Embed Size (px)
Citation preview

SINGLE-MOLECULE ANALYSIS
A ribosome in action Susanne brakmann
The manufacture of proteins by ribosomes involves complex interactions of diverse nucleic-acid and protein ligands. Single-molecule studies allow us, for the first time, to follow the synthesis of full-length proteins in real time.
Protein synthesis involves a complex interplay of various cellular components. Ribosomes are the cell’s protein-production factories, and interact with messenger RNA (the template), amino-acylated transfer RNAs (which act as adaptors between mRNA and amino-acid residues) and diverse co-factors (for the ini-tiation of synthesis, elongation of the nascent chain and release of the mature polypeptide). In this issue, Uemura et al.1 report the use of an extremely sensitive single-molecule detection technique to observe this process at unprec-edented resolution: the stepwise synthesis of a single protein (page 1012).
Ribosomes are evolutionarily conserved molecular nanomachines with a diameter of about 25 nanometres and a molecular weight of around 2.5 megadaltons. In functional terms, they are amino-acid polymerase enzymes with an RNA ‘heart’. They accelerate the rate of pro-tein synthesis by at least one millionfold, owing exclusively to entropic effects that involve the positioning of aminoacyl-tRNAs, the shielding of the reaction from bulk solvent and the organ-ization of their own active site2,3. Ribo somes also check the quality of their polypeptide products — inaccurate amino-acid sequences
could result in an altered three-dimensional protein structure and even cellular toxicity.
Structural analyses of functional ribosome complexes have formed the basis of a consist-ent biochemical model for the mechanism of protein synthesis4, suggesting that this process depends on large-scale conformational changes in the ribosome. But the nature, timescale and magnitude of these dynamic changes have remained undefined.
Traditional biophysical investigations of dynamic molecular processes yield ensem-ble-averaged data. These may hide crucial information owing to the presence of asyn-chronous conformational states and varying stages of the enzymatic reaction. By contrast, single-molecule studies circumvent the need for synchronous molecular behaviour and the use of uniform samples, thus allowing the identification and direct characterization of transient, rare and so physiologically relevant events. With the use of sensitive fluorophores, single-molecule spectroscopy and force-based techniques, researchers can observe changes in distance on the subnanometre scale with a temporal resolution in the millisecond range.
To study the details of translation, for
example, fluorophores can be chemically coupled to tRNA molecules. Consequently, after bind-ing to the ribosome, the fluorescently labelled tRNAs ‘report’ with high sensitivity on how they are selected by the ribosome, their motion within the ribosome and even conformational changes in the ribosome.
Indeed, single-molecule analyses of fluo-rescently labelled tRNAs have allowed the monitoring of tRNA selection and (single) peptide-bond formation in real time, as well as studies of tRNA dynamics on the ribosome during elongation, and single-molecule force measurements have helped to determine the strength of ribosome–mRNA interactions5. Although such studies have revealed previ-ously unknown details of tRNA selection and catalytically important ribosomal states, the findings were limited to the initial stages of translation. A desirable extension of this is the investigation of several rounds of peptide-bond formation — for instance, study of the continu-ity of synthesis (processivity) or its accuracy (translational fidelity).
The first study to attempt this6 used specifi-cally designed, single hairpin-shaped mRNA sequences tethered by their ends to optical tweezers, and determined their transloca-tion along the ribosome. The translocation measured three mRNA bases (one codon) and occurred as a series of movement–pause–movement events, with pause durations ranging from a fraction of a second to several seconds. The ‘dwell’ time at each codon was the time taken for tRNA selection and peptide-bond formation, which can be correlated to the sequence context.
Uemura and colleagues1 now take these methods even further. Their approach is based on a set of tRNAs labelled with distinct fluoro-phores, allowing their immediate identifica-tion after ribosomal binding and so real-time ‘reading’ of the underlying mRNA sequence. By watching tRNA substrates instead of the template mRNA, the authors were able to fol-low tRNA binding and transit on individual translating ribosomes during multiple rounds of chain elongation in the synthesis of short peptides (4–13 amino acids long). Moreover, they showed that only two tRNA molecules simultaneously bind to the ribosome, allow-ing them to explore the mechanism of tRNA transit in relation to other reactants (Fig. 1).
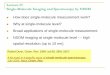
What is of utmost importance is that Uemura et al. were able to study translation at physiologically relevant (micromolar) con-centrations of tRNA and other factors using extremely small reaction vessels known as zero-mode waveguides, which were gener-ated from nano photonic metal films and illu-minated from below by a laser. The resulting observation volumes of 10–21 litres were there-fore occupied by, on average, not more than one molecule (Fig. 1). Because the duration of binding of the correct fluorescently labelled tRNA to the ribosome was much longer than the timescales associated with the freely
Figure 1 | Close-up view of a zero-mode waveguide vessel. Uemura and colleagues’ set-up1 consisted of a nanostructured metal film called a zero-mode waveguide, containing circular holes of diameter 50–200 nanometres. Using this, they analysed sample volumes as small as 10–21 litres to detect individual fluorescent tRNA molecules against the background signal from freely diffusing tRNA molecules. The binding of fluorescently labelled, cognate tRNA molecules — each labelled with a different fluorophore — within the ribosomal active site causes an elevation in fluorescent emission on the corresponding colour channel.
New polypeptide chain
mRNA
Fluorophores
tRNA
Ribosome
Zero-modewaveguide
Laserexcitation
Emission
Active site
Codon
987
NATURE|Vol 464|15 April 2010 NEWS & VIEWS
© 20 Macmillan Publishers Limited. All rights reserved10

diffusing tRNA molecules, genuine signals were reliably distinguished from low and con-stant background.
