Embed Size (px)
Citation preview

Appl Microbiol Biotechnol (2005) 67: 322–335DOI 10.1007/s00253-004-1806-0
MINI-REVIEW
Sergi Maicas . José Juan Mateo
Hydrolysis of terpenyl glycosides in grape juice and other fruit
juices: a review
Received: 28 July 2004 / Revised: 30 September 2004 / Accepted: 19 October 2004 / Published online: 6 January 2005# Springer-Verlag 2005
Abstract The importance of monoterpenes on varietalflavour of must and other fruit juices has been reviewed.These compounds were mainly found linked to sugarmoieties in grape juice and wines, showing no olfactorycharacteristics. In this way, analytical techniques developedto study these compounds, in both free or glycosidicallyforms, are discussed. Mechanisms to liberate terpenes werestudied, making a comparative study between acidic andenzymic hydrolysis of terpene glycosides; as enzymichydrolysis seems to be the most natural way to liberateterpenes, the ability to use glycosidases from grapes, yeasts,bacterial or exogenous, i.e. fungal commercial preparations,were reviewed. Re-arrangements of terpenes after acidichydrolysis of glycoconjugated are discussed as well aspotential adverse effects of enzyme preparations.
Introduction
Research over the last decades has revealed that a greatnumber of plant-tissue flavour compounds are glycosi-lated and accumulate as non-volatile and flavourless gly-coconjugates (Stahl-Bishop et al. 1993; Winterhalter andSkouroumounis 1997; Mateo and Jimenez 2000). Althoughresults in literature had long suggested the occurrence ofglycosidically bound flavour compounds in plants, the firstclear evidence was found in 1969 by Francis and Allock in
rose (Francis and Allcock 1969). The work of Cordonnierand Bayonove (1974), suggesting the occurrence in grapesof monoterpenes (important flavour compounds) as glyco-conjugates on the basis of enzymatic works, was laterconfirmed by identification of glycosides (Williams et al.1982). These findings opened a new field of intensive re-search on the chemistry of glycoconjugated flavour com-pounds to exploit this important flavour source present inboth plants and fruit tissues.
Glycoconjugates of flavour compounds are present inseveral fruits such as grapes (Williams et al. 1982; Gunata etal. 1985), apricot (Krammer et al. 1991; Salles et al. 1991),peach (Krammer et al. 1991), yellow plum (Krammer et al.1991), quince (Lutz and Winterhalter 1992), sour cherry(Schwab et al. 1990), passion fruit (Chassagne et al. 1996;Winterhalter 1990), kiwi (Young and Paterson 1995),papaya (Heidlas et al. 1984; Schwab et al. 1989), pineapple(Wu et al. 1991), mango (Sakho et al. 1997), lulo (Suarez etal. 1991), raspberry (Pabst et al. 1991) and strawberry(Roscher et al. 1997). The occurrence of glycosidicallybound volatiles is typically two to eight times greater thanthat of their free counterparts (Gunata et al. 1985; Krammeret al. 1991). Moreover, most norisoprenoids in fruit, someof which are precursors of very potent flavour compounds,have been detected mainly in glycosidic forms. This, to-gether with the low aroma threshold and sensory propertiesof aglycones, makes the glycosidic compounds an im-portant potential source of flavour volatiles during fruitjuice processing.
Some aglycones are already odourous when releasedfrom glycosides. They can therefore contribute to the floralaroma of some wines (Mateo and Jimenez, 2000), grapes(Gunata et al. 1993), apricots (Chairote et al. 1981), peaches(Engel et al. 1988) and tea (Ogawa et al. 1997). This is thecase of monoterpenes such as geraniol, nerol and linalool,which possess mainly floral attributes and low odourthresholds (100–400 ppb, Rapp and Mandery 1986).
Terpene compounds belong to the secondary plantconstituents, the biosynthesis of which begins with acetyl-CoA (Manitto 1980; Fig. 1). Microorganisms are also ableto synthesize terpene compounds (Hock et al. 1984), but the
S. MaicasDepartment of Food Science and Technology,Universidad Cardenal Herrera-CEU,Seminari s/n,46113 Montcada, Spain
S. Maicas . J. Mateo J.Department of Microbiology and Ecology,Universitat de València,Dr. Moliner 50,46100, Burjassot, SpainTel.: +34-96-3983145Fax: +34-96-3983099e-mail: [email protected]
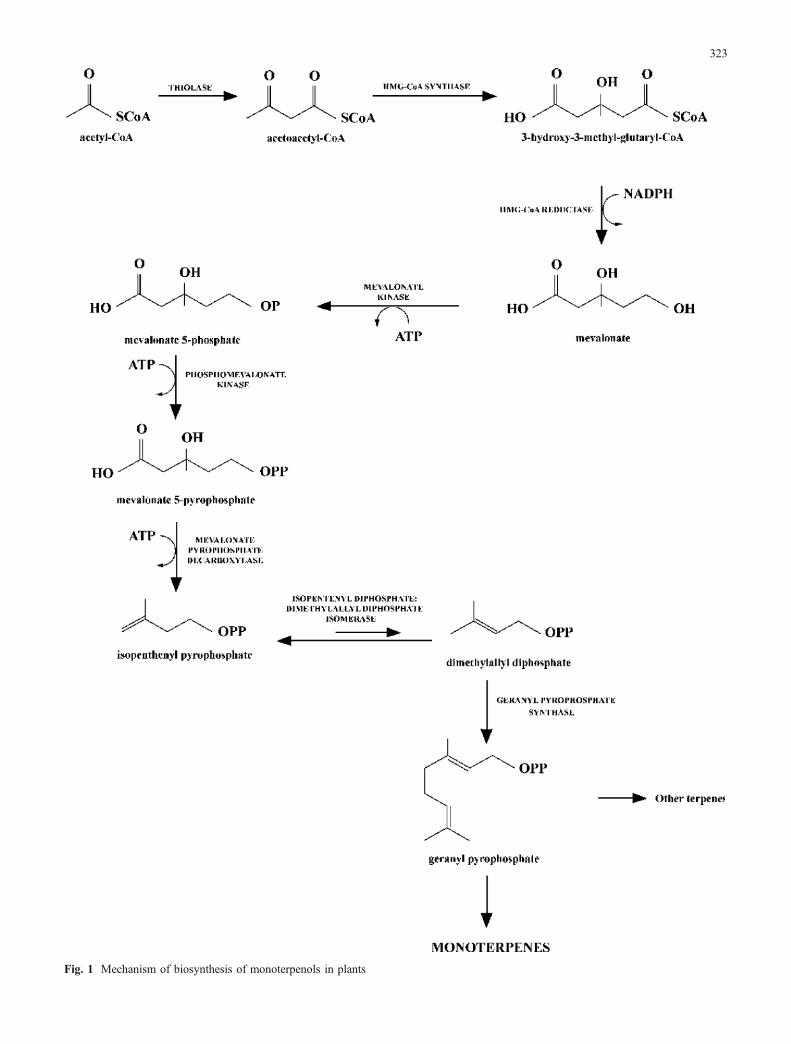
Fig. 1 Mechanism of biosynthesis of monoterpenols in plants
323

formation of terpenes by Saccharomyces cerevisiae has notyet been observed (Rapp and Mandery 1986). Severalauthors have shown that terpenes play a significant role inthe varietal flavour of wines by means of their transforma-tion to other compounds (Cordonnier and Bayonove 1974;Gunata et al. 1985; Wilson et al. 1986).
Classification of monoterpenes
Three types of categories of monoterpenes exist in planttissues, with some interrelationships among the catego-ries. On the top of the complex are the free aroma com-pounds, commonly dominated by linalool, geraniol andnerol, together with the pyran and furan forms of the lin-alool oxides. However, depending on how the juice hasbeen treated and on certain factors (which may include cli-mate), many additional monoterpenes can be found in thisgroup, i.e. citronellol, α-terpineol, hotrienol, nerol oxide,myrcenol, the ocimenols, plus several other oxides, alde-hydes and hydrocarbons. In wines, several monoterpeneethyl ethers and acetate esters have also been found amongthe free aroma compounds.
