Embed Size (px)
Citation preview

Shine Jr, J.M. et a/. Proc: ISSCT, V& 25, 2005
COMPARISON OF FIVE ISOLATES OF SUGARCANE BROWN RUST AND DIFFERENTIAL REACTION ON SIX SUGARCANE CLONES
JAMES M. SHINE ~ r ' , JACK C. COMSTOCK~ and JACK L. DEAN^ 'sugar Cane Growers Cooperative of Florida,
Post Ofice Box 666, Belle Glade, Florida 33430, USA jmshine@scgc. org
USDA-ARS Sugarcane Field Station, Canal Point, Florida 33438 3~eceased
KEYWORDS: Puccinia melanocephala, Saccharurn spp., Rust Races, Single-Degree of Freedom Comparisons, AUDPC,
Abstract
THE OCCURRENCE of pathological races of sugarcane brown rust (Puccinia melanocephala H. and P. Sydow) in Florida was first reported in 1984. Cultivars that were resistant to the disease in the variety development program displayed susceptibility to the disease once expanded commercially. A study was conducted over two years to determine the effect on yield of six cultivars of sugarcane to five isolates of the sugarcane rust pathogen suspected to be differing pathological races. Sugarcane plants were grown in pots and inoculated under conditions favouring disease development while preventing infection of non-inoculated control plants. Plant height, estimates of percent infection and whole-plant disease ratings were recorded bi-weekly for 4 months during the late winter and spring of 1991 and 1992. Final plant height and fresh weight were measured at the end of the experiment each year. Areas under the disease progress curves (AUDPC) for plant height, percent infection and disease rating were calculated. Analysis of variance and single-degree-of-freedom comparisons were used to interpret results for all response variables. These analyses indicated the existence of four pathogenic races of the brown rust pathogen in this experiment. These data coupled with field observations lead to the hypothesis that pathogenic variants of P. melanocephala develop within the sugarcane production region and parallel the development of new commercial sugarcane cultivars. The diversification of commercial cultivars and identification of horizontal resistance mechanisms will reduce losses to sugarcane brown rust in the Florida sugarcane industry.
Introduction
There has been much argument over the existence of races of the causal agent of sugarcane brown rust, Puccina melanocephala H. and P. Sydow, in sugarcane production regions of the world. Sugarcane rust was first reported in the Americas in 1978 (Dean et al., 1979). A report in 1984 presented data on one variety of sugarcane, CP 79-1 580. This variety was originally rated as resistant to the disease, and was later observed to be susceptible at some outfield test sites while appearing resistant at others (Dean and Purdy, 1984). A second observation of a change in the amount of disease observed on a cultivar was observed two years after the release of CP 78-1247 (Raid, 1989).
The cultivar was widely planted soon after its release to commercial growers due to its superior agronomic performance in trials. Significant sugarcane rust was observed on the cultivar early in 1988 and yields at harvest were 40% of expected yield as determined from experiments conducted on the same farms three years prior (Raid et al., 199 1). In yet another example, CP 72- 12 10 displayed resistance to rust when it was released in 1980 and, by 1986, it occupied more than 50% of the total acreage planted to sugarcane in Florida (Glaz and Coale, 1992).
During the spring of 1987, rust was severe and widespread on this variety although not uniformly distributed over the entire south Florida production region. Losses on CP 72-1210 were estimated to be as high as 39% of its historical yield in rust-infected plant cane (Raid et al., 1991). The importance of CP 72- 1210 declined rapidly because of the effects of sugarcane rust.

Shine Jr, J.M. et a/. Proc. ISSCT, Vol. 25, 2005
Economic yield losses due to rust experienced by farmers growing CP 72-1210 stimulated diversification of cultivars planted in Florida. Cultivars planted to replace CP 72- 12 10 were rated 'resistant' to rust at release. Since 1989, two of the so-called resistant vasieties, CP 74-2005 and CL 73-239 have shown a susceptible reaction to the disease in localised areas. Yield losses due to rust on planting of the formerly resistant CL 73-239 were estimated to range from 20-30%. The cultivar CP 74-2005 showed a highly susceptible reaction in localised aseas. None-the-less, the cultivars were accepted as resistant and are now considered susceptible. Conversely, CL 41-223 was grown on 10% of the planted acreage when rust was first observed in Florida (Glaz, 1981). Sugarcane rust was a major factor contributing to the decision to totally replace this cultivar. Since the removal of this cultivar fiom commercial production, plantings of this cultivar in small plots show little rust development relative to other susceptible varieties planted in the same area. These observations have led pathologists and breeders in Florida to believe that there are pathogenic races of sugarcane rust present (Dean and Purdy, 1984; Raid, 1989). Changes in the distribution of rust races appear to correspond with changes in cultivars grown, generally increasing in importance with increasing extent of new cultivars.
Discussions on the existence of pathological races of the sugarcane rust fungus have been argued since it was first reported in the Americas. Arguments presented by Whittle (1980) suggested that the term race should only be used in pathosystems involving vertical resistance where a gene-for-gene resistance relationship can be demonstrated; and that this is not probable in the sugarcane/sust pathosystem since sugarcane is a complex polyploid. The existence of races must be evidenced by a significant differential reaction to the disease between two isolates and two clones and it is on this basis that we are claiming to demonstrate the existence of races regardless of the genetic basis. The existence of races bears importance in breeding strategies employed to develop host plant resistance.
The objectives of this study were to:
1) determine whether or not there are pathogenic races of sugarcane rust in Florida;
2) identifjr, if possible, some of the pathogenic races, if they exist; and
3) determine if identification of pathogenic races is possible using traditional qualitative ratings of percent TVD leaf infection, overall severity and quantitative measures of plant height compared to non-inoculated control and final fresh weight of above ground plant pasts.
