Embed Size (px)
Citation preview

Somatostatin 2A Receptor Is Expressedby Enteric Neurons, and by Interstitial
Cells of Cajal and Enterochromaffin-LikeCells of the Gastrointestinal Tract
CATIA STERNINI,1,2,3* HELEN WONG,1 S. VINCENT WU,1,2 ROBERTO DE GIORGIO,1,2
MOON YANG,1,2 JOSEPH REEVE, JR.,1,2 NICHOLAS C. BRECHA,1,2,3
AND JOHN H. WALSH1,2
1Department of Medicine, CURE Digestive Diseases Research Center,University of California, Los Angeles, California 90073
2West Los Angeles Veterans Administration Medical Center, Los Angeles, California 900733Department of Neurobiology, University of California, Los Angeles, California 90095
ABSTRACTSomatostatin exerts multiple effects by activating distinct G protein-coupled receptors.
Here we report the cellular sites of expression of the somatostatin subtype 2A (sst2A) receptorin the rat enteric nervous system by using a C-terminus-specific, affinity-purified antiserumand immunohistochemistry. Antibody specificity was confirmed by the cell surface staining ofhuman embryonic kidney 293 cells expressing the sst2A receptor, the lack of staining of cellsexpressing the somatostatin subtype 2B receptor, and the abolition of staining by preincubat-ing the antiserum with the C-terminus peptide used for immunization, sst2A(361-369). The sst2Areceptor antibody recognized a broad 80 kDa band on Western blots of membranes preparedfrom cells transfected with sst2A receptor cDNA; following receptor membrane deglycosyla-tion, the antibody detected an additional 40 kDa band. In the enteric nervous system, the sst2Aantibody primarily stained neurons of the myenteric and submucosal plexuses, and abundantfibers distributed to the muscle, mucosa, and vasculature. Immunoreactive staining was alsoobserved in non-neuronal cells, including presumed interstitial cells of Cajal of the intestineand enterochromaffin-like cells of the stomach. Fibers expressing sst2A receptor immunoreac-tivity were often in close proximity to D cells of the gastric and intestinal mucosa.Colocalization of somatostatin and sst2A receptor immunoreactivities was not observed inendocrine cells nor in enteric neurons. Double-label immunohistochemistry revealed colocal-ization of sst2A and vasoactive intestinal peptide immunoreactivities in enteric neurons. Themultiple types of cells expressing the sst2A receptor, including enteric neurons and non-neuronal structures, in addition to the relationship between somatostatin and sst2A receptorelements, provide evidence that the sst2A receptor mediates somatostatin effects in thegastrointestinal tract via neuronal and paracrine pathways. J. Comp. Neurol. 386:396–408,1997. r 1997 Wiley-Liss, Inc.
Indexing terms: inhibitory neurons; neuronal pathway; paracrine pathway; vasoactive intestinal
peptide
Somatostatin (SOM) is a tetradecapeptide widely distrib-uted in the nervous and endocrine systems (Larsson, 1985;Epelbaum, 1986; Yamada and Chiba, 1989; Epelbaum etal., 1994). It is a well-established inhibitory neurotransmit-ter/neuromodulator in the nervous system, and it has awell-known neurohormonal regulatory role. SOM may alsoact as an autocrine and paracrine regulatory factor. In thedigestive system, SOM is expressed in neurons, fibers, andendocrine D cells (Costa et al., 1977, 1980; Ekblad et al.,1987, 1988). SOM inhibits endocrine and exocrine secre-
tion, electrolyte and enzyme secretion, electrolyte trans-port, intestinal nutrient absorption, muscle contraction,
Grant sponsor: NIH and VA Medical Research Funds; Grant number: DK41301, DK 17294.
*Correspondence to: Catia Sternini, M.D., CURE Digestive DiseasesResearch Center, West Los Angeles Veterans Administration MedicalCenter, Bldg. 115, Rm. 224, 11301 Wilshire Blvd., Los Angeles, CA 90073.E-mail: [email protected]
Received 23 January 1997; Revised 2 May 1997; Accepted 5 May 1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 386:396–408 (1997)
r 1997 WILEY-LISS, INC.

and cell proliferation (Reichlin, 1983; McIntosh, 1985;Yamada and Chiba, 1989).
SOM functions are initiated by the interaction withmembrane-bound receptors on multiple target tissues,perhaps by activating different receptor subtypes (Lewin,1992; Law et al., 1995). Five distinct SOM receptor geneshave been cloned (McIntosh, 1985; Yamada and Chiba,1989; Bruno et al., 1992; O’Carroll et al., 1992; Yasuda etal., 1992). Their corresponding receptors, named sst1 tosst5, belong to the superfamily of G protein-coupled recep-tors with seven transmembrane domains (Bell and Rei-sine, 1993; Hoyer et al., 1995; Patel et al., 1995; Reisineand Bell, 1995). They exhibit high affinity for the endog-enous ligands, SOM-14 and its N-terminal extended form,SOM-28. The sst2 receptor features two separate isoforms,the sst2A and sst2B receptors, as a result of alternativesplicing of the sst2 gene. These isoforms differ in theircytoplasmic C-terminal sequence (Vanetti et al., 1992).SST receptors are coupled to a variety of cellular effectorsand mediate the inhibition of adenylyl cyclase activity andof Ca21 conductance, the stimulation of protein tyrosinephosphatase, and the potentiation of K1 conductance (Lawet al., 1995; Patel et al., 1995; Reisine and Bell, 1995). SSTreceptors are characterized by marked differences in theiraffinities for synthetic SOM analogues (Patel and Srikant,1994; Rossowski and Coy, 1994; Law et al., 1995). Theseobservations indicate that distinct sst receptor subtypesare likely to mediate different SOM functions.
The overall distribution of SOM receptor binding siteshas been described in the brain and pituitary, and theexpression of all five sst receptor mRNAs has been demon-strated in the central nervous system (Breder et al., 1992;Krantic et al., 1992; Kong et al., 1994; Beaudet et al.,1995). Furthermore, a recent study has reported thecellular localization of sst2A receptors in the central ner-vous system (Dournaud et al., 1996). However, there is apaucity of studies characterizing SOM receptor bindingsites and sst receptor mRNA expression in the gastrointes-tinal tract (Reyl and Lewin, 1982; Arilla et al., 1984;Yamada et al., 1992), and their cellular distributions,which are required to understand the function and mode ofaction of SOM, are still unknown. The present study wasundertaken to: (a) identify the cellular sites of expressionof the sst2 receptor isoform, sst2A, in the rat enteric nervoussystem by using immunohistochemistry with an affinity-purified polyclonal antiserum directed to its C-terminalportion; and (b) elucidate the relationship between thesst2A receptor and its ligand, SOM, by means of double-labeling immunofluorescence approaches. Double-labelingwas also employed to determine whether enteric neuronsexpressing the sst2A receptor subtype contain immunoreac-tivity for the bioactive peptide, vasoactive intestinal poly-peptide (VIP), a marker for enteric secretomotor anddescending inhibitory motor neurons (Furness and Costa,1987; Furness et al., 1992). This study shows that the sst2Areceptor is localized to distinct populations of entericneurons that innervate a variety of targets as well as tonon-neuronal cells including interstitial cells of Cajal andenterochromaffin-like (ECL) cells.
MATERIALS AND METHODS
Antibody preparation
Polyclonal antibodies were raised in New Zealand whitefemale rabbits against a peptide with the amino acid
sequence corresponding to the C-terminal region of themouse sst2A (YLNGDLQTSI, sst2A(361-369), antiserum #9431).The peptide was synthesized on an Advanced ChemtechPeptide Synthesizer (Model 396; Louisville, KY), and waspurified by reverse-phase, high-pressure liquid chromatog-raphy. The composition of the peptide was verified by massspectroscopy, and purity was evaluated by high-perfor-mance capillary electrophoresis. Fractions with greaterthan 90% purity were pooled and lyophilized. The peptidewas conjugated to keyhole limpet hemocyanin (Calbio-chem, La Jolla, CA) using bisdiazotized benzidine, whichcouples through tyrosine residues. The efficiency of theconjugation was approximately 70% as determined byincorporation of iodinated peptide into the conjugate. Theconjugate was emulsified with an equal volume of com-plete Freund’s adjuvant. Tri-immunol vaccine (0.5 ml;Lederle, Pearl River, NY) was injected intramuscularly atthe first immunization. Three rabbits were immunizedwith 2 ml of the emulsion containing approximately 500mg of conjugate in 15–20 sites at 6–8 week intervals. Theanimals were boosted with approximately 200 mg ofconjugate peptide in incomplete Freund’s adjuvant. Ani-mals were bled from the central ear artery 6–8 days aftereach booster injection.
Antiserum screening by ELISA
The peptide used for immunizations (500 mg/well) wascoated onto 96 well plates by incubation with 50 ml of 10mg/ml peptide solution in phosphate-buffered saline (PBS)overnight at 4°C. Plates were treated with 1% bovineserum albumin in PBS containing 0.01% Tween 20 for onehour at room temperature, washed and incubated with 100ml of diluted antisera (1:100–1:1,000,000). Following onehour incubation at 37°C, plates were washed, incubatedwith goat anti-rabbit IgG conjugated with horseradish-peroxidase (100 ml of 1:1,000 dilution) for one hour at roomtemperature, washed, and then incubated with 100 ml ofo-phenylenediamine dihydrochloride in 0.1 M citrate buffer,pH 5.0 containing 0.015% H2O2 for 10 minutes at 37°C.Reactions were stopped by the addition of 50 ml of 10%H2SO4. Absorbance was measured at 492 nm.
