Embed Size (px)
Citation preview

Journal of Luminescence 102–103 (2003) 469–475
Study of the effect of simulated space environment on phageT7 and isolated T7 DNA thin films
Andrea Feketea, Istv!an F .oldv!arib,*, M!arton Heged .usa, K!aroly M !odosa,Gy .orgyi Ront !oc, G!asp!ar Kov!acsc, Attila B!ercesc, !Agnes P!eterb
a Institute of Biophysics and Radiation Biology, Semmelweis University, Budapest, POB 263, H-1444, HungarybResearch Institute for Solid State Physics and Optics, Hungarian Academy of Sciences, Konkoly-Thege u. 29-33,
Budapest 1121, HungarycMTA-SE Research Group for Biophysics, Semmelweis University, Budapest, Hungary
Abstract
Phage and uracil response (PUR) experiment is part of the ROSE consortium selected by the European Space Agency
for the first mission on the EXPOSE facility of the International Space Station. Its main goal is to examine and quantify
the effect of specific space conditions (e.g. solar UV radiation, vacuum) on nucleic acid models. To achieve this, an
improved method was elaborated for the preparation of DNA and bacteriophage thin films. The homogeneity of the
films was controlled by UV spectroscopy and microscopy. To provide experimental evidence for the hypothesis that
interplanetary transfer of life is possible, phage T7 and isolated T7 DNA thin films have been exposed to selected space
conditions: intense UVC radiation (l ¼ 254 nm) and high vacuum (10�5mbar). The effects of DNA hydration,
conformation and packing on UV radiation damage were examined. Characteristic changes in the absorption spectrum,
in the electrophoretic pattern of DNA and the decrease of the amount of PCR products have been detected indicating
the photodamage of isolated and intraphage DNA. The paper summarizes the spectroscopical data.
r 2003 Elsevier Science B.V. All rights reserved.
PACS: 87.14G; 87.15M; 87.50G
Keywords: Thin films; DNA; Bacteriophage; UV radiation; Space vacuum
1. Introduction
The European Space Agency (ESA) developed amulti-user exposure facility with the acronymEXPOSE [1], and this exobiology research plat-form will be taken to the International SpaceStation (ISS). The ISS provides a unique oppor-tunity for in-depth experimental programs in
various fields of biology. To study the responsesof organisms to the space environment (ROSE), aconsortium of scientists has been formed to studyphotobiological processes in the simulated radia-tion climate of planets, and the probabilities andlimitations for life to be distributed among objectsof our solar system. The experiments selected forthe first flight during the early utilization period ofthe ISS are summarized in Ref. [2]. One of them isphage and uracil response (PUR) with theobjective of studying the sensitivity of biologicallyeffective UV radiation to ozone, and the assay
*Corresponding author. Tel.: +36-1-392-2627; fax: +36-1-
392-2223.
E-mail address: [email protected] (I. F .oldv!ari).
0022-2313/03/$ - see front matter r 2003 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0022-2313(02)00581-1

systems are: bacteriophage T7, phage T7 DNA,uracil. In this paper, we focus on the first twoassay systems. We report here the results of thepreliminary experiments on the responses to solarUV radiation and space vacuum. Solar UVradiation has been found to be the most deleter-ious factor of space, as tested with dried prepara-tions of viruses and bacterial and fungal spores [3].The reason for this is the highly energetic UVCand vacuum UV radiation that is directly absorbedby the DNA. The atmosphere effectively absorbsthese harmful UV ranges, and they do not reachthe surface of the Earth. In space vacuumsolutions/suspensions cannot be examined, there-fore an improved method of Foldvari et al. [4] hasbeen developed to produce good quality thinlayers of DNA and phage T7. To provideexperimental evidence, testing the hypothesis thatinterplanetary transfer of life is possible, phage T7and isolated T7 DNA have been exposed toselected space conditions: intense UV radiationand high vacuum. The following effects have beenanalysed: change in the UV spectra, and change inthe electrophoretic pattern. The amount of da-mages in DNA/phage was determined by quanti-tative PCR. This paper summarizes thespectroscopical data, the detailed results of theother methods will be published elsewhere.