Materials and methods Rmt isolates
Five isolates of sugarcane rust were collected from sources that were suspected to represent individual pathogenic races of P. melanocephala. The rust isolates were iCP72-1210, iCP78-1247, iB4362, iCL41-223, and iCL73-239. The isolates were named based on the cultivar from which they were collected and expected to be the best adapted. Collection and increase of isolates from single uredia was attempted, but abandoned 'since there were questions regarding the possibility of selecting a different genetic variant than that generally observed in the field.
Isolate iCL41-223 was collected by Mike lrey (United States Sugar Corporation) in 1984 fiom infected fields. The cultivar was grown on a declining number of acres at the time and suffered severe losses to the disease. Isolate iCP72-1210 was collected in the spring of 1990. This isolate was collected from an area where a number of varieties were being grown, some of which were known to be moderately susceptible to the disease. The disease was more intense on CP 72-1210, while expression of the disease on other susceptible cultivars was vely light. Isolate iB4362 was collected from the USDA Sugarcane Field Station at Canal Point and was suspected to be different because other cultivars used in the experiment, with the exception of CP 72-1210, appeared resistant to it.
It would be plausible for several pathogenic races to be present at Canal Point, since a large sugarcane germplasin collection is grown on the experiment station and conditions favour rust development year-round. The iCP78-1247 isolate was collected in 1990 from fields of severely infected CP 78-1247 located south of Pahokee, Florida. This was two years after the first severe epidemic on the cultivar. This area appeared to be a focus for spread of the disease throughout the production region. Disease development on the 'parent' cultivar was notably more severe than other susceptible cultivars in the area, such as CP 72-1210. This was the first assurance on a commercial basis that pathogenic races exist. The isolate collected from CL 73-239 came from fields located south of Canal Point and east of Pahokee in 1990. Expression of the disease occurred earlier in the yeas than that normally observed and did not occur in neighbouring fields of CP 72- 121 0.
639

Shine Jr, J.M. eS a/, Proc. ISSCT, Vol. 25, 2005
Each isolate was collected and stored in liquid nitrogen. Urediospore germination was detelmined to be low before storage (in the order of 10%) and decreased over a shost period of time in storage. The isolates were collected from whole leaves that were cut in the field in the morning, brought to the laboratory, air-dried and spores collected using a custom-made vacuum aspirator. The spores were typically collected froin the leaf associated with the youngest visible collar. This leaf has pustules just beginning the most productive phase of the asexual reproductive cycle with little secondary infection and necrosis associated with the lesion (Shine et al., 1989). The leaves were sometimes washed to remove dust and debris, and then held at room temperature overnight in a humid chalnber and the spores again collected the next morning following air-diying.
Each isolate was increased on the 'parent' cultivar grown in 8 litre containers. A suspension of approximately lo5 sporeslml was prepared in distilled water immediately prior to inoculation. Sugascane plants were inoculated by placing a 0.5 inL drop of spore suspension in the whorl of each shoot and spraying an atomised spore suspension over the whole plant. The plants were held in a dew chamber overnight then removed to an isolated, shaded chamber. These plants were re-inoculated on a twice-weekly basis to maintain a source of inoculum throughout the experiment. This process was repeated for each isolate, handling each isolate separately to avoid cross infection and mixing of pure types. The spores were collected from these plants using a hand-held mouth-operated vacuum aspirator. Spores were collected in gelatin capsules, stored at room temperature and used for inoculum within two days after collection.
Plant material Sugarcane plants of six cultivars were germinated from single-eyed seed-pieces and transplanted
into 8-litre containers after germination. Plants were selected for uniformity in size and vigour. The soil medium was a mixture of one past pine bark, one past sand, and one part peat. A slow release festiliser (~smocote@ 13-13-1 3) was added to this mix prior to planting. Plants were fertilised with a complete liquid fertiliser ( ~ r a c i t e ~ 20-20-20) bi-weekly throughout the experiment.
The experiment was a split-plot design. Each cultivar/isolate treatment was replicated 4 times with 3-plant sub-plots using the aforementioned 5 isolates of P, rnelcznocephala, a non-inoculated control and six sugarcane cultivars. Isolates were main plots in the experiment. The sugarcane cultivars were CP 72- 1210, CP 78-1247, CP 74-2005, B4362, CL 41-223, and CL 73-239. The six main plots were randomly arranged on each of four carts, serving as replicates, in the photoperiod house at Calla1 Point. This allowed the plants to be moved in and out of the building for exposure to normal sunlight during the day and moving them in at night to prevent dew formation on the leaves. The plants could also be moved into the chambers to avoid rainfall and damaging winds. The chambers were maintained at 26°C to provide favourable conditions for disease development.
Care was taken not to wet the foliage while irsigating to prevent infection by spores that may have fallen on the plants from the adjacent cane-growing area or cross infection from neighbouring plots. Each plot was surrounded by a tarpaulin to reduce wind-borne spread of loose spores from plot to plot. The non- inoculated plot in each replicate was maintained to determine the amount of cross-infection and extent of disease development. The measurements taken on the non-inoculated control provide the basis for determining yield loss.
