Embed Size (px)
Citation preview

Anal Bioanal Chem (2006) 385: 1265–1280DOI 10.1007/s00216-006-0582-3
SPECIAL ISSUE PAPER
María Teresa Doménech-Carbó . Laura Osete-Cortina .Juana de la Cruz Cañizares . Fernando Bolívar-Galiano .Julio Romero-Noguera . María Antonia Fernández-Vivas .Inés Martín-Sánchez
Study of the microbiodegradation of terpenoid resin-basedvarnishes from easel painting using pyrolysis–gaschromatography–mass spectrometry and gaschromatography–mass spectrometryReceived: 15 March 2006 / Revised: 24 May 2006 / Accepted: 29 May 2006 / Published online: 7 July 2006# Springer-Verlag 2006
Abstract The alterations produced by microbiologicalattack on terpenoid resin-based varnishes from panel andcanvas paintings have been evaluated using pyrolysis–gaschromatography–mass spectrometry (Py–GC–MS) and gaschromatography–mass spectrometry (GC–MS). The pro-posed methods include the on-line derivatisation of dryingoils and diterpenoid resins using hexamethyldisilazaneduring pyrolysis and the application of methyl chlorofor-mate as a derivatisation reagent for triterpenoid resins inGC–MS. Two types of specimens, consisting of model oilmedium prepared from linseed oil and model spiritvarnishes prepared from colophony and mastic resinsdissolved in turpentine, have been used as referencematerials. For a series of specimens upon which differentgenera of bacteria and fungi were inoculated andencouraged to grow, analyses indicated that no mecha-nisms that commonly occur during the attack of enzymeson drying oils and terpenoid biodegraders were observed tooccur in the oil medium and varnishes studied. Thus, thedegradation pathways observed in the performed trialsusually occur as consequence of natural ageing. Specific
trials consisting of the application of biocides to unin-oculated colophony varnish resulted in the identification ofprocesses that produce undesirable degradation of thevarnish due to interactions between the biocide and thevarnish components. Finally, the studied biocides—Biotin,New-Des and Nipagine—generally exhibited good inhibit-ing effects on the microorganisms studied, although someinteresting differences were found between them regardingthe application method and type of biocide.
Keywords Py–GC–MS . Terpenoid resin .Hexamethyldisilazane . Biocide .Painting microbiodegradation
Introduction
One of the major problems facing analysts and conserva-tors of easel paintings relates to assessing integrity theintegrity of the painting in question, since conservationtreatments are dictated by the degree of preservation.Therefore, it is important to identify the causes ofdeterioration and the mechanisms involved in theseprocesses. Special attention has been paid to the microbi-ological agents that act on the different components of thepainting, in particular on organic components. The effectsproduced by these agents have been described extensively.In particular, biodeterioration of the canvas support due tobiocolonization by microorganisms, which are able tohydrolyse the cellulosic and proteinaceous materials in thesupport, results in a reduction in the mechanical strength ofthe support. Spots due to coloured metabolic byproducts arealso frequently observed. Fading and detachment of thepaint layer are other degradation effects caused by thegrowth of bacteria and fungi in the paint layers. Fatty acidsand glycerol have been reported to occur as breakdownproducts of lipids in coloured spots resulting from themetabolic action of fungi. Lipids easily undergo autoxida-
M. T. Doménech-Carbó (*) . L. Osete-Cortina .J. de la Cruz CañizaresHeritage Conservation Institute,Polytechnical University of Valencia,Camino de Vera s/n,46022 Valencia, Spaine-mail: [email protected]
F. Bolívar-Galiano . J. Romero-NogueraDepartment of Painting, Faculty of Fine Arts,University of Granada,Avda. Andalucía s/n,18071 Granada, Spain
M. A. Fernández-Vivas . I. Martín-SánchezDepartment of Microbiology, Faculty of Sciences,University of Granada,Campus Fuentenueva,18071 Granada, Spain

tion and form very destructive high-energy free radicals andperoxides [1]. The biocolonization of varnishes by fungi hasbeen described as being concomitant to the presence ofmicrocrack patterns that affect the film coating [2, 3].
Despite the fact that works devoted to evaluating theeffects of biodeterioration resulting from the colonization ofstone monuments and wall paintings by microorganisms andthe effects of biocide application are often found in theliterature, only a few works that deal with the biodeterio-ration of easel paintings and materials used in polychromedsculptures have been reported. Fungi species such asTrichoderma, Aspergillius, Penicillium [4, 5],Cladosporium,Fusarium [5], Alternaria[5, 6], Paecilomyces and Sordaria[6], together with the Bacillus and Streptomyces genera ofbacteria [6], have been identified in polychromed sculpturesand canvas painting collections. These microorganismsutilize the organic compounds present in the painting as asource of carbon or release byproducts from their metabolismonto the substrate on which they are growing [7]. In thatsense, the ability of some fungi to convert medium chain fattyacids or triacylglycerols containing these acids to methylketones with one carbon atom less, which is termed “ketonicrancidity” [8, 9], is well-known. As done for easel paintings,Giuliani and Nugari [10] studied the effects of microbialattack on ancient textiles using cultural analyses, microscopyand firefly bioluminescent assays of adenosine triphosphate(ATP). Biofilm formation on the original substrate due tobacterial metabolic activity has also been reported [6].
Presumed mechanisms of action of resistance to biocidesused as preservatives, sanitizers and disinfectants in thefield of cultural heritage conservation have also beenproposed in the literature [11]. Resistance strategies such asinhibitor inactivation, target site alteration, and target siteexclusion are described. Maupeou et al. [12] discussedproblems associated with conserving decorated surfacessuch as mural paintings, and they studied biocidetreatments applied to polychromy. FT-IR studies on thebiodeterioration of amber resin from art collections havebeen carried out by Fenn et al. [13, 14].
Terpenoid resins such asmastic, dammar, elemi, sandarac,copal or rosin resins have been traditionally used forpreparing pictorial varnishes. Colophony is another name forrosin that is widely used in the fields of art history andheritage conservation. This name is used not only in generaltreatises on conservation science and painting materials butalso in current papers devoted to the characterisation ofmaterials used in pictorial varnishes that have been publishedin specialised scientific journals [15, 16]. Vernice liquida,which consisted of sandarac and linseed or walnut oil, andvernice comune, which was made of colophony or oleoresinand linseed oil, were commonly used from the Middle Agesonwards [15, 17–20]. Gettens and Stout [15] establish that avarnish widely used from the ninth up to the late fifteenthcentury was made by dissolving mastic and colophony ormastic, sandarac and colophony in linseed oil. Spiritvarnishes, in which terpenoid resins were dissolved in spiritsof wine and turpentine, were introduced in around thesixteenth century [16, 21–26]. The triterpenoid resin dammarappears to have been first used as a varnish as recently as
1829 [16]. In addition, terpenoid resins have been alsoemployed as additional ingredients in binding media [17, 23].
Gas chromatography (GC) has been applied to the studyof drying oils and terpenoid compounds used in the finearts. Such analysis is based on the formation of trimethylsi-lyl, methyl or ethyl esters from the carboxylic groups presentin the drying oils and the terpenoid components of the resindue to the application of trimethylsilylation [27], methyla-tion [28–35] or ethylation [36, 37] reagents. Pyrolysis hasalso been applied to the analysis of terpenoid compoundsand drying oils present in artworks. This technique has beendirectly coupled to a mass spectrometer (direct temperature-resolved mass spectrometry) [28, 38] or combined with gaschromatography–mass spectrometry [39–48]. Pyrolysis–derivatization thermochemolysis has been performed withtetramethylammonium hydroxide (TMAH) [27, 49–59]. Inparticular, this methodology has been used in a biodeteriora-tion study focused on the molecular characterization oflignin in spruce wood that has been attacked by Phaner-ochaete chrysosporium and Trametes versicolor [60].Methods based on the formation of trimethylsilyl ethersand esters, which use HMDS as the derivatisation reagent[61, 62] instead of TMAH in order to improve the detectionof hydroxylated compounds and to avoid isomerization andunwanted alkylation reactions, have been also proposed.
The general aim of the present work was to evaluate thebiodegradations of a drying oil (linseed oil), a diterpenoidresin (colophony) and a triterpenoid resin (mastic) sincethese are the main components of the terpenoid resin-basedoil varnishes traditionally used to coat easel paintings.Although other materials present in paintings, such asproteins or polysaccharides, are the principal targetcompounds in microbial attacks, studies into the degreeof preservation of paintings demonstrate that varnishesconsisting of terpenoid resins and oils can undergoalterations upon microbial action, as indicated earlier,which is why this study focuses on these materials. Theeffect of biocides employed as disinfectants on colophonyvarnish has also been studied using GC–MS in combina-tion with methyl chloroformate (for mastic) and Py–GC–MS in combination with on-line derivatisation with HMDS(for linseed oil and colophony). The proposed analyticalmethod was applied to samples extracted from a series ofspecimens upon which different microorganisms had beeninoculated.