The real-time monitoring of single trans-lating ribosomes is truly a breakthrough for studying the process of translation, how its accuracy is controlled and the conformational changes that it involves in ribosomes. It should now be only a small step to elucidating the cor-relation between tRNA binding and changes in ribosomal conformation, to probing the dynamics of translational control, and to study-ing the synthesis and folding of full-length pro-teins in real time. Beyond that, Uemura and colleagues’ impressive demonstration1 of how their powerful technique (originally developed for ‘next-generation’ DNA sequencing7) can be used to answer biological questions will play
a decisive part in studying other processes, including RNA sequencing, the sequencing of genomic sites modified by methyl groups, and various time-dependent regulatory processes. ■
Susanne Brakmann is in the Department of Chemical Biology, Technische Universität Dortmund, D-44227 Dortmund, Germany.e-mail: [email protected]
1. Uemura, S. et al. Nature 464, 1012–1017 (2010).2. bieling, P., beringer, M., Adio, S. & Rodnina, M. V. Nature
Struct. Mol. Biol. 13, 423–428 (2006).3. Schmeing, T. M. & Ramakrishnan, V. Nature 461,
1234–1242 (2009).4. Ehrenberg, M. http://nobelprize.org/nobel_prizes/
chemistry/laureates/2009/cheadv09.pdf (2009).5. Marshall, R. A., Echeverría Aitken, C., Dorywalska, M. &
Puglisi, J. D. Annu. Rev. Biochem. 77, 177–203 (2008).6. Wen, J.-D. et al. Nature 452, 598–603 (2008).7. Eid, J. et al. Science 323, 133–138 (2009).
INFORMATION SCIENCE
Guaranteed randomnessValerio Scarani
You have received a device that is claimed to produce random numbers, but you don’t trust it. Can you check it without opening it? In some cases, you can, thanks to the bizarre nature of quantum physics.
From the stock market to the weather, we are surrounded by processes that are best described by unpredictable, random elements. But randomness is a notoriously difficult property to test. Worse still, when it is used to protect personal details, the random elements must be private. On page 1021 of this issue, Pironio and co-workers1 describe a method for obtaining numbers that are guaranteed to be random and private from an unknown process, provided that the numbers are certi-fied as being derived from measurements on quantum systems. Some may surmise that this claim is trivial, because quantum physics has long been known to produce random-ness. Some may even surmise that this claim is wrong, on the basis of the idea illustrated by the Dilbert comic strip ‘Tour of Accounting’ reproduced here. But the result is both new and correct.
Starting with the comic strip, Dilbert’s guide is uttering a scientific truth: the list 9, 9, 9, 9, 9, 9
is as valid an output of a generator of random numbers as is 1, 2, 3, 4, 5, 6 or 4, 6, 7, 1, 3, 8. In fact, in a long enough sequence of lists, any list of numbers should appear with the same frequency. Classic, ‘black-box’, tests of random-ness exploit this idea: they check for the relative frequencies of lists. But, in practice, no such test can distinguish a sequence generated by a truly random process from one generated by a suit-able deterministic algorithm that repeats itself after, for example, 1023 numbers. More over, in this way, whether the numbers are private cannot be checked: even if the sequence had initially been generated by a truly random pro-cess, it could have been copied several times, and the random generator may just be reading from a record.
It seems therefore that only a careful characterization of the process — that is, an ‘open-box’ test — can guarantee random-ness, especially of the private kind. But nature sometimes dares to go where abstract thinking
alone cannot. Pironio and colleagues1 take an alternative route for generating guaranteed, private randomness in their study, using one of the most remarkable phenomena of quantum physics: the violation of the ‘Bell inequalities’2, which has been observed in numerous differ-ent experiments in the past three decades3.
To understand what Bell inequalities are, first consider two quantum systems, for exam-ple two photons emitted by the same source and propagating away from one another. Con-sider further that a measurement is made of each of the photons, which are now spatially distant. For example, the polarization of each photon can be measured, assigning the value 0 if a photon is transmitted through a polar-izer and the value 1 if it is reflected. For certain specific sources, the outcomes of the two meas-urements are not independent. For instance, when the two polarizers are set in the same direction, only the pairs of outcomes (0, 1) and (1, 0) are observed: the photons are never both transmitted or both reflected. Such cor-relations between distant events are striking, raising the question of where the connection is. There is surely no communication between the two photons, because the signal would need to propagate faster than light. The only plausible hypothesis therefore is that the photons leave the source with a common ‘list of instructions’, which dictates the outcomes of each possible measurement.
Bell inequalities are criteria that, when applied, allow this latter hypothesis to be proved false. If the statistics of the measure-ment outcomes violate the inequalities, then the observed correlations cannot arise from a pre-established common list. Quantum corre-lations violate Bell inequalities and thus can-not come from a pre-established list either, and there is no classical mechanism that explains those correlations.
So how can random numbers be obtained using Bell inequalities? Following on from the example above, many pairs of photons are taken, and the measurement procedure is repeated: two sequences of 0s and 1s are produced, one at each measurement location. If these sequences violate Bell inequalities, they are guaranteed to be private random numbers: random, because there was no information about them before their generation; and private, because, given that the information did not exist, nobody else could have had access to it.
This line of qualitative reasoning has been known for many years. However, only recently have physicists been realizing and using its full power by stressing that Bell inequalities depend solely on the statistics of the observed outcomes and not on the description of the physical system or on the measurements that are carried out on it. In other
988
NATURE|Vol 464|15 April 2010NEWS & VIEWS
© 20 Macmillan Publishers Limited. All rights reserved10