats. J. Nutr. 2012, 142, 690–697.
22. Poudyal, H.; Panchal, S.K.; Ward, L.C.; Brown, L. Effects of ALA, EPA and DHA in
high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24,
1041–1052.

Mar. Drugs 2015, 13 804
23. Satija, A.; Hu, F. Cardiovascular benefits of dietary fiber. Curr. Atheroscler. Rep. 2012, 14,
505–514.
24. Slavin, J.L. Position of the American Dietetic Association: Health implications of dietary fiber.
J. Am. Diet. Ass. 2008, 108, 1716–1731.
25. Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited.
FASEB J. 2008, 22, 659–661.
26. Bocanegra, A.; Bastida, S.; Benedi, J.; Rodenas, S.; Sanchez-Muniz, F.J. Characteristics and
nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258.
27. Jensen, G.M.; Pedersen, C.; Kristensen, M.; Frost, G.; Astrup, A. Review: Efficacy of alginate
supplementation in relation to appetite regulation and metabolic risk factors: Evidence from animal
and human studies. Obes. Rev. 2013, 14, 129–144.
28. Kristensen, M.; Knudsen, K.E.; Jorgensen, H.; Oomah, D.; Bugel, S.; Toubro, S.; Tetens, I.;
Astrup, A. Linseed dietary fibers reduce apparent digestibility of energy and fat and weight gain in
growing rats. Nutrients 2013, 5, 3287–3298.
29. Domozych, D.S.; Ciancia, M.; Fangel, J.U.; Mikkelsen, M.D.; Ulvskov, P.; Willats, W.G. The cell
walls of green algae: A journey through evolution and diversity. Front. Plant Sci. 2012, 3,
doi:10.3389/fpls.2012.00082.
30. Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.;
Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J.
Nutr. 2010, 104 (Suppl. 2), S1–S63.
31. Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.;
Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl.
Acad. Sci. USA 2004, 101, 15718–15723.
32. Nagai, N.; Ito, Y. Effect of magnesium ion supplementation on obesity and diabetes mellitus in
Otsuka Long-Evans Tokushima Fatty (OLETF) rats under excessive food intake. J. Oleo Sci. 2013,
62, 403–408.
33. Volpe, S.L. Magnesium, the metabolic syndrome, insulin resistance, and type 2 diabetes mellitus.
Crit. Rev. Food Sci. Nutr. 2008, 48, 293–300.
34. Sales, C.H.; Santos, A.R.; Cintra, D.E.; Colli, C. Magnesium-deficient high-fat diet: Effects on
adiposity, lipid profile and insulin sensitivity in growing rats. Clin. Nutr. 2013, 33, 879–888.
35. Zicha, J.; Dobesova, Z.; Behuliak, M.; Kunes, J.; Vaneckova, I. Preventive dietary potassium
supplementation in young salt-sensitive Dahl rats attenuates development of salt hypertension by
decreasing sympathetic vasoconstriction. Acta Physiol. (Oxf.) 2011, 202, 29–38.
36. Nobre, J.L.; Lisboa, P.C.; Santos-Silva, A.P.; Lima, N.S.; Manhaes, A.C.; Nogueira-Neto, J.F.;
Cabanelas, A.; Pazos-Moura, C.C.; Moura, E.G.; de Oliveira, E. Calcium supplementation reverts
central adiposity, leptin, and insulin resistance in adult offspring programed by neonatal nicotine
exposure. J. Endocrinol. 2011, 210, 349–359.
37. Miao, X.; Wang, Y.; Sun, J.; Sun, W.; Tan, Y.; Cai, L.; Zheng, Y.; Su, G.; Liu, Q. Zinc protects
against diabetes-induced pathogenic changes in the aorta: Roles of metallothionein and nuclear
factor (erythroid-derived 2)-like 2. Cardiovasc. Diabetol. 2013, 12, doi:10.1186/1475-2840-12-54.

Mar. Drugs 2015, 13 805
38. Whelton, S.P.; Hyre, A.D.; Pedersen, B.; Yi, Y.; Whelton, P.K.; He, J. Effect of dietary fiber intake
on blood pressure: A meta-analysis of randomized, controlled clinical trials. J. Hypertens. 2005, 23,
475–481.
39. King, D.E.; Mainous, A.G.; Egan, B.M.; Woolson, R.F.; Geesey, M.E. Fiber and C-reactive protein
in diabetes, hypertension, and obesity. Diabetes Care 2005, 28, 1487–1489.
40. Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011,
29, 415–445.
41. Ajani, U.A.; Ford, E.S.; Mokdad, A.H. Dietary fiber and C-reactive protein: Findings from National
Health and Nutrition Examination Survey data. J. Nutr. 2004, 134, 1181–1185.
42. Kumar, S.A.; Brown, L. Seaweeds as potential therapeutic interventions for the metabolic
syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 299–308.
43. Galvez, J.; Rodríguez-Cabezas, M.E.; Zarzuelo, A. Effects of dietary fiber on inflammatory bowel
disease. Mol. Nutr. Food Res. 2005, 49, 601–608.
44. De Simone, G.; di Lorenzo, L.; Costantino, G.; Buonissimo, S.; Moccia, D. Echocardiographic
indexes of left ventricular contractility. Effect of load manipulation in arterial hypertension.
Jpn. Heart J. 1988, 29, 151–160.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article
distributed under the terms and conditions of the Creative Commons Attribution license
(http://creativecommons.org/licenses/by/4.0/).





























