Embed Size (px)
Citation preview

Spliceostatin hemiketal biosynthesis in Burkholderiaspp. is catalyzed by an iron/α-ketoglutarate–dependent dioxygenaseAlessandra S. Eustáquio1, Jeffrey E. Janso, Anokha S. Ratnayake, Christopher J. O’Donnell, and Frank E. Koehn
Natural Products Laboratory, Worldwide Medicinal Chemistry, Pfizer Worldwide Research and Development, Groton, CT 06340
Edited by Jerrold Meinwald, Cornell University, Ithaca, NY, and approved July 15, 2014 (received for review May 6, 2014)
Spliceostatins are potent spliceosome inhibitors biosynthesized bya hybrid nonribosomal peptide synthetase−polyketide synthase(NRPS−PKS) system of the trans-acyl transferase (AT) type. Bur-kholderia sp. FERM BP-3421 produces hemiketal spliceostatins,such as FR901464, as well as analogs containing a terminal carbox-ylic acid. We provide genetic and biochemical evidence for hemi-ketal biosynthesis by oxidative decarboxylation rather than thepreviously hypothesized Baeyer–Villiger oxidative release postu-lated to be catalyzed by a flavin-dependent monooxygenase(FMO) activity internal to the last module of the PKS. Inactivationof Fe(II)/α-ketoglutarate–dependent dioxygenase gene fr9P led toloss of hemiketal congeners, whereas the mutant was still able toproduce all major carboxylic acid-type compounds. FMO mutants,on the other hand, produced both hemiketal and carboxylic acidanalogs containing an exocyclic methylene instead of an epoxide,indicating that the FMO is involved in epoxidation rather thanBaeyer–Villiger oxidation. Moreover, recombinant Fr9P enzymewas shown to catalyze hydroxylation to form β-hydroxy acids,which upon decarboxylation led to hemiketal FR901464. Finally,a third oxygenase activity encoded in the biosynthetic gene clus-ter, the cytochrome P450 monooxygenase Fr9R, was assigned asa 4-hydroxylase based on gene inactivation results. Identificationand deletion of the gene involved in hemiketal formation allowedus to generate a strain—the dioxygenase fr9P− mutant—that accu-mulates only the carboxylic acid-type spliceostatins, which are aspotent as the hemiketal analogs, when derivatized to increase cellpermeability, but are chemically more stable.
thailanstatin | natural product | bacteria | secondary metabolite
The spliceosome, a multi-megadalton ribonucleoprotein complexthat is involved in mRNA processing in eukaryotic cells, has
emerged as a promising target in cancer therapy (1). Mis-regulation of mRNA splicing and mutations in the splicingmachinery has been observed in a variety of cancers. Splicingmodulation with small molecules provides a potential avenue fortreatment (2, 3). Three classes of bacterial natural products (Fig.1A)—pladienolide/FD-895 (4–6), spliceostatin/FR901464 (7, 8),and herboxidiene (GEX1A) (9)—have been shown to modulatesplicing activity by binding to the SF3b spliceosome subunit (10–12), exhibiting low to subnanomolar half-maximal inhibitoryconcentration (IC50) values against different cancer cell lines.The mode of action (MoA) and potent cytotoxicity of theseagents led to efforts in advancing spliceosome inhibitors asantitumor drugs. Results of a phase I clinical trial of a semi-synthetic analog of pladienolide D, E7107, have been recentlypublished (13), providing early clinical evidence for spliceo-some inhibition as a potentially viable MoA to treat cancer.The challenging polyketide structures of spliceosome inhib-
itors coupled with clinical potential have inspired the syntheticchemistry community. As a result, several total synthesis routesto pladienolide, spliceostatin, and herboxidiene classes of naturalproducts have been reported (for the most recent, see refs. 14–16). For spliceostatins, the shortest described synthetic routecontains 20 total steps, with the longest linear sequence being 10
steps (14). Harnessing the biosynthetic potential of the producingorganisms for compound production and analog generation is anattractive alternative to chemical synthesis. Pladienolide (17)and herboxidiene (18) are biosynthesized by different Streptomycesspecies via a canonical type I polyketide synthase (PKS) system,whereas biosynthesis of spliceostatins in two Burkholderia strains(FERM BP-3421 and MSMB 43) is encoded by highly homol-ogous, hybrid trans-acyl transferase (AT) PKS–nonribosomalpeptide synthetase (NRPS) gene clusters (19, 20).Hemiketal-bearing compounds such as FR901464 (Fig. 1B, 4)
were described from strain FERM BP-3421 in 1997 (8). FERMBP-3421 had been identified as Pseudomonas sp. no. 2663 butwas recently reclassified as Burkholderia (21, 22). A more stable,semisynthetic methyl ketal of 4 was later shown to inhibit thespliceosome, and it was thus termed spliceostatin A (Fig. 1A)(11, 23). Subsequently, an analog bearing a terminal carboxylicacid (3) was reported from Burkholderia sp. MSMB 43 (20, 24).Although 3 belongs to the same family of compounds as 4 and 5,the name thailanstatin A was used when reporting compound 3,as it had been isolated from a Burkholderia thailandensis-likestrain. It has recently been shown that strain FERM BP-3421produces carboxylic acids 1, 2, and 3 in addition to hemiketals 4and 5 (22, 25), whereas only 3 and two chlorohydrin adducts of 3have been isolated from MSMB 43 (20). We chose to use theterm spliceostatin in this manuscript to refer to all congeners 1–5
Significance
Spliceostatins are bacterial natural products that show prom-ising anticancer activity. Understanding how the bacteriummakes spliceostatins will aid efforts toward a sustainable routefor their production. Moreover, altering the chemical structureof a natural product is usually necessary to improve its phar-maceutical properties. For example, the parent spliceostatinmolecule contains an unstable hemiketal chemical group.Contrary to previous hypotheses, we report on the identifica-tion of a dioxygenase enzyme responsible for hemiketal bio-synthesis. Deletion of the corresponding dioxygenase gene ledto a strain that produces exclusively spliceostatin congenersthat are more stable than, and as active as, the parent com-pound, when derivatized to increase cell permeability. Thestrain generated in this study will be the basis for futuredevelopment.
Author contributions: A.S.E., J.E.J., and A.S.R. designed research; A.S.E., J.E.J., and A.S.R.performed research; A.S.E., J.E.J., A.S.R., C.J.O., and F.E.K. analyzed data; and A.S.E. wrotethe paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Freely available online through the PNAS open access option.
Data deposition: The DNA sequence of the fr9 biosynthetic gene cluster reported in thispaper has been deposited in the GenBank database (accession no. KJ461964).1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1408300111/-/DCSupplemental.
E3376–E3385 | PNAS | Published online August 5, 2014 www.pnas.org/cgi/doi/10.1073/pnas.1408300111
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0

in an attempt to unify the terminology of this compound class.Spliceostatins bearing a terminal carboxylic acid have beenreported to inhibit mRNA splicing in vitro as effectively as thecorresponding hemiketal, while having improved stability (20).We have shown that the free acids themselves are less cytotoxicthan the hemiketals against cancer cell lines. However, activity isrescued when carboxylic acid compounds are derivatized (e.g., asmethyl esters or propyl amides) presumably due to improved cellpermeability (22). As such, the carboxyl analog series representsan attractive chemical lead for anticancer drug discovery.We were particularly intrigued by how hemiketals are bio-
synthesized, and compelled by the opportunity to leverage bio-synthetic knowledge to block hemiketal formation and accumulatestable carboxylic acid analogs in fermentation. The biosynthetichypothesis proposed by Tang and coworkers (19, 26) embodies anunprecedented Baeyer–Villiger oxidation catalyzed by a flavin-
dependent monooxygenase (FMO) domain present in the lastmodule of the trans-AT PKS (Fig. 1C). If this is correct, domainskipping may explain the formation of acids over hemiketals, asproposed by Liu et al. (20). However, we envisioned oxidativedecarboxylation as a plausible alternative hypothesis, in whichcarboxylic acid compounds would be biosynthesized first, followedby hydroxylation and decarboxylation to form the hemiketalcongeners (Fig. 1D). We herein assign functions to three oxy-genases encoded in the spliceostatin biosynthetic gene cluster andprovide genetic and biochemical evidence for the oxidativedecarboxylation hypothesis.
ResultsGenome Sequencing and Identification of the Spliceostatin BiosyntheticGene Cluster. We independently sequenced the genome ofBurkholderia sp. FERM BP-3421 using next-generation technologies
O
OOH
O
COOH
Herboxidiene (GEX1A)
A
DO
HOO
RCOOH
4
OOH
HOO
R
PKS
3
1 2
ACP9
S
R
O
OH
OH
ACP9
S
O
R
O
HO
HO
CO2
R
O
OH
H+
O
R
OH
FMO9
O2
DH9 / TE9 4
B
DH9
oxidative
decarboxylation
OOH
HOO
O
NH
AcOO
FR901464 (4), R=HFR901465 (5), R=OH
R
O
RO
COOHSpliceostatin C (2), R=HThailanstatin A (3), R=OH
OCOOHSpliceostatin B (1)
OOCH3
HOO
O
NH
AcOO
Spliceostatin A
A
A
O
NH
AcOO
E
R=
Pladienolide D (R1=H, R2=H, R3=OH, R4=Ac)FD-895 (R1=CH3, R2=OH, R3=H, R4=Ac)
E7107 (R1=H, R2=H, R3=OH, R4= )N NO
OR1
O
O OR4
OHOH
OR2
R3
C
O
R
COOH
TE9 2, 3
1
SO
HO
R
ACP9
H+
SO
O
R
ACP9
H2O
H
H
H
H
H
HH
HH
H
3
157911
13
15
1'3'
5'
R
HO
OHH
Fig. 1. Structures of spliceosome inhibitors and hypotheses for spliceostatin biosynthesis. (A) Structural classes of spliceosome inhibitors. Spliceostatin A is a morestable, semisynthetic analog of natural product FR901464 (11, 23). (B) Known spliceostatin analogs that are the subject of this study. (C) Baeyer–Villiger oxidationhypothesis for spliceostatin hemiketal formation according to Zhang et al. (19) (Upper); biosynthesis of acids over hemiketals by FMO domain skipping, adaptedfrom Liu et al. (20) (Lower). ACP, acyl carrier protein; FMO, flavin-dependent monooxygenase; DH, dehydratase; TE, thioesterase. The subscript 9 indicates the PKSmodule in which the enzyme activities are present. (D) Alternative, oxidative decarboxylation hypothesis. The R group is indicated in C.