Affinity purification of antisera
AH Sepharose beads (Pharmacia Fine Chemicals, Bel-mont, CA) were prepared according to the manufacturer’srecommendation. Five mg of receptor peptide, 500 ml of 1M ECDI (N-ethyl-N’ 3-dimethylaminopropyl) carbodiimideHCl (K & K Laboratory, Cleveland, OH), and 10 ml of100,000 cpm 125I-labeled peptide (which was used as aconjugation efficiency marker) were added to the beadsuspension. Following an overnight incubation at roomtemperature on a shaker, beads were washed with 0.1 Macetic acid in 1 M NaCl, followed by 0.14 M NaHCO3 in 1 MNaCl, then resuspended in 5 ml of 0.05 M phosphate-buffered saline (PBS). The peptide-conjugation efficiencywas approximately 60%. IgGs were initially prepared fromthe antisera by ammonium sulfate precipitation, and 2 mlof dialyzed IgG were incubated with the peptide-conju-gated bead suspension overnight at room temperature.Beads were washed three times with PBS at the end of thisincubation. Affinity-purified IgG was subsequently elutedwith 0.1 M citrate buffer, pH 3.2. Fractions were tested forimmunoreactivity by ELISA. Immunoreactive fractionswere pooled, dialyzed in 0.05 M PBS, and stored at -70°Cuntil use (Walsh and Wong, 1987).
SST2A RECEPTOR EXPRESSION IN THE GUT 397

Development of stable cell lines expressingrat sst2A or sst2B
Clones for sst2A and sst 2B receptors were isolated fromSprague-Dawley rat genomic DNA (Clontech, Palo Alto,CA) and gastric ECL cDNA libraries (provided by Dr.George Sachs, VA West Los Angeles Medical Center, LosAngeles, CA), respectively. After confirmation with theoriginal published DNA sequences (Kluxen et al., 1992;Vanetti et al., 1992), the open reading frame of subtype 2A(1.3 kb) or subtype 2B (1.2 kb) was subcloned into amammalian expression vector pCEP4 (Invitrogen, SanDiego, CA). Two receptor subtype constructs were thentransfected into human embryonic kidney 293 cell lines(HEK-293, ATCC CRL 1573) by lipofectamine (Life Tech-nologies, Gaithersburg, MD). Cells expressing individualreceptor subtypes were selected based on their resistanceto hygromycin (500 unit/ml) and on their binding to125I-SOM-14 (Amersham, 2000 Ci/mmol). Single-cell clonesused in the present study express similar numbers ofreceptors (1–2 x 105 sites/cell) and comparable Kd values(SOM-14: 260.5 nM) for both receptor subtypes (unpub-lished observations). HEK-293 cells transfected with sst2Aor sst2B cDNA were used to assess the specificity of thesst2A antibody used in the present study.
Western blotting
Transfected cells were lysed in Laemmli buffer. Aliquots(20 mg total protein/lane) were separated by SDS-PAGEon a 4–15% polyacrylamide gel (Bio-Rad, Hercules, CA)and transferred to nitrocellulose. Filters were first incu-bated with blocking solution (3% nonfat dry milk in PBS,0.1% Tween 20) for one hour at room temperature. Theywere then incubated with the receptor antiserum at a finaldilution of 1:3,000 overnight at room temperature, washed,and incubated with goat anti-rabbit IgG conjugated toperoxidase at 1:7,500 dilution for one hour at room tempera-ture. Immunoreactive bands were detected by using anAmersham ECL detection kit (Amersham, ArlingtonHeight, Ill) according to the manufacturer’s protocol. As acontrol, the receptor antiserum was preadsorbed with 1mg of C-terminus peptide, sst2A(361-369), per ml of antibodysolution for one hour at 37°C, and used in place of thereceptor antibody. For receptor membrane deglycosyla-tion, PNGase F and endoglycosidase H (BioLabs, Beverly,CA) were used according to the manufacturer’s recommen-dations. An equal volume of radioimmune precipitation(RIPA) buffer (150 mM NaCl, 50 mM Tris-HCl, pH 8.5, 5mM EDTA, 1% Nonidet P40, 0.1% sodium deoxycholate,0.1% SDS) was added to 200 mg/100 ml of protein fromeach cell membrane preparation and incubated overnightat 4°C with 20 ml of PNGase F (500,000 U/ml) or 40 ml ofendoglycosidase H (500,000 U/ml). The extent of deglyco-sylation was then evaluated by Western blotting analysis.
Tissue preparation
Care and handling of the animals were approved by theAnimal Research Committees of the Veterans Administra-tion Medical Center, West Los Angeles, in accordance withall NIH guidelines. Adult Sprague-Dawley rats of bothsexes were deeply anesthetized with 30% chloral hydratei.p. For whole-mount preparations, animals were eutha-nized by cardiac incision. Specimens of the ileum (distal)and colon (proximal) were dissected, thoroughly washedwith saline, opened along the mesenteric region, slightly
stretched and pinned onto a wax plate (mucosa down), andimmersion-fixed in 4% paraformaldehyde in 0.1 M phos-phate buffer, pH 7.4 (PB) or Zamboni’s fixative (2% parafor-maldehyde, 15% saturated picric acid solution in PB) at4°C for 4–12 hours. The following day, tissue was eitherplaced in 25% sucrose in PB for 12–24 hours for cryoprotec-tion and sectioned flat with a sliding microtome from theserosa surface through the mucosa at 25–30 µm thickness(Sternini et al., 1995), or was prepared for layer separation(Sternini et al., 1987). In this latter case, tissue wasdehydrated in graded alcohols, cleared in xylene, rehy-drated in alcohol, and washed in PB. After scraping off themucosa, the submucosa was separated from the musclelayer.
For preparation of cryostat sections, deeply anesthe-tized rats were transcardially perfused with 50 ml of PB,followed by 500 ml of 4% paraformaldehyde in 0.1M PB orZamboni’s fixative at room temperature. The stomach,proximal duodenum, distal ileum, and proximal colon wereremoved and rinsed in PBS. Tissue was postfixed for onehour at room temperature, thoroughly washed in PBS, andplaced in 25% sucrose in PBS overnight at 4°C. Transversesections of 12 µm were cut perpendicularly to the lumenwith a cryostat and collected onto gelatin-coated slides(Sternini et al., 1987; Sternini and Anderson, 1992).
Immunohistochemistry
Transfected HEK-293 cells, grown on coverslips, wererinsed in 0.1 M PB, fixed in 4% paraformaldehyde for 15minutes, washed in PB, and then incubated in primaryantibodies diluted in 0.3% Triton X-100/PB containing 10%normal goat serum, overnight at 4°C. For this study, anaffinity-purified antibody raised to a synthetic peptide(YNSKTGEEDTMAWV) corresponding to a specific regionof the C-terminal portion of rat sst2B receptor (sst2B(330–343),antibody #9578), was also used. SST2B receptor antibodyspecificity was determined by ELISA, Western blotting,and immunohistochemistry (unpublished observations).Affinity-purified antibodies were used at 1:50–1:500 dilu-tions. Cells were then washed and incubated with goatanti-rabbit IgG conjugated with isothiocyanate fluorescein(FITC, American Qualex, La Mirada, CA, or JacksonImmuno Labs, West Grove, PA) at 1:50 dilution in 0.3%Triton X-100/PB with 10% normal goat serum for twohours at room temperature, washed, and mounted onslides. Cells were examined using a Zeiss Laser ScanningMicroscope 410 with a krypton/argon laser (Carl Zeiss,Inc., Thornwood, NY), and attached to a Zeiss Axiovert 100microscope with PlanApo 100 x 1.4 na objective. Imageswere collected at a magnification zoom of 1.5–3.5x. Typi-cally, 10–20 optical sections were taken with a z-axis of 0.5µm. Images were processed and labeled using AdobePhotoshop 3.0.5 (Adobe Systems, Mountain View, CA).
Tissue sections and whole-mounts were processed by theimmunofluorescence or the avidin-biotin complex (ABC)technique for single-labeling, or the immunofluorescencemethod for double-labeling, as previously described (Ster-nini et al., 1987; Goehler et al., 1988; Sternini and Ander-son, 1992). For these studies, affinity-purified (1:50-1:100dilutions) and nonaffinity-purified (1:500-1:1,000 dilu-tions) sst2A receptor antibodies were used. Whole-mountsand microtome sections were processed as free-floatingspecimens, and cryostat sections were processed on theslides. For the immunofluorescence method, tissues werewashed in PB, incubated in primary antibodies for 18–48
398 C. STERNINI ET AL.

hours at 4°C, followed by a two-hour incubation in goatanti-rabbit IgG conjugated with FITC at 1:50 dilution,washed and, if free-floating, were mounted and thencoverslipped. For the ABC method, tissues were washed inPB, pretreated in a 0.01%–0.1% H2O2 solution for 15minutes, and incubated in primary antibodies for 18–48hours at 4°C; this was followed by a two-hour incubation ina 1:100 biotinylated goat anti-rabbit IgG (Vector Laborato-ries, Burlingame, CA) and a two-hour incubation in ABCsolution (Vectastain Elite ABC kit, Vector Laboratories).The immunoreaction was visualized by using 3’,3’-diamino-benzidine with 0.01% H2O2. Tissues were dehydrated withgraded alcohol, cleared in xylene, and coverslipped withKrystalon or Permount. Whole-mounts and microtomesections were mounted on gelatin-coated slides, air-dried,exposed to 0.5% osmium tetraoxide for 10 seconds, dehy-drated, and cleared in xylene before coverslipping. Fordouble-label immunofluorescence, sections were incubatedin a mixture of rabbit sst2A receptor antiserum and mousemonoclonal SOM (SOM6, 1:100 dilution) or VIP (VIP55,1:1,500 dilution) antibody overnight at 4°C, followed by atwo-hour incubation at room temperature with a mixtureof goat anti-rabbit IgG coupled with FITC (1:50, JacksonImmuno Labs) and goat anti-mouse IgG conjugated withtetramethyl rhodamine (TRITC; 1:100; Jackson ImmunoLabs); the sections were then coverslipped with a mixtureof glycerol/PB. Anti-rabbit and anti-mouse IgG were affin-ity-purified and absorbed against mouse and rabbit serum,respectively, to minimize cross-reaction of the secondaryantibodies with the inappropriate antigen-antibody com-plex. Primary and secondary antibodies were diluted in0.5% Triton X-100 in 0.1 M PB containing 10% normal goatserum. Specificity controls included preadsorption of thesst2A receptor, SOM, and VIP antibodies with the appropri-ate peptides (10 µM) for 16–24 hours at 4°C. SOM and VIPantibodies have been previously validated for immunohis-tochemical use in tissue (Wong et al., 1990, 1996). Fordouble-labeling experiments, controls were included toassess that the primary antisera did not cross-react whenmixed and that the secondary antibodies recognized theappropriate antigen-antibody complexes, as previouslydescribed (Goehler et al., 1988). Experimental and controltissues were taken from the same animal and processedsimultaneously.
