Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 2005, p. 3139–3150 Vol. 187, No. 90021-9193/05/$08.00�0 doi:10.1128/JB.187.9.3139–3150.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Staphylococcus intermedius Produces a Functional agr AutoinducingPeptide Containing a Cyclic Lactone
Guangyong Ji,1* Wuhong Pei,1 Linsheng Zhang,1† Rongde Qiu,1 Jianqun Lin,1 Yvonne Benito,2Gerard Lina,2 and Richard P. Novick3*
Department of Microbiology and Immunology, Uniformed Services University of the Health Sciences, 4301 Jones Bridge Rd.,Bethesda, Maryland 208141; Centre National de Reference des Staphylocoques, INSERM E0230,
IFR62 Laennec, 7 rue Guillaume Paradin, 69372 Lyon Cedex 08, France2; andMolecular Pathogenesis Program, Skirball Institute of Biomolecular Medicine,
New York University Medical Center, 540 First Ave.,New York, New York 100163
Received 31 July 2004/Accepted 19 January 2005
The agr system is a global regulator of accessory functions in staphylococci, including genes encodingexoproteins involved in virulence. The agr locus contains a two-component signal transduction module that isactivated by an autoinducing peptide (AIP) encoded within the agr locus and is conserved throughout thegenus. The AIP has an unusual partially cyclic structure that is essential for function and that, in all but onecase, involves an internal thiolactone bond between a conserved cysteine and the C-terminal carboxyl group.The exceptional case is a strain of Staphylococcus intermedius that has a serine in place of the conservedcysteine. We demonstrate here that the S. intermedius AIP is processed by the S. intermedius AgrB protein togenerate a cyclic lactone, that it is an autoinducer as well as a cross-inhibitor, and that all of five other S.intermedius strains examined also produce serine-containing AIPs.
The agr locus, a global regulator of genes involved in patho-genesis and other accessory functions (27), is widely conservedamong staphylococci (2) and has homologs in other species(11). It consists of two divergent operons, driven by promotersP2 and P3, respectively (6, 22, 24, 26). The P2 operon containsfour genes: agrA, -B, -C, and -D. AgrA and -C comprise atwo-component signaling module, of which AgrC is the recep-tor and AgrA the response regulator (22), which, when acti-vated, upregulates the transcription of both the P2 and the P3operons. The P3 transcript, RNAIII, encodes delta-hemolysinand is the effector of the agr response (5, 24). The AgrC ligandis an autoinducing peptide (AIP) (9, 14, 15) that is proteolyti-cally processed by AgrB from a propeptide encoded by agrDand probably also secreted by AgrB (9, 37). Although struc-turally conserved, agrB, -D, and -C have diverged widely amongstaphylococci, giving rise to multiple specificity groups, inwhich heterologous AIP-receptor interactions are inhibitory.The mature AIPs are seven to nine amino acids long and havevariable amino acid sequences. All of the seven native staph-ylococcal AIPs thus far analyzed, including the four knownStaphylococcus aureus variants plus strains of Staphylococcuswarneri, Staphylococcus epidermidis, and Staphylococcus lug-dunensis (7, 8, 23, 25), contain a five-amino-acid thiolactone
ring, essential for function, in which a conserved cysteine isattached to the C-terminal carboxyl by a thioester linkage.AgrD has been sequenced for ca. 20 other non-S. aureus staph-ylococci, and all but one are predicted to contain a cysteine atthe same position (2), suggesting that the mature AIPs wouldeach contain the same thiolactone ring (8, 9, 16–18, 25). Theexception is a strain of Staphylococcus intermedius that is pre-dicted to contain a serine in place of the usual cysteine (2, 20).AIPs with a serine replacing cysteine and therefore having alactone rather than a thiolactone ring have been synthesized invitro. These do not activate the cognate receptor, and theyinhibit agr activation in heterologous combinations (17, 18).This raises three questions. (i) Do other S. intermedius strainsproduce serine-containing AIPs and do they produce an AIP,or is the serine the result of a missense mutation resulting in aninactive agrD gene? (ii) If a lactone AIP is produced, is it afunctional autoinducer? (iii) Finally, can an S. aureus AgrBthat normally processes a cysteine-containing pro-AIP alsoprocess a serine-containing pro-AIP and vice versa?
We have analyzed here the serine-producing S. intermediusCCM5739 plus five other S. intermedius strains and have foundthat all have a serine in the conserved position in AgrD andthat culture supernatants of each of the six S. intermediusstrains inhibit agr activation in S. aureus, suggesting that eachproduces an AIP. We determined the specificity and the in-volvement of S. intermedius AgrB (AgrB-Si) in the processingof S. intermedius AgrD (AgrD-Si) to produce the mature lac-tone AIP. For strain CCM5739, tandem mass spectroscopy hasconfirmed that the AIP contains a lactone ring (10), and weshow that this AIP and also that produced by ATCC 29663 areself-activators and that all six are cross-inhibitors. In addition,an S. aureus agr group I strain with a mutation in agrD thatwould replace the conserved cysteine with a serine does not
* Corresponding author. Mailing address for R. P. Novick: Depart-ments of Microbiology and Medicine, Molecular Pathogenesis Pro-gram, Skirball Institute of Biomolecular Medicine, New York Univer-sity Medical Center, 540 First Ave., New York, NY 10016. Phone:(212) 263-6290. Fax: (212) 263-5711. E-mail: [email protected]. Mailing address for G. Ji: Department of Microbiology andImmunology, Uniformed Services University of the Health Sciences,4301 Jones Bridge Rd., Bethesda, MD 20814. Phone: (301) 295-9621.Fax: (301) 295-1545. E-mail: [email protected].
† Present address: Division of Pediatric Oncology, The Johns Hop-kins University, 1650 Orleans St., Baltimore, MD 21231.
3139
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

produce any detectable AIP, whereas an S. intermedius strainwith cysteine replacing the AIP serine produces a mature thio-lactone AIP that retains (weak) self-activation, as well asstrong cross-inhibition activity.
MATERIALS AND METHODS
Bacterial strains and growth conditions. Staphylococcal strains and plasmidsused in the present study are listed in Table 1. Escherichia coli strains JM109 andDH5� were used for cloning.
S. aureus cells were grown in CYGP broth (21) with shaking at 37°C. Super-natants of cultures in late exponential phase or post-exponential phase werecentrifuged, filter sterilized, and used as a source of group-specific AIPs. Gen-erally, a 1/10 volume of such supernatants contained sufficient AIP for fullactivation or inhibition. Overnight cultures on GL plates (21) were routinelyused as inocula. Cell growth was monitored with a Klett-Summerson colorimeterwith a green (540-nm) filter (Klett, Long Island City, NY). Antibiotics used forplasmid maintenance were erythromycin (10 �g/ml) and tetracycline (5 �g/ml).
Transformation of S. intermedius. Attempts to directly transform S. interme-dius with DNA isolated from S. aureus by either electroporation (30) or proto-plast transformation (21) failed. S. intermedius was successfully transformed byfirst amplifying the intact plasmid by using PCR, followed by ligation of the PCR
products. This material could then be used for protoplast transformation (21).The success of this method depends on the use of wholly PCR-generated DNA,which seems to be taken up across species barriers, possibly owing to lack ofmethylation and/or other modifications.