Eustáquio et al. PNAS | Published online August 5, 2014 | E3377
BIOCH
EMISTR
YPN
ASPL
US
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0
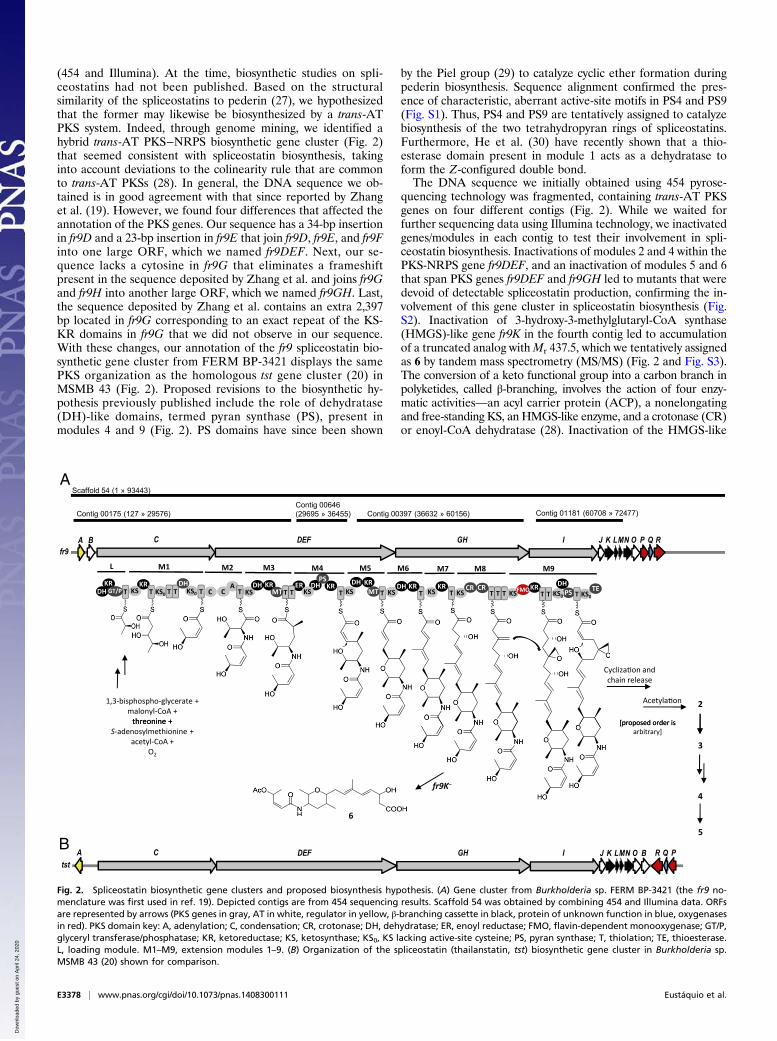
(454 and Illumina). At the time, biosynthetic studies on spli-ceostatins had not been published. Based on the structuralsimilarity of the spliceostatins to pederin (27), we hypothesizedthat the former may likewise be biosynthesized by a trans-ATPKS system. Indeed, through genome mining, we identified ahybrid trans-AT PKS−NRPS biosynthetic gene cluster (Fig. 2)that seemed consistent with spliceostatin biosynthesis, takinginto account deviations to the colinearity rule that are commonto trans-AT PKSs (28). In general, the DNA sequence we ob-tained is in good agreement with that since reported by Zhanget al. (19). However, we found four differences that affected theannotation of the PKS genes. Our sequence has a 34-bp insertionin fr9D and a 23-bp insertion in fr9E that join fr9D, fr9E, and fr9Finto one large ORF, which we named fr9DEF. Next, our se-quence lacks a cytosine in fr9G that eliminates a frameshiftpresent in the sequence deposited by Zhang et al. and joins fr9Gand fr9H into another large ORF, which we named fr9GH. Last,the sequence deposited by Zhang et al. contains an extra 2,397bp located in fr9G corresponding to an exact repeat of the KS-KR domains in fr9G that we did not observe in our sequence.With these changes, our annotation of the fr9 spliceostatin bio-synthetic gene cluster from FERM BP-3421 displays the samePKS organization as the homologous tst gene cluster (20) inMSMB 43 (Fig. 2). Proposed revisions to the biosynthetic hy-pothesis previously published include the role of dehydratase(DH)-like domains, termed pyran synthase (PS), present inmodules 4 and 9 (Fig. 2). PS domains have since been shown
by the Piel group (29) to catalyze cyclic ether formation duringpederin biosynthesis. Sequence alignment confirmed the pres-ence of characteristic, aberrant active-site motifs in PS4 and PS9(Fig. S1). Thus, PS4 and PS9 are tentatively assigned to catalyzebiosynthesis of the two tetrahydropyran rings of spliceostatins.Furthermore, He et al. (30) have recently shown that a thio-esterase domain present in module 1 acts as a dehydratase toform the Z-configured double bond.The DNA sequence we initially obtained using 454 pyrose-
quencing technology was fragmented, containing trans-AT PKSgenes on four different contigs (Fig. 2). While we waited forfurther sequencing data using Illumina technology, we inactivatedgenes/modules in each contig to test their involvement in spli-ceostatin biosynthesis. Inactivations of modules 2 and 4 within thePKS-NRPS gene fr9DEF, and an inactivation of modules 5 and 6that span PKS genes fr9DEF and fr9GH led to mutants that weredevoid of detectable spliceostatin production, confirming the in-volvement of this gene cluster in spliceostatin biosynthesis (Fig.S2). Inactivation of 3-hydroxy-3-methylglutaryl-CoA synthase(HMGS)-like gene fr9K in the fourth contig led to accumulationof a truncated analog withMr 437.5, which we tentatively assignedas 6 by tandem mass spectrometry (MS/MS) (Fig. 2 and Fig. S3).The conversion of a keto functional group into a carbon branch inpolyketides, called β-branching, involves the action of four enzy-matic activities—an acyl carrier protein (ACP), a nonelongatingand free-standing KS, an HMGS-like enzyme, and a crotonase (CR)or enoyl-CoA dehydratase (28). Inactivation of the HMGS-like
A
PS
2
Scaffold 54 (1 » 93443)
TT KSTDH KR
KS TDH
T KSKR
KST KSKR
TKR
KS DHT T KS0 T T T KSA
CKR
TDH ER
KST TKR KR
T T TKR
DH C
PSTEMT
DH CR CRDHGT/P FMOMT
L M1 M2 M3 M4 M5 M6 M7 M8 M9
1,3-bisphospho-glycerate +malonyl-CoA +
threonine + [proposed order is
Cycliza on and chain release
Acetyla on
KS0KS0 KS0
Contig 00175 (127 » 29576)Contig 00646 (29695 » 36455) Contig 00397 (36632 » 60156) Contig 01181 (60708 » 72477)
DHGT/P
4
3
threonine +S-adenosylmethionine +
acetyl-CoA +O2
[proposed order is arbitrary]
B
fr9K−
6
5
Fig. 2. Spliceostatin biosynthetic gene clusters and proposed biosynthesis hypothesis. (A) Gene cluster from Burkholderia sp. FERM BP-3421 (the fr9 no-menclature was first used in ref. 19). Depicted contigs are from 454 sequencing results. Scaffold 54 was obtained by combining 454 and Illumina data. ORFsare represented by arrows (PKS genes in gray, AT in white, regulator in yellow, β-branching cassette in black, protein of unknown function in blue, oxygenasesin red). PKS domain key: A, adenylation; C, condensation; CR, crotonase; DH, dehydratase; ER, enoyl reductase; FMO, flavin-dependent monooxygenase; GT/P,glyceryl transferase/phosphatase; KR, ketoreductase; KS, ketosynthase; KS0, KS lacking active-site cysteine; PS, pyran synthase; T, thiolation; TE, thioesterase.L, loading module. M1–M9, extension modules 1–9. (B) Organization of the spliceostatin (thailanstatin, tst) biosynthetic gene cluster in Burkholderia sp.MSMB 43 (20) shown for comparison.
E3378 | www.pnas.org/cgi/doi/10.1073/pnas.1408300111 Eustáquio et al.
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0
gene did not furnish a spliceostatin analog containing a ketogroup in place of the exocyclic methylene/epoxide, but insteadled to apparent interruption of biosynthesis at the time ofβ-branching (Fig. 2). This result is in accord with the establishedsubstrate specificity of trans-AT KSs (31). Canonical cis-AT PKSsevolved by gene duplication, resulting in KS domains that code fora particularmetabolite sharing high sequence identity, and allowingthe production of structurallymodified natural products by domain/module engineering [albeit with possible reduction in reaction rates(32)]. In contrast, trans-ATKSdomains aremore dissimilar and canbe phylogenetically grouped by the type of substrates processed(e.g., β-hydroxylated, α-methylated, α,β-unsaturated, β-branched)(28), indicating that successful PKS engineering toward structuremodification of polyketides biosynthesized by trans-AT PKSs mayrequire KS swap or mutagenesis.