Biodegradation of drying oils and terpenoid resins
Oil media
Different pathways for the bioconversion of triacylglycerols,which are present in oils and fats of vegetal and animalorigin, due to the enzymatic action of fungi have beendescribed in the literature. These pathways enable fungi toeliminate toxic molecules [9]. The proposed processesinvolve hydrolysis, oxidation, reduction, decarboxylation oresterification, and result in a decrease in the polarity of thecarboxyl group. The metabolic effects of fatty acids are
1266

dependent on pH, concentration and chain length. On theother hand, the ability of fungi to use triacylglycerols as asource of carbon for growth depends on the fatty acidcomposition, so triacylglycerols that contain saturated longchain fatty acids yield fewer growth products than thosecontaining medium chain fatty acids. Similarly, more growthis obtained for liquid substrates than for solid ones [9].
Studies into the natural degradation and biodegradationof oils have established that two different mechanismsoccurs. A mechanism based on classical free radicalreactions has been proposed for the photoageing of oils[63], whereas a mechanism based on reactions catalysed bylipoxygenase enzymes—commonly found in fungi—havebeen implicated in biodegradation phenomena such asrancidity [9]. Two different biodegradation pathways havebeen proposed. The first one is a process related to thepresence of double bonds in the chains of triacylglycerols;this is thought to consist of an oxidation leading to theformation of triacylglycerol hydroperoxides [64] and then ahydrolytic process which yields free fatty acid hydroper-oxides. A third step yields different end-products: ketones,alcohols, aldehydes, ethers and short and medium chainfatty mono and diacids, among others [27, 65, 66]. Asecond pathway involves hydrolysis of the triacylglycerolsto form free long and medium chain fatty acids andglycerol. In the natural ageing of oil paints, this reactionoccurs very quickly to some degree, but much longer timesare required for it to occur extensively [67]. Following this,two processes are possible: i) photooxidation or enzymaticoxidation of the free long chain fatty acids to form free fattyacid hydroperoxydes and then other pertinent end-products[27]; ii) the formation of methyl ketones and secondaryalcohols from free medium chain fatty acids resulting frommetabolism by moulds [9]. Scheme 1 summarizes thereaction pathways involved in the biodegradation of oils.
Terpenoid resins
Various works that focus on the biochemistry of resin acidbiodegradation have been reported in the literature. Thefirst works tried to isolate the bacteria that degrade resinacid using a mixture of alkaline salts of several pimarane-and abietane-type resin acids along with oleic and palmiticfatty acids [68]. Other works are based on the character-ization of microorganisms that occur in soils, wood andnatural water systems and that degrade resin acids [69].Several groups of bacteria from the Commamonas, Pseu-domonas, Sphingomonas, Zooglea and Alcaligenes generaare able to degrade a variety of organic substrates. SomeArthrobacter spp. are also reported to be degraders ofdehydroabietic and methyldehydroabietic acid. Variousbacteria from the Bacillus, Arthrobacter, Pseudomonas andFlavobacterium genera as well as Escherichia coli andAlcaligenes eutrophus have been identified as biodegradersthat use dehydroabietic acid as their sole carbon source,whereas Alcaligenes spp. have been found that use abieticacid as their sole carbon source [70]. Yu et al. [71] haveidentified several Sphingomonas spp. and Pseudomonasspp. that degrade abietic and dehydroabietic acid as well asseveral Pseudomonas spp. that use pimaranes as a carbonsource together with aliphatic and aromatic hydrocarbons.Mortiella isabellina, Chaetomium cochlioes, Corticumsasaki and Fomes annosus fungi are also reported todegrade dehydroabietic acid [69].
Mechanisms for the bacteriological degradation ofditerpenoids have been reported in the literature, whichshow that levopimaric, palustric and dehydroabietic acidsare degradated more rapidly than abietic and neoabieticacids, whereas pimaric and isopimaric acids are the mostresistant to degradation [69]. Several biological degradationpathways through which abietanes are broken down bymicroorganisms have been reported in the literature [69].
Scheme 1 Scheme of the proposed pathways for oil biodegradation
1267
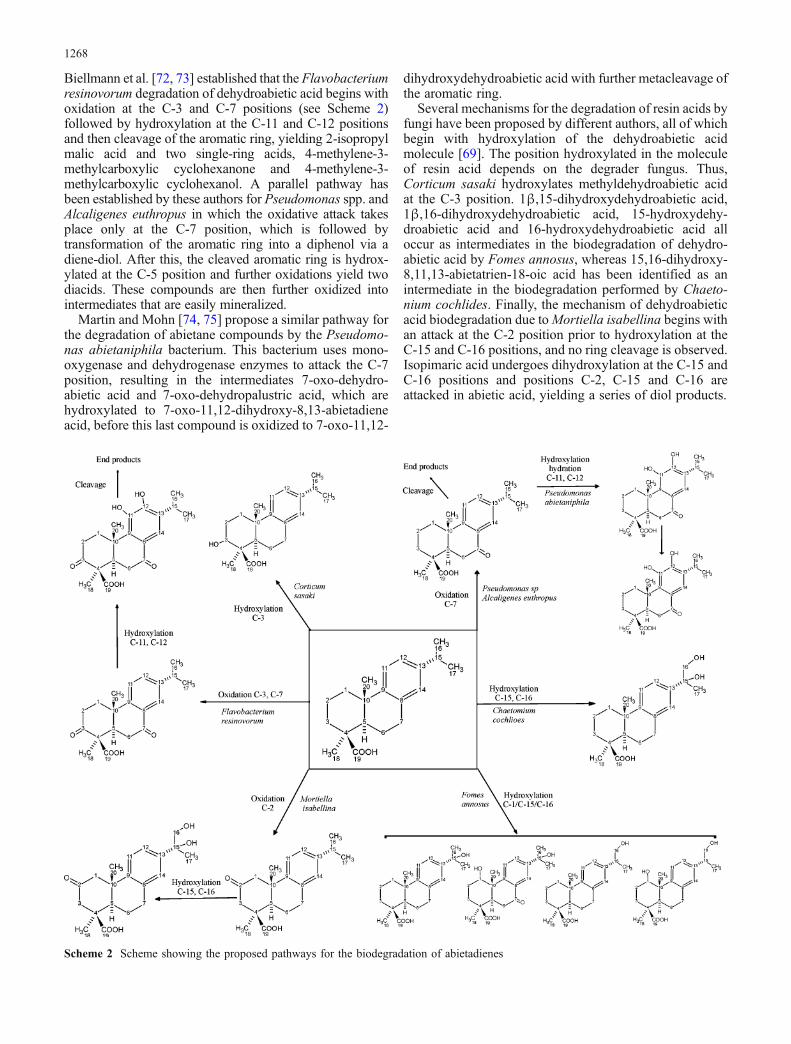
Biellmann et al. [72, 73] established that the Flavobacteriumresinovorum degradation of dehydroabietic acid begins withoxidation at the C-3 and C-7 positions (see Scheme 2)followed by hydroxylation at the C-11 and C-12 positionsand then cleavage of the aromatic ring, yielding 2-isopropylmalic acid and two single-ring acids, 4-methylene-3-methylcarboxylic cyclohexanone and 4-methylene-3-methylcarboxylic cyclohexanol. A parallel pathway hasbeen established by these authors for Pseudomonas spp. andAlcaligenes euthropus in which the oxidative attack takesplace only at the C-7 position, which is followed bytransformation of the aromatic ring into a diphenol via adiene-diol. After this, the cleaved aromatic ring is hydrox-ylated at the C-5 position and further oxidations yield twodiacids. These compounds are then further oxidized intointermediates that are easily mineralized.
Martin and Mohn [74, 75] propose a similar pathway forthe degradation of abietane compounds by the Pseudomo-nas abietaniphila bacterium. This bacterium uses mono-oxygenase and dehydrogenase enzymes to attack the C-7position, resulting in the intermediates 7-oxo-dehydro-abietic acid and 7-oxo-dehydropalustric acid, which arehydroxylated to 7-oxo-11,12-dihydroxy-8,13-abietadieneacid, before this last compound is oxidized to 7-oxo-11,12-
dihydroxydehydroabietic acid with further metacleavage ofthe aromatic ring.
Several mechanisms for the degradation of resin acids byfungi have been proposed by different authors, all of whichbegin with hydroxylation of the dehydroabietic acidmolecule [69]. The position hydroxylated in the moleculeof resin acid depends on the degrader fungus. Thus,Corticum sasaki hydroxylates methyldehydroabietic acidat the C-3 position. 1β,15-dihydroxydehydroabietic acid,1β,16-dihydroxydehydroabietic acid, 15-hydroxydehy-droabietic acid and 16-hydroxydehydroabietic acid alloccur as intermediates in the biodegradation of dehydro-abietic acid by Fomes annosus, whereas 15,16-dihydroxy-8,11,13-abietatrien-18-oic acid has been identified as anintermediate in the biodegradation performed by Chaeto-nium cochlides. Finally, the mechanism of dehydroabieticacid biodegradation due toMortiella isabellina begins withan attack at the C-2 position prior to hydroxylation at theC-15 and C-16 positions, and no ring cleavage is observed.Isopimaric acid undergoes dihydroxylation at the C-15 andC-16 positions and positions C-2, C-15 and C-16 areattacked in abietic acid, yielding a series of diol products.
