-
7/28/2019 Signature of ocean warming in global fisheries
catch.pdf
1/5
LETTERdoi:10.1038/nature12156
Signature of ocean warming in global fisheries catchWilliam W.
L. Cheung 1, Reg Watson 2 & Daniel Pauly 3
Marine fishes and invertebrates respond to oceanwarming
throughdistribution shifts, generally to higher latitudes and
deeper waters.Consequently, fisheries should be affected by
tropicalization of catch 14 (increasing dominance of warm-water
species). However,a signature of such climate-change effects on
global fisheries catchhasso farnot been detected.Herewe
reportsuchan index,the meantemperature of thecatch (MTC), that is
calculated from the averageinferred temperature preference of
exploited species weighted by their annual catch. Our results show
that, after accounting for theeffects of fishing and large-scale
oceanographic variability, globalMTC increased at a rate of 0.19
degrees Celsius per decade between1970 and 2006, and non-tropical
MTC increased at a rate of 0.23
degreesCelsiusper decade.In tropicalareas,MTCincreased initially
because of the reduction in the proportion of subtropical
speciescatches, but subsequently stabilized as scope for further
tropicaliza-tion of communities became limited. Changes in MTC in
52 largemarineecosystems, covering themajority of the worlds
coastal andshelf areas, are significantly and positively related to
regionalchanges in sea surface temperature 5 . This study shows
that oceanwarming has already affected global fisheries in the past
four dec-ades, highlighting the immediate need to develop
adaptation plansto minimize the effect of such warming on the
economy and foodsecurity of coastal communities, particularly in
tropical regions 6,7 .
Assessing the effects of climate change on marine fisheries is
oneof the challenges for sustainable management of marine
ecosystems.Marine biota respond to ocean warming through changes in
distribu-
tions andabundance3,4,8
, phenology 9
andbodysize10
, leadingtoalterationofcommunitystructure 4,8 andtrophic
interactions 11, andultimatelyaffect-ing fisheries7. Studies
assessing the potential effects of climate changeon globalfisheries
under scenarios of greenhousegasemissions predicta large-scale
redistribution of maximum fisheries catch potential 12,13
and increased vulnerability of manycoastal fisheries to
climatechange,particularly in the tropics 6. Climate change effects
on some fisherieshave been detected14,15 . For example, the rapid
increase in catches of red mullet ( Mullus barbatus), a warm-water
species, around the UK issuggested to be relatedto ocean warming 15
. However, a signatureof theeffect of climate change on global
fisheries has so far not been demon-strated. Because marine
fisheries contribute to the economy and foodsecurity of many
coastal communities, fisheries responses to climatechange need to
be better understood to inform the development of effective
management and adaptation policies7.
Shifts in distributions of exploited stocks areexpected to
affect theiravailabilityto fisheries. Spatialdistributionsof marine
fishesand inver-
tebrates are strongly dependent on the relationship between
physio-logical optima and limits under different temperatures,
oxygen levelsand other biotic and abiotic conditions 16,17 .
Organisms living in tem-peratures outside their thermal optima
experience reduced aerobicscope, negatively affecting their growth
and reproduction, and ulti-mately reducing their abundance 18 .
Incontrast, some species may findthat areas previously unsuitable
for their survival have become morefavourable. Consequently, the
distribution margins and centroids of many marine fishes and
invertebrates shift following changes in oceanconditions 16 , with
a tropicalization of species compositions occurring as the ocean
warms1. Concomitantly, species with limited dispersalpotential or a
narrow range of temperature tolerance, such as in semi-enclosed
seas and tropics, will decrease in overall abundance18,19 .
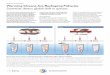
Werepresent the temperaturepreferenceof speciesin fisheries
catch
by an index called mean temperature of the catch (MTC) (Fig. 1).