Second, there are the polyhydroxylated forms of themonoterpenes, or free odourless polyols. A most significantfeature of the polyols is that, although these compoundsmake no direct contribution to the aroma, some of them arereactive and can break down with great ease to give pleas-ant and potent volatiles, e.g. diendiol (3,7-dimethylocta-1,5-diene-3,7-diol) can give hotrienol and nerol oxide(Williams et al. 1980).
Third, there are the glycosidically conjugated forms ofthe monoterpenes which also make no direct contributionto the aroma of the grape. Glycosides are, in most cases,more abundant than the unglycosilated forms of individualmonoterpenes and polyols (Mateo and Jimenez 2000).
Structure of glycosides
Glycosidically bound volatiles identified in fruits and plantsare highly complex and diverse, especially the aglyconemoiety. The sugar parts consist of β-D-glucopyranosidesand different diglycosides: 6-O-α-L-arabinofuranosyl-β-D-glucopyranosides, 6-O-α-L-arabinopyranosyl-β-D-glu-copyranosides (vicianosides), 6-O-α-L-rhamnopyranosyl-β-D-glucopyranoside (rutinosides), 6-O-β-D-apiofuranosyl-β-D-glucopyranosides, 6-O-β-D-glucopyranosyl-β-D-glu-copyranosides and 6-O-β-D-xilopyranosyl-β-D-glucopy-ranosides (primeverosides) (Gunata et al. 1985; Vasserotet al. 1995). Figure 2 shows main glycosides found inMuscat grapes. In rare cases, trisaccaharide glycoconju-gates have been isolated (Winterhalter and Skouroumounis1997). The aglycon part is often formed with terpenols, butlinalool oxides, terpene diols and triols can also been found.However, other flavour precursors can occur such as linearor cyclic alcohols, e.g. hexanol, phenylethanol, benzyl al-cohol, C-13 norisoprenoids, phenolic acids and probably
volatile phenols such as vanillin (Fig. 3; Allen et al. 1989;Schwab et al. 1990; Sarry and Gunata 2004).
If we consider in grape juice, only glycosides with themost flavourant aglycons (i.e. with the same state of oxi-dation than linalool) are the most abundant are apiosylgly-cosides (up to 50% according to grape variety), followed byrutinosides (6–13%) and finally, glucosides (4–9%). Amore accurate analysis indicates that all glycosides are notpresent in all cultivars, and that their proportions also differaccording to grapes (Bayonove et al. 1993). The glycosideflavour potential from grapes remains, unfortunately, quitestable during winemaking and in young wines as well.
Isolation of glycosides
Isolation of glycosides by selective retention of the com-pounds on a solid-phase adsorbent is a commonly usedtechnique. By washing the adsorbent with water followingthe adsorption step, free sugars and other polar constituentscan be removed, while the less polar glycosides are retained.Elution with an organic solvent then gives the glycosidicfraction. Different methodologies have been proposed toextract glycoside precursors from grape juices and wines.Williams et al. (1982) used glass column chromatographycontaining C-18 reversed-phase adsorbent to extract glyco-sides from juice or de-alcoholised wine. After washing withwater and eluting free compounds with 20% aqueous aceticacid, precursors were eluted in two fractions with 30%aqueous acetic acid and methanol. A modification of themethodology has been proposed by using 1 g solid phaseextraction C-18 cartridges: hydrophylic compounds wereeluted with water, free terpenes with dichloromethane andglycosides with methanol (Di Stefano 1991). This methodhas been improved in the last years, but it has a disadvan-tage, because separation is different depending on thecommercial origin of the cartridges (R. Di Stefano, personalcommunication). A second approach to the problem hasbeen proposed by Montpellier researchers by using Amber-lite XAD-2 resin, because it possesses an excellent capacityfor adsorption of free terpenols from grape juice (Gunataet al. 1985). This resin had been previously used to isolatenaringin and limonin from grape juices. Once wine has beeneluted through resin, free compounds were eluted withpentane and glycosides with ethyl acetate. Amberlite XAD-2 resin displays extraction capacities similar to those ofcoated octadecyl silica. Furthermore, it has the advantageof being sold in large particle sizes; it is thus possible touse it in a wide preparative column at atmospheric pres-sure. A modification of this method has been suggested,and free compounds were eluted with pentane:dichlor-omethane to improve extraction (Di Stefano 1989). Never-theless, even with extensive washing of the adsorbentbeds, both Amberlite XAD-2 and Amberlite XAD-16 (theAmberlite XAD-16 resin has a surface area and capacitygreater than that of Amberlite XAD-2) retained free glu-cose. Both of these adsorbents had the disadvantage ofretaining free glucose in addition to the adsorbed glyco-sides (Williams et al. 1995).
324

Reversed-phase silica gel has been found to be a par-ticularly suitable adsorbent for the isolation of glycosidicterpenes (Williams et al. 1995). The commercial availabilityof this adsorbent in uniform, pre-packed cartridges was anadditional advantage in development of the isolation step.
A method has been proposed to extract glycosilat-ed flavour precursors from grape juices by microwaves(Bureau et al. 1996). This method was compared to themethod using a column of Amberlite XAD-2 resin. Al-though a further purification of the extracts was required,this method provided the advantages of rapidity, ease ofextraction and possibility of extract berries without de-seeding or crushing.
Glycosides fractionation
Williams et al. (1982) laid stress on the existence, in thearomatic varieties of grapes, of two classes of precursors ofC-10 and C-13 components with different polarity, separ-able by chromatography on C-18 cartridges. The methodfrom these authors described of the separation of twoglycosides classes, the absorption of de-alcoholised wineon C-18 cartridges of adequate size to the wine volumeused, the elimination of hydrophylic components withwater, the elution of more polar precursors with 30% aceticacid and the elution of monohydroxilated terpenic alcoholswith methanol. The components in both fractions wererecuperated by evaporation of solvent and subjected toanother purification or hydrolysis.
Fig. 2 Main monoterpenecompounds in grape juice andwines. Numbers like Table 1
325

The critical points of this method are: (1) the incompleteelimination of volatile components during the evaporationof wine alcohol under vacuum and (2) the partial recuper-ation of the precursors with an acidic solvent, with theopportunity of hydrolytic adulterations and chemical trans-formations during the solvent evaporation.
In addition, Strauss et al. (1988) fractionated by dopletcountercurrent chromatography the C-10, C-13 and aro-matic ring precursors in must previous isolation on C-18 RPcolumn, and Bitteur et al. (1989) described an analyticalHPLC method with C-18 column to fractionate aromaprecursors, using acetonitrile and water as solvents.
Following developments of chromatographic techniquesin totally liquid phase (Skouroumounis and Winterhalter1994) led to the fractionation of the precursors in leaves ofRenan Riesling by using multilayer coil countercurrentchromatography. In this case, the precursors were pre-viously isolated on Amberlite XAD-2 resin column. Thistechnique has been used to isolate two novel terpenoidglucose esters from Riesling wines (Bonnlander et al.1998).
Thinking it helpful to also bring the fractionation tech-niques into laboratories without instruments for chro-matography in totally liquid phase, an attempt to create aseparation system for volatile precursors, using commer-cial C-18 RP columns and inert solvents, has been per-
formed. The glycosides of mono-, di- and trihydroxilatedterpene and norisoprenoid alcohols, as well the relatedshikimate pathway ones, have been isolated on C-18 RPcartridges and fractionated in classes having different po-larity, with eluents at increasing percentages of methanol.The benzyl alcohol glycosides appear the most polar, whilethose of terpene monohydroxilated alcohols and of ge-ranic acid the least polar. The terpene diols, linalool furanoidand pyranoid oxides, as well norisoprenoid precursors, showintermediate polarity and place themselves in well-fixedfractions according their polarity (Mateo et al. 1997).
Acidic hydrolysis of terpene glycosides
Acidic hydrolysis of terpene glycosides can induce a mo-lecular rearrangement of the monoterpenols, which aretransformed in other compounds. Nevertheless, these waysto liberate terpenes simulate the reactions taking placeduring ageing of wines, and the different terpenic alcoholswere produced in similar quantitative rations.
