Embed Size (px)
Citation preview

Page 1 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
Critical review
For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
AbstractIntroductionSex steroid hormones, androgens and oestrogens regulate diverse ph-ysiological processes by interacting with their intracellular protein rece-ptors and modulating the expression of target genes. Classical sex steroid hormone receptors belong to the nu-clear receptor superfamily, which forms the largest known family of tr-anscription factors in eukaryotes. A common feature of sex steroid hor-mone receptors genes is the inciden-ce of alternative splicing, a process that generates multiple variants fro-m a single mRNA precursor molecu-le. This process facilitates the prod-uction of functionally distinct protei-ns and contributes to proteome div-ersity in higher eukaryotes. Herein we will critically review current inf-ormation about the diversity of oest-rogen receptors and androgen rece-ptors variants, discussing their stru-ctural features and physiological functions.ConclusionThe identification of structural varia-nts of both ERs and ARs in healthy testis and a broad range of vertebra-te species, highlights the newly reve-aled complexity of SSHRs signalling mechanisms in this tissue.
Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles
S Laurentino1, P Pinto2, S Correia3, JE Cavaco3, AVM Canário2, S Socorro3*
IntroductionSex steroid hormones, namely andro-gens and oestrogens, regulate multiple reproductive and non-reproductive processes by interacting with their cognate specific receptor protein and regulating the expression of target genes. Classical sex steroid hormone receptors (SSHRs) belong to the nu-clear receptor superfamily, which represents the largest known family of transcription factors in eukaryotes. In addition, SSHRs are present in a variety of species ranging from nem-atodes to humans1.
For several years, the existence of a single oestrogen receptor (ER) was generally accepted. The discovery of a second type of ER (ERβ) in rats, hu-mans and other species2–4 profoundly changed the outlook of ER signalling, stimulating a new wave of research on ERs that was aimed at discovering their specific roles and interactions. While the two ER subtypes (ERα and ERβ) have a typical steroid receptor structure, they are encoded by genes located on different chromosomes5,6, sharing only 41%–65% of overall amino acid identity depending on the species being considered2,4. Never-theless, both receptors exhibit simi-lar binding affinity to 17β-oestradiol and display binding characteristics recognised for ER proteins; specifi-cally, relative binding affinity for natural and synthetic oestrogens in addition to anti-oestrogenic com-pounds is higher than the binding affinity for androgens, progoestro-gens or corticosteroids7. However, selective modulators functioning as agonists or antagonists have been identified for ERα and ERβ proteins8.
In contrast to ERs, only a single g-
ene encoding a receptor for andro-gens has been found in mammals. The human androgen receptor (AR) is a protein with approximately 919 amino acids encoded by a gene lo-cated on the X chromosome9. The precise length of ARs is variable due to the existence of two variable poly-glutamine and polyglycine stretches in the N-terminal region of the pro-tein9, which modulates AR transacti-vation10,11, and thus, its functionality. These stretches have been associated with diseases, such as spinal and bulbar muscular atrophy and prostate can-cer, in addition to male infertility12,13.
A common event described in the structural molecular biology of SSHR genes is the occurrence of alternative splicing14. Alternative splicing is a widespread process in higher eu-karyotes that facilitates the genera-tion of distinct structural mRNA variants from a single gene, which in turn may produce functionally distinct protein isoforms in cell- or tissue-specific contexts15,16. This critical review aims to provide an overview of existing information about alternative splice variants of ERs and ARs in the testis, from their molecular biology to phys-iological roles.
* Corresponding authorEmail: [email protected] CeRA—Centre of Reproductive Medicine and
Andrology, Institute of Reproductive and Regenerative Biology, University Hospital of Muenster, Albert-Schweitzer-Campus 1, Building D11, 48149, Muenster, Germany
2 CCMAR—Centre of Marine Sciences, Universidade do Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
3 CICS-UBI—Health Sciences Research Center, University of Beira Interior, Av. Infante D. Henrique, 6200-506 Covilhã, Portugal
Nucl
eic
Aci
ds &
Mol
ecul
ar
Biol
ogy
DiscussionThe authors have referenced some of their own studies in this review. These referenced studies have been conducted in accordance with the Declaration of Helsinki (1964) and the protocols of these studies have been approved by the relevant ethics committees related to the ins-

For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Page 2 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
Steroid hormone receptor structure and general mechanism of actionAll steroid receptors share commo-n structural features with the follow-ing functional domains encoded by distinctly separate exons 1–8 (Figure 1)1: amino-terminal domain (NTD
or A/B region), DNA-binding domain (DBD or C region), hinge region (D region) and ligand binding doma-in (LBD or E region). The NTD, which contains the transcription activation function 1 (AF1), is the most variable region17. In contrast, DBD is the most conserved domain being character-ised by the presence of two zinc- fin-gers, which are responsible for bindi-ng receptors to DNA at specific nucle-otide sequences called hormone res-ponse elements (HREs)18. DBD is lin-ked to LBD through the hinge region, which is a poorly conserved domain that often contains the nuclear
localisation signal. LBD, which is less conserved than DBD, is responsible for ligand binding, dimerisation and interaction with heat shock proteins, in addition to containing the ligand-dependent transactivation function 2 (AF2)19. The F region of yet unknown function is only present in ERs.