2. Experimental
The preparation of bacteriophage T7 wascarried out according to the method described inRef. [5], while the isolation of T7 DNA ispublished in Ref. [6]. To obtain good opticalquality DNA thin films, the isolated T7 DNA wasdissolved in aqueous solution containing 0.01–0.1M NaCl or LiCl (5–20 ng/ml DNA). Ethanolwas added to the solution step by step until theethanol concentration came close to the amountneeded for precipitation of DNA (about 50%).The obtained quasi-colloidal solution was depos-ited onto the silica window (16mm diameter) ofthe measuring cell by centrifugation at 11000g inspecial centrifuge tubes (3� 20min centrifugationwith gradual increase of ethanol concentration upto 80%). The homogeneity of the obtained DNA
films were checked by polarization and phasecontrast microscopy (Olympus SM20 and ReichertMeF2 microscope). The estimated thickness ofthe films used in this experiment was about20–200 nm. The relative humidity (r.h.) of theDNA films was adjusted according to Falk et al.[7] resulting in B, A, C or D conformations. Thesamples were allowed to equilibrate for at least 3days at RT. Films of intact T7 phages wereprepared similarly but without ethanol. Afterdissolution from the film about 60% of the phagesremained active as it was determined using E. coli
B host cells by plaque counting [8].Absorption spectra and difference spectra were
recorded with a UNICAM UV/VIS spectrophot-ometer (equipped with Vision 3.4 software). Forthin film measurements, special vacuum-tightsample holders were used. The absorption spectrawere corrected for reflected light from the surfaceand for the scattered light applying the relationE ¼ kl�n; with empirical fitting for k and n: Theintegrity of isolated DNA and phage T7 waschecked by dissolving the layers and analysingneutral (for ds breaks) and alkaline agarose gels(for ss breaks). It was shown that our thin layerpreparation method did not damage the DNAstructure. To quantify DNA lesions as a result ofselected space parameters, a quantitative PCRmethod was developed based on the amplificationof a 555 bp (base pair) and a 3826 bp segments ofT7 DNA. The details of the method will bedescribed elsewhere. UVC irradiation of the thinlayers was performed in sandwich form in thespecial, vacuum tight sample holder by a Tungs-ram 15W germicidal lamp with a fluence rate of1W/m2 (254 nm). The effect of vacuum 10�5mbar(1 week) was tested at the Space SimulationFacilities, German Aerospace Center, DLR, Co-logne.
3. Results and discussion
3.1. DNA and phage thin layers
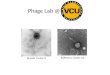
Fig. 1 shows typical microscopic picture ofphage T7 thin layer (upper part) and NaDNAthin film (lower part) produced by an improved
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475470

method of Foldvari et al. [4]. An enlargement scale640:1 and a Nomarski interface was applied toobtain pictures with large contrast. The phage filmshows a uniform, microcrystal pattern, and thephage particles are apparently embedded in saltcrystals while the NaDNA film posses no micro-scopic crystallinity. However, the covering of bothfilms is homogeneous, and the films retaineduniformity under different r.h. conditions. Com-pared to NaDNA, LiDNA films are more prone toself-associate, giving rise to microscopic crystallinedomains in the latter. The microscopic picture ofthe films explains the observed low amount ofscattered light and the fact that the n values
obtained from fitting varied from 1.5 to 2.8 insteadof the expected 4.
3.2. Effects of vacuum and DNA conformation
Earlier space experiments have shown [9] thatup to 70% of bacterial and fungal spores surviveshort-term (10 days) exposure to space vacuum, ifthe spores are embedded in chemical protectantssuch as salt crystals, sugars or if they are exposedin thick layers. Therefore, we examined the role ofsalts (NaCl and LiCl) and the thickness of layer aswell. Changing the initial phage/DNA concentra-tion in the suspension, we found that the thicknessof layer can be linearly increased in the 20–500 nmrange as was indicated by the UV spectra. Bychanging the type of salt and the r.h. of the filmsthe conformation of the DNA can be varied. Invivo, the DNA is normally tightly compacted intochromatin via association with histone and non-histon proteins [10]. Phage T7 contains a double-stranded DNA packed in a superhelical structurebetween a protein shell and core [11,12] thatresembles the chromosomal DNA arrangement inmammalian cells, and it possess a slightly distortedB conformation. Hence, it can be used as achromosome model from the point of view ofresponses to space parameters.The absorption spectra of NaDNA film at
different conformations are presented in Fig. 2A.The spectra are the averages of four independentmeasurements after light scattering correction.Small but repeatable differences were obtainedamong the spectra belonging to different confor-mations. These differences are owing to thelinkage of the different counterions and theamount of water. At the fully hydrated state,the DNA contains 20–30 water molecules pernucleotide pairs, and there are about 2–6 watermolecules per nucleotide pair that are very difficultto remove from the DNA [13–15]. In thecondensed state and in nucleosomes, DNA main-tains its closely associated water but the overallextent of water surrounding DNA is reduced [16].The absorption spectra of phage T7 films showdifferences from DNA below 220 nm where theproteins have distinct absorption. The phage layerwas dissolved from the film in buffer solution and
Fig. 1. Reichert MeF2 reflexion microscopic picture of intact
phage T7 thin film (upper part) and of isolated T7 DNA thin
layer (lower part). Enlargement scale is 640:1 and a Nomarski
interface was applied.
