Embed Size (px)
Citation preview

Cell Tissue Res (1985) 242:197-209 Cell and Tissue Research �9 Springer-Verlag 1985
Structural and biochemical differentiation of the guinea-pig colon during foetal development Teresa Smith*, Kevin Christianson**, Raymond Moss*, and David Bailey** Departments of Electron Microscopy* and Surgery**, St. George's Hospital Medical School, London, Great Britain
Summary. We have studied some aspects of the morphologi- cal and biochemical differentiation of the foetal guinea-pig colonic epithelium. At day 40 the epithelium was organised in ridges and appeared pseudo-stratified. Folding of the epithelium, followed by villus formation, occurred between days 45 and 55, and by day 50 mucus-secreting goblet cells appeared at the bases of the colonic villi. By day 55 most epithelial cells, including goblet cells, possessed numerous microvilli which, by day 65, had become organised into well developed brush-borders. Between day 55 and term (day 65-68) mucosal depth increased markedly and the co- lon attained its final glandular morphology.
Biochemical studies showed the specific activities of the microvillar hydrolases to be much lower in the washed co- lon than in either foetal meconium or small intestine at all times during development. Furthermore, a membrane fraction highly enriched in microvillus hydrolase activities was prepared from foetal colonic meconium using tech- niques originally devised to isolate the foetal small intestinal microvillus membrane. This meconial subfraction was al- most identical in polypeptide composition to the highly- purified foetal small intestinal microvillus membrane. Iden- tification of the colonic microvillus membrane was ham- pered by the absence of reliable membrane markers. Never- theless, a fraction 14-fold enriched in aminopeptidase activi- ty was prepared from day 40 foetal colon and its polypep- tide composition compared by SDS-PAGE to that of the small intestinal microvillus membrane at the same age.
Key words: Foetal colon Guinea-pig - Meconium - Mi- crovillus membrane
Surprisingly little is known about the biochemical develop- ment of the mammalian colon, most biochemical studies having focussed on the small intestine and its differentia- tion-specific microvillar hydrolases (reviewed by Grand et al. 1976). These 'markers ' have assisted in the isolation and characterisation of the small intestinal microvillus membrane as a pure fraction for biochemical analysis dur- ing both foetal and postnatal development. Although the
Send offprint requests to." Dr. D.S. Bailey, Department of Surgery, St. George's Hospital Medical School, Cranmer Terrace, London SWt7 ORE, Great Britain
luminal surface of the adult mammalian colon possesses a clearly defined brush-border membrane, the latter ex- presses fewer differentiation markers than that of the small intestine. Its isolation has therefore remained somewhat en- igmatic (Biber et al. 1983).
In the human, there is strong evidence that some charac- teristics of the foetal colon are shared by colonic neoplasms. Glycogen storage, a characteristic feature of the foetal intes- tine (Lev and Weisberg 1969; Rousset et al. 1979), is also associated with the growth of human intestinal carcinoma cell lines both in vitro and in vivo (Rousset et al. 1980). Sucrase-isomaltase, a small intestinal microvillar hydrolase (Semenza et al. 1983), is also expressed during early devel- opment of the human foetal colon and in some colonic tumours (Zweibaum et al. 1983). The demonstration of these and other oncofoetal characteristics and their obvious clinical importance (Goldenberg 1976) necessitates further detailed studies of mammalian colonic development.
Colonic development has been examined in several ex- perimental systems using the rat (Helander 1973, 1975; Brackett and Townsend 1980; Colony and Neutra 1983) and chick (Lira and Low 1977). However, some aspects of gastrointestinal development differ between species with long and short gestation periods. In animals with short gestation periods, such as the mouse and rat, much intesti- nal maturation takes place after birth when the young are suckling. In contrast, both the human and the guinea-pig have relatively long gestation periods (266 and 68 days re- spectively), during which time the foetal intestine achieves considerable maturity (Grand et al. 1976; Moog and Ortiz 1960; Bailey et al. 1984). The structural and enzymatic dif- ferentiation of the human foetal colon have been reported recently (Bell and Williams 1982; Lacroix et al. 1984).
In the present study we have used a variety of techniques to investigate certain aspects of the structural organisation of the guinea-pig colon during foetal and early postnatal development. Some biochemical studies of the enzymatic differentiation of the foetal guinea-pig colon are presented, together with an analysis of foetal intestinal meconium. We have also isolated a microvillus membrane-enriched frac- tion from the 40-day foetal colon. The polypeptides of this membrane fraction have been analysed by one-dimensional sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) and have been compared with previous re- sults from our laboratory concerning the foetal guinea-pig small intestinal microvillus membrane (Bailey et al. 1984).

198
Materials and methods
Animals. Foetuses from timed-pregnant guinea-pigs (Dun- kin-Hartley strain) were exposed through a longitudinal in- cision in the maternal abdominal wall and a small incision in the uterine horn, under methoxyflurane anaesthesia (Penthrane, Abbott Laboratories, Queenborough, Kent). The entire foetal colon was excised, briefly washed with buffer (300 mM mannitol, 5 mM HEPES/KOH, pH 7.4) and immediately processed for light microscopy, electron microscopy or biochemical analysis.
Microscopy. Specimens for LM, TEM and SEM were ex- cised and immediately fixed in 50 mM cacodylate-buffered 3% glutaraldehyde (pH 7.2) for 12 h at room temperature. Post-fixation for 1 h in 1% osmium tetroxide in cacodylate buffer was followed by a buffer rinse and dehydration in graded ethanol/aqueous uranyl acetate. Propylene oxide acted as the intermediate fluid and specimens were embed- ded in Spurr's low viscosity resin. Sections were cut with a glass knife using a Reichert ultramicrotome. For TEM, sections were collected on 200 mesh copper grids, counter- stained with aqueous uranyl acetate and lead citrate (Stem- pak and Ward 1964; Sato 1968), and examined in a Philips 301 electron microscope. For LM, 2-~t resin-embedded sec- tions were stained with hot toluidine blue (Richardson et al. 1960) for 60 sec and examined in a Nikon light microscope.
Specimens for SEM were post-fixed for 1 h using 50 mM cacodylate-buffered 1% osmium tetroxide, rinsed with buffer and dehydrated through graded concentrations of ethanol and ethanol/freon. Specimens were critical-point dried, mounted on aluminium stubs, sputter-coated with gold and examined in a Coates and Welter field emission scanning electron microscope.
To identify intracellular glycogen, 2-1a resin-embedded sections of 50-day-old foetal colon were stained with peri- odic acid-Schiff reagent (BDH, Southampton) both with and without pretreatment with a-amylase according to the method of Lev and Weisberg (1969).
Enzymatic determinations. Aminopeptidase (E.C. 3.4.11.2) and dipeptidyl peptidase IV (E.C. 3.4.14.X) assays were performed as described by Wachsmuth et al. (1966) and Nagatsu et al. (1979), respectively. Alkaline phosphatase (E.C. 3.13.3.1) was assayed according to the method of Engstr6m (1961) as modified by Graham and Ford (1983). Sucrase (E.C. 3.2.1.48), enterokinase (E.C. 3.4.21.9) and protein were assayed by the methods of Dahlqvist (1968), Baratti et al. (1973), and Lowry et al. (1951) respectively.
Microvillus membrane preparation from foetal colon. Vesicu- larised microvillus membranes were prepared from 40-day foetal guinea-pig colon by cation precipitation, performed as described by Kessler et al. (1978) with the modifications of Gains and Hauser (1981), followed by isopycnic sucrose density gradient centrifugation as described in detail by Bai- ley et al. (1984) for foetal guinea-pig small intestine. This technique is similar to that originally used to prepare both the human small intestinal microvillus membrane by Schmitz et al. (1973) and the human foetal colon microvillus membrane by Lacroix et al. (1984), with the exception that the divalent cation used is Mg 2+ not C a 2 +
Briefly, colons from 20 40-day-old foetal guinea-pigs were excised and opened along their length with fine surgical scissors. All further operations were carried out
at 4 ~ C. Gentle washing in homogenisation buffer (300 mM mannitol, 5 mM HEPES/KOH pH 7.4) to remove any loosely-associated meconium was followed by homogeni- sation in a Waring blender (full speed, 3 min, in 150 ml buffer) and filtration through 40-11 nylon mesh. Two frac- tions were produced by differential centrifugation, a total membrane fraction and a microvillus-enriched membrane fraction.