Supporting their structural simi-larities, SSHRs modulate numerous and specific responses in a large vari-ety of cells that share a general mech-anism of action, which has been clarified in detail through molecular biology research20.
A simplified model of steroid ac-tion in target cells is presented in Figure 2. After biosynthesis, steroid hormones reach target cells via the blood stream and due to their lipo-philic nature, they cross the cell membrane by simple diffusion, being retained with high affinity and speci-ficity by their nuclear receptors. In the absence of the hormones, the re-ceptors are mostly located in the cy-toplasm and are transcriptionally inactive due to being associated with heat shock proteins (HSPs). The most widely accepted theory states that ligand-binding promotes a set of conformational modifications in the receptor structure, which causes the displacement of inhibitory HSPs and facilitates receptor translocation to the nucleus1. Upon binding to the hormone response elements of target genes, hormone–receptor complexes induce remodelling of chromatin and interact with transcriptional machin-ery, thereby activating or repressing the transcription of target genes20. Steroid hormone receptors may also establish protein–protein interac-tions with other sequence-specific transcription factors or establish cross-talk with signal transduction pathways that are activated by extra-cellular signals via membrane recep-tors [e.g., the G-protein coupled receptor (GPCR)] and the activation of protein kinase cascades21–23. These represent the so-called non-genomic
Figure 1: Schematic of the structural and functional domains of steroid hor-mone receptor proteins (a) and the coding of exons 1–8 in relation to each func-tional domain of human ER (b) and AR (c) genes. The highly conserved regions, domain C and E containing DNA-binding and ligand-binding domains, respec-tively, are indicated by shaded boxes. Empty boxes represent the divergent re-gions A/B, D and F. Note that ER is the only steroid receptor where the F domain is present. The function of each domain is indicated. AF, transcriptional activa-tion function; NLS, nuclear localisation signal; HSP, heat shock protein.
titution in which they were perform-ed. All human subjects, in these refe-renced studies, gave informed conse-nt to participate in these studies. Animal care was in accordance with the institution guidelines.

Page 3 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
genes and signalling mechanisms involved in both classical- and membrane-mediated effects. Detailed research on the mechanism of steroid actions in each particular cell type of the testis may lead to the development of new and more efficient options for the prevention and treatment of male reproductive disorders.
Alternatively spliced variants of ER and AR in the testisAlternative splicing is a mechanism in which several mRNA transcripts are generated from a single gene giv-ing rise to functionally distinct pro-teins. Thus, it plays a major role in the generation of proteomic and functional diversity in metazoa15. Indeed, more than 50% of human genes undergo alternative splicing, which seems to be a process that is quite prevalent in SSHR genes14. The process of alternative splicing in-volves several distinct mechanisms, including mainly the deletion of ex-ons in addition to the use of ‘intronic’ exons and alternative splice sites (Figure 3).
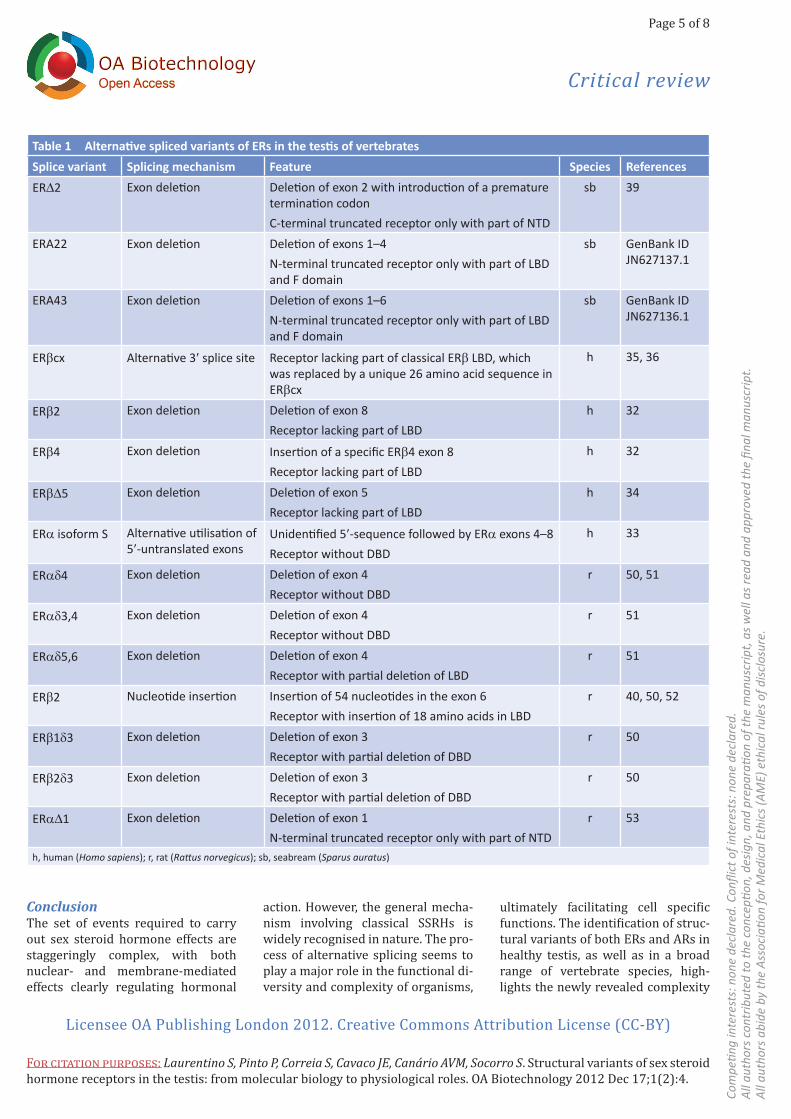