Scheme 2 Scheme showing the proposed pathways for the biodegradation of abietadienes
1268

Experimental
Solvents, reagents, culture media and biocides
Analytical reagents and reference materials The followingreagents were used to treat the samples: hexamethyldisila-zane (HMDS) and methyl chloroformate (MCF) (purity99%) (Sigma-Aldrich, Steinheim, Germany). Absolute pyr-idine and chloroform at 98% for GC (Acros, Cambridge,MA, USA). The absolute methanol used for analysis wasfrom Carlo Erba, Rodano, Italy. The sodium hydrogencar-bonate used for analysis was from Panreac, Barcelona,Spain. Turpentine was from Guinama, Valencia, Spain.Tween 80, a polyoxyethylene (20) sorbitan monolaurate,was from Aldrich Chemicals, Milwaukee, WI, USA. Thediterpenoid resin of colophony was supplied by RCM(Barcelona, Spain), the triterpenoid resin of mastic wassupplied by Agar-Agar (Vigo, Spain), and the sun-thickenedlinseed oil was supplied by Caremi Pigmentos (Sevilla,Spain).
Culture media TSB media (Scharlau Chemie, Barcelona,Spain) was used for bacteria cultures and CM liquidmedium (yeast extract 0.5%, malt extract 0.5% andglucose 1%) was used for fungi cultures. Solid mediawere obtained by adding 2 wt% of Agar-Agar (Sigma, St.Louis, MO, USA) to the previously described liquidmedia.
Biocides Three microbiocides were tested: Biotin N(METATIN N 5810/101) (ÁCIMA Chemical Industries,Buchs, Switzerland), New Des (STERAMINA H) (C.T.S.Madrid, Spain) and Nipagine (Phase, Firenze, Italy).
Instrumentation and procedures
Experiments were carried out with an Agilent 6890 N GC(Agilent Technologies, Palo Alto, CA, USA) coupled to anAgilent 5973 N mass spectrometer, and equipped with bothon-column and pyrolysis injection systems. Capillarycolumn HP-5 MS (stationary phase 5% phenyl–95%methylpolysiloxane, 30 m×0.25 mm I.D., 0.25 μm filmthickness) was used in order to get adequate separation ofcomponents.
Ions were generated by electron ionisation (70 eV) in theionisation chamber of the mass spectrometer. The massspectrometer was scanned from m/z 20 to m/z 800, with acycle time of one second. Agilent Chemstation softwareG1701CA MSD was used for GC–MS control, peakintegration and mass spectral evaluation. The tuning of themass spectrometer was checked using perfluorotributyl-amine (PFTBA). EI mass spectra were acquired in the totalion monitoring mode and the peak area (TIC) data wereused to obtain peak area percentage values [49]. Thetemperatures of the interface and the source were 280 °Cand 150 °C, respectively. The Wiley Library of MassSpectra was used to identify compounds. This libraryprovides a lot of information on oils, diterpenoid and
triterpenoid compounds. Additionally, prior work per-formed in our laboratory, in which a wide collection ofunaged and aged proteins, drying oils and terpenoidcompounds were analysed, provided abundant informationon the components of most of these materials, so that wehave at our disposal a complete in-house library of massspectra specific to methyl, ethyl and trimethilsilyl deriva-tives of short, medium chain saturated and unsaturated fattyacids, mono-, sesqui-, di- and triterpenoid compounds andtheir oxidized products, along with other compoundsusually present in artworks [31, 37, 62]. The data containedin the in-house library of mass spectra are in goodagreement with the results reported from specialists in thefield of artwork analysis [49, 56, 88].
Analysis of the mastic model varnish was carried outusing the GC–MS system. The chromatographic conditionsused were: initial temperature of the gas chromatograph170 °C, a gradient of 5 °C/min up to 185 °C, ramping up at15 °C/min to 250 °C, and finally a gradient of 5 °C/min to295 °C, which was held for 20 minutes. The carrier gas wasHe with an inlet pressure of 72.5 kPa and a split ratio of1:20.
Experiments on linseed oil and colophony model varnishspecimens were carried out with an integrated systemcomprising a CDS Pyroprobe 1000 heated filamentpyrolyser (Analytical Inc., New York, USA) and theabove-described gas chromatograph.
The chromatographic conditions used in the pyrolysis ofspecimens containing colophony and linseed oil were asfollows: the initial oven temperature of 100 °C wasincreased at 5 °C/min to 295 °C, and then held for 8 min.The helium gas flow was set to 1.2 and 1.6 ml/min,respectively, for colophony and linseed oil. The inletpressure of the carrier gas was 67.5 and 72.5 kPa,respectively. The electronic pressure control was set toconstant flow mode with vacuum compensation. Thepyrolysis interface and the inlet were set to 250 °C. Thesamples were injected in the split mode (split ratio 1:80).
A CLIMAS culture chamber (Génesis Instrumentación,Madrid, Spain) was used for incubation trials.
Derivatization of samples
Off-line derivatisation
The derivatisation procedure was based on a methodologyapplied previously for the analysis of triterpenoid resins,which was developed in our laboratory [31]. Samples weredissolved in 50 μl methanol–pyridine (4:1, v/v). Then10 μl MCF were added. The reaction mixture was vortexedfor about 10 s and the derivatives were extracted in 50 μlchloroform containing 1% MCF. After that, 50 μl of asaturated solution of NaHCO3 was added and the mixturewas shaken carefully. A 1-μl aliquot of the organic phasewas injected into the GC–MS system.
1269

On line derivatisation–pyrolysis
Samples scraped from the specimens using a scalpel wereplaced in a microquartz pyrolysis tube, and then two smallportions of quartz wool were introduced on both sides ofthe quartz tube to avoid undesirable displacements of thesample. Afterwards, 5–10 μl of HMDS were added. Lastly,the sample was placed in the pyrolysis coil and introducedinto the pyrolysis interface, which was kept at 250 °C andpyrolyzed at 600 °C for 10 s.
Preparation of test specimens
A series of test specimens were prepared by dissolving 90 gof colophony in 100 ml of turpentine. The freshly preparedmodel varnish was applied by brushing three successivefine layers onto glass slides of a standard size (24×80 mm).The specimens were then dried at room temperature fortwo weeks. These supports reproduce the varnishedsurface, so that any potential interferences produced bythe organic materials present in artworks are avoided.
A second series of linseed oil specimens was preparedby spreading this product on glass slides so that a thin filmwas formed in a similar way to that described above forcolophony model varnish. The specimens were dried atroom temperature for four weeks.
A third series of test specimens was prepared bydissolving 57 g of mastic in 100 ml of turpentine. Thespecimens were then dried at room temperature forfour weeks.
Each series consisted of three different batches ofinoculated specimens, including the complete collectionof microorganisms studied, which were prepared inDecember 2003, January 2004 and June 2004. Eachbatch was prepared with different strains of every species.Additionally, three different areas, each ~20 mm2, wereinoculated on the specimens prepared for each microor-ganism and terpenoid resin studied in the two first batches,and four different areas were inoculated on the specimensprepared for each microorganism in the third batch, so thatten replicates were obtained for the biodegradated resinsand linseed oil in total for each microorganism and liningmaterial.
Blank samples were taken by scraping a scalpel acrossthe model varnish spread on the glass slides, avoiding theinoculated area. The varnish removed was then ground inan agate mortar and finally split into quarters until theamount of sample was suitable for analysis (about 1 g). Thesame procedure was carried out to retrieve samples fromthe inoculated area.
Microorganism inoculation and incubation
The microorganisms studied were recognized biodeteriora-tion agents, selected after an extensive review of theliterature [76–84]. Fungi and bacteria that could originatebiodeterioration processes in colophony varnish were
chosen. All of the species studied are ubiquitoussaprophytes, commonly found in the air, and they camefrom stock collections belonging to the Spanish Collectionof Type Cultures (CECT, Colección Española de CultivosTipo, Universidad de Valencia, Valencia, Spain). Themicroorganisms used were:
– Fungi: Aspergillus niger (An)(CECT−2088), Penicil-lium chrysogenum (Pc)(CECT−2306), Trichodermapseudokoningii (Tp) (CECT−2937), Cladosporiumcladosporoides(Cc)CECT
– Bacteria: Streptomyces cellulofans (Sc)(CECT 3242),Bacillus amyloliquefaciens (Ba)(CECT 493), Arthro-bacter oxydans (Ao)(CECT 386)
Lyophilized collection stocks were hydrated in CM(fungi) and TSB (bacteria) and incubated for 18 h in aculture chamber (28 °C). Afterwards, they were inoculatedonto solid CM medium (fungi) and TSA (bacteria). After48 h, these cultures were used to prepare dense microbialsuspensions on distilled water, which were inoculated ontospecimens containing the studied oil and terpenoidmaterials after performing centrifugation to eliminate anypossible remaining culture media.