Inoculation
The inoculation procedure followed was similar to that repor-ted by Comstoclc et al. (1 992). Plants were inoculated with spores of each respective isolate by preparing a suspension in distilled water of approximately 8x 1 o4 spores1mL. The carts were rolled indoors and the plants to be inoculated covered with plastic bags. The spore suspension was sprayed over the entire plant by reaching up into the bag. The bags were left on the plants overnight to maintain leaf wetness. The non-inoculated plants were not covered to minimise disease development arising from spores previously deposited on leaves which might germinate under the humid conditions created by bagging. The bags were removed the morning following inoculation and the carts returned to the outdoors. This process was initially completed twice a week until an epidemic was established. The inoculation procedure was then repeated weekly to perpetuate the epidemic on the inoculated plants. /
Data collection and analysis
The experiment was repeated over a two-year period. The first year, inoculation of treatments began February 25, 1991 and ended May 8, 199 1. Final height measurements were taken June 3, 199 1, and fresh weights were determined June 6, 1991. The second year, inoculation began on February 3, 1992 and
Shine Jr, J.M. ef a/. Proc. ISSCT, Vol. 25, 2005
ended June 2, 1992. Final heights were determined June 10, 1992, and fresh weight determinations made June 12, 1992. Inoculation treatments were started as soon as sufficient inoculu~n was available. The experiment was discontinued both years when temperature conditions (>30°C) prohibited inocul~un production and disease development.
Sugarcane plant height measurements were taken prior to the first inoculation by measuring from the soil surface to the youngest visible collar on the oldest tiller. Height measurements were continued semi-monthly for the first 60 days of the experiment and weekly thereaftel-. The measuring frequency was increased when the plants began to grow faster. The objective was to measure the height at the rate of new leaf emergence. Ratings were taken when rust pustules became visible on inoculated plants and continued every two weeks throughout the experiment.
Two rating parameters were used; the first parameter (% infection) was an estimate of the percent area occupied by lesions on the leaf associated with the youngest visible collar (top visible dewlap leaf, TVD). The second parameter (rating) was an overall plant rating that classified the severity of infection on a scale of 1 to 10. The severity rating scale was defined by Coinstock et al. (1992) where 1 = no apparent infection and 10 = Inany productive pustules on younger foliage and necrosis of older leaves due to the disease. Whole-plant fresh weights were taken at the end of the experiment by cutting all plant parts above the soil line in the container and weighing them on an electronic balance.
The area under the disease progress cullre (AUDPC) was calculated for % infection and rating over the duration of the experiment. AUDPC was calculated to integrate the bi-weekly determinations made over the period of the experiment. Plant height was adjusted for the non-inoculated control by subtracting the mean height of three plants of each cultivar within each replicate from the mean height of the three plants of the non-inoculated treatment within each respective replicate.
This value could then be used as a quantitative measure of disease development and an AUDPC value could be calculated. Analysis of variance was perfosmed on fresh weight, AUDPC height, AUDPC % infection, and rating.
Single-degree-of-freedom contrasts for all possible pairs of isolates and clones were made using the F-test and significant differences were determined (P = 0.01). Each year was analysed separately. The objective of this experiment was to identify which isolate/cultivar pairs reacted significantly differently from one another using both the qualitative rating parameters and the quantitative growth parameters.
Results and discussion The hypothesis was that there are pathogenic variants in the brown rust pathogen population in the
Florida sugarcane production region. The goal of this study was to determine whether or not races exist in Florida by focusing on the quantitative parameters of weight and height as much as possible. Percent infection and disease severity ratings were included in the analysis, since they are commonly used parameters for assessing the severity of sugarcane rust diseases.
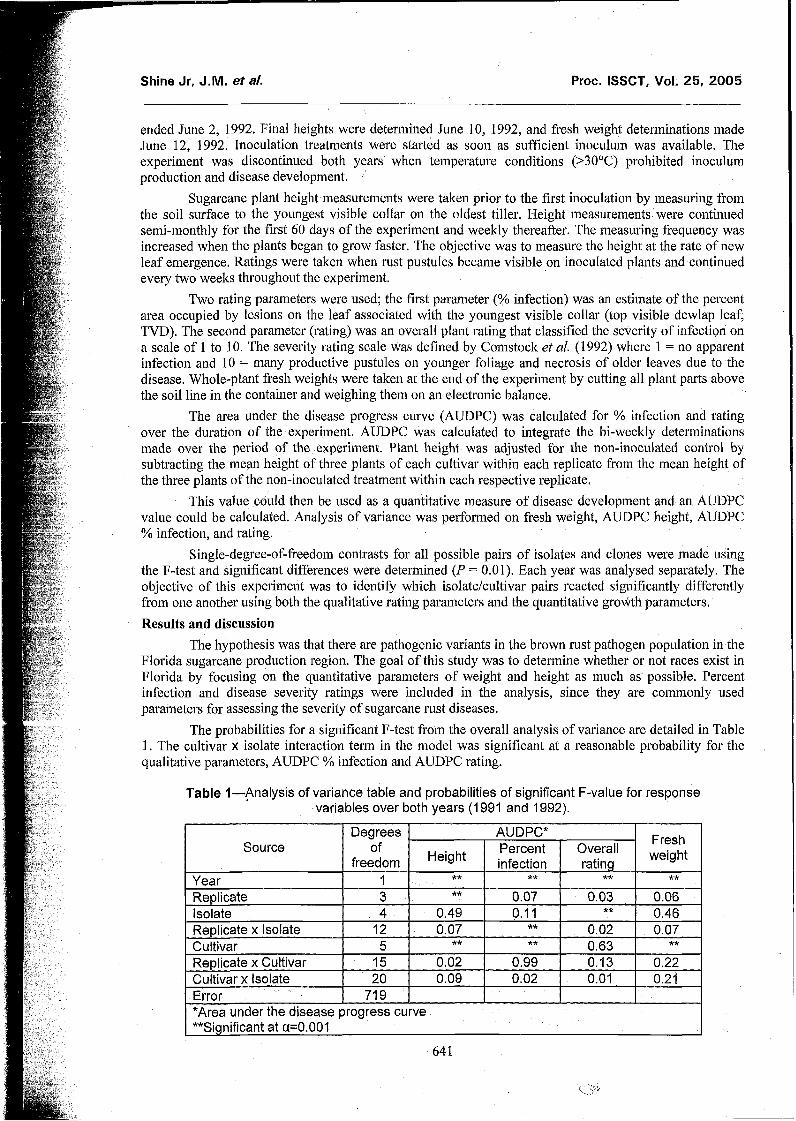
The probabilities for a significant F-test froin the overall analysis of variance are detailed in Table 1. The cultivar x isolate interaction term in the inodel was significant at a reasonable probability for the qualitative parameters, AUDPC % infection and AUDPC rating.