The total membrane fraction was prepared by direct centrifugation of one half of the filtered homogenate at high speed (Sorvall RC5B centrifuge, SS34 rotor, 20000 rpm, 30 rain). The microvillus-enriched membrane fraction was prepared by adjusting the other half of the homogenate to 10mMMgC12, allowing non-microvillus membrane fragments to aggregate for 30 min, and then re- moving the aggregated membranes by low speed centrifuga- tion (Sorvall RC5B centrifuge, SS34 rotor, 5000rpm, 10 min). Microvillus-enriched membranes were then har- vested by high speed centrifugation of the resulting superna- rant (Sorvall RC5B centrifuge, SS34 rotor, 20000 rpm, 30 min). The resulting pellet was dispersed by passage through a gauge 19 syringe needle and applied to a 15-55% (w/w) sucrose density gradient. Isopycnic centrifugation (Beckman L8 centrifuge, SW40Ti rotor, 35000 rpm, 16 h) was followed by fractionation of the gradient into 1 ml frac- tions, and each fraction was assayed for aminopeptidase and alkaline phosphatase activities, as well as density (using an Abbe refractometer) and protein content. Equal volumes of certain fractions were then centrifuged (Beckman L8 cen- trifuge, 50Ti rotor, 45000 rpm, 90 min) and the pelleted material analysed by SDS-PAGE.
Analysis ofJoetal meconium. The separated colons and small intestines of several 60-day foetal guinea-pigs were carefully washed with homogenisation buffer (300 mM mannitol, 5 mM HEPES/KOH pH 7.4) to prepare mucosal and me- conial fractions. All subsequent operations were carried out at 4 ~ C. Colonic meconium and meconium-free small intes- tinal tissue were then homogenised separately in homogen- isation buffer using a Waring blender (full speed, 3 rain), and the homogenates adjusted to 10 mM MgC1 z. Aggrega- tion was allowed to proceed for 30 min, after which time the supernatant was clarified by low-speed centrifugation (Sorvall RC5B centrifuge, SS34 rotor, 5000 rpm, 10 rain). The resulting supernatants were then centrifuged at high speed (Sorvall RC5B centrifuge, SS34 rotor, 20000 rpm, 30 min), and the resulting pellets dispersed by passage through a gauge 19 syringe needle and applied to two identi- cal 15-55% (w/w) sucrose density gradients. Isopycnic cen- trifugation (Beckman L8 centrifuge, SW28 rotor, 25000 rpm, 16 h) was followed by fractionation of the gra- dients into 3 ml fractions. Each fraction was assayed for aminopeptidase activity, as well as density and protein con- tent. Specific fractions were then centrifuged (Beckman L8 ultracentrifuge, 50Ti rotor, 45000 rpm, 90 min) and the pel- leted material analysed by SDS-PAGE.
SDS-polyacrylamide gel electrophoresis. One-dimensional SDS-PAGE was performed using slabs of 7.5% polyacryl- amide as described by Laemmli (1970) under reducing con- ditions. Protein molecular weights were estimated by refer- ence to the migration of standard proteins (carbonic anhyd- rase, 29000 daltons; ovalbumin, 45 000 daltons; bovine se- rum albumin, 68 000 daltons; rabbit muscle phosphorylase