Several drops (50 μl) of the suspension containing themicroorganisms were applied onto the support using amicropipette, so that they covered a defined area (~20 mm2/drop) of the solidified model varnish, and then the inoculatedtest specimen was placed in the incubator.
Previous experiments helped us to establish the optimalconditions for incubating the model varnish specimensbeforehand. The experimental conditions in which theincubation was carried out were 28 °C and 60% relativehumidity.
Biocide treatments
Experiments to test for chemical changes in the modelvarnish test specimens after biocide treatments were alsoperformed. Three biocides were selected: Biotin N, NewDes and Nipagine. Biotin N and New Des were applied as a5% m/v aqueous solution onto the surfaces of the varnishtest specimens, whereas a 0.1% m/v aqueous solution ofNipagine was prepared, to which Tween 80 was added atconcentration of 0.01% m/v, and then this mixture wasapplied onto the solidified model varnish specimens. Twoapplications were made with the help of a thin pencil in allcases. Additionally, a fourth series of test specimens wereprepared in which a 1%m/v solution of Nipagine in ethanolwas mixed with the freshly prepared varnish and thus aliquid varnish was obtained containing the biocide at aconcentration of 0.1% m/v. After this, the varnish wasapplied to the glass slides using a thin pencil, and the testspecimens prepared were dried in the same way asdescribed earlier for the series of pure varnish testspecimens. Finally, the specimens were stored for35 days at 28 °C and 60% RH.
1270
Results and discussion
Assessment of biodegradation experiments
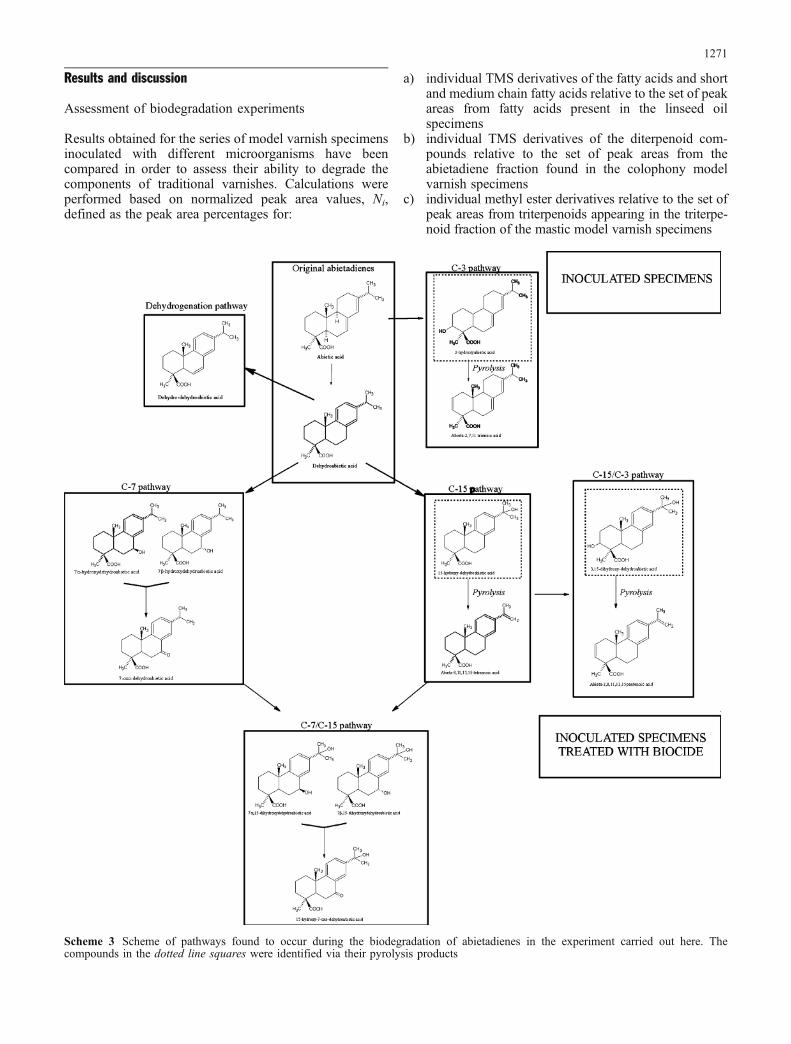
Results obtained for the series of model varnish specimensinoculated with different microorganisms have beencompared in order to assess their ability to degrade thecomponents of traditional varnishes. Calculations wereperformed based on normalized peak area values, Ni,defined as the peak area percentages for:
a) individual TMS derivatives of the fatty acids and shortand medium chain fatty acids relative to the set of peakareas from fatty acids present in the linseed oilspecimens
b) individual TMS derivatives of the diterpenoid com-pounds relative to the set of peak areas from theabietadiene fraction found in the colophony modelvarnish specimens
c) individual methyl ester derivatives relative to the set ofpeak areas from triterpenoids appearing in the triterpe-noid fraction of the mastic model varnish specimens
Scheme 3 Scheme of pathways found to occur during the biodegradation of abietadienes in the experiment carried out here. Thecompounds in the dotted line squares were identified via their pyrolysis products
1271

Ni is defined as follows:
Ni ¼ Ai
Pn
i¼1Ai
� 100 (1)
Here Ai is the peak area of each methyl or TMS derivativeof the n components found in the chromatogram orpyrogram.
In order to simplify the analysis, the Ni values weregrouped into k sets corresponding to:
a) the three categories of saturated fatty acids, unsaturatedfatty acids and short/medium chain fatty acidsoccurring in linseed oil specimens
b) the original abietadiene and pimaradiene acids andtheir oxidation products identified in the varnishspecimens studied and formed via each degradationpathway, for the colophony specimens [85, 86].
Thus, a new parameter, Nk, was defined as the sum of thenormalised peak area values of:
a) the derivatives of the fatty acids included in each oneof the three established categories
b) the derivatives of the pimaradiene and abietadieneacids and their oxidation compounds occurring in eachspecific pathway
Nk ¼Xm
j¼1
Nj; (2)
where Nj is one of the m TMS derivatives correspondingto:
a) the kth set of fatty acids.b) the kth set of original pimaradiene, abietadiene and
dehydroabietic acids and their oxidation productsinvolved in the same pathway (Scheme 3).
In order to discriminate between the changes incomposition due to the microbial attack and other changesdue to natural or thermal ageing that take place in theculture chamber during incubation, the value Nko, corre-sponding to the blank (a sample extracted from anuninoculated area of the specimen) in each specimen wassubtracted from the Nk value for the sample extracted fromthe inoculated area of the specimen.
ΔNk ¼ Nk � Nko (3)
The resulting ΔNk values are indicative of the change incomposition that occurs during the incubation process, andthey allow us to evaluate the biodegradation that occurredin the specimens tested.
A total of ten replicates for each model varnish wereused. The repeatability obtained in the experimentalconditions used to perform the analyses and the relativestandard deviations (calculated using the formula RSD=
(standard deviation/mean of the normalized peakarea)×100%) were in the range 1–5% for all thecompounds analysed. From statistical mathematics, therelative standard deviation of the Nk value is similar tothat estimated for the individual normalised peak areapercentage Nj.
Assessment of the effects of biodegradation dueto microorganisms incubated on linseed oil
Figure 1 shows a pyrogram for a sample of linseed oilextracted from an area of incubated specimen in whichBacillus amyloliquefaciens was inoculated. The pyrogramis dominated by the long chain fatty acids palmitic acid(14) and stearic acid (17) together with weak peakscorresponding to docosanoic acid (21) and eicosanoic acids(19). The presence of oleic acid (9-octadecenoic acid) andlinoleic acid (9,12-octadecadienoic acid) is indicative ofthe relatively young age of the linseed oil tested. The largeamount of oleic acid found in the specimens analysed isdue to the lower reactivity exhibited by this monounsatu-rated fatty acid compared to the doubly unsaturated linoleicacid.
The occurrence of peaks corresponding to a series ofshort/medium chain fatty mono- and diacids (1–11, 13) inthe chromatogram shows that oxidation processes tookplace during the incubation of the specimens. It isinteresting to note that a series of short/medium chainmonounsaturated monoacids have been also found in thechromatograms from samples of blank and inoculatedspecimens (2, 4, 6 and 8). Their presence in the chromato-gram could be due to the occurrence of dehydrogenationreactions during the pyrolysis process rather than biodeg-radation processes, as suggested by the similarity of theresults to those obtained by Colombini et al. [87] frompyrolysed shellac resin.