Thisglobal index builds on previous use of temperature preferences
toevaluate potential effects of climate change on fisheries locally
14,20 . Wepropose thatoceanwarmingleadsto increasedcatches
ofwarmer-waterspecies and decreased catches of colder-water
species, resulting in an
1 Changing Ocean Research Unit, Fisheries Centre, The University
of British Columbia, Vancouver, British ColumbiaV6T 1Z4, Canada. 2
Institute for Marine and Antarctic Studies, University of
Tasmania,Taroona, Tasmania 7001, Australia. 3 Sea Around Us
Project, The University of British Columbia, Vancouver, British
Columbia V6T 1Z4, Canada.
T r o p
i c a
l
T e m p e r a
t e
S u
b t r o p
i c a
l
Temperature ( C)
M T C ( C )
M T C ( C )
M T C ( C )
MTC
MTC MTC MTC
Spp. 1Spp. 2Spp. 3
Spp. 4
Multi-decadal ocean warming (SST)
0 5 10 15 20 25 30 0 5 10 15 20 25 30 0 5 10 15 20 25 30
Year
Oceantemperature
Latitude Decade 1 Decade 2 Decade 3
0 5 10 15 20 25 30
0 5 10 15 20 25 30
5 10 15 20 25 300
5 10 15 20 25 300
5 10 15 20 25 300
10 15 20 25 300 5
Figure 1 | Changes in catch speciescomposition in relation to
oceanwarming and the resulting changesin MTC. Species distributions
arerelated to ocean temperature(coloured bars) and
temperaturepreferences of the exploited species(grey curves).
Increase and decreasein abundance due to ocean warming are
indicated by green curves and thereduction in area under the grey
curves, respectively. The verticalblack and red arrows represent
MTCin the initial and subsequent decades,respectively. D MTC
represents thedifference in MTC relative to theinitial decade.
Species localextinction and invasion because of warming are
indicated by red andgreen dotted curves, respectively.Theexpected
changes in MTC over timeare shown on the right.
1 6 M AY 2 0 1 3 | V O L 4 9 7 | N AT U R E | 3 6 5Macmillan
Publishers Limited. All rights reserved201 3
http://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/doifinder/10.1038/nature12156
-
7/28/2019 Signature of ocean warming in global fisheries
catch.pdf
2/5
increase in MTC. We further propose that the general
relationshipbetween MTC and sea surface temperature (SST) is
modified in thetropics, where catch compositions consist of both
tropical and sub-tropical species. Warming would cause the initial
decrease in the pro-portionof catchesof subtropicalspecies,
resultingin increases in MTCbecause of the poleward shift in the
equatorial range boundary of subtropical species. Such initial
increases in MTC would then stabilizeas catches become dominated by
tropical species, but the scope forfurther tropicalization is
limited 16 . Further warming, to levels that
exceed the temperature preference and tolerance of tropical
species,is then expectedto reducetheirabundance 12 without
changesin MTC.
We calculated MTC as the average of the inferred
temperaturepreference of exploited species (990 species in total)
weighted by theirannual catch between 1970 and 2006, for 52 large
marine ecosystems(LMEs) that account for most of the worlds
fisheries (Methods). Toattribute changes in MTC to ocean warming,
we used a generalizedadditive mixed model (GAMM) with an ensemble
of different combi-nations of factors to account for the potential
effects of large-scale
*
Rate of change in SST ( C yr 1 )
0.04 0.02 0.02 0.04 0.06 0.080
1.0
0.5
2.0
a
b
c
2.0
0.5
1.0
1.0
0.0
1.5
1.0
0.0
1.0
13
1
13
13
02
2
0.0
1.01.5
0.01.5
0.40.0
0.4
0.8
0.0
0.6
0.4
0.20.6
0.4
0.20.6
0.4
0.0
0.3
1.0
0.5
1.5
1.00.00.6
0.40.0
0.4
0.2
0.4
0.0
0.2
0.20.0
0.3
0.3
0.0
2
10
2
20
2
20
13
1
0.2
0.2
0.50.5
0.0
1.0
1.5
0.5
3
1
1
12
1
1
131
13 10
2
2
0
224
02
0.2
0.2
0.2
0.00.6
0.0
0.6
0.6
0.4
0.0
1.5
1.5
0.0
10
2
1
1.2
0.2
0.4
1.0
0.5
1.0
0.0
1
21
1
21
0.2
0.0
0.0
24
20
24
20
0.0
0.2
0.2
0.0
0.3
Figure 2 | Changes in MTC andSST of 52 LMEs between 1970
and2006. Changeovertime( x axis;yearsbetween 1970 and 2006) in
MTCanomalies relative to the mean of thetime series ( y axis; u C).