Nucleotide sequencing. The agr locus of S. intermedius ATCC 29663 wassequenced by two methods. First, a PCR product was made with oligonucleotidesWP4 and WP6 as primers (oligonucleotides used in the present study are listedin Table 2), and chromosomal DNA prepared from S. intermedius ATCC 29663as a template. The PCR product was directly sequenced. The DNA sequence wasused to design primers for the second sequencing method as follows: chromo-somal DNA was digested with both HindIII and EcoRI enzymes. The resultingDNA fragments were separated on agarose gels, followed by Southern blothybridization with either PCR probe A generated with oligonucleotides WP4 andWP11 as primers and chromosomal DNA as a template or PCR probe B am-plified from chromosomal DNA with primers WP14 and WP6. Southern blothybridization was performed accordingly (28). Two chromosomal DNA frag-ments (ca. 1.7 and 2.0 kb) were detected by using probe A and probe B,respectively. DNA fragments within either 1.7- or 2.0-kb regions were excisedfrom agarose gels, purified, and then ligated into the HindIII and EcoRI sites ofan E. coli cloning vector, pGEM-3zf (Promega). PCR products were then madeby using oligonucleotides WP7 and T7 promoter primer (located on the vector)as primers and the ligation mixtures (pGEM-3zf plus 1.7-kb DNA fragment) as
TABLE 1. Staphylococcal strains and plasmids used in this study
Plasmid or strain Genotype and descriptiona Source or reference
PlasmidspRN5543 Cloning vector 24pRN5548 Vector carrying a staphylococcal inducible Pbla promoter 24pRN6441 Cloning vector 22pRN6683 S. aureus group I agr P3-blaZ fusion 22pRN7062 Shuttle plasmid, AIP-I tester 14pRN7105 Shuttle plasmid, AIP-II tester 14pRN7106 Shuttle plasmid, AIP-III tester 14pRN7128 Shuttle plasmid, AIP-IV tester 14pGJ2001 S. aureus agrB-I in pRN5548 37pGJ2002 S. aureus agrB-II in pRN5548 38pGJ4002 S. aureus agrD-I in pRN6441 38pLZ4001 S. aureus agrD-II in pRN6441 38pLZ4003 S. aureus agrD-III in pRN6441 This studypLZ4005 agrD-I with double His6 tags in pRN6441 (HDH) 37pLZ4006 agrD-I with C27S mutation and double His6 tags in pRN6441
(HDHcs)This study
pWP1004 S. intermedius promoter P3-blaZ fusion in pRN5543 This studypWP1101 S. intermedius agrB-Si in pRN5548 This studypWP1102 agrB-Si and S. intermedius agrD-Si in pRN5548 This studypWP1103 T7 epitope-agrD-Si-His6 (TDsiH) in pRN5548 This studypWP1104 agrB-Si and TDsiH in pRN5548 This studypWP1105 agrD-Si mutation (S27C) in pWP1104 This studypWP1106 agrD-Si mutation (S27A) in pWP1104 This study
StrainsRN4220 Mutagenized derivative of S. aureus that readily accepts foreign DNA 21RN6390B S. aureus group I agr� laboratory strain 24RN6734 �13 lysogen of RN6390 This studyRN6911 RN6390B �agr 1057–4546::tetM 24RN7206 �13 lysogen of RN6911 This studyRN8463 S. aureus group III agr� strain 8SA502A S. aureus group II agr� strain 8GJ2035 RN6911(pI524) 37ATCC 29663 S. intermedius strain American Type Culture CollectionCCM5739 S. intermedius strain from the Czechoslovak Collection of
Microorganisms2
RN9515 S. intermedius strain Novick labRN9161 S. intermedius strain W. KloosRN9167 S. intermedius strain W. KloosRN9169 S. intermedius strain W. Kloos
a agr groups, including individual genes and products, are indicated by roman numerals. Thus, agr-II means the agr locus from a group II S. aureus strain; AIP-IIImeans the AIP produced by agr group III strains; AgrB-I refers to the AgrB protein of S. aureus agr group I, etc. The suffix or subscript “Si” is used to indicate thatS. intermedius is the source of the gene or gene product.
3140 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

a template or by using oligonucleotides WP8 and T7 promoter primer as primersand the ligation mixtures (pGEM-3zf plus 2.0-kb DNA fragment) as templates.Both PCR products were used for DNA sequencing. We note that all PCRproducts were amplified with PfuTurbo high-fidelity DNA polymerase (Strat-agene), and the PCR products from three independent PCRs were generatedand sequenced to confirm the lack of PCR errors. Four other S. intermedius agrDgenes were sequenced by using oligonucleotides SiB-F3 and SiC-R5, from con-served positions in agrB and agrC, respectively, and rnaIII in S. intermediusCCM5739 was sequenced by using primers SaR3-F1 and SiB-R3.
agr activation and inhibition reporter gene assays. Two methods were used forthese assays, (i) Assays were performed with bacterial cultures in early exponen-tial phase (�2 � 108 cells/ml) by using a �-lactamase reporter gene readout. Foractivation assays, 80 �l of cells (9 � 108 cells/ml) were treated in duplicate withvarious amounts of culture supernatant containing AIP at ca. 60 nM, as esti-mated by comparing its activity with that of a sample of synthetic AIP of knownconcentration, and incubated with shaking at 37°C for 60 or 90 min in aTHERMOmax microplate reader (Molecular Devices) with monitoring of celldensity at an optical density at 650 nm, followed by determination of agr activa-tion by the �-lactamase–nitrocefin assay (9). Nitrocefin was purchased fromBecton Dickinson (Franklin Lakes, NJ). For inhibition assays, 80 �l of cells (9 �108 cells/ml) was treated in duplicate with AIP-containing culture supernatants inthe presence of the group-specific wild-type AIP agonist at a concentration of100 nM for 90 min, followed by a �-lactamase assay. An agonist concentration of100 nM is a saturating but not oversaturating dose of activator, which generatesmaximal activation against which to test various concentrations of heterologousAIPs. (ii) Assays with S. aureus reporter cells containing plasmid pRN6683 (22)with the S. aureus P3-blaZ fusion were done according to the method describedpreviously (8, 9). The same method was used for the AIP activity assays with S.intermedius reporter cells harboring plasmid pWP1004 (S. intermedius P3-blaZ)(Table 1). In brief, culture supernatants were prepared from either S. intermediuswild-type cells grown in CYGP media at 37°C for 6 h or S. aureus cells containingthe cloned S. intermedius agr gene(s) under the control of Pbla promoter grownat 37°C to 70 Klett units, followed by induction with 0.5 mM methicillin at 37°Cfor 5 h. The supernatants were filtered with 0.22-�m-pore-size filters. To 45 �l ofculture of S. intermedius ATCC 29663(pWP1004) (�40 and �100 Klett units for
activation and inhibition assays, respectively), 5 �l of supernatant prepared (ordiluted in CYGP medium) was added, and the mixtures were incubated at 37°Cfor 55 min (activation) or 80 min (inhibition) in a VERSAmax microplate reader(Molecular Devices). Diluted cultures (10% in CYGP plus 5 nM sodium azide)were mixed with equal volumes of nitrocefin solution, and the �-lactamaseactivities were measured as described previously (8, 9).
Agr induction by S. intermedius culture supernatants. The stationary-phasefiltered bacterial supernatant from an overnight 25-ml culture of S. intermediusstrain CCM5739 was lyophilized. Lyophilized brain heart infusion broth was usedas a control. From 50 ml of a 1-h culture, 25 ml was poured into a flask containingthe lyophilized supernatant and 25 ml was poured into another flask containingthe lyophilized brain heart infusion. Then, 1-ml aliquots were drawn at 0, 30, 60,and 90 min, and the cell densities were measured. The 1-ml samples were usedto prepare RNA for Northern blot hybridization. Equal masses of cells were usedfor each time point.
Plasmid constructions. Plasmid pWP1004 was constructed as follows. Theputative P2/P3 promoter region of S. intermedius was amplified by PCR usingoligonucleotides WP12 and SINT11 as primers and chromosomal DNA as atemplate. The PCR product was cloned into E. coli cloning vector pGEM-T(Promega) in both orientations. The resulting plasmid pGEM-T-P2P3 was di-gested with EcoRI and SpeI, and the DNA fragment containing the P2/P3promoter region was then cloned into the EcoRI/NheI sites of pRN5543, fol-lowed by insertion of an EcoRI fragment from pRN6683 containing the staph-ylococcal blaZ gene into the EcoRI site of the resulting plasmid.
Plasmid pWP1101 was constructed by cloning a PCR product amplified fromthe chromosomal DNA using primers SINT1 and SINT5 into the XbaI site ofpRN5548. Plasmid pWP1102 was made by cloning a PCR product amplified fromthe chromosomal DNA using primers SINT1 and SINT2 into the XbaI/EcoRIsites of pRN5548. To construct pWP1103, a PCR product was prepared by usingoligonucleotides SINT3 and SINT4 as primers and S. intermedius chromosomalDNA as a template. The EcoRI/BspHI-digested PCR product was then clonedinto the EcoRI/BspHI sites of pLZ4012 (38). Plasmid pWP1104 was made bycloning a PCR product amplified from chromosomal DNA by using primersSINT1 and SINT5 into the XbaI site of pWP1103.