Several variants of both ERα and ERβ (Table 1) have been found in the testis of vertebrates32–38 and, more re-cently, new exon-deleted ERα tran-script variants have been isolated from piscine testis39. Many of these transcripts seem to modulate oestro-genic actions and may play a role in the regulation of spermatogenesis. Some transcripts display cell-depen-dent expression pattern during sper-matogenesis, either being absent or restricted to haploid cells32,36. In addi-tion, variants lacking an intact DBD or unaltered ligand-binding capabil-ity have been described as dominant negative receptors that block ERα, ERβ or both ERα and ERβ signalling pathways34,35,40. An alternatively spliced isoform of ERβ lacking exon 5 and predicted to have part of LBD missing displayed ligand-independent nuclear localisation, inhibiting 17β-oestradiol-stimulated transactivation both in ERβ and ERα34.
receptors may exert particular and tissue-specific effects that depend on genetic, cellular and physiological circumstances.
Nevertheless, the manner in which SSHRs function in healthy and dereg-ulated reproductive processes has not been completely resolved. There is much more to learn about steroid hormone actions in testicular cells that regulate spermatogenesis; spe-cifically, knowledge remains lacking about the set of steroid activated
pathways of steroid hormone action and have been connected with rapid effects, because they do not depend on the activation of RNA transcrip-tion or translation into protein and usually involve the activation of protein kinase cascades24,25, mobili-sation of intracellular calcium26,27, increased cyclic AMP (cAMP) concen-trations28,29 and modulation of nitric oxide release30,31, amongst others. Through the fine tuning of all these dif-ferent mechanisms, steroid hormone
Steriodhormones
Receptor
1
2
G-protein coupledreceptor
5Adenylylcyclase
cAMP
P
P P
Protein-kinasecascade
PLC
DNA
DNA
DNA
TF
TF
TFNucleusProtein-kinase
cascade
Growth factorreceptor
3
4
Cytoplasm
Membrane-boundsteroid receptor
PKAIP3 DAG
PKC
Ca2+
PDNA
Figure 2: Molecular mechanism of the action of steroid hormone receptors. After entering the cell, steroid hormones bind to their specific intracellular receptors, and hormone–receptor complexes are translocated to the nucleus, where they bind to DNA as dimmers modulating gene expression (1). Alternatively, in the nucleus, hormone-bound receptors may interact with tran-scription factors, which in turn bind to their responsive elements on the DNA, thereby controlling gene expression (2). Hormone independent mechanisms may involve steroid-hormone receptor phosphorylation and activation, which is triggered by protein-kinase cascade in response to growth factors binding to their membrane receptor. Phosphorylated steroid receptors enter the nucleus and bind to DNA, regulating gene expression (3). Steroid hormones may also bind membrane located steroid receptors, triggering the activation of protein-kinase cascades that result in the phosphorylation of transcription factors that bind to their own response elements in the genome, thereby controlling gene expression (4). There are also steroid hormone effects that do not depend on transcription and protein synthesis, which are usually mediated by intracellular secondary messengers produced in response to the activation of G-protein cou-pled receptors by steroids (5). TF, transcription factor; cAMP, cyclic AMP; PKA, protein kinase A; PLC, phospholipase C; IP3, Inositol 1,4,5-triphosphate; DAG, dyacilglycerol; PKC, protein kinase C.