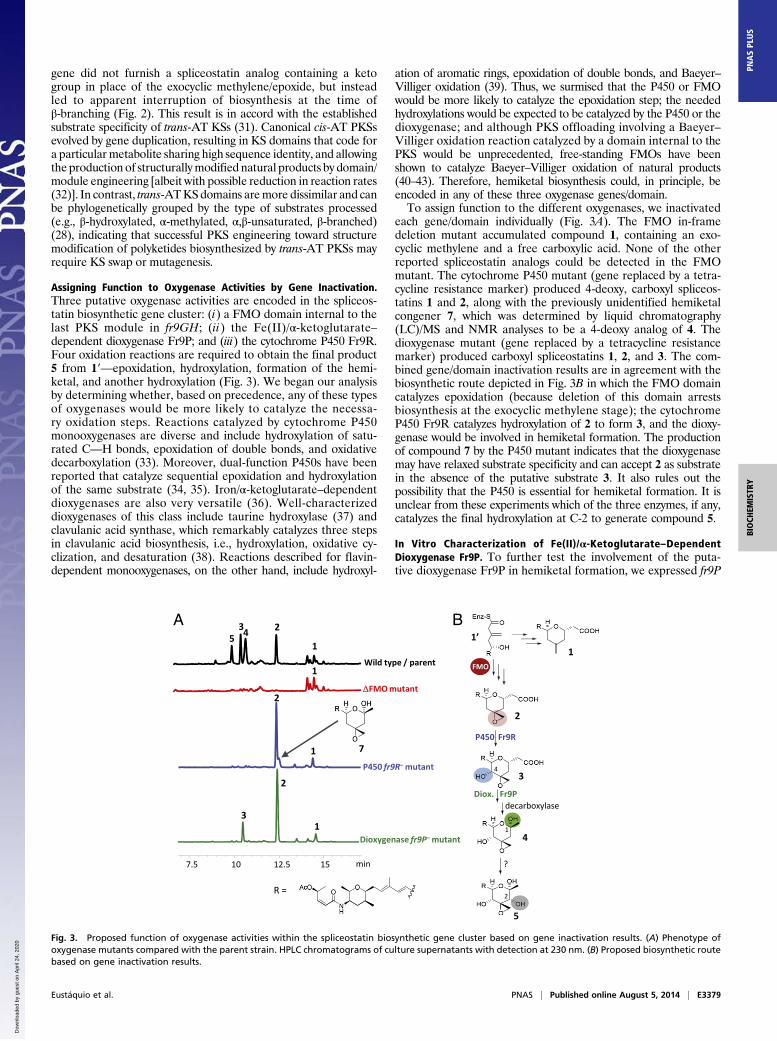
Assigning Function to Oxygenase Activities by Gene Inactivation.Three putative oxygenase activities are encoded in the spliceos-tatin biosynthetic gene cluster: (i) a FMO domain internal to thelast PKS module in fr9GH; (ii) the Fe(II)/α-ketoglutarate–dependent dioxygenase Fr9P; and (iii) the cytochrome P450 Fr9R.Four oxidation reactions are required to obtain the final product5 from 1′—epoxidation, hydroxylation, formation of the hemi-ketal, and another hydroxylation (Fig. 3). We began our analysisby determining whether, based on precedence, any of these typesof oxygenases would be more likely to catalyze the necessa-ry oxidation steps. Reactions catalyzed by cytochrome P450monooxygenases are diverse and include hydroxylation of satu-rated C—H bonds, epoxidation of double bonds, and oxidativedecarboxylation (33). Moreover, dual-function P450s have beenreported that catalyze sequential epoxidation and hydroxylationof the same substrate (34, 35). Iron/α-ketoglutarate–dependentdioxygenases are also very versatile (36). Well-characterizeddioxygenases of this class include taurine hydroxylase (37) andclavulanic acid synthase, which remarkably catalyzes three stepsin clavulanic acid biosynthesis, i.e., hydroxylation, oxidative cy-clization, and desaturation (38). Reactions described for flavin-dependent monooxygenases, on the other hand, include hydroxyl-
ation of aromatic rings, epoxidation of double bonds, and Baeyer–Villiger oxidation (39). Thus, we surmised that the P450 or FMOwould be more likely to catalyze the epoxidation step; the neededhydroxylations would be expected to be catalyzed by the P450 or thedioxygenase; and although PKS offloading involving a Baeyer–Villiger oxidation reaction catalyzed by a domain internal to thePKS would be unprecedented, free-standing FMOs have beenshown to catalyze Baeyer–Villiger oxidation of natural products(40–43). Therefore, hemiketal biosynthesis could, in principle, beencoded in any of these three oxygenase genes/domain.To assign function to the different oxygenases, we inactivated
each gene/domain individually (Fig. 3A). The FMO in-framedeletion mutant accumulated compound 1, containing an exo-cyclic methylene and a free carboxylic acid. None of the otherreported spliceostatin analogs could be detected in the FMOmutant. The cytochrome P450 mutant (gene replaced by a tetra-cycline resistance marker) produced 4-deoxy, carboxyl spliceos-tatins 1 and 2, along with the previously unidentified hemiketalcongener 7, which was determined by liquid chromatography(LC)/MS and NMR analyses to be a 4-deoxy analog of 4. Thedioxygenase mutant (gene replaced by a tetracycline resistancemarker) produced carboxyl spliceostatins 1, 2, and 3. The com-bined gene/domain inactivation results are in agreement with thebiosynthetic route depicted in Fig. 3B in which the FMO domaincatalyzes epoxidation (because deletion of this domain arrestsbiosynthesis at the exocyclic methylene stage); the cytochromeP450 Fr9R catalyzes hydroxylation of 2 to form 3, and the dioxy-genase would be involved in hemiketal formation. The productionof compound 7 by the P450 mutant indicates that the dioxygenasemay have relaxed substrate specificity and can accept 2 as substratein the absence of the putative substrate 3. It also rules out thepossibility that the P450 is essential for hemiketal formation. It isunclear from these experiments which of the three enzymes, if any,catalyzes the final hydroxylation at C-2 to generate compound 5.
In Vitro Characterization of Fe(II)/α-Ketoglutarate–DependentDioxygenase Fr9P. To further test the involvement of the puta-tive dioxygenase Fr9P in hemiketal formation, we expressed fr9P
min7.5 10 12.5 15
Dioxygenase fr9P– mutant
P450 fr9R– mutant
FMO mutant
Wild type / parent
R =
7
45
1
1
31
2
2
A B
1
23
FMO
P450 Fr9R
Diox. Fr9P
?
decarboxylase
2
3
4
1’
2
4
1
1
5
Fig. 3. Proposed function of oxygenase activities within the spliceostatin biosynthetic gene cluster based on gene inactivation results. (A) Phenotype ofoxygenase mutants compared with the parent strain. HPLC chromatograms of culture supernatants with detection at 230 nm. (B) Proposed biosynthetic routebased on gene inactivation results.
Eustáquio et al. PNAS | Published online August 5, 2014 | E3379
BIOCH
EMISTR
YPN
ASPL
US
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0
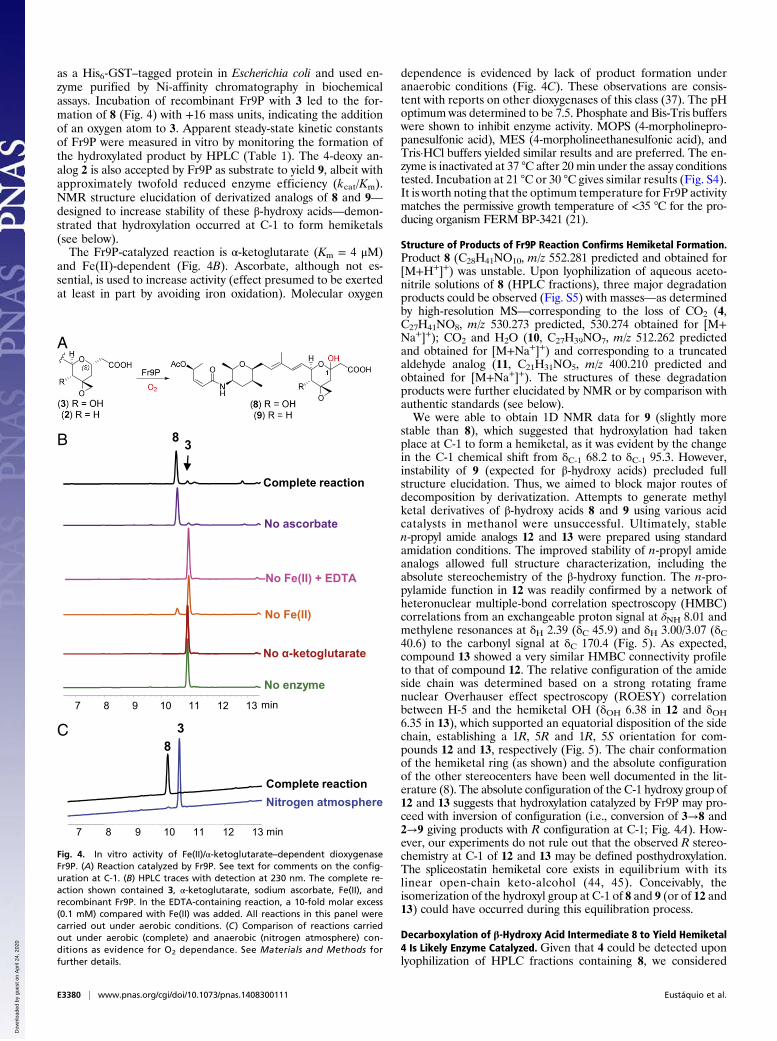
as a His6-GST–tagged protein in Escherichia coli and used en-zyme purified by Ni-affinity chromatography in biochemicalassays. Incubation of recombinant Fr9P with 3 led to the for-mation of 8 (Fig. 4) with +16 mass units, indicating the additionof an oxygen atom to 3. Apparent steady-state kinetic constantsof Fr9P were measured in vitro by monitoring the formation ofthe hydroxylated product by HPLC (Table 1). The 4-deoxy an-alog 2 is also accepted by Fr9P as substrate to yield 9, albeit withapproximately twofold reduced enzyme efficiency (kcat/Km).NMR structure elucidation of derivatized analogs of 8 and 9—designed to increase stability of these β-hydroxy acids—demon-strated that hydroxylation occurred at C-1 to form hemiketals(see below).The Fr9P-catalyzed reaction is α-ketoglutarate (Km = 4 μM)
and Fe(II)-dependent (Fig. 4B). Ascorbate, although not es-sential, is used to increase activity (effect presumed to be exertedat least in part by avoiding iron oxidation). Molecular oxygen
dependence is evidenced by lack of product formation underanaerobic conditions (Fig. 4C). These observations are consis-tent with reports on other dioxygenases of this class (37). The pHoptimumwas determined to be 7.5. Phosphate and Bis-Tris bufferswere shown to inhibit enzyme activity. MOPS (4-morpholinepro-panesulfonic acid), MES (4-morpholineethanesulfonic acid), andTris·HCl buffers yielded similar results and are preferred. The en-zyme is inactivated at 37 °C after 20 min under the assay conditionstested. Incubation at 21 °C or 30 °C gives similar results (Fig. S4).It is worth noting that the optimum temperature for Fr9P activitymatches the permissive growth temperature of <35 °C for the pro-ducing organism FERM BP-3421 (21).