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475 471

we found B60% of the phages remained activedetermined by infecting E:coli B host cells. Itshould be emphasized that the biological activityof the phage exists only in the case of uninjuredDNA–protein interaction [12]. Exposure to va-cuum (10�5mbar, 1 week) of NaDNA films causeda very similar effect to dehydration (0% r.h.) in thespectra of DNA thin films, B30% hyperchromi-city (at 260 nm) was measured. In phage T7 thinlayer B30% hypochromicity was found, also withno significant structural change in the spectrum.Under the effect of vacuum, the volume of thesample contracts, decreasing the average interhe-lical spacing. Using polarization microscopy, wefound that microscopic patterns of salt crystallitesformed as the film dehydrates, but the filmsretained uniformity.The conformation of DNA is controlled by
many different factors, including DNA hydration
[15,17], DNA packing interaction [15], salt con-centration and the DNA counterion [18]. NaDNAundergoes several conformational changes as afunction of r.h.: denatured state, D conformation(very low r.h.), B-like form (below 45% r.h.), Aconformation (between 45% and 92% r.h.), Bconformation (above 92% r.h.) [14]. The A formdiffers significantly from the B form in structure,and exhibits unique base-stacking characteristics,the large base tilt causes interstrand base overlap.This can be the reason that the calculateddifference spectra between B and A conformationof NaDNA film showed relatively large differences(5–15%) at characteristic wavelengths (210, 230,260, and 310 nm). LiDNA, on the other hand,undergoes a change in conformation C to B over65% r.h., and no changes with further increases inr.h. We found a smaller change in the differencespectra of B and C conformations (0.7–11% atdifferent wavelengths), reflecting that the B and Cconformations are structurally similar. The pack-ing order in NaDNA film samples seems to berelatively homogeneous for a given DNA hydra-tion. In contrast, the packing in LiDNA filmsvaries between different samples even at the samer.h. The observed 260 nm absorbance of theNaDNA and LiDNA thin films increase byB30% upon dehydration, and the observedhyperchromism is very similar to the hyperchro-micity measured by ‘‘melting’’ in solution [12],but we found it to be fully reversible. In contrastto the denatured conformation in solutions,the disordered, D conformation of solid DNAin the dehydrated state still maintains a certainrigid structure, the parallel stacking of the basesare largely lost, but the double helical strandsremain entwined. Upon sufficient rehydration, thisstructure returns to the original (B, A or C)conformation by the rapid rematching betweencomplementary bases.
3.3. Effect of UVC radiation
The effect of UVC radiation was measured onNaDNA and phage T7 films. Fig. 2B representsthe UV difference spectra of UVC irradiatedphage thin layers, the samples were irradiatedand measured in sandwich form at 0% r.h. Since
Fig. 2. (A) Absorption spectra of isolated T7 phage NaDNA
thin film at different relative humidities resulting in different
conformations after light scattering correction. (B) difference
spectra of UVC (254 nm) irradiated phage T7 thin layers. The
samples were irradiated and measured in sandwich form, the
data are the averages of four measurements.