199
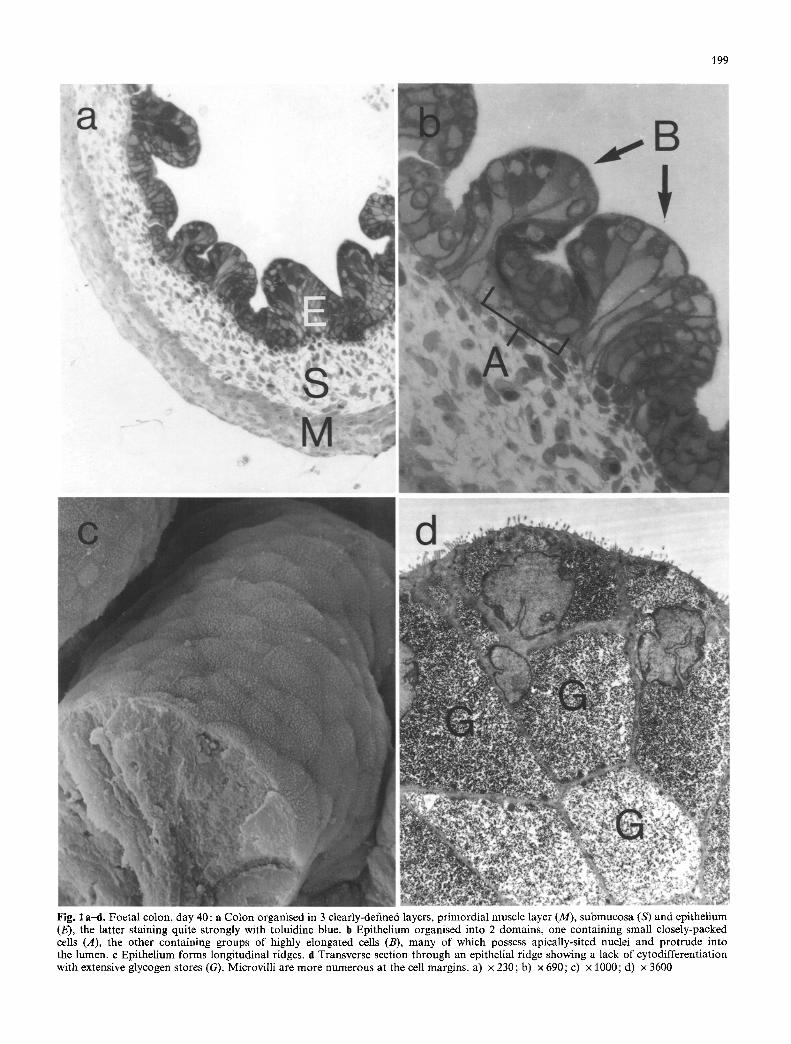
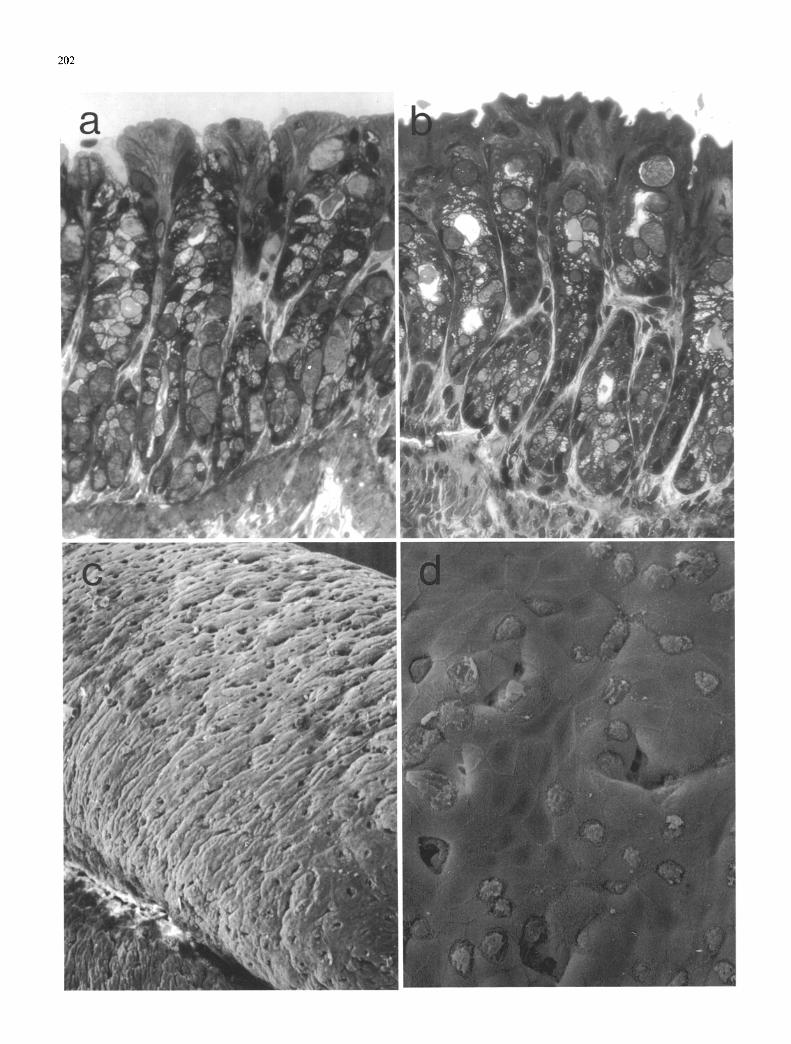
Fig. I a--d. Foetal colon, day 40: a Colon organised in 3 clearly-defined layers, primordial muscle layer (M), submucosa (S) and epithelium (E), the latter staining quite strongly with toluidine blue. b Epithelium organised into 2 domains, one containing small closely-packed cells (A), the other containing groups of highly elongated cells (B), many of which possess apically-sited nuclei and protrude into the lumen, e Epithelium forms longitudinal ridges, d Transverse section through an epithelial ridge showing a lack of cytodifferentiation with extensive glycogen stores (G). Microvilli are more numerous at the cell margins, a) • 230; b) • 690; c) • 1000; d) • 3600
200
201
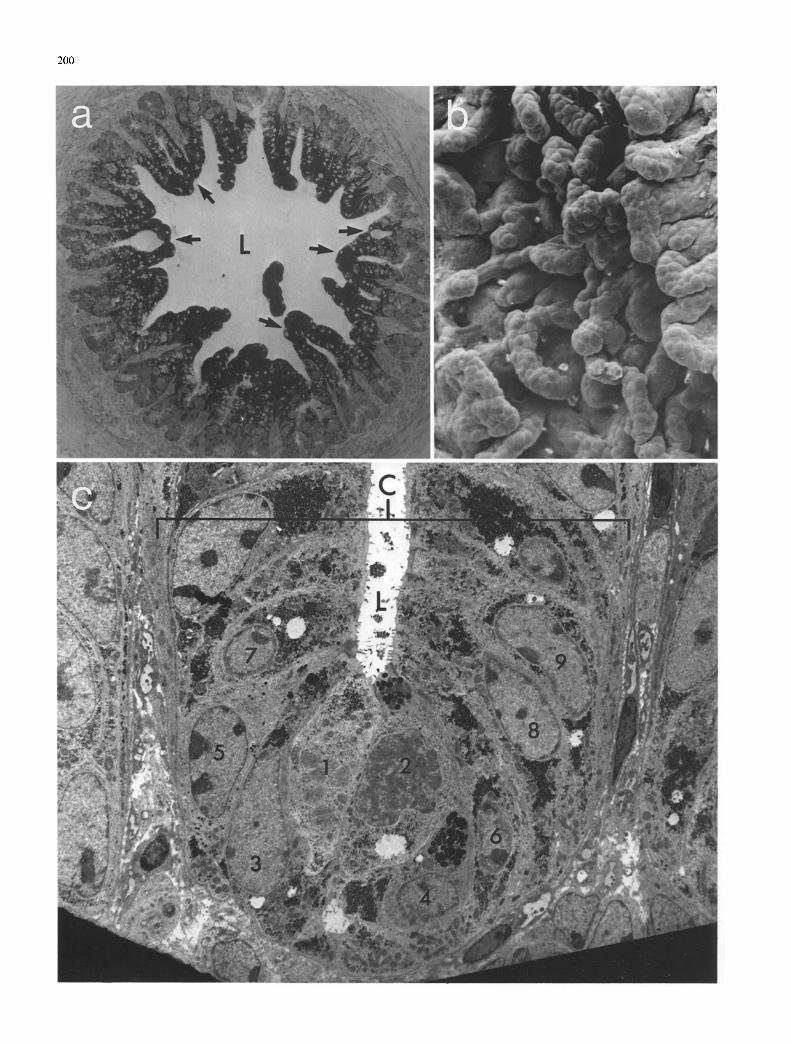
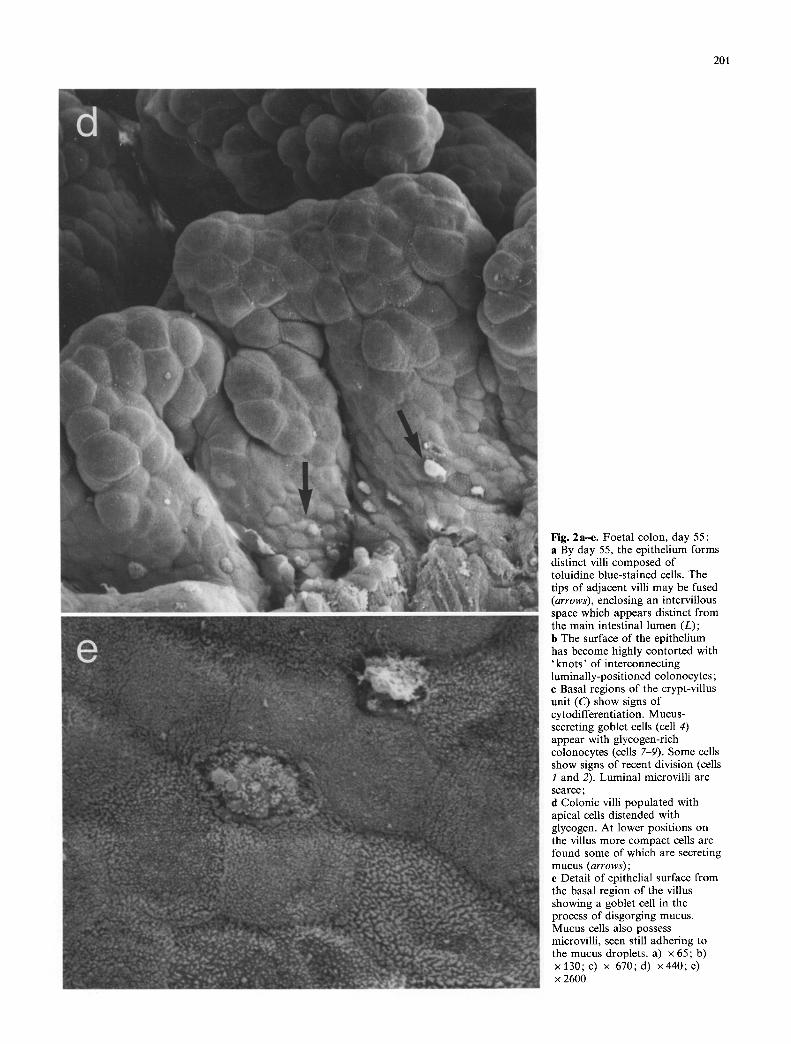
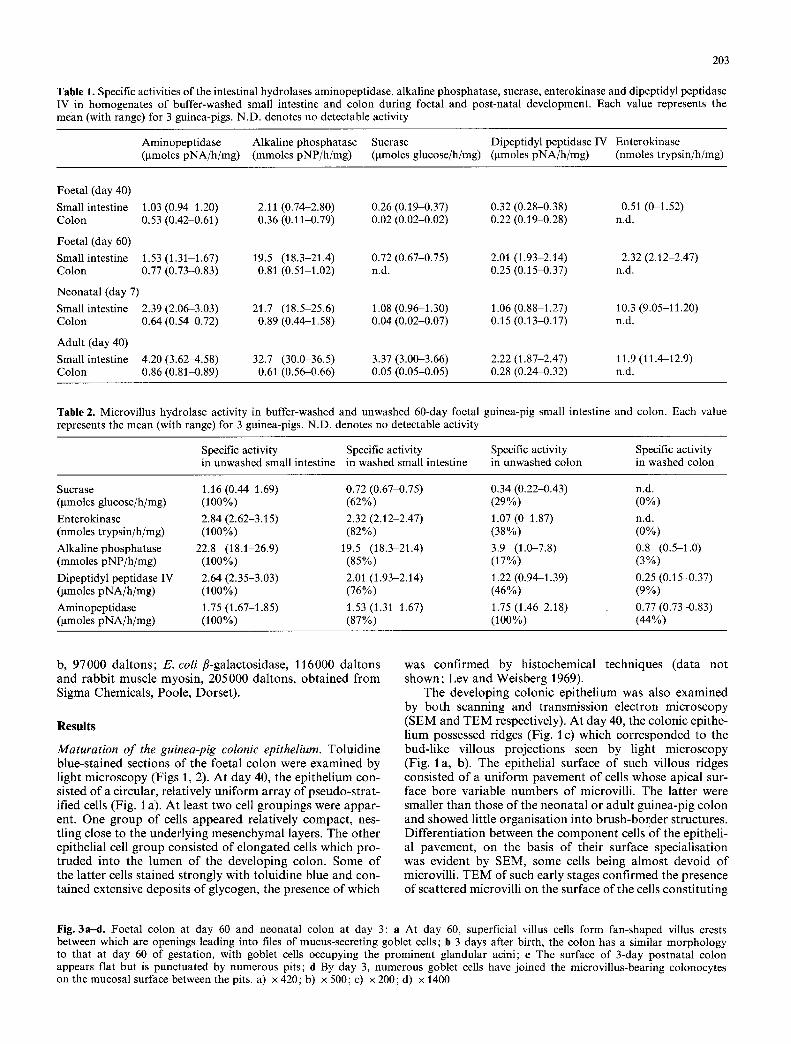
Fig. 2a-e. Foetal colon, day 55: a By day 55, the epithelium forms distinct villi composed of toluidine blue-stained cells. The tips of adjacent villi may be fused (arrows), enclosing an intervillous space which appears distinct from the main intestinal lumen (L); b The surface of the epithelium has become highly contorted with 'knots ' of interconnecting luminally-positioned colonocytcs; c Basal regions of the crypt-villus unit (C) show signs of cytodifferentiation. Mucus- secreting goblet cells (cell 4) appear with glycogen-rich colonocytes (cells 7-9). Some cells show signs of recent division (cells 1 and 2). Luminal microvilli are scarce; d Colonic villi populated with apical cells distended with glycogen. At lower positions on the villus more compact cells are found some of which are secreting mucus (arrows); e Detail of epithelial surface from the basal region of the villus showing a goblet cell in the process of disgorging mucus. Mucus cells also possess microvilli, seen still adhering to the mucus droplets, a) x 65; b) x l 3 0 ; c ) x 670;d) x440 ;e ) x 2600
202
203
Table l. Specific activities of the intestinal hydrolases aminopeptidase, alkaline phosphatase, sucrase, enterokinase and dipeptidyl peptidase IV in homogenates of buffer-washed small intestine and colon during foetal and post-natal development. Each value represents the mean (with range) for 3 guinea-pigs. N.D. denotes no detectable activity
Aminopeptidase Alkaline phosphatase Sucrase Dipeptidyl peptidase IV Enterokinase (p-moles pNA/h/mg) (mmoles pNP/h/mg) (p-moles glucose/h/mg) (p.moles pNA/h/mg) (nmoles trypsin/h/rag)
Foetal (day 40) Small intestine 1.03 (0.94-1.20) Colon 0.53 (0.42-0.61)
Foetal (day 60) Small intestine 1.53 (1.31-1.67) Colon 0.77 (0.73-0.83)
2.11 (0.74-2.80) 0.26 (0.19-0.37) 0.32 (0.28-0.38) 0.51 (0-1.52) 0.36 (0.11-0.79) 0.02 (0.02-0.02) 0.22 (0.19-0.28) n.d.