Structure of Products of Fr9P Reaction Confirms Hemiketal Formation.Product 8 (C28H41NO10, m/z 552.281 predicted and obtained for[M+H+]+) was unstable. Upon lyophilization of aqueous aceto-nitrile solutions of 8 (HPLC fractions), three major degradationproducts could be observed (Fig. S5) with masses—as determinedby high-resolution MS—corresponding to the loss of CO2 (4,C27H41NO8, m/z 530.273 predicted, 530.274 obtained for [M+Na+]+); CO2 and H2O (10, C27H39NO7, m/z 512.262 predictedand obtained for [M+Na+]+) and corresponding to a truncatedaldehyde analog (11, C21H31NO5, m/z 400.210 predicted andobtained for [M+Na+]+). The structures of these degradationproducts were further elucidated by NMR or by comparison withauthentic standards (see below).We were able to obtain 1D NMR data for 9 (slightly more
stable than 8), which suggested that hydroxylation had takenplace at C-1 to form a hemiketal, as it was evident by the changein the C-1 chemical shift from δC-1 68.2 to δC-1 95.3. However,instability of 9 (expected for β-hydroxy acids) precluded fullstructure elucidation. Thus, we aimed to block major routes ofdecomposition by derivatization. Attempts to generate methylketal derivatives of β-hydroxy acids 8 and 9 using various acidcatalysts in methanol were unsuccessful. Ultimately, stablen-propyl amide analogs 12 and 13 were prepared using standardamidation conditions. The improved stability of n-propyl amideanalogs allowed full structure characterization, including theabsolute stereochemistry of the β-hydroxy function. The n-pro-pylamide function in 12 was readily confirmed by a network ofheteronuclear multiple-bond correlation spectroscopy (HMBC)correlations from an exchangeable proton signal at δNH 8.01 andmethylene resonances at δH 2.39 (δC 45.9) and δH 3.00/3.07 (δC40.6) to the carbonyl signal at δC 170.4 (Fig. 5). As expected,compound 13 showed a very similar HMBC connectivity profileto that of compound 12. The relative configuration of the amideside chain was determined based on a strong rotating framenuclear Overhauser effect spectroscopy (ROESY) correlationbetween H-5 and the hemiketal OH (δOH 6.38 in 12 and δOH6.35 in 13), which supported an equatorial disposition of the sidechain, establishing a 1R, 5R and 1R, 5S orientation for com-pounds 12 and 13, respectively (Fig. 5). The chair conformationof the hemiketal ring (as shown) and the absolute configurationof the other stereocenters have been well documented in the lit-erature (8). The absolute configuration of the C-1 hydroxy group of12 and 13 suggests that hydroxylation catalyzed by Fr9P may pro-ceed with inversion of configuration (i.e., conversion of 3→8 and2→9 giving products with R configuration at C-1; Fig. 4A). How-ever, our experiments do not rule out that the observed R stereo-chemistry at C-1 of 12 and 13 may be defined posthydroxylation.The spliceostatin hemiketal core exists in equilibrium with itslinear open-chain keto-alcohol (44, 45). Conceivably, theisomerization of the hydroxyl group at C-1 of 8 and 9 (or of 12 and13) could have occurred during this equilibration process.
Decarboxylation of β-Hydroxy Acid Intermediate 8 to Yield Hemiketal4 Is Likely Enzyme Catalyzed. Given that 4 could be detected uponlyophilization of HPLC fractions containing 8, we considered
No Fe(II) + EDTA
3
No Fe(II)
No ascorbate
Complete reaction
8
A
B
No enzyme
No α-ketoglutarate
min7 8 9 10 11 12 13
C
min7 8 9 10 11 12 13
Nitrogen atmosphereComplete reaction
38
Fig. 4. In vitro activity of Fe(II)/α-ketoglutarate–dependent dioxygenaseFr9P. (A) Reaction catalyzed by Fr9P. See text for comments on the config-uration at C-1. (B) HPLC traces with detection at 230 nm. The complete re-action shown contained 3, α-ketoglutarate, sodium ascorbate, Fe(II), andrecombinant Fr9P. In the EDTA-containing reaction, a 10-fold molar excess(0.1 mM) compared with Fe(II) was added. All reactions in this panel werecarried out under aerobic conditions. (C) Comparison of reactions carriedout under aerobic (complete) and anaerobic (nitrogen atmosphere) con-ditions as evidence for O2 dependance. See Materials and Methods forfurther details.
E3380 | www.pnas.org/cgi/doi/10.1073/pnas.1408300111 Eustáquio et al.
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0

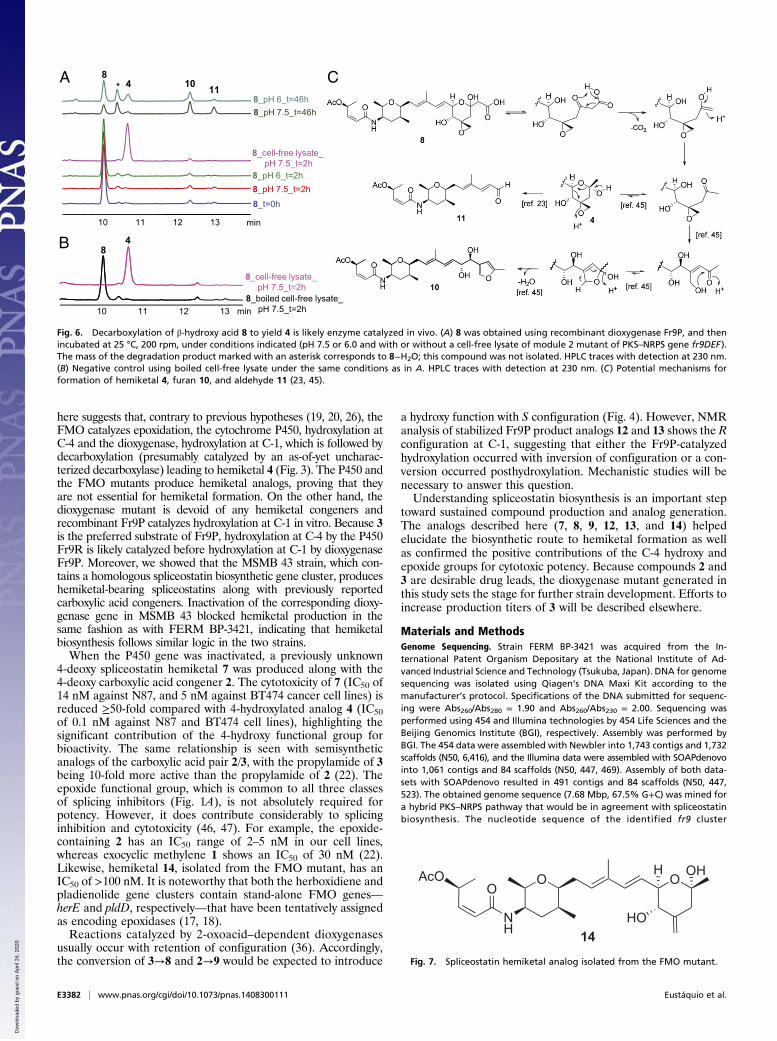
whether nonenzymatic decarboxylation of 8 is the in vivo route of4 production. The half-life of 8 at 25 °C (temperature used forcompound production) was determined to be 17 h at pH 7.5 and24 h at pH 6.0 (Fig. S6). Degradation products include the furan10 and the truncated aldehyde 11 (Fig. 6). Hemiketal 4 is a minorcomponent in both cases, with its formation being favored atlower pH, as it would be expected for nonenzymatic de-carboxylation. In contrast, incubation of 8 with a cell-free lysateof module 2 mutant of PKS–NRPS gene fr9DEF (chosen for itslack in spliceostatin production) leads nearly exclusively to 4,suggesting a role for a (as of yet) uncharacterized decarboxylase(Fig. 6). A candidate decarboxylase is the product of fr9Q, theonly gene with unassigned, putative function in the spliceostatingene cluster (Fig. 2). Fr9Q shares sequence similarity to hypo-thetical proteins/putative cyclases and was proposed by Zhanget al. (19) to play a role in the formation of the first tetrahy-dropyran ring of spliceostatins. However, because pyran synthasedomains in modules 4 and 9 are likely to catalyze the two nec-essary cyclizations, Fr9Q may well be the missing decarboxylase.A potential mechanism for formation of 4 includes decarboxylationof a β-ketoacid followed by recyclization (Fig. 6C). Alternatively,the decarboxylase may be present outside of the spliceostatingene cluster. Further experiments will be necessary to testthese hypotheses.