Experiments on both whole juice and monoterpeneglycosides isolated from juice have demonstrated thatsignificantly different patterns of volatile monoterpenes areproduced when each is hydrolysed at different pH values.Furthermore, there appears to be a pH-dependent inter-
Fig. 3 Structure of some gly-coconjugated aroma compounds
326

relationship among several of the grape monoterpenes.Thus, e.g., the isomerics ocimenols appear to be formedhydrolytically in juice at pH 1 at the expense of linalool,nerol and geraniol, the last three compounds being pH 3products (Williams et al. 1982).
For acidic hydrolysis, samples were dissolved in tartratebuffer at pH 3.2 or water to which perchloric acid was addedto give pH of 1.0. After removing any residual volatilecompound remaining, solutions were heated on a steambath for 15 min, cooled and resulting free volatile com-pounds extracted and analysed by GC. Mateo et al. (1997)used tartaric acid 5 g l−1, NaCl 10% with a pH of 3.0(adjusted with 1M NaOH) to dissolve samples and thenheated them for 60 min in a boiling-water bath. R. DiStefano (personal communication) used H2SO4 0.1 N toadjust the pH to 1.0 and heated te solutions for 30 min in aboiling-water bath.
Significant differences can be seen in the patterns ofvolatile monoterpenoids produced from material by hy-drolysis at pH 1.0 and pH 3.0. It is apparent that the moreacidic conditions bring out extensive rearrangements ofmonoterpenoids (Williams et al. 1982). On hydrolysis at pH3.0, linalyl, geranyl and nerylβ-D-glucosides each gave thesame major products, linalool and α-terpineol. Addition-ally, several compounds were observed in the hydrolysis of
linalyl and geranyl glucosides that were not given by theneryl derivative, as geraniol, (E)- and (Z)-ocimene, α-ter-pinene and myrcene. α-Terpenyl β-D-glucoside gave noacyclic monoterpenes, and α-terpineol was its predominanthydrolysis product. In contrast with the reaction at pH 3.0,the precursor material at low pH gave little linalool, andno geraniol or nerol was detected. While α-terpineol wasstill a significant product at pH 1.0, it was now accom-panied by many other monoterpenoids not given by theprecursors at pH 3.0 (Williams et al. 1982).
Acid hydrolysis products are not diagnostic of the mono-terpene aglycon composition of the grape precursors. Thus,while the grape glycosides are made up predominantly ofgeranyl, linalyl and neryl derivatives and only trace quan-tities of α-terpenyl glycosides, the hydrolysis products atpH 3.0 are dominated by linalool and α-terpineol, withgeraniol relatively less abundant.
Most of the compounds given under hydrolytic condi-tions at pH 3.0 are the free terpenes of the juice (Cordonnierand Bayonove 1974; Sarry and Gunata 2004). However,many of those products developed at pH 1.0 were seen inthe headspace of Muscat juice heated for 15 min at 70°Cand pH 3.2 (Williams et al. 1982). It has been observed thatprolonged heating of juice at pH 3.0 ultimately altered thesensory character by imparting a eucalyptus-like aroma,
Fig. 4 Acid catalysed re-arrangement of monoterpenes
327

attributable to the presence of excessive quantities of 1,8-cineole in the headspace composition of the juice. Theoccurrence of pH 1.0 products 1-terpineol, 4-terpineol, β-terpineol and myrcenol in cognac can also be accounted forby hydrolytic degradation of grape monoterpene glyco-sides during wine distillation.
The content of various monoterpene components changeduring bottle storage or rather during the maturation of wineby means of acid-catalysed reactions (Rapp and Mandery1986; Di Stefano 1989). In the case of several components(e.g. linalool, geraniol, hotrienol and isomers of linalooloxide), an obvious decrease in concentration can be ascer-tained during the course of ageing. In addition to this, com-pounds are formed which are not present in young wines:among others the formation of cis- and trans-1,8 terpine.During the maturation of wine, monoterpene alcohols canbe formed from linalool. According to these results, linaloolis transformed in an aqueous acid medium toα-terpineol bycyclation, to hydroxy-linalool through hydration in theseventh position and to geraniol and nerol by a nucleophilic1,3-transition (allyl) (Fig. 4) (Rapp and Mandery 1986).
Enzymic hydrolysis of terpene glycosides
Terpene glycosides can also be hydrolysed by an enzymicway, a more interesting way because it produces a more‘natural’ flavour in the wine (Cordonnier and Bayonove
1974; Gunata et al. 1985; Mateo and Jimenez 2000; Gunata2002). The glycosidase flavour potential from grape re-mains, unfortunately, quite stable during winemaking andin youngwines as well. So, to enrichwine flavour by releaseof free aromatic compounds from natural glycoside precur-sors, particularly pathways are required. Mainly, enzymichydrolysis of glycosides is carried out with various en-zymes which act sequentially according to two steps: firstly,α-L-rhamnosidase, α-L-arabinosidase or β-D-apiosidasemake the cleavage of the terminal sugar and rhamnose,arabinose or apiose and the corresponding β-D-glucosidesare released; subsequently liberation of monoterpenol takesplace after action of a β-D-glucosidase (Fig. 5; Gunata etal. 1988). Nevertheless, one-step hydrolysis of disaccha-ride glycosides has also been described; enzymes catalys-ing this reaction have been isolated from tea leaves (Ogawaet al. 1997) and grapes (Gunata et al. 1998). This one stepreaction occurs through the cleavage of the aglycone link-age which yields a disaccharide and aglycone, the identityof which have been confirmed by HPLC and GC/MS(Ogawa et al. 1997; Gunata et al. 1998) (Fig. 6).
Studies regarding the mechanism of the enzymes whichparticipate in the enzymic hydrolysis of terpene glyco-sides, individual or sequentially, have been made onglycoside fraction of the grapes or on different syntheticglycosides.
Enzymic hydrolysis of glycoside extracts from Muscat,Riesling, Semillon, Chardonnay, Sauvignon and Sirah
Fig. 5 Sequential enzymatic hydrolysis of disaccharidic flavour precursors
328

varieties have provoked the liberation not only of terpenes,but also C-13 norisoprenoids such as 3-oxo-α-ionol and 3-hydroxy-β-damascenona (Gunata et al. 1990a). Thesecompounds are totally glycosilated in the grape and, asopposed to terpenes, they are found in the same quantities inall the grape varieties, aromatics or neutral, and they arecapable of awarding certain typicity to the wine flavour,because they have lower threshold values than terpenes, andthey contribute characteristic aromatic features (Razungleset al. 1987).
Plant glycosidases
Most work has been devoted to grape glycosidases, becausethe first evidence of multiple forms of glycosidic flavourprecursors was found in this fruit. The potential use ofenzymic systems of grapes to liberate terpenes from grapejuices or wines has been always the subject of differentworks regarding the enzymic hydrolysis of terpene glyco-sides.
Grapes have an enzyme with β-glucosidase activity(Table 1; Aryan et al. 1987; Gunata et al. 1990a,b; DiStefano and García 1995) but only low α-rhamnosidase, α-arabinosidase or β-xylosidase activities have been detected(Gunata et al. 1990a); the presence of β-apiosidase ingrapes has not yet been confirmed (Gunata 2002). On the
other hand, grape β-glucosidase was not quite stable andshowed low activity at grape juice or wines pH values(Lecas et al. 1991).
β-Glucosidase was isolated from mature grapes (cv.Muscat of Alexandria) and partially purified by chroma-tography on Ultrogel AcA 44 and DEAE-Sepharose CL-6B. Two distinct proteins with β-glucosidase activity wereisolated but, except for their mass, no differences occurredin other biochemical characteristics; the optimum pH ofactivity was 5.0 and the optimum temperature 45°C. Fornatural substrates, the two enzymes had a high affinity forgeranyl β-D-glucoside and were inhibited by glucose,gluconolactone, Ca2+, Cu2+ and p-chloromercuribenzoate(Lecas et al. 1991). Similarly, certain β-glucosidase activityhas been observed in grape leaves (Biron et al 1988; Gunataet al. 1990a; Di Stefano and García 1995).