Table I-Analysis of variance table and probabilities of significant F-value for response variables over both years (1 991 and 1992).
64 1
< ) *
*Area under the disease progress curve **Significant at a=0.001
Shine Jr, J.M. et a/. Proc. ISSCT, Vol. 25, 2005
The interaction was also significant at P = 0.09 probability level for pri~nary tiller height. The interaction was not significant at a reasonable (P < 0.10) level of probability for fiesh weight. The weight parameter was highly variable primarily due to the young age of the plants. There was a significant difference between years for all parameters measured. The difference between years is likely due to the onset of elevated temperatures in early spring of Year 1 which limited disease development and the duration of the epidemic relative to Year 2. Significant differences in the height and weight parameters between cultivars were obsewed due to variety characteristics.
Differences among isolates averaged across all clones would be significant if at least one isolate caused colnparatively inore or less damage (i.e. more or less virulent) relative to the other isolates. The isolates that were targeted appeared to be similar in virulence and exhibited differential infection across the cultivars. Overall, the extent of damage caused by each isolate was silnilar on average across all clones, resulting in no differeilce between isolates for all parameters except AUDPC Rating.
Host specificity of races is identified by interaction, in the simplest case, betiveen two cultivars and two isolates. An example is where Cultivar A is susceptible to Isolate A and resistant to Isolate B and Cultivar B is resistant to Isolate A and susceptible to Isolate B. This interaction would be highly significant if it were a simple matter of resistance and high susceptibility in the strictest sense. The reality is that, while some sugarcane cultivars are highly resistant to sugarcane rust, none have deinonstrated irninunity.
This supports arguments presented by Whittle (1980) and Robinson (1976), namely, tllht vertical resistance genes may not be involved in the host resistance mechanisms in this pathosystem. Also, rust disease development is limited to periods of relatively cool (15-28OC), humid environmental conditions. These conditions are typically only present for a few months early in the growth of the crop in Florida. Susceptible plants begin to grow more or less nor~nally once epidemics decline in the summer months.
I , Differences between pairs of isolates and the disease response between isolate/cultivar pairs is a matter of degree and therefore difficult to detect, especially on mature plants. This is why the evaluations were done in the spring and the experiment was terminated when the weather became too hot for urediospore germination and disease development.
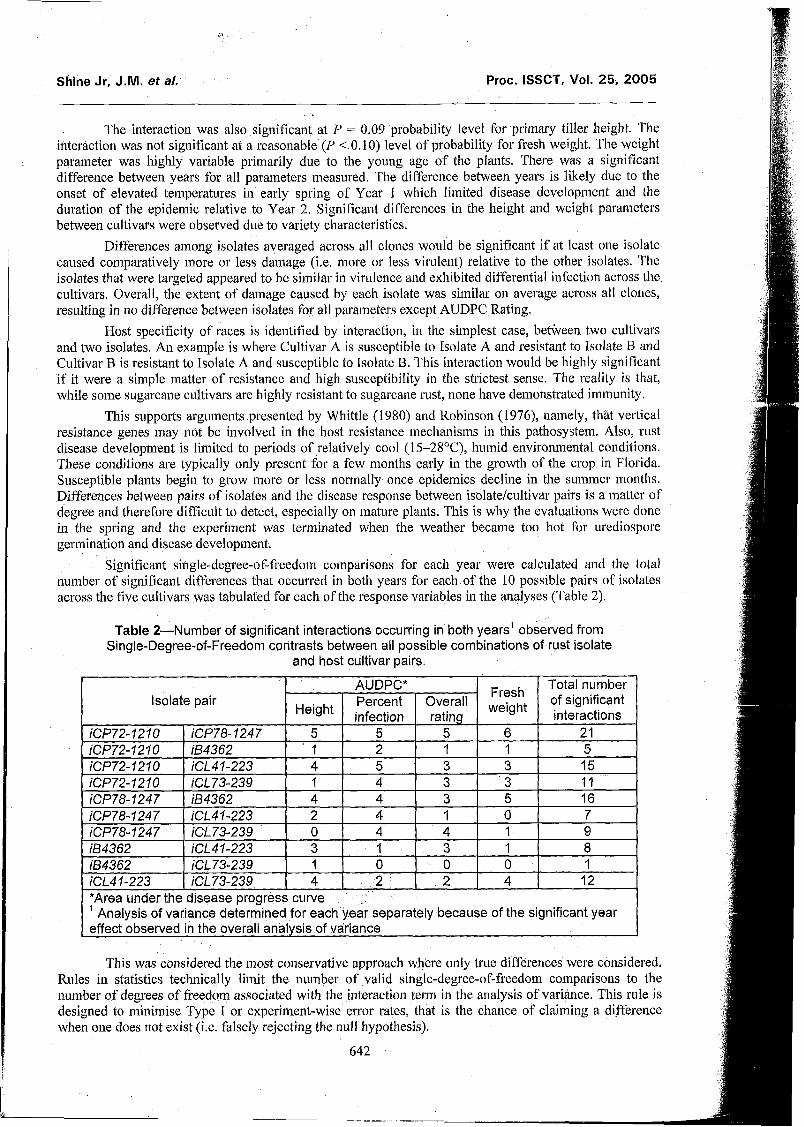
Significant single-degree-of-fi-eedom comparisolis for each year were calculated and the total number of signi.ficant differences that occurred in both years for each of the 10 possible pairs of isolates across the five cultivars was tabulated for each of the response variables in the analyses (Table 2).