For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Page 4 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
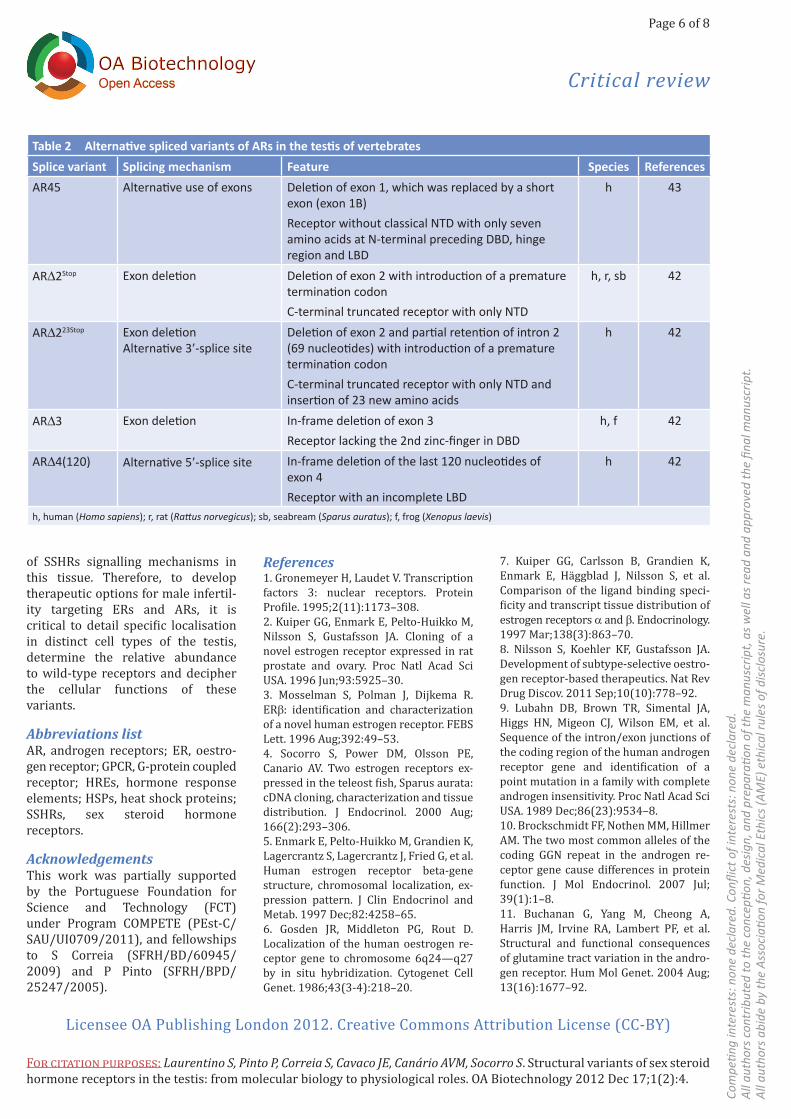
Although the precise role of AR variants in the context of androgen signalling in testicular cells requires exploration in future studies, cellular and molecular studies have charac-terised the function of similar alter-native forms of AR found in tissues other than testis. Many of these truncated AR isoforms, even with deletion of part or entire LBD, are constitutively active, serving as ligand-independent transcription factors and being able to promote the transcription of AR-dependent genes44–46. Jagla et al.47 described a variant with aberrant retention of intron 2 and insertion of 23 amino acids, which, even though devoid of genomic actions, exhibits a cytoplas-matic function affecting the activity of transcription factors NF-B and AP-1. Another curious variant without DBD was described as being localised at the cell membrane, in addition to being required for the optimal tran-scriptional activity of AR48. The AR45 variant, which was detected in the testis, lacks the entire region encoded by exon 1 and generates a protein with DBD, hinge region, and LBD, which are preceded by a short N-terminal region composed of seven amino acid encoded by a short alternative exon (exon 1B)43. This variant seems to act as a negative regulator of classical AR function, and its mechanism of AR inhibition was suggested to involve the formation of AR–AR45 heterodi-mers43. Moreover, in addition to their identification and structural charac-terisation, it has been shown that some AR variants have relevant phys-iological functions in regulating pros-tate cancer cell growth and in the development of androgen-resistant phenotype43–45,47–49.
Recent findings on the identifica-tion of new AR and ER variants in the testis have demonstrated an unex-plored complexity of steroid hor-mone actions, opening new lines of research to delineate the ER/AR tran-scriptome, as well as potentially new signalling pathways.
of intron 2, corresponding to the addition of 23 amino acids and a pre-mature stop codon in the translated product. The partial retention of in-tron 2 resulted from the use of an al-ternative splicing site upstream of the standard site. This study42 also demonstrated the presence of some of these AR variants in evolutionarily distant vertebrate species and in a broad range of human tissues, suggesting a relevant physiological role for the putative protein isoforms that were generated. Moreover, this was the first report to clearly demon-strate that the occurrence of AR splice variants is not restricted to pathological tissues. It is worth not-ing that until 2011, only one AR vari-ant had been identified in human testis43. Table 2 summarises the in-formation about the identified testic-ular AR splice variants, splicing mechanisms and features of the puta-tive proteins.