Site-directed mutagenesis of the agrD-Si gene was done by using PCR primers
TABLE 2. Oligonucleotides used in this study
Primer Nucleotide sequence (533)a Locationb
WP4 CATCAAATATGCACCAGCAGC agrB-Si (118–138)�WP6 CATAATCATGACGGAACTTGCGCA agrC-I (3201–3224)†WP7 GTATGGTGGTGGTACGATGCATGC agrB-Si (796–809)‡WP8 CGCAACGACTTTAACGATTGAACGTG agrC-Si (1246–1271)‡WP11 GAAAAGGTAACAACCTAGCGC agrC-Si (1416–1436)‡WP12 CCATCACCAATGTGATGATG P3-Si (109–129)‡WP14 TAGCGTGCGGGTTCGTCA agrC-Si (1525–1542)‡SINT1 GCTCTAGAAGGAGGAGAGCCGT SD of agrB-Si (317–330)‡ (XbaI)SINT2 GCGAATTCACACTTTCTAACTTT agrD-Si (1036–1050)‡ (EcoRI)SINT3 GCGAATTCACATGAGAATTTTAGAAG agrD-Si (897–914)‡ (EcoRI)SINT4 CGATTCATGATTAATGATGATGATGATGATGTTTTTCCTCTTCTAA
CAACTCAGCagrD-Si (1010–1033)‡ (His6 � 1 stop codon)
(BspHI)SINT5 GCTCTAGATGTTAAACTTCCTCC agrB-Si (885–899)‡ (XbaI)SINT6 GCTAGAATCCCAACGTGTACTGGATTTTTCG agrD-Si (962–992)‡ (AGT3TGT)SINT8 AAAAGTACCGATTGATTGG agrD-Si (943–961)‡SINT11 CGGCTCTCCTCCTTGTTT P2-Si (312–329)‡SINT15 GCTAGAATCCCAACGGCTACTGGATTTTTCG agrD-Si (962–992)‡ (AGT3TGT)GJ#28 CTAATGATGATGATGATGATGTTCGTGTAATTGTGTAATTC agrD-I (2459–2479)† (His6 � 1 stop codon)GJ#45 GTAAATGAAGTCCATGGAATAATAG Sequence around the NcoI site of pRN5548 (NcoI)GJ#56 GCTCTAGAAGCTATTACATTATTACC Before the SD of agrD-I (2297–2314)† (XbaI)CS-1 GCTTATAGTACTAGTGACTTC agrD-I (2411–2431)† (TGT3AGT)SiB-F3 TATGCACCAGCAGCTACGAAAAAAAG agrB-Si (125–150)�SiC-R5 TTTCCAATTTTATACACAC agrC-Si (475–493)�SaR3-F1 GGCCGCGAGCTTGGGAGGG agr-I rnaIII (1059–1077)†SiB-R3 CTTTTTTTCGTAGCTGCTGGTGCATA Complementary to SiB-F3RNAIII-F ACTGAGTCCAAGGAAACTAACT agr-I rnaIII (1259–1280)†RNAIII-R TAGATCACAGAGAT agr-I rnaIII (1558–1571)†T7 primer TAATACGACTCACTATAGGG T7 promoter sequencing primer (Promega)
a Added enzyme sites are underlined. His6 plus 1 stop codon are indicated in italics.b �, Nucleotide numbering of S. intermedius CCM5739 (GenBank accession no. AF346723); †, nucleotide numbering of S. aureus agr-I (GenBank accession no.
X52543); ‡, nucleotide numbering of S. intermedius ATCC 29663 agr (GenBank accession no. AY557375).
VOL. 187, 2005 S. INTERMEDIUS agr AUTOINDUCING PEPTIDE 3141
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

containing desired mutations and the ExSite PCR-based site-directed mutagen-esis method according to the manufacturer’s instruction (Stratagene). PCR prod-ucts were generated by using T4 polynucleotide kinase (MBI Fermentas) phos-phorylated oligonucleotides SINT6 and SINT8, or SINT15 and SINT8, asprimers and pWP1104 as a template. The PCR products were purified fromagarose gels and then ligated, resulting in plasmids pWP1105 and pWP1106,respectively. pLZ4003 carrying the S. aureus group III agrD gene was constructedby cloning a ClaI fragment of pRN6963 (8) into the ClaI site of pRN6441.
Plasmid pLZ4006 that carries the S. aureus group I agrD (agrD-I) gene with thecysteine codon in the AIP coding region changed to a serine codon and with sixhistidine codons fused at both its 3 and 5 ends was constructed as follows. APCR product was generated by using primers GJ#56 and GJ#28, with pRN6852(9) as a template. The PCR product was then digested with XbaI and cloned intopRN5548 XbaI and EcoRI (blunted with Klenow) sites. The resulting plasmidwas then used as a template to generate a PCR product using oligonucleotidesGJ#45 and CS-1 as primers. The PCR products were digested with ScaI andNcoI and cloned into the ScaI/NcoI sites of pGJ4004 (37).
All PCR products used for plasmid construction were amplified with PfuTurbohigh-fidelity DNA polymerase (Stratagene). The DNA sequences of the clonedwild-type and mutated genes in the constructed plasmids were confirmed.
Whole-cell lysates and Northern blotting for RNAIII expression. Whole-celllysates were prepared as described previously (13). Cultures were centrifuged,fixed in acetone-ethanol (1:1), and washed in N-tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid (TES)–sucrose buffer. Equalized cell samples were
incubated on ice for 30 min with lysostaphin (150 �g/ml) in TES-sucrose buffer(20% sucrose, 20 mM Tris [pH 7.6], 10 mM EDTA, 50 mM NaCl) and shakenfor 1 h with proteinase K (50 �g/ml; Sigma) and 2% sodium dodecyl sulfate(SDS) at 4°C. For Northern blotting, the same amount of cell lysate was elec-trophoresed through a 0.66 M formaldehyde–1% agarose gel in morpholinepro-panesulfonic acid buffer (28). Nucleic acids were transferred to a nitrocellulosemembrane (Amersham) with a VacuGene apparatus (Pharmacia) in 20� SSC (3M NaCl plus 0.3 M sodium citrate [pH 7]) and fixed under UV light. Themembrane was preincubated for 2 h at 52°C in 2� Denhardt solution (0.02%bovine serum albumin, 0.02% Ficoll, 0.02% polyvinylpyrrolidone, 0.05 M EDTA[pH 8], 0.2% SDS, and 5� SSC, with sonicated and heat-denatured salmonsperm DNA at 100 �g/ml) and 10% dextran (Sigma) and then hybridized over-night with a 32P-labeled DNA probe in hybridization solution. 32P-labeledRNAIII probes were prepared by PCR using oligonucleotide primers RNAIII-Fand RNAIII-R, and pRN6735 DNA as a template. In labeling reactions, dATPconcentration was reduced to 2 �M, and the reaction mixture contained 50 �Ciof [�-32P]dATP (Amersham; 1 Ci 37 GBq). The blot was exposed to a storagephosphor screen (Molecular Dynamics).
Western blot hybridization. S. aureus cells expressing various genes under thecontrol of Pbla were grown and induced with 0.5 mM methicillin for 4 h at 37°C.The cultures were then mixed with an equal volume of ice-cold solution (50%acetone and 50% ethanol). The fixed cells were washed with 1� SMM (21) andsuspended in 1� SMM plus lysostaphin (Sigma). After 1 h of incubation at 37°C,cells were lysed by sonication. Whole-cell lysates were separated by 16% poly-
FIG. 1. Comparison of the AgrD amino acid sequences from S. aureus and S. intermedius. The amino acid sequences were analyzed by theClone Manager software (Sci-Ed Software, Durham, NC). Dash lines represent gaps generated by the analysis program. The identical amino acidresidues are dark shaded, and the AIP sequences in AgrDs are in boldface. The amino acid sequences of AgrD proteins can be accessed throughthe NCBI protein database under NCBI accession no. CAA36782 (S. aureus group I, Sa I) (22), AAB63265 (S. aureus group II, Sa II) (8),AAB63268 (S. aureus group III, Sa III) (8), AAG03056 (S. aureus group IV, Sa IV) (7), AY871105 (RN9161), AY871106 (RN9167), AY871107(RN9169), AAL65836 (CCM5739) (2), AF346723 (RN9515), and AAS66746 (ATCC 29663). The amino acid sequences of AgrD proteins can beaccessed through the NCBI protein database under NCBI accession no. CAA36782 (S. aureus group I, Sa I) (22), AAB63265 (S. aureus group II,Sa II) (8), AAB63268 (S. aureus group III, Sa III) (8), AAG03056 (S. aureus group IV, Sa IV) (7), AAC38294 (S. epidermidis, Se) (35), AAA71977(S. lugdunensis, Sl) (34), and AAS66746 (S. intermedius ATCC 29663, Si).