Table 2-Number of significant interactions occurring in both years' observed from Single-Degree-of-Freedom contrasts between all possible combinations of rust isolate
and host cultivar pairs.
This was considered the most conservative approach where only true differences were considered. Rules in statistics technically limit the number of valid single-degree-of-heedoln comparisons to the number of degrees of freedom associated with the interaction term in the analysis of variance. This rule is designed to minimise Type I or experiment-wise error rates, that is the chance of claiming a difference when one does not exist (i.e. falsely rejecting the null hypothesis).
Shine Jr, J.M. eta/, Proc. ISSCT, Vol. 25, 2005
Furthermore, specific co~nparisons should tl~eoretically be pre-identified prior to review of the data, in other words by randoln chance. An evaluation of every combination is needed in order to test the existence of one or more races. Computer simulation of datasets with similar means and standard deviation were conducted to test for the experiment-wise error rate (unpublished data). These simulations indicated that this inethod produced error rates on the order of 5% using data similar to the fresh weight parameter and on the order of 3% on data siinilar to the rating parameter. The approach talten in this study was designed to establish statistical support for interpretation of response variables and for determining if different physiological races of the pathogen were present in Florida.
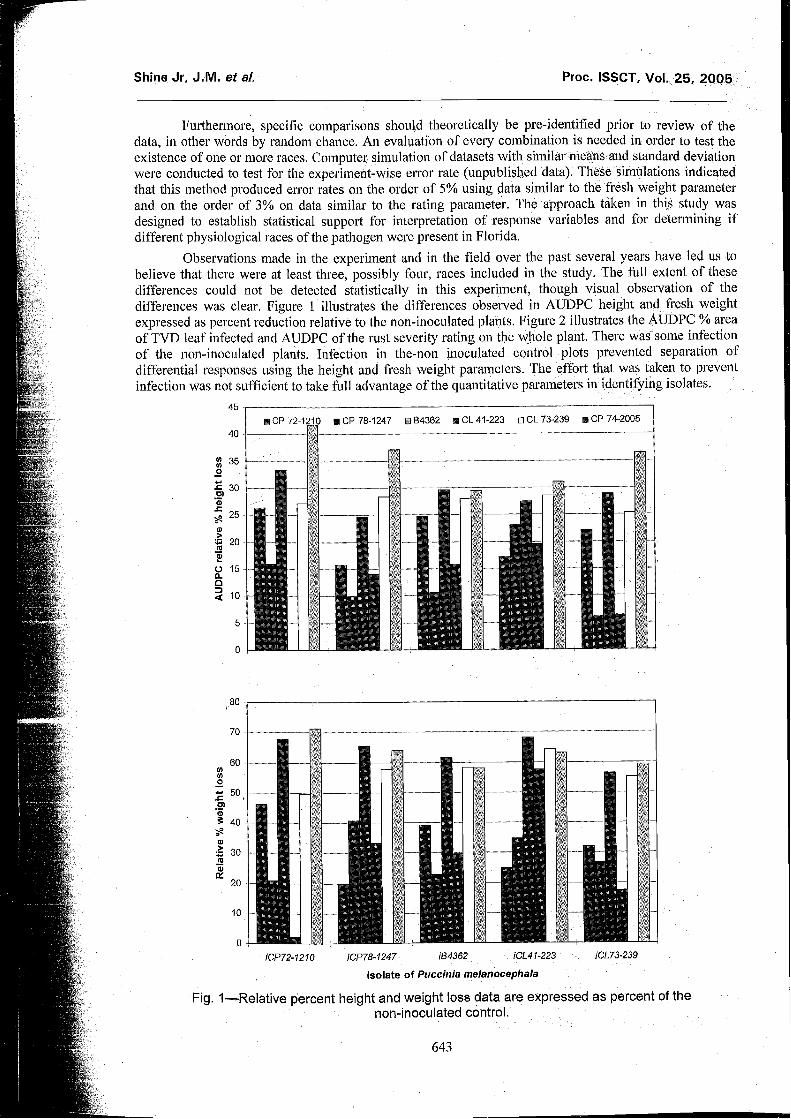
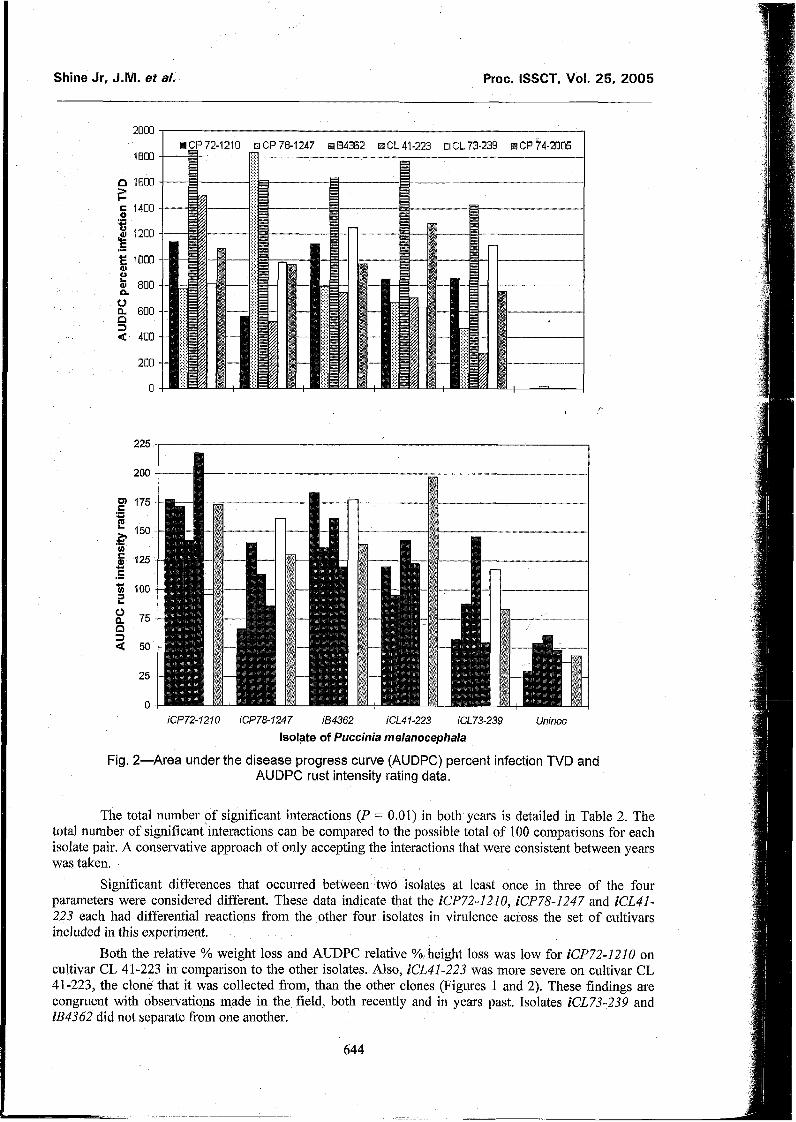
Observations made in the experiment and in the field over the past several years have led us to believe that there were at least three, possibly four, races included in the study. The full extent of these differences could not be detected statistically in this experiment, though visual observation of tlie differences was clear, Figure 1 illustrates the differences observed in AUDPC height and fresh weight expressed as percent reduction relative to the non-inoculated plants. Figure 2 illustrates the AUDPC % area of TVD leaf infected and AUDPC of the rust severity rating on the whole plant. There was some infection of the non-inoculated plants. Infection in the-non inoculated control plots prevented separation of differential responses using the height and fresh weight parameters. The effort that was taken to prevent infection was not sufficient to take full advantage of the quantitative parameters in identifying isolates.
Isolate of Puceinia melanocephak
Fig. I-Relative percent height and weight loss data are expressed as percent of the non-inoculated control.
Shine Jr, J.M. et a/. Proc. ISSCT, Vol. 25, 2005
tij loo 2
iCP72-1210 iCP78-1247 i54362 iCL41-223 iCL73-239 Uninoc
Isolate of Puccinia melanocephala