In general, β-glucosidases with a vegetal origin show alow activity on monoglucosides of terpenes with a tertiaryalcohol group (linalool, α-terpineol), and they are onlycapable to hydrolyse monoglucosides of terpenes with aprimary alcohol group (geraniol, nerol, citronellol) (Aryanet al. 1987; Gunata et al. 1990a).
Endoglycosidase, active on disaccharidic flavour pre-cursors in fruits, has been isolated and partiatilly character-ized from grape skins (Gunata et al. 1998). Structure of thisenzyme differs from that obtained from tea and could be amembrane bound protein. Moreover, grape endoglycosi-
Fig. 6 GC-MS chromatogram of terpenes obtained by enzymic hydrolysis of glycosides. Numbers like Table 1
329

dase also seems to possess β-glucosidase activity (Gunataet al. 1998).
Microbial glycosidases
Glycosidases with oenological implications have beenwidely reported in distant taxomical species such as yeasts,bacteria and fungi (Grossmann et al. 1987; Gunata et al.1990b; Boido et al. 2002).
Yeast glycosidases
Yeasts of the Hansenula species isolated from ferment-ing must were reported to have an inducible β-glucosi-dase activity, but this enzyme was inhibited by glucose(Grossmann et al. 1987). Other yeast strains such as Can-dida molischiana (Gonde et al. 1985) and C. wickerhamii(Leclerc et al. 1984) also possess activities towards variousβ-glucosides, and they were little influenced by the nature
of aglycon (Gunata et al. 1990b). β-Glucosidase fromC. molischiana was immobilized to Duolite A-568 resin,showing similar physicochemical properties to those offree enzyme. The immobilized enzyme was found to bevery stable under wine conditions and could be used re-peatedly for several hydrolyses of bound aroma (Gueguenet al. 1997). Endomyces fibuliger also produces extra-cellular β-glucosidase when grown in malt extract broth(Brimer et al. 1998).
Screening 370 strains belonging to 20 species of yeasts,all of the strains of the species Debaryomyces castelli, D.hansenii, D. polymorphus, Kloeckera apiculata and Han-senula anomala showed β-glucosidase activity (Rosi et al.1994). A strain of D. hansenii exhibited the highestexocellular activity, and some wall-bound and intracellularactivity and its synthesis, occurring during exponentialgrowth, was enhanced by aerobic conditions and repressedby high glucose concentration. The optimum condition forthis enzyme was pH 4.0–5.0 and 40°C. This enzyme wasimmobilized using a one-step procedure on hydroxyapa-tite. The immobilized enzyme exhibited a lower activity
Table 1 Results obtained byhydrolysis of terpene glyco-sides treated with differentenzymatic preparations. Dataare normalized to 100 inuntreated wines
No. Terpenes Grapeskin
Saccharomycescerevisiaeglucosidase
Klerzyme200 Exogenousglycosidases
Hemicellulase
PectinolC
RohapectC
Sweetwine
Drywine
1 trans-Furan linalooloxide
108.2 122.9 247.3 174.3 943.1 109.7 129.7
2 cis-Furanlinalool oxide
124.3 296.8 149.2 133.8 456.1 87.0 117.6
3 Linalool 101.2 204.6 113.6 111.0 187.8 107.5 108.84 Hotrienol 93.3 113.2 130.8 87.15 Neral6 α-Terpineol 230.0 150.8 190.8 113.8 141.77 Geranial8 trans-Pyran linalool
oxide124.9 109.3 109.1 113.1 104.2 104.4
9 cis-Pyran linalooloxide
104.5 >88.9 136.3 111.4 98.0
10 Citronellol 177.6 176.6 551.9 239.8 131.8 146.5 335.411 Nerol 593.0 847.6 424.5 720.0 1,173.2 190.6 642.012 Geraniol 381.9 833.4 491.8 633.2 806.2 107.0 359.113 Diol I 104.2 91.4 151.9 112.7 152.3 113.8 98.014 Endiol 133.015 Diol II 101.4 112.5 174.7 114.0 124.3 100.1 97.616 Hydroxy-cityronellol 152.6 159.0 178.4 552.917 8-Hydroxy-dihydro-
linalool275.9 230.8 152.9 397.6 105.9 93.2
18 Hydroxy-nerol 141.419 trans-8-Hydroxy-
linalool133.4 466.0 1,343.0 431.5
20 Hydroxy-geraniol 140.2 475.021 cis-8-Hydroxy-
linalool271.8 688.9 4,083.0 990.0 916.9
22 Geranic acid 362.6 522.8 410.223 Triol
330

than the purified free enzyme, but was much more stablethan the enzyme in cell-free supernatant (Riccio et al.1999). These studies have shown the ability of severalwine yeasts to hydrolyse terpenoids, norisoprenoids andbenzenoids glycosides; among wine yeasts Hanseniasporauvarum was able to hydrolyse both glycoconjugated formsof pyranic and furanic oxides of linalool (Fernandez et al.2003) Other authors have also shown the important role ofnon-Saccharomyces species in releasing glycosidid boundfraction of grape aroma components (Esteve et al. 1998;Mendes et al. 2001).
Finally, the situation regarding S. cerevisiae is morecomplex, because this yeast is capable of modifying theterpenic profile of the wine; thus, it can produce citronellolfrom geraniol and nerol; the intensity of this transformationdepends on the yeast strain used (Dugelay et al. 1992;Hernandez et al. 2003). Other authors propose a morecomplex scheme: geraniol was transformed by these yeastsinto geranyl acetate, citronellyl acetate and citronellol,while nerol was transformed into neryl acetate; in addition,geraniol was transformed into linalool and nerol wascyclized to α-terpineol at must pH (Di Stefano et al. 1992).
Few data are available regarding glycosidase activitiesof oenological yeast strains and the technological proper-ties of the enzymes. Low α-rhamnosidase, α-arabinosidaseor β-apiosidase activities were detected in S. cerevisiae(Delcroix et al. 1994). Nevertheless, data on β-glucosidaseactivity on Saccharomyces are contradictory. First resultsshowed that these yeasts had a very low activity (Gunataet al. 1990c), but Delcroix et al. (1994) found three eno-logical strains showing high β-glucosidase activity. Onthe other hand, Darriet et al. (1988) have shown thatoxidases located in the periplasmic space of a strain of S.cerevisiae were able to hydrolyse monoterpene glucosidesof Muscat grapes; they found also that the activity of thisβ-glucosidase was glucose independent. Mateo and DiStefano (1998) have detected β-glucosidase activity indifferent Saccharomyces strains on the basis of its hydro-lytic activity on para-nitrophenyl-β-D-glucoside (pNPG)and terpene glucosides of Muscat juice (Table 1). Thisenzymic activity is induced by the presence of bound β-glucose as a carbon source in the medium and seems to bea characteristic of the yeast strain. This β-glucosidase isassociated with the yeast cell wall is quite glucose in-dependent but is inhibited by ethanol. These results couldopen new pathways regarding other glycosidase activitiesin S. cerevisiae; α-rhamnosidase, α-arabinosidase or β-apiosidase activities could be induced in wine yeast bychanging the composition of the medium including induc-tive compounds, as well as in filamentous fungi (Shoseyovet al. 1990; Dupin et al. 1992).
Bacterial glycosidases
Although little information is available regarding the β-glucosidase activity of lactic acid bacteria (LAB) involvedin wine-making, Oenococcus oeni is the species generallyrecognized as beneficial for final aroma compound
(Henick-Klling 1995; Maicas et al. 1999). This bacteriumis involved in the malolactic fermentation (MLF), theconversion of the malic acid in wine to lactic acid andcarbon dioxide. This reaction, generally associated to afinally equilibrated good wine (Fleet 1993), can also beperformed by other LAB (Lactobacillus and Pediococcus),although it is usually performed by O. oeni (Davis et al.1985).