19.5 (18.3-21.4) 0.72 (0.67-0.75) 2.01 (1.93-2.14) 2.32 (2.12-2.47) 0.81 (0.51-1.02) n.d. 0.25 (0.15~0.37) n.d.
Neonatal (day 7) Small intestine 2.39 (2.06-3.03) 21.7 (18.5-25.6) 1.08 (0.96-1.30) 1.06 (0.88-1.27) 10.3 (9.05-11.20) Colon 0.64 (0.54-0.72) 0.89 (0.44-1.58) 0.04 (0.02-0.07) 0.15 (0.13-0.17) n.d.
Adult (day 40) Small intestine 4.20 (3.62-4.58) 32.7 (30.0 36.5) 3.37 (3.00-3.66) 2.22 (1.822.47) 11.9 (11.4-12.9) Colon 0.86 (0.81-0.89) 0.61 (0.56-0.66) 0.05 (0.05-0.05) 0.28 (0.24-0.32) n.d.
Table 2. Microvillus hydrolase activity in buffer-washed and unwashed 60-day foetal guinea-pig small intestine and colon. Each value represents the mean (with range) for 3 guinea-pigs. N.D. denotes no detectable activity
Specific activity Specific activity Specific activity Specific activity in unwashed small intestine in washed small intestine in unwashed colon in washed colon
Sucrase 1.16 (0.44-1.69) 0.72 (0.67-0.75) 0.34 (0.22-0.43) n.d. (p.moles glucose/h/mg) (100%) (62%) (29%) (0%) Enterokinase 2.84 (2.62-3.15) 2.32 (2.12-2.47) 1.07 (0-1.87) n.d. (nmoles trypsin/h/mg) (100%) (82%) (38%) (0%) Alkaline phosphatase 22.8 (18.1-26.9) 19.5 (18.3-21.4) 3.9 (1.0-7.8) 0.8 (0.5-1.0) (mmoles pNP/h/mg) (100%) (85%) (17%) (3%) Dipeptidyl peptidase IV 2.64 (2.35-3.03) 2.01 (1.93-2.14) 1.22 (0.94-1.39) 0.25 (0.15-0.37) (p-moles pNA/h/mg) (100%) (76%) (46%) (9%) Aminopeptidase 1.75 (1.6%1.85) 1.53 (1.31-1.67) 1.75 (1.46-2.18) 0.77 (0.73-0.83) (p-moles pNA/h/mg) (100%) (87%) (100%) (44%)
b, 97000 daltons; E. coli fl-galactosidase, 116000 daltons and rabbit muscle myosin, 205000 daltons, obtained from Sigma Chemicals, Poole, Dorset).
Results
Maturation of the guinea-pig colonic epithelium. Toluidine blue-stained sections of the foetal colon were examined by light microscopy (Figs 1, 2). At day 40, the epithelium con- sisted of a circular, relatively uniform array of pseudo-strat- ified cells (Fig. 1 a). At least two cell groupings were appar- ent. One group of cells appeared relatively compact, nes- tling close to the underlying mesenchymal layers. The other epithelial cell group consisted of elongated cells which pro- truded into the lumen of the developing colon. Some of the latter cells stained strongly with toluidine blue and con- tained extensive deposits of glycogen, the presence of which
was confirmed by histochemical techniques (data not shown; Lev and Weisberg 1969).
The developing colonic epithelium was also examined by both scanning and transmission electron microscopy (SEM and TEM respectively). At day 40, the colonic epithe- lium possessed ridges (Fig. 1 c) which corresponded to the bud-like villous projections seen by light microscopy (Fig. 1 a, b). The epithelial surface of such villous ridges consisted of a uniform pavement of cells whose apical sur- face bore variable numbers of microvilli. The latter were smaller than those of the neonatal or adult guinea-pig colon and showed little organisation into brush-border structures. Differentiation between the component cells Of the epitheli- al pavement, on the basis of their surface specialisation was evident by SEM, some cells being almost devoid o f microvilli. TEM of such early stages confirmed the presence of scattered microvilli on the surface of the cells constituting
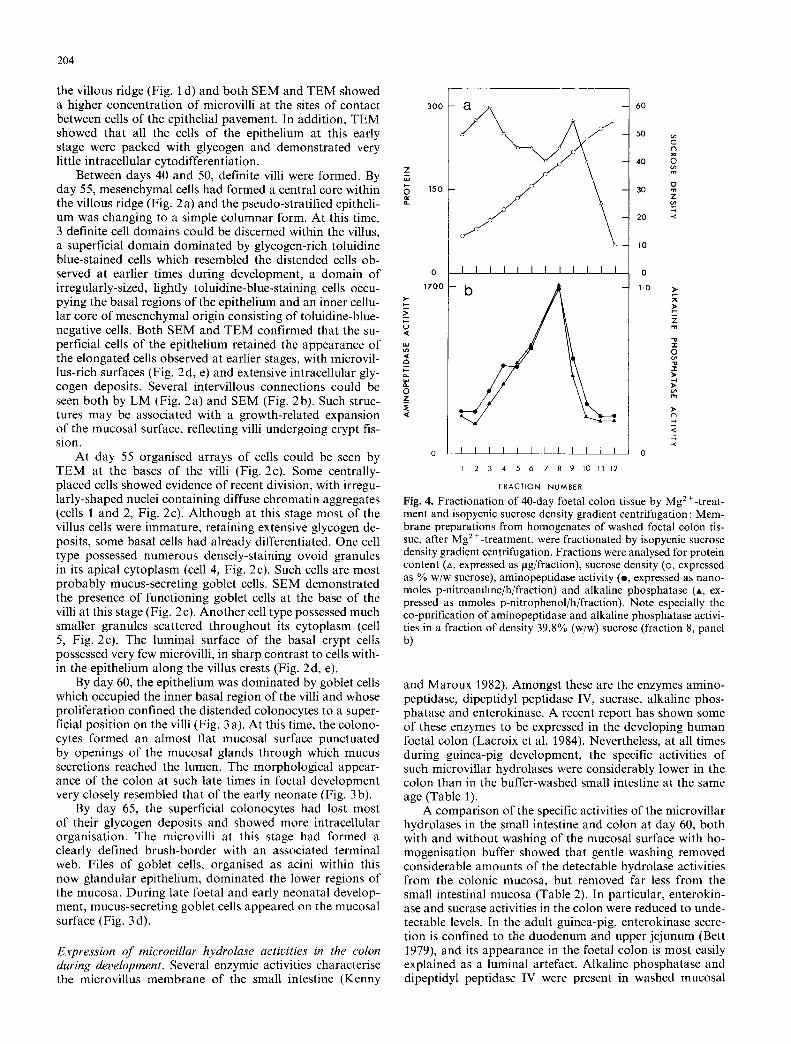
Fig. 3a-d. Foetal colon at day 60 and neonatal colon at day 3: a At day 60, superficial villus cells form fan-shaped villus crests between which are openings leading into files of mucus-secreting goblet cells; b 3 days after birth, the colon has a similar morphology to that at day 60 of gestation, with goblet cells occupying the prominent glandular acini; e The surface of 3-day postnatal colon appears flat but is punctuated by numerous pits; d By day 3, numerous goblet cells have joined the microvillus-bearing colonocytes on the mucosal surface between the pits. a) x 420; b) x 500; c) • 200; d) x 1400
204
the villous ridge (Fig. 1 d) and both SEM and TEM showed a higher concentration o f microvilli at the sites of contact between cells of the epithelial pavement. In addition, TEM showed that all the cells o f the epithelium at this early stage were packed with glycogen and demonstrated very little intracellular cytodifferentiation.