Rate of SSTchange is shown by the colour scale.a , Eurasia; b, the
Americas; c, Asiaand Oceania. Red lines represent
tropical LMEs. To highlight thenonlinear trends of MTC
anomaliesin this figure, each MTC time serieswas fitted with a
spline smoothing function using GAMM: grey line,mean; shaded area,
95% confidenceinterval.
RESEARCH LETTER
3 6 6 | N AT U R E | V O L 4 9 7 | 1 6 M AY 2 0 1 3Macmillan
Publishers Limited. All rights reserved201 3
-
7/28/2019 Signature of ocean warming in global fisheries
catch.pdf
3/5
oceanographic variability and fishing effort, both of which are
knownto affect fisheries strongly 21,22 . In addition, we repeated
the analysisusing a subset of 698 species with their temperature
preference pre-dicted from the above and an alternative approach to
species distri-bution modelling (Supplementary Information).
Overall, theMTC in the52 LMEs from
theensembleGAMManalysisincreased at an average rate of 0.19u C per
decade between 1970 and2006 (Fig. 2), andthe MTCin non-tropical
LMEs increasedat a rate of 0.23 u C per decade. Specifically, MTC
increased consistently in LMEsin the northeast Pacific Ocean (0.48
u C per decade) and the northeastAtlantic Ocean (0.49 u C per
decade), where SST increased by 0.20 and0.26 u C per decade,
respectively. For each LME, and on the basis of theresults fromthe
ensemble GAMMs, we ranked the relative importanceof SST, fishing
effort and large-scale oceanographic indices as factorsaccounting
for the changes in MTC over time. The results suggestedthat there
was no significant difference between these factors, in
termsofthefrequency distribution ofLMEs withdifferent
rankings(KruskalWallis rank-sumtest, P . 0.05; Supplementary
Fig.1). However, therewere regionaldifferences in their
importance(SupplementaryTable1).
MTC changes in the52 LMEs were significantly related to
therateof SST changesbetween1970 and 2006 (P , 0.05; Fig. 3 and
Supplemen-tary Table 2). The relationships remained significant
when the bestmodels (lowest Akaike information criteria) were
selected instead of the model ensemble (Supplementary Table 2) and
when the twenty-fifthand seventy-fifth percentilesof the
temperaturepreferenceprofiles
ofthe species, insteadofthe median values, wereusedin
calculatingtheMTC(SupplementaryFig. 2).Using thealternativespecies
distributionmodelling approach to calculate species temperature
preference hasno significant effect on the relationship between
changes in MTC andSST (P . 0.1; Methods and Supplementary
Information).
TropicalLMEs showed, overall, an asymptoticpattern
ofMTCchange,with a reduction in the proportion of subtropical
species in catches(Fig. 3). Average MTC from 14 tropical LMEs
increased rapidly between 1970 and 1980 by around 0.6u C, and
subsequently stabilizedat around 26 u C (Fig. 3). Moreover, the
average temperature tolerancerange, calculated from thedifferencein
temperature betweenthe twenty-fifth and seventy-fifth percentilesof
the temperaturepreferenceprofileof each species (Supplementary
Information), decreased significantly
during the period. Subtropical species had wider temperature
tole-rances than did tropical species17 (Supplementary Fig. 3),
providing further evidence to support the hypothesis that fisheries
catches in thetropics are becoming inimicalto subtropicalspecies.