In contrast to the information available about ERs, data about alter-natively spliced forms of AR is almost entirely associated with cancer and cases of androgen insensitivity syn-drome41. To date, limited knowledge about the presence of alternative forms of AR in non-pathological tis-sues, including testis, exists. However, a recent report by our research group42, using a reverse transcrip-tion-polymerase chain reaction (RT-PCR) strategy, identified four alternatively spliced isoforms of AR in human testis: i) an isoform with the deletion of exon 2 introducing a premature stop codon, which is ex-pected to result in a truncated pro-tein mainly consisting of NTD, ii) an isoform with an in-frame deletion of 120 nucleotides of exon 4, iii) an iso-form with an in-frame deletion of exon 3, which encodes the second zinc-finger in DBD and iv) an isoform lacking exon 2 but retaining 69 bp
AG guragu yuray Yrich ag
PPTBranch site 3′ splice site5′ splice site
Exon Intron Exon
Cons�tu�vesplicing
Exondele�on
Intronreten�on
Alterna�ve5′ss
Alterna�ve3′ss
Pre-mRNA mRNA
(a)
(b)
G
Figure 3: Common mechanisms of alternative splicing in eukaryotic genes (a). Basal splice signals (ss): 5′ splice site, branch site, polypyrimidine tract (PPT) and 3′ splice site (b). Constitutive splicing and types of alternative splicing: exon deletion, intron retention, alternative 3′ ss and alternative 5′ ss. Constitutive ex-ons are shown in blue, alternatively spliced exons are shown in yellow, introns are represented by solid lines and dashed lines indicate splicing options.
Page 5 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
ultimately facilitating cell specific functions. The identification of struc-tural variants of both ERs and ARs in healthy testis, as well as in a broad range of vertebrate species, high-lights the newly revealed complexity
action. However, the general mecha-nism involving classical SSRHs is widely recognised in nature. The pro-cess of alternative splicing seems to play a major role in the functional di-versity and complexity of organisms,
ConclusionThe set of events required to carry out sex steroid hormone effects are staggeringly complex, with both nuclear- and membrane-mediated effects clearly regulating hormonal
Table 1 Alternative spliced variants of ERs in the testis of vertebratesSplice variant Splicing mechanism Feature Species ReferencesERD2 Exon deletion Deletion of exon 2 with introduction of a premature
termination codonC-terminal truncated receptor only with part of NTD
sb 39
ERA22 Exon deletion Deletion of exons 1–4 N-terminal truncated receptor only with part of LBD and F domain
sb GenBank ID JN627137.1
ERA43 Exon deletion Deletion of exons 1–6N-terminal truncated receptor only with part of LBD and F domain
sb GenBank ID JN627136.1
ERβcx Alternative 3′ splice site Receptor lacking part of classical ERβ LBD, which was replaced by a unique 26 amino acid sequence in ERβcx
h 35, 36
ERβ2 Exon deletion Deletion of exon 8Receptor lacking part of LBD
h 32
ERβ4 Exon deletion Insertion of a specific ERβ4 exon 8Receptor lacking part of LBD
h 32
ERβD5 Exon deletion Deletion of exon 5Receptor lacking part of LBD
h 34
ERα isoform S Alternative utilisation of 5′-untranslated exons
Unidentified 5′-sequence followed by ERα exons 4–8Receptor without DBD
h 33
ERαd4 Exon deletion Deletion of exon 4Receptor without DBD
r 50, 51
ERαd3,4 Exon deletion Deletion of exon 4Receptor without DBD
r 51
ERαd5,6 Exon deletion Deletion of exon 4Receptor with partial deletion of LBD
r 51
ERβ2 Nucleotide insertion Insertion of 54 nucleotides in the exon 6Receptor with insertion of 18 amino acids in LBD
r 40, 50, 52
ERβ1d3 Exon deletion Deletion of exon 3Receptor with partial deletion of DBD
r 50
ERβ2d3 Exon deletion Deletion of exon 3Receptor with partial deletion of DBD
r 50
ERαD1 Exon deletion Deletion of exon 1N-terminal truncated receptor only with part of NTD
r 53
h, human (Homo sapiens); r, rat (Rattus norvegicus); sb, seabream (Sparus auratus)
For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Page 6 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
7. Kuiper GG, Carlsson B, Grandien K, Enmark E, Häggblad J, Nilsson S, et al. Comparison of the ligand binding speci-ficity and transcript tissue distribution of estrogen receptors α and β. Endocrinology. 1997 Mar;138(3):863–70.8. Nilsson S, Koehler KF, Gustafsson JA. Development of subtype-selective oestro-gen receptor-based therapeutics. Nat Rev Drug Discov. 2011 Sep;10(10):778–92.9. Lubahn DB, Brown TR, Simental JA, Higgs HN, Migeon CJ, Wilson EM, et al. Sequence of the intron/exon junctions of the coding region of the human androgen receptor gene and identification of a point mutation in a family with complete androgen insensitivity. Proc Natl Acad Sci USA. 1989 Dec;86(23):9534–8.10. Brockschmidt FF, Nothen MM, Hillmer AM. The two most common alleles of the coding GGN repeat in the androgen re-ceptor gene cause differences in protein function. J Mol Endocrinol. 2007 Jul; 39(1):1–8.11. Buchanan G, Yang M, Cheong A, Harris JM, Irvine RA, Lambert PF, et al. Structural and functional consequences of glutamine tract variation in the andro-gen receptor. Hum Mol Genet. 2004 Aug; 13(16):1677–92.