Between days 40 and 50, definite villi were formed. By day 55, mesenchymal cells had formed a central core within the villous ridge (Fig. 2 a) and the pseudo-stratified epitheli- um was changing to a simple columnar form. At this time, 3 definite cell domains could be discerned within the villus, a superficial domain dominated by glycogen-rich toluidine blue-stained cells which resembled the distended cells ob- served at earlier times during development, a domain of irregularly-sized, lightly toluidine-blue-staining cells occu- pying the basal regions of the epithelium and an inner cellu- lar core of mesenchymal origin consisting of toluidine-blue- negative cells. Both SEM and TEM confirmed that the su- perficial cells of the epithelium retained the appearance of the elongated cells observed at earlier stages, with microvil- lus-rich surfaces (Fig. 2d, e) and extensive intracellular gly- cogen deposits. Several intervillous connections could be seen both by LM (Fig. 2a) and SEM (Fig. 2b). Such struc- tures may be associated with a growth-related expansion of the mucosal surface, reflecting villi undergoing crypt fis- sion.
At day 55 organised arrays of cells could be seen by TEM at the bases o f the villi (Fig. 2c). Some centrally- placed cells showed evidence of recent division, with irregu- larly-shaped nuclei containing diffuse chromatin aggregates (cells 1 and 2, Fig. 2c). Although at this stage most of the villus cells were immature, retaining extensive glycogen de- posits, some basal cells had already differentiated. One cell type possessed numerous densely-staining ovoid granules in its apical cytoplasm (cell 4, Fig. 2c). Such cells are most probably mucus-secreting goblet cells. SEM demonstrated the presence of functioning goblet cells at the base of the villi at this stage (Fig. 2e). Another cell type possessed much smaller granules scattered throughout its cytoplasm (cell 5, Fig. 2c). The luminal surface of the basal crypt cells possessed very few microvilli, in sharp contrast to cells with- in the epithelium along the villus crests (Fig. 2d, e).
By day 60, the epithelium was dominated by goblet cells which occupied the inner basal region of the villi and whose proliferation confined the distended colonocytes to a super- ficial position on the villi (Fig. 3 a). At this time, the colono- cytes formed an almost flat mucosal surface punctuated by openings of the mucosal glands through which mucus secretions reached the lumen. The morphological appear- ance of the colon at such late times in foetal development very closely resembled that of the early neonate (Fig. 3 b).
By day 65, the superficial colonocytes had lost most of their glycogen deposits and showed more intracellular organisation. The microvilli at this stage had formed a clearly defined brush-border with an associated terminal web. Files of goblet cells, organised as acini within this now glandular epithelium, dominated the lower regions of the mucosa. During late foetal and early neonatal develop- ment, mucus-secreting goblet cells appeared on the mucosal surface (Fig. 3 d).
Expression of microvillar hydrolase activities in the colon during development. Several enzymic activities characterise the microvillus membrane of the small intestine (Kenny
3 0 0
Z
150
a,.
1 7 0 0
w
- a ,
I I I t I I I I 11 I I
I I I I I I I I I I I I I 2 3 4 5 6 7 8 9 10 11 12
FRACTION NUMBER
- - 6 O
~o
40 ~ m
3o ~ Z
--i 2 0 <
1 0
0
- - 1.0 >
<
---i -<
0
Fig. 4. Fractionation of 40-day foetal colon tissue by Mg 2 +-treat- ment and isopycnic sucrose density gradient centrifugation: Mem- brane preparations from homogenates of washed foetal colon tis- sue, after Mg z +-treatment, were fractionated by isopycnic sucrose density gradient centrifugation. Fractions were analysed for protein content (A, expressed as gg/fraction), sucrose density (o, expressed as % w/w sucrose), aminopeptidase activity (e, expressed as nano- moles p-nitroaniline/h/fraction) and alkaline phosphatase (A, ex- pressed as mmoles p-nitrophenol/h/fraction). Note especially the co-purification of aminopeptidase and alkaline phosphatase activi- ties in a fraction of density 39.8% (w/w) sucrose (fraction 8, panel b)
and Maroux 1982). Amongst these are the enzymes amino- peptidase, dipeptidyl peptidase IV, sucrase, alkaline phos- phatase and enterokinase. A recent report has shown some of these enzymes to be expressed in the developing human foetal colon (Lacroix et al. 1984). Nevertheless, at all times during guinea-pig development, the specific activities of such microvillar hydrolases were considerably lower in the colon than in the buffer-washed small intestine at the same age (Table 1).
A comparison of the specific activities of the microvillar hydrolases in the small intestine and colon at day 60, both with and without washing of the mucosal surface with ho- mogenisation buffer showed that gentle washing removed considerable amounts of the detectable hydrolase activities from the colonic mucosa, but removed far less from the small intestinal mucosa (Table 2). In particular, enterokin- ase and sucrase activities in the colon were reduced to unde- tectable levels. In the adult guinea-pig, enterokinase secre- tion is confined to the duodenum and upper jejunum (Bett 1979), and its appearance in the foetal colon is most easily explained as a luminal artefact. Alkaline phosphatase and dipeptidyl peptidase IV were present in washed mucosal
a b c d e f
205
210,- 2 0 0 "
-,, 2 0 5
-.127
-, 8 0
62,-
~24g -. 47
40,-
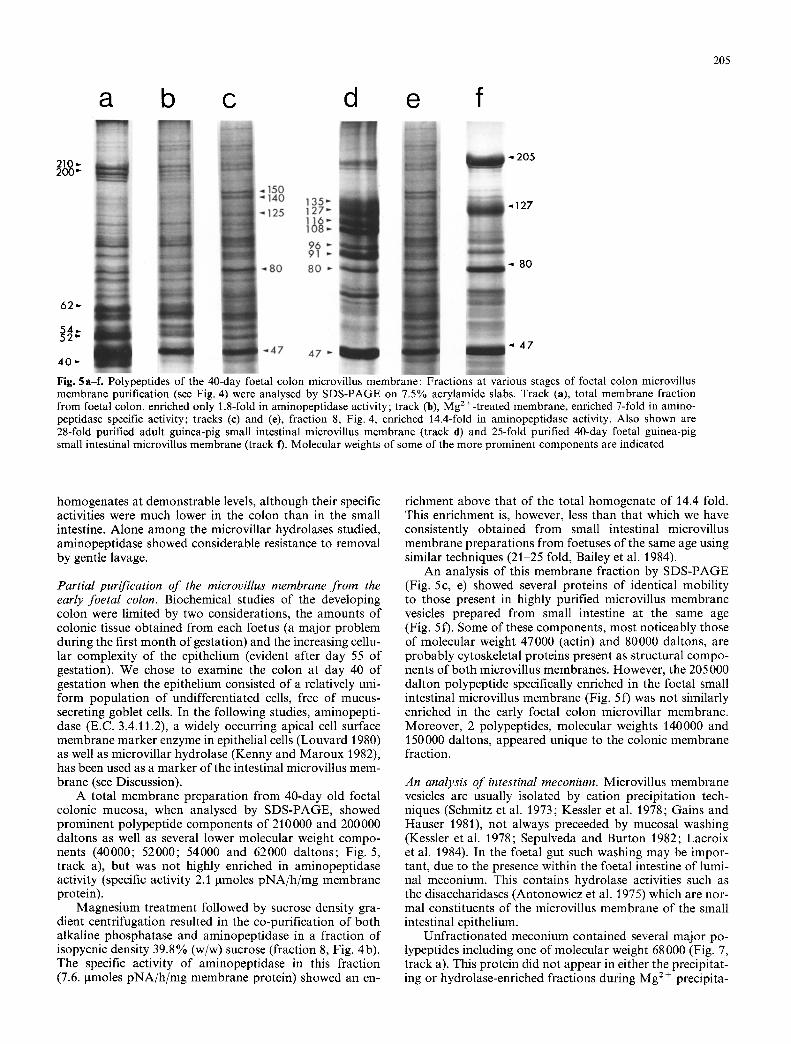
Fig. 5a-f. Polypeptides of the 40-day foetal colon microvillus membrane: Fractions at various stages of foetal colon microvillus membrane purification (see Fig. 4) were analysed by SDS-PAGE on 7.5% acrylamide slabs. Track (a), total membrane fraction from foetal colon, enriched only 1.8-fold in aminopeptidase activity; track (b), Mg 2 +-treated membrane, enriched 7-fold in amino- peptidase specific activity; tracks (e) and (e), fraction 8, Fig. 4, enriched 14.4-fold in aminopeptidase activity. Also shown are 28-fold purified adult guinea-pig small intestinal microvillus membrane (track d) and 25-fold purified 40-day foetal guinea-pig small intestinal microvillus membrane (track f). Molecular weights of some of the more prominent components are indicated
homogenates at demonstrable levels, although their specific activities were much lower in the colon than in the small intestine. Alone among the microvillar hydrolases studied, aminopeptidase showed considerable resistance to removal by gentle lavage.