RESULTS
Western blotting
The sst2A receptor antiserum #9431 recognized a broadprotein band with a molecular weight of approximately 80kDa in membranes prepared from HEK-293 cells trans-fected with sst2A receptor cDNA (Fig. 1A). This antiserumdetected an additional band at approximately 40 kDamolecular weight when membranes were treated withPNGase F, which cleaves between the innermost GlcNAcand asparagine residues of high mannose and complexoligosaccharides from N-linked glycoproteins (Fig. 1B), butnot with endoglycosidase H (not shown), which cleaves thechitobiose core of high mannose and some hybrid oligosac-charides from glycoproteins. These bands were not de-tected when the antibody was preadsorbed with the syn-thetic peptide (Fig. 1C,D). Similarly, the sst2B receptorantibody #9578 detected a broad band at approximately 80kDa molecular weight using HEK-293 cells transfectedwith sst2B receptor cDNA, and an additional band of
approximately 40 kDa molecular weight following deglyco-sylation with PNGase F, but not with endoglycosidase H(unpublished observations).
Immunocytochemical localization of sst2A
receptor in HEK-293 transfected cells
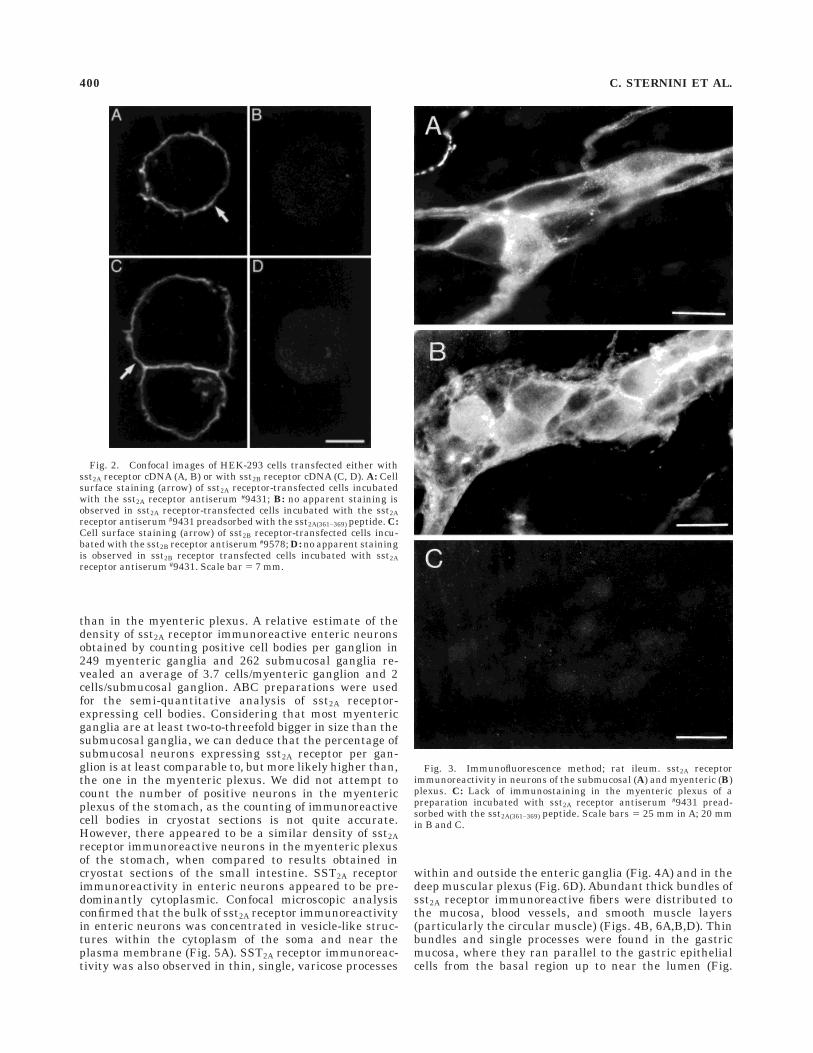
The anti-sst2A receptor antiserum #9431 stained theplasma membrane of HEK-293 cells transfected with sst2Areceptor cDNA (Fig. 2A), but not HEK-293 cells trans-fected with sst2B receptor cDNA (Fig. 2D). SST2A receptorcell surface staining was abolished by preincubation of the9431 antiserum with the sst2A(361-369) peptide (Fig. 2B), butnot with the sst2B(330-343) peptide (not shown). HEK-293cells transfected with sst2B receptor cDNA were stainedwith the sst2B receptor antiserum 9578 (Fig. 2C); stainingwas confined to the plasma membrane and was abolishedby preincubation of 9578 antiserum with the sst2B(330-343)peptide, but not by preadsorption with the sst2A(361-369) (notshown).
Cellular localization of sst2A
receptor immunoreactivity in the ratgastrointestinal tract
The sst2A receptor 9431 antiserum stained cell bodiesand fibers in the myenteric and submucosal plexuses of theintestine (Figs. 3A,B, 4A,B) as well as in the myentericplexus of the stomach (not shown). Immunoreactivity wasnot present in tissue incubated with the antiserum pread-sorbed with its homologous peptide (Fig. 3C). In general,sst2A receptor immunoreactive staining in enteric neuronsappeared to be more intense in the submucosal plexus
Fig. 1. Western blot of membranes prepared from HEK-293 cellstransfected with sst2A receptor cDNA and incubated with either sst2Areceptor antiserum #9431 (A, B) or with sst2A receptor 9431 antiserumpreadsorbed with sst2A(361–369) peptide (C, D). B, D: Membranes treatedwith PNGase F. The sst2A receptor antiserum labels a broad band atapproximately 80 kDa (A, B); following glycosylation with PNGase F(B), the antiserum also labels a band at approximately 40 kDa. Notethe lack of immunoreactive bands in membranes incubated with theantiserum preadsorbed with the peptide (C, D). Molecular size mark-ers in kDa (k) are shown on the left.
SST2A RECEPTOR EXPRESSION IN THE GUT 399
than in the myenteric plexus. A relative estimate of thedensity of sst2A receptor immunoreactive enteric neuronsobtained by counting positive cell bodies per ganglion in249 myenteric ganglia and 262 submucosal ganglia re-vealed an average of 3.7 cells/myenteric ganglion and 2cells/submucosal ganglion. ABC preparations were usedfor the semi-quantitative analysis of sst2A receptor-expressing cell bodies. Considering that most myentericganglia are at least two-to-threefold bigger in size than thesubmucosal ganglia, we can deduce that the percentage ofsubmucosal neurons expressing sst2A receptor per gan-glion is at least comparable to, but more likely higher than,the one in the myenteric plexus. We did not attempt tocount the number of positive neurons in the myentericplexus of the stomach, as the counting of immunoreactivecell bodies in cryostat sections is not quite accurate.However, there appeared to be a similar density of sst2Areceptor immunoreactive neurons in the myenteric plexusof the stomach, when compared to results obtained incryostat sections of the small intestine. SST2A receptorimmunoreactivity in enteric neurons appeared to be pre-dominantly cytoplasmic. Confocal microscopic analysisconfirmed that the bulk of sst2A receptor immunoreactivityin enteric neurons was concentrated in vesicle-like struc-tures within the cytoplasm of the soma and near theplasma membrane (Fig. 5A). SST2A receptor immunoreac-tivity was also observed in thin, single, varicose processes
within and outside the enteric ganglia (Fig. 4A) and in thedeep muscular plexus (Fig. 6D). Abundant thick bundles ofsst2A receptor immunoreactive fibers were distributed tothe mucosa, blood vessels, and smooth muscle layers(particularly the circular muscle) (Figs. 4B, 6A,B,D). Thinbundles and single processes were found in the gastricmucosa, where they ran parallel to the gastric epithelialcells from the basal region up to near the lumen (Fig.
Fig. 2. Confocal images of HEK-293 cells transfected either withsst2A receptor cDNA (A, B) or with sst2B receptor cDNA (C, D). A: Cellsurface staining (arrow) of sst2A receptor-transfected cells incubatedwith the sst2A receptor antiserum #9431; B: no apparent staining isobserved in sst2A receptor-transfected cells incubated with the sst2Areceptor antiserum #9431 preadsorbed with the sst2A(361–369) peptide. C:Cell surface staining (arrow) of sst2B receptor-transfected cells incu-bated with the sst2B receptor antiserum #9578; D: no apparent stainingis observed in sst2B receptor transfected cells incubated with sst2Areceptor antiserum #9431. Scale bar 5 7 mm.
Fig. 3. Immunofluorescence method; rat ileum. sst2A receptorimmunoreactivity in neurons of the submucosal (A) and myenteric (B)plexus. C: Lack of immunostaining in the myenteric plexus of apreparation incubated with sst2A receptor antiserum #9431 pread-sorbed with the sst2A(361–369) peptide. Scale bars 5 25 mm in A; 20 mmin B and C.