Moreover,averageSST change in the tropical LMEs increased
consistently, at a rate of 0.14 u C per decade.
A number of factors indicate that MTC is a valid proxy to
examinechanges in composition of catches in a region in relation to
the tem-perature preference of the exploited animals. First, there
was no dif-ference (analysis of covariance (ANCOVA), P . 0.1;
Supplementary Table 4) between using catch data and scientific
bottom trawl survey data in the calculated rate of change in MTC in
North Sea (Fig. 4 andMethods). Because the scientific survey data
set can be viewed as areliable indicator of relative abundance of
animals in the ocean, ourresults therefore support the use of catch
data to detect climate changesignature in fisheries. However,
because the availability of survey-based data was limited to a few
well-studied regions23 , we were unableto use such data to analyse
global fisheries. The relationship betweenthe thermal preference of
the species and the environmental temper-ature also corroborates
the results of regional-scalestudies using survey data14,24 .
Second, fishing efforts in many LMEs have been increasing
con-tinuously since the1970s. This coincided with increases in SST,
result-ing in strong correlation between changes in SST and fishing
effort insome LMEs. However, there is no evidence that fishing
systematically alters MTC. Specifically, significant but weak
relationships betweenmaximum body size (positively related to
vulnerability to fishing, ingeneral) and the temperature preference
of exploited species is foundin only 19 LMEs, with the majority
(13) of them showing a positiverelationship,suggesting that the
increasing MTCtrend wasnota result
0.15
0.10
0.05
0.00
0.05
0.10
0.15
0.01 0.01 0.03 0.05
R a
t e o
f c
h a n g
e i n M T C ( C y r
1 )
Rate of change in SST ( C yr 1 )
1:1 line
0.6
0.4
0.2
0.0
0.2
0.4
0.6
25.0
25.2
25.4
25.6
25.8
26.0
26.2
26.4b
a
1970 1980 1990 2000
M T C ( C )
S S T an
om
al y
( C )
Year
MTC
SST
Figure 3 | Relationship between rates of change in MTC and SST
between1970and2006 in52 LMEs. a , Rate of change of MTC was
calculated from theensemble GAMM in each LME(slope of
linearregression, P , 0.005,R2 5 0.19
(coefficient of determination)). The black line shows the mean
and the grey lines delineate the 95% confidence interval.b, Changes
in MTC (red) and SSTanomalies (grey) in tropical LMEs. The dashed
lines are fitted with asymptoticand linear models for the MTC and
SST anomalies, respectively.
1.5
1.0
0.5
0.0
0.5
1.0
1.5
1970 1980 1990 2000Year
From survey data
From catch
M T C a n o m a
l y ( C )
Figure 4 | Comparison of MTC calculated from fisheries catch
data andrelative abundance calculated from scientific survey data.
MTC is expressedas an anomaly relative to the mean of the time
series, and relative abundance isexpressedrelative tothe average
between 1970 and2006. Thetypeof data used hadno effect on the rate
of change in MTC (ANCOVA, P . 0.1).
LETTER RESEARCH
1 6 M AY 2 0 1 3 | V O L 4 9 7 | N AT U R E | 3 6 7Macmillan
Publishers Limited. All rights reserved201 3
-
7/28/2019 Signature of ocean warming in global fisheries
catch.pdf
4/5
of the depletion of large fish by fishing that was reported by
many fisheries25 (Supplementary Information).
Third, our findings are not an artefact of the quality of
fisheries andenvironmentaldata, whichvarybetween regions. In four
of fiveareas inwhich misreporting of the level of catch was
previously estimated, theestimated rates of changein MTC were not
significantly different fromthosecalculated from data sets
withoutcorrectionfor misreporting. Forthe only area that showed a
significant difference in rate of MTC
change, MTCs calculated from both data sets show significant
increaseandthequalitative trend thusremains robust. Serious
over-reportingof fisheriescatches byChinasince themid 1980shave
been documented 26 .Recalculating MTC in the South China Sea, the
East China Sea and theYellow Sea using catch data from 19701985
increased the goodnessof fit of its significant relationship with
SST. The use of different SSTclimatology to calculatespecies
temperaturepreference, moreover,hadno significant effect on the
findings (Supplementary Table 3).