Partial purification of the microvillus membrane from the early foetal colon. Biochemical studies of the developing colon were limited by two considerations, the amounts of colonic tissue obtained from each foetus (a major problem during the first month of gestation) and the increasing cellu- lar complexity of the epithelium (evident after day 55 of gestation). We chose to examine the colon at day 40 of gestation when the epithelium consisted of a relatively uni- form population of undifferentiated cells, free of mucus- secreting goblet cells. In the following studies, aminopepti- dase (E.C. 3.4.11.2), a widely occurring apical cell surface membrane marker enzyme in epithelial ceils (Louvard 1980) as well as microvillar hydrolase (Kenny and Maroux 1982), has been used as a marker of the intestinal microvillus mem- brane (see Discussion).
A total membrane preparation from 40-day old foetal colonic mucosa, when analysed by SDS-PAGE, showed prominent polypeptide components of 210000 and 200000 daltons as well as several lower molecular weight compo- nents (40000; 52000; 54000 and 62000 daltons; Fig. 5, track a), but was not highly enriched in aminopeptidase activity (specific activity 2.1 lamoles pNA/h/mg membrane protein).
Magnesium treatment followed by sucrose density gra- dient centrifugation resulted in the co-purification of both alkaline phosphatase and aminopeptidase in a fraction of isopycnic density 39.8% (w/w) sucrose (fraction 8, Fig. 4b). The specific activity of aminopeptidase in this fraction (7.6. txmoles pNA/h/mg membrane protein) showed an en-
richment above that of the total homogenate of 14.4 fold. This enrichment is, however, less than that which we have consistently obtained from small intestinal microvillus membrane preparations from foetuses of the same age using similar techniques (21-25 fold, Bailey et al. 1984).
An analysis of this membrane fraction by SDS-PAGE (Fig. 5c, e) showed several proteins of identical mobility to those present in highly purified microvillus membrane vesicles prepared from small intestine at the same age (Fig. 5 f). Some of these components, most noticeably those of molecular weight 47000 (actin) and 80000 daltons, are probably cytoskeletal proteins present as structural compo- nents of both microvillus membranes. However, the 205 000 dalton polypeptide specifically enriched in the foetal small intestinal microvillus membrane (Fig. 5 f) was not similarly enriched in the early foetal colon microvillar membrane. Moreover, 2 polypeptides, molecular weights 140000 and 150000 daltons, appeared unique to the colonic membrane fraction.
An analysis of intestinal meconium. Microvillus membrane vesicles are usually isolated by cation precipitation tech- niques (Schmitz et al. 1973; Kessler et al. 1978; Gains and Hauser 1981), not always preceeded by mucosal washing (Kessler et al. 1978; Sepulveda and Burton 1982; Lacroix et al. 1984). In the foetal gut such washing may be impor- tant, due to the presence within the foetal intestine of lumi- nal meconium. This contains hydrolase activities such as the disaccharidases (Antonowicz et al. 1975) which are nor- mal constituents of the microvillus membrane of the small intestinal epithelium.
Unfractionated meconium contained several major po- lypeptides including one of molecular weight 68 000 (Fig. 7, track a). This protein did not appear in either the precipitat- ing or hydrolase-enriched fractions during Mg 2+ precipita-
206
100 - 6 0
- 5 0 c n
-
I . --
U
< 4 0 m
~_ 3 0 Z
0 z 2 0
O - 10
I I I I I I I [ [ I I [ I
I 2 3 4 5 6 7 8 9 10 11 12 13
FRACTION NUMBER
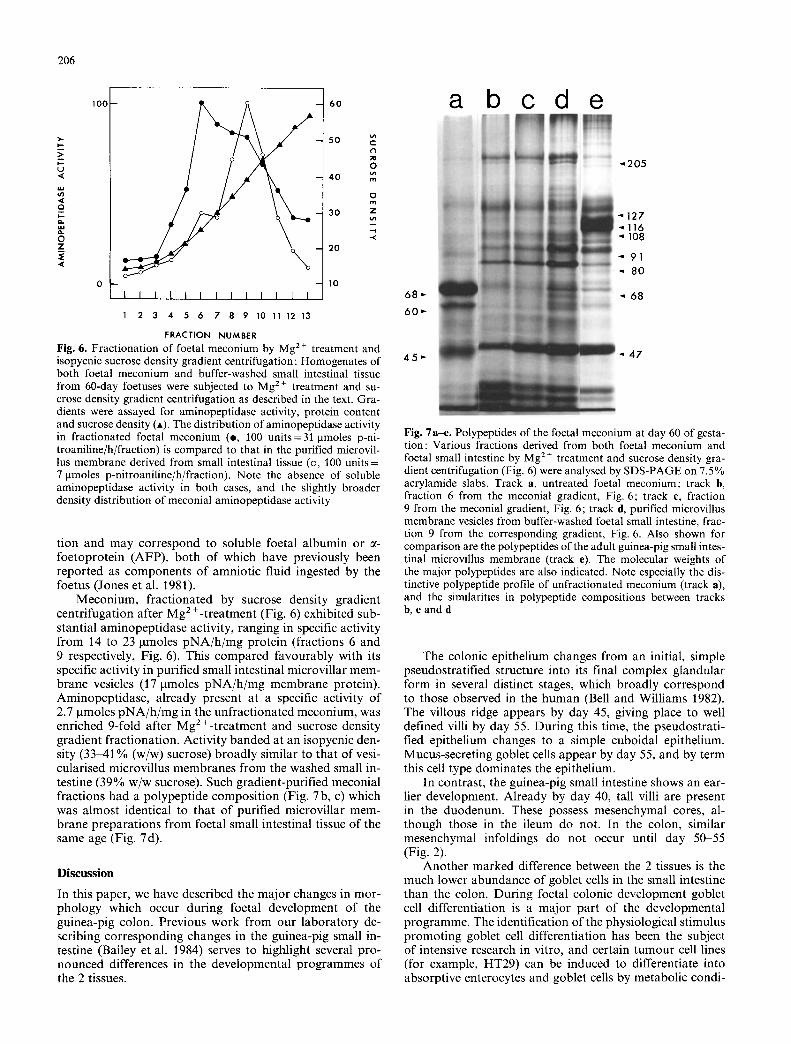
Fig. 6. Fractionation of foetal meconium by Mg 2 + treatment and isopycnic sucrose density gradient centrifugation : Homogenates of both foetal meconium and buffer-washed small intestinal tissue from 60-day foetuses were subjected to Mg 2+ treatment and su- crose density gradient centrifugation as described in the text. Gra- dients were assayed for aminopeptidase activity, protein content and sucrose density (A). The distribution of aminopeptidase activity in fractionated foetal meconium (e, 100 units=31 gmoles p-ni- troaniline/h/fraction) is compared to that in the purified microvil- lus membrane derived from small intestinal tissue (o, 100 units = 7 gmoles p-nitroaniline/h/fraction). Note the absence of soluble aminopeptidase activity in both cases, and the slightly broader density distribution of meconial aminopeptidase activity
tion and may correspond to soluble foetal albumin or ~- foetoprotein (AFP), both of which have previously been reported as components of amniotic fluid ingested by the foetus (Jones et al. 1981).
Meconium, fractionated by sucrose density gradient centrifugation after Mg 2+-treatment (Fig. 6) exhibited sub- stantial aminopeptidase activity, ranging in specific activity from 14 to 23 gmoles pNA/h /mg protein (fractions 6 and 9 respectively, Fig. 6). This compared favourably with its specific activity in purified small intestinal microvillar mem- brane vesicles (17 gmoles pNA/h /mg membrane protein). Aminopeptidase, already present at a specific activity of 2.7 gmoles pNA/h /mg in the unfractionated meconium, was enriched 9-fold after Mg 2+-treatment and sucrose density gradient fractionation. Activity banded at an isopycnic den- sity (33-41% (w/w) sucrose) broadly similar to that of vesi- cularised microvillus membranes from the washed small in- testine (39% w/w sucrose). Such gradient-purified meconial fractions had a polypeptide composition (Fig. 7b, c) which was almost identical to that of purified microvillar mem- brane preparations from foetal small intestinal tissue of the same age (Fig. 7d).