3142 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
acrylamide Tris-Tricine SDS-polyacrylamide gel electrophoresis (29) and elec-trophoretically transferred onto a Protran nitrocellulose membrane (Schleicher& Schuell). After incubation at 4°C overnight in blocking buffer (Tris-bufferedsaline plus 0.05% Tween 20 and 5% bovine serum albumin [TBST]) (28), themembranes were incubated in blocking buffer containing anti-T7-tag monoclonalantibody (1:5,000 dilution; Novagen) for 1 h at room temperature. The mem-branes were washed extensively with TBST buffer, probed with horseradishperoxidase-conjugated goat anti-mouse antibody (Amersham), and detected withECL Plus Western blotting detection kit (Amersham), followed by exposure toKodak Biomax MR film.
The S. intermedius agr nucleotide sequence. The nucleotide sequences of the S.intermedius ATCC 29663 agr locus (GenBank accession number AY557375),agrD genes (RN9161, GenBank accession number AY87105; RN9167, GenBankaccession number AY87106; RN9169, GenBank accession numberAY87107; andRN9515, GenBank accession number AF346723), and hld coding sequence of S.intermedius CCM5739 (GenBank accession number AY860843) have been de-posited in the GenBank.
RESULTS
The S. intermedius agrD sequences. We determined theDNA sequence of agrD genes for three uncharacterized S.intermedius strains kindly provided by Wesley Kloos, two morefrom our strain collection, and a sixth strain, ATCC 29663, byusing the PCR methods described in Materials and Methods.
These are shown in Fig. 1A with the four groups of the S.aureus AgrDs for comparison. AgrD-Si is highly conserved andthere are two variants among the six strains. It is not known atpresent whether this variation affects activation specificity.Shown in boldface are the predicted sequences of the S. inter-medius AIPs, one of which, that of CCM5739, has been con-firmed as a cyclic lactone with the above sequence by massspectroscopy (10). Note that the C-terminal processing site isabsolutely conserved among these strains, as well as among 30other staphylococci, whereas the N-terminal processing site isnot conserved among other staphylococci (2). The latter isclearly conserved, however, among these S. intermedius strains.
The AgrB-Si sequence. Sequencing of a 2,084-bp segment ofthe S. intermedius ATCC 29663 agr locus revealed that thisportion of the locus was identical to the published partialsequence of the same region of S. intermedius CCM5739 (2).Comparison of the AgrB-Si sequence to that of other staphy-lococci showed that AgrB-Si was clearly related to other staph-ylococcal AgrBs (Fig. 1B). Interestingly, the N-terminal region,two regions that are highly hydrophilic and are proposed to betwo transmembrane segments (37), and a small C-terminalregion were significantly similar. The divergent N-terminal halfof AgrC (data not shown) was also clearly a member of theAgrC receptor family, having significant similarities with otherAgrC receptor domains (2, 20), but with little overall conser-vation, providing no obvious indication of how it would beactivated by a lactone AIP.
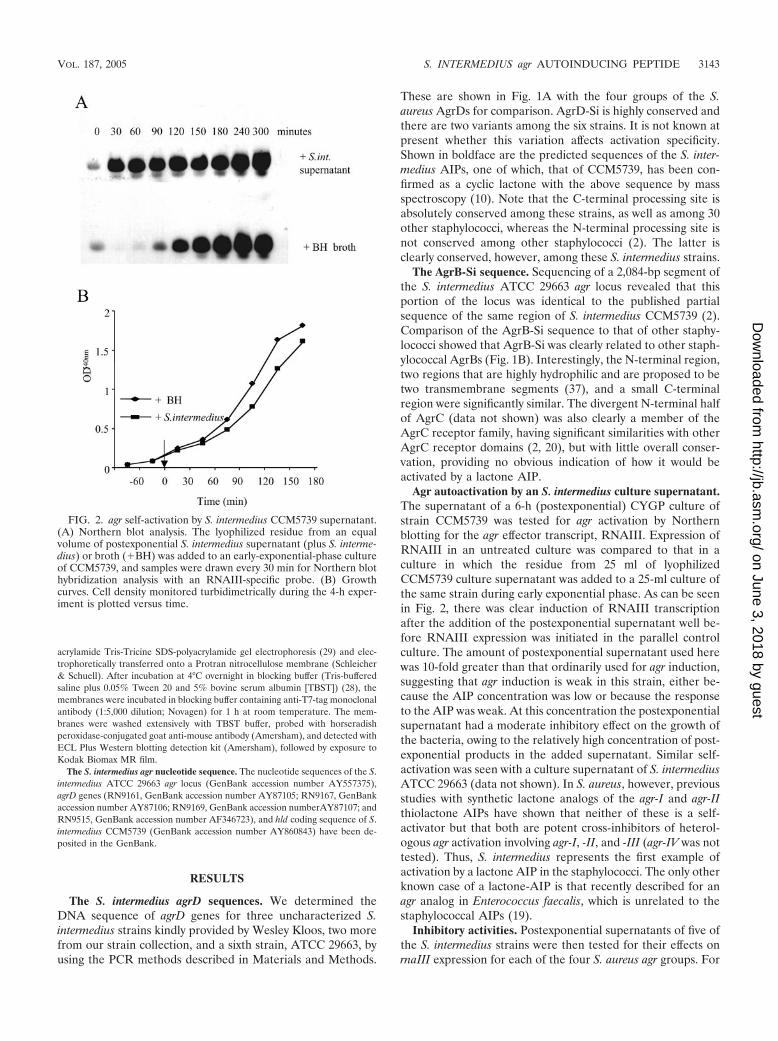
Agr autoactivation by an S. intermedius culture supernatant.The supernatant of a 6-h (postexponential) CYGP culture ofstrain CCM5739 was tested for agr activation by Northernblotting for the agr effector transcript, RNAIII. Expression ofRNAIII in an untreated culture was compared to that in aculture in which the residue from 25 ml of lyophilizedCCM5739 culture supernatant was added to a 25-ml culture ofthe same strain during early exponential phase. As can be seenin Fig. 2, there was clear induction of RNAIII transcriptionafter the addition of the postexponential supernatant well be-fore RNAIII expression was initiated in the parallel controlculture. The amount of postexponential supernatant used herewas 10-fold greater than that ordinarily used for agr induction,suggesting that agr induction is weak in this strain, either be-cause the AIP concentration was low or because the responseto the AIP was weak. At this concentration the postexponentialsupernatant had a moderate inhibitory effect on the growth ofthe bacteria, owing to the relatively high concentration of post-exponential products in the added supernatant. Similar self-activation was seen with a culture supernatant of S. intermediusATCC 29663 (data not shown). In S. aureus, however, previousstudies with synthetic lactone analogs of the agr-I and agr-IIthiolactone AIPs have shown that neither of these is a self-activator but that both are potent cross-inhibitors of heterol-ogous agr activation involving agr-I, -II, and -III (agr-IV was nottested). Thus, S. intermedius represents the first example ofactivation by a lactone AIP in the staphylococci. The only otherknown case of a lactone-AIP is that recently described for anagr analog in Enterococcus faecalis, which is unrelated to thestaphylococcal AIPs (19).
Inhibitory activities. Postexponential supernatants of five ofthe S. intermedius strains were then tested for their effects onrnaIII expression for each of the four S. aureus agr groups. For
FIG. 2. agr self-activation by S. intermedius CCM5739 supernatant.(A) Northern blot analysis. The lyophilized residue from an equalvolume of postexponential S. intermedius supernatant (plus S. interme-dius) or broth (�BH) was added to an early-exponential-phase cultureof CCM5739, and samples were drawn every 30 min for Northern blothybridization analysis with an RNAIII-specific probe. (B) Growthcurves. Cell density monitored turbidimetrically during the 4-h exper-iment is plotted versus time.