The FMO Mutant Is Capable of Producing a Hemiketal Analog. To testthe oxidative decarboxylation hypothesis further, we searched forhemiketal congeners in the FMO mutant. First, we showed thatthe known exocyclic methylene derivative 1 produced by theFMO mutant is accepted as substrate by dioxygenase Fr9P invitro (Fig. S7). We were then indeed able to isolate and elucidatethe structure of the previously unidentified hemiketal analog 14(Fig. 7) from the FMO mutant, proving that the FMO is notessential for hemiketal formation in vivo. Although Tang et al.(26) demonstrated that the FMO can catalyze Baeyer–Villigeroxidation in vitro, the truncated, surrogate substrate that wasused lacked a double bond. Thus, it is reasonable to speculatethat the reaction observed in vitro is not the preferred reaction invivo. Indeed, the indirect evidence provided here suggests therole for the FMO as an epoxidase.
Production of Spliceostatins by Burkholderia sp. MSMB 43 andDeletion of the Dioxygenase Gene tstP. As mentioned above, therecently described tst gene cluster from Burkholderia sp. MSMB43 is highly homologous to the fr9 gene cluster. Likewise, the tstcluster encodes three oxygenases, i.e., FMO (OX) domain inTstGH, cytochrome P450 TstR, and dioxygenase TstP (Fig. 2)(20). However, only analogs containing a carboxyl moiety havebeen described from the MSMB 43 strain, i.e., 3, and two chlo-rohydrin adducts of 3, one of them containing an isobutyrylgroup instead of the distal acetyl group (20). In addition to 3, wefound that MSMB 43 also produces carboxyl analog 2 andhemiketal congeners 4 and 5 when fermented under the sameconditions as FERM BP-3421 (Fig. S8A). The production ofboth hemiketal- and carboxyl-containing spliceostatins by MSMB43 suggests that all three oxygenases in the tst cluster are active.To test whether the dioxygenase TstP has a role in hemiketalformation, an in-frame deletion of tstP was performed in MSMB43. Similar to the inactivation of fr9P in FERM BP-3421, the tstPdeletion mutant produced only 2 and 3 in fermentation; nohemiketal-containing spliceostatins were detected in the mutant(Fig. S8B). These results suggest that TstP is involved in hemi-ketal formation in MSMB 43 and that spliceostatin biosynthesisfollows the same logic in the two homologous pathways.
DiscussionSplicing inhibition offers a potential avenue for cancer treat-ment. Among the natural products known to target the spliceo-some, spliceostatins are of particular interest to us due to theirpotent bioactivity and intriguing biosynthesis. The other twoknown spliceosome inhibitor classes, pladienolides and herbox-idiene, are biosynthesized by Streptomyces spp. via a modularcis-AT PKS, while spliceostatins are produced by Burkholderiaspp. using the more recently described and evolutionary distincttrans-AT PKS system. Among the peculiarities of trans-AT PKSsis the large range of module varieties, including unusual domainorders, tandem domains, and atypical domain types (28). Severalfeatures of the spliceostatin PKS worth noting include (Fig. 2):(i) a TE domain in module 1 that has been recently shown to actas a dehydratase, generating a Z-configured double bond (30); (ii)two DH-like domains (termed PS) in modules 4 and 9 that areproposed to catalyze formation of the tetrahydropyran rings asdemonstrated during pederin biosynthesis (29); (iii) the integratedCR domains in module 8, as observed in pederin biosynthesis,and known to catalyze dehydration and decarboxylation duringβ-branching; and finally (iv) the FMO domain contained inmodule 9 of the PKS, which has been proposed to catalyze Baeyer–Villiger oxidative release to generate hemiketal congeners (19,20, 26).Besides the FMO, two other oxygenase activities are encoded
in the spliceostatin biosynthetic gene cluster, i.e., the cytochromeP450 fr9R and the Fe(II)/α-ketoglutarate–dependent dioxyge-nase fr9P as stand-alone genes. The combined evidence presented
Table 1. Apparent kinetic constants for Fr9P
Substrate Km, μM kcat, min−1 kcat/Km, μM−1·min−1Relative enzymeefficiency, %
3 60 ± 14 86 ± 9 1.43 1002 74 ± 7 60 ± 6 0.81 57
The kinetic values (at 30 °C, pH 7.5) are average of triplicate measure-ments ± SD.
OH
H
CH3
HO
H
R OH
HN Pr
O145
R = OH (12)R = H (13)
Key ROESY correlations
O
O
H
HH
H
HO H
H
H
H
HOH H
O
NHCH3
H H
H H
Selected HMBC correlations of then-propyl group in 12
8.0145.9
3.00 3.07
40.6170.4
1
4
5
6.384.335.67
5.24
3.3395.7
56.741.3
135.5
126.3
2.27
1.4971.4
46.2
2.314.55
68.4
2.73
2.39
Fig. 5. Key ROESY correlations depicting the relative configuration of 12 and 13 along with selected HMBC data and chemical shift assignments for compound 12.
Eustáquio et al. PNAS | Published online August 5, 2014 | E3381
BIOCH
EMISTR
YPN
ASPL
US
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0
here suggests that, contrary to previous hypotheses (19, 20, 26), theFMO catalyzes epoxidation, the cytochrome P450, hydroxylation atC-4 and the dioxygenase, hydroxylation at C-1, which is followed bydecarboxylation (presumably catalyzed by an as-of-yet uncharac-terized decarboxylase) leading to hemiketal 4 (Fig. 3). The P450 andthe FMO mutants produce hemiketal analogs, proving that theyare not essential for hemiketal formation. On the other hand, thedioxygenase mutant is devoid of any hemiketal congeners andrecombinant Fr9P catalyzes hydroxylation at C-1 in vitro. Because 3is the preferred substrate of Fr9P, hydroxylation at C-4 by the P450Fr9R is likely catalyzed before hydroxylation at C-1 by dioxygenaseFr9P. Moreover, we showed that the MSMB 43 strain, which con-tains a homologous spliceostatin biosynthetic gene cluster, produceshemiketal-bearing spliceostatins along with previously reportedcarboxylic acid congeners. Inactivation of the corresponding dioxy-genase gene in MSMB 43 blocked hemiketal production in thesame fashion as with FERM BP-3421, indicating that hemiketalbiosynthesis follows similar logic in the two strains.When the P450 gene was inactivated, a previously unknown
4-deoxy spliceostatin hemiketal 7 was produced along with the4-deoxy carboxylic acid congener 2. The cytotoxicity of 7 (IC50 of14 nM against N87, and 5 nM against BT474 cancer cell lines) isreduced ≥50-fold compared with 4-hydroxylated analog 4 (IC50of 0.1 nM against N87 and BT474 cell lines), highlighting thesignificant contribution of the 4-hydroxy functional group forbioactivity. The same relationship is seen with semisyntheticanalogs of the carboxylic acid pair 2/3, with the propylamide of 3being 10-fold more active than the propylamide of 2 (22). Theepoxide functional group, which is common to all three classesof splicing inhibitors (Fig. 1A), is not absolutely required forpotency. However, it does contribute considerably to splicinginhibition and cytotoxicity (46, 47). For example, the epoxide-containing 2 has an IC50 range of 2–5 nM in our cell lines,whereas exocyclic methylene 1 shows an IC50 of 30 nM (22).Likewise, hemiketal 14, isolated from the FMO mutant, has anIC50 of >100 nM. It is noteworthy that both the herboxidiene andpladienolide gene clusters contain stand-alone FMO genes—herE and pldD, respectively—that have been tentatively assignedas encoding epoxidases (17, 18).Reactions catalyzed by 2-oxoacid–dependent dioxygenases
usually occur with retention of configuration (36). Accordingly,the conversion of 3→8 and 2→9 would be expected to introduce
a hydroxy function with S configuration (Fig. 4). However, NMRanalysis of stabilized Fr9P product analogs 12 and 13 shows the Rconfiguration at C-1, suggesting that either the Fr9P-catalyzedhydroxylation occurred with inversion of configuration or a con-version occurred posthydroxylation. Mechanistic studies will benecessary to answer this question.Understanding spliceostatin biosynthesis is an important step
toward sustained compound production and analog generation.The analogs described here (7, 8, 9, 12, 13, and 14) helpedelucidate the biosynthetic route to hemiketal formation as wellas confirmed the positive contributions of the C-4 hydroxy andepoxide groups for cytotoxic potency. Because compounds 2 and3 are desirable drug leads, the dioxygenase mutant generated inthis study sets the stage for further strain development. Efforts toincrease production titers of 3 will be described elsewhere.