Discussion
In this paper, we have described the major changes in mor- phology which occur during foetal development of the guinea-pig colon. Previous work from our laboratory de- scribing corresponding changes in the guinea-pig small in- testine (Bailey et al. 1984) serves to highlight several pro- nounced differences in the developmental programmes of the 2 tissues.
6 8 " "
6 0 "
a b c d e
",, 2 0 5
" 1 2 7 " "116 " 108
" 9 1
~, 8 0
~, 6 8
45,- -. 47
Fig. 7a-e. Polypeptides of the foetal meconium at day 60 of gesta- tion: Various fractions derived from both foetal meconium and foetal small intestine by Mg 2§ treatment and sucrose density gra- dient centrifugation (Fig. 6) were analysed by SDS-PAGE on 7.5 % acrylamide slabs. Track a, untreated foetal meconium; track b, fraction 6 from the meconial gradient, Fig. 6; track c, fraction 9 from the meconial gradient, Fig. 6; track d, purified microvillus membrane vesicles from buffer-washed foetal small intestine, frac- tion 9 from the corresponding gradient, Fig. 6. Also shown for comparison are the polypeptides of the adult guinea-pig small intes- tinal microvillus membrane (track e). The molecular weights of the major polypeptides are also indicated. Note especially the dis- tinctive polypeptide profile of unfractionated meconium (track a), and the similarities in polypeptide compositions between tracks b, c and d
The colonic epithelium changes from an initial, simple pseudostratified structure into its final complex glandular form in several distinct stages, which broadly correspond to those observed in the human (Bell and Williams 1982). The villous ridge appears by day 45, giving place to well defined villi by day 55. During this time, the pseudostrati- fled epithelium changes to a simple cuboidal epithelium. Mucus-secreting goblet cells appear by day 55, and by term this cell type dominates the epithelium.
In contrast, the guinea-pig small intestine shows an ear- lier development. Already by day 40, tall villi are present in the duodenum. These possess mesenchymal cores, al- though those in the ileum do not. In the colon, similar mesenchymal infoldings do not occur until day 50-55 (Fig. 2).
Another marked difference between the 2 tissues is the much lower abundance of goblet cells in the small intestine than the colon. During foetal colonic development goblet cell differentiation is a major part of the developmental programme. The identification of the physiological stimulus promoting goblet cell differentiation has been the subject of intensive research in vitro, and certain tumour cell lines (for example, HT29) can be induced to differentiate into absorptive enterocytes and goblet cells by metabolic condi-

207
tioning (Pinto et al. 1982). Such mechanisms may also oper- ate at a developmental level.
The morphology of the adult colon and small intestine differ in that the adult colon does not posses villi. However, we have observed villus formation in the foetal colon, as have several other workers (Johnson 1913; Bell and Wil- liams 1982; Colony and Neutra 1983). A point which re- quires further clarification is the generation of the final adult morphology, during the process of which separate villi are converted into a flat mucosal floor, punctuated by intramucosal pits or crypts (Fig. 3 c).
Our studies using SEM parallel those of other workers. The clinical use of SEM to investigate human colonic dis- ease, recently reviewed by Siew (1983), has demonstrated an inverse correlation between cell surface microvillus den- sity and colonic malignancy (Cooke et al. 1984). Our studies have clearly shown a lower incidence of luminal microvilli at early times during development of the foetal guinea-pig colon. Indeed, variation in the concentration of surface mi- crovilli may occur between adjacent cells within the epitheli- um, presumably reflecting varying degrees of differentiation between neighbouring cells. SEM represents a powerful tool with which to examine such changes in cell surface mor- phology.
Our attempts to isolate the foetal colonic microvillus membrane, using methods which we have successfully used to isolate that of the foetal small intestine, have been com- plicated by the presence of colonic meconium and the ab- sence of demonstrable marker enzymes. In the latter re- spect, the foetal guinea-pig colon resembled the adult guinea-pig colon (Biber et al. 1983).
It was clear from our early experiments that consider- able luminal activities of the microvillar hydrolases were present in the foetal colon, but that most were effectively removed by gentle washing with homogenisation buffer which did not disrupt the surface of the colonocytes. The luminal material is meconium, some of which passes from the small intestine into the colon by peristalsis and has a yellow colour due to bile pigments secreted from the de- veloping liver. Inspection of the gut at various stages during development showed a progressive movement of the yellow meconium through the small intestine towards the colon. By day 60 of gestation, the colon contained substantial amounts of meconium which was difficult to remove entire- ly from the mucosal surface.
Preliminary analysis of unfractionated foetal meconium by 1-dimensional SDS-PAGE showed the presence of large amounts of a 68000 dalton polypeptide (Fig. 6a), either AFP or foetal albumin. These proteins are major compo- nents of amniotic fluid (Jones et al. 1979). We have not yet pursued a full characterisation of meconium by 2-di- mensional SDS-PAGE, one way to unequivocally distin- guish between AFP and foetal albumin. It would be inter- esting to know whether amniotic fluid is passing through the foetal intestine at early stages of physiological develop- ment since human amniotic fluid is known to contain sever- al hormones which are trophic for intestinal maturation, including steroids (Homoki et al. 1983), insulin (Casper and Benjamin i980; Spellacy et al. 1973), and epidermal growth factor (Dailey et al. 1978). Furthermore, extracts from rat amniotic fluid have been reported to induce intestinal crypt formation (Calvert et al. 1983).
In addition to soluble components such as AFP/albu- min, we have isolated a particulate fraction from meconium
which is both enriched in microvillus hydrolase activities and which has an almost identical polypeptide composition to that of the foetal small intestinal microvillus membrane at the same age (Fig. 7). This suggests the presence of lumi- nal microvillar fragments which may travel through the developing intestinal tract at the onset of peristalsis. These undoubtedly would pose a problem in studies of microvillus membrane differentiation in the colon, since the latter mem- brane may be contaminated by adsorbed polypeptides de- rived from the small intestinal microvillus membrane, trans- ported into the colon via the meconium. Even sophisticated techniques such as indirect immunodetection to identify antigenic determinants expressed at the microvillus mem- brane (Zweibaum et al. 1983) would be susceptible to misin- terpretation if meconial components bound extensively to the colonic membrane. In fact, if this were true, the only reliable way to identify endogenous microvillar components in the colonic epithelium would be by biosynthetic studies in segments of foetal colon isolated from intestinal contin- uity.
We chose to examine day 40 of colonic development in order to minimise meconial adsorption problems, making the assumption that peristalsis was unlikely to occur in the absence of well defined muscle layers (Fig. 1). Microvillar membrane preparation, using methods which are reportedly successful for the human foetal colon (purification factor not reported, Lacroix et al. 1984) proved much less success- ful than we had expected, purifying the putative colonic membrane only 14 fold above the homogenate on the basis of aminopeptidase activity enrichment. Exactly similar methods using small intestinal tissue of the same age have proved highly successful in our hands, resulting in 21-25 fold purification of this membrane (Bailey et al. 1984). We know that microvilli are indeed quite abundant in the colon at day 40, suggesting that the cause for this relatively poor purification must reside in the methodology employed. Ca- tion precipitation methods rely on the presence of negative- ly-charged side-chains expressed at the membrane surface (Booth and Kenny 1974). I f the colonic microvillus mem- brane does not possess such a high concentration of sialic- acid rich glycoproteins at the membrane surface, such ca- tion-precipitation methods for microvillus membrane prep- aration from the colon may not be as effective as they are for preparing the microvillus membrane of the small intestine.
In our preparation, we noticed the copurification of aminopeptidase and alkaline phosphatase in a membrane fraction which may correspond to the foetal colonic micro- villus membrane (Fig. 4). In many species, alkaline phos- phatase is a marker for the foetal colonic microvillus mem- brane (Lev and Orlic 1974; Colony and Neutra 1983). Aminopeptidase has also been reported as a marker for the apical plasmamembrane domain of several epithelial cells (discussed by Louvard 1980). Moreover, in our experi- ments, aminopeptidase was to a large extent retained by the colon after gentle washing (Table 2), suggesting that this enzyme may be a true component of the epithelium. However, its presence in the foetal colonic microvillus mem- brane has not yet been demonstrated using histochemical techniques.
Analyses of the colonic microvillus membrane fraction and the microvillus membrane of the small intestine at day 40 by SDS-PAGE (Fig. 5) illustrated basic similarities in the structure of the 2 intestinal microvillus membranes, but

208
also pointed to specific differences in polypeptide composi- tion. Fur ther characterisation of the component polypep- tides of the colonic microvillar membrane awaits its purifi- cation to homogeneity.
In conclusion, our results have illustrated the develop- ment of the foetal guinea-pig colon at histological, enzymic and molecular levels. The observed developmental changes, when compared with those occurring in the small intestine at the same age, highlight the early structural and functional divergence of these 2 regions of the gastrointestinal tract.
