Embed Size (px)
Citation preview

Camp. Biochem. Physiol. Vol. 81C, No. 2, PP. 323-328, 1985 Printed in Great Britain
0306-4492/85 $3.00 + 0.00 0 1985 Pergamon Press Ltd
SENSITIVITY OF THE RESTING SODIUM CONDUCTIVITY OF LEECH RETZIUS NERVE
CELLS TO TETRODOTOXIN
BOGDAN B. BELESLIN
Department of Pathological Physiology, Medical Faculty, Dr Subotika 1, 11000 Belgrade, Yugoslavia
Abstract-l. Simultaneously, the effect of sodium-free medium and tetrodotoxin (3 x lo-* M/ml) were investigated on some passive electrophysiological properties of leech Retzius nerve cells.
2. Complete replacement of Na’ with Tris or addition of tetrodotoxin to the leech Ringer was followed by an increase of input resistance in contrast to the cell-to-cell interaction which was not affected by such a procedure.
3. At the same time tetrodotoxin was not able to block repetitive spike activity. 4. The data imply the existence of two types of Na+ channel in leech Retzius nerve cells.
INTRODUCTION
Ionic current membranes can be distinguished by ion substitution experiments, tracer flux measurements or by inhibition studies with specific blocking agents. One of the most significant achievements made by experiments using the voltage clamp technique is the identification of the sodium inward and potassium outward current in the squid giant axon (Hodgkin and Huxley, 1952). Later on, a number of in- vestigators, using additional selective blocking agents like tetrodotoxin (TTX), tetraethylammonium (TEA) 4-aminopyridine (4-AP), cobalt, lanthanum, vera- pamil and others, demonstrated that the conductance changes of excitable membranes were more numerous than previously thought. The existence of the Ca*+ current (Fatt and Ginsborg, 1958; Hagiwara and Naka, 1964) and now the hydrogen current (Thomas and Meech, 1982) are recognized.
In studies of the action of TTX on identified neurones Beleslin and MihailoviC (1967) and Klein- haus and Prichard (1976, 1983) found that the ex- citability of the Retzius nerve cells of the leech was not depressed. On the other hand, it is well known that the resting potential of Retzius nerve cells of medical and horse leeches are lower than in other neurones (Hagiwara and Morita, 1962; Lent, 1972; Kayser and Lent, 1977; Beleslin, 1977). Theoretically, this can be due to the low resting potassium perme- ability or the high membrane permeability to sodium. In view of these possibilities it was interesting to examine the effect of TTX on resting conductances and compare it with the effect of a sodium-free medium.
MATERIALS AND METHODS
Experiments were carried out on the Retzius nerve cells in the first four abdominal ganglia of the horse leech (Huemopis sunguisugu). Usually, the ring of the three ganglia was placed in a lucite chamber containing 2 ml Ringer solution. The electrical arrangement for current clamping was similar to that previously described (Beleslin, 1971). In order to calculate input and coupling resistances
constant current pulses were applied to both sides of the junction alternately, assuming that the delta network is equivalent for a pair of cells in an electrotonic junction (Bennett, 1966). In this circumstance the change of input resistances of directly polarized cells (R,, , R,,, R;, , R g2) and transfer resistances (R,2, R,,, Ri2, R;,) and conductances (C,, CL) can be given by the following relation
R,,R,, - R:z R;,R;, - R;; R,, - RI, R;2 - Rh
= G;/G,.
Leech Ringer had the standard composition. This Ringer solution was made by replacing NaCl with Tris Cl. Tetrodo- toxin (Sankyo) was added to the fluid in the bath or to the Ringer in the reservoir. To change the solution the chamber was flushed continuously with a volume of fluid at least five times that of the chamber volume. The perfusion rate was kept low so that implanted microelectrodes remained inside the cells during the perfusion.
All experiments were performed at room temperature (2s24°C).
RESULTS
The initial object of the work was to estimate the resting sodium conductance in Retzius nerve cells taking into the account current spread through the electrotonic junction. When the neurones were ex- posed to the Na+ deficient solution the membrane potential immediately hyperpolarized and remained without significant change throughout the 30min exposure. The size of the electrotonic potentials in this fluid increased gradually until they reached a constant steady value within 10min. So in order to ensure that the neurones had reached a steady state in the test fluid at least 10 min were allowed after flushing the solution before measurements were made.
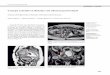
Figure 1 illustrates the main differences between superimposed potential changes in a pair of Retzius cells taken in leech and Tris Ringer. The cells were first equilibrated in leech Ringer, and then soaked in sodium-free medium for 15 min and again returned in the initial medium. The cells were stimulated by rectangular pulses of intensities up to 2 x 10m8 A.
323