It has been demonstrated that O. oeni was able to cleavethe glucose moiety from the major red wine anthocyanin,malvidin-3-glucoside, to use it as a carbon source. Releaseof glycosylated volatile precursors in Tannat wine was alsoobserved (Boido et al. 2002), and only minimal O. oeniglycosidase activity was noted in Viognier glycosidic ex-tracts (McMahon et al. 1999).
The effective β-glucosidase activity of some strains ofO. oeni has been recently reported (Grimaldi et al. 2000;Boido et al. 2002; Barbagallo et al. 2004; D’Incecco et al.2004). Mansfield et al. (2002) have also detected theproduction of β-glucosidase enzymes in several strains ofO. oeni, although cultures of the same strains failed tohydrolyse native grape glycosides.
The study of the hydrolysis of wine aroma precursors(linalool, α-terpineol, nerol and geraniol) during MLF hasbeen carried out with someO. oeni starter cultures in modelwine solutions by Ugliano et al. (2003). The liberation ofglycosidically bound aroma compounds was assessed forvarious strains, which also performed the MLF. Althoughthe quantity of released precursors was strain dependent, thelarge release of glycosylated aroma compounds observedduring their experiments suggests that O. oeni can activelycontribute to the changes of sensory characteristics of wineafter MLF through the hydrolysis of aroma precursors.
Fungal glycosidases
Taking into account that enzymic systems of grapes are notsuitable to hydrolyse terpene glycosides in grape juice orwine and that more studies are needed regarding the abilityof S. cerevisiae and lactic acid bacteria to produce all theenzymes which take part in this process, several exogenousenzymes, mainly with a fungal origin, have been developedto liberate terpenes in wines. They are the so-called ex-ogenous (fungal) glycosidases. Comparing 34 differentenzymic preparations, the most suitable ones to be usedduring winemaking process are those which posses allβ-D-glucopyranosidase, α-L-arabinofuranosidase, α-L-rhamno-pyranosidase and β-D-apiofuranosidase activities (Table 1;Cordonnier et al. 1989).
With regard to the oenological uses of exogenous en-zymes, the non-selectivity of the glycosidases in RohapectC, together with its relative tolerance to ethanol, suggest apossible application in releasing any glycosidically boundterpenes present in wines. But even for this application, thehigh pH values required for the enzymes in this prepara-tion (5.0–6.0) to function effectively limits their practicalutility. The great sensitivity to glucose of β-glucosidase ofRohapect C and several other commercially non-plant gly-
331

cosidases precludes any application of these enzymes injuice processing (Aryan et al. 1987).
Aspergillus niger β-glucosidases from two differentcommercial preparations do not show the same hydrolysisrates towards linalyl β-D-glucosides. After 24 h of incu-bation, β-glucosidase from the Hemicellulase preparation(Gist Brocades, France) hydrolysed the substrate com-pletely, whereas β-glucosidase from the Pectinol VP prep-aration (Röhm, Germany) hydrolysed only one third of it.α-Terpenyl β-D-glucoside was not a substrate for A. nigerβ-glucosidase from Pectinol preparation; however, thissubstrate was hydrolysed by A. niger β-glucosidase fromHemicellulase preparation. Such variations in both com-mercial enzymic preparations are probably due to the useof different strains of A. niger (Gunata et al. 1990b).
The ability of purified enzymes, synthetic substrates andsuitable analytical methods are needed for the rigorousprogress in the field of enzymic hydrolysis of terpenyl gly-cosides (Williams et al. 1982; Gunata et al. 1988; Bitteuret al. 1989; Voirin et al. 1990, 1992; Skouroumounis et al.1995). Enzymes have been isolated from fungal enzymicpreparations, vegetal extracts or synthetic culture mediainoculated with fungal cultures and have been purified bydifferent chromatographic techniques (gel filtration, ion-exchange chromatography, affinity chromatography, chro-matofocusing).
β-Apiosidase has been partially purified by filtrationchromatography on Ultrogel AcA 44 and ion exchangechromatography on DEAE-Sepharose CL-6B from A. nigercultures. The sugar linkage (1→6) of apiosylglucosides ofnerol, geraniol and pyran linalool oxide was cleaved by theenzyme with liberation of the corresponding monogluco-sides. Furthermore, apiosylglucosides of furan linalooloxide did not appear to be a good substrate forβ-apiosidase,unlike other apiosylglucosides. The production of theenzyme by A. niger is inducible, as it is only produced whenapiin (2-O-β-D-apiofuranosyl-β-D-glucoside of apigenin)was present in the culture medium as the carbon source. Inthe same way, peptone (2% w/v) and Tween 80 (0.15% v/v)activate β-apiosidase synthesis. With regard to the activity,the optimum pH and temperature were, respectively, 5.6and 50°C, and different ions have inhibitory effects on β-apiosidase activity. Conversely, the enzyme was inhibitedneither by glucose nor by ethanol (Dupin et al. 1992).
An α-L-arabinofuranosidase from A. niger was purifiedfrom a commercial crude preparation of HemicellulaseREG2 by gel filtration on Ultrogel AcA 44, ion exchangechromatography on DEAE-Sepharose CL-6B and affinitychromatography on Concanavalin A-Ultrogel AcA 22.The optimum pH of the enzyme was 3.9, and thetemperature of maximal activity 60°C. This arabinosidasewas active against monoterpenyl α-L-arabinofuranosyl-glucosides from grapes by liberating monoterpenyl β-D-glucosides and arabinose regardless the structure of theaglycon moiety (Gunata et al. 1990b). This enzyme hasbeen immobilised on chitosan by conjugation; this methodproved to be the better one as it ensures good biocatalyst
activity and better stability than the reported for the freeenzyme (Spagna et al. 1998).
An α-L-rhamnopyranosidase from a naringinase com-mercial preparation from Penicillium sp. and freed fromcontaminating β-D-glucosidase activity by chromatofocus-ing on PBE 94 using a pH gradient. Rhamnosidase isoen-zymes were eluted at pH 6.2 and 5.7, respectively (Gunataet al. 1988).
A strain of A. niger excreted into the medium a β-glu-cosidase which was partially purified by affinity chroma-tography. The enzyme was found to hydrolyse naturalglycosides and was competitively inhibited by glucose. ThepH optimum was 3.4, and ethanol enhanced the activity.Immobilization of fungus using Ca2+ alginate beads en-abled an increase in enzyme production in a continuousfermentation (Shoseyov et al. 1988). Later, an endo-β-glucosidase fom A. niger was immobilized to acrylic beads(Shoseyov et al. 1990) or to γ-alumina activated withdodecamethylendiamine and glutaraldehyde in sequence(Fu-Mian et al. 1994). GC-MS analysis of the wines in-dicated that the enzyme treatment increased concentrationsof free monoterpene alcohols.
Aspergillus oryzae was found to secrete two distinctβ-glucosidases when it was grown in liquid culture. Themajor form was highly inhibited by glucose, but the minorform, which was induced on quercitin, exhibited hightolerance to glucose or gluconolactone inhibition. The en-zyme was optimally active at 50°C and pH 5.0. It exhibitsexoglucanase activity and was able to release flavour com-pounds (Riou et al. 1998). Similar results were obtained inA. niger (Gunata and Vallier 1999).
These enzymes are present only in low quantities in themajority of commercial fungal enzymic preparations, main-ly regarding β-D-apiosidase activity (Dupin et al 1992).
Determination of free volatile compond terpenols, nori-soprenoids and volatile phenols indicates that concentrationin enzyme-treated wines highly increase, not only in aro-matic varieties, but also in neutral ones. Enhancementswidely varied according to the original grapes, from 265%to 2,000% (Gunata et al. 1990c).
Nevertheless, multiple forms of glycosidases could bepresent in an enzymic preparation. So, chromatofocusingon PBE 94 of a crude enzyme preparation from A. nigershowed the presence of multiple forms of β-apiosidase, β-glucosidase, α-rhamnosidase and α-arabinosidase (Gunataet al. 1997).
Enzymes are effective during some days from theiraddition to the wine, while glycosidic precursors remainalmost unchanged along fermentation. The method usedactually to enrich wine in volatile compounds is onlyeffective when dry wines are used, because β-glucosidasein fungal enzymic preparations is hardly inhibited by glu-cose. The hydrolysis of terpene glycosides is not completedin sweet wines obtained fromMuscat de Frontignan grapes,breaking in the monoglucoside stay due to inhibition ofβ-glucosidase by the presence of glucose (Gunata et al.1990b).