Materials and MethodsGenome Sequencing. Strain FERM BP-3421 was acquired from the In-ternational Patent Organism Depositary at the National Institute of Ad-vanced Industrial Science and Technology (Tsukuba, Japan). DNA for genomesequencing was isolated using Qiagen’s DNA Maxi Kit according to themanufacturer’s protocol. Specifications of the DNA submitted for sequenc-ing were Abs260/Abs280 = 1.90 and Abs260/Abs230 = 2.00. Sequencing wasperformed using 454 and Illumina technologies by 454 Life Sciences and theBeijing Genomics Institute (BGI), respectively. Assembly was performed byBGI. The 454 data were assembled with Newbler into 1,743 contigs and 1,732scaffolds (N50, 6,416), and the Illumina data were assembled with SOAPdenovointo 1,061 contigs and 84 scaffolds (N50, 447, 469). Assembly of both data-sets with SOAPdenovo resulted in 491 contigs and 84 scaffolds (N50, 447,523). The obtained genome sequence (7.68 Mbp, 67.5% G+C) was mined fora hybrid PKS–NRPS pathway that would be in agreement with spliceostatinbiosynthesis. The nucleotide sequence of the identified fr9 cluster
min10 11 12 13
8_t=0h
8_pH 7.5_t=2h8_pH 6_t=2h
8_cell-free lysate_pH 7.5_t=2h
8_pH 7.5_t=46h8_pH 6_t=46h
84*
84
A
B
C10 11
8_cell-free lysate_pH 7.5_t=2h
8_boiled cell-free lysate_pH 7.5_t=2hmin10 11 12 13
Fig. 6. Decarboxylation of β-hydroxy acid 8 to yield 4 is likely enzyme catalyzed in vivo. (A) 8 was obtained using recombinant dioxygenase Fr9P, and thenincubated at 25 °C, 200 rpm, under conditions indicated (pH 7.5 or 6.0 and with or without a cell-free lysate of module 2 mutant of PKS–NRPS gene fr9DEF).The mass of the degradation product marked with an asterisk corresponds to 8−H2O; this compound was not isolated. HPLC traces with detection at 230 nm.(B) Negative control using boiled cell-free lysate under the same conditions as in A. HPLC traces with detection at 230 nm. (C) Potential mechanisms forformation of hemiketal 4, furan 10, and aldehyde 11 (23, 45).
O
NH
OO OH
HO14
AcO H
Fig. 7. Spliceostatin hemiketal analog isolated from the FMO mutant.
E3382 | www.pnas.org/cgi/doi/10.1073/pnas.1408300111 Eustáquio et al.
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0

from FERM BP-3421 has been deposited to GenBank under accessionno. KJ461964.
Gene Inactivation in Burkholderia sp. FERM BP-3421. The FMO domain wasinactivated by in-frame deletion. All other mutants were generated byreplacing the targeted gene with a tetracycline-resistance marker. In-activation was performed by homologous recombination. Inactivationplasmids (see SI Materials and Methods for plasmid construction) based onpEX100T (48) or pEX18Tc (49) were transferred into FERM BP-3421 by con-jugation from E. coli S17.1, i.e., FERM BP-3421 and E. coli S17-1 containingthe plasmid for gene inactivation were separately cultivated in 10 mL ofLuria–Bertani (LB) medium at 30 °C, 200 rpm orbital shaking, overnight (withthe E. coli culture containing tetracycline, 10 μg/mL). Two milliliters of the E.coli culture was harvested by centrifugation, washed twice with 2 mL of LB(to remove antibiotics), and resuspended in 0.2 mL of fresh LB. Two millilitersof the Burkholderia culture was harvested by centrifugation and resus-pended in 0.2 mL of fresh LB. The two cell suspensions were carefully mixedand spread onto LB agar without antibiotics. Two negative control plates,one containing only E. coli and the other, only FERM BP-3421, treated asabove were prepared. Plates were incubated overnight at 30 °C. A “loopful”of cells from the conjugation plate was then streaked for single coloniesonto selection LB plates and incubated at 30 °C for 2 d. Single colonies werestreaked again on selection plates and incubated at 30 °C for 2 d. In the caseof gene replacement with a tetracycline marker, selection plates containedgentamycin (10 μg/mL, for removing E. coli after conjugation), tetracycline(25 μg/mL, for selection of mutants), and sucrose [5% (wt/vol), for counter-selection of the vector backbone]. Double-crossover mutants with the de-sired gene replacement were obtained directly. In the case of in-framedeletion (FMO domain), selection plates contained gentamycin (10 μg/mL)and tetracycline (25 μg/mL) to select for a single-crossover mutant, whichwas then streaked onto LB agar without supplements and incubated at 30 °Cfor 2 d. The single-crossover mutant was then passed three times through LBagar containing 5% (wt/vol) sucrose. Single colonies were replica plated onLB agar with or without tetracycline. Out of 38 tet-sensitive clones, only 2were confirmed as the desired double-crossover mutants (in-frame deletion),18 were reversion to wild type, and the others did not show any amplicon. Inall cases, mutants were confirmed by colony PCR (RED Taq; Sigma).
Gene Inactivation in Burkholderia sp. MSMB 43. Strain MSMB 43 was obtainedfrom the Centers for Disease Control and Prevention (CDC). MSMB 43 wasisolated by investigators at the Menzies School of Health Research, CharlesDarwin University (Casuarina, NT, Australia) (24) and is currently being de-scribed as “Burkholderia humptydooensis.” The inactivation plasmid (SIMaterials and Methods) was transferred into MSMB 43 by conjugation fromE. coli S17-1 as described above for FERM BP-3421. Selection plates containedgentamicin (10 μg/mL) and tetracycline (50 μg/mL) to select for a single-crossover mutant, which was then streaked onto LB agar without supple-ments and incubated at 37 °C for 2 d. The single-crossover mutant was thenpassed three times through LB agar without supplements. A total of 219single colonies was replica plated on LB agar with or without tetracycline(50 μg/mL). Six colonies were tetracycline sensitive; three were confirmed asthe desired double-crossover mutants (in-frame deletion), and three werereversion to wild type. All mutants were confirmed by colony PCR (REDTaq; Sigma).
Cultivation Conditions and Metabolite Analysis. Burkholderia sp. FERM BP-3421 and MSMB 43 were routinely cultured in LB broth or agar medium(Becton Dickinson) at 30 °C, unless otherwise stated. Frozen stocks wereprepared with 20% (vol/vol) glycerol and kept at −80 °C. For metaboliteanalyses, seed medium (10 g/L polypeptone, 5 g/L yeast extract, 5 g/L sodiumchloride) was inoculated with a frozen stock, and the culture was incubatedat 30 °C, 220 rpm overnight. This first seed culture was used to inoculatea second stage seed at 10% (vol/vol), which was incubated as above. Pro-duction medium (10 g/L soluble starch, 10 g/L glycerol, 5 g/L glucose, 10 g/LHySoy soypeptone, 5 g/L corn steep liquor, 2 g/L ammonium sulfate, 0.06 g/Lmagnesium sulfate heptahydrate, 2 g/L calcium carbonate) was inoculatedwith seed culture at 2.5% (vol/vol) and incubated at 25 °C, 200 rpm for 3 d.When 2S4G production medium (40 g/L glycerol, 20 g/L HySoy soypeptone,2 g/L ammonium sulfate, 0.1 g/L magnesium sulfate heptahydrate, 2 g/Lcalcium carbonate) was used, cultures were harvested at day 5. Seed cultureswere grown either in 14-mL, polypropylene round-bottom tubes (BD) con-taining 5 mL of medium or in 250-mL Erlenmeyer flasks containing 50 mL ofmedium. Tetracycline (25–50 μg/mL) was added where appropriate. Pro-duction cultures were grown either in 250-mL Erlenmeyer flasks containing50 mL of medium or in 2.8-L Fernbach flasks containing 0.5 L of medium. No
antibiotics were added to production cultures. The fermentation was pro-cessed with 5% (wt/vol) Diaion HP-20 resin as follows. Wet, activated HP-20was added to the culture at the end of fermentation, and the mixture wasshaken for 30 min on a tilting platform or shaker. After centrifugation, thesupernatant was decanted and the cell/resin pellet extracted twice with1/2 vol ethyl acetate. After removing solvent under reduced pressure,the crude extract was dissolved in DMSO to make a 2× sample for HPLCand LC/MS analyses. Alternatively, a 0.5-mL aliquot of the production culturewas centrifuged to remove cells, and the supernatant was mixed with 0.5 mLof acetonitrile. After centrifugation and filtering, the sample was analyzedby HPLC and/or LC/MS. HPLC analytic conditions were as follows: column,reversed-phase C18 YMC ODS-A, 4.6 × 150 mm, 5 μm; mobile phase A, 0.01%trifluoroacetic acid in water; mobile phase B, 0.01% trifluoroacetic acid inacetonitrile; gradient, 10–100% B over 19 min; flow rate, 1.0 mL/min; tem-perature, not controlled; detection, diode array detector (DAD) 230 nm;injection volume, 5–25 μL; instrument, Agilent 1100 series HPLC. LC/MSconditions were as follows: column, reversed-phase C8 Acquity UPLC BEH, 2.1mm × 100 mm, 1.7 μm; mobile phase A, 0.1% formic acid in water; mobilephase B, 0.1% formic acid in acetonitrile; gradient, 20% B for 0.5 min, 20–80% B for 1.5 min, 80–100% B for 3 min, 100% B for 5 min, 100–20% B for0.1 min, 20% B for 1.4 min; flow rate, 0.4 mL/min; instrument, WatersAcquity UPLC-LCT Premier TOF MS with alternating positive-ion and nega-tive-ion full scan (100–2,000 mass units) mode.