324 BOGDAN B. BELESLIN
Cell I Cell II
Leech Ringer
Trts Ringer
Recovery
I--l
50 msec
Fig. 1. Superimposed potential levels in directly polarized (V,,, VZZ) and adjacent cells (I’,,, V,,) taken in leech Ringer, Tris Ringer and during recovery. The cells were alternatively stimulated by hyper- polarizing current pulses (ii and i2) from the left (Cell I) and right side (Cell II) of the junction.
When the hyperpolarized current pulses were passed though one microelectrode appreciable potential changes were recorded in directly polarized and adja- cent cells no matter from which side of the electro- tonic junction the cells were stimulated. The steady state current voltage relation taken 200msec after stimulation for the same cells is shown in Fig, 2. Clearly, in both cells an increase of input resistance occurred, but it was more pronounced in the first cell (42%) compared with the second (28%). At the same time the coupling resistance increased only 12% in the forward direction and 10% in the backward direction. During the next 15 min a partial recovery was ob- tained.
According to the preceding observation, during the resting state there is a sodium conductance, whose sensitivity to TTX was tested. An example of the experiment is illustrated in Fig. 3. The cells were treated with 3 x IO-* M/ml TTX.
From the computed values based on the current voltage relationship for the same cells (Fig. 4) the following conclusions can be drawn:
(1) in the presence of 3 x IO-* M/ml TTX the increase of the input resistance of the pair of cells was 68 and 18% respectively.
(2) an electrotonic spread from one to other cells in the forward and backward directions was present.
(3) the effect of TTX was reversible since in leech Ringer an almost complete recovery of the passive electrophysiological properties was obtained.
From Fig. 3 it is seen that spontaneous spike activity was not inhibited by TTX. Having shown that TTX did not block spontaneous activity, it was interesting to re-examine its effect on evoked repeti- tive spike activity obtained with long-lasting stimu- lation.
TTX, even 60min after application, was not effective in inhibiting repetitive spike activity (Fig. 5).

TTX and resting Na+ conductance
-
I v s_o1 AW OS
-

326 BOGDAN B. BELFSLIN
Leech Ringer Cell I Cell IL
IO-’ A IO-’ A
l TTX
IO-’ A .
lO-8 A
mV
-lOOmV
Fig. 4. Plot of the voltage levels in the two cells at the termination of current strength obtained from the records in Fig. 3.
DISCUSSION
The results reported in this paper show that the resting sodium conductance of Retzius nerve cells of the horse leech is sensitive to the effect of TTX in contrast to the sodium conductance during spike electrogenesis. At the same time a cell to cell inter- action was present.
In the past 30 years much of our knowledge about permeability changes has been gained through the study of the interaction of blocking agents with ionic channels. In this study tetrodotoxin has been used to investigate the sensitivity of resting sodium conduc- tance to its action. The decrease of the resting conductance in the presence of TTX is not surprising, since it is a highly specific agent, used by many investigators to separate sodium inward current from the calcium inward current or non-specific influx of Ca2+ (Katz and Miledi, 1967; Geduldig and Gruener, 1970; Baker et al., 1971; Kostyuk, 1980). However, the presence of TTX-sensitive sodium channels in many tissues including rat skeletal denervated fibres (Albuquerque and Thesleff, 1968), chick embryonic heart muscle fibres (Sperelakis and Shigenobu, 1972) some molluscan neurones (Kostyuk and Krishtal, 1977) has been confirmed. By checking the changes of the spontaneous discharges of spike potentials of Retzius nerve cells of the leech Beleslin and Mihailovic (1967) found that TTX at a concentration of 10d5 g/ml did not have an effect. Later on, Klein-
haus and Prichard (1976) gave evidence that the Retzius nerve cell membrane possessed a significant number of channels which can carry sodium current but cannot be blocked by TTX. A subsequent analy- sis demonstrated a substantial difference in sensitivity to TTX of the sodium-dependent excitation mech- anism in normal mature neurones juxtaposed in the same ganglion (Kleinhaus and Pritchard, 1983). Tetrodotoxin had little or no effect on the excitability of Retzius nerve cells in contrast to sensory neurones which responded to light tactile, sustained pressure and noxious stimuli to the skin. They postulated the existence of two types of sodium channel in normal fully differentiated leech neurones. The present results support the view that they are present in the same Retzius nerve cells.
The resting sodium conductance in Retzius nerve cells was established (Beleslin, 1968). Recent experi- ments performed with K+ and Na+ sensitive micro- electrodes in the presence of the Na+-K+ pump inhibitor ouabain, allowed calculation of the perme- ability of the Retzius cell membrane to K+ and Na+ (Deitmer and Schlue, 1981, 1983). The estimated ratio of the Na+ to K+ permeability coefficient was 0.11 indicating that the resting membrane is about nine times more permeable to K+ than to Nat. The increase of the input resistance without blocking repetitive spike activity, obtained in the current ex- periments with TTX, suggests that its effect is specific