VOL. 187, 2005 S. INTERMEDIUS agr AUTOINDUCING PEPTIDE 3143
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
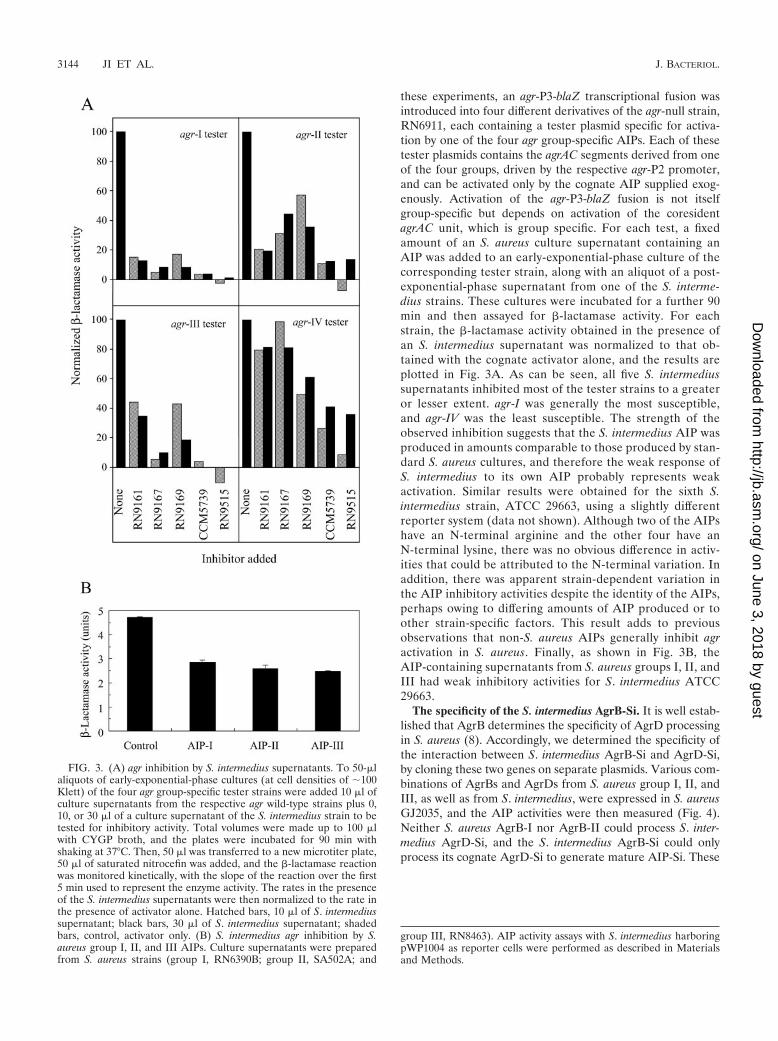
these experiments, an agr-P3-blaZ transcriptional fusion wasintroduced into four different derivatives of the agr-null strain,RN6911, each containing a tester plasmid specific for activa-tion by one of the four agr group-specific AIPs. Each of thesetester plasmids contains the agrAC segments derived from oneof the four groups, driven by the respective agr-P2 promoter,and can be activated only by the cognate AIP supplied exog-enously. Activation of the agr-P3-blaZ fusion is not itselfgroup-specific but depends on activation of the coresidentagrAC unit, which is group specific. For each test, a fixedamount of an S. aureus culture supernatant containing anAIP was added to an early-exponential-phase culture of thecorresponding tester strain, along with an aliquot of a post-exponential-phase supernatant from one of the S. interme-dius strains. These cultures were incubated for a further 90min and then assayed for �-lactamase activity. For eachstrain, the �-lactamase activity obtained in the presence ofan S. intermedius supernatant was normalized to that ob-tained with the cognate activator alone, and the results areplotted in Fig. 3A. As can be seen, all five S. intermediussupernatants inhibited most of the tester strains to a greateror lesser extent. agr-I was generally the most susceptible,and agr-IV was the least susceptible. The strength of theobserved inhibition suggests that the S. intermedius AIP wasproduced in amounts comparable to those produced by stan-dard S. aureus cultures, and therefore the weak response ofS. intermedius to its own AIP probably represents weakactivation. Similar results were obtained for the sixth S.intermedius strain, ATCC 29663, using a slightly differentreporter system (data not shown). Although two of the AIPshave an N-terminal arginine and the other four have anN-terminal lysine, there was no obvious difference in activ-ities that could be attributed to the N-terminal variation. Inaddition, there was apparent strain-dependent variation inthe AIP inhibitory activities despite the identity of the AIPs,perhaps owing to differing amounts of AIP produced or toother strain-specific factors. This result adds to previousobservations that non-S. aureus AIPs generally inhibit agractivation in S. aureus. Finally, as shown in Fig. 3B, theAIP-containing supernatants from S. aureus groups I, II, andIII had weak inhibitory activities for S. intermedius ATCC29663.
The specificity of the S. intermedius AgrB-Si. It is well estab-lished that AgrB determines the specificity of AgrD processingin S. aureus (8). Accordingly, we determined the specificity ofthe interaction between S. intermedius AgrB-Si and AgrD-Si,by cloning these two genes on separate plasmids. Various com-binations of AgrBs and AgrDs from S. aureus group I, II, andIII, as well as from S. intermedius, were expressed in S. aureusGJ2035, and the AIP activities were then measured (Fig. 4).Neither S. aureus AgrB-I nor AgrB-II could process S. inter-medius AgrD-Si, and the S. intermedius AgrB-Si could onlyprocess its cognate AgrD-Si to generate mature AIP-Si. These
FIG. 3. (A) agr inhibition by S. intermedius supernatants. To 50-�laliquots of early-exponential-phase cultures (at cell densities of �100Klett) of the four agr group-specific tester strains were added 10 �l ofculture supernatants from the respective agr wild-type strains plus 0,10, or 30 �l of a culture supernatant of the S. intermedius strain to betested for inhibitory activity. Total volumes were made up to 100 �lwith CYGP broth, and the plates were incubated for 90 min withshaking at 37°C. Then, 50 �l was transferred to a new microtiter plate,50 �l of saturated nitrocefin was added, and the �-lactamase reactionwas monitored kinetically, with the slope of the reaction over the first5 min used to represent the enzyme activity. The rates in the presenceof the S. intermedius supernatants were then normalized to the rate inthe presence of activator alone. Hatched bars, 10 �l of S. intermediussupernatant; black bars, 30 �l of S. intermedius supernatant; shadedbars, control, activator only. (B) S. intermedius agr inhibition by S.aureus group I, II, and III AIPs. Culture supernatants were preparedfrom S. aureus strains (group I, RN6390B; group II, SA502A; and
group III, RN8463). AIP activity assays with S. intermedius harboringpWP1004 as reporter cells were performed as described in Materialsand Methods.
3144 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

results indicated that the interaction between AgrB-Si andAgrD-Si was specific.
Cross-processing of S-to-C and C-to-S mutants of AgrD.Since AgrB determines the specificity of AIP processing (8), itis very likely that AgrB catalyzes the formation of the cyclicthiolactone bond in the S. aureus AIPs and, therefore, also thecyclic lactone bond in the S. intermedius AIPs. We thereforetested the possibility that AgrB-I can catalyze the formation ofthe AIP-I lactone analog by mutationally replacing the cysteinecodon (TGT) in agrD-I with a serine codon (AGT), yieldingagrD-I (C28S). Since the synthetic AIP-I lactone is a potentcross-inhibitor, and possibly a very weak self-activator, culturesupernatants of the mutant strain were then tested for inhibi-tion of agr activation in the other S. aureus agr groups, as wellas for self-activation of agr-I. As shown in Fig. 5, no inhibitoryactivity was detected, nor did this supernatant activate the agr-Ireporter or either of the other two. For these tests, we usedonly the group I, II, and III reporter strains, since these wouldhave detected any AIP produced. Note that activation of thegroup I reporter was seen only in track C, containing the
wild-type agrD-I and agrB-I plasmids. No activation was seenwith either the group II or III reporters. Conversely, inhibitionof either the group II or group III reporters (tracks C) was seenonly with the wild-type agrD-B combination. We thereforeconclude that AgrB-I cannot process a serine-containingAgrD-I.
We next prepared and tested the corresponding S27C mu-tant in AgrD-Si, in which the serine was replaced by a cysteineand also an S27A mutation. As noted, S. intermedius AgrB-Si
FIG. 4. Interaction between AgrB and AgrD. Conditioned mediawere prepared from S. aureus GJ2035 expressing various combinationsof AgrB and AgrD, and the AIP activities were measured by using S.intermedius containing pWP1004 as reporter cells. (A and B) Test foractivation of S. intermedius agr by AIPs prepared from cells coexpress-ing S. aureus AgrB-I and AgrB-II or S. intermedius AgrB-Si and the S.intermedius AgrD-Si (A) and test for inhibition of S. intermedius agr byAIPs prepared from bacteria coexpressing the S. intermedius AgrB-Siand S. aureus AgrD-I, AgrD-II, or AgrD-III (B). Reporter cells grownin the absence of AIP were used as controls. Values are means fromthree independent experiments with standard errors as indicated.