Fig. 2-Area under the disease progress curve (AUDPC) percent infection TVD and AUDPC rust intensity rating data.
The total number of significant interactions (P = 0.01) in both years is detailed in Table 2. The total number of ~i~nificant'interactions can be compared to the possible total of 100 comparisons for each isolate pair. A conselvative approach of only accepting the interactions that were consistent between years was taken.
Significant differences that occurred between two isolates at least once in three of the four parameters were considered different. These data indicate that the iCP72-1210, iCP78-1247 and iCL41- 223 each had differential reactions from the other four isolates in virulence across the set of cultivars included in this experiment.
Both the relative % weight loss and AUDPC relative % height loss was low for iCP72-1210 on cultivar CL 41-223 in comparison to the other isolates. Also, ECL41-223 was more severe on cultivar CL 41-223, the clone that it was collected from, than the other clones (Figures 1 and 2). These findings are congruent with observations made in the field, both recently and in years past. Isolates iCL73-239 and iB4362 did not separate from one another.

Shine Jr, J.M. et a/. Proc. ISSCT, Vol. 25, 2005
We suspected that the isolate iCL73-239 could be closely related to iCP78-1247. There is evidence to segregate them from each other and insuff~cient evidence to clearly list them as the same. Field observations indicate that the cultivar CL 73-239 did not sustain economic rust loss until approximately two years following the severe epidethic of rust on CP 78-1247, which caused major economic loss (Raid et al., 1991). CP 78-1247 was planted over a wide area, though not on a large acreage while CL 73-239 was being extended in range and acreage during the same period. CL 73-239 had been considered resistant to the disease when growers began to expand plantings of this variety. The number of spores produced throughout the areas nearby on cultivar CP 78-1247 certainly provided sufficient opportunity for any suitable variant in the pathogen to become established on the new host cultivar.
Similarly, we suspected a relationship between the iCL41-223 and iB4362 isolates, since both of the parent cultivars succumbed when rust first appeared in the ,Florida sugarcane production region. These data segregate the isolates indicating that they are likely to be different. iCL73-239 did not separate from iB4362 indicating these isolates might be the same. The cultivar B4362 has been considered a universal susceptible (i.e. susceptible to all races and strains of the rust pathogen). It is very possible that the parent host plant where the isolate was collected was infected with the iCL73-239 isolate and therefore did not exhibit differences in this experiment. We conclude these were four races of the rust pathogen included in this study.