Finally, the occurrence of the 2,3-bis[(TMS)oxy]propylester of hexadecanoic acid (20), the diglyceride of thepalmitic acid, is another interesting finding. The presenceof this compound indicates that total and partial hydrolysisreactions of the original triacylglycerols, forming a cross-liked structure in the dried linseed oil, took place in thespecimens analysed.
Figure 2 summarizes the ΔNk values corresponding tothe three categories of short, medium and long chain fattyacids considered. They were obtained from the mean valueof the normalized peak area, Ni, of the TMS derivative ofeach fatty acid and its degradation products that occurs inthe model oil medium. The values for the short/mediumchain fatty acids (ΔNsa) formed upon the oxidation orhydrolysis of the triacylglycerols depended on the type ofmicroorganism inoculated onto the specimen. Samplesfrom specimens inoculated with the bacterium Bacillusamyloliquefaciens and the fungi Cladosporium cladospor-oides and Trichoderma pseudokoningii exhibited a reducedlevels of these acids (<4%). Based on the pathwaysproposed by other authors and presented in Sect. 2.1, thisnegative shift has been tentatively ascribed to the
1272

predominant transformation of triacylglycerols via enzy-mic mechanisms promoted by microorganism metabolism,in which intermediate molecules consisting of short/medium chain fatty acids yield more volatile ketones andalcohols as end-products. These compounds should be lostwhen the specimen is dried during incubation.
Significant decreased unsaturated fatty acid contents(ΔNua) in the inoculated samples compared to the blankswere expected as result of intense microbial or fungalactivity. However, for both bacteria and fungi, the Nua
values were found to be similar in both the blank andinoculated samples from the specimens, suggesting thatmicroorganism attack on the lipidic substrate was weak.
Assessment of the effects of biodegradation dueto microorganisms incubated on colophony modelvarnish
Figure 3 shows a pyrogram for a sample of the modelvarnish extracted from the area of the specimen inoculatedwith Bacillus amyloliquefaciens.
Based on the pathways proposed by other authors andpresented in Sect. 2.2, the results obtained suggest thatthree oxidation pathways involving the C-3, C-15 and C-7positions took place in the inoculated varnish specimensanalysed (see Scheme 3). The first pathway, involving theC-15 and C-3 positions, probably begins with hydroxyl-ation at the C-15 position. Abieta-8,11,13,15-tetraenoicacid, formed from 15-hydroxydehydroabietic acid as adehydration product during the pyrolysis [56], wasidentified in the chromatograms. Further hydroxylation at
Fig. 2 Bar chart summarizingΔNk values obtained fromthe specimens prepared withlinseed oil
Fig. 1 Pyrogram corresponding to a sample of the linseed oilspecimen. 1, Hexanoic acid, TMS ester; 2, heptenoic acid , TMSester; 3, heptanoic acid, TMS ester; 4, octenoic acid, TMS ester; 5,octanoic acid, TMS ester; 6, nonenoic acid, TMS ester; 7, nonanoicacid, TMS ester; 8, decenoic acid, TMS ester; 9, decanoic acid, TMSester; 10, octanedioic acid, di-TMS ester; 11, nonanedioic acid, di-TMS ester; 12, tetradecanoic acid, TMS ester; 13, decanedioic acid,
di-TMS ester; 14, hexadecanoic acid, TMS ester; 15, 9-octadecenoicacid, TMS ester; 16, isomer of 9-octadecenoic acid, TMS ester; 17,octadecanoic acid, TMS ester; 18, 9,12-octadecadienoic acid, TMSester; 19, eicosanoic acid, TMS ester; 20, hexadecanoic acid acid,2,3-bis[(TMS)oxy]propyl, TMS ester; 21, docosanoic acid, TMSester. Identification was achieved using the Wiley Mass SpectraLibrary
1273

C-3 position yields 3,15-dihydroxydehydroabietic acid,identified from its pyrolytic dehydration product abieta-2,8,11,13,15-pentenoic acid [56].
An alternative biodegradation pathway involving theC-7 position is also tentatively proposed. Both 7α- and7β-hydroxydehydroabietic acid (isomers) were identifiedin the chromatograms. In a second step, these compoundsare oxidized to 7-oxodehydroabietic acid. No diphenols,bicyclic diketones or short chain mono and diacids andrelated oxidised compounds that are formed upon cleavageof the aromatic ring occurred in the samples analysed,according to the bacteria degradation models proposedpreviously [69].
A third biodegradation pathway, yielding 3-hydroxy-abietic acid from abietic acid, is proposed to have takenplace in the specimens studied. Abieta-2,7,11-trienoic acid,a dehydration product of the 3-hydroxyabietic acid [56]formed during the pyrolysis process, has been identified.
A fourth degradation pathway is also proposed from theresults obtained in the specimens studied. This mechanismimplies the dehydrogenation of dehydroabietic acid to formthe dehydrodehydroabietic acid. This process, previouslyreported by Scalarone et al. [88, 89], takes place inartificially and naturally aged varnish film samples ofcolophony.
It should be noted that all of the pathways that occurredin the specimens analysed here have been described asprocesses that usually take place in naturally and artificiallyaged diterpenoid varnishes prepared as thin films [49, 88].
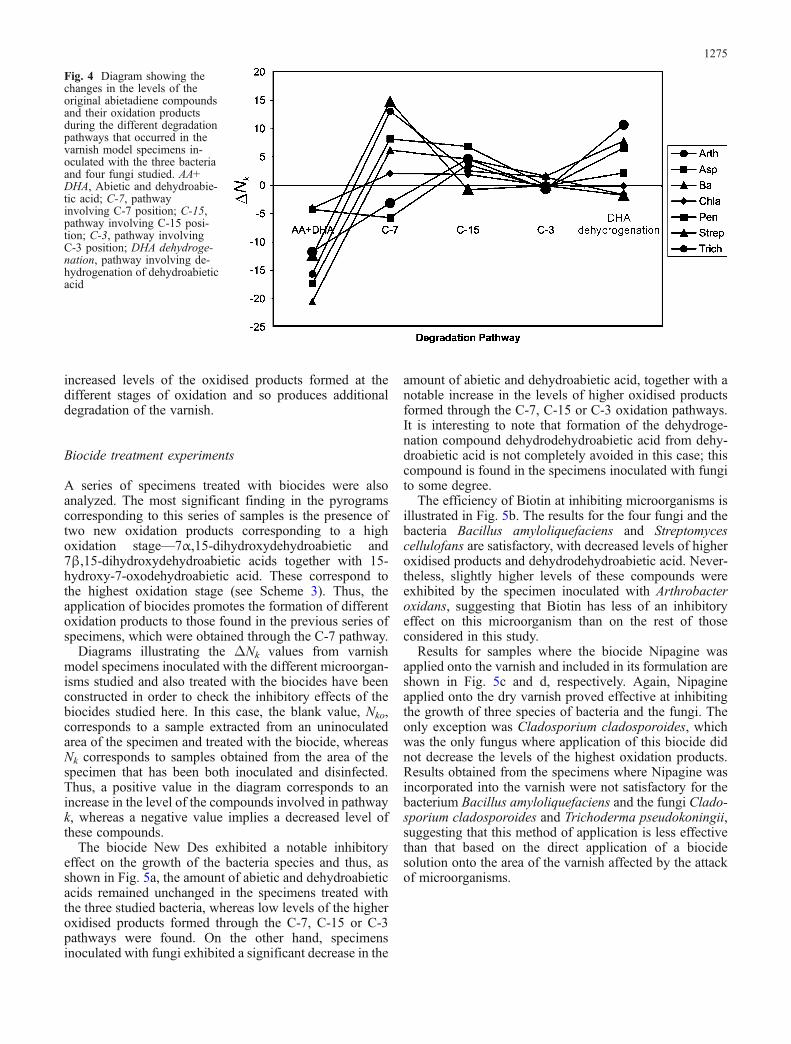
Figure 4 summarizes the ΔNk values, calculated fromthe set of abietadiene compounds from the model varnish
and their oxidation products, for the different pathways.The results obtained suggest that the bacteria Bacillusamyloliquefaciens and Streptomyces cellulofans and thefungi Aspergillius niger, Cladosporium cladosporoidesand Trichoderma pseudokoningii have similar effects onthe model varnish: oxidation through the C-7 position is thepredominant pathway together with the dehydrogenation ofdehydroabietic acid. Oxidation processes involving theC-15/C-3 positions and the C-3 position are less popular.The specimens inoculated with the bacterium Arthrobacteroxydans and the fungus Penicillium chrysogenum exhib-ited different behaviour from the other specimens; the C-7pathway was not significantly enhanced.
In addition to the previous comments, it should be notedthat no significant difference was observed between thebehaviour of fungi and that of bacteria in terms of theoxidation processes promoted on abietadiene acids. More-over, a decrease in the content of pimaradiene acid wasobserved not only in the specimens inoculated with fungi—as was previously reported for some specific biodegraderfungi on diterpenoid substrates—but was also observed inthe specimens inoculated with bacteria.