Although fisheries catch statistics are usually not reported at
stock level, an analysis using simulated species and stock
distributions, tem-perature preferencesand,subsequently, MTCchanges
showedthat theexistence of stock structure andstock-specific
temperaturepreferencesdid not significantly bias the rate of change
of MTC calculated fromaggregated species-level data (P . 0.1;
Supplementary Information).
Potential phenotypicand evolutionary responsesof species to
warming,if occurring, would reduce the rate of change in MTC. The
accuracy of theSST data setvaried between regions as a result of
differentdensitiesofmeasurements.Given these uncertainties,
thefactthat ouranalysis isable to detect a signature
ofclimatechangein globalfisheries highlightsthe robustness of the
underlying relationship.
Overall, ourresults suggest that changein thecomposition of
marinefisheriescatch is significantlyrelated to temperaturechange
in theocean,with an increasing dominance of catches of warmer
waters species athigher latitudes anda decrease in theproportion of
catches of subtropicalspecies in the tropics. Such changes in catch
composition have directimplicationsforcoastal fishing communities,
particularly thosein tropicaldeveloping countries, which tend to be
socioeconomically vulnerable totheeffects of climate change6.
Continuedwarmingin the tropics to a level
that exceeds the thermal tolerance of tropical species may
largely reducecatch potential in this region 12. This highlights
the need to prioritizeresources to develop adaptation plans
immediately to minimize impactson the economy and food security of
tropical coastal communities.
METHODS SUMMARYThemappedfisheries landingsdataweresourcedfromthe
SeaAroundUs project 27 ,which utilized a rangeof
sourcesincludingthe Food
andAgricultureOrganizations(FAO)fisheriesdatabase supplementedby
regional datasets (SupplementaryInfor-mation). We inferred the
thermal preference of each species on the basis of itsmodelled
distribution 28,29 (Supplementary Table 5 and Supplementary
Information).The MTC was computed fromthe average inferred
temperature preference of 990species of exploited fishes and
invertebrates weighted by their annual catch:
MTCyr ~P ni T iC i, yrP n
iC i, yr
1
Here C i ,yr is catch of species i in a specific region in year
yr, T i is the mediantemperature preference of species i and n is
the total number of species. Similarly,MTC was calculated using
survey data in the North Sea from equation (1) with C i ,yrreplaced
by the relative abundance for species i in year yr. Relative
abundance
datawereobtainedfromtheICESInternationalBottomTrawlSurvey.A totalof
55 speciescommon to both the catch database and the survey data set
were included in thisanalysis.We tested theeffectof the useof catch
or surveydata in estimating changesin MTC over time using ANCOVA,
accounting for temporal autocorrelation.
Weusedthe GAMM(R packageMuMIn)witha full model,MTC 5 s(Effort)
1s(OI) 1 Year,where sisa splinesmoothingfunction (Supplementary
Information)with a temporalautocorrelation term (R functioncorAR1),
allowingfor a lagof upto three years in the response of MTC to
these two variables and the time effectexpressed by Year. To
estimate the average rate of change in MTC from thecoefficients of
this time effect, we did not include a spline smoothing functionfor
Year. Fishing effort data were sourced from the Sea Around Us
project30
(Supplementary Information).
Full Methods and any associated references are available in the
online version ofthe paper .
Received 13 October 2012; accepted 5 April 2013.
1. Cheung, W. W. L. et al. Climate change induced
tropicalization of marinecommunities in Western Australia. Mar.
Freshw. Res. 63, 415427 (2012).
2. Wernberg, T. etal. An extreme climaticeventalters marine
ecosystem structure ina global biodiversity hotspot. Nature Clim.