332

Potential adverse effects of enzyme preparations
When enzymic preparations have been used to improve thearomatic characteristics of the wines, undesirable odourshave been sometimes detected, even if liberation of ter-penes has been produced. These wines show high concen-trations in vinyl-phenols (4-vinyl-phenol, 4-vinyl-guayacol),reaching concentrations up to 1 mg l−1. The presence ofthese compounds is due to the action of two different enzy-mic systems. Firstly, a cinnamyl esterase from the enzymicpreparation hydrolyses cinnamic esters, the main phenolicderivatives in grape juices; so, the medium enriches in p-coumaric and ferulic acids, which are subsequently de-carboxylated by yeast cinnamate decarboxylase producingvinyl-phenols. It had been impossible to explain the for-mation of these undesirable compounds in wines when en-zymic preparations were used in winemaking (mainly forclarification processes) until this conjugate action of en-zymes was discovered. Based on these data, industries havedeveloped new fungal enzymic preparations with no cin-namyl esterase activity (Gunata 1993; Sefton and Williams1994).
Hydrolytic activity of fungal enzyme preparations to-wards anthocyanins was reported as early as 1955 (Huang1955). Enzymic activities from commercial preparationsmay potentially degrade anthocianins, composed of antho-cyanidin glycosilates (Yang and Steele 1958; Blom 1983).The de-coloration effect of these enzymes has been reportedfor raspberry (Jiang et al. 1990), strawberry (Rwahahizi andWrolstad 1988), blackberry (Yang and Steele 1958) andgrape anthocyanins (Fu-mian et al. 1994; Wightman et al.1997). So, the choice of enzyme preparations for flavourenhancement in fruit juices and derived beverages shouldbe made carefully. Glycosidases that are highly active onanthocyanins may have specific applications for the de-coloration of some orange juice or wines containing lowlevels of anthocyanins pigments.
On the other hand, fungal enzyme preparations generateoxidation artefacts during the hydrolysis of glycosides(Sefton and Williams 1994); this fact implies that, perhaps,the review of all the information regarding the use of theseenzymic preparations will be necessary.
Trends and prospects
The presence of terpenes, in their different forms, in grapejuices and wines represents an enormous potential in a wayto increase the varietal characteristics of the wines, contri-buting the final product with higher fruit-like character-istics. Actually, researchers have sufficient information andtools to study the presence of terpenes and their evolutionin grape juices and wines, but it is not yet possible totranslate all of the acquired knowledge to the wineries,because an efficient methodology to improve the terpenecontent of all the wines present in the market has not beenfound.
Several approaches through genetic engineering tech-niques will doubtless be used in the near future to overcome,
by using recently discovered glucosidases, the limitationson the exploitation of the glycosylated aroma source in theprocessing of fruit juices and derived beverages. On theother hand, construction of fungal strains expressing theseproteins involved in flavour liberation may enable theproduction of tailor made glycosidases without undesir-able activities.
So, great work remains to be made in a way to obtain amethodology, probably of enzymic nature, which allowsthat wine-consumer sense of smell can, at last, appreciatethe whole organoleptic richness of the product that he hason his hand or in his mouth.
Acknowledgements This work was supported by Conselleria deCultura, Educació i Esport, Generalitat Valenciana (Spain) projectGV04B-175.
References
Allen MS, Lacey MJ, Brown WV, Harris RLN (1989) Sauvignonblanc varietal aroma. Aust Grapegrower Winemaker 292:51–56
Aryan AP, Wilson B, Strauss CR, Williams PJ (1987) Theproperties of glycosidases of Vitis vinifera and comparison oftheir β-glucosidase activity with that of exogenous enzymes.An assessment of possible applications in enology. Am J EnolViticult 38:182–188
Barbagallo RN, Spagna G, Palmeri R, Restuccia C, Giudici P (2004)Selection, characterization and comparison of β-glucosidasefrom mould and yeasts employables for enological applications.Enzyme Microbiol Technol 35:58–66
Bayonove C, Gunata Y, Sapis JC, Baumes RL, Dugelay I, Grassin C(1993) L’aumento degli aromi nel vino mediante l’uso deglienzimi. Vignevini 9:33–36
Biron C, Cordonnier C, Glory O, Gunata Z, Sapis JC (1988) Etudedans le raisin de l’activité β-glucosidase. Conn Vigne Vin22:125–134
Bitteur S, Gunata Z, Brillouet JM, Bayonove C, Cordonnier R(1989) GC and HPLC of grape monoterpenyl glycosides. J SciFood Agric 47:341–352
Blom H (1983) Partial characterization of a thermostable anthocya-nin-β-glycosidase from Aspergillus niger. Food Chem 12:197–204
Boido E, Lloret A, Medina K, Carrau F, Dellacasa E (2002) Effect ofβ-glucosidase activity of Oenococcus oeni on the glycosylatedflavor precursors of Tannat wine during malolactic fermenta-tion. J Agric Food Chem 50:2344–2349
Bonnlander B, Baderschnider B, Messerer M, Winterhalter P (1998)Isolation of two novel terpenoid glucose esters from Rieslingwine. J Agric Food Chem 46:1474–1478
Brimer L, Nout MJR, Tuncel G (1998) Glycosidase (amygdalaseand linamarase) from Endomyces fibuliger (LU677): formationand crude enzyme properties. Appl Microbiol Biotechnol 49:182–188
Bureau S, Razungles A, Baumes R, Bayonove C (1996) Glycosy-lated flavor precursor extraction by microwaves from grapejuice and grapes. J Food Sci 61:557–561
Chairote G, Rodriguez F, Crouzet J (1981) Characterization ofadditional volatile flavor components of apricot. J Agric FoodChem 46:1898–1901
Chassagne D, Crouzet JC, Bayonnove CL, Brillouet JM, BaumesRL (1996) 6-O-α-D-Arabinopyranoyl-β-D-glucopyranosidesas aroma precursors from passion fruit. Phytochemistry 41:1497–1500
Cordonnier R, Bayonove C (1974) Mise en evidence dans la baie deraisin, variété Muscat d’Alexandrie, de monotèrpenes liés,révélables par une ou plusieurs encimes du fruti. CR Acad SciParis 278:3387–3390
333

Cordonnier RE, Gunata Z, Baumes RL, Bayonove CL (1989)Recherche d’un materiel enzymatique adapté à l’hydrolyse desprécurseurs d’arômes de nature glucosidique du raisin. ConnVigne Vin 23:7–23
Darriet P, Boidron JN, Dubourdieu D (1988) L’hydrolyse deshétérosides terpéniques du Muscat a Petit Grains par lesenzymes périplasmiques de Saccharomyces cerevisiae. ConnVigne Vin 22:189–195
Davis CR, Wibowo D, Eschenbruch R, Lee TH, Fleet GH (1985)Practical implications of malolactic fermentation: a review. AmJ Enol Viticult 36:290–301
Delcroix A, Gunata Z, Sapis JC, Salmon JM, Bayonove C (1994)Glycosidase activities of three enological yeast strains duringwine making. Effect on the terpenol content of Muscat wine.Am J Enol Viticult 45:291–296
Di Stefano R (1989) Evoluzione dei composti terpenici liberi eglucosidici e degli actinidioli durante la conservazione deimosti e dei vini in funzione del pH. Riv Vitic Enol 2:11–23