Acknowledgements. We would like to thank Prof. J. Hermon-Tay- lor for his advice and encouragement during this work. Financial support was provided by the Wellcome Trust, the Nuffield Founda- tion and the Cancer Research Campaign of Great Britain.
References
Antonowicz I, Ishida S, Shwachman H (1975) Studies in meconi- urn: disaccharidase activities in meconium from cystic fibrosis patients and controls. Pediatrics 56:782-787
Bailey DS, Cook A, McAIlister G, Moss M, Mian N (1984) Struc- tural and biochemical differentiation of the mammalian small intestine during development. J Cell Sci 72:195-212
Baratti J, Maroux S, Louvard D (1973) Effect of ionic strength and calcium ions on the activation of trypsinogen by enterokin- ase: a modified test for the quantitative evaluation of this en- zyme. Biochim Biophys Acta 321:632-638
Bell L, Williams L (1982) A scanning and transmission electron microscopical study of the morphogenesis of human colonic villi. Anat Embryol 165 : 437-455
Bett NJ (1979) Regulation of enterokinase synthesis in animal and human small intestine by luminal signals: its implication in upper gastrointestinal surgery. Br J Surg 66 : 708-711
Biber J, Rechkemmer G, Bodmer M, Schroder P, Haase W, Murer H (1983) Isolation of basolateral membranes from columnar cells of the proximal colon of the guinea-pig. Biochim Biophys Acta 735 : 1-11
Booth AG, Kenny AJ (1974) A rapid method for the preparation of microvilli from rabbit kidney. Biochem J 142:575-581
Brackett KA, Townsend SF (1980) Organogenesis of the colon in rats. J Morphol 163 : 191-201
Calvert R, Lehoux J-G, Arsenault P, Menard D (1983) Extracts of rat amniotic fluid contain a potent inducer of intestinal crypt formation. Anat Rec 205 : 27-37
Casper D J, Benjamin F (1980) Immunoreactive insulin in amniotic fluid. Obstet Gynecol 35 : 389-393
Colony PC, Neutra MR (1983) Epithelial differentiation in the fetal rat colon: plasma membrane phosphatase activities. Dev Biol 97 : 349 363
Cooke T, Kirkham N, Stainthorp DH, Inman C, Goeting N, Tay- lor I (1984) Detection of early neoplastic changes in experimen- tally induced colorectal cancer using scanning electron micros- copy and cell kinetic studies. Gut 25:748 755
Dahlqvist A (1968) Assay of intestinal disaccharidases. Anal Bio- chem 22:99-107
Dailey GE, Kraus JW, Orth DN (1978) Homologous radioimmu- noassay for human epidermal growth factor (urogastrone). J Clin Endocrinol Metab 46:929-936
EngstrSm L (1961) Studies on calf intestinal alkaline phosphatase. I. Chromatographic purification, microheterogeneity and some other properties of the purified enzyme. Biochim Biophys Acta 52 : 36M8
Gains N, Hauser H (1981) Detergent-induced proteolysis of rabbit intestinal brush-border vesicles. Biochim Biophys Acta 646:211-217
Goldenberg DM (1976) Oncofetal and other tumour-associated antigens of the human digestive system. In: Morson BC (ed) Current Topics in Pathology 63. Springer, Berlin pp 289-342
Graham J, Ford TC (1983) Enzymic and chemical assays compati- ble with iodinated density gradient media. In: Rickwood D (ed) Iodinated density gradient media: a practical approach. IRL Press, Oxford. pp 195-216
Grand RJ, Watkins JB, Torti FM (1976) Development of the hu- man gastrointestinal tract. A review. Gastroenterology 70:790-810
Helander HR (1973) Morphological studies on the development of the rat colonic mucosa. Acta Anat 85:153-176
Helander HF (1975) Enzyme patterns and protein absorption in rat colon during development. Acta Anat 91:330~349
Homoki J, Roitman E, Shackleton CHL (1983) Characterisation of the major steroids present in amniotic fluid obtained between the 15th and 17th weeks of gestation. J Steroid Biochem 19:1061 1068
Johnson FK (1913) The development of the mucous membrane of the large intestine and vermiform process of the human em- bryo. Am J Anat 14:187-233
Jones MI, Spragg SP, Webb T (1981) Detection of proteins in human amniotic fluid using two-dimensional gel electrophore- sis. Biol Neonate 39:171-t77
Kenny A J, Maroux S (1982) Topology of microvillar membrane hydrolases of kidney and intestine. Physiol Rev 62:91-128
Kessler M, Acuto O, Storelli C, Murer H, Muller M, Semenza G (1978) A modified procedure for the rapid preparation of efficiently-transporting vesicles from small intestinal brush- border membranes. Biochim Biophys Acta 506:136-154
Lacroix B, Kedinger M, Simon-Assmann P, Rousset M, Zweibaum A, Haffen K (1984) Developmental pattern of brush-border enzymes in the human fetal colon. Correlation with some mor- phogenetic events. Early Hum Dev 9:95 103
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227 : 680-685
Lev R, Orlic D (1974) Histochemical and radioautographic studies of normal human foetal colon. Histochemistry 39:301-311
Lev R, Weisberg H (1969) Human foetal epithelial glycogen: a histochemical and electronmicroscopic study. J Anat 105:337 349
Lim S-S, Low FN (1977) Scanning electron microscopy of the developing alimentary canal in the chick. Am J Anat 150:149-174
Louvard D (1980) Apical membrane aminopeptidase appears at site of cell-cell contact in cultured kidney epithelial cells. Proc Natl Acad Sci 77:4132-4136
Lowry OH, Rosebrough N J, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265-275
Moog F, Ortiz E (I 960) The functional differentiation of the small intestine VII. The duodenum of the foetal guinea-pig, with a note on the growth of the adrenals. J Embryol Exp Morphol 8:182-194
Nagatsu T, lwase K, Kasahara Y, Kubono K, Sakakibara S, Aoyama H, Izawa Y (1979) Correlation between X-prolyl di- peptidyl aminopeptidase and serum amine oxidase in serum of patients with post-burn keloids. Clin Chem 25 : 376-378
Pinto M, Appay M-D, Simon-Assmann P, Chevalier G, Dracapoli N, Fogh J, Zweibaum A (t982) Enterocytic differentiation of cultured human colon cancer ceils by replacement of glucose by galactose in the medium. Biol Cell 44:193-196
Richardson KC, Jarrett L, Finke EH (1960) Embedding in epoxy resins for ultrathin sectioning in electronmicroscopy. Stain Technol 35 : 313-323
Rousset M, Dussaulx E, Chevalier G, Zweibaum A (1980) Growth- related glycogen levels of human intestine carcinoma cell lines grown in vitro and in vivo in nude mice. J Nat Cancer Inst 65 : 885-889
Rousset M, Robine-Leon S, Dussaulx E, Chevalier G, Zweibaum A (1979) Glycogen storage in foetal and malignant epithelial cells of the human colon. Front Gastroenterol Res 4:80-85
Sato T (1968) A modified method for lead staining of thin sections. Jpn J Electr Microsc (Tokyo) 17 : 158 159

209
Schmitz J, Preiser H, Maestracci D, Ghosh BK, Cerda J J, Crane RK (1973) Purification of the human intestinal brush-border membrane. Biochim Biophys Acta 323:98-112
Semenza G, Brunner J, Wacker H (1983) Biosynthesis and assem- bly of the largest and major intrinsic polypeptide of the small intestinal brush-border. In: Porter R, Collins GM (eds) Brush- border membranes, CIBA Foundation Symposium 95. Pitman Books, London, pp 92-112
Sepulveda RV, Burton KA (1982) 7-Glutamyl transferase activity in the pig proximal colon during early postnatal development. FEBS Letts 139:171-173
Siew S (1983) The application of scanning electron microscopy in the clinical investigation of the human colon. Scann Electr Microsc IV : 1911-1929
Spellacy WN, Buhi WC, Bradley B, Kolsinger KK (1973) Mater- nal, foetal and amniotic fluid levels of glucose, insulin and growth hormone. Obstet Gynecol 41:323-331
Stempak JG, Ward RT (1964) An improved staining method for electron microscopy. J Cell Biol 22:697-701
Wachsmuth ED, Fritze I, Pfleiderer G (1966) An aminopeptidase occurring in pig kidney. I. An improved method of preparation: physical and enzymic properties. Biochemistry 5 : 169-174
Zweibaum A, Triadou N, Kedinger M, Augeron C, Robine-Leon S, Pinto M, Rousset M, Haffen K (1983) Sucrase-isomaltase: a marker of foetal and malignant epithelial cells of the human colon. Int J Cancer 32:407-412
Accepted March 10, 1985