Observations by pathologists working in Florida sugarcane concur that there are sugarcane clones considered susceptible now that did not contract the disease a few years after release. Changes have been observed earlier in the agronomic test phases of potentially new commercial varieties (Dean and Purdy, 1984). This indicates that there is sufficient inoculum produced on popular susceptible cultivars to develop new pathological races. Any one of these races could predominate within a year or two following the release of a variety.
The magnitude of economic losses depends on the extent of area the cultivar is planted on when a new highly virulent race develops. This means that physiological races that coincide with new sugarcane genotypes are developing parallel to the development of varieties. The hypothesis is then presented that pathogenic rust variants appearing in Florida are generated within the Florida production area and are not simply imported froin the outside as was the case when the initial occui-sence of the disease was reported in 1978 (Purdy et al., 1985).
The existence of differentially interactive pathogenic races is, in many respects, an academic question. Evidence from Australia supports the existence of only one pathogenic rust type within their sugarcane industty (Taylor, 1992). There have been reports from other regions (Comstock, 1988; Lui, 1982) that there are races present in those productipn regions, while the possibility of the existence of races of the rust pathogen has been denied by at least one researcher (Whittle, 1980) based on the hypothesis that there is no established gene-for-gene relationship.
Regardless of the genetic basis, changes are occurring in the Florida rust pathogen population that influences the severity of the disease on previously resistant sugarcane cultivars. The diversification of cultivars in production, the identification of horizontal resistance in the sugarcane genome, and incorporating this characteristic in coinmercial sugarcane cultivars may reduce and possibly eliminate economic losses due to sugarcane brown rust in Florida. Sugarcane breeding prograins must continue to strive for durable.resistance to the disease in sugarcane. Genes such as Brzll (Asnaghi et al., 2004), a major resistance gene to brown rust that is so fas efficient toward all tested isolates of P. melanocephala (Asnaghi et al., 2001), may contribute to achieve this goal.
REFERENCES Asnaghi, C., D'Hont, A., Glaszmann, J.-C. and Rott, P. (2001). Resistance of sugarcane cultivar R570
to Puccinia melanocephala isolates from different geographic locations. Plant disease, 85(3): 282-286.
Asnaghi C., Roques, D., Ruffel, S., Kaye, C., Hoarau, J.-Y., TClismart, H., Girard, 3.-C., Raboin, L.M., Risterucci, A.M., Grivet, L. and D'Hont A. (2004). Targeted mapping of sugarcane rust resistance gene (Brul) using bulked segregant analysis and RFLP markers. Theor. Appl. Genet., 108: 759-764.
Comstock, J.C. (I 988). Rust: Varietal differences in uredinospore production. In: Hawaiian Sugar Planters Association Expt. Sta. Rep. 1987,4042.
Comstock, J.C., Shine Jr, J.M. and Raid, R.N. (1992). Effect of rust on sugarcane growth and biomass. Plant disease, 76: 175-177.

Shine Jr, J.M. et a/. Proc. ISSCT, Vol. 25, 2005
Dean, J.L. and Purdy, L.H. (1984). Races of the sugar cane rust fungus, Puccinia melanocephala, found in Florida. Sugar Cane, (1): 15,16.
Dean, J.L., Tai, P.Y.P. and Todd, E,H. (1979). Sugarcane rust in Florida. Sugar Jownal, 42(2): 10.
Glaz, B. (1981). Florida's 1981 sugiu. cane variety census. Sugar y Azucar, 76(12): 37-40.
Glaz, B and Coale, F.J. (1 992). Sugar cane variety census: Florida 1991. Sugar y Azucar, 87(1): 32-35.
Lui, L.J, (1982). Culture of Puccinia nzelanocephala on detached leaves and uprights of sugarcane in Puerto Rico. Jour. Agric. Univ. P.R., 66: 168-176.
Purdy, L.H., Krupa, S.V. and Dean, J.L. (1985). Introduction of sugarcane rust into the Americas and its spread into Florida. Plant Disease, 69: 689-693.
Raid, R.N. (1989). Physiological specialisation in sugarcane rust (Puccinia melanocephala) in Florida. Plant Disease, 73: 183 (Disease Note).
Raid, R.N., Anderson, D.L. and Coale, F.J. (1991). Yield loss incited by Puccinia ~nilanoce~hala on a highly susceptible sugarcane cultivar in Florida. (Abstr.) Phytopathology, 8 1 : 699.
Raid, R.N., Comstock, J.C., Shine Jr, J.M. and Anderson, D.L. (1991). Effect of sugarcane rust on yield components of CP72-1210 plant cane. Univ. Fla. Belle Glade EREC Res. Rep. EV-1991-5: 1-7.
Robinson, R.A. (1976). Plant Pathosystems. Springer-Verlag, Berlin.
Shine, J.M., Chew, V. and Miller, J.D. (1989). A method for determining the spore production of sugarcane rust, (Puccinia melanocephala). Jour. of Amer. Soc. of Sugarcane Tech., 9: 30-37.
Taylor, P.W.J. (1992). Evidence for the existence of a single race of common rust caused by Puccinia melanocephala, in Australian sugar cane cultivars. Aust. J. Agric. Res., 43: 443-450.
Whittle, A.M. (1980). Do races exist in pathogens of sugarcane? Proc. First Inter-Am. Sugar Cane Seminar, Cane Diseases, 16-1 9. (Inter-American Transport and Equipment Company. Miami, Florida).