References1. Gronemeyer H, Laudet V. Transcription factors 3: nuclear receptors. Protein Profile. 1995;2(11):1173–308.2. Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA. Cloning of a novel estrogen receptor expressed in rat prostate and ovary. Proc Natl Acad Sci USA. 1996 Jun;93:5925–30.3. Mosselman S, Polman J, Dijkema R. ERβ: identification and characterization of a novel human estrogen receptor. FEBS Lett. 1996 Aug;392:49–53.4. Socorro S, Power DM, Olsson PE, Canario AV. Two estrogen receptors ex-pressed in the teleost fish, Sparus aurata: cDNA cloning, characterization and tissue distribution. J Endocrinol. 2000 Aug; 166(2):293–306.5. Enmark E, Pelto-Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, Fried G, et al. Human estrogen receptor beta-gene structure, chromosomal localization, ex-pression pattern. J Clin Endocrinol and Metab. 1997 Dec;82:4258–65.6. Gosden JR, Middleton PG, Rout D. Localization of the human oestrogen re-ceptor gene to chromosome 6q24—q27 by in situ hybridization. Cytogenet Cell Genet. 1986;43(3-4):218–20.
of SSHRs signalling mechanisms in this tissue. Therefore, to develop therapeutic options for male infertil-ity targeting ERs and ARs, it is critical to detail specific localisation in distinct cell types of the testis, determine the relative abundance to wild-type receptors and decipher the cellular functions of these variants.
Abbreviations listAR, androgen receptors; ER, oestro-gen receptor; GPCR, G-protein coupled receptor; HREs, hormone response elements; HSPs, heat shock proteins; SSHRs, sex steroid hormone receptors.
AcknowledgementsThis work was partially supported by the Portuguese Foundation for Science and Technology (FCT) under Program COMPETE (PEst-C/SAU/UI0709/2011), and fellowships to S Correia (SFRH/BD/60945/ 2009) and P Pinto (SFRH/BPD/ 25247/2005).
Table 2 Alternative spliced variants of ARs in the testis of vertebrates Splice variant Splicing mechanism Feature Species ReferencesAR45 Alternative use of exons Deletion of exon 1, which was replaced by a short
exon (exon 1B)Receptor without classical NTD with only seven amino acids at N-terminal preceding DBD, hinge region and LBD
h 43
ARD2Stop Exon deletion Deletion of exon 2 with introduction of a premature termination codonC-terminal truncated receptor with only NTD
h, r, sb 42
ARD223Stop Exon deletionAlternative 3′-splice site
Deletion of exon 2 and partial retention of intron 2 (69 nucleotides) with introduction of a premature termination codonC-terminal truncated receptor with only NTD and insertion of 23 new amino acids
h 42
ARD3 Exon deletion In-frame deletion of exon 3Receptor lacking the 2nd zinc-finger in DBD
h, f 42
ARD4(120) Alternative 5′-splice site In-frame deletion of the last 120 nucleotides of exon 4 Receptor with an incomplete LBD
h 42
h, human (Homo sapiens); r, rat (Rattus norvegicus); sb, seabream (Sparus auratus); f, frog (Xenopus laevis)

Page 7 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
34. Inoue S, Ogawa S, Horie K, Hoshino S, Goto W, Hosoi T, et al. An estrogen recep-tor beta isoform that lacks exon 5 has dominant negative activity on both ERalpha and ERbeta. Biochem Biophys Res Commun. 2000 Dec;279(3):814–9.35. Ogawa S, Inoue S, Watanabe T, Orimo A, Hosoi T, Ouchi Y, et al. Molecular clon-ing and characterization of human estro-gen receptor betacx: a potential inhibitor ofestrogen action in human. Nucleic Acids Res. 1998 Aug;26(15):3505–12.36. Saunders PT, Millar MR, Macpherson S, Irvine DS, Groome NP, Evans LR, et al. ERbeta1 and the ERbeta2 splice variant (ERbetacx/beta2) are expressed in dis-tinct cell populations in the adult human testis. J Clin Endocrinol Metab. 2002 Jun; 87(6):2706–15.37. Scobie GA, Macpherson S, Millar MR, Groome NP, Romana PG, Saunders PT. Human oestrogen receptors: differential expression of ER alpha and beta and the identification of ER beta variants. Steroids. 2002 Nov;67(12):985–92.38. Shoda T, Hirata S, Kato J, Hoshi K. Cloning of the novel isoform of the estro-gen receptor beta cDNA (ERbeta isoform M cDNA) from the human testicular cDNA library. J Steroid Biochem Mol Biol. 2002 Oct;82(2-3):201–8.39. Pinto PI, Teodósio R, Socorro S, Power DM, Canário AV. Structure, tissue distri-bution and estrogen regulation of splice variants of the sea bream estrogen recep-tor alpha gene. Gene. 2012 Jul;503(1): 18–24.40. Maruyama K, Endoh H, Sasaki-Iwaoka H, Kanou H, Shimaya E, Hashimoto S, et al. A novel isoform of rat estrogen receptor beta with 18 amino acid insertion in the ligand binding domain as a putative dom-inant negative regular of estrogen action. Biochem Biophys Res Commun. 1998 May;246(1):142–7.41. Dehm SM, Tindall DJ. Alternatively spliced androgen receptor variants. Endocr Relat Cancer. 2011 Sep;18(5): R183–96.42. Laurentino SS, Pinto PI, Tomás J, Cavaco JE, Sousa M, Barros A, et al. Identification of androgen receptor vari-ants in testis from humans and other ver-tebrates. Andrologia. 2012 Jun 26.43. Ahrens-Fath I, Politz O, Geserick C, Haendler B. Androgen receptor function is modulated by the tissue-specific AR45 variant. FEBS J. 2005 Jan;272(1): 74–84.