The lack of specific compounds reported to beintermediates or end biodegradation products associatedwith the enzymatic attacks of specific diterpenoid biode-graders leads us to conclude that the bacteria and fungiconsidered in our study do not act as specific biodegradersof diterpenoid substrates. Nevertheless, the results obtainedhere suggest that the presence of these microorganismsfavours, in general, the oxidative processes that usuallyoccur in substrates prepared as thin films. This results in
Fig. 3 Pyrogram corresponding to a sample of model colophonyvarnish inoculated with Bacillus amyloliquefaciens. 1, Pimaric acid,TMS ester; 2, Sandaracopimaric acid, TMS ester; 3, isopimaric acid,TMS ester; 4, dehydrodehydroabietic acid, TMS ester, 5, dehydroabieticacid, TMS ester; 6, abietic acid, TMS ester; 7, abieta-2,8,11,13,15-
pentenoic acid, TMS ester; 8, abieta-8,11,13,15-tetraenoic acid, TMSester; 9, 7β-hydroxydehydroabietic acid, TMS ether, TMS ester; 10, 7α-hydroxydehydroabietic acid, TMS ether, TMS ester; 11, abieta-2,7,13-trienoic acid, TMS ester; 12, 15-hydroxydehydroabietic acid, TMS ether,TMS ester; 13, 7-oxo-dehydroabietic acid, TMS ester
1274
increased levels of the oxidised products formed at thedifferent stages of oxidation and so produces additionaldegradation of the varnish.
Biocide treatment experiments
A series of specimens treated with biocides were alsoanalyzed. The most significant finding in the pyrogramscorresponding to this series of samples is the presence oftwo new oxidation products corresponding to a highoxidation stage—7α,15-dihydroxydehydroabietic and7β,15-dihydroxydehydroabietic acids together with 15-hydroxy-7-oxodehydroabietic acid. These correspond tothe highest oxidation stage (see Scheme 3). Thus, theapplication of biocides promotes the formation of differentoxidation products to those found in the previous series ofspecimens, which were obtained through the C-7 pathway.
Diagrams illustrating the ΔNk values from varnishmodel specimens inoculated with the different microorgan-isms studied and also treated with the biocides have beenconstructed in order to check the inhibitory effects of thebiocides studied here. In this case, the blank value, Nko,corresponds to a sample extracted from an uninoculatedarea of the specimen and treated with the biocide, whereasNk corresponds to samples obtained from the area of thespecimen that has been both inoculated and disinfected.Thus, a positive value in the diagram corresponds to anincrease in the level of the compounds involved in pathwayk, whereas a negative value implies a decreased level ofthese compounds.
The biocide New Des exhibited a notable inhibitoryeffect on the growth of the bacteria species and thus, asshown in Fig. 5a, the amount of abietic and dehydroabieticacids remained unchanged in the specimens treated withthe three studied bacteria, whereas low levels of the higheroxidised products formed through the C-7, C-15 or C-3pathways were found. On the other hand, specimensinoculated with fungi exhibited a significant decrease in the
amount of abietic and dehydroabietic acid, together with anotable increase in the levels of higher oxidised productsformed through the C-7, C-15 or C-3 oxidation pathways.It is interesting to note that formation of the dehydroge-nation compound dehydrodehydroabietic acid from dehy-droabietic acid is not completely avoided in this case; thiscompound is found in the specimens inoculated with fungito some degree.
The efficiency of Biotin at inhibiting microorganisms isillustrated in Fig. 5b. The results for the four fungi and thebacteria Bacillus amyloliquefaciens and Streptomycescellulofans are satisfactory, with decreased levels of higheroxidised products and dehydrodehydroabietic acid. Never-theless, slightly higher levels of these compounds wereexhibited by the specimen inoculated with Arthrobacteroxidans, suggesting that Biotin has less of an inhibitoryeffect on this microorganism than on the rest of thoseconsidered in this study.
Results for samples where the biocide Nipagine wasapplied onto the varnish and included in its formulation areshown in Fig. 5c and d, respectively. Again, Nipagineapplied onto the dry varnish proved effective at inhibitingthe growth of three species of bacteria and the fungi. Theonly exception was Cladosporium cladosporoides, whichwas the only fungus where application of this biocide didnot decrease the levels of the highest oxidation products.Results obtained from the specimens where Nipagine wasincorporated into the varnish were not satisfactory for thebacterium Bacillus amyloliquefaciens and the fungi Clado-sporium cladosporoides and Trichoderma pseudokoningii,suggesting that this method of application is less effectivethan that based on the direct application of a biocidesolution onto the area of the varnish affected by the attackof microorganisms.
Fig. 4 Diagram showing thechanges in the levels of theoriginal abietadiene compoundsand their oxidation productsduring the different degradationpathways that occurred in thevarnish model specimens in-oculated with the three bacteriaand four fungi studied. AA+DHA, Abietic and dehydroabie-tic acid; C-7, pathwayinvolving C-7 position; C-15,pathway involving C-15 posi-tion; C-3, pathway involvingC-3 position; DHA dehydroge-nation, pathway involving de-hydrogenation of dehydroabieticacid
1275

Fig. 5 Diagram showing thechanges in the levels of theoriginal abietadiene compoundsand their oxidation productsduring the different degradationpathways that occurred in thevarnish model specimens in-oculated with the three bacteriaand four fungi studied and alsotreated with (a) New Des, (b)Biotin, (c) Nipagine-Tween, and(d) Nipagine. AA+DHA, abieticand dehydroabietic acid; C-7,pathway involving C-7 position;C-15, pathway involving C-15position; C-3, pathway involv-ing; dDHA, pathway involvingdehydrogenation of dehydro-abietic acid
1276

Assessment of the effects of biodegradation dueto microorganisms incubated on mastic varnish
Figure 6 shows a chromatogram for a mastic specimeninoculated with Trichoderma pseudokoningii. GC–MSanalysis resulted in the identification of the compoundspresent in the mastic specimens, which included not onlythe original triterpenoids but also the related dehydroge-nation and oxidization products.
The free triterpenoid fraction of mastic was characterizedby the occurrence of hexakisnor-dammaran-3,20-dione (3),nor-olean-17-en-3-one (9), hydroxydammarenone (11),moronic acid (13), 20,24-epoxy-25-hydroxy-dammaren-3-one (14), oleanonic acid (15) and masticadienonic acid (16),together with small amounts of degradation products such as3-methoxy-28-norolean-12-ene (5), olean-18-en-3-one (7),olean-Δ18-en-3-one (8), 28-norolean-12,17-dien-3-one (10)and 3β-hydroxy-28-norolean-17-en-6-al (12), as well as twoalteration products corresponding to peaks (4) and (6) whosestructures could not be completely elucidated.
The values of ΔNi obtained for the set of microorgan-isms tested were widely scattered, which suggests that nospecific biodegradation pathways took place under theexperimental conditions applied. Table 1 summarizes thevalues of ΔNi obtained for the set of specimens analysed.The values of ΔNi obtained from the analyses of samplesextracted from the specimens inoculated with Arthrobacteroxydans revealed slight decreases of ~2–6% in thecontents of dammarane and the mono- and diketone-typeoleanane triterpenoids together with an increase in theunidentified degradation compound. Bacillus amylolique-faciens specimens exhibited notable positive values for
olean-18-en-3-one and 28-norolean-12,17-dien-3-one—degradation products from the ketone type—in parallelwith a 5.46% reduction in the content of the originaltriterpenoid from the ketone type, nor-olean-17-en-3-one.Decreases in the Ni values for the original hydroxyl-ketone-and carboxylic acid-type triterpenoids hydroxydammare-none and masticadienonic acid were found in thespecimens treated with Streptomyces cellulofans, alongwith a decrease in that for nor-olean-17-en-3-one. In-creased Ni values for the degradation compound 20,24-epoxy-25-hydroxy-dammaran-3-one in the Penicillium-chrysogenum and Trichoderma pseudokoningii specimenswere also observed. ΔNi values of <4% were obtained forthe triterpenoids identified in the samples from thespecimens treated with fungi, except from the slightincrease in the Ni values obtained for an unidentifiedalteration compound in the Cladosporium chladosporidesspecimen, the degradation compound 20, 24-epoxy-25-hydroxy-dammaran-3-one in the Trichoderma pseudo-koningii specimen, and those for Penicillium chrysogenum.Lastly, it is interesting to note that specimens inoculatedwith Penicillium chrysogenum behaved in a differentmanner, exhibiting a significant increase of in the originaltriterpenoids in parallel with a notable decrease in thecharacteristic oxidized and altered compounds.