Change 3, 7882 (2013).
3. Perry, A. L., Low, P. J., Ellis, J. R. & Reynolds, J. D.
Climate change and distributionshifts in marine fishes. Science
308, 19121915 (2005).
4. Dulvy,N.K.etal. Climatechangeand deepening of theNorthSea
fishassemblage:a biotic indicator of warming seas. J. Appl.
Ecol.45, 10291039 (2008).
5. Belkin, I. M. Rapid warming of large marine ecosystems. Prog.
Oceanogr. 81,207213 (2009).
6. Allison, E.H.et al. Vulnerability of national economies to
the impacts of climatechange onfisheries. Fish Fish. 10, 173196
(2009).
7. Sumaila,U.R.,Cheung,W.W. L.,Lam,V. W.Y.,Pauly,D.&
Herrick,S.Climatechangeimpacts on the biophysics and economics of
world fisheries. NatureClim. Change1, 449456 (2011).
8. Simpson,S. D. etal. Continental shelf-wide responseof a
fishassemblageto rapidwarming of the sea. Curr. Biol.21, 15651570
(2011).
9. Edwards, M. & Richardson, A. J. Impact of climate change
on marine pelagicphenology and trophic mismatch. Nature 430, 881884
(2004).
10. Cheung, W. W. L. et al. Shrinking of fishes exacerbates
impacts of global oceanchanges on marine ecosystems. Nature Clim.
Change 3, 254258 (2013).
11. Harley, C. D. G. Climate change, keystone predation,and
biodiversityloss. Science334, 11241127 (2011).
12. Cheung, W. W. L. et al. Large-scale redistribution of
maximum fisheries catch
potential in the global ocean under climate change. Glob. Change
Biol. 16, 2435(2010).13. Blanchard, J. et al. Potential
consequences of climate change for primary
production and fish production in large marine ecosystems. Phil.
Trans. R. Soc. B367, 29792989 (2012).
14. Pinsky,M. & Fogarty,M. Lagged social-ecological
responses to climate and rangeshifts in fisheries. Clim. Change
115, 883891 (2012).
15. Cheung, W. W. L., Pinnegar, J., Merino, G., Jones, M. C.
& Barange,M. Review ofclimate change impacts on marine
fisheries in theUK and Ireland. Aquat.Conserv.Mar. Freshwat.
Ecosyst. 22, 368388 (2012).
16. Sunday,J. M., Bates, A. E. & Dulvy, N. K. Global
analysis of thermal tolerance andlatitude in ectotherms. Proc. R.
Soc. Lond. B278, 18231830 (2011).
17. Po rtner, H. O. & Farrell, A. P. Physiology and climate
change. Science 322,690692 (2008).
18. Po rtner,H. O. & Knust,R. Climatechangeaffects
marinefishesthrough theoxygenlimitation of thermal tolerance.
Science 315, 9597 (2007).
19. Ben Rais Lasram, F. et al. The Mediterranean Sea as a
cul-de-sac for endemicfishes facing climate change. Glob. Change
Biol. 16, 32333245 (2010).
20. Collie,J. S.,Wood,A. D. & Jeffries, H. P. Long-term
shifts in thespecies composition
of a coastal fish community. Can. J. Fish. Aquat. Sci.65,
13521365 (2008).21. McGowan, J. A., Cayan, D. R. & Dorman, L.
M. Climate-ocean variability andecosystem response in the northeast
Pacific. Science 281, 210217 (1998).
22. Pauly,D. etal. Towardssustainability inworldfisheries.
Nature 418, 689695(2002).23. Costello, C. etal. Statusand solutions
forthe worldsunassessedfisheries. Science ,
(2012).24. Howell, P. & Auster, P. J. Phaseshift in an
estuarine finfishcommunityassociated
with warming temperatures. Mar. Coast. Fish. 4, 481495
(2012).25. Cheung, W. W. L., Watson, R., Morato, T., Pitcher, T. J.