Di Stefano R(1991) Proposition d’une methode de preparation del’echantillon pour la determination des terpenes libres etglycosides des raisins et des vins. Bull OIV 4:721–725
Di Stefano R, Magiorotto G, Gianotti S (1992) Transformazioni dinerolo e geraniolo indotte dai lieviti. Riv Vitic Enol 42:43–49
Di Stefano R, Garcia E (1995) Attività idrolitica di preparati en-zimatici da foglie e da acini di Moscato sui glicosidi terpenici.Riv Vitic Enol 48:53–58
D’Incecco N, Bartowsky E, Kassara S, Lante A, Spettoli P,Henschke P (2004) Release of glycosidically bound flavourcompounds of Chardonnay by Oenococcus oeni during ma-lolactic fermentation. Food Microbiol 21:257–266
Dugelay I, Gunata Z, Sapis JC, Baumes R, Bayonove C (1992)Etude de l’origine du citronellol dans les vins. J Int Sci VigneVin 26:177–184
Dupin I, Gunata Z, Sapis JC, Bayonove C, M’Bairaroua O, TapieroC (1992) Production of a β-apiosidase by Aspergillus niger.Partial purification, properties and effect on terpenyl apiosyl-glucosides from grape. J Agric Food Chem 40:1886–1891
Engel KH, Flath RA, Butter RG, Mon TR, Ramming DW, TeranishiR (1988) Investigation of volatile constituents in nectarines. 1.Analytical and sensory characterization of aroma componentsin some nectarine cultivars. J Agric Food Chem 36:549–553
Esteve B, Manzanares P, Ramon D, Querol A (1998) The role ofnon-Saccharomyces yeasts in industrial wine making. IntMicrobiol 1:143–148
Fernandez M, Di Stefano R, Briones A (2003) Hydrolysis andtransformation of terpene glycosides from Muscat must bydifferent yeast species. Food Microbiol 20:35–41
Fleet GH (1993) Wine microbiology and biotechnology. Harwood,Switzerland
Francis MJO, Allcock C (1969) Geraniol β-D-glucoside: occurrenceand synthesis in rose flowers. Phytochemistry 8:1339–1347
Fu-Mian C, Pifferi PG, Setti L, Spagna G, Martino A (1994)Immobilizzazione di un’antocianasi (β-glucosidasi) da Asper-gillus niger. Ital J Food Sci 1:31–42
Gonde P, Ratomahenina R, Arnaud A, Galzy P (1985) Purificationand properties of the exocellular β-glucosidase of Candidawickerhamii (Zikes) Meyer and Yarrow capable of hydrolysingsoluble cellodextrines. Can J Biochem Cell Biol 63:1160–1166
Grimaldi A, McLean H, Jiranek V (2000) Identification and partialcharacterization of glycosidic activities of commercial strains ofthe lactic acid bacterium, Oenococcus oeni. Am J Enol Viticult51:362–369
Grossmann M, Rapp A, Rieth W (1987) Enzymatische freisetzunggebundener aromastoffe in wein. Dtsch Lebensm Rdsch 83:7–12
Gueguen Y, Chemardin P, Pien S, Arnaud A, Galzy P (1997)Enhancement of aromatic quality of Muscat wine by the use ofimmobilized β-glucosidase. J Biotechnol 55:151–156
Gunata YZ, Bayonove CL, Baumes RL, Cordonnier RE (1985) Thearoma of grapes. I. Extraction and determination of free andglycosidically bound fraction of some grape aroma compo-nents. J Chromatogr 331:83–90
Gunata Z, Bitteur S, Brillouet JM, Bayonove C, Cordonnier R(1988) Sequential enzymic hydrolysis of potentially aromaticglycosides from grape. Carbohydr Res 184:139–149
Gunata Z, Bayonove C, Tapiero C, Cordonnier R (1990a) Hydrol-ysis of grape monoterpenyl β-D-glucosides by various β-glucosidases. J Agric Food Chem 38:1232–1236
Gunata Z, Brillouet JM, Voirin S, Baumes B, Cordonnier R (1990b)Purification and some properties of an α-L-arabinofuranosidasefrom Aspergillus niger. Action on grape monoterpenyl arab-inofuranosyl glucosidases. J Agric Food Chem 38:772–776
Gunata Z, Dugelay I, Sapis JC, Baumes R, Bayonove C (1990c)Action des glycosidases exogènes au cours de la vinification:Liberation de l’arôme à partir des précurseurs glycosidiques. JInt Sci Vigne Vin 24:133–144
Gunata Z (1993) Etude et exploitation par voie enzymatique desprecurseurs d’arômes du raisin de nature glycosidique. RevOenol 74:22–27
Gunata Z, Dugelay I, Sapis JC, Baumes R, Bayonove C (1993) Roleof the enzymes in the use of the flavour potential from grapeglycosides in winemaking. In: Schreier P, Winterhalter P (eds)Progress in flavour precursor studies. Alured, Carol Stream, pp219–234
Gunata Z, Dugelay I, Vallier MJ, Sapis JC, Bayonove C (1997)Multiple forms of glycosidases in an enzyme preparation fromAspergillus niger: partial characterization of an apiosidase.Enzyme Microbiol Technol 21:39–44
Gunata Z, Blondeel C, Vallier MJ, Lepoutre JP, Sapis JC, WatanabeN (1998) An endoglycosidase from grape berry skin of cv. M.Alexandrie, hydrolysing potentially aromatic disscharide glyco-sides. J Agric Food Chem 46:2748–2753
Gunata Z, Vallier MJ (1999) Production of highly glucose-tolerantextracellular β-glucosidase by three Aspergillus strains. Bio-technol Lett 21:219–223
Gunata Z (2002) Flavor enhancement in fruit juices and derivedbeverages by exogenous glycosidases and consequences of theuse of enzyme preparations. In: Whitaker JR (ed) Handbook offood enzymology. Marcel Dekker, New York, pp 303–330
Heidlas H, Lehr M, Idstein H, Schreier P (1984) Free and boundterpene compounds in papaya (Carica papaya L.) fruit pulp. JAgric Food Chem 32:1020–1021
Henick-Klling T (1995) Control of malo-lactic fermentation in wine:energetics, flavour modification and methods of starter culturepreparation. J Appl Bacteriol 79(Suppl):29–37
Hernandez LF, Espinosa JC, Fernandez M, Briones A (2003)β-Glucosidase activity in a Saccharomyces cerevisiae winestrain. Int J Food Microbiol 80:171–176
Hock R, Benda J, Schreier P (1984) Formation of terpenes by yeastsduring alcoholic fermentation. Z Lebensm Unters Forsch 179:450–452
Huang HT (1955) Fruit colour destruction: decolorization ofanthocianins by fungal enzymes. J Agric Food Chem 3:141–146
Jiang J, Paterson A, Piggott JR (1990) Effects of pectolytic enzymetreatments on anthocyanins in raspberry juice. Int J Food SciTechnol 25:596–600
Krammer G, Winterhalter P, Schwab M, Schreier P (1991) Gly-cosidically aroma bound compounds in the fruits os Prunusspecies: apricot (P. armeniaca L.), peach (P. persica L.), yellowplum (P. domestica L. ssp syriaca). J Agric Food Chem 39:778–781
Lecas M, Gunata Z, Sapis JC, Bayonove C (1991) Purification andpartial characterization of β-glucosidase from grape. Phyto-chemistry 30:451–454
Leclerc M, Arnaud A, Ratomahenina R, Galzy P, Nicolas M (1984)The enzyme system in a strain of Candida wickerhamii Meyerand Yarrow participating in the hydrolysis of cellodextrins. JGen Appl Microbiol 30:509–521
Lutz A, Winterhalter P (1992) Isolation of additional carotenoidmetabolites from quince fruit (Cydonia oblonga Mill). J AgricFood Chem 40:1116–1120
334

Maicas S, Gil JV, Pardo I, Ferrer S (1999) Improvement of volatilecomposition of wines by controlled addition of malolacticbacteria. Food Res Int 32:491–496
Manitto P (1980) Byosynthesis of natural products. Ellis Horwood,Chichester
Mansfield AK, Zoecklein BW, Whiton RS (2002) Quantification ofglycosidase activity in selected strains of Brettanomyces brux-ellensis and Oenococcus oeni. Am J Enol Viticult 53:303–307