400 C. STERNINI ET AL.
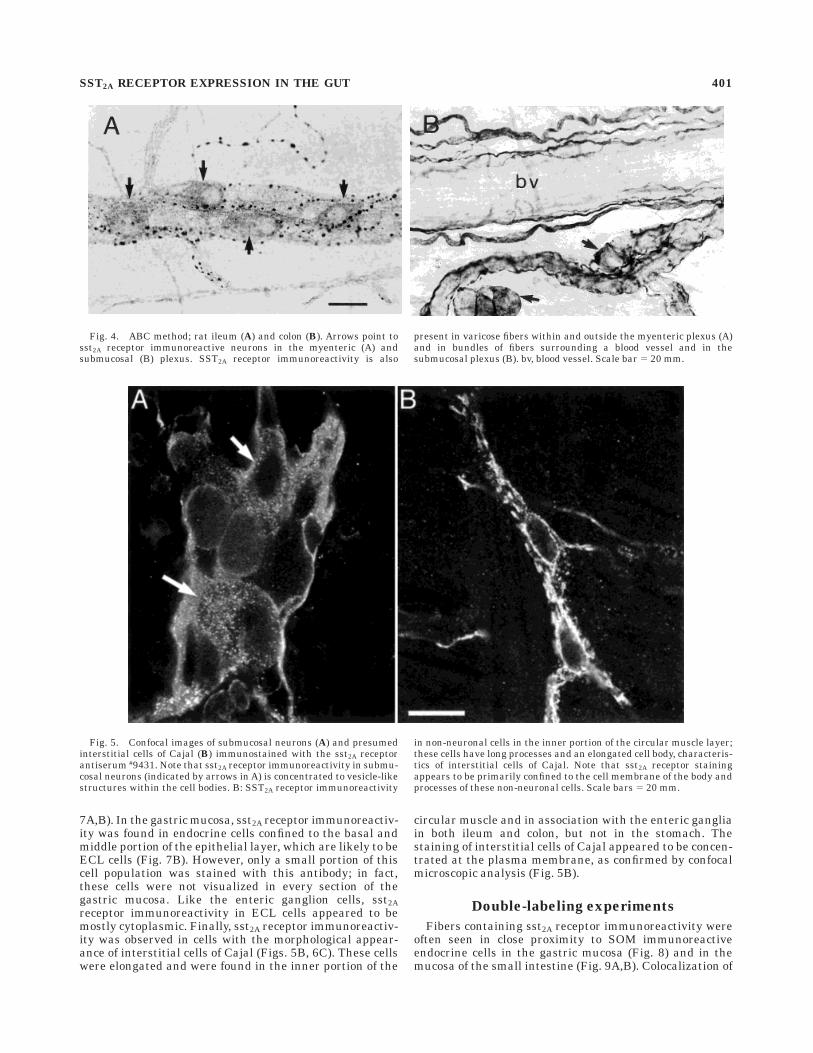
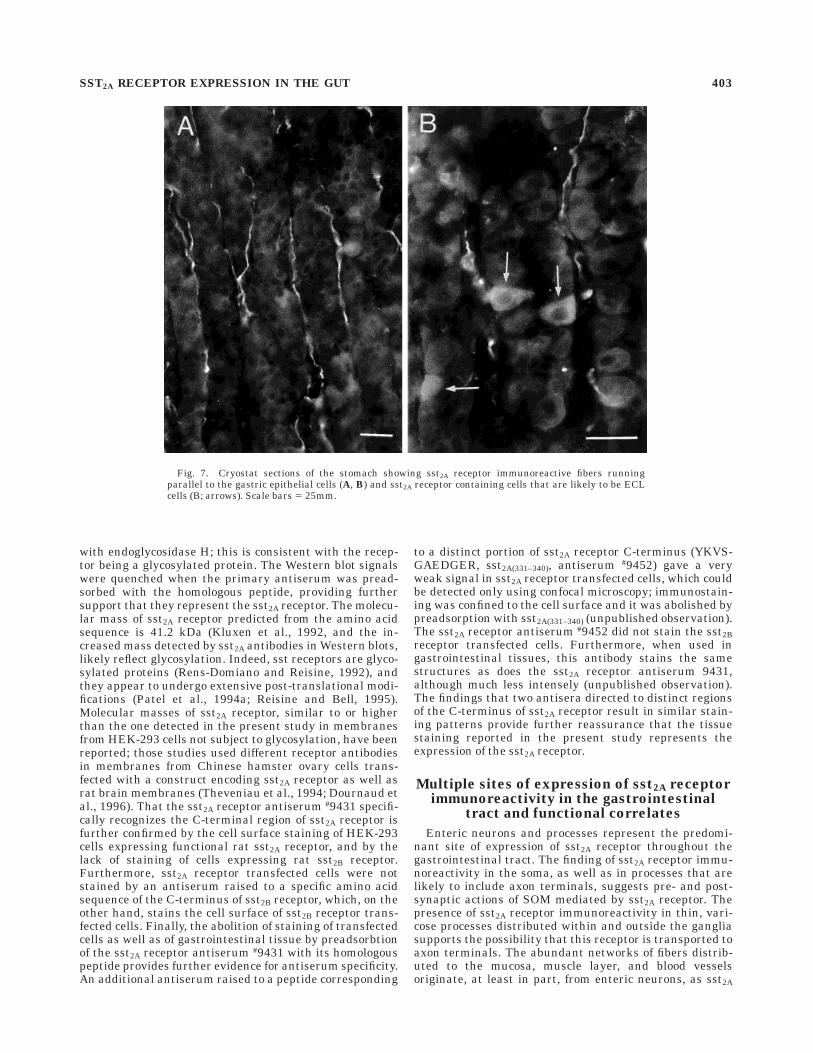
7A,B). In the gastric mucosa, sst2A receptor immunoreactiv-ity was found in endocrine cells confined to the basal andmiddle portion of the epithelial layer, which are likely to beECL cells (Fig. 7B). However, only a small portion of thiscell population was stained with this antibody; in fact,these cells were not visualized in every section of thegastric mucosa. Like the enteric ganglion cells, sst2Areceptor immunoreactivity in ECL cells appeared to bemostly cytoplasmic. Finally, sst2A receptor immunoreactiv-ity was observed in cells with the morphological appear-ance of interstitial cells of Cajal (Figs. 5B, 6C). These cellswere elongated and were found in the inner portion of the
circular muscle and in association with the enteric gangliain both ileum and colon, but not in the stomach. Thestaining of interstitial cells of Cajal appeared to be concen-trated at the plasma membrane, as confirmed by confocalmicroscopic analysis (Fig. 5B).
Double-labeling experiments
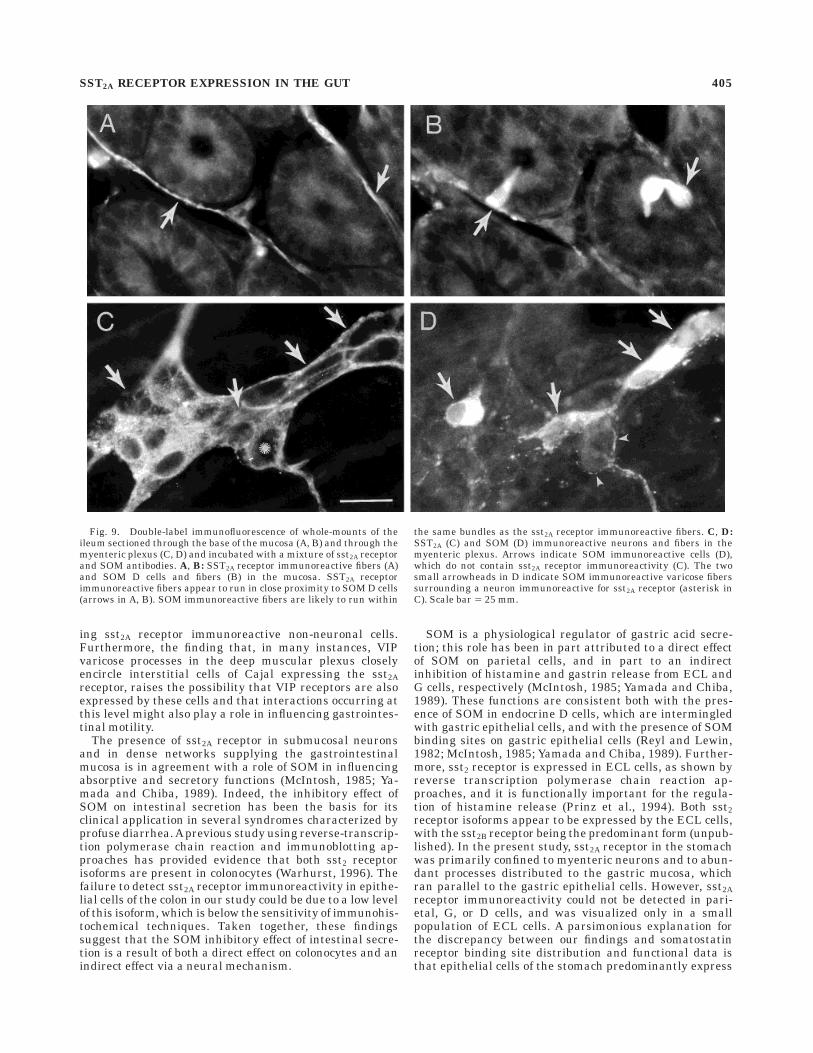
Fibers containing sst2A receptor immunoreactivity wereoften seen in close proximity to SOM immunoreactiveendocrine cells in the gastric mucosa (Fig. 8) and in themucosa of the small intestine (Fig. 9A,B). Colocalization of
Fig. 4. ABC method; rat ileum (A) and colon (B). Arrows point tosst2A receptor immunoreactive neurons in the myenteric (A) andsubmucosal (B) plexus. SST2A receptor immunoreactivity is also
present in varicose fibers within and outside the myenteric plexus (A)and in bundles of fibers surrounding a blood vessel and in thesubmucosal plexus (B). bv, blood vessel. Scale bar 5 20 mm.
Fig. 5. Confocal images of submucosal neurons (A) and presumedinterstitial cells of Cajal (B) immunostained with the sst2A receptorantiserum #9431. Note that sst2A receptor immunoreactivity in submu-cosal neurons (indicated by arrows in A) is concentrated to vesicle-likestructures within the cell bodies. B: SST2A receptor immunoreactivity
in non-neuronal cells in the inner portion of the circular muscle layer;these cells have long processes and an elongated cell body, characteris-tics of interstitial cells of Cajal. Note that sst2A receptor stainingappears to be primarily confined to the cell membrane of the body andprocesses of these non-neuronal cells. Scale bars 5 20 mm.