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475472

UVC radiation is strongly absorbed in DNA/nucleoproteins, to quantify the UV effect thesamples must be exposed in thin and homogeneouslayers. The photochemical reactions of nucleicacids in the 200–300 nm (4–6 eV) range have beenextensively studied, the most frequent types ofDNA lesions are bipyrimidine photoproducts,mostly cyclobutane-type pyrimidine dimers(CPD)s and (6-4) photoproducts (6-4) PDs (seereviews [19,20]).The effect of UVC radiation on thin films can be
followed by the changes in the UV absorptionspectrum. As an example, the difference spectra ofthe phage T7 thin films under UVC radiation areshown in Fig. 2B. In order to compare the changesoccurring in thin films and in DNA/phage solu-tions, we choose the OD at 264 nm for character-ization of the UVC damage. Fig. 3 shows thechanges at 264 nm as a function of the incidentdose of UVC radiation in phage T7 and T7NaDNA thin films (A), and in isolated T7 DNA
and phage T7 solutions (B). In solution (Fig. 3B),the observed change in OD was more significant inphage T7 than in isolated T7 DNA. However, itcan be seen in Fig. 3A that in thin layers bothphage T7 and isolated DNA behave differently.The difference at 264 nm is positive in films andnegative in solution. In addition, the slope andsaturation of the dose-effect curves are alsodifferent. UVC irradiated phage T7 layer andisolated T7 DNA show saturation in the examinedUVC dose range. In order to achieve measurabledifferences in the spectra the used dose ranges aresomewhat different in solution and in thin films.However, the UVC-induced changes do notsignificantly depend on the conformational stateof DNA, we obtained very similar dose-effectcurves for the different DNA conformations (B, Aor C). This observation is in agreement with thedata of Patrick and Rahn [22].The reasons of the observed differences between
films and solutions can be manifolded. Themeasured change at 264 nm originated from thecompeting hypochromicity due to the decrease ofpyrimidine monomers, and characteristic to theformation of CPDs and (6-4)PDs and hyperchro-micity due to partial denaturation of the DNAstructure. In solution, the effect of CPD and(6-4)PD production is dominant (Fig. 3B). Earlierwe found by lesion specific antibodies [6,21] asignificant decrease in the yield of CPD and(6-4)PD in UVC irradiated isolated T7 DNAsolutions compared to intraphage DNA. The sametendency was obtained by UV difference spectro-scopy (Fig. 3B). For the same amount of templateDNA, the average lesion frequency, determined byPCR in UVC irradiated T7 DNA film, is abouttwice that in aqueous T7 DNA. The amount ofamplified product in DNA film was identical tothat in aqueous DNA for the same amount ofunirradiated DNA. Therefore, the increased UVdamage in DNA thin film compared to isolatedDNA solution can be originated from the tightlypacked DNA when the molecules are stacked oneach other, and the formation of other photo-products than CPDs and (6-4)PDs is favoured (e.g.DNA-DNA cross-links and 5-thyminil-5,6-dihy-drothymine ‘‘spore’’ product). These photopro-ducts do not cause decrease at 264 nm. In Refs.
Fig. 3. The effect of UVC (254 nm) radiation on phage T7 and
isolated T7 DNA thin layers (A) and on T7 phage suspension
and in isolated T7 DNA solution (B). The UVC effect is
characterized by the change in OD at 264 nm (expressed in %)
and is represented in function of incident UVC dose.
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475 473

[6,21], we demonstrated that in phage T7/T7 DNAsolution mostly CPDs and (6-4)PDs are producedby UVC radiation. The PCR is blocked not onlyby CPDs and (6-4)PDs, but also by some otherbulky lesions, and therefore all DNA damages canbe detected. Apparently, in thin films the effect ofhyperchromicity caused by denaturation owing tothe formation of all types of photoproducts islarger than the hypochromicity due to only theformation of CPDs and (6-4)PDs. The reason whyphage T7 in thin layer is less UV sensitive thanisolated T7 DNA may be the structure of thephage thin film (Fig. 1), phage molecules areembedded in salt crystals and crystalline salt canprovide protection against UVC radiation.The explanation for the observed saturation
tendency is that under the influence of shortwavelength UV radiation not only the bipyrimi-dine photoproducts (mostly CPDs and (6-4)PDs)are induced but the reversion of them can also beprovoked by the same photons [6,19–21]. Thesaturation tendency at higher doses reflects akinetics where the formation-reversion of thephotoproducts approaches an equilibrium. As-suming these photo-induced reactions as statisticalevents, the chance for a number of nucleoprotein/DNA molecules to survive the radiation damageduring the irradiation can be a reality. Thedifferent dose range necessary for the spectro-scopical detection in solution/thin film can beexplained by the fact that in thin layers the DNA/phage molecules stack up on each other, thisstacking causes shielding of molecules below thetop few layers. Therefore, the probability of theDNA molecule below the top few layers interact-ing with the photons is small. Fig. 3 indicatesthe incidence UVC intensities in kJ/m2 that are notcorrected for the shielding effect. Assuming amonolayer of DNA of 2 nm thick [23], in ourfilms 10–100 DNA/phage layers are above eachother. However, only the conditions of exposure inmonolayer warrant that statistically all cellsreceive the same radiation dose. Clusters ofcells or multilayer cells do not meet this condition,but when we address the question of survival inspace an exposure in cellular multilayers or largecell clusters may be more realistic than mono-layers.