to modulate G protein-coupled receptors via activation of protein kinase A and pro-tein kinase C pathways. Steroids. 1999 Jan-Feb;64(1-2):64–75.25. Lee YF, Lin WJ, Huang J, Messing EM, Chan FL, Wilding G, et al. Activation of mitogen-activated protein kinase path-way by the antiandrogen hydroxyfluta-mide in androgen receptor-negative prostate cancer cells. Cancer Res. 2002 Nov;62(21):6039–44.26. Lyng FM, Jones GR, Rommerts FF. Rapid androgen actions on calcium sig-naling in rat sertoli cells and two human prostatic cell lines: similar biphasic re-sponses between 1 picomolar and 100 nanomolar concentrations. Biol Reprod. 2000 Sep;63(3):736–47.27. Morley P, Whitfield JF, Vanderhyden BC, Tsang BK, Schwartz JL. A new, nonge-nomic estrogen action: the rapid release of intracellular calcium. Endocrinology. 1992 Sep;131(3):1305–12.28. Aronica SM, Kraus WL, Katzenellenbogen BS. Estrogen action via the cAMP signaling pathway: stimulation of adenylate cyclase and cAMP-regulated gene transcription. Proc Natl Acad Sci USA. 1994 Aug;91(18):8517–21.29. Merkle D, Hoffmann R. Roles of cAMP and cAMP-dependent protein kinase in the progression of prostate cancer: cross-talk with the androgen receptor. Cell Signal. 2011 Mar;23(3):507–15.30. Koizumi H, Yu J, Hashimoto R, Ouchi Y, Okabe T. Involvement of androgen recep-tor in nitric oxide production induced by icariin in human umbilical vein endothe-lial cells. FEBS Lett. 2010 Jun;584(11): 2440–4.31. Stefano GB, Prevot V, Beauvillain JC, Cadet P, Fimiani C, Welters I, et al. Cell-surface estrogen receptors mediate cal-cium-dependent nitric oxide release in human endothelia. Circulation. 2000 Apr; 101(13):1594–7.32. Aschim EL, Saether T, Wiger R, Grotmol T, Haugen TB. Differential distri-bution of splice variants of estrogen re-ceptor beta in human testicular cells suggests specific functions in spermato-genesis. J Steroid Biochem Mol Biol. 2004 Sep;92(1-2):97–106.33. Hirata S, Shoda T, Kato J, Hoshi K. The novel isoform of the estrogen receptor-alpha cDNA (ERalpha isoform S cDNA) in the human testis. J Steroid Biochem Mol Biol. 2002 Mar;80(3): 299–305.
12. Davis-Dao CA, Tuazon ED, Sokol RZ, Cortessis VK. Male infertility and varia-tion in CAG repeat length in the androgen receptor gene: a meta-analysis. J Clin Endocrinol Metab. 2007 Nov;92(11): 4319–26.13. Kumar R, Atamna H, Zakharov MN, Bhasin S, Khan SH, Jasuja R. Role of the androgen receptor CAG repeat polymor-phism in prostate cancer, and spinal and bulbar muscular atrophy. Life Sci. 2011 Mar;88(13-14):565–71.14. Hirata S, Shoda T, Kato J, Hoshi K. Isoform/variant mRNAs for sex steroid hormone receptors in humans. Trends Endocrinol Metab. 2003 Apr;14(3): 124–9.15. Nilsen TW, Graveley BR. Expansion of the eukaryotic proteome by alternative splicing. Nature. 2010 Jan;463(7280): 457–63.16. Elliott DJ, Grellscheid SN. Alternative RNA splicing regulation in the testis. Reproduction. 2006 Dec;132(6):811–9.17. Tzukerman MT, Esty A, Santiso-Mere D, Danielian P, Parker MG, Stein RB, et al. Human estrogen receptor transactiva-tional capacity is determined by both cel-lular and promoter context and mediated by two functionally distinct intramolecu-lar regions. Mol Endocrinol. 1994 Jan; 8(1):21–30.18. Schwabe JW, Neuhaus D, Rhodes D. Solution structure of the DNA-binding do-main of the oestrogen receptor. Nature. 1990 Nov;348(6300):458–61.19. Danielian PS, White R, Lees JA, Parker MG. Identification of a conserved region required for hormone dependent tran-scriptional activation by steroid hormone receptors. EMBO J. 1992 Mar;11(3): 1025–33.20. Beato M, Klug J. Steroid hormone re-ceptors: an update. Hum Reprod Update. 2000 May-Jun;6(3):225–36.21. Nilsson S, Mäkelä S, Treuter E, Tujague M, Thomsen J, Andersson G, et al. Mechanisms of estrogen action. Physiol Rev. 2001 Oct;81(4):1535–65.22. Pi M, Parrill AL, Quarles LD. GPRC6A mediates the non-genomic effects of ste-roids. J Biol Chem. 2010 Dec;285(51): 39953–64.23. Prossnitz ER, Maggiolini M. Mechanisms of estrogen signaling and gene expression via GPR30. Mol Cell Endocrinol. 2009 Sep;308(1-2):32–8.24. Kelly MJ, Lagrange AH, Wagner EJ, Rønnekleiv OK. Rapid effects of estrogen

For citation purposes: Laurentino S, Pinto P, Correia S, Cavaco JE, Canário AVM, Socorro S. Structural variants of sex steroid hormone receptors in the testis: from molecular biology to physiological roles. OA Biotechnology 2012 Dec 17;1(2):4.