FIG. 5. Inability of AgrB-I to process a serine-containing AgrD-Imutant. Using a cloned agrD-I derivative, the cysteine codon, TGT,was replaced by a serine codon, AGT, and the mutant agrD was clonedinto a vector between N- and C-terminal His6 tags. The resultingconstruct was tested in vivo in the presence or absence of an agrB-I-containing plasmid and compared to the native agrD-I, also containingthe N- and C-terminal His tags, for the production of agr-activating or-inhibiting substances by using agr reporter strains with a �-lactamasereadout. (A) For activation tests, the culture was grown for 90 min inthe presence of the supernatant to be tested for activation and thenassayed. (B) For inhibition tests, a sample of a cognate supernatant(activator) (i.e., group II supernatant for the group II reporter andgroup III supernatant for the group III reporter) was added at thesame time as the supernatant to be tested for inhibition, and theculture then grown for 90 min and assayed for �-lactamase. For eachof the five sets of tests shown, the columns are labeled as follows: A,wild-type AgrD; B, AgrD, C28S; C, wild-type AgrD plus AgrB-I; D,AgrD, C28S plus AgrB-I [the reporters were RN6390B(pRN6683) forgroup I, SA502A(pRN6683) for group II, and RN8463(pRN6683) forgroup III as described previously (8, 9)].
VOL. 187, 2005 S. INTERMEDIUS agr AUTOINDUCING PEPTIDE 3145
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

is quite similar to the other known AgrBs (2), so that itsdivergent sites may be informative with respect to the pro-AIPprocessing mechanism and, especially, with regard to the abil-ity to process a serine-containing pro-AIP. As shown in Fig. 6,the AgrD-Si serine-to-cysteine mutant, AgrD-Si(S27C), couldbe processed by AgrB-Si to generate a mature mutant AIP, as
determined by AIP activity assays (Fig. 6A to D). However,activation of the S. intermedius agr response by the mutantAIP-Si was much weaker than that by wild-type AIP-Si. TheAgrD-Si serine-to-alanine mutant, AgrD-Si(S27A), did notgenerate any functionally detectable AIP (Fig. 6). The inhibi-tory activities of the S27C mutant AIP-Si (presumably a thio-
FIG. 6. Processing of the wild-type and the serine-to-cysteine mu-tant AgrD-Si by AgrB-Si. (A to D) AIP activity assays. S. aureus cellswere grown and induced. After centrifugation, the culture superna-tants were used as the conditioned media (either concentrated ordiluted with CYGP medium) to perform either AIP activation assayswith S. intermedius containing pWP1004 as reporter cells (A) or AIPinhibition assays with RN6390B(pRN6683) (S. aureus group I) (B),SA502A(pRN6683) (S. aureus group II) (C), and RN8463(pRN6683)(S. aureus group III) (D) reporter cells. Undiluted conditioned mediawere equal to 100% AIP. Conditioned media were prepared fromcultures of cells coexpressing AgrB-Si and the wild-type AgrD-Si (F),AgrD-Si(S27C) (Œ), or AgrD-Si(S27A) (■). Values are means fromthree independent experiments with the standard errors as indicated.(E) Western blot hybridization analysis. S. aureus cells coexpressingAgrB-Si and wild-type AgrD-Si, AgrD-Si(S27C), or AgrD-Si(S27A)were grown and induced, and the cell cultures were mixed with etha-nol-acetone (1:1). The mixtures were centrifuged, and the cells werewashed and lysed. Whole-cell lysates were separated by SDS-polyacryl-amide gel electrophoresis and transferred to nitrocellulose mem-branes. The membranes were then probed with an anti-T7 tag mono-clonal antibody.
3146 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

lactone molecule) on the agr responses in group II but not ingroup I and III S. aureus strains were stronger than those of thewild-type AIP-Si (Fig. 6B to D).
To demonstrate that AgrB-Si was involved in the proteolyticprocessing of AgrD-Si, we performed Western blot hybridiza-tion analysis with an anti-T7 monoclonal antibody as a probe.We note that AgrD-Si used in these experiments was doublytagged with a T7 epitope at the N terminus and a six-histidinestretch (His6) at the C terminus, which would facilitate thedetection of AgrD-Si, as well as of its potential processingintermediate(s), by Western blot analyses with commerciallyavailable antibodies. We note that cells expressing the doublytagged AgrD-Si and AgrB-Si produced an amount of AIP-Sicomparable to those expressing the wild-type AgrD-Si andAgrB-Si, indicating that the addition of these tags had noeffects on the AgrD-Si processing and AIPSi secretion (datanot shown). As shown in Fig. 6E, a band with a molecular massof ca. 9 kDa corresponding to the calculated molecular mass ofthe doubly tagged AgrD-Si (9,205 Da) was detected in the lanecontaining a lysate of cells lacking agrB-Si or containing AgrD-Si(S27A) and producing the tagged AgrD-Si. In the presenceof the wild-type AgrB-Si or AgrD-Si(S27C), an anti-T7 anti-body-responding band with a molecular mass of ca. 6 kDa wasdetected. Although this band probably corresponds to a pro-cessing intermediate previously observed with S. aureusAgrD-I (18), we have not been able to obtain sufficient mate-rial for confirmation by mass spectroscopy. These results sug-gested that AgrB-Si could process both the serine and thecysteine containing AgrD-Si to generate AIPs that had theability to activate the S. intermedius and inhibit the S. aureusagr responses.
Hemolytic activities and agr function. agr is widely con-served among the staphylococci (2), although its function hasbeen defined in only two species, S. aureus and S. epidermidis(9, 36). Given that the S. intermedius lactone AIP is an agrautoinducer, it follows that agr is likely to be functional in S.intermedius, which would make S. intermedius the third. A verypreliminary picture of the hemolytic activities of S. intermediussupports this, although a definitive conclusion would requirethe construction of an agr knockout. The production of delta-hemolysin, which is encoded by rnaIII, is generally an indica-tion of agr self-activation. We have determined the DNA se-quence of agr rnaIII in an S. intermedius strain, CCM5739, andidentified a typical staphylococcal delta-hemolysin readingframe (3). An alignment of delta-hemolysin sequences fromstaphylococci is shown in Table 3. The NCBI accession num-bers are as follows: AAW55662 (S. intermedius), CAA11542 (S.simulans) (33), LESAD (S. aureus) (3), CAA11541 (S. epider-midis) (33), CAA11543 (S. warneri 1) (33), and CAA11544 (S.warneri 2) (33).
The results of a simple test for the production of delta-hemolysin and other hemolysins, involving cross-streaking onsheep blood agar against RN4220, are shown in Fig. 7. At leastfour of the five S. intermedius strains shown—RN9161,RN9167, 9168, and 9515—produce delta-hemolysins that aredetectable by this test. The fifth strain, CCM5739 (shown asRN9423 in the picture), is also likely to produce delta-hemo-lysin, since agr is autoinducible in this strain and encodes delta-hemolysin, although it is not identifiable in the CCM5739 he-molytic pattern. Hemolysins are typically agr upregulated, butthe group of S. intermedius strains is rather heterogeneous withrespect to hemolytic activities. Thus, three produce beta-hemoly-sin, although in considerably different amounts, and none pro-duces detectable amounts of alpha-hemolysin, but CCM5739 hasthe strongest hemolytic activity of any of the five, probably pro-ducing at least two different hemolysins, neither of which is typicalof the S. aureus hemolysins as characterized by this simple test.These results are consistent with agr functionality but certainly donot prove it. Experiments are in progress to isolate an agr knock-out to obtain a definitive view of what is regulated by agr in S.intermedius.