4. Conclusions
In order to study the responses to simulatedspace conditions an improved method has beendeveloped to prepare DNA and bacteriophagethin films. The optical quality of the films obtainedby centrifugal deposition permits to follow subtledifferences in the UV spectra owing to vacuum(dehydration), different conformations and UVradiation. The effect of UVC radiation on thinfilms was followed by changes in OD at 264 nmshowing a saturation tendency in nucleoproteinfilms. Assuming that the formation reversion ofthe photoproducts are statistical events, during theirradiation the chance for a number of nucleopro-tein molecules to survive the radiation damage canbe a reality.
Acknowledgements
We thank Erzsebet Szakacs, Monika Drabbantand Marta Varszegi for their excellent technicalassistance. This work was supported by grantsfrom the Ministry of Welfare (ETT, 230/2000 and229/2001) as well as PRODEX grant Impl. Cont.No. 14526/00/IVL/SFe(IC).
References
[1] W. Schulte, P. Baglioni, R. Demets, in: Proceedings of
the First European Workshop for Exo-/Astrobiology,
Frascati, Italy, EAS-SP 496, 2001, 183pp.
[2] G. Horneck, Adv. Space Res. 22 (1998) 317.
[3] G. Horneck, Origins Life 23 (1993) 37.
[4] I. Foldvari, A. Fekete, G. Corradi, J. Biochem. Biophys.
Methods 5 (1981) 319.
[5] S. Gaspar, K. Modos, G. Ronto, in: L. Fedina (Ed.),
Advances in Physiological Science, Pergamon, Oxford,
1981, p. 141.
[6] A. Fekete, A.A. Vink, S. Gaspar, A. Berces, K. Modos,
G. Ronto, L. Roza, Photochem. Photobiol. 68 (1998) 527.
[7] M. Falk, K.A. Hartman, R.C. Lord, J. Am. Chem. Soc.
(1963) 391.
[8] G. Ronto, S. Gaspar, P. Grof, J. Photochem. Photobiol.
12 (1992) 285.
[9] G. Horneck, Adv. Space Res. 23 (1999) 381.
[10] D.S. Goodsell, Trend Biochem. Sci. 16 (1999) 203.
[11] G. Ronto, M.M. Agamalyan, G.M. Drabkin, L.A. Feigin,
Y.M. Lvov, Biophys. J. 43 (1983) 309.
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475474

[12] A. Fekete, G. Ronto, L.A. Feigin, V.V. Tikhonchev,
K. Modos, Biophys. Struct. Mech. 9 (1982) 1.
[13] J.F. Ward, Progr. Nucleic Acid Res. Mol. Biol. 35 (1988) 95.
[14] N.J. Tao, S.M. Lindsay, Biopolymers 28 (1989) 203.
[15] S.M. Lindsay, S.A. Lee, J.W. Powell, T. Weidlich,
C. DeMarco, G.D. Lewen, N.J. Tao, Biopolymers 27
(1988) 1015.
[16] M.M. Struck, A. Klug, T. Richmond, J. Mol. Biol. 224
(1992) 253.
[17] W. Saenger, W.H. Hunter, O. Kennard, Nature 324 (1986)
325.
[18] Z. Shakked, G. Guerstein-Guzikevich, M. Eisenstein,
F. Frolow, D. Rabinovch, Nature 342 (1989) 456.
[19] J. Cadet, C. Anselmino, T. Douki, L. Voiturez,
J. Photochem. Photobiol. B 15 (1992) 277.
[20] H. Gorner, J. Photochem. Photobiol. B 26 (1994) 117.
[21] A. Fekete, A.A. Vink, S. Gaspar, K. Modos, A. Berces,
G. Ronto, L. Roza, Photochem. Photobiol. 69 (1999) 545.
[22] M.H. Patrick, D.M. Gray, Photochem. Photobiol. 24
(1976) 507.
[23] M. Folkard, K.M. Prise, B. Vojnovich, B. Brocklehurst,
B.D. Michael, Int. J. Radiat. Biol. 76 (2000) 763.
A. Fekete et al. / Journal of Luminescence 102–103 (2003) 469–475 475





















![Lamda phage[1]](https://img.pdfslide.net/doc/110x75/58cedaba1a28abd4098b6283/lamda-phage1.jpg)