Production of Recombinant Fr9P Enzyme in and Purification from Escherichiacoli. The codon-optimized Fr9P gene was synthesized and ligated into theNcoI–HindIII sites of pGS-21a (GenScript) to generate pAE-PF16. Recombi-nant, His6-GST tagged Fr9P protein was produced in and purified from E. coliBL21(DE3) after transformation with plasmid pAE-PF16. A typical purifica-tion was as follows: two 2.8-L Fernbach flasks containing 0.5 L of medium(Terrific broth with 100 mg/L ampicillin) were each inoculated with 20 mLof an overnight LB culture and incubated at 200 rpm, 25 °C. When theOD600 reached ∼0.9, cells were induced with 0.2 mM isopropyl β-D-1-thiogalactopyranoside, and incubation was resumed at 25 °C and 200 rpm.After 18–20 h, cells were harvested by centrifugation and frozen at −80 °C.The cell pellet was resuspended in ∼50 mL of ice-cold lysis buffer (10 mMphosphate buffer, pH 7.4; 500 mM NaCl; 20 mM imidazole; 10% glycerol;lysozyme, 1 mg/mL; 0.5% Tween 20; 20 mM β-mercaptoethanol) and in-cubated on ice for 30 min. Following sonication on ice, the cell lysate wascentrifuged at 21,000 × g and 4 °C for 45 min. The supernatant was trans-ferred to a new tube and centrifuged again at 21,000 × g and 4 °C for30 min. Five milliliters of Ni-NTA resin slurry (Qiagen) were added to thesupernatant fraction (clear lysate) contained in a small beaker on ice andgently stirred for 1 h. The suspension was transferred to a Falcon tube andcentrifuged at 1,000 × g and 4 °C for 10 min. The supernatant was discardedand the resin washed three times, each with 30 mL of ice-cold wash buffer(10 mM phosphate buffer, pH 7.4; 500 mM NaCl; 40 mM imidazole; 10%glycerol; 20 mM β-ME) followed by centrifugation at 1,000 × g and 4 °C for10 min. The resin was transferred to a disposable column and washed threemore times, each with 2.5 mL of wash buffer. The enzyme was eluted with3× 2.5-mL elution buffer (10 mM phosphate buffer, pH 7.4; 500 mM NaCl;250 mM imidazole; 10% glycerol; 20 mM β-ME). The buffer was exchangedto 50 mM MOPS, pH 7.5, using a PD-10 column and the solution concen-trated using a Vivaspin column with molecular weight cutoff of 30 kDa.Storage buffer contained 50 mMMOPS, pH 7.5, 2 mM DTT, and 10% glycerol(for storage at −80 °C) or 50% glycerol (for storage at −20 °C). The yield ofpurified enzyme was ∼25 mg per liter of culture.
Fr9P Enzyme Assay and Determination of Michaelis–Menten Kinetic Constants.Fr9P assays were carried out in MOPS buffer (50 mM) at pH 7.5. Note thatphosphate and Bis-Tris buffers were found to inhibit enzyme activity (Fig.S4C). For assays presented in Fig. 4, the complete reaction contained 3 (0.1mM), α-ketoglutarate (0.2 mM), sodium ascorbate (0.02 mM), NH4Fe(II)SO4
(0.01 mM), and Fr9P (0.4–1.6 μM); reactions were incubated for 1–2 h atroom temperature (21 °C). For the reaction carried out under anaerobicconditions, 0.925 mL of MOPS buffer (contained in a capped glass vial) wasbubbled with nitrogen for 10 min at 37 °C. At the same time, a α-ketoglu-tarate stock solution (10 mM) was likewise bubbled with nitrogen for 10 minat 37 °C. After bringing the buffer to room temperature, reagents wereadded using a needle syringe in the following order: (i) a master mix ofsodium ascorbate, Fe(II) and 3, and (ii) Fr9P. The mixture was bubbled withnitrogen for an additional 10 min at room temperature before addingα-ketoglutarate to initiate the reaction under nitrogen atmosphere (finalvolume, 1 mL). For determination of kinetic constants, recombinant Fr9P(0.2 μM) was incubated with spliceostatin substrate (10–500 μM), α-ketoglutarate
Eustáquio et al. PNAS | Published online August 5, 2014 | E3383
BIOCH
EMISTR
YPN
ASPL
US
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0

(0.002–1 mM), sodium ascorbate (0.1 mM), and NH4Fe(II)SO4 (0.05 mM) in 50 mMMOPS buffer (pH 7.5) at 30 °C for 1 min. Reactions were then immediatelytransferred to ice, quenched with 1 vol of acetonitrile, filtered, and analyzed byHPLC as above (25-μL injections). Product area under the curve was used to cal-culate conversion (micromolar product per minute) using a 3 standard curve.Michaelis–Menten constants were calculated from Lineweaver–Burke plots.
Incubation of 8 with a Cell-Free Lysate of the NRPS− Mutant. The NRPS− mu-tant of FERM BP-3421 was grown in 50 mL of 2S4G spliceostatin productionmedium as described above. Cells were harvested after 3 d and frozen at−80 °C. The cell pellet was defrosted and resuspended in 5 mL of MOPSbuffer, pH 7.5. Cells were lyzed by sonication on ice. After centrifugation at21,000 × g for 20 min, the supernatant was transferred to a new tube on ice.8 was obtained using recombinant Fr9P. Twenty microliters of the cell-freelysate was added to 8 (0.1 mM) contained in 200 μL of Fr9P reaction mixtureand incubated at 25 °C and 200 rpm for 2 h. Boiled cell-free lysate was usedas negative control, in which case formation of 4 was not observed. Afterquenching with 1 vol of MeCN, centrifuging, and filtering, the reaction wasanalyzed by HPLC and LC/MS.
Isolation and Structure Elucidation of Hemiketal 14 from the ΔFMO Mutant.The ΔFMO mutant of FERM BP-3421 was cultured in 1 L of 2S4G productionmedium as described above. After removing cells by centrifugation (20 minat 4,500 × g), wet, activated HP-20 [5% (wt/vol)] was added to the super-natant, and the mixture was mixed for 30 min on an orbital shaker. Thecompound-bound resin was collected by centrifugation and extracted twicewith 0.5 L of ethyl acetate each. The dried, crude extract (0.5 g) was purifiedby preparative, reversed-phase HPLC (column: Phenomenex Luna C18, 150 ×21.2 mm, 5 μm; mobile phase A: 0.02% acetic acid in water; mobile phase B:
0.02% acetic acid in acetonitrile; gradient: 30% B for 1.5 min, 30–80% B over8.5 min, 80–100% B over 0.5 min, 100% B for 2 min, 100–30% B over 0.5 min;flow rate: 27 mL/min). The fraction with retention time of 7.1–7.6 min wascollected and freeze-dried to afford 14 (51% pure at 215 nm). This fractionwas submitted to a second round of reverse-phase HPLC purification (col-umn: Phenomenex Luna C18, 150 × 21.2 mm, 5 μm; mobile phase A: 0.1%formic acid in water; mobile phase B: 0.1% formic acid in acetonitrile; gra-dient: 33% B for 10 min, 33–100% B over 0.5 min, 100% B for 2 min, 100–33% B over 0.5 min; flow rate: 27 mL/min). The fraction with retention timeof 5.75–6.75 min was collected and freeze-dried to afford 14 (11.8 mg, 94%pure by UV at 215 nm) as a white powder. High-resolution electrosprayionization mass spectrometry, m/z 492.2961, error −0.1 mDa for C27H42NO7
[M+H]+. For 1D and 2D NMR dataset, see Table S1.Refer to SI Materials and Methods for further methods.
ACKNOWLEDGMENTS. We thank Jay E. Gee (CDC) and Bart J. Currie(Menzies School of Health Research) for providing strain MSMB 43; H. P.Schweizer (Colorado State University) for vectors pEX18Tc and pEX100T;Xidong Feng and Keiko Tabei (Groton Center of Chemistry, Pfizer) for high-resolution MS and MS/MS analyses; Melissa Wagenaar (Groton Center ofChemistry, Pfizer) for purification of 7 and 14; Kathleen A. Farley, Dennis P.Anderson, and Guoyun Bai (Groton Center of Chemistry, Pfizer) for NMRsupport; My-Hanh Lam and Frank Loganzo (Oncology, Pfizer) for cytotoxicitydata; Ellen Murphy for initial assembly of the 454 sequence; Brian Dougherty,Baohong Zhang, Paul Fracasso, and Wen He (Computational Sciences, Pfizer) forhelp with Illumina sequencing; Andrew Tomaras (Biotherapeutics, Pfizer) forsupportive discussions on Burkholderia genetics; and Haiyin He, Li-Ping Chang,Greg Steele, Min He, Chakrapani Subramanyam, Andreas Maderna, RussellDushin, Edmund Graziani, and many others for helpful discussions during thecourse of these studies.
1. Bonnal S, Vigevani L, Valcárcel J (2012) The spliceosome as a target of novel anti-
tumour drugs. Nat Rev Drug Discov 11(11):847–859.2. Butler MS (2013) Remediating cancer via splicing modulation. J Med Chem 56(17):
6573–6575.3. Kaida D, Schneider-Poetsch T, Yoshida M (2012) Splicing in oncogenesis and tumor
suppression. Cancer Sci 103(9):1611–1616.4. Mizui Y, et al. (2004) Pladienolides, new substances from culture of Streptomyces
platensis Mer-11107. III. In vitro and in vivo antitumor activities. J Antibiot (Tokyo)
57(3):188–196.5. Sakai T, Asai N, Okuda A, Kawamura N, Mizui Y (2004) Pladienolides, new substances
from culture of Streptomyces platensisMer-11107. II. Physico-chemical properties and
structure elucidation. J Antibiot (Tokyo) 57(3):180–187.6. Seki-Asano M, et al. (1994) Isolation and characterization of a new 12-membered
macrolide FD-895. J Antibiot (Tokyo) 47(12):1395–1401.7. Nakajima H, et al. (1996) New antitumor substances, FR901463, FR901464 and
FR901465. II. Activities against experimental tumors in mice and mechanism of action.