& Pauly, D. Intrinsic
vulnerability in the global fish catch. Mar. Ecol. Prog. Ser.
333, 112 (2007).26. Watson,R. & Pauly,D. Systematicdistortions
inworldfisheriescatchtrends. Nature
414, 534536 (2001).27. Watson, R., Kitchingman, A., Gelchu,A.
& Pauly,D. Mapping global fisheries:
sharpening our focus. Fish Fish. 5, 168177 (2004).28. Jones,M.,
Dye,S.,Pinnegar,J., Warren,R. & Cheung,W. W. L. Modelling
commercial
fish distributions: prediction and assessment using different
approaches. Ecol.Modell. 225, 133145 (2012).
29. Rockmann, C., Dickey-Collas, M., Payne,M. R. & van Hal,
R. Realized habitats ofearly-stage North Sea herring: looking for
signals of environmental change. ICES J. Mar. Sci.68, 537546
(2011).
30. Watson, R. A. et al. Global marine yield halved as fishing
intensity redoubles. FishFish. (2012).
Supplementary Information is available in the online version of
the paper .
Acknowledgements W.W.L.C. acknowledges funding support from the
NationalGeographic Society and the Natural Sciences and Engineering
Research Council ofCanada. R.W. and D.P. were supported by the Pew
CharitableTrust through the SeaAround Us project. We are grateful
to S. Pauly and D. Palomares for reviewing themanuscript and
providing the Aquamap distributions from FishBase,
respectively.
Author Contributions W.W.L.C. and D.P. designed the study.
W.W.L.C. conducted theanalysis. R.W.and D.P.providedthe
fisheriescatchand effortdata fromthe SeaAroundUs project. All
authors contributed to the writing of the manuscript.
Author Information Reprints and permissions information is
available atwww.nature.com/reprints . The authors declare no
competing financial interests.Readersare welcome to commenton the
online version of thepaper . Correspondenceand requests for
materials should be addressed to
W.W.L.C.([email protected]) .
RESEARCH LETTER
3 6 8 | N AT U R E | V O L 4 9 7 | 1 6 M AY 2 0 1 3Macmillan
Publishers Limited. All rights reserved201 3
http://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/reprintshttp://www.nature.com/doifinder/10.1038/nature12156mailto:[email protected]:[email protected]://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/reprintshttp://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/doifinder/10.1038/nature12156http://www.nature.com/doifinder/10.1038/nature12156
-
7/28/2019 Signature of ocean warming in global fisheries
catch.pdf
5/5
METHODSGlobal catch and effort data. We use the terms fisheries
catch and fisherieslandings interchangeably, but strictly these
both refer here to landings; the weightof marine fishes and
invertebrates that were caught and retained. The mappedfisheries
landings and effort data were sourced from the Sea Around Us
project27
(Supplementary Information). Landings data were not used to
estimate temper-ature preference ofthe species, andtemperature data
were notusedto infer speciesdistributions. Effort data were
standardized and collated on the basis of fishing boat engine power
(watts) and fishing days30 (Supplementary Information).Inferring
species thermal preference. We inferred the thermal preference of
each species from its modelled distribution (Supplementary Table
5). First, wemodelled the present (19702000) distribution of each
species using an algorithmdevelopedby theSea AroundUs project
anddocumented inref. 28.The algorithmestimated the relative
abundance (on a 309 3 309 latitudelongitude grid) of aspecies in
each spatial cell. Input parameters for each species considered in
themodel included the species maximum and minimum depth limits,
northern andsouthern latitudinal range limits, an index of
association to major habitat types(seamounts, estuaries, inshore,
offshore, continental shelf, continental slope andthe abyssal) and
known occurrence boundaries. For pelagic species, seasonal(summer
and winter) distributions wereconsidered. The parametervalues of
eachspecies were derived from data in online databases, mainly
FishBase (http://www.fishbase.org ) and SeaLifeBase
(http://www.sealifebase.org ). Catch and tem-perature data were not
used to model the current species distributions. Eachmodelled
species distribution was normalized and overlaid over the SST
climato-logy from the Hadley Centre SST data set for 1970200029 .