DISCUSSION
Since S. intermedius was classified as a new staphylococcalspecies in 1972, this bacterium has been isolated from variousanimals (12). S. intermedius produces toxins, hemolysins, coag-ulase and maybe other virulence factors that have been sug-gested to cause food poisoning in humans (1) and diseasesranging from abscesses to mastitis and endocarditis in dogs (4,31). Occasionally, S. intermedius causes infection in humans(32). Although S. intermedius is, in most respects, a typicalstaphylococcal species, it differs strikingly in one respect: itcontains a serine in place of the cysteine that is absolutelyconserved in the AIPs of all of the 14 other staphylococcalspecies thus far analyzed. All of the six S. intermedius strainsstudied produce a nonomeric agrD lactone AIP. The predictedAgrB-Si, AgrD-Si, and the N-terminal portion of AgrC-Si fromthe sequenced S. intermedius agr locus were clearly members ofthe widespread family of agr gene products. The AgrB-Si spe-cifically processed AgrD-Si to produce mature AIP-Si, and italso had one interesting feature that was different from otherAgrBs, i.e., its abilities to process both the wild-type and amutant AgrD-Si, in which the serine residue in the AIP-Siregion was replaced by a cysteine residue, to produce matureAIPs. In contrast, the AgrB-I could process only the nativecysteine containing AgrD-I but not cysteine-to-serine mutantsto generate mature AIP. We note that chemically synthesizedlinear AIPs have neither activation nor inhibition activities (9,17). These results imply that AgrB-Si can catalyze the forma-tion of both ester and thioester bonds in AIP-Si.
The agr autoinducing activity has been demonstrated for twoof the strains, and the production of a cyclic lactone was con-firmed. Cross-inhibition has been demonstrated for all sixstrains. Four of the six strains clearly produce delta-hemolysin,and the fifth probably does so as well, and the overall hemolyticpatterns are consistent with agr functionality. Given that theonly agr autoinducing and cross-inhibiting substances yet iden-tified are the cyclic AIPs, we suggest that S. intermedius pro-
TABLE 3. Alignment of delta-hemolysin sequences
Strain Sequence
S. intermedius .................................MAGDIISTIVDFIKLIAETVKKFTKS. simulans .....................................MAGDIVGTIGEFVKLIIETVQKFTQKS. aureus .........................................MAQDIISTIGDLVKWIIDTVNKFTKS. epidermidis .................................MMAADIISTIGDLVKWIIDTVNKFKKS. warneri I .....................................MAADIISTIGDLVKLIINTVKKFQKS. warneri II....................................MTADIISTIGDFVKWILDTVKKFTK
VOL. 187, 2005 S. INTERMEDIUS agr AUTOINDUCING PEPTIDE 3147
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

duces lactone AIPs that are active as autoinducers and cross-inhibitors and that agr has a functional role in S. intermedius.
An enduring question is why evolution has favored the de-velopment of cyclic AIPs in staphylococci and enterococci,since cyclic peptides are not used in any of the other knownpeptide-based receptor-ligand signaling systems described todate. We do not have any compelling hypothesis for this. It wasinitially thought that activation of the receptor would involvethe formation of a covalent bond with the AIP. Although thishas been ruled out, the possibility of a transient covalent bond
has not been ruled out (14). It has also been suggested that thecyclic configuration could confer stability toward environmen-tal proteases; this would have to be tested by a comparison oflinear peptides in other systems with the staphylococcal andenterococcal cyclic ones. Since lactone- and thiolactone-con-taining peptides would have very different intrinsic stabilities, itis difficult to argue that intrinsic stability is relevant here.
There is increasing evidence that AgrB is necessary andsufficient for the processing and secretion of the agr AIPs (8, 9,37–39) and that the thiolactone configuration is necessary for
FIG. 7. Hemolytic patterns. Cultures to be tested were grown overnight on GL agar, cross-streaked on sheep blood agar against a culture ofRN4220, incubated overnight at 37°C, and then incubated for 6 h at 4°C. The patterns can be interpreted as follows: the “hot-cold” beta-hemolysinappears as a partially turbid zone, as seen with RN4220, which produces only beta-hemolysin. Delta-hemolysin is synergistic with beta-hemolysinand is seen as a clearing where the two hemolysins intersect; this is best seen with RN9515 (top left). Coproduction of beta-hemolysin anddelta-hemolysin is seen as a clearing next to the streak within a wider beta-hemolysin zone (best illustrated with RN9169, bottom left) and, lessstrongly, with RN9167 (bottom right). RN9161 produces only delta-hemolysin and quite weakly. RN6734 produces a very strong alpha-hemolysinzone, as shown by the characteristic antagonism between alpha-hemolysin and beta-hemolysin. It also produces delta-hemolysin, as shown by theclearer zone where the beta-hemolysin and delta-hemolysin zones intersect. RN7206 produces a very weak alpha-hemolysin zone, as shown by itsinhibition by beta-hemolysin. The CCM5739 (labeled RN9423) pattern could represent a beta-hemolysin zone equivalent to that of RN9169 plusa very strong delta-hemolysin zone, which obscures that of beta-hemolysin. The interaction of CCM5739 (labeled RN9423) with the RN4220beta-hemolysin zone is very atypical and is not interpretable according to our understanding of the activities of the S. aureus hemolysins.
3148 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

activation of AgrC. Given that at least one of the S. aureusAgrBs cannot process a serine-containing variant of its pro-AIP, the ability of the S. intermedius AgrBs to do so and theability of S. intermedius AgrC to be activated by the lactoneAIP have interesting implications regarding the evolutionaryparadigm that we have favored to account for the apparentcovariation in AgrB, -D, and -C, i.e., that the sequence of theAIP, the processing activity of AgrB, and the receptor speci-ficity of AgrC would have to evolve in concert, so as to preservethe three-way specificity of the system. Several experimentspresented here and elsewhere address this point. We haveshown here that a cysteine to serine mutation (in AgrD-I)inactivates the system because AgrB-I cannot process thepropeptide and, previously, that neither the synthetic AIP-Inor -II lactone can activate the cognate AgrC (17, 18). We haveobserved, however, that a mutant S. intermedius AgrD propep-tide in which the serine has been replaced by a cysteine can beprocessed to generate an AIP, as demonstrated by testingculture supernatants for AIP activity. This AIP, which wouldcontain a cysteine in place of the native serine, is weakly activeas an autoinducer, although potent as a cross-inhibitor of S.aureus agr.
Given that the replacement of a cysteine by a serine or viceversa can occur by a single point mutation, the spontaneousoccurrence of a C-to-S or S-to-C switch would have a reason-able probability. If we assume that the S. intermedius propep-tide arose by a C-S point mutation and that the S. intermediusagrB and -C subsequently evolved to accommodate the change,it seems reasonable to imagine that S. intermedius AgrB “re-members” how to process a cysteine-containing propeptideand that S. intermedius AgrC “remembers” how to respond toa thiolactone AIP. On the other hand, AgrB-I of S. aureuswould never have encountered a serine-containing AgrD andAgrC-I of S. aureus would never have encountered a self-codedlactone AIP. Were a C-S mutation to occur in S. aureus, therest of the locus would have to evolve so as to restore activityto the system. Since most or all S. intermedius strains have alactone AIP, one would have to postulate that the divergence,giving rise to S. intermedius, which is the canine equivalent ofS. aureus, would probably have occurred after the putativeC-to-S mutation in the AIP gene. According to Kloos’ hypoth-esis of coevolution of staphylococci and mammals (W. E.Kloos, unpublished data), S. intermedius would probably havediverged from S. aureus coincidentally with the divergence ofthe line leading to the carnivores from that leading to theprimates, ca. 55 million years ago.
ACKNOWLEDGMENTS
This study was supported by NIH grants RO1AI30138 andRO1AI42783 to R.P.N. and by RO1AI46445 to G.J.
We are grateful to Brian Weinrick for measuring the inhibitoryactivities of the S. intermedius supernatants.
REFERENCES
1. Becker, K., B. Keller, C. von Eiff, M. Bruck, G. Lubritz, J. Etienne, and G.Peters. 2001. Enterotoxigenic potential of Staphylococcus intermedius. Appl.Environ. Microbiol. 67:5551–5557.
2. Dufour, P., S. Jarraud, F. Vandenesch, T. Greenland, R. P. Novick, M. Bes,J. Etienne, and G. Lina. 2002. High genetic variability of the agr locus inStaphylococcus species. J. Bacteriol. 184:1180–1186.
3. Fitton, J. E., A. Dell, and W. V. Shaw. 1980. The amino acid sequence of thedelta haemolysin of Staphylococcus aureus. FEBS Lett. 115:209–212.
4. Girard, C., and R. Higgins. 1999. Staphylococcus intermedius cellulitis andtoxic shock in a dog. Can. Vet. J. 40:501–502.
5. Janzon, L., and S. Arvidson. 1990. The role of the delta-lysin gene (hld) inthe regulation of virulence genes by the accessory gene regulator (agr) inStaphylococcus aureus. EMBO J. 9:1391–1399.