J Antibiot (Tokyo) 49(12):1204–1211.8. Nakajima H, Takase S, Terano H, Tanaka H (1997) New antitumor substances,
FR901463, FR901464 and FR901465. III. Structures of FR901463, FR901464 and
FR901465. J Antibiot (Tokyo) 50(1):96–99.9. Miller-Wideman M, et al. (1992) Herboxidiene, a new herbicidal substance from
Streptomyces chromofuscus A7847. Taxonomy, fermentation, isolation, physico-
chemical and biological properties. J Antibiot (Tokyo) 45(6):914–921.10. Kotake Y, et al. (2007) Splicing factor SF3b as a target of the antitumor natural
product pladienolide. Nat Chem Biol 3(9):570–575.11. Kaida D, et al. (2007) Spliceostatin A targets SF3b and inhibits both splicing and nu-
clear retention of pre-mRNA. Nat Chem Biol 3(9):576–583.12. Hasegawa M, et al. (2011) Identification of SAP155 as the target of GEX1A (Her-
boxidiene), an antitumor natural product. ACS Chem Biol 6(3):229–233.13. Eskens FA, et al. (2013) Phase I pharmacokinetic and pharmacodynamic study of the
first-in-class spliceosome inhibitor E7107 in patients with advanced solid tumors. Clin
Cancer Res 19(22):6296–6304.14. Ghosh AK, Chen ZH (2013) Enantioselective syntheses of FR901464 and spliceostatin
A: Potent inhibitors of spliceosome. Org Lett 15(19):5088–5091.15. Kumar VP, Chandrasekhar S (2013) Enantioselective synthesis of pladienolide B and
truncated analogues as new anticancer agents. Org Lett 15(14):3610–3613.16. Lagisetti C, et al. (2014) Pre-mRNA splicing-modulatory pharmacophores: The total
synthesis of herboxidiene, a pladienolide-herboxidiene hybrid analog and related
derivatives. ACS Chem Biol 9(3):643–648.17. Machida K, et al. (2008) Organization of the biosynthetic gene cluster for the poly-
ketide antitumor macrolide, pladienolide, in Streptomyces platensisMer-11107. Biosci
Biotechnol Biochem 72(11):2946–2952.18. Shao L, Zi J, Zeng J, Zhan J (2012) Identification of the herboxidiene biosynthetic gene
cluster in Streptomyces chromofuscus ATCC 49982. Appl Environ Microbiol 78(6):
2034–2038.19. Zhang F, et al. (2011) Cloning and elucidation of the FR901464 gene cluster revealing
a complex acyltransferase-less polyketide synthase using glycerate as starter units.
J Am Chem Soc 133(8):2452–2462.
20. Liu X, et al. (2013) Genomics-guided discovery of thailanstatins A, B, and C as pre-mRNA splicing inhibitors and antiproliferative agents from Burkholderia thai-landensis MSMB43. J Nat Prod 76(4):685–693.
21. Nakajima H, et al. (1996) New antitumor substances, FR901463, FR901464 andFR901465. I. Taxonomy, fermentation, isolation, physico-chemical properties and bi-ological activities. J Antibiot (Tokyo) 49(12):1196–1203.
22. He H, et al. (2014) Cytotoxic spliceostatins from Burkholderia sp. and their semi-synthetic analogues. J Nat Prod, in press.
23. Motoyoshi H, et al. (2004) Structure-activity relationship for FR901464: A versatilemethod for the conversion and preparation of biologically active biotinylated probes.Biosci Biotechnol Biochem 68(10):2178–2182.
24. Gee JE, et al. (2008) Recovery of a Burkholderia thailandensis-like isolate from anAustralian water source. BMC Microbiol 8:54–58.
25. Liu X, Biswas S, Tang GL, Cheng YQ (2013) Isolation and characterization of spli-ceostatin B, a new analogue of FR901464, from Pseudomonas sp. No. 2663. J Antibiot(Tokyo) 66(9):555–558.
26. Tang MC, He HY, Zhang F, Tang GL (2013) Baeyer-Villiger oxidation of acyl carrierprotein-tethered thioester to acyl carrier protein-linked thiocarbonate catalyzed bya monooxygenase domain in FR901464 biosynthesis. ACS Catal 3(3):444–447.
27. Piel J (2002) A polyketide synthase-peptide synthetase gene cluster from an un-cultured bacterial symbiont of Paederus beetles. Proc Natl Acad Sci USA 99(22):14002–14007.
28. Piel J (2010) Biosynthesis of polyketides by trans-AT polyketide synthases. Nat ProdRep 27(7):996–1047.
29. Pöplau P, Frank S, Morinaka BI, Piel J (2013) An enzymatic domain for the formationof cyclic ethers in complex polyketides. Angew Chem Int Ed Engl 52(50):13215–13218.
30. HeHY, TangMC, Zhang F, TangGL (2014) cis-Double bond formation by thioesterase andtransfer by ketosynthase in FR901464 biosynthesis. J Am Chem Soc 136(12):4488–4491.
31. Jenner M, et al. (2013) Substrate specificity in ketosynthase domains from trans-ATpolyketide synthases. Angew Chem Int Ed Engl 52(4):1143–1147.
32. Khosla C, Kapur S, Cane DE (2009) Revisiting the modularity of modular polyketidesynthases. Curr Opin Chem Biol 13(2):135–143.
33. Meunier B, de Visser SP, Shaik S (2004) Mechanism of oxidation reactions catalyzedby cytochrome p450 enzymes. Chem Rev 104(9):3947–3980.
34. Kudo F, Motegi A, Mizoue K, Eguchi T (2010) Cloning and characterization of thebiosynthetic gene cluster of 16-membered macrolide antibiotic FD-891: Involvementof a dual functional cytochrome P450 monooxygenase catalyzing epoxidation andhydroxylation. ChemBioChem 11(11):1574–1582.
35. Anzai Y, et al. (2008) Functional analysis of MycCI and MycG, cytochrome P450 en-zymes involved in biosynthesis of mycinamicin macrolide antibiotics. Chem Biol 15(9):950–959.
36. Prescott AG, Lloyd MD (2000) The iron(II) and 2-oxoacid-dependent dioxygenases andtheir role in metabolism. Nat Prod Rep 17(4):367–383.
37. Eichhorn E, van der Ploeg JR, Kertesz MA, Leisinger T (1997) Characterization ofα-ketoglutarate-dependent taurine dioxygenase from Escherichia coli. J Biol Chem272(37):23031–23036.
38. Zhang Z, et al. (2000) Structural origins of the selectivity of the trifunctional oxy-genase clavaminic acid synthase. Nat Struct Biol 7(2):127–133.
39. Walsh CT, Wencewicz TA (2013) Flavoenzymes: Versatile catalysts in biosyntheticpathways. Nat Prod Rep 30(1):175–200.
E3384 | www.pnas.org/cgi/doi/10.1073/pnas.1408300111 Eustáquio et al.
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0

40. Du L, Lou L (2010) PKS and NRPS release mechanisms. Nat Prod Rep 27(2):255–278.41. Gibson M, Nur-e-alam M, Lipata F, Oliveira MA, Rohr J (2005) Characterization of
kinetics and products of the Baeyer-Villiger oxygenase MtmOIV, the key enzyme ofthe biosynthetic pathway toward the natural product anticancer drug mithramycinfrom Streptomyces argillaceus. J Am Chem Soc 127(50):17594–17595.
42. Seo MJ, Zhu D, Endo S, Ikeda H, Cane DE (2011) Genome mining in Streptomyces.Elucidation of the role of Baeyer-Villiger monooxygenases and non-heme iron-dependent dehydrogenase/oxygenases in the final steps of the biosynthesis of pen-talenolactone and neopentalenolactone. Biochemistry 50(10):1739–1754.
43. Tibrewal N, et al. (2012) Baeyer-Villiger C-C bond cleavage reaction in gilvocarcin andjadomycin biosynthesis. J Am Chem Soc 134(44):18181–18184.
44. Thompson CF, Jamison TF, Jacobsen EN (2001) FR901464: Total synthesis, proofof structure, and evaluation of synthetic analogues. J Am Chem Soc 123(41):9974–9983.
45. Albert BJ, Sivaramakrishnan A, Naka T, Czaicki NL, Koide K (2007) Total syntheses,fragmentation studies, and antitumor/antiproliferative activities of FR901464 and itslow picomolar analogue. J Am Chem Soc 129(9):2648–2659.
46. Effenberger KA, et al. (2014) Coherence between cellular responses and in vitrosplicing inhibition for the anti-tumor drug pladienolide B and its analogs. J Biol Chem289(4):1938–1947.
47. Villa R, et al. (2013) Stabilized cyclopropane analogs of the splicing inhibitor FD-895.J Med Chem 56(17):6576–6582.
48. Schweizer HP, Hoang TT (1995) An improved system for gene replacement and xylEfusion analysis in Pseudomonas aeruginosa. Gene 158(1):15–22.
49. Hoang TT, Karkhoff-Schweizer RR, Kutchma AJ, Schweizer HP (1998) A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonasaeruginosa mutants. Gene 212(1):77–86.
Eustáquio et al. PNAS | Published online August 5, 2014 | E3385
BIOCH
EMISTR
YPN
ASPL
US
Dow
nloa
ded
by g
uest
on
Apr
il 24
, 202
0