SST2A RECEPTOR EXPRESSION IN THE GUT 401

sst2A receptor and SOM immunoreactivities was not ob-served in enteric ganglion cells (Fig. 9C,D). Finally, SOM-containing varicose fibers often closely encircled sst2Areceptor immunoreactive enteric ganglion cells (Fig. 9C,D).
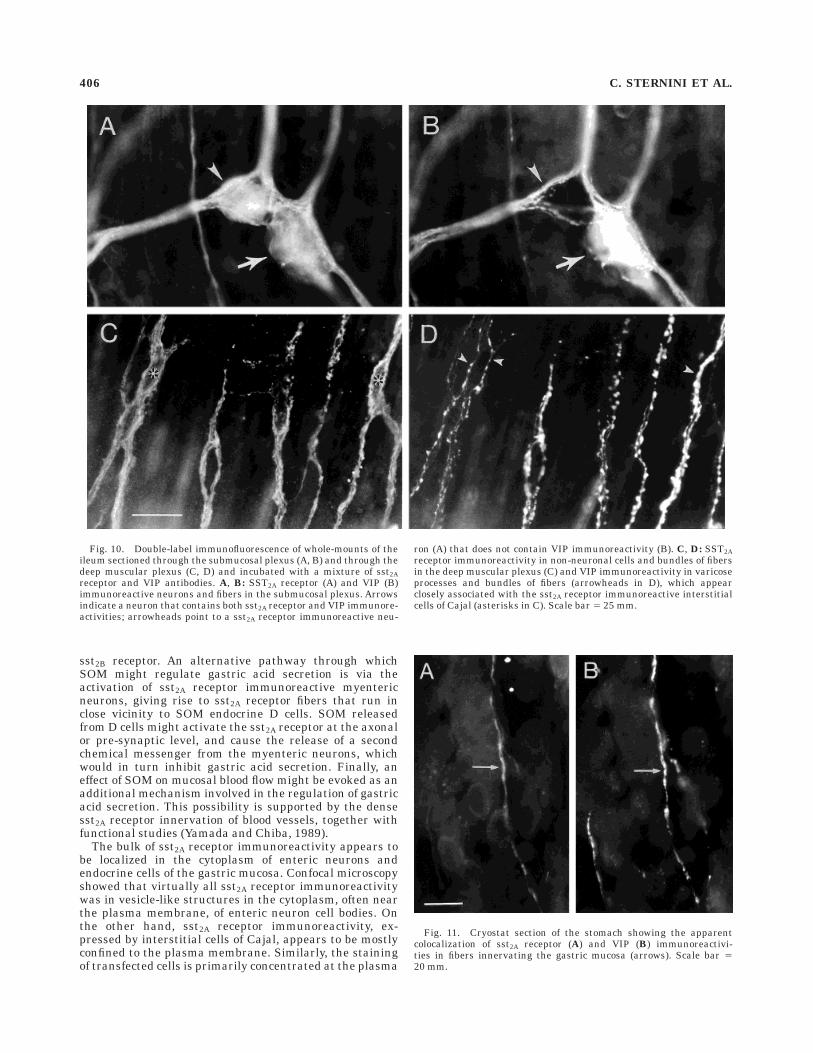
SST2A receptor and VIP immunoreactivities often colocal-ized in enteric neurons, particularly in the submucosalplexus (Fig. 10A,B). Approximately 25% of VIP immunore-active myenteric neurons and approximately 40% of VIPimmunoreactive submucosal neurons were immunostainedwith the sst2A receptor antiserum. SST2A receptor and VIPimmunoreactivities also appeared to colocalize in varicosefibers and in thin bundles of fibers in the deep muscularplexus of the intestine (Fig. 10C,D) and in the gastricmucosa (Fig. 11A,B). Finally, varicose VIP immunoreac-tive processes were observed in close proximity to andsometimes encircling sst2A receptor immunoreactive inter-stitial cells of Cajal in the inner portion of the circular musclelayer (Fig. 10C,D).
Specificity of sst2A receptor immunostainingin tissue specimens
Preadsorption of the sst2A receptor antibody with thesynthetic peptide abolished the staining in tissue speci-mens (Fig. 3C). Preadsorption of the sst2A receptor anti-body with SOM or VIP peptide did not affect the sst2A
receptor immunostaining. Similarly, an excess of SOM orVIP peptide blocked the SOM or VIP staining, respectively,whereas an excess of sst2A receptor peptide fragment didnot modify the SOM or VIP immunostaining. Controls fordouble-labeling confirmed that the sst2A receptor antise-rum and the SOM or VIP antibody do not cross-react whenmixed, and that the secondary antisera bind only to theappropriate antigen-antibody complexes.
DISCUSSION
This study demonstrates that sst2A receptor immunore-activity is quite abundant in the gastrointestinal tract.The sst2A receptor is expressed by neuronal and non-neuronal structures, which are often in close proximity toSOM-containing elements; this indicates that SOM re-leased by neurons or endocrine D cells can activate thesst2A receptor over close distance, thus influencing avariety of gastrointestinal functions via local circuits.
Specificity of the antiserum
Several lines of evidence indicate specificity of the 9431antiserum. First, it detects a broad band of membranes, ofapproximately 80 kDa in Western blot, of HEK-293 cellstransfected with sst2A receptor, as well as an additional 40kDa band, following treatment with PNGase F but not
Fig. 6. SST2A receptor immunoreactivity in whole-mounts of theileum (A, C, D) and a cryostat section of the upper duodenum (B).Networks of sst2A receptor immunoreactive fibers surround the base ofthe intestinal glands (A). B: Thick bundles of sst2A receptor immunore-active fibers associated with blood vessels (bv) of the submucosa andbase of mucosa. C: SST2A receptor immunoreactivity in elongated
non-neuronal cells (arrows) with long processes in the inner portion ofthe circular muscle; these are likely to be interstitial cells of Cajal. D:Networks of fibers and thin, varicose (arrowheads) processes immuno-reactive for sst2A receptor in the deep muscular plexus. Calibrationbars 5 25 mm in A,B,D; 20 mm in C.
402 C. STERNINI ET AL.
with endoglycosidase H; this is consistent with the recep-tor being a glycosylated protein. The Western blot signalswere quenched when the primary antiserum was pread-sorbed with the homologous peptide, providing furthersupport that they represent the sst2A receptor. The molecu-lar mass of sst2A receptor predicted from the amino acidsequence is 41.2 kDa (Kluxen et al., 1992, and the in-creased mass detected by sst2A antibodies in Western blots,likely reflect glycosylation. Indeed, sst receptors are glyco-sylated proteins (Rens-Domiano and Reisine, 1992), andthey appear to undergo extensive post-translational modi-fications (Patel et al., 1994a; Reisine and Bell, 1995).Molecular masses of sst2A receptor, similar to or higherthan the one detected in the present study in membranesfrom HEK-293 cells not subject to glycosylation, have beenreported; those studies used different receptor antibodiesin membranes from Chinese hamster ovary cells trans-fected with a construct encoding sst2A receptor as well asrat brain membranes (Theveniau et al., 1994; Dournaud etal., 1996). That the sst2A receptor antiserum #9431 specifi-cally recognizes the C-terminal region of sst2A receptor isfurther confirmed by the cell surface staining of HEK-293cells expressing functional rat sst2A receptor, and by thelack of staining of cells expressing rat sst2B receptor.Furthermore, sst2A receptor transfected cells were notstained by an antiserum raised to a specific amino acidsequence of the C-terminus of sst2B receptor, which, on theother hand, stains the cell surface of sst2B receptor trans-fected cells. Finally, the abolition of staining of transfectedcells as well as of gastrointestinal tissue by preadsorbtionof the sst2A receptor antiserum #9431 with its homologouspeptide provides further evidence for antiserum specificity.An additional antiserum raised to a peptide corresponding
to a distinct portion of sst2A receptor C-terminus (YKVS-GAEDGER, sst2A(331–340), antiserum #9452) gave a veryweak signal in sst2A receptor transfected cells, which couldbe detected only using confocal microscopy; immunostain-ing was confined to the cell surface and it was abolished bypreadsorption with sst2A(331–340) (unpublished observation).The sst2A receptor antiserum #9452 did not stain the sst2Breceptor transfected cells. Furthermore, when used ingastrointestinal tissues, this antibody stains the samestructures as does the sst2A receptor antiserum 9431,although much less intensely (unpublished observation).The findings that two antisera directed to distinct regionsof the C-terminus of sst2A receptor result in similar stain-ing patterns provide further reassurance that the tissuestaining reported in the present study represents theexpression of the sst2A receptor.
Multiple sites of expression of sst2A receptorimmunoreactivity in the gastrointestinal
tract and functional correlates
Enteric neurons and processes represent the predomi-nant site of expression of sst2A receptor throughout thegastrointestinal tract. The finding of sst2A receptor immu-noreactivity in the soma, as well as in processes that arelikely to include axon terminals, suggests pre- and post-synaptic actions of SOM mediated by sst2A receptor. Thepresence of sst2A receptor immunoreactivity in thin, vari-cose processes distributed within and outside the gangliasupports the possibility that this receptor is transported toaxon terminals. The abundant networks of fibers distrib-uted to the mucosa, muscle layer, and blood vesselsoriginate, at least in part, from enteric neurons, as sst2A
Fig. 7. Cryostat sections of the stomach showing sst2A receptor immunoreactive fibers runningparallel to the gastric epithelial cells (A, B) and sst2A receptor containing cells that are likely to be ECLcells (B; arrows). Scale bars 5 25mm.