The predicted speciesdistributionis considered to be representative
of thedistributionof relativeabund-ance. The temperature preference
profile at SST bini ( pi) of each species wascalculated from the
total relative abundance, K i , and range area, Ai , at SST
bini(Supplementary Information): pi 5 (K i/ Ai)/P i (K i/ Ai).
The median and the twenty-fifth and seventy-fifth percentiles of
the temper-ature preference were calculated. The modelled species
distribution and temper-ature preference represents an average of
the species life stages. The analysis wasrepeated with alternative
estimate climatologies for 19511960 and 19912000.The choice of
climatology has no significant effect (ANCOVA, P . 0.1) on
therelationship between rate of change in MTC and rate of change in
SST(Supplementary Table 3). The analysis is also repeated using an
alternativeapproach to species distribution modelling
(Supplementary Information).Calculation of MTC. The MTC was
computed from the average inferred tem-perature preference of 990
species of exploited fishes and invertebrates weightedby their
annual catch:
MTCyr ~P ni T iC i, yrP ni C i, yr
1
Here C i ,yr is catch of species i in a specific region in year
yr, T i is the mediantemperature preference of species i and n is
the total number of species.
In the North Sea LME, MTC was calculated from equation (1) with
C i ,yrreplaced by the relative abundance for species i in year yr.
Relative abundancedata were obtained from the ICES International
Bottom Trawl Survey. A total of
55 species that coexist in both the catch database and the
survey data set wereincluded in this analysis. We tested whether
the changes in MTC calculated fromcatchdata and scientific survey
datawere different, using ANCOVA and account-ing for temporal
autocorrelation.Large-scale oceanographic indices. Indices
representing large-scale oceano-graphic conditions for each of the
six ocean basins were included in this study (Supplementary
Information). For LMEs in the Pacific Ocean, their relationshipwith
the Pacific decadal oscillation was used. For LMEs in the North
Atlantic andSouth Atlantic, the North Atlantic oscillation index
and, respectively, the dipoleindex were used. For LMEs in the
Indian Ocean, the Indian Ocean dipole index was used. For LMEs in
the Arctic, the summer sea-ice extent was used. For LMEsin the
Southern Ocean, the Antarctic Oscillation index was used.Accounting
for the effects of fishing and large-scale oceanographic
indices.For changes in MTC in each LME, we accounted for the effect
of changes in totalfishing effort (E ) and the corresponding
large-scale oceanographic index (OI). Wethenuseda GAMM (R package
MuMIn)witha full model,MTC 5 s(Effort 1 s(OI)1 Year, where s
represents a spline smoothing function (Supplementary
Infor-mation), allowingfor a lagof upto three years inthe response
of MTC to these two variablesand the time effect Year, and
includingan autoregressiveterm to accountfortemporal
autocorrelation. To estimate theaveragerateof change inMTC fromthe
coefficients of the time effect, we did not include a spline
smoothing functionfor the variable Year. These models were analysed
using a multi-model analysisframework (R package MuMIn). There were
significant autocorrelations of 1-yrlag in the MTC time series for
all LMEs (P , 0.05). Rates of MTC changes (coef-ficientof Year)
wereestimated using a multi-modelensemblemean withweighting (W )
calculated from W i 5 exp[(AICmin 2 AICi)/2], where AICmin is the
bestmodel (that with minimum Akaike information criterion) and AIC
i was basedon alternative model i. In addition, the rates of MTC
change were also estimatedfrom the model with the lowest Akaike
information criterion. The relationshipbetween the rates of change
in MTC and SST remained significant with thesealternative
estimates.
LETTER RESEARCH
ll bl h d ll h d201
http://www.fishbase.org/http://www.fishbase.org/http://www.sealifebase.org/http://www.sealifebase.org/http://www.fishbase.org/http://www.fishbase.org/