Conclusions
A method for evaluating the effect of microbiologicalattack on the chemical compositions of terpenoid-based oilvarnishes has been proposed, which is based in the Py–
Fig. 6 TIC chromatogram of methyl ester derivatives in a sample ofmastic extracted from the area of an incubated specimen upon whichthe Trichoderma pseudokoningii microorganism was inoculated. 1,Hexadecanoic acid methyl ester; 2, octadecanoic acid methyl estera;3, hexakisnor-dammaran-3,20-dioneb; 4, unknown compound; 5, 3-methoxy-28-norolean-12-enec; 6, unknown compound; 7, olean-18-en-3-onec; 8, olean-Δ18-en-3-onec; 9, 28-norolean-17-en-3-onea;
10, 28-norolean-12,17-dien-3-onec; 11, hydroxydammarenoneb; 12,3β-hydroxy-28-norolean-17-en-6-alc; 13, moronic acid methylestera; 14, 20,24-epoxy-25-hydroxy-dammaren-3-oneb; 15, oleano-nic acid methyl estera; 16, masticadienonic acid methyl esterb.Identification of compounds: a Wiley Mass Spectra Library; b massspectra published in [90]; c mass spectra published in [91, 92]
1277
GC–MS technique with on-line derivatization with HMDS,and the GC–MS technique using methyl chloroformate asthe derivatisation reagent.
The results obtained from the Py–GC–MS analyses oflinseed oil specimens inoculated with the bacteria indicatethat microbial attack hardly affects the chemical composi-tion of the drying oil.
Analyses performed on a series of model colophonyvarnish specimens inoculated with three different bacteriaand four different fungi usually found in paintingcollections suggest that the most common degradationpathway is an oxidation process that involves hydroxyl-ation and further oxidation of abietadienoic acids at the C-7position, together with the dehydrogenation of dehydro-abietic acid. Thus, no mechanism commonly found tooccur during enzymatic attacks from specific diterpenoidbiodegraders was observed, and the degradation pathwaysthat occurred in the performed trials were usually the onesthat occur in thin-film substrates due to natural ageing.
Nevertheless, these results regarding the biodegradationpathways that occurred in the experiments performed hereare of a speculative nature and further work is required tocorroborate them.
The analysis of specimens where biocides were appliedto uninoculated varnish permitted the processes resulting inundesirable degradation of the varnish due to interactionsbetween the biocide and the varnish components to beinvestigated. In this case, 7α, 15-dihydroxydehydroabieticacid, 7β, 15-dihydroxydehydroabietic acid and 15-hy-droxy-7-oxodehydroabietic acid were identified, which areoxidation products that were not found in the specimensthat had not been treated with biocide. This suggests thatthe application of biocide products results in differentoxidation processes to those that occur as result of thebiodegradation.
The GC–MS method enabled the identification of anumber of oxidation products formed from the originaltriterpenoid compounds during resin incubation due to the
Table 1 Values of ΔNi corresponding to the changes in the levels of triterpenoid compounds from the inoculated specimens due to thebiodegradation process
Triterpenoid ΔNI
Arthrobacteroxydans
Bacillusamyloliquefaciens
Streptomycescellulofans
Aspergilliusniger
Cladosporyumcladosporoydes
Penicilliumchrysogenum
Trichodermapseudokoningii
Mono and diketonetypeHexakisnor-dammaran-3,20-dione
−2.54 −0.65 −1.86 0.24 0.70 <0.01 −3.80
Olean-18-en-3-onea 0.91 1.48 2.13 0.49 −1.05 −5.52 0.41Olean-Δ18-en-3-onea −4.09 −0.17 1.39 −1.84 −1.83 −9.38 −2.14Nor-olean-17-en-3-one
−6.55 −5.46 8.49 2.35 2.79 −23.25 1.80
28-Norolean-12,17-dien-3-onea
−8.57 6.38 1.84 4.16 −1.27 −28.60 0.82
Methoxy type3-Methoxy-28-norolean-12-enea
0.27 0.46 2.69 0.60 −0.29 <0.01 −2.64
Hydroxy-ketone typeHydroxydammarenone 0.05 0.19 −4.30 −0.23 −0.07 −6.67 −0.193-Hydroxy-28-norolean-17-en-6-ala
−1.91 <0.01 <0.01 <0.01 <0.01 <0.01 <0.01
20,24-Epoxy-25-hydroxy-dammaran-3-onea
<0.01 <0.01 −6.84 −3.85 <0.01 7.08 6.44
Carboxylic acid typeMoronic acid −0.22 −2.64 <0.01 −0.86 1.78 47.46 <0.01Oleanonic acid 1.03 <0.01 <0.01 −1.41 <0.01 <0.01 <0.01Masticadienonic acid 0.22 1.33 −5.97 −1.98 −2.02 46.49 −1.96Alteration compoundDegradationcompound
7.76 0.48 2.09 3.03 −2.82 <0.01 1.53
Degradationcompound
−0.33 −1.39 0.34 −0.69 4.06 −13.44 −0.27
aDegradation product
1278

increase in temperature and relative humidity, resulting inaccelerated ageing of the resin. A comparison betweensamples extracted from the inoculated and uninoculatedareas of the specimens showed that microbial attack hardlyaffects the chemical composition of the resin, and enabledthe stability of the major triterpenoids to be studied.Bacteria species exhibit slightly higher values of ΔNi foraltered compounds, suggesting that these microorganismsare more effective at attacking triterpenoid resins thanfungi, with the exception of Penicillium chrysogenum,whose high values of ΔNi show that this fungus has astrong inhibitory effect on the oxidation processes thatoccur in the resin.
Acknowledgements Financial support is gratefully acknowledgedfrom the National Spanish “I+D+I MCYT” Projects BQU2001-2776-C03-01/03 and CTQ2005-09339-CO3-01, which were alsosupported by FEDER funds and “I+D Generalitat Valenciana”project GV04B-441.
References
1. Rebrikova L (1978) A study of microflora of museums textilesand methods of their desinfection and prophylaxis. In: Proc 5thTriennial Meeting of the ICOM Committee for Conservation,1–8 October 1978, Zagreb, pp 156–159
2. Bergeaud C, Hulot JF, Roche A (1997) La dégradation despeintures sur toile. Méthode d’examen des alterations. EcoleNationale du Patrimoine, Paris, pp 87–89
3. Nicolaus K (1999) Handbuch der Gemälderestaurierung.Könemann, Köln, pp 204–207
4. Briski F, Krstic D, Jagic R (2001) Stud Conserv 46:14–225. Mayumi I, Koyano M (1991) Int Biodeter Biodegr 28:23–356. Arai H, Matsumura N, Murakita H (1990) Induced foxing by
components found in foxed areas. In: Proc 9th TriennialMeeting of the ICOM Committee for Conservation, 26–31August 1990, Dresden, Germany, pp 801–803
7. Janda K (2005) Int Biodeter Biodegr 55:149–1528. Kinderlerer JL (1993) Int Biodeter 32:213–2249. Kinderlerer JL (1994) Int Biodeter 33:345–35410. Giuliani MR, Nugari MP (1993) A case of fungal biodeteriora-
tion on an ancient textile. In: Preprints of 10th TriennialMeeting of the ICOM Committee for Conservation, 22–27August 1993, Washington, DC, pp 305–307
11. Chapman JS (2003) Int Biodeter Biodegr 51:133–13812. Maupéou CD, Volfovsky C, Mortamet JG (1990) Architecture et
décors peints. Ministère de la Culture de la Communication desGrands Travaux et du Bicentenaire, Amiens, France, pp 14–20
13. Waddington J, Fenn J (1988) Collection Forum 4:25–3114. Williams RS, Waddington JB, Fenn J (1990) Collection Forum
6:65–7515. Arteni SC, Sánchez-Posada M (1984) From handcraft to mass
production: notes on the manufacture of oil painting materials.In: Proc 7th Triennial Meeting of the ICOM Committee forConservation, 10–14 Sept 1984, Copenhagen, Denmark
16. Sabin AH (1927) The industrial and artistic technology of paintvarnish. Wiley, New York p 437
17. Cennini C (1988) El libro del arte. Akal, Madrid, p14118. da Vinci L (1995) Tratado de pintura, 3rd edn. Akal, Madrid, p 43319. Gettens RJ, Stout GL (1966) Painting materials: a short
encyclopaedia. Dover, New York, pp 34–3520. Palomino A (1947) El museo pictórico y escala óptica. Aguilar,