6. Janzon, L., S. Lofdahl, and S. Arvidson. 1989. Identification and nucleotidesequence of the delta-lysin gene, hld, adjacent to the accessory gene regu-lator (agr) of Staphylococcus aureus. Mol. Gen. Genet. 219:480–485.
7. Jarraud, S., G. J. Lyon, A. M. Figueiredo, L. Gerard, F. Vandenesch, J.Etienne, T. W. Muir, and R. P. Novick. 2000. Exfoliatin-producing strainsdefine a fourth agr specificity group in Staphylococcus aureus. J. Bacteriol.182:6517–6522.
8. Ji, G., R. Beavis, and R. P. Novick. 1997. Bacterial interference caused byautoinducing peptide variants. Science 276:2027–2030.
9. Ji, G., R. C. Beavis, and R. P. Novick. 1995. Cell density control of staphy-lococcal virulence mediated by an octapeptide pheromone. Proc. Natl. Acad.Sci. USA 92:12055–12059.
10. Kalkum, M., G. J. Lyon, and B. T. Chait. 2003. Detection of secretedpeptides by using hypothesis-driven multistage mass spectrometry. Proc.Natl. Acad. Sci. USA 100:2795–2800.
11. Kleerebezem, M., L. E. Quadri, O. P. Kuipers, and W. M. de Vos. 1997.Quorum sensing by peptide pheromones and two-component signal-trans-duction systems in gram-positive bacteria. Mol. Microbiol. 24:895–904.
12. Kloos, W. E. 1980. Natural populations of the genus Staphylococcus. Annu.Rev. Microbiol. 34:559–592.
13. Kornblum, J. S., S. J. Projan, S. L. Moghazeh, and R. P. Novick. 1988. Arapid method to quantitate non-labeled RNA species in bacterial cells. Gene63:75–85.
14. Lyon, G. J., P. Mayville, T. W. Muir, and R. P. Novick. 2000. Rational designof a global inhibitor of the virulence response in Staphylococcus aureus,based in part on localization of the site of inhibition to the receptor-histidinekinase, AgrC. Proc. Natl. Acad. Sci. USA 97:13330–13335.
15. Lyon, G. J., J. S. Wright, A. Christopoulos, R. P. Novick, and T. W. Muir.2002. Reversible and specific extracellular antagonism of receptor-histidinekinase signaling. J. Biol. Chem. 277:6247–6253.
16. Lyon, G. J., J. S. Wright, T. W. Muir, and R. P. Novick. 2002. Key determi-nants of receptor activation in the agr autoinducing peptides of Staphylococ-cus aureus. Biochemistry 41:10095–10104.
17. Mayville, P., G. Ji, R. Beavis, H. Yang, M. Goger, R. P. Novick, and T. W.Muir. 1999. Structure-activity analysis of synthetic autoinducing thiolactonepeptides from Staphylococcus aureus responsible for virulence. Proc. Natl.Acad. Sci. USA 96:1218–1223.
18. McDowell, P., Z. Affas, C. Reynolds, M. T. Holden, S. J. Wood, S. Saint, A.Cockayne, P. J. Hill, C. E. Dodd, B. W. Bycroft, W. C. Chan, and P. Williams.2001. Structure, activity and evolution of the group I thiolactone peptidequorum-sensing system of Staphylococcus aureus. Mol. Microbiol. 41:503–512.
19. Nakayama, J., Y. Cao, T. Horii, S. Sakuda, and H. Nagasawa. 2001. Chem-ical synthesis and biological activity of the gelatinase biosynthesis-activatingpheromone of Enterococcus faecalis and its analogs. Biosci. Biotechnol. Bio-chem. 65:2322–2325.
20. Novick, R. P. 2003. Autoinduction and signal transduction in the regulationof staphylococcal virulence. Mol. Microbiol. 48:1429–1449.
21. Novick, R. P. 1991. Genetic systems in staphylococci. Methods Enzymol.204:587–636.
22. Novick, R. P., S. J. Projan, J. Kornblum, H. F. Ross, G. Ji, B. Kreiswirth, F.Vandenesch, and S. Moghazeh. 1995. The agr P2 operon: an autocatalyticsensory transduction system in Staphylococcus aureus. Mol. Gen. Genet.248:446–458.
23. Novick, R. P., H. F. Ross, A. M. S. Figueiredo, G. Abramochkin, and T. W.Muir. 2000. Activation and inhibition of the staphylococcal Agr system.Science 287:391a.
24. Novick, R. P., H. F. Ross, S. J. Projan, J. Kornblum, B. Kreiswirth, and S.Moghazeh. 1993. Synthesis of staphylococcal virulence factors is controlledby a regulatory RNA molecule. EMBO J. 12:3967–3975.
25. Otto, M., R. Sussmuth, G. Jung, and F. Gotz. 1998. Structure of the pher-omone peptide of the Staphylococcus epidermidis agr system. FEBS Lett.424:89–94.
26. Peng, H. L., R. P. Novick, B. Kreiswirth, J. Kornblum, and P. Schlievert.1988. Cloning, characterization, and sequencing of an accessory gene regu-lator (agr) in Staphylococcus aureus. J. Bacteriol. 170:4365–4372.
27. Recsei, P., B. Kreiswirth, M. O’Reilly, P. Schlievert, A. Gruss, and R. P.Novick. 1986. Regulation of exoprotein gene expression in Staphylococcusaureus by agar. Mol. Gen. Genet. 202:58–61.
28. Sambrook, J., and D. W. Russell. 2001. Molecular cloning: a laboratorymanual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y.
29. Schagger, H., and G. von Jagow. 1987. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the rangefrom 1 to 100 kDa. Anal. Biochem. 166:368–379.
30. Schenk, S., and R. A. Laddaga. 1992. Improved method for electroporationof Staphylococcus aureus. FEMS Microbiol. Lett. 73:133–138.
VOL. 187, 2005 S. INTERMEDIUS agr AUTOINDUCING PEPTIDE 3149
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

31. Smith, A. N., S. T. Finn-Bodner, and A. R. Dillon. 2000. Left ventricularoutflow tract to left atrial fistula associated with endocarditis in a dog. J. Am.Anim. Hosp. Assoc. 36:133–136.
32. Tanner, M. A., C. L. Everett, and D. C. Youvan. 2000. Molecular phyloge-netic evidence for noninvasive zoonotic transmission of Staphylococcus in-termedius from a canine pet to a human. J. Clin. Microbiol. 38:1628–1631.
33. Tegmark, K., E. Morfeldt, and S. Arvidson. 1998. Regulation of agr-depen-dent virulence genes in Staphylococcus aureus by RNAIII from coagulase-negative staphylococci. J. Bacteriol. 180:3181–3186.
34. Vandenesch, F., S. J. Projan, B. Kreiswirth, J. Etienne, and R. P. Novick.1993. Agr-related sequences in Staphylococcus lugdunensis. FEMS Micro-biol. Lett. 111:115–122.
35. Van Wamel, W. J., G. van Rossum, J. Verhoef, C. M. Vandenbroucke-Grauls,and A. C. Fluit. 1998. Cloning and characterization of an accessory gene
regulator (agr)-like locus from Staphylococcus epidermidis. FEMS Microbiol.Lett. 163:1–9.
36. Vuong, C., F. Gotz, and M. Otto. 2000. Construction and characterization of anagr deletion mutant of Staphylococcus epidermidis. Infect. Immun. 68:1048–1053.
37. Zhang, L., L. Gray, R. P. Novick, and G. Ji. 2002. Transmembrane topologyof AgrB, the protein involved in the post-translational modification of AgrDin Staphylococcus aureus. J. Biol. Chem. 277:34736–34742.
38. Zhang, L., and G. Ji. 2004. Identification of staphylococcal AgrB segment(s)responsible for group-specific processing of AgrD by gene swapping. J.Bacteriol. 186:6706–6713.
39. Zhang, L., J. Lin, and G. Ji. 2004. Membrane anchoring of the AgrDN-terminal amphipathic region is required for its processing to produce aquorum sensing pheromone in Staphylococcus aureus. J. Biol. Chem. 279:19448–19456.
3150 JI ET AL. J. BACTERIOL.
on June 3, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
















![Neutral Citation Number: [2018] EWHC 3139 (QB)](https://img.pdfslide.net/doc/110x75/6205381108631c7abc062464/neutral-citation-number-2018-ewhc-3139-qb.jpg)