SST2A RECEPTOR EXPRESSION IN THE GUT 403

receptor immunoreactivity is present in both myentericand submucous neurons. However, from this study wecannot exclude that a component of these fibers originatesfrom extrinsic sources. Indeed, sst2A receptor immunoreac-tivity has been reported in the dorsal motor nucleus of thevagus (Dournaud et al., 1996), and it is expressed inneurons of the dorsal root ganglia (unpublished observa-tions); this raises the possibility that efferent and afferentcomponents of the innervation of the gastrointestinal tractexpress sst2A receptor. The distribution of sst2A receptorimmunoreactive processes is consistent with the sst2Areceptor being expressed by functionally distinct entericneurons. Indeed, sst2A receptor immunoreactive fibers aredistributed to all the layers of the gastrointestinal tractfrom the mucosa to the muscularis externa and to thevasculature. The enteric ganglia, which form the entericnervous system, provide the bulk of the innervation of thegastrointestinal tract and are responsible for maintainingits functions (Furness and Costa, 1987). Myenteric gangliaare primarily involved in the control of the peristalticreflex (Furness and Costa, 1987), and submucosal gangliaare mostly concerned with mucosa function and blood flowregulation (Bornstein and Furness, 1988; Song et al.,
1992). Enteric neurons comprise sensory neurons, interneu-rons, inhibitory and excitatory motor and secretomotorneurons (Furness and Costa, 1987; Furness et al., 1992;Gershon et al., 1994), which can be distinguished on thebasis of their morphological appearance, projections, andneurochemistry. The presence of sst2A receptor immunore-activity in a large proportion of neurons containing immu-noreactivity for VIP provides evidence that sst2A receptorenteric neurons comprise neurons of the descending inhibi-tory pathway, which is responsible for generating smoothmuscle relaxation and includes motor neurons and inter-neurons, as well as submucosal secretomotor neurons(Furness and Costa, 1987). In fact, VIP is an establishedimmunocytochemical marker for these types of entericneurons (Furness et al., 1992).
SOM is involved in the regulation of peristalsis andinterdigestive motility, as indicated by the presence ofSOM immunoreactive neurons in the myenteric plexusand by functional studies (McIntosh, 1985; Yamada andChiba, 1989), even though the mechanism of this action isstill largely unknown. Its overall effect on gastrointestinaltransit appears to be inhibitory (Johansson et al., 1978;Teitelbaum et al., 1984), and it is likely to involve therelease of another substance. The expression of sst2Areceptor by VIP-containing myenteric neurons is consis-tent with the hypothesis that SOM causes muscle relax-ation by activating inhibitory neurons (Furness and Costa,1979). The relaxation effect of SOM in isolated smoothmuscle preparation has also been attributed to a loss ofcholinergic tone, due to the inhibition of acetylcholinerelease caused by SOM (Guillemin, 1976; Furness andCosta, 1979; Yau et al., 1983). This would suggest thatsst2A receptor is expressed by cholinergic, excitatory neu-rons. The non-VIP immunoreactive sst2A receptor neuronsmight comprise cholinergic and perhaps non-cholinergicexcitatory ganglion cells. We can hypothesize that SOMaction on gastrointestinal transit results from the activa-tion of distinct populations of neurons, including neuronsof both the inhibitory descending pathway (which use VIPand nitric oxide as transmitters) and the stimulatoryascending pathway (cholinergic and noncholinergic, thelatter using tachykinins as transmitters) (Furness et al.,1992). A neuronal mechanism for SOM effect on gastroin-testinal transit is supported by the close vicinity of neu-rons containing SOM or sst2A receptor immunoreactivity,and by the observation that varicose SOM immunoreactivefibers often encircle sst2A receptor immunoreactive neu-rons in the enteric ganglia. An additional pathway throughwhich SOM could influence gastrointestinal motility isrepresented by the non-neuronal cells expressing sst2Areceptor that have the morphological appearance of inter-stitial cells of Cajal. These cells are found in the innerportion of the circular muscle of the intestine where thedeep muscular plexus is located, and they are intercon-nected to neighboring smooth muscle cells of the circularmuscle layer as well as in proximity to the enteric ganglia(Gabella, 1974; Thuneberg, 1982). The interstitial cells ofCajal have been regarded as an important site of regula-tion of smooth muscle contraction, and have been impli-cated in signaling transmission from nerves to muscle andas mediators of neurotransmission (Daniel and Berezin,1992; Sanders, 1996). The presence of sst2A in these specialnon-neuronal cell types, which are often closely associatedwith SOM-containing fibers, supports the possibility thatSOM influences smooth muscle activity by directly activat-
Fig. 8. Double-label immunofluorescence of a cryostat section ofthe rat stomach. Simultaneous visualization of sst2A receptor immuno-reactive fibers (in green) and SOM immunoreactive D cells (in red)visualized with FITC/TRITC dual band filter set. Note the closeapposition of a sst2A receptor fiber to a SOM D cell. Scale bar 5 40 mm.
404 C. STERNINI ET AL.
ing sst2A receptor immunoreactive non-neuronal cells.Furthermore, the finding that, in many instances, VIPvaricose processes in the deep muscular plexus closelyencircle interstitial cells of Cajal expressing the sst2Areceptor, raises the possibility that VIP receptors are alsoexpressed by these cells and that interactions occurring atthis level might also play a role in influencing gastrointes-tinal motility.
The presence of sst2A receptor in submucosal neuronsand in dense networks supplying the gastrointestinalmucosa is in agreement with a role of SOM in influencingabsorptive and secretory functions (McIntosh, 1985; Ya-mada and Chiba, 1989). Indeed, the inhibitory effect ofSOM on intestinal secretion has been the basis for itsclinical application in several syndromes characterized byprofuse diarrhea.Aprevious study using reverse-transcrip-tion polymerase chain reaction and immunoblotting ap-proaches has provided evidence that both sst2 receptorisoforms are present in colonocytes (Warhurst, 1996). Thefailure to detect sst2A receptor immunoreactivity in epithe-lial cells of the colon in our study could be due to a low levelof this isoform, which is below the sensitivity of immunohis-tochemical techniques. Taken together, these findingssuggest that the SOM inhibitory effect of intestinal secre-tion is a result of both a direct effect on colonocytes and anindirect effect via a neural mechanism.
SOM is a physiological regulator of gastric acid secre-tion; this role has been in part attributed to a direct effectof SOM on parietal cells, and in part to an indirectinhibition of histamine and gastrin release from ECL andG cells, respectively (McIntosh, 1985; Yamada and Chiba,1989). These functions are consistent both with the pres-ence of SOM in endocrine D cells, which are intermingledwith gastric epithelial cells, and with the presence of SOMbinding sites on gastric epithelial cells (Reyl and Lewin,1982; McIntosh, 1985; Yamada and Chiba, 1989). Further-more, sst2 receptor is expressed in ECL cells, as shown byreverse transcription polymerase chain reaction ap-proaches, and it is functionally important for the regula-tion of histamine release (Prinz et al., 1994). Both sst2receptor isoforms appear to be expressed by the ECL cells,with the sst2B receptor being the predominant form (unpub-lished). In the present study, sst2A receptor in the stomachwas primarily confined to myenteric neurons and to abun-dant processes distributed to the gastric mucosa, whichran parallel to the gastric epithelial cells. However, sst2Areceptor immunoreactivity could not be detected in pari-etal, G, or D cells, and was visualized only in a smallpopulation of ECL cells. A parsimonious explanation forthe discrepancy between our findings and somatostatinreceptor binding site distribution and functional data isthat epithelial cells of the stomach predominantly express
Fig. 9. Double-label immunofluorescence of whole-mounts of theileum sectioned through the base of the mucosa (A, B) and through themyenteric plexus (C, D) and incubated with a mixture of sst2A receptorand SOM antibodies. A, B: SST2A receptor immunoreactive fibers (A)and SOM D cells and fibers (B) in the mucosa. SST2A receptorimmunoreactive fibers appear to run in close proximity to SOM D cells(arrows in A, B). SOM immunoreactive fibers are likely to run within
the same bundles as the sst2A receptor immunoreactive fibers. C, D:SST2A (C) and SOM (D) immunoreactive neurons and fibers in themyenteric plexus. Arrows indicate SOM immunoreactive cells (D),which do not contain sst2A receptor immunoreactivity (C). The twosmall arrowheads in D indicate SOM immunoreactive varicose fiberssurrounding a neuron immunoreactive for sst2A receptor (asterisk inC). Scale bar 5 25 mm.
SST2A RECEPTOR EXPRESSION IN THE GUT 405
sst2B receptor. An alternative pathway through whichSOM might regulate gastric acid secretion is via theactivation of sst2A receptor immunoreactive myentericneurons, giving rise to sst2A receptor fibers that run inclose vicinity to SOM endocrine D cells. SOM releasedfrom D cells might activate the sst2A receptor at the axonalor pre-synaptic level, and cause the release of a secondchemical messenger from the myenteric neurons, whichwould in turn inhibit gastric acid secretion. Finally, aneffect of SOM on mucosal blood flow might be evoked as anadditional mechanism involved in the regulation of gastricacid secretion. This possibility is supported by the densesst2A receptor innervation of blood vessels, together withfunctional studies (Yamada and Chiba, 1989).
The bulk of sst2A receptor immunoreactivity appears tobe localized in the cytoplasm of enteric neurons andendocrine cells of the gastric mucosa. Confocal microscopyshowed that virtually all sst2A receptor immunoreactivitywas in vesicle-like structures in the cytoplasm, often nearthe plasma membrane, of enteric neuron cell bodies. Onthe other hand, sst2A receptor immunoreactivity, ex-pressed by interstitial cells of Cajal, appears to be mostlyconfined to the plasma membrane. Similarly, the stainingof transfected cells is primarily concentrated at the plasma
Fig. 10. Double-label immunofluorescence of whole-mounts of theileum sectioned through the submucosal plexus (A, B) and through thedeep muscular plexus (C, D) and incubated with a mixture of sst2Areceptor and VIP antibodies. A, B: SST2A receptor (A) and VIP (B)immunoreactive neurons and fibers in the submucosal plexus. Arrowsindicate a neuron that contains both sst2A receptor and VIP immunore-activities; arrowheads point to a sst2A receptor immunoreactive neu-
ron (A) that does not contain VIP immunoreactivity (B). C, D: SST2Areceptor immunoreactivity in non-neuronal cells and bundles of fibersin the deep muscular plexus (C) and VIP immunoreactivity in varicoseprocesses and bundles of fibers (arrowheads in D), which appearclosely associated with the sst2A receptor immunoreactive interstitialcells of Cajal (asterisks in C). Scale bar 5 25 mm.