Madrid, p 74521. Mills JS, White R (1987) The organic chemistry of museum
objects. Butterworths, London, p 150
22. Hiscox GD, Hopkins AA (1997) El recetario industrial.Gustavo Gili, Mexico, p 961
23. Mills JS, White R (1987) The organic chemistry of museumobjects. Butterworths, London, p 89
24. Walch K, Koller J, Baumer U, Fischer CH (1996) The redlaquer-work in the cabinet of miniatures in the MunichResidenz, Vol II. Preprints of 11th Triennial Meeting ICOMCommittee for Conservation, 1–6 September 1996, Edinburgh,UK, p 920
25. Orellana F (1755) Tratado de barnices y charoles. Imprenta J.García, Valencia, Spain
26. Hurst GH (1901) Painter’s colours, oils, and varnishes, apractical manual. Charles Griffin & Company, London (cited inMills JS, White R (1987) The organic chemistry of museumobjects. Butterworths, London, p 36)
27. van den Berg JDJ, van den Berg KJ, Boon JJ (2002) JChromatogr A 950:195–211 (and references therein)
28. Van der Doelen GA, van den Berg KJ, Boon JJ (1998) StudConserv 43:249–264
29. Papageorgiou VP, Bakola-Christianopoulou MN, ApazidouKK, Psarros EE (1997) J Chromatogr A 769:263–273
30. Mills JS, White R (1977) Stud Conserv 22:12–3131. de la Cruz-Cañizares J, Doménech-Carbó MT, Gimeno-
Adelantado JV, Mateo-Castro R, Bosch-Reig F (2005) JChromatogr A 1093:177–194
32. Mills JS (1966) Stud Conserv 11:92–10733. Schilling MR, Khanjian HP, Carson DM (1997) Techne 5:71–7834. Pancella R, Bart R (1989) Z Kunsttechnol Konservierung
3:101–11135. Koller J, Baumer U (1995) Z Kunsttechnol Konservierung 9:
286–29536. Nowik W (1995) Stud Conserv 40:120–12637. Mateo R, Doménech-Carbó MT, Peris-Martínez V, Gimeno-
Adelantado JV, Bosch-Reig F (1997) J Chromatogr A778:373–381
38. van der Berg KJ, van der Horst J, Boon JJ, Sudmeijer OO(1998) Tetrahedron Lett 39:2645–2648
39. Wright MM, Wheals BB (1987) J Anal Appl Pyrol 11:195–21140. Challinor JM (1990) J Anal Appl Pyrol 18:233–24441. Chiavari G, Galetti GC, Lanterna G, Mazzeo R (1993) J Anal
Appl Pyrol 24:227–24242. van den Berg JDJ, Boon JJ, van den Berg KJ, Fiedler I, Miller
MA (1998) Anal Chem 70:1823–183043. Challinor JM (1996) J Anal Appl Pyrol 37:185–19744. Wright MM, Wheals BB, J Anal Appl Pyrol (1987) 11:195–21145. Shedrinsky AM, Stone RE, Baer NS (1991) J Anal Appl Pyrol
20:229–23846. Stevanato R, Rovea M, Carbini M, Favretto D, Traldi P (1997)
Rapid Commun Mass Spectrom 11:286–29447. Shedrinsky AM, Wampler T, Indictor N, Baer NS (1989) J Anal
Appl Pyrol 15:393–41248. Bocchini P, Traldi P (1998) J Mass Spectrom 33:1053–106249. van der Berg KJ, Pastorova I, Spetter L, Boon JJ (1996) State of
oxidation of diterpenoid Pinaceae resins in varnish, wax liningMaterial, 18th century resin oil paint, and a recent copperresinate glaze, Vol II. Preprints of 11th Triennial MeetingICOM Committee for Conservation, 1–6 September 1996,Edinburgh, UK, p 930
50. Pastorova I, van der Berg KJ, Boon JJ, Verhoeven JW (1997) JAnal Appl Pyrol 43:41–57
51. van der Berg KJ, van der Horst J, Boon JJ (1999) Recognitionof copals in aged resin/oil paints and varnishes, Vol II. Preprintsof 12th Triennial Meeting of the ICOM Committee forConservation, 29 Aug–3 Sept 1999, Lyon, France, p 855
52. van der Werf ID, van der Berg KJ, Schmitt S, Boon JJ (2000)Stud Conserv 45:1–18
53. van der Berg KJ, Ossebaar J, Van Keulen H (2002) Analysis ofcopal resins in 19th century oil paints and resin/oil varnishes.In: Van Grieken R, Janssens P, Van’t dack K, Meersman L (eds)Art. University of Antwerp, Antwerp
1279

54. Prati S, Smith S, Chiavari G (2004) Chromatographia 59:227–23155. Challinor JM (2001) J Anal Appl Pyrol 61:3–3456. Anderson K, Winans RE (1991) Anal Chem 63:2901–290857. Hardell H, Nilvebrant NO (1991) J Anal Appl Pyrol 52:1–1458. Scalarone D, Lazzari M, Chiantore O (2001) J Anal Appl Pyrol
58–59:503–51259. Cappitelli F, Learner T, Chiantore O (2002) J Anal Appl Pyrol
63:339–34860. Vane CH (2003) Int Biodeter Biodegr 51:67–7561. Chiavari G, Fabbri D, Prati S (2002) Chromatographia
55:611–61662. Osete-Cortina L, Doménech-Carbó MT (2005) J Chromatogr A
1065:265–27863. vand der Weerd J, van Loon A, Boon JJ (2005) Stud Conserv
50:3–2264. Wexler H (1964) Chem Rev 64:591–61165. Mallégol J, Lemaire J, Gardette JL (2000) Progr Org Coat
39:107–11366. Mills JS, White R (1987) The organic chemistry of museum
objects. Butterworths, London, pp 33–3467. Erhardt D, Tumosa CS, Mecklenburg MF (2005) Stud Conserv
50:143–15068. Hemingway RW, Greaves H (1973) Tappi 56:189–19269. Liss SN, Bicho PA, Saddler JN (1997) Can J Microbiol
75:599–611 (and references cited therein)70. Cross BE, Myers PL (1968) Biochem J 108:303–31071. Yu Z, Stewart GR, Mohn WW (2000) Appl Environ Microb
66:5148–515472. Biellmann F, Branlant G, Gero-Robert M, Poiret M (1973)
Tetrahedron 29:1237–124173. Biellmann JF, Branlant G, Gero-Robert M, Poiret M (1973)
Tetrahedron 29:1227–123674. Mohn WW, Martin VJJ, Yu Z (1999) Wat Sci Tech 40:273–28075. Martin VJ, Mohn WW (2000) J Bacteriol 182:3784–379376. Ciferri O (1999) Appl Environ Microbiol 65:879–885
77. Giacobini C, Firpi M (1981) Problemi di microbiologia neidipinti su tela. In: Atti del Convenzione sul Restauro delleOpere d’Arte, Opificio delle Pietre Dure e Laboratorio diRestauro di Firenze, Polistampa, Florence, Italy, p 203–211
78. Giacobini C, De Cicco MA, Tiglie I, Accardo G (1988)Actinomycetes and biodeterioration in the field of fine art. In:Houghton DR, Smith RN, Eggins HOW (eds) Biodeterioration,vol. 7. Elsevier, New York, pp 418–423
79. Giacobini C, Pedica M, Spinucci M (1991) 31 Problems andfuture projects on the study of biodeterioration: mural andcanvas paintings. In: Proceedings of the 1st InternationalConference on the Biodeterioration of Cultural Property.Macmillan India, New Delhi, India, pp 275–286
80. Ross RT (1963) Adv Appl Microbiol 5:217–23481. Seves AM, Sora S, Ciferri O (1996) Int Biodeter Biodegr
37:215–22482. Strelczyc A (1981) Paintings and sculptures. In: Rose AH (ed)
Microbialdeterioration. Academic, London, pp 203–23483. Walsh J (2001) Int Biodeter Biodegr 48:16–2584. Zyska BJ (2002) Int Biodeter Biodegr 49:73–8385. Proefke ML, Kenneth L, Rinehart L (1992) Am Soc Mass
Spectrom 3:582–58986. Sandermann W (1960) Naturharze, terpentinöl, tallöl. Springer,
Berlin Heidelberg New York87. Colombini MP, Bonaduce I, Gautier G (2003) Chromatographia
58:357–36388. Scalarone D, Lazzari M, Chiantore O (2002) J Anal Appl Pyrol
64:345–36189. Scalarone D, Van der Horst J, Boon JJ, Chiantore O (2003) J
Mass Spectrom 38: 607–61790. van der Doelen GA (1999) PhD thesis. University of Amsterdam,
Amsterdam91. Assimopoulou AN, Papageorgiou VP (2005) Biomed Chromatogr
19:586–60592. Assimopoulou AN, Papageorgiou VP (2005) Biomed Chromatogr
19:285–311
1280






![a n a l ysis Journal of Rosarin and Mirunalini J Bioanal Biomed 2011 · PDF file · 2018-01-09Rosarin and Mirunalini J Bioanal Biomed 2011, 3:4 ... biomedical applications [2]. Ag](https://img.pdfslide.net/doc/110x75/5ab139a47f8b9ac3348c1a36/a-n-a-l-ysis-journal-of-rosarin-and-mirunalini-j-bioanal-biomed-2011-2018-01-09rosarin.jpg)



![Publications of Prof. Dr. Ron M.A. Heeren · Anal. Bioanal. Chem. (2019) 412 2277–2289 [DOI] LipostarMSI: comprehensive, vendor-neutral software solu-tion for visualisation, data](https://img.pdfslide.net/doc/110x75/607f6ec6e01cf8332536e5df/publications-of-prof-dr-ron-ma-heeren-anal-bioanal-chem-2019-412-2277a2289.jpg)


