Page 8 of 8
Com
petin
g in
tere
sts:
non
e de
clar
ed. C
onfli
ct o
f int
eres
ts: n
one
decl
ared
.Al
l aut
hors
con
trib
uted
to th
e co
ncep
tion,
des
ign,
and
pre
para
tion
of th
e m
anus
crip
t, as
wel
l as r
ead
and
appr
oved
the
final
man
uscr
ipt.
All a
utho
rs a
bide
by
the
Asso
ciati
on fo
r Med
ical
Eth
ics (
AME)
eth
ical
rule
s of d
isclo
sure
.
Critical review
Licensee OA Publishing London 2012. Creative Commons Attribution License (CC-BY)
rat testis. J Mol Endocrinol. 2010 Sep; 45(3):147–59.51. Friend KE, Resnick EM, Ang LW, Shupnik MA. Specific modulation of es-trogen receptor mRNA isoforms in rat pi-tuitary throughout the estrous cycle and in response to steroid hormones. Mol Cell Endocrinol. 1997 Aug;131(2):147–55.52. Petersen DN, Tkalcevic GT, Koza-Taylor PH, Turi TG, Brown TA. Identification of estrogen receptor beta2, a functional variant of estrogen receptor beta expressed in normal rat tissues. Endocrinology. 1998 Mar;139(3):1082–92.53. Ishii H, Kobayashi M, Sakuma Y. Alternative promoter usage and alternative splicing of the rat estrogen receptor alpha gene generate numerous mRNA variants with distinct 5’-ends. J Steroid Biochem Mol Biol. 2010 Jan;118(1-2):59–69.
variant of the androgen receptor de-tected in a metastatic prostate cancer ex-hibits exclusively cytoplasmic actions. Endocrinology. 2007 Sep;148(9):4334–43.48. Yang X, Guo Z, Sun F, Li W, Alfano A, Shimelis H, et al. Novel membrane-associ-ated androgen receptor splice variant potentiates proliferative and survival re-sponses in prostate cancer cells. J Biol Chem. 2011 Oct;286(41):36152–60.49. Tepper CG, Boucher DL, Ryan PE, Ma AH, Xia L, Lee LF, et al. Characterization of a novel androgen receptor mutation in a relapsed CWR22 prostate cancer xeno-graft and cell line. Cancer Res. 2002 Nov;62(22):6606–14.50. Bois C, Delalande C, Nurmio M, Parvinen M, Zanatta L, Toppari J, et al. Age- and cell-related gene expression of aromatase and estrogen receptors in the
44. Dehm SM, Schmidt LJ, Heemers HV, Vessella RL, Tindall DJ. Splicing of a novel androgen receptor exon generates a con-stitutively active androgen receptor that mediates prostate cancer therapy resis-tance. Cancer Res. 2008 Jul;68(13): 5469–77.45. Guo Z, Yang X, Sun F, Jiang R, Linn DE, Chen H, et al. A novel androgen receptor splice variant is up-regulated during pros-tate cancer progression and promotes an-drogen depletion-resistant growth. Cancer Res. 2009 Mar;69(6):2305–13.46. Hu R, Isaacs WB, Luo J. A snapshot of the expression signature of androgen re-ceptor splicing variants and their distinc-tive transcriptional activities. Prostate. 2011 Nov;71(15):1656–67.47. Jagla M, Fève M, Kessler P, Lapouge G, Erdmann E, Serra S, et al. A splicing



![Steroid Receptors in Human Lung Cancer1 · [CANCER RESEARCH 45, 4206-4214, September 1985] Steroid Receptors in Human Lung Cancer1 Craig W. Seattle, Nancy W. Hansen, and Paul A. Thomas2](https://img.pdfslide.net/doc/110x75/60e9c9b1d253595c6007e030/steroid-receptors-in-human-lung-cancer1-cancer-research-45-4206-4214-september.jpg)

