Fig. 11. Cryostat section of the stomach showing the apparentcolocalization of sst2A receptor (A) and VIP (B) immunoreactivi-ties in fibers innervating the gastric mucosa (arrows). Scale bar 520 mm.
406 C. STERNINI ET AL.

membrane. SOM receptor binding sites have been re-ported in the cytosolic compartment in addition to theplasma membrane (Lewin, 1992). Intracellular localiza-tions have also been reported for other types of G-protein-coupled receptors (Levey et al., 1991, 1993; Arvidsson etal., 1995). The cytoplasmic sst2A receptor immunoreactiv-ity may represent a soluble receptor that differs from themembrane receptor. Alternatively, sst2A receptor immuno-reactivity may be translocated from the plasma membraneto intracellular sites due to a continuous activation of thereceptor by SOM, which is in agreement with recentobservations that sst receptors internalize upon agoniststimulation (Hofland et al., 1995). Intracellular sst2A recep-tor immunoreactivity could also represent recycling recep-tor, or receptor that is moving to the plasma membranefrom the Golgi complex.
The expression of sst2A receptor by different types ofstructures within the gastrointestinal tract is consistentwith the existence of different regulatory pathways. Theactivation of sst receptors and the formation of ligand-receptor complexes are associated with distinct signalingpathways, including inhibition of adenylyl cyclase andinhibition of cellular Ca21 entry via voltage-sensitive Ca21
channels (Law et al., 1995). Although sst2 receptor selec-tive agonists cause inhibition of adenylyl cyclase activityin brain membranes (Raynor and Reisine, 1992), func-tional coupling of sst2 receptor could not be demonstratedin cell lines (Rens-Domiano and Reisine, 1992; Yamada etal., 1992; Law et al., 1993; Buscail et al., 1994). However,coupling of sst2A or sst2B receptor splice variant to adenylylcyclase has been reported (Reisine et al., 1993; Hoyer etal., 1994; Patel et al., 1994b). A better understanding of themechanisms and regulation of SOM function will beprovided by clarification of the intracellular mechanismsthrough which sst2A and sst2B mediate SOM action.
In summary, this study demonstrates that (a) the sst2Areceptor is expressed by different types of cells in thegastrointestinal tract, including functionally distinct en-teric neurons, interstitial cells of Cajal, and ECL cells; (b)sst2A immunoreactive neurons innervate a variety of tar-gets; and (c) sst2A receptor expressing structures are inclose vicinity to SOM-containing neurons and D cells.These findings support the hypothesis that SOM releasedfrom neurons and endocrine D cells in the gut exerts itseffects on target tissues via different routes, includingneuronal and non-neuronal pathways, by activating thesst2A receptor. The relationship between SOM and sst2Areceptor immunoreactive elements is consistent with aparacrine route of action. By contrast, the lack of colocaliza-tion of SOM and sst2A receptor immunoreactivities in thesame structures argues against an autocrine mechanismof action. Finally, the localization of sst2A receptor immuno-reactivity in the cytoplasm and at the cell surface raisesthe question of whether the sst2A receptor exists in differ-ent forms in distinct structures of the gastrointestinaltract, or whether the intracellular expression of sst2Areceptor observed in enteric neurons and ECL cells repre-sents internalization of the receptor upon SOM stimula-tion.
ACKNOWLEDGMENT
The authors would like to thank Katherine Wen for hertechnical assistance.
LITERATURE CITED
Arilla, E., M.P. Lopez-Ruiz, L.G. Guijarro, J.C. Prieto, A. Gomez-Pan, andB. Hirst (1984) Characterization of somatostatin binding sites incytosolic fraction of rat intestinal mucosa. Biochim. Biophys. Acta802:203–208.
Arvidsson, U., M. Riedl, S. Chakrabarti, L. Vulchanova, J.H. Lee, A.H.Nakano, X. Lin, H.H. Loh, P.Y. Law, M.W. Wessendorf, and R. Elde(1995) The kappa-opioid receptor is primarily post-synaptic: Combinedimmunohistochemical localization of the receptor and endogenousopioids. Proc. Natl. Acad. Sci. USA 92:5062–5066.
Beaudet, A., D. Greenspun, J. Raelson, and G.S. Tannenbaum (1995)Patterns of expression of SSTR1 and SSTR2 somatostatin receptorsubtypes in the hypothalamus of the adult rat: Relationship to neuroen-docrine function. Neuroscience 65:551–561.
Bell, G.I., and T. Reisine (1993) Molecular biology of somatostatin recep-tors. TINS 16:34–38.
Bornstein, J.C., and J.B. Furness (1988) Correlated electrophysiologicaland histochemical studies of submucous neurons and their contributionto understanding enteric neural circuits. J. Auton. Nerv. Syst. 25:1–13.
Breder, C.D., Y. Yamada, K. Yasuda, S. Seino, C.B. Saper, and G.I. Bell(1992) Differential expression of somatostatin receptor subtypes inbrain. J. Neurosci. 12:3920–3934.
Bruno, J.F., Y. Xu, J. Song, and M. Berelowitz (1992) Molecular cloning andfunctional expression of a brain specific somatostatin receptor. Proc.Natl. Acad. Sci. USA 89:11151–11155.
Buscail, L., N. Delesque, J-P. Esteve, N. Saint-Laurent, H. Prats, P. Clerc,D. Robberecht, G. Bell, C. Liebow, A. Schally, N. Vaysse, and C. Susinin(1994) Stimulation of tyrosine phosphatase and inhibition of cellproliferation by somatostatin analogues: Mediation by human somato-statin receptor subtypes SST1 and SST2. Proc. Natl. Acad. Sci. USA91:2315–2319.
Costa, M., Y. Patel, J.B. Furness, and A. Arimura (1977) Evidence that someintrinsic neurons of the intestine contain somatostatin. Neurosci. Lett.6:215–222.
Costa, M., J.B. Furness, I.J. Llewellyn-Smith, B. Davies, and J. Oliver(1980)An immunohistochemical study of the projections of somatostatin-containing neurons in the guinea-pig intestine. Neuroscience 5:841–852.
Daniel, E.E., and I. Berezin (1992) Interstitial cells of Cajal: Are they majorplayers in control of gastrointestinal motility? J. Gastrointest. Mot.4:1–24.
Dournaud, P., Y.Z. Gu, A. Schonbrunn, J. Mazella, G.S. Tannenbaum, andA. Beaudet (1996) Localization of the somatostatin receptor sst2A in ratbrain using a specific anti-peptide antibody. J. Neurosci. 16:4468–4478.
Ekblad, E., R. Ekman, R. Håkanson, and F. Sundler (1988) Projections ofpeptide-containing neurons in rat colon. Neuroscience 27:655–674.
Ekblad, E., C. Winther, R. Ekman, R. Håkanson, and F. Sundler (1987)Projections of peptide-containing neurons in rat small intestine. Neuro-science 20:169–188.
Epelbaum, J. (1986) Somatostatin in the central nervous system: Physiol-ogy and pathological modifications. Prog. Neurobiol. 27:63–100.
Epelbaum, J., P. Dournaud, M. Fodor, and C. Viollet (1994) The neurobiol-ogy of somatostatin. Crit. Rev. Neurobiol. 8:25–44.
Furness, J.B., and M. Costa (1979) Actions of somatostatin on excitatoryand inhibitory nerves in the intestine. Eur. J. Pharmacol. 56:69–74.
Furness, J.B., and M. Costa (1987) The Enteric Nervous System. Edin-burgh: Churchill-Livingstone.
Furness, J.B., J.C. Bornstein, R. Murphy, and S. Pompolo (1992) Roles ofpeptides in transmission in the enteric nervous system. TINS 15:66–71.
Gabella, G. (1974) Special muscle cells and their innervation in themammalian small intestine. Cell Tiss. Res. 153:63–77.
Gershon, M.D.,A.L. Kirchgessner, and P.R. Wade (1994) Functional anatomyof the enteric nervous system. In L.R. Johnson (ed): Physiology of theGastrointestinal Tract. New York: Raven Press, 3rd ed., pp. 381–422.
Goehler, E.L., C. Sternini, and N. Brecha (1988) Calcitonin gene-relatedpeptide immunoreactivity in the biliary pathway and liver of the guineapig: Distribution and colocalization with substance P. Cell Tiss. Res.253:145–150.
Guillemin, R. (1976) Somatostatin inhibits the release of acetylcholineinduced electrically in the myenteric plexus. Endocrinology 99:1653–1654.
Hofland, L.J., P.M. van Koetsveld, M. Waaijers, J. Zuyderwijk, W.A.Breeman, and S.W.J. Lamberts (1995) Internalization of the radioiodin-ated somatostatin analog [125I- Tyr3]Octreotide by mouse and human
SST2A RECEPTOR EXPRESSION IN THE GUT 407

pituitary tumor cells: Increase by unlabeled octreotide. Endocrinology136:3698–3706.
Hoyer, D., H. Lubbert, and C. Bruns (1994) Molecular pharmacology ofsomatostatin receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 350:441–453.
Hoyer, D., G.I. Bell, M. Berelowitz, J. Epelbaum, W. Feniuk, P.P.A.Humphrey, A.M. O’Carroll, Y.C. Patel, A. Schonbrunn, J.E. Taylor, andT. Reisine (1995) Classification and nomenclature of somatostatinreceptors. Trends Pharmacol. Sci. 13:61–69.