COMPARAISON DE ClNQ ISOLATS DE LA ROUILLE BRUNE DE LA CANNE A SUCRE ET REACTION DIFFERENTIELLE SUR SIX VARIETES COMMERCIALES
JAMES M. SHINE ~ r ' , JACK C. COMSTOCK* et JACK L. DEAN^ 'sugar Cane Growers Cooperative of Florida, Post OfJice Box 666, Belle Glade, Florida 33430, USA
2 ~ ~ ~ ~ - ~ ~ ~ Sugarcane Field Station, Canal Point, Florida 33438
3~ecede'
MOTS CLES: Puccinla ~nelanocephala, Saccharurn spp,, Races de Rouille, Comparaisons de Degre Simple de Liberti?, Surfaces sous la Courbe de Progression de la Maladie.
RCsumC
LA PRESENCE de races pathologiq~les de la rouille brune de la canne a sucre (Puccinia nzelanocephala H. et P Sydow) a 6te rapport6e pour la premiere fois en Floride en 1984. Les variktes qui s76taient montrkes rdsistantes a la rouille brune au cours du programme de dBveloppement varietal o l~t d6iriontr6 une sensibilitk la ~naladie des qu'elles ont Bti: exploit6es si 1'6chelle industrielle. Une 6tude a 6tC entreprise sur deux ans afin de determiner I'effet de cinq isolats du pathogene, soup$onnCs d'&tre des races pathologiques differentes, sur le rendeinent de six variet6s. Des plants de canne a sucre ollt Bt6 cultives en pots et inocul6s sous des conditions favorables au dkveloppement de la maladie, tandis que les plants tdmoins, non inoculds, etaient pr6serv6s de I'infection. La hauteur de la plante, le pourcentage d'infection et la reaction des plantes entihes a la maladie ont CtC notes deux fois par semaine pendant 4 mois A la fin de 17hiver et a~r printe~nps de 1991 a 1992. La hauteur totale de la plante ainsi que le poids eais oilt BtC mesures chaque annee a la fin de l'exp6rimentation. Les surfaces sous les courbes de progression de la lnaladie pour la hauteur de la plante, le pourcentage d'infection et la rCaction a la maladie ont 6tC calculCes. L7ana1yse de variance et les comparaisons du degrC simple de liberti: ont 6tC utiliskes pour interprkter les r6sultats de tous les paramhtres variables. Les resultats de cette Ctude ont indiqu6 I'existence de quatre races pathologiques de la rouille brune. Ces donnkes ajout6es aux observations faites au champ ont men6 a
646

Shine Jr, J.M. eta/. Proc. ISSCT, Vol. 25, 2005
I'hypothese que les variants pathogdniques de P melanocephala se dCveloppent dans la rBgion de production canniere en parallhle avec le developpe~nent des nouvelles varietds commerciales. La diversification de ces vari6tes et l'identification du mecanisme de rksistance horizontale permettront de rkduire les pertes dues a la rouille biune dans l'industrie de la canne St sucre en Floride.
COMPARACION DE CINCO AISLAMIENTOS DE ROYA CAFE EN REACCION DIFERENCIAL EN SEIS CLONES DE C A ~ A DE A Z ~ C A R
JAMES M. SHINE Jrl, JACK C. COMSTOCK~ y JACK L. DEAN^ 'sugar Cane Growers Cooperative of Florida, Post O f f e Box 666, Belle Glade, Florida 33430, USA
[email protected] . 2 ~ S ~ ~ - ~ ~ ~ Sugarcane Field Station, Canal Point, Florida 33438
3~eceased
PALABRAS CLAVES: Pticcinia melanocephala, Saccharum spp., Razas de Roya, Comparaciones de un Solo Grado de Libertad, AUDPC.
Resumen LA PRESENC~A de razas patologicas de la roya cafe de la caiia de azucar (Puccinia melanocephala H. y P. Sydow ) en la Florida, se registr6 por primera vez en 1984. Los cultivares que eran resistentes a la enfermedad en el programa de desamollo de variedades resultaron luego susceptibles cuando se sembraron co~ne~cialmente. Por tanto se realiz6 este estudio por msis de dos aiios para determinar el efecto sobre la produccicin, de seis cultivares de c@a de azucar a cinco aislalnientos de la roya de la caiia de azucar que se sospechaban correspondian a razas diferentes del pat6geno. Las plantas de caiia de azucar se cultivaron en macetas y fueron inoculadas bajo condiciones de invernadero que favorecian el desarrollo de la enferlnedad, mientras que se evit6 la infeccibn en plantas que no fueron inoculadas como control. La altura de las plantas asi como estimaciones de 10s porcentajes de infecci6n y grado de infecci6n de toda la planta se tomaron cada dos semanas durante cuatro meses a1 final del invierno y pritnavera de 1991 y 1992. La altura final de las plantas y su peso fiesco se tomason cada afio a1 final del experimento. Luego se calcularon las areas bajo las cusvas de progreso de la enfermedad (AUDPC) para la altura de la planta, porcentaje de.infecci6n y el grado de infeccion de la enferlnedad. Se efectuaron ansilisis de variancia y comnparaciones con un solo-grado-de-libertad para interpretas 10s resultados de todas las variables de respuesta. Estos anlilisis indicaron la presencia de cuatro razas patogknicas de la roya cafe en el experiinento. Estos resultados junto con las observaciones de campo condujeron a la hip6tesis de que las variantes patogenicas de P. melanocephala se desarrollan dentro de una regi6n paralelamente con el desarrollo de nuevos cultivares comerciales de caiia de azucar. La diversificaci6n de cultivares comerciales y la identificacion de mecanistnos de resistencia horizontal podrian conducir a inenores perdidas debidas a la roya caf6 en la industsia de caiia de azucar de la Florida.





