TTX and resting Na+ conductance
Cell I Cell II
Leech Ringer
327
TTX
Recovery
I 2 ::
I 6
9 0 -
Fig. 5. Repetitive spike activity of a pair of Retzius nerve cells taken in leech Ringer in the presence of TTX (3 x IO-* M/ml) and during the recovery.
to the membrane conductance. For more accurate analyses it was necessary to calculate transverse membrane resistance of a unit area of membrane, which is inversely related to the resting conductance of the membrane. In other words, it was necessary to compute only transverse membrane resistance with- out the internal longitudinal resistance. However, since the coupling resistance did not change markedly, it can be concluded that TTX affected only the resting resistive properties of the membrane, since the internal longitudinal resistance in general is a part of the coupling resistance.
It is difficult to tell why TTX blocks resting sodium conductance but not the sodium conductance during spike electrogenesis. One possibility would be that TTX sensitive Nat channels, containing carboxyl groups, that according to Freeman et al. (1983) are critical, are located on the soma in contrast to the
TTX insensitive Na+ channels, without carboxyl groups, located in the axon hillock. However, the present data does not allow us to make any con- clusion about the molecular organization of the Na+ channels in the Retzius nerve cells of the leech.
Acknowledgement-This work was supported by the Research Council of Serbia.
REFERENCES
Albuquerque E. X. and Thesleff S. (1968) A comparative study of membrane properties of innervated and chroni- cally denervated fast and slow skeletal muscle of the rat. Acta physiol. stand. 13, 471480.
Barker P. F., Hodgkin A. L. and Ridgway R. B. (1971) Depolarization and calcium entry in squid giant axons. J. Physiol. Land. 218, 709-755.
Beleslin B. B. (1968) Studija spontanih prainjenja poten-

328 B~GIJAN B. BELESLIN
cijala Siljaka Retziusovih ganghjskih kelija pijavica po- mocu mikroelektrode. (MSc. Thesis) Inst. za pato-fiziol. Med. fak. Beograd.
Beleslin B. B. (1971) Effects of different external media on the leech ganglion cells interaction. Period. hiol. 73, 63-61.
Beieslin B. B. (1977) Lack of electrotonic transmission between Retzius nerve cells in the fifth and sixth free ganglion of the horse leech Haemopis sanguisuga. Comp. Biochem. Physiol. 56A, 509-512.
Beleslin B. B. and Mihailovii- Lj. (1967) Effect of tetrodo- toxin on the spontaneous discharge of spike potentials of the nerve cells of Retzius in a segmental ganglion of Leech. Iu~o&x~. physiol. phurmacol. Acta 3, 85-86.
Bennett M.‘V. L. (i966) Physiology of electrotonic junc- tions. Ann. N. Y. Acad. Sci. 137. 509-539.
Deitmer J. W. and Schlue W. R. (1981) Measurements of the intracellular potassium activity of Retzius cells in the leech central nervous system. J. exp. Biol. 91, 87- 101.
Deitmer J. W. and Schlue W. R. (1983) Intracellular Na+ and Cal+ in leech Retzius neurones during inhibition of the Na+-K+ pump. P$tigers Arch. 397, 195-201.
Fatt P. and Ginsborg B. (1958) The ionic requirements for the production of action potentials in crustacean muscle fibres. J. Physiol., Land. 142, 516543.
Freeman S. E., Gray P. J. and Bladen M. P. (1983) Chemical modification of the sodium channel in the guinea pig atrium. Proc. Int. Union Physiol. Sci. 15, 385.08.
Geguldig D. and Gruener R. (1970) Voltage clamp of the Aplysia giant neurone: early sodium and calcium current. J. Physiol., Lond. 211, 211-244.
Hagiwara S. and Morita H. (1962) Electrotonic trans-
mission between two nerve cells in leech ganglion. .I. Neurophysiol. 25, 721-73 1,
Hagiwara S. and Naka K. I. (1964) The initiation of spike potential in barnacle muscle fibres under low intracellular Cazi. J. gen. Physiol. 48, 141-162.
Hodgkin A. L. and Huxley A. F. (1952) Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. .I. Physiol., Land. 116, 449-472.
Katz B. and Miledi R. (1967) Tetrodotoxin and neuro- muscular transmission. Proc. R. Sot. B 167, 8-22.
Keyser K. and Lent C. M. (1977) On the neuronal homol- ogies within the central nervous system of leeches. Camp. Biochem. Physiol. $%A, 285-297.
Kleinhaus A. L. and Prichard J. W. (1976) Sodium de- pendent tetrodotoxin resistant action potentials in a leech neuron. Brain Res. 102, 368-373.
Kleinhaus A. L. and Prichard J. W. (1983) Differential action of tetrodotoxin on identified leech neurons. Camp. Biochem. Physiol. 74C, 2 I I-2 18.
Kostyuk P. G. (1980) Calcium ionic channels in electrically excitable membrane. ~euroscielzce 5, 945-959.
Kostyuk P. G. and Krishtal 0. A. (1977) Aeparation of sodium and calcium current in the somatic membrane of mollusc neurones. J. Physiol., Lond. 270, 545-568.
Lent C. M. (1972) Electrophysiology of Retzius’ cells of segmental ganglia in the horse leech Haemopis marmorata (Say). Camp. Biochem. Physiol. 42A, 857-862.
Sperelakis N. and Shigenobu K. (1972) Changes in mem- brane properties of chick embryonic heart during devel- opment. J. gen. Physiol. 60, 430453.
Thomas R. C. and Meech R. W. (1982) Hydrogen ion currents and intracellular pH in depolarized voltage- clamped snail neurones. Nature, Lond. 299, 826828.