Johansson, C., S. Efendic, O. Wisen, K. Uvnas-Wallensten, and R. Luft(1978) Effects of short-term somatostatin infusion on the gastric andintestinal propulsion in humans. Scand. J. Gastroenterol. 13:481–483.
Kluxen, F.W., C. Bruns, and H. Lubbert (1992) Expression cloning of a ratbrain somatostatin cDNA. Proc. Natl. Acad. Sci. USA 89:4618–4622.
Kong, H., A.M. De Paoli, C.D. Breder, K. Yasuda, G.I. Bell, and T. Reisine(1994) Differential expression of messenger RNAs for somatostatinreceptor subtypes SSTR1, SSTR2 and SSTR3 in adult rat brain:Analysis by RNA blotting and in situ hybridization histochemistry.Neuroscience 59:175–184.
Krantic, S., R. Quirion, and G. Uhl (1992) Somatostatin receptors. In A.Bjorklund, T. Hokfelt, and M.J. Kuhar (eds): Handbook of ChemicalNeuroanatomy Neuropeptide Receptors in the CNS. Amsterdam:Elsevier Science, Vol. III, pp. 321–3446.
Larsson, L.I. (1985) Distribution and morphology of somatostatin cells. InP.Y.C. Patel and G.S. Tannenbaum (eds): Somatostatin. New York:Plenum, pp. 383–402.
Law, S.F., D. Woulfe, and T. Reisine (1995) Minireview. Somatostatinreceptor activation of cellular effector systems. Cell. Signal. 7:1–8.
Law, S., K. Yasuda, G. Bell, and T. Reisine, (1993) Gia3 and Goa selectivelyassociate with the cloned somatostatin receptor SSTR2. J. Biol. Chem.268:10721–10727.
Levey, A.I., C.A. Kitt, W.F. Simmonds, D.L. Price, and M.R. Brann (1991)Identification and localization of muscarinic acetylcholine receptorproteins in brain with subtype-specific antibodies. J. Neurosci. 11:3218–3226.
Levey, A.I., S.M. Hersch, D.B. Rye, R.G. Sunahara, H.B. Niznik, C.A. Kitt,D.L. Price, R. Maggio, M.R. Brann, and B.J. Ciliax (1993) Localizationof D1 and D2 dopamine receptors in brain with subtype-specific antibod-ies. Proc. Natl. Acad. Sci. USA 90:8861–8865.
Lewin, M.J.M. (1992) The somatostatin receptors in the GI tract. Annu.Rev. Physiol. 54:455–468.
McIntosh, C.H.S. (1985) Minireview. Gastrointestinal somatostatin: Distri-bution, secretion and physiological significance. Life Sci. 37:2043–2058.
O’Carroll, A.M., S.J. Lolait, M. Konig, and L.C. Mahan (1992) Molecularcloning and expression of a pituitary somatostatin receptor via preferen-tial affinity for somatostatin 28. Mol. Pharmacol. 42:939–946.
Patel, Y.C., and C.B. Srikant (1994) Subtype selectivity of peptide analogsfor all five cloned human somatostatin receptors (hsstR1-5). Endocrinol-ogy 135:2814–2817.
Patel, Y.C., R. Panetta, E. Escher, M. Greenwood, and C.B. Srikant (1994a)Expression of multiple somatostatin receptor genes in AtT-20 cells.Evidence for a novel somatostatin-28 selective receptor subtype. J. Biol.Chem. 269:1506–1509.
Patel, Y.C., M.T. Greenwood, A. Warszynka, R. Panetta, and C.B. Srikant(1994b) All five cloned somatostatin receptors (hsstR1-5) are function-ally coupled to adenylyl cyclase. Biochem. Biophys. Res. Commun.198:605–612.
Patel, Y.C., M.T. Greenwood, R. Panetta, L. Demchyshyn, H. Niznik, andC.B. Srikant (1995) Minireview. The somatostatin receptor family. LifeSci. 57:1249–1265.
Prinz, C., G. Sachs, J.H. Walsh, D.H. Coy, and S.V. Wu (1994) Thesomatostatin receptor subtype on rat enterochromaffin-like cells. Gas-troenterology 107:1067–1074.
Raynor, K., and T. Reisine (1992) Differential coupling of somatostatin1receptors to adenyll cyclase in the rat striatum vs. the pituitary andother regions of the rat brain. J. Pharmacol. Expl. Ther. 260:841–848.
Reichlin, S. (1983) SRIF. New Engl. J. Med. 309:1495–1563.Reisine, T., and G.I. Bell (1995) Molecular properties of somatostatin
receptors. Commentary. Neuroscience 67:777–790.
Reisine, T., H. Kong, K. Raynor, H. Yano, J. Takeda, K. Yasuda, and G.I. Bell(1993) Splice variant of the somatostatin receptor 2 subtype, somatosta-tin receptor 2B, couples to adenylyl cyclase. Mol. Pharmacol. 44:1008–1015.
Rens-Domiano, S., and T. Reisine (1992) Structural analysis and functionalrole of the carbohydrate component of rat brain SRIF receptors. J. Biol.Chem. 266:20094–20102.
Reyl, F.J., and M.J.M. Lewin (1982) Intracellular receptor for somatostatinin gastric mucosal cells: Decomposition and reconstitution of somatosta-tin-stimulated phosphoprotein phosphatases. Proc. Natl. Acad. Sci.USA 79:978–982.
Rossowski, W., and D. Coy (1994) Specific inhibition of rat pancreaticinsulin and glucagon release by receptor-selective somatostatin ana-logs. Biochem. Biophys. Res. Commun. 205:341–346.
Sanders, K.M. (1996) Special reports and reviews. A case for interstitialcells of Cajal as pacemakers and mediators of neurotransmission in thegastrointestinal tract. Gastroenterology 111:492–515.
Song, Z-M., S.J.H. Brookes, P.A. Steele, and M. Costa (1992) Projectionsand pathways of submucous neurons to the mucosa of the guinea pigsmall intestine. Cell Tiss. Res. 269:87–98.
Sternini, C., and K. Anderson (1992) Calcitonin gene-related peptide-containing neurons in the digestive system: Differential distributionand expression pattern. Somatos. Mot. Res. 9:45–59.
Sternini, C., J.R. Reeve, Jr., and N. Brecha (1987) Distribution andcharacterization of calcitonin gene-related peptide immunoreactivity inthe digestive system of normal and capsaicin-treated rats. Gastroenter-ology 93:852–862.
Sternini, C., D. Su, P.D. Gamp, and N.W. Bunnett (1995) Cellular sites ofexpression of the neurokinin-1 receptor in the rat gastrointestinal tract.J. Comp. Neurol. 358:531–540.
Teitelbaum, D.H., T.M. O’Dorisio, W.E. Perkins, and T.S. Gaginella (1984)Somatostatin modulation of peptide-induced acetylcholine release inguinea pig ileum. Am. J. Physiol. 246:G509–G514.
Theveniau, M., K. Yasuda, G.I. Bell, and T. Reisine (1994) Immunologicaldetection of isoforms of SRIF receptor subtype, sstR2. J. Neurochem.63:447–455.
Thuneberg, L. (1982) Interstitial cells of Cajal: Intestinal pacemaker cells?Adv. Anat. Embryol. Cell Biol. 71:1–130.
Vanetti, M., M. Kouba, X. Wang, G. Vogt, and V. Hollt (1992) Cloning andexpression of a novel mouse somatostatin receptor (sstR2B). FEBS311:290–294.
Walsh, J.H., and H.C. Wong (1987) Radioimmunoassay of gastrointestinalpolypeptides. In C. Patrono and B.A. Peskar (eds): Handbook ofExperimental Pharmacology: Radioimmunoassay in Basic and ClinicalPharmacology. New York: Springer-Verlag, Vol. 82, pp. 313–350.
Warhurst, G., N.B. Higgs, H. Fakhoury, A.C. Warhurst, J. Garde, and D.H.Coy (1996) Somatostatin receptor subtype 2 mediates somatostatininhibition of ion secretion in rat distal colon. Gastroenterology 111:325–333.
Wong, H.C., J.H. Walsh, H. Yang, Y. Tache, and A.M.J. Buchan (1990) Amonoclonal antibody to somatostatin with potent immunoneutralizingactivity. Peptides 11:707–712.
Wong, H.C., C. Sternini, K. Lloyd, R. De Giorgio, and J.H. Walsh (1996)Monoclonal antibody to VIP: Production, characterization, immunoneu-tralizing activity, and usefulness in cytochemical staining. Hybridoma15:133–139.
Yamada, T., and T. Chiba (1989) Somatostatin. In S.T. Schultz and G.M.Makhlouf (eds): Handbook of Physiology: The Gastrointestinal SystemII. Oxford: Oxford University Press, pp. 431–453.
Yamada, Y., S.R.T. Post, K. Wang, H.S. Tager, G.I. Bell, and S. Seino (1992)Cloning and functional characterization of a family of human somatosta-tin receptors expressed in brain, gastrointestinal tract and kidney. Proc.Natl. Acad. Sci. USA 89:251–255.
Yasuda, K., S. Rens-Domiano, C.D. Breder, S.F. Law, C.B. Saper, T. Reisine,and G.I. Bell (1992) Cloning of a novel somatostatin receptor, SSTR3,coupled to adenylyl cyclase. J. Biol. Chem. 267:20422–20428.
Yau, W.M., P.F. Lingle, and M.L. Youther (1983) Modulation of cholinergicneurotransmitter release from myenteric plexus by somatostatin. Pep-tides 4:49–53.
408 C. STERNINI ET AL.





![Research Paper The human somatostatin receptor type 2 as ... · unstable rep orter gene expression [10] . Furthermore, a heterogeneous cell population will be generated, as cells](https://img.pdfslide.net/doc/110x75/5f2aedb955b91e04354cc70e/research-paper-the-human-somatostatin-receptor-type-2-as-unstable-rep-orter.jpg)






