Mateo JJ, Gentilini N, Huerta T, Jiménez M, Di Stefano R (1997)Fractionation of glycosilated aroma precursors of grapes andwines. J Chromatogr A 778:219–224
Mateo JJ, Di Stefano R (1998) Enological properties ofβ-glucosidasein wine yeasts. Food Microbiol 14:583–591
Mateo JJ, Jimenez M (2000) Monoterpene in grape juice and wines.J Chromatogr A 881:557–567
McMahon H, Zoecklein BW, Fugelsang K, Jasinski Y (1999)Quantification of glycosidase activities in selected yeasts andlactic acid bacteria. J Ind Microbiol Biotechnol 23:198–203
Mendes A, Climaco MC, Mendes A (2001) The role of non-Saccharomyces species in releasing glycosidic bound fractionof grape aroma components—a preliminary study. J ApplMicrobiol 91:67–71
Ogawa K, Yasuyuki I, Guo W, Watanabe N, Usui T, Dong S, TongQ, Sakata K (1997) Purification of a β-primeverosidaseconcerned with alcoholic aroma formation in tea leaves (cv.Shuixian) to be processed to oolong tea. J Agric Food Chem45:877–882
Pabst A, Barron B, Etievant P, Schreier P (1991) Studies on theenzymatic hydrolysis of bound aroma constituents fromraspberry fruit pulp. J Agric Food Chem 39:173–175
Rapp A, Mandery H (1986) Wine aroma. Experientia 42:873–884Razungles A, Bayonove CL, Cordonnier RE, Baumes RL (1987)
Investigation on the carotenoids of the mature grape. Vitis26:183–191
Riccio P, Rossano R, Vinella M, Domizio P, Zito F, Sanseverino F,D’Elia A, Rosi I (1999) Extraction and immobilization in onestep of two β-glucosidases released from a yeast strain ofDebaryomyces hansenii. Enz Microb Technol 24:123–129
Riou C, Salmon JM, Vallier MJ, Gunata Z, Barre P (1998)Purification, characterization and substrate specifity of novelhigh glucosidase from Aspergillus orzyae. Appl Environ Mi-crobiol 64:3607–3614
Roscher R, Herderich M, Steffen JP, Schreier P, Schwab W (1997)2,5-dimethyl-4-hydroxy-3(2H)-furanone 6′-o-malonyl β-D-glu-copyranosidase in strawberry fruits. Phytochemistry 43:155–159
Rosi I, Vinella M, Domizio P (1994) Characterization of β-glu-cosidase activity in yeasts of enological origin. J Appl Bacteriol77:519–527
Rwahahizi S, Wrolstad RE (1988) Effects of mold contaminationand ultrafiltration on the color stability of strawberry juice andconcentrate. J Food Sci 53:857–861
Sakho M, Chassagne D, Crouzet J (1997) African mango gly-cosidically bound volatile compounds. J Agric Food Chem 45:883–888
Salles C, Jallageas JC, Fournier F., Tabet JC, Crouzet JC (1991)Apricot glycosidically bouns volatile components. J Agric FoodChem 39:1979–1983
Sarry JE, Gunata Z (2004) Plant and microbial glycoside hydrolases:Volatile release from glycosidic aroma precursors. Food Chem87:509–521
Schwab W, Mahr C, Schreier P (1989) Studies on the enzymatichydrolysis of bound aroma components from carica papayafruit. J Agric Food Chem 37:1009–1012
SchwabW, Scheller G, Schreier P (1990) Glycosidically bound aromacomponents from sour cherry. Phytochemistry 29:607–612
Sefton MA, Williams PJ (1994) Generation of oxidation artifactsduring the hydrolysis of norisoprenoid glycosides by fungalenzyme preparations. J Agric Food Chem 39:1994–1997
Shoseyov O, Bravdo BA, Ikan R, Chet I (1988) Endo-β-glucosidasefrom Aspergillus niger grown on monoterpene glycoside con-taining medium. Phytochemistry 27:1973–1976
Shoseyov O, Bravdo BA, Goldman A, Cohen S, Shoseyov L, IkanR (1990) Immobilized endo-β-glucosidase enriches flavour ofwine and passion fruit juice. J Agric Food Chem 38:1387–1390
Skouroumounis GK, Winterhalter P (1994) Glycosidically boundnorisoprenoids from Vitis vinifera cv. Riesling leaves. J AgricFood Chem 42:1066–1072
Skouroumounis GK, Massy-Westropp RA, Sefton MA, Williams PJ(1995) Synthesis of glucosides related to grape and wine aromaprecursors. J Agric Food Chem 43:974–980
Spagna G, Andreani F, Salatelli F, Romagnoli D, Casarini D, PifferiPG (1998) Immobilization of the glycosidases: α-L-arabinofur-anosidase and β-D-glucopyranosidase from Aspergillus nigeron a chitosan derivative to increase the aroma of wine. Part II.Enzyme Microbiol Technol 23:413–421
Stahl-Bishop E, Intert F, Holthuijzen J, Stengele M, Schulz G (1993)Glycosidically bound volatiles—a review 1986–1991. FlavFrag J 8:61–80
Strauss CR, Wilson B, Williams PJ (1988) Novel monoterpene diolsand diol glycosides in Vitis vinifera grapes. J Agric Food Chem36:569–573
Suarez M, Duque C, Wintoch H, Schreier P (1991) Glycosidicallybound aroma compounds from the pulp and the peelings ofLulo fruit (Solanum vetissimum D). J Agric Food Chem 39:1643–1645
Ugliano M, Genovese A, Moio L (2003) Hydrolysis of wine aromaprecursors during malolactic fermentation with four commer-cial starter cultures of Oenococcus oeni. J Agric Food Chem51:5073–5078
Vasserot Y, Arnaud A, Galzy P (1995) Monoterpenol glycosides inplants and their biotechnological transformation. Acta Biotech-nol 15:77–95
Voirin S, Baumes R, Bayonove C, M’Bairaroua O, Tapiero C (1990)Synthesis and NMR spectral properties of grape monoterpenylglycosides. Carbohydr Res 207:39–56
Voirin S, Baumes R, Sapis JC Bayonove C (1992) Analyticalmethod for monoterpene glycosides in grape and wine. Part 2.Qualitative and quantitative determination of glycosides ingrape. J Chromatogr 595:269–281
Wightman JD, Steven FP, Watson BT, Wrolstad RE (1997) Someeffects of processing enzymes on anthocyanins and phenolics inPinot Noir and Cabernet Sauvignon wines. Am J Enol Viticult48:39–47
Williams PJ, Strauss CR, Wilson B (1980) Hydroxylated linaloolderivatives as precursors of volatile monoterpenes of Muscatgrapes. J Agric Food Chem 28:766–771
Williams PJ, Strauss CR, Wilson B, Massy-Westropp RA (1982)Use of C18 reversed-phase liquid chromatography for the iso-lation of monoterpene glycosides and nor-isoprenoid precursorsfrom grape juice and wines. J Chromatogr 235:471–480
Williams PJ, Cynkar W, Francis IL, Gray JD, Iland PG, Coombe BG(1995) Quantification of glycosides in grapes, juices, and winesthrough a determination of glycosyl glucose. J Agric FoodChem 43:121–128
Wilson B, Strauss CR, Williams PJ (1986) The distribution of freeand glycosidically bound monoterpenes among skin, juice andpulp fractions of some white grape varieties. Am J Enol Viticult37:107–111
Winterhalter P (1990) Bound terpeniods in the juice of the purplepassion fruit (Passiflora edulis Sims.). J Agric Food Chem38:455–460
Winterhalter P, Skouroumounis GK (1997) Glycoconjugated aromacompounds: occurrence, role and biotechnological transforma-tion. In: Scheper T (ed) Advances in biochemical engineering/biotechnology. Springer, BerlinHeidelbergNewYork, pp 74–105
Wu P, Kuo MC, Hartman TG, Roten RT, Ho CT (1991) Free andglycosidically bound aroma components in pineapple (Ananascomosus L. Merr.). J Agric Food Chem 39:170–172
Yang HY, Steele WF (1958) Removal of excessive anthocyaninpigment by enzyme. Food Technol 12:517–519
Young H, Paterson VJ (1995) Characterization of bouns flavourcomponents in kiwi fruit. J Sci Food Agric 68:257–260